Embed Size (px)
Citation preview

Available online at www.sciencedirect.com
08 (2007) 238–246www.elsevier.com/locate/yexnr
Experimental Neurology 2
Amperometric measures of age-related changes in glutamateregulation in the cortex of rhesus monkeys
Jorge E. Quintero a,b,c, Brian K. Day a,b,c, Zhiming Zhang a,b, Richard Grondin a,b,Michelle L. Stephens a,b,c, Peter Huettl a,b,c, François Pomerleau a,b,c,
Don M. Gash a,b, Greg A. Gerhardt a,b,c,⁎
a Department of Anatomy and Neurobiology, University of Kentucky, Lexington, KY, USAb Morris K. Udall Parkinson's Disease Research Center of Excellence, University of Kentucky, Lexington, KY, USA
c Center for Microelectrode Technology, University of Kentucky, Lexington, KY, USA
Received 22 June 2007; revised 1 August 2007; accepted 7 August 2007Available online 22 August 2007
Abstract
L-glutamate (glutamate) is the principal excitatory neurotransmitter of the central nervous system and is involved in altered neural functionduring aging and in neurodegenerative diseases. Relatively little is known about the mechanisms of glutamate signaling in the primate brain, inpart, because there is an absence of a method capable of rapidly measuring glutamate in either a non-clinical or a clinical setting. We haveaddressed this paucity of information by measuring extracellular glutamate at 1 Hz in the pre-motor and motor cortices of young, middle-aged, andaged monkeys using a minimally invasive amperometric recording method. In the motor cortex, mean resting glutamate levels were five timeshigher in the aged group compared to the young group while the pre-motor cortex showed an increasing trend in resting glutamate levels that wasnot statistically significant. In addition, we measured rapid, phasic glutamate release after local pressure-ejection of nanoliter volumes of eitherisotonic 70 mM potassium (to stimulate glutamate release) or 1 mM glutamate (to study glutamate uptake) into the pre-motor and motor cortex. Inthe pre-motor cortex, we measured reproducible glutamate uptake signals that had a significantly decreased (47%) rate of glutamate uptake in agedanimals compared to young animals. However, following a 70 mM potassium delivery, we did not observe any consistent changes in evokedrelease between young versus aged animals. Using these non-clinical microelectrodes to measure glutamate signaling in the brain, our resultssupport the hypothesis that the glutamatergic system undergoes reorganization with aging of the central nervous system.© 2007 Published by Elsevier Inc.
Keywords: Resting glutamate levels; Rhesus monkeys; Motor cortex; Enzyme-based microelectrode; Amperometry; Non-human primates; Translational research
Introduction
Glutamate is the predominant excitatory neurotransmitter inthe mammalian central nervous system (CNS). Thus, alterationsin the glutamatergic system contribute to a variety of neuro-degenerative diseases (Greenamyre and Young, 1989; Green-amyre, 2001; Beart and O'Shea, 2007), as well as changes in theCNS during aging (Strolin et al., 1990; Morrison, 2003).
⁎ Corresponding author. Center for Microelectrode Technology, University ofKentucky, 306 Whitney-Hendrickson Building, Lexington, KY 40536-0098,USA. Fax: +1 859 257 5310.
E-mail address: [email protected] (G.A. Gerhardt).
0014-4886/$ - see front matter © 2007 Published by Elsevier Inc.doi:10.1016/j.expneurol.2007.08.002
Normally, presynaptic receptors and glutamate transporterson neurons and glia work in concert to tightly regulateextracellular glutamate levels and control glutamate neurotrans-mission. However, age-related decreases in N-methyl-D-asparticacid (NMDA) and α-amino-5-hydroxy-3-methyl-4-isoxazolepropionic acid (AMPA) receptors (Wenk et al., 1989; Tamaruet al., 1991; Magnusson and Cotman, 1993a,b; Gazzaley et al.,1996; Hof et al., 2002; Bai et al., 2004) and transporterexpression (Nickell et al., 2007) may increase the susceptibilityof neurons to excitotoxic damage (Liu et al., 1996; Brewer,2000; Arundine and Tymianski, 2003). Potentially, extracellularglutamate levels could serve as an indicator of neuronal diseasesassociated with aging or age-related neurodegeneration exceptthat the relationship between changes in the nervous system and

Fig. 1. Microelectrodes used to measure glutamate from the neocortex ofmonkeys. The double pair of microelectrode sites serves as a backup redundancyas well as the basis for a self-referencing configuration. For all experiments, theceramic microelectrode received a coating of enzyme solution on the lower pairof microelectrode sites and an enzyme-free solution on the upper pair of sites.Micropipettes were placed so that the tip was centered among the four sites (see⁎ on figure). The Pt electrode sites are 15×333 μm in size with a 30 μm spacingbetween them and a 100 μm spacing between pairs. Scale bar: 100 μm.
239J.E. Quintero et al. / Experimental Neurology 208 (2007) 238–246
resting glutamate levels is unclear. Some reports indicate anage-related decrease in extracellular glutamate levels (Strolinet al., 1990; Kaiser et al., 2005) while others have not identifieda difference (Segovia et al., 2001).
The neurons in the pre-motor and motor cortex form acomplex network of glutamatergic interneurons, afferent pro-jections, and pyramidal neurons that project to several CNSareas that include the striatum and spinal cord. Motor deficitsassociated with pre-motor and motor cortical reorganization areone of the hallmark symptoms of neurodegenerative diseasesand age-related neurodegenerative diseases including amyo-trophic lateral sclerosis and Parkinsons's disease (Kuninobuet al., 1993; Sabatini et al., 2000; Lefaucheur, 2005). Forexample, in Parkinsons's disease, the dopaminergic neuronsarising from the substantia nigra undergo a loss of function thatindirectly results in diminished activity through the thalamo-cortical pathway that leads to a subsequent decrease in gluta-matergic output from the motor cortex (Wichmann and DeLong,1996). Thus, the role the pre-motor and motor cortex play inage-related neurodegenerative diseases along with their super-ficial location in the non-human primate brain make theseregions critical targets for studying the mechanisms ofglutamatergic neurotransmission in vivo.
The changes that the non-human primate CNS undergoeswith age remain mostly unknown even though informationgathered from this model is invaluable to an understanding of thehuman CNS. While most in vivo studies that examine theglutamatergic system have been in rodents, only a few studieshave been in non-human primates (Graham et al., 1989; Klinget al., 1993; Bradberry, 2000; Kodama et al., 2002; Galvan et al.,2003) with investigators employing the technique of micro-dialysis in these latter studies to measure glutamate neurotrans-mission. Although the technique of microdialysis is widespreadand is used in the clinic for neurochemical monitoring in humans(Mendelowitsch et al., 1998; Alves et al., 2005; Berg et al.,2006), it suffers from two main limitations. One, the large size ofthe microdialysis probe imparts extensive damage to the sur-rounding tissue area and necessitates the sampling of an area atleast 200 μm away from the probe (Borland et al., 2005), andtwo, the low sampling rate required for measurements hinder thetechnique's effectiveness in describing the rapid temporal natureof signaling events for fast neurotransmitters such as glutamate(Timmerman and Westerink, 1997).
