Embed Size (px)
Citation preview

An
Fa
b
a
ARRAA
KMBOZA
1
smch2c(2
e
jP
h0
Aquatic Toxicology 166 (2015) 21–28
Contents lists available at ScienceDirect
Aquatic Toxicology
j ourna l ho me pa ge: www.elsev ier .com/ locate /aquatox
n in vivo evaluation of acute toxicity of cobalt ferrite (CoFe2O4)anoparticles in larval-embryo Zebrafish (Danio rerio)
arooq Ahmad a, Xiaoyi Liu a, Ying Zhou a,b,∗, Hongzhou Yao a
College of Chemical Engineering, Zhejiang University of Technology, Hangzhou, ChinaResearch Center of Analysis and Measurement, Zhejiang University of Technology, Hangzhou, China
r t i c l e i n f o
rticle history:eceived 12 April 2015eceived in revised form 3 July 2015ccepted 6 July 2015vailable online 10 July 2015
eywords:agnetic nanoparticles
iochemical changexidative stressebrafish embryopoptosis
a b s t r a c t
The broad spectrum applications of CoFe2O4 NPs have attracted much interest in medicine, environ-ment and industry, resulting in exceedingly higher exposures to humans and environmental systemsin succeeding days. Their health effects and potential biological impacts need to be determined for riskassessment. Zebrafish (Danio rerio) embryos were exposed to environmentally relevant doses of nano-CoFe2O4 (mean diameter of 40 nm) with a concentration range of 10–500 �M for 96 h. Acute toxic endpoints were evaluated by survival rate, malformation, hatching delay, heart dysfunction and tail flexureof larvae. Dose and time dependent developmental toxicity with severe cardiac edema, down regulationof metabolism, hatching delay and tail/spinal cord flexure and apoptosis was observed. The biochemicalchanges were evaluated by ROS, Catalase (CAT), Lipid peroxidation (LPO), Acid phophatase (AP) and Glu-tatione s- transferase (GST). An Agglomeration of NPs and dissolution of ions induces severe mechanicaldamage to membranes and oxidative stress. Severe apoptosis of cells in the head, heart and tail regionwith inhibition of catalase confirms ROS induced acute toxicity with increasing concentration. Increasedactivity of GST and AP at lower concentrations of CoFe2O4 NPs demonstrates the severe oxidative stress.
Circular dichroism (CD) spectra indicated the weak interactions of NPs with BSA and slight changesin �-helix structure. In addition, CoFe2O4 NPs at lower concentrations do not show any considerableinterference with assay components and analytical instruments. The results are possible elucidation ofpathways of toxicity induced by these particles, as well as contributing in defining the protocols for riskassessment of these nanoparticles.© 2015 Elsevier B.V. All rights reserved.
. Introduction
Cobalt ferrite nanoparticles (NPs) are one of the most exten-ively used magnetic NPs in medicine. Cobalt ferrite (CoFe2O4)agnetic NPs have far reaching applications in imaging (MRI),
ancer treatment (Sincai et al., 2001), ferrofluids, drug delivery,yperthermia (Guglielmo et al., 2010), diagnostics (Hong et al.,013) and electronic devices because of their high permeability,oercivity, moderate magnetization, high saturation magnetizationSanpo et al., 2013) and physiochemical stability (Guglielmo et al.,
010).Routine use of CoFe2O4 NPs results in vast disposal into thenvironment, raising an alarming situation (Novak et al., 2013)
∗ Corresponding author at: Research Center of Analysis and Measurement, Zhe-iang University of Technology, 18 Chaowang Road, Hangzhou 310032, Zhejiangrovince, China.
E-mail address: [email protected] (Y. Zhou).
ttp://dx.doi.org/10.1016/j.aquatox.2015.07.003166-445X/© 2015 Elsevier B.V. All rights reserved.
for the global ecosystem. These released CoFe2O4 NPs can undergobiodegradation, resulting in possible release of Co2+ and Fe3+ ionsinto the environment. Co2+ and Fe3+ ions can induce ROS mediatedoxidative stress (Novak et al., 2013; Sun et al., 2013), cytotoxicity,genotoxicity, inflammations and oxidation of proteins and mem-brane disruptions (Papis et al., 2007; Ponti et al., 2009; Azaria et al.,2011; Luo et al., 2014). Therefore, there is pressing need to study thepossible toxic effects of CoFe2O4 NPs and subsequent dissolution ofions from these NPs into the environment.
Cobalt ferrite NPs induces toxic responses (cyto & genotoxicity)in organisms on different trophic levels from producers (Photosyn-thetic algae) to consumers (humans and pigs, etc.) (Barhoumi andDewez, 2013; Azaria et al., 2013; Kapilevich et al., 2010). Cobaltferrite (CoFe2O4) NPs causes cleavage furrow regression and pro-motes cancerous cells by paralyzing the immune system (Colognato
et al., 2007). CoFe2O4 NPs induce respiratory disruptions in guineapigs (Kapilevich et al., 2010) and human lungs as well. CoFe2O4NPs promote ROS dependent oxidative stress in liver (Human &mice), kidney, intestine, lymphoblasts cell lines and primary mouse
2 Toxico
dNfis(a
nmtsepeiebTiapm(hlrcFu2
ci(cdwf
2
2
(Aw
2
iFb
2
sbSCenaw
2 F. Ahmad et al. / Aquatic
endritic-cells (Azaria et al., 2013; Hwang et al., 2012). CoFe2O4Ps also proved lethal to artificial and biological membranes. Thesendings are substantial with regard to environmental health andafety issues concerned with CoFe2O4 NPs application in medicineDrasler et al., 2014). These NPs also interfere with lipid metabolismnd embryogenesis (Guglielmo et al., 2010; Marmorato et al., 2011).
Physicochemical properties like colloidal stability, purity, inert-ess, size, shape and charge etc. also determine the fate and toxicityechanisms (Luyts et al., 2013) of NPs. The scientific world has
he dilemma of lacking prior characterizing of the NPs for toxicitytudy, which not only reduces the significance of these studies orven totally senseless (Krug, 2014). This necessitates the study ofrinciple interactions in the light of their physicochemical param-ters at biological interfaces (Chen et al., 2013). Nanomaterials alsonterfere with analytical techniques due to their inherent prop-rty of fluorescence/absorbance, not only with assay components,ut also with analytes (Ong et al., 2014; Maccormack et al., 2012).here is a pressing need for prior testing of their physicochem-
cal properties, possible interference with analytical techniquesnd assays. Zebrafish as an alternative to animal experiment (3Rsrinciple; Reduction, Replacement and Refinement) have receiveduch attention in acute fish toxicity test with ISO standard 15088
Braunbeck et al., 2005; Lammer et al., 2009; ISO, 2007). Zebrafishave long been used as a model vertebrate research animal in the
ife sciences because of similarities with the human genome, theirapid well characterize development stages and transparent lowost embryos with high fecundity (Kane et al., 1996; Fako andurgeson, 2009; Hill et al., 2005). Recently, Zebrafish have beensed in toxicity and risk assessment of nanomaterials (Wang et al.,010).
The aim of this study was (i) to gain insights into the biochemi-al mechanisms underlying the acute toxic effects of nano-CoFe2O4n relation to their physicochemical properties utilizing the 3Rsreduction, refinement and replacement) alternative animal prin-iple, (ii) role of ROS in acute nanotoxicity and malformation ofeveloping Zebrafish, (iii) possible interference of CoFe2O4 NPsith the assay components and procedure of removing such inter-
erences.
. Material and method
.1. Chemicals
Powdered CoFe2O4 NPs were purchased from RegentaHangzhou, China). All other chemicals were purchased fromladdin (Shanghai, China). All chemicals used in the present studyere at least of analytical grade or relevant biochemical grade.
.2. Physicochemical characterization of nanoparticles
Purity of providing NPs was measured by ICPMS. Physicochem-cal properties, surface coating by FTIR, morphology and size byESEM, crystalline shape and size by XRD and magnetic propertiesy VSM was determined (Supplementary material S-1).
.3. Physicochemical properties in Medium
Dynamic light scattering measurements of the hydrodynamicize and zeta potential for CoFe2O4 nanoparticles were determinedy Beckman Coulter Delsa (TM) Nano C (Nano Zeta Potential andubmicron Particle Size Analyzer). The amount and rate of leachedo2+ and Fe3+ ions from the CoFe2O4 NPs in E3 medium was
stimated by ICPMS measurement over a period of 96 h. The super-atant was taken after 4, 12, 24, 48, 72 and 96 h, twice centrifugedt 10,000 rpm for 20 min. After centrifugation, the supernatantas acid digested followed by ICPMS (ICP-MS; Elan DRC-e; Perkinlogy 166 (2015) 21–28
Elmer, USA) measurement. The rate of agglomeration and sedimen-tation of different concentrations of cobalt ferrite (CoFe2O4) NPsin E3 medium with the passage of time until 96 h was evaluatedby double beam UV/Vis spectrophotometer (JASCO-V550, Japan) at540 nm wavelength.