In this report, we employed the electrochemical method ofamperometry in conjunction with enzyme-based microelec-trodes (Burmeister et al., 2000; Burmeister and Gerhardt, 2001;Burmeister et al., 2002) to characterize normal glutamateregulation during aging in the neocortex of anesthetized rhesusmonkeys, and we established a foundation for examiningglutamate neurotransmission in a clinical setting. These studiesexamined three components of the glutamatergic system: first,resting extracellular glutamate levels in both the pre-motor andmotor cortices of young, middle-aged, and aged rhesusmonkeys; second, the glutamate uptake transport systemfollowing the delivery of exogenous glutamate; and last, rapidkinetics of glutamate release and uptake after local applicationof high potassium stimulation.
Materials and methods
Animals
Twelve adult female rhesus monkeys (Macaca mulatta) wereused for this study and divided into three groups: young (5.4±0.1 years old, mean±S.E.M., n=4), middle-aged (17.4±0.6 yearsold, n=4), and aged (26.2±0.3 years old, n=4) (DeRousseau,1985). Animals were maintained in a 12 h/12 h light/dark cyclewith glutamate measurements occurring during the daytime.Glutamate measurements were conducted once for six animals,but for one young, three middle-aged, and two aged animals, therecordings were repeated because of missing data points in thefirst trial.
All procedures were conducted in the Laboratory AnimalFacilities of the University of Kentucky, fully accredited by theAssociation for Assessment and Accreditation of LaboratoryAnimal Care International (AAALACI). The Animal Care andUse Committee of the University of Kentucky approved allprotocols. In addition, veterinarians skilled in the health careand maintenance of non-human primates supervised all animalcare. Prior studies have not found relationships betweenmenstrual cycle and age-related changes in neurotransmittersystems (unpublished data). Therefore, no attempts were madein the present study due to the small sample size.
Microelectrodes
Ceramic-based microelectrodes were assembled, enzyme-coated, and used in a self-referencing mode as previouslydescribed (Burmeister and Gerhardt, 2001; Burmeister et al.,2002; Day et al., 2006). Briefly, multisite (4 platinum sites,15 μm×333 μm each) microelectrodes (Fig. 1) were dip-coated

240 J.E. Quintero et al. / Experimental Neurology 208 (2007) 238–246
in Nafion® and then dried in an oven at 175 °C for 4 min. Next,the lower pair of microelectrode sites was coated with a smallamount (b1 μL) of 1% glutamate oxidase (Associates of CapeCod, East Falmouth, MA)/1% bovine-serum albumin (BSA)/0.125% glutaraldehyde while the upper pair of microelectrodesites was coated with an enzyme-free solution of 1% BSA/0.125% glutaraldehyde creating a self-referencing microelec-trode. All coated microelectrodes were allowed to cure aminimum of 48 h prior to use. On the day of the experiment, themicroelectrodes were calibrated with glutamate in vitro at 37 °Cin a phosphate buffered solution (pH 7.4) to generate a standardcurve (sensitivity N4 pA/μM), to determine the limit ofdetection (=3 times the signal to noise ratio; b1.0 μM), andto assess the selectivity of glutamate relative to the endogenouselectroactive compound, ascorbic acid (N30:1, glutamate:ascorbic acid, in the non-self reference recording mode).
Surgical procedures
All procedures were conducted using sterile field conditionsin fully equipped operating rooms specifically designed to meetthe specifications of hospital surgical suites. Following sedationwith ketamine hydrochloride (∼20 mg/kg; i.m.) plus atropinesulfate (∼0.04 mg/kg; i.m.), each animal was placed in amagnetic resonance imaging (MRI)-compatible stereotaxicapparatus in a ventral–lateral position and maintained underanesthesia with the use of isoflurane (∼1.5%) while the animalbreathed on its own. Coordinates for microelectrode implanta-tion were determined by MRI for each monkey prior to thesurgery. Coordinates for the pre-motor cortex were: AP:21.0 mm–25.0 mm from the ear bars; L: 9.0 mm–14.0 mmfrom midline. Meanwhile, coordinates for the motor cortexwere AP: 15.0 mm–20.0 mm from the ear bars; L: 10.0 mm–11.0 mm from the midline. Small (2–3 mm diameter) holeswere drilled in the skull directly over the targeted areas. Alateral pocket (5 mm×50 mm) was made in the fascia at theright posterior extent of the incision using blunt dissection and aAg/AgCl reference electrode (Model RE-5B, BioanalyticalSystems, West Lafayette, IN) was inserted into the pocket andheld in position by sutures (3.0 silk). Sterile physiological saline(0.9% NaCl) irrigation was used to maintain ionic contactbetween the reference electrode and the exposed brain. Allanimals recovered from the surgery without incident.
In vivo glutamate measurements
Solutions were delivered using micropipettes, formed from1.0 mm o.d., 0.58 mm i.d. glass (A-M Systems, Everett, WA),with an inside tip diameter of 10–15 μm. Micropipettes wereattached to the microelectrode array with the micropipette tipcentered among the four microelectrode sites at a distance of70–110 μm. Prior to implantation, the micropipette was filledwith either a 70 mM potassium solution (70 mM KCl, 79 mMNaCl, 2.5 mM CaCl2, pH 7.0–7.4) or a 1 mM glutamatesolution [1 mM glutamate in physiological (0.9%) saline, pH7.0–7.4]. The fully prepared microelectrode/micropipette as-semblies then were implanted into the brain based on individual
coordinates predetermined with MRI for each monkey. Afterinitially advancing the center of the microelectrode array to thecortical surface, the assembly was sequentially lowered to therecording depths (−1.0 mm to −3.0 mm below the surface ofthe brain in 0.5 mm increments) by a microdrive (MO-8;Narishige, East Meadow, NY) to obtain multiple measurementsfrom individual monkeys. A counterbalanced design was usedin this study where glutamate release from one hemisphere wasmeasured after a depolarizing stimulation with 70 mM KCl andglutamate uptake, from the other hemisphere, after the deliveryof 1 mM glutamate. For each solution, a separate microelec-trode/micropipette assembly was used. In each hemisphere, thepre-motor cortex was recorded in one pass, and then the micro-electrode was removed from the brain, rinsed with physiologicalsaline, and implanted in the motor cortex for a second pass.
The microelectrodes were given 15–30 min to equilibrate perimplantation prior to the start of local solution applications anda 2- to 5-min equilibration period between varying depths.Resting glutamate levels were assessed at each depth in realtime by self-referencing methods (Burmeister and Gerhardt,2001). The 70 mM KCl solutions or 1 mM glutamate solutionswere ejected at 1-min intervals at each depth until at least two tofive reproducible glutamate responses were recorded. Deliveryof solution volumes (25 nL up to 400 nL; 0.1 s–3 s, 2–12 p.s.i.)were controlled by a pressure-ejection system (Picospritzer II;Parker Hannifin Corp., Cleveland) and monitored using astereomicroscope fitted with a reticule (Friedemann andGerhardt, 1992). Extracellular glutamate levels were measuredusing constant potential amperometry (+0.7 V vs. Ag/AgClreference) controlled by a FAST16 electrochemical recordingsystem (Quanteon, LLC, Nicholasville, KY).
Statistical analysis
Using the FAST16 system for on-line signal assessment anda custom Excel-based software for off-line signal analysis,glutamate signaling dynamics were monitored and differenceswere compared among the age groups.
In the hemispheres where 70 mM KCl-evoked responseswere measured, resting glutamate level measures were calcu-lated for each depth by a constant, applied potential paired-channel subtraction (Day et al., 2006). Briefly, at the end ofequilibration, a stable glutamate recording of 1-min durationwas evaluated to calculate the resting glutamate levels (GluRest;in μM) using the equation:
GluRest ¼ ðIþGluOx � I�GluOxÞ=mþGluOx ð1Þ
where I is the average current measured from both enzyme-coatedmicroelectrode sites (+GluOx) and both non-enzyme-coatedelectrode sites (−GluOx) and m is the slope of the standard curvecalculated from the calibration of each glutamate sensitive micro-electrode prior to the experiment. Average glutamate concentra-tions over these time intervals were calculated at each depth ineach brain region.
Signals within the amplitude range of 3–15 μM wereanalyzed in experiments where 70 mM KCl was used. In
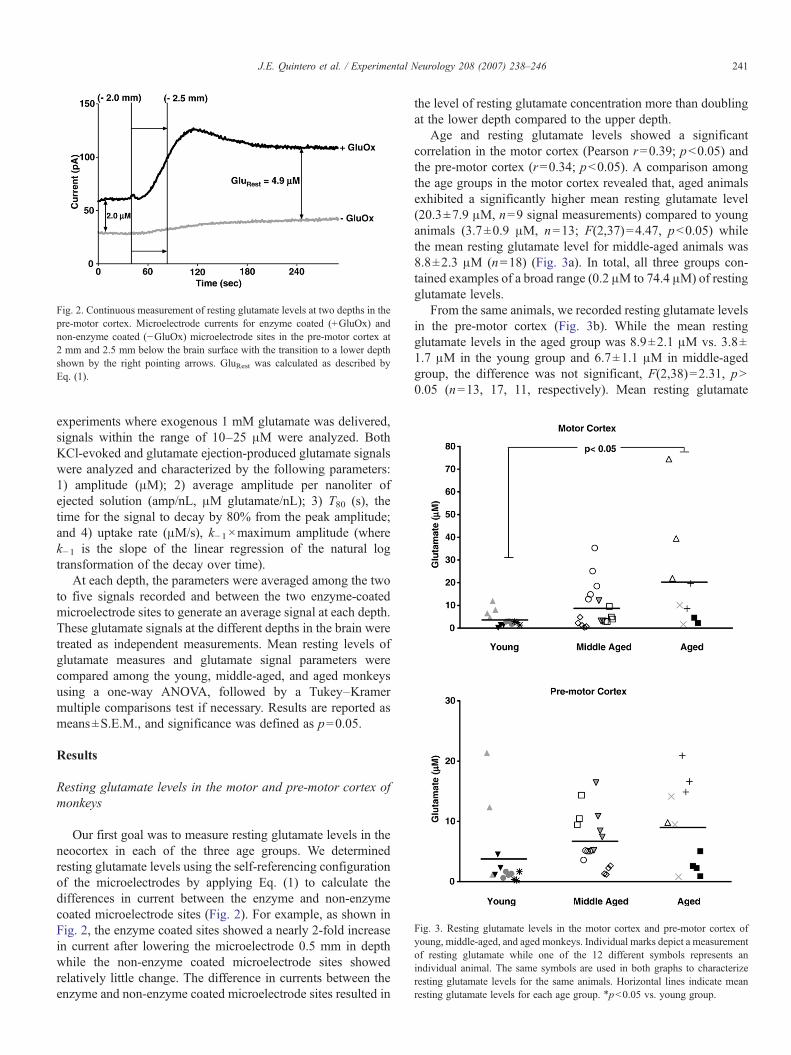
Fig. 2. Continuous measurement of resting glutamate levels at two depths in thepre-motor cortex. Microelectrode currents for enzyme coated (+GluOx) andnon-enzyme coated (−GluOx) microelectrode sites in the pre-motor cortex at2 mm and 2.5 mm below the brain surface with the transition to a lower depthshown by the right pointing arrows. GluRest was calculated as described byEq. (1).
Fig. 3. Resting glutamate levels in the motor cortex and pre-motor cortex ofyoung, middle-aged, and aged monkeys. Individual marks depict a measurementof resting glutamate while one of the 12 different symbols represents anindividual animal. The same symbols are used in both graphs to characterizeresting glutamate levels for the same animals. Horizontal lines indicate meanresting glutamate levels for each age group. ⁎pb0.05 vs. young group.
241J.E. Quintero et al. / Experimental Neurology 208 (2007) 238–246
experiments where exogenous 1 mM glutamate was delivered,signals within the range of 10–25 μM were analyzed. BothKCl-evoked and glutamate ejection-produced glutamate signalswere analyzed and characterized by the following parameters:1) amplitude (μM); 2) average amplitude per nanoliter ofejected solution (amp/nL, μM glutamate/nL); 3) T80 (s), thetime for the signal to decay by 80% from the peak amplitude;and 4) uptake rate (μM/s), k−1×maximum amplitude (wherek−1 is the slope of the linear regression of the natural logtransformation of the decay over time).
At each depth, the parameters were averaged among the twoto five signals recorded and between the two enzyme-coatedmicroelectrode sites to generate an average signal at each depth.These glutamate signals at the different depths in the brain weretreated as independent measurements. Mean resting levels ofglutamate measures and glutamate signal parameters werecompared among the young, middle-aged, and aged monkeysusing a one-way ANOVA, followed by a Tukey–Kramermultiple comparisons test if necessary. Results are reported asmeans±S.E.M., and significance was defined as p=0.05.