2.4. Fish husbandry, embryo collection and exposure
Wild-type Zebrafish (AB) were raised according to standardbreeding protocols (28 ± 0.5 ◦C with 14:10 day/night photoperiod)in a recirculation system in an Environmental science facility ofZhejiang University of Technology. Reverse osmosis (pH 6.5–7.5)filtered water was supplied to the recirculation system with con-ductivity of 450–1000 s cm−1. Zebrafish were fed twice daily withlive Artemia (Jiahong Feed Co., Tianjin, China) and a dry flake diet(Zeigler, Aquatic Habitats, Apopka Florida, USA). The developmentstatus of Zebrafish embryos and larvae were observed with anInverted Microscope (Nikon, Japan). Zebrafish embryos used forchemical exposure were obtained from spawning adults in tanksovernight with the sex ratio of 1:1. Embryos were collected within1 h after the light switched on and rinsed in E3 medium. The fer-tilized and normal embryos were inspected and staged for thefollowing experiment under a stereomicroscope (Nikon, Japan)[Westerfield, 1995]. The Zebrafish embryos were exposed to differ-ent concentrations (0, 10, 62.5,125, 250, 500 �M) of CoFe2O4 NPsfor a period of 96 h. This range is selected from the LC50 value deter-mined by range finding test of CoFe2O4 NPs toxicity according tothe guidelines of OECD (2012).
2.5. Mortality, hatching, heart rate and malformation
Mortality and malformations as toxicity endpoint were analyzedin embryo/larvae daily (Choi et al., 2013). The E3 solution (50 mL)was replaced every 24 h until 96 hpf. The mortality of embryos orlarval fish was determined by examining their movement, heart-beat and blood circulation via inverted microscope (Nikon, Japan),and the results were recorded daily. The heartbeat of Zebrafishembryos was recorded in 48-96 hpf, Heart rates of 10 anesthetizedembryos were directly observed for 30 s, and the rate per minutededuced from this. Embryos were kept at room temperature for30 min to allow the heartbeat to assume a steady rate prior to man-ual counting. Counts were taken at 24, 48, and 72 hpf. At 72 and96 hpf, hatching and the presence of physical abnormalities wererecorded, respectively. Each experiment was performed indepen-dently in triplicate.
2.6. Cell assault measurement
To measure the cell assault caused by CoFe2O4 NPs is eitherapoptosis or necrosis, the embryos were stained by a mixtureof fluorescent dyes i.e., Acridine orange (AO) and ethidium bro-mide (EtBr). AO and EtBr are differentially uptake by live (intactand destabilized membrane) and dead (destabilized membrane)cells, indicating apoptosis and necrosis respectively. AO permeateinto the membranes (normal/apoptotic) producing green fluores-cence by intercalating with DNA while EtBr only permeate into theruptured membranes and produce orange fluorescence by inter-calating with DNA (McGahon et al., 1995). Embryos were exposedto CoFe2O4 NPs (10, 62.5, 125, 250 and 500 �M) for 96 hpf. Thenembryos were rinsed three times with PBS and incubated in a mix-ture of AO and EtBr (5 mg/mL, 1:1) for 30 min in the dark at 28 ◦C,