Results
Resting glutamate levels in the motor and pre-motor cortex ofmonkeys
Our first goal was to measure resting glutamate levels in theneocortex in each of the three age groups. We determinedresting glutamate levels using the self-referencing configurationof the microelectrodes by applying Eq. (1) to calculate thedifferences in current between the enzyme and non-enzymecoated microelectrode sites (Fig. 2). For example, as shown inFig. 2, the enzyme coated sites showed a nearly 2-fold increasein current after lowering the microelectrode 0.5 mm in depthwhile the non-enzyme coated microelectrode sites showedrelatively little change. The difference in currents between theenzyme and non-enzyme coated microelectrode sites resulted in
the level of resting glutamate concentration more than doublingat the lower depth compared to the upper depth.
Age and resting glutamate levels showed a significantcorrelation in the motor cortex (Pearson r=0.39; pb0.05) andthe pre-motor cortex (r=0.34; pb0.05). A comparison amongthe age groups in the motor cortex revealed that, aged animalsexhibited a significantly higher mean resting glutamate level(20.3±7.9 μM, n=9 signal measurements) compared to younganimals (3.7±0.9 μM, n=13; F(2,37)=4.47, pb0.05) whilethe mean resting glutamate level for middle-aged animals was8.8±2.3 μM (n=18) (Fig. 3a). In total, all three groups con-tained examples of a broad range (0.2 μM to 74.4 μM) of restingglutamate levels.
From the same animals, we recorded resting glutamate levelsin the pre-motor cortex (Fig. 3b). While the mean restingglutamate levels in the aged group was 8.9±2.1 μM vs. 3.8±1.7 μM in the young group and 6.7±1.1 μM in middle-agedgroup, the difference was not significant, F(2,38)=2.31, pN0.05 (n=13, 17, 11, respectively). Mean resting glutamate
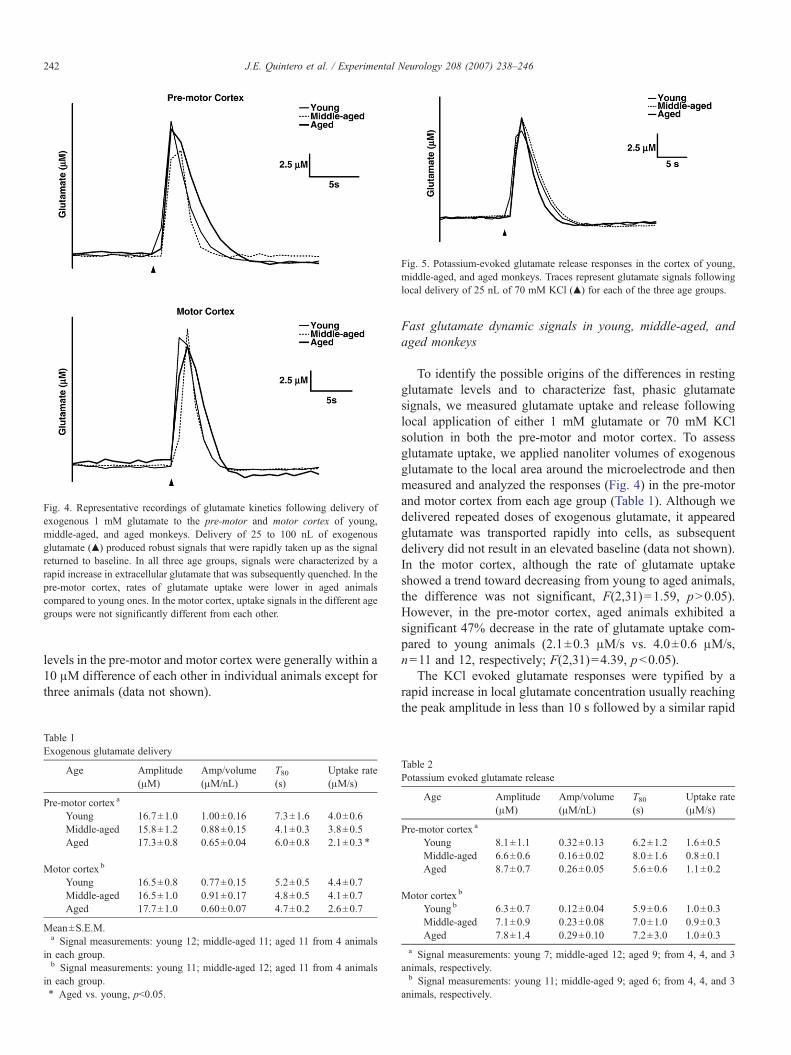
Fig. 5. Potassium-evoked glutamate release responses in the cortex of young,middle-aged, and aged monkeys. Traces represent glutamate signals followinglocal delivery of 25 nL of 70 mM KCl (▲) for each of the three age groups.
Fig. 4. Representative recordings of glutamate kinetics following delivery ofexogenous 1 mM glutamate to the pre-motor and motor cortex of young,middle-aged, and aged monkeys. Delivery of 25 to 100 nL of exogenousglutamate (▲) produced robust signals that were rapidly taken up as the signalreturned to baseline. In all three age groups, signals were characterized by arapid increase in extracellular glutamate that was subsequently quenched. In thepre-motor cortex, rates of glutamate uptake were lower in aged animalscompared to young ones. In the motor cortex, uptake signals in the different agegroups were not significantly different from each other.
242 J.E. Quintero et al. / Experimental Neurology 208 (2007) 238–246
levels in the pre-motor and motor cortex were generally within a10 μM difference of each other in individual animals except forthree animals (data not shown).
Table 1Exogenous glutamate delivery
Age Amplitude(μM)
Amp/volume(μM/nL)
T80(s)
Uptake rate(μM/s)
Pre-motor cortex a
Young 16.7±1.0 1.00±0.16 7.3±1.6 4.0±0.6Middle-aged 15.8±1.2 0.88±0.15 4.1±0.3 3.8±0.5Aged 17.3±0.8 0.65±0.04 6.0±0.8 2.1±0.3 ⁎
Motor cortex b
Young 16.5±0.8 0.77±0.15 5.2±0.5 4.4±0.7Middle-aged 16.5±1.0 0.91±0.17 4.8±0.5 4.1±0.7Aged 17.7±1.0 0.60±0.07 4.7±0.2 2.6±0.7
Mean±S.E.M.a Signal measurements: young 12; middle-aged 11; aged 11 from 4 animals
in each group.b Signal measurements: young 11; middle-aged 12; aged 11 from 4 animals
in each group.⁎ Aged vs. young, pb0.05.
Fast glutamate dynamic signals in young, middle-aged, andaged monkeys
To identify the possible origins of the differences in restingglutamate levels and to characterize fast, phasic glutamatesignals, we measured glutamate uptake and release followinglocal application of either 1 mM glutamate or 70 mM KClsolution in both the pre-motor and motor cortex. To assessglutamate uptake, we applied nanoliter volumes of exogenousglutamate to the local area around the microelectrode and thenmeasured and analyzed the responses (Fig. 4) in the pre-motorand motor cortex from each age group (Table 1). Although wedelivered repeated doses of exogenous glutamate, it appearedglutamate was transported rapidly into cells, as subsequentdelivery did not result in an elevated baseline (data not shown).In the motor cortex, although the rate of glutamate uptakeshowed a trend toward decreasing from young to aged animals,the difference was not significant, F(2,31)=1.59, pN0.05).However, in the pre-motor cortex, aged animals exhibited asignificant 47% decrease in the rate of glutamate uptake com-pared to young animals (2.1±0.3 μM/s vs. 4.0±0.6 μM/s,n=11 and 12, respectively; F(2,31)=4.39, pb0.05).
The KCl evoked glutamate responses were typified by arapid increase in local glutamate concentration usually reachingthe peak amplitude in less than 10 s followed by a similar rapid
Table 2Potassium evoked glutamate release
Age Amplitude(μM)
Amp/volume(μM/nL)
T80(s)
Uptake rate(μM/s)
Pre-motor cortex a
Young 8.1±1.1 0.32±0.13 6.2±1.2 1.6±0.5Middle-aged 6.6±0.6 0.16±0.02 8.0±1.6 0.8±0.1Aged 8.7±0.7 0.26±0.05 5.6±0.6 1.1±0.2
Motor cortex b
Young b 6.3±0.7 0.12±0.04 5.9±0.6 1.0±0.3Middle-aged 7.1±0.9 0.23±0.08 7.0±1.0 0.9±0.3Aged 7.8±1.4 0.29±0.10 7.2±3.0 1.0±0.3
a Signal measurements: young 7; middle-aged 12; aged 9; from 4, 4, and 3animals, respectively.b Signal measurements: young 11; middle-aged 9; aged 6; from 4, 4, and 3
animals, respectively.

243J.E. Quintero et al. / Experimental Neurology 208 (2007) 238–246
decrease in the signal (Fig. 5). Direct delivery of KCl to the areaaround the electrode generated varying sized signal responses;however, for the purpose of these studies, we only compared thesignal parameters of responses that had amplitudes within therange of 3–15 μM (Table 2). However, these signals did notreveal significant differences in the parameters we tested.
Discussion
We successfully used a novel, glutamate-detecting micro-electrode to measure at 1 Hz both resting glutamate levels andglutamate neurotransmission in the cortex of anesthetized rhesusmonkeys. We found resting glutamate levels were significantlyelevated in the motor cortex of aged animals compared to youngones, while resting glutamate levels in the pre-motor cortexshowed a correlation to age, there was not a significant dif-ference among the age groups. In conjunction, we analyzed therapid kinetics of glutamate signals, and in the pre-motor cortexregion, the rate of glutamate uptake was slower in aged animalscompared to young animals. Other rapid measurements ofexogenous glutamate- or potassium-induced glutamate releaseand uptake revealed a glutamatergic neurotransmission systemwith robust, reproducible signals in the neocortex in each agegroup; however, we did not identify any additional significantdifferences among the age groups. Correlation analyses betweenage or resting glutamate levels and uptake rates or signal ampli-tudes were inconclusive.
While only a few studies have examined glutamate levels inthe brain of monkeys, the values we report are in agreement withconcentrations measured for resting glutamate levels in the brainof monkeys and other species (see Table 3). The values for restingglutamate levels appeared to cover a broad range, but almost all ofthe valueswere in the limits of previously reportedmeasurements.In particular, only one depth measurement from an aged animal(379P) showed a relatively high resting glutamate level (74.4μM)compared to published results; we included it in the resultsbecause we could not identify a criterion for excluding themeasurement. Animal 379P's surgery and recording wereunremarkable, and the responses to 70mMKCl, 1 mM glutamatestimulation, and other resting glutamate levels were comparableto recordings from other animals. In addition, to our knowledge,
Table 3Resting glutamate levels in the brain
Species Glutamateconcentrations a
Location
Rat 3.0 Striatum Miele et al. (1996)1.6 Striatum Day et al. (2006)7.3, 44.9 Striatum, prefrontal cx Rutherford et al.
(2007)Monkey ∼1–2 Prefrontal cx Kodama et al. (2002)
28.7 Striatum Galvan et al. (2003)3.7 to 20.3 Pre-motor, motor cx This study
Human 20.3 Hippocampus During and Spencer (1993)17 Frontal cortex Reinstrup et al. (2000)2.6 Neocortex Cavus et al. (2005)8.3, 9.6 Motor cortex Kaiser et al. (2005)
a Mean resting glutamate levels in μM.
this is the first report examining in vivo resting glutamate levels atmultiple depths and brain regions in a primate aging paradigm.This novelty coupled with the limits on access to primates made itnecessary to present all of the available valid results. Still, werealize future studies are necessary to address the origin and extentof glutamatergic reorganization in the aging primate brain.
From this study, it is unclear what the physiological effects ofelevated resting glutamate levels are. Resting glutamate levelsin the two brain regions we examined were of comparablemagnitude within each age group, and in some cases withineach animal. Although elevated resting glutamate levels of thismagnitude may be by themselves insufficient to induce neuro-toxic damage (Mangano and Schwarcz, 1983; Obrenovitchet al., 2000), over time, raised levels may increase a neuron'ssusceptibility to excitotoxic injury, and excitotoxicity is linkedwith a number of neurodegenerative diseases includingAlzheimer's disease, amyotrophic lateral sclerosis, and Hun-tington's disease. In particular, elevated levels of glutamatewere found in epileptogenic hippocampi prior to and followingseizure activity (During and Spencer, 1993). In addition to theincrease in resting glutamate levels with age, we also noted thatthe values of resting glutamate levels in the middle-aged andaged monkeys showed a wider distribution and overall vari-ability compared to the young monkeys. Age-related variabilityis a common feature of aging (Grondin et al., 2003; Fridrikssonet al., 2006), and further studies may help determine if restingglutamate levels might serve as markers of aging or neurode-generative damage in the primate nervous system.