followed by three times rinses in PBS. Stained embryos were exam-ined using fluorescence microscopy (Nikon, Tokyo, Japan). WholeBody fluorescence was quantified using ImageJ (V. 1.49, NIH, USA)software.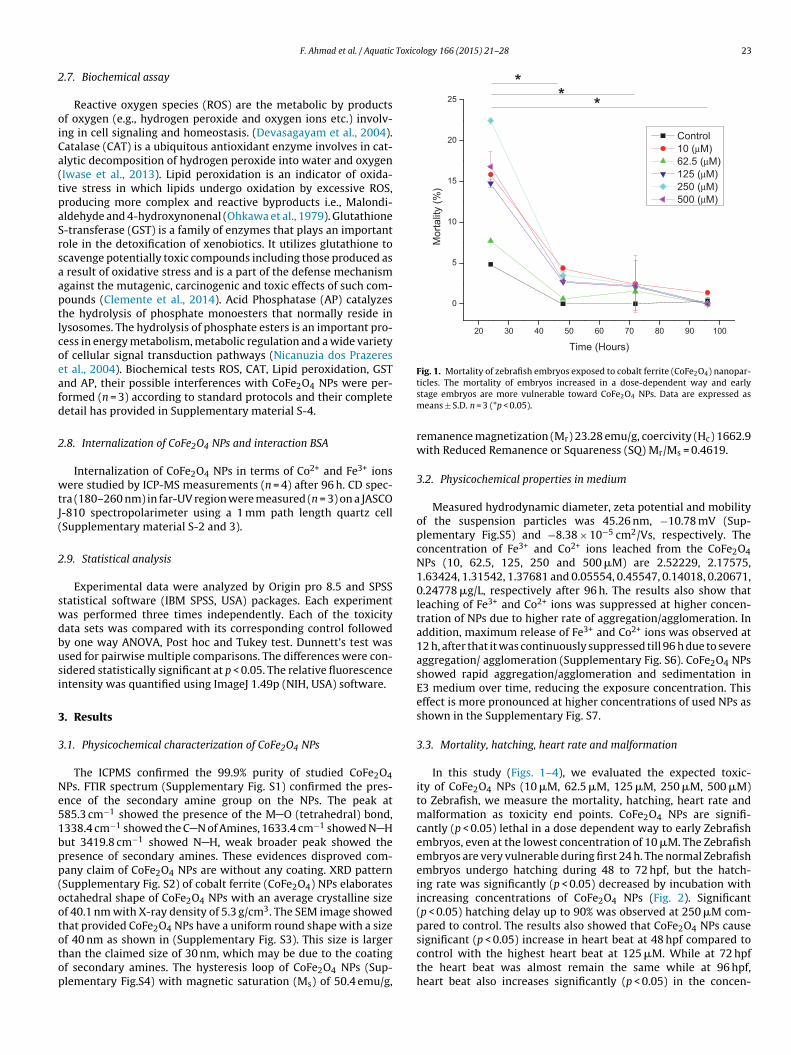
Toxicology 166 (2015) 21–28 23
2
oiCa(tpaSrsaaptlcoeafd
2
wtJ(
2
swdbusi
3
3
Ne51bpp(oototop
20 30 40 50 60 70 80 90 100
0
5
10
15
20
25
Mor
talit
y (%
)
Time (Hours)
Control 10 (µM) 62.5 (µM) 125 (µM) 250 (µM) 500 (µM)
** *
Fig. 1. Mortality of zebrafish embryos exposed to cobalt ferrite (CoFe2O4) nanopar-
F. Ahmad et al. / Aquatic
.7. Biochemical assay
Reactive oxygen species (ROS) are the metabolic by productsf oxygen (e.g., hydrogen peroxide and oxygen ions etc.) involv-
ng in cell signaling and homeostasis. (Devasagayam et al., 2004).atalase (CAT) is a ubiquitous antioxidant enzyme involves in cat-lytic decomposition of hydrogen peroxide into water and oxygenIwase et al., 2013). Lipid peroxidation is an indicator of oxida-ive stress in which lipids undergo oxidation by excessive ROS,roducing more complex and reactive byproducts i.e., Malondi-ldehyde and 4-hydroxynonenal (Ohkawa et al., 1979). Glutathione-transferase (GST) is a family of enzymes that plays an importantole in the detoxification of xenobiotics. It utilizes glutathione tocavenge potentially toxic compounds including those produced as
result of oxidative stress and is a part of the defense mechanismgainst the mutagenic, carcinogenic and toxic effects of such com-ounds (Clemente et al., 2014). Acid Phosphatase (AP) catalyzeshe hydrolysis of phosphate monoesters that normally reside inysosomes. The hydrolysis of phosphate esters is an important pro-ess in energy metabolism, metabolic regulation and a wide varietyf cellular signal transduction pathways (Nicanuzia dos Prazerest al., 2004). Biochemical tests ROS, CAT, Lipid peroxidation, GSTnd AP, their possible interferences with CoFe2O4 NPs were per-ormed (n = 3) according to standard protocols and their completeetail has provided in Supplementary material S-4.
.8. Internalization of CoFe2O4 NPs and interaction BSA
Internalization of CoFe2O4 NPs in terms of Co2+ and Fe3+ ionsere studied by ICP-MS measurements (n = 4) after 96 h. CD spec-
ra (180–260 nm) in far-UV region were measured (n = 3) on a JASCO-810 spectropolarimeter using a 1 mm path length quartz cellSupplementary material S-2 and 3).