One finding that may offer a possible explanation for amechanism responsible for the age-related increase in restingglutamate levels was the ∼50% decrease in the rate of gluta-mate uptake in the pre-motor cortex and the trend in reducedglutamate uptake in the motor cortex. The local, fast delivery ofexogenous glutamate to the brain allowed us to mimic endog-enous release events and measure how rapidly cells removeglutamate from the extracellular space. The important con-tributors toward maintaining a tight regulation of extracellularglutamate are the glial and neuronal glutamate transporters (Sealand Amara, 1999). Two glial transporters, the high-affinitysodium-dependent GLAST and GLT, account for the removalof a majority of the glutamate from the extracellular space(Rothstein et al., 1996; Tanaka et al., 1997), and among thesetransporters, GLAST shows a decreased cell surface expressionin aged rats compared to young rats (Nickell et al., 2007). If acomparable change in the surface expression of glial glutamatetransporters occurs in monkeys, this might account in part forthe elevated resting glutamate levels and/or the decreased rate ofglutamate uptake in the neocortex of aged monkeys. However,multiple components of the glutamatergic system may changewith age and these could differentially affect resting glutamatelevels and glutamate uptake in the pre-motor and motor cortices.
While we identified an age-related decrease in the rate ofglutamate uptake following the delivery of exogenous gluta-mate, we did not identify any significant differences followingpotassium stimulation. Had we examined the kinetics of higheramplitude signals, we might have detected significant differ-ences in glutamatergic dynamics (Nickell et al., 2005). However,

244 J.E. Quintero et al. / Experimental Neurology 208 (2007) 238–246
the amplitude range of glutamate signals we analyzed reflectedthe signal size that represented the majority of the elicitedendogenous glutamate signals following high potassiumstimulation. In addition, the use of anesthetized animalsmay have contributed to the blunting of any depolarization-dependent differences in glutamate release among the agegroups. Previous reports suggest that isoflurane may decreaseglutamate release while increasing glutamate uptake in mam-mals (Hudspith, 1997; Larsen et al., 1998; Zuo, 2001; Ritz et al.,2006). However, isoflurane is a commonly used anesthetic inclinical surgery so any potential attenuation of the responsesmay be affected comparably in patients in an operating room.
Although the technique of microdialysis is the standardmethod of measuring neurochemicals in the brain, in comparison,enzyme-basedmicroelectrodes provide useful advantages.Micro-dialysis probes create damage to the surrounding tissue as faraway as 1.4 mm from the implant site (Clapp-Lilly et al., 1999),alter neurotransmitter release and uptake mechanisms in thesurrounding tissue (Bungay et al., 2003; Borland et al., 2005), andmeasure glutamate that may not represent one that is the result ofneuronally derived synaptic release (Timmerman and Westerink,1997). By comparison, in chronically implanted ceramicmicroelectrodes, the tissue damage is confined to 50–100 μmfrom the microelectrode (Rutherford et al., 2007), and glutamatesignals measured with electrochemical microelectrodes are atleast, in part, sodium channel-dependent (Kulagina et al., 1999;Day et al., 2006). The limited disruption of tissue along with thesmaller size of these microelectrodes compared to microdialysisprobes enables us to place the microelectrode in greater proximityto synapses and more closely measure synaptic overflowcompared to microdialysis. Finally, although effective advanceshave increased the temporal resolution of microdialysis fromminutes to ∼60 s (Shou et al., 2006), microdialysis continues tohave an insufficient time resolution to measure fast neuronalsignaling events. However, the technology we describe here canresolve stimulus-evoked glutamate release on at least a second-by-second basis, creating an opportunity to examine in detail theactions of fast-acting neurotransmitters.
One other important aspect of this study was the successfulinitial characterization of resting glutamate levels and rapidglutamate signaling in non-human primates. In the future, neuro-chemical studies on non-human primates will grow in importancebecause as Bradberry (2000) suggests, with the increased study ofhuman brain function by noninvasive imaging methods, we willrelymore onmethods that can provide a correlative explanation ofthe changes in imaging signals based on quantitative chemicalanalysis. These future studies likely will use the more clinicallyrelevant awake non-human primate model since their brainstructure, size, and function more closely resemble that ofhumans. A further advantage of studying the unaesthetizedmonkey, aside from reducing the potential confound of anesthesiaon glutamate transmission, is that the awake model presents theopportunity to measure neurochemical changes in the brain whilethe animal is behaving.
Additionally, we conducted these studies under conditionssimilar to hospital surgical suites with the objective of laying afoundation for measuring rapid glutamate signals in the clinic.
We have designed and developed the equipment and proceduresnecessary to measure the glutamate in an operating room, andonly a few refinements may be necessary for similar measure-ments in humans. The glutamate measurements in this studywere confined to the more superficial areas of the brain, butcurrently, we are continuing to develop and fabricate micro-electrodes that will reach the deeper structures of the brain. Thetranslation of this technique from non-clinical research to theclinic offers the possibility for a better understanding of the roleglutamate and other neurotransmitters play in the process ofneurodegenerative diseases and disease states in humans.
In summary, we employed a minimally invasive technique inan operating room setting to measure consistently and reliablycortical glutamate levels and to examine age-related changes inthe glutamatergic system in a non-human primate model. Weshowed that this technique has the sensitivity to measure tonic (asreflected in the resting glutamate levels) as well as phasic(following KCl stimulation) glutamate release and uptake (fol-lowing exogenous glutamate application). This advancement willallow others to conduct detailed examinations of the neurochem-istry of primates in situ. Further studies will help elucidate therelationship between age-related elevations in resting glutamatelevels and both normal aging and age-related neurodegeneration.
Acknowledgments
Support providedbyUSPHSAG00242,NS39787,AG013494,DA017186; NSF DBI 0352848.
Disclosure statement: Greg A. Gerhardt, PhD is the principalowner of Quanteon LLC.
References
Alves, O.L., Bullock, R., Clausen, T., Reinert, M., Reeves, T.M., 2005.Concurrent monitoring of cerebral electrophysiology and metabolism aftertraumatic brain injury: an experimental and clinical study. J. Neurotrauma.22, 733–749.
Arundine, M., Tymianski, M., 2003. Molecular mechanisms of calcium-dependent neurodegeneration in excitotoxicity. Cell Calcium 34, 325–337.
Bai, L., Hof, P.R., Standaert, D.G., Xing, Y.D., Nelson, S.E., Young, A.B.,Magnusson, K.R., 2004. Changes in the expression of the NR2B subunitduring aging in macaque monkeys. Neurobiol. Aging 25, 201–208.
Beart, P.M., O'Shea, R.D., 2007. Transporters for L-glutamate: an update on theirmolecular pharmacology and pathological involvement. Br. J. Pharmacol. 150,5–17.
Berg, A., Bellander, B.M., Wanecek, M., Gamrin, L., Elving, A., Rooyackers,O., Ungerstedt, U., Wernerman, J., 2006. Intravenous glutamine supple-mentation to head trauma patients leaves cerebral glutamate concentrationunaffected. Intensive Care Med. 32, 1741–1746.
Borland, L.M., Shi, G., Yang, H., Michael, A.C., 2005. Voltammetric study ofextracellular dopamine near microdialysis probes acutely implanted in thestriatum of the anesthetized rat. J. Neurosci. Methods 146, 149–158.
Bradberry, C.W., 2000. Applications of microdialysis methodology in nonhumanprimates: practice and rationale. Crit. Rev. Neurobiol. 14, 143–163.
Brewer, G.J., 2000. Neuronal plasticity and stressor toxicity during aging. Exp.Gerontol. 35, 1165–1183.
Bungay, P.M., Newton-Vinson, P., Isele, W., Garris, P.A., Justice, J.B., 2003.Microdialysis of dopamine interpreted with quantitative model incorporat-ing probe implantation trauma. J. Neurochem. 86, 932–946.
Burmeister, J., Pomerleau, F., Palmer, M., Day, B., Huettl, P., Gerhardt, G.,2002. Improved ceramic-based multisite microelectrode for rapid measure-ments of L-glutamate in the CNS. J. Neurosci. Methods 119, 163.

245J.E. Quintero et al. / Experimental Neurology 208 (2007) 238–246
Burmeister, J.J., Gerhardt, G.A., 2001. Self referencing ceramic based multisitemicroelectrodes for the detection and elimination of interferences from themeasurement of L-glutamate and other analytes. Anal. Chem. 73, 1037–1042.
Burmeister, J.J., Moxon, K., Gerhardt, G.A., 2000. Ceramic-based multisitemicroelectrodes for electrochemical recordings. Anal. Chem. 72, 187–192.
Cavus, I., Kasoff, W.S., Cassaday, M.P., Jacob, R., Gueorguieva, R., Sherwin,R.S., Krystal, J.H., Spencer, D.D., Abi-Saab, W.M., 2005. Extracellularmetabolites in the cortex and hippocampus of epileptic patients. Ann.Neurol. 57, 226–235.
Clapp-Lilly, K.L., Roberts, R.C., Duffy, L.K., Irons, K.P., Hu, Y., Drew, K.L.,1999. An ultrastructural analysis of tissue surrounding a microdialysisprobe. J. Neurosci. Methods 90, 129–142.
Day, B.K., Pomerleau, F., Burmeister, J.J., Huettl, P., Gerhardt, G.A.,2006. Microelectrode array studies of basal and potassium-evoked release ofL-glutamate in the anesthetized rat brain. J. Neurochem. 96, 1626–1635.
DeRousseau, C.J., 1985. Aging in the musculoskeletal system of rhesus monkeys:II. Degenerative joint disease. Am. J. Phys. Anthropol. 67, 177–184.
During, M.J., Spencer, D.D., 1993. Extracellular hippocampal glutamate andspontaneous seizure in the conscious human brain. Lancet 341, 1607–1610.
Fridriksson, J., Morrow, K.L., Moser, D., Baylis, G.C., 2006. Age-relatedvariability in cortical activity during language processing. J. Speech Lang.Hear. Res. 49, 690–697.
Friedemann, M.N., Gerhardt, G.A., 1992. Regional effects of aging ondopaminergic function in the Fischer-344 rat. Neurobiol. Aging 13, 325–332.
Galvan, A., Smith, Y., Wichmann, T., 2003. Continuous monitoring ofintracerebral glutamate levels in awake monkeys using microdialysis andenzyme fluorometric detection. J. Neurosci. Methods 126, 175–185.
Gazzaley, A.H., Siegel, S.J., Kordower, J.H., Mufson, E.J., Morrison, J.H.,1996. Circuit-specific alterations of N-methyl-D-aspartate receptor subunit 1in the dentate gyrus of aged monkeys. Proc. Natl. Acad. Sci. U. S. A. 93,3121–3125.
Graham, W.C., Robertson, R.G., Sambrook, M.A., Crossman, A.R., 1989.Extracellular amino acid levels in the globus pallidus of the consciousprimate studied by intracerebral microdialysis. Br. J. Pharmacol. 98Suppl:819P., 819P.
Greenamyre, J.T., 2001. Glutamatergic influences on the basal ganglia. Clin.Neuropharmacol. 24, 65–70.
Greenamyre, J.T., Young, A.B., 1989. Excitatory amino acids and Alzheimer'sdisease. Neurobiol. Aging 10, 593–602.
Grondin, R., Cass, W.A., Zhang, Z., Stanford, J.A., Gash, D.M., Gerhardt, G.A.,2003. Glial cell line-derived neurotrophic factor increases stimulus-evokeddopamine release and motor speed in aged rhesus monkeys. J. Neurosci. 23,1974–1980.
Hof, P.R., Duan, H., Page, T.L., Einstein, M., Wicinski, B., He, Y., Erwin, J.M.,Morrison, J.H., 2002. Age-related changes in GluR2 and NMDAR1glutamate receptor subunit protein immunoreactivity in corticocorticallyprojecting neurons in macaque and patas monkeys. Brain Res. 928,175–186.
Hudspith, M.J., 1997. Glutamate: a role in normal brain function, anaesthesia,analgesia and CNS injury. Br. J. Anaesth. 78, 731–747.
Kaiser, L.G., Schuff, N., Cashdollar, N., Weiner, M.W., 2005. Age-relatedglutamate and glutamine concentration changes in normal human brain: 1HMR spectroscopy study at 4 T. Neurobiol. Aging 26, 665–672.
Kling, A.S., Tachiki, K., Lloyd, R., 1993. Neurochemical correlates of theKluver–Bucy syndrome by in vivo microdialysis in monkey. Behav. BrainRes. 56, 161–170.
Kodama, T., Hikosaka, K., Watanabe, M., 2002. Differential changes inglutamate concentration in the primate prefrontal cortex during spatialdelayed alternation and sensory-guided tasks. Exp. Brain Res. 145, 133–141.
Kulagina, N.V., Shankar, L., Michael, A.C., 1999. Monitoring glutamate andascorbate in the extracellular space of brain tissue with electrochemicalmicrosensors. Anal. Chem. 71, 5093–5100.
Kuninobu, N., Ann, C.M., Neil, W.K., 1993. Patterns of neuronal degenerationin the motor cortex of amyotrophic lateral sclerosis patients. ActaNeuropathol. 86, 55–64.
Larsen, M., Valo, E.T., Berg-Johnsen, J., Langmoen, I.A., 1998. Isofluranereduces synaptic glutamate release without changing cytosolic free calciumin isolated nerve terminals. Eur. J. Anaesthesiol. 15, 224–229.
Lefaucheur, J.P., 2005. Motor cortex dysfunction revealed by corticalexcitability studies in Parkinson's disease: influence of antiparkinsoniantreatment and cortical stimulation. Clin. Neurophysiol. 116,244–253.
Liu, Z., Stafstrom, C.E., Sarkisian, M., Tandon, P., Yang, Y., Hori, A., Holmes,G.L., 1996. Age-dependent effects of glutamate toxicity in the hippocampus.Brain Res. Dev. Brain Res. 97, 178–184.
Magnusson, K.R., Cotman, C.W., 1993a. Age-related-changes in excitatoryamino-acid receptors in 2 mouse strains. Neurobiol. Aging 14,197–206.
Magnusson, K.R., Cotman, C.W., 1993b. Effects of aging on Nmda and Mk801binding-sites in mice. Brain Res. 604, 334–337.
Mangano, R.M., Schwarcz, R., 1983. Chronic infusion of endogenousexcitatory amino acids into rat striatum and hippocampus. Brain Res.Bull. 10, 47–51.
Mendelowitsch, A., Sekhar, L.N., Wright, D.C., Nadel, A., Miyashita, H.,Richardson, R., Kent, M., Shuaib, A., 1998. An increase in extracellularglutamate is a sensitive method of detecting ischaemic neuronal damageduring cranial base and cerebrovascular surgery. An in vivo microdialysisstudy. Acta Neurochir. 140, 349–356.
Miele, M., Berners, M., Boutelle, M.G., Kusakabe, H., Fillenz, M., 1996. Thedetermination of the extracellular concentration of brain glutamate usingquantitative microdialysis. Brain Res. 707, 131–133.
Morrison, J.H., 2003. Aging and mammalian cerebral cortex: monkeys tohumans. Alzheimer Dis. Assoc. Disord. 17 (Suppl 2:S51-3), S51–S53.
Nickell, J., Pomerleau, F., Allen, J., Gerhardt, G.A., 2005. Age-related changesin the dynamics of potassium-evoked L-glutamate release in the striatum ofFischer 344 rats. J. Neural Transm. 112, 87–96.
Nickell, J., Salvatore, M., Pomerleau, F., Apparsundaram, S., Gerhardt, G.A.,2007. Reduced plasma membrane surface expression of GLAST mediatesdecreased glutamate regulation in the aged striatum. Neurobiol. Aging,doi:10.1016/j.neurobiolaging.2006.07.015.
Obrenovitch, T.P., Urenjak, J., Zilkha, E., Jay, T.M., 2000. Excitotoxicity inneurological disorders — the glutamate paradox. Int. J. Dev. Neurosci. 18,281–287.
Reinstrup, P., Stahl, N., Mellergard, P., Uski, T., Ungerstedt, U., Nordstrom, C.H.,2000. Intracerebral microdialysis in clinical practice: baseline values forchemical markers during wakefulness, anesthesia, and neurosurgery. Neuro-surgery 47, 701–709.
Ritz, M.F., Schmidt, P., Mendelowitsch, A., 2006. Effects of isoflurane onglutamate and taurine releases, brain swelling and injury during transientischemia and reperfusion. Int. J. Neurosci. 116, 191–202.
Rothstein, J.D., Dykes-Hoberg, M., Pardo, C.A., Bristol, L.A., Jin, L., Kuncl,R.W., Kanai, Y., Hediger, M.A., Wang, Y., Schielke, J.P., Welty, D.F., 1996.Knockout of glutamate transporters reveals a major role for astroglialtransport in excitotoxicity and clearance of glutamate. Neuron 16,675–686.
Rutherford, E.C., Pomerleau, F., Huettl, P., Stromberg, I., Gerhardt, G.A.,2007. Chronic second-by-second measures of L-glutamate in the centralnervous system of freely moving rats. J. Neurochem. 102, 712–722.
Sabatini, U., Boulanouar, K., Fabre, N., Martin, F., Carel, C., Colonnese, C.,Bozzao, L., Berry, I., Montastruc, J.L., Chollet, F., Rascol, O., 2000. Corticalmotor reorganization in akinetic patients with Parkinson's disease: afunctional MRI study. Brain 123, 394–403.
Seal, R.P., Amara, S.G., 1999. Excitatory amino acid transporters: a family influx. Annu. Rev. Pharmacol. Toxicol. 39, 431–456.
Segovia, G., Porras, A., Del Arco, A., Mora, F., 2001. Glutamatergicneurotransmission in aging: a critical perspective. Mech. Ageing Dev.122, 1–29.
Shou, M., Ferrario, C.R., Schultz, K.N., Robinson, T.E., Kennedy, R.T., 2006.Monitoring dopamine in vivo by microdialysis sampling and on-lineCE-laser-induced fluorescence. Anal. Chem. 78, 6717–6725.
Strolin, B.M., Cini, M., Fusi, R., Marrari, P., Dostert, P., 1990. The effects ofaging on MAO activity and amino acid levels in rat brain. J. Neural Transm.,Suppl. 29, 259–268.
Tamaru, M., Yoneda, Y., Ogita, K., Shimizu, J., Nagata, Y., 1991. Age-relateddecreases of the N-methyl-D-aspartate receptor complex in the rat cerebral-cortex and hippocampus. Brain Res. 542, 83–90.

246 J.E. Quintero et al. / Experimental Neurology 208 (2007) 238–246
Tanaka, K., Watase, K., Manabe, T., Yamada, K., Watanabe, M., Takahashi, K.,Iwama, H., Nishikawa, T., Ichihara, N., Kikuchi, T., Okuyama, S.,Kawashima, N., Hori, S., Takimoto, M., Wada, K., 1997. Epilepsy andexacerbation of brain injury in mice lacking the glutamate transporter GLT-1. Science 276, 1699–1702.
Timmerman, W., Westerink, B.H., 1997. Brain microdialysis of GABA andglutamate: what does it signify? Synapse 27, 242–261.
Wenk, G.L., Pierce, D.J., Struble, R.G., Price, D.L., Cork, L.C., 1989. Age-related changes in multiple neurotransmitter systems in the monkey brain.Neurobiol. Aging 10, 11–19.
Wichmann, T., DeLong, M.R., 1996. Functional and pathophysiological modelsof the basal ganglia. Curr. Opin. Neurobiol. 6, 751–758.
Zuo, Z., 2001. Isoflurane enhances glutamate uptake via glutamate transportersin rat glial cells. Neuroreport 12, 1077–1080.





