.9. Statistical analysis
Experimental data were analyzed by Origin pro 8.5 and SPSStatistical software (IBM SPSS, USA) packages. Each experimentas performed three times independently. Each of the toxicity
ata sets was compared with its corresponding control followedy one way ANOVA, Post hoc and Tukey test. Dunnett’s test wassed for pairwise multiple comparisons. The differences were con-idered statistically significant at p < 0.05. The relative fluorescencentensity was quantified using ImageJ 1.49p (NIH, USA) software.
. Results
.1. Physicochemical characterization of CoFe2O4 NPs
The ICPMS confirmed the 99.9% purity of studied CoFe2O4Ps. FTIR spectrum (Supplementary Fig. S1) confirmed the pres-nce of the secondary amine group on the NPs. The peak at85.3 cm−1 showed the presence of the M O (tetrahedral) bond,338.4 cm−1 showed the C N of Amines, 1633.4 cm−1 showed N Hut 3419.8 cm−1 showed N H, weak broader peak showed theresence of secondary amines. These evidences disproved com-any claim of CoFe2O4 NPs are without any coating. XRD patternSupplementary Fig. S2) of cobalt ferrite (CoFe2O4) NPs elaboratesctahedral shape of CoFe2O4 NPs with an average crystalline sizef 40.1 nm with X-ray density of 5.3 g/cm3. The SEM image showedhat provided CoFe2O4 NPs have a uniform round shape with a size
f 40 nm as shown in (Supplementary Fig. S3). This size is largerhan the claimed size of 30 nm, which may be due to the coatingf secondary amines. The hysteresis loop of CoFe2O4 NPs (Sup-lementary Fig.S4) with magnetic saturation (Ms) of 50.4 emu/g,ticles. The mortality of embryos increased in a dose-dependent way and earlystage embryos are more vulnerable toward CoFe2O4 NPs. Data are expressed asmeans ± S.D. n = 3 (*p < 0.05).
remanence magnetization (Mr) 23.28 emu/g, coercivity (Hc) 1662.9with Reduced Remanence or Squareness (SQ) Mr/Ms = 0.4619.
3.2. Physicochemical properties in medium
Measured hydrodynamic diameter, zeta potential and mobilityof the suspension particles was 45.26 nm, −10.78 mV (Sup-plementary Fig.S5) and −8.38 × 10−5 cm2/Vs, respectively. Theconcentration of Fe3+ and Co2+ ions leached from the CoFe2O4NPs (10, 62.5, 125, 250 and 500 �M) are 2.52229, 2.17575,1.63424, 1.31542, 1.37681 and 0.05554, 0.45547, 0.14018, 0.20671,0.24778 �g/L, respectively after 96 h. The results also show thatleaching of Fe3+ and Co2+ ions was suppressed at higher concen-tration of NPs due to higher rate of aggregation/agglomeration. Inaddition, maximum release of Fe3+ and Co2+ ions was observed at12 h, after that it was continuously suppressed till 96 h due to severeaggregation/ agglomeration (Supplementary Fig. S6). CoFe2O4 NPsshowed rapid aggregation/agglomeration and sedimentation inE3 medium over time, reducing the exposure concentration. Thiseffect is more pronounced at higher concentrations of used NPs asshown in the Supplementary Fig. S7.
3.3. Mortality, hatching, heart rate and malformation
In this study (Figs. 1–4), we evaluated the expected toxic-ity of CoFe2O4 NPs (10 �M, 62.5 �M, 125 �M, 250 �M, 500 �M)to Zebrafish, we measure the mortality, hatching, heart rate andmalformation as toxicity end points. CoFe2O4 NPs are signifi-cantly (p < 0.05) lethal in a dose dependent way to early Zebrafishembryos, even at the lowest concentration of 10 �M. The Zebrafishembryos are very vulnerable during first 24 h. The normal Zebrafishembryos undergo hatching during 48 to 72 hpf, but the hatch-ing rate was significantly (p < 0.05) decreased by incubation withincreasing concentrations of CoFe2O4 NPs (Fig. 2). Significant(p < 0.05) hatching delay up to 90% was observed at 250 �M com-pared to control. The results also showed that CoFe2O4 NPs cause
significant (p < 0.05) increase in heart beat at 48 hpf compared tocontrol with the highest heart beat at 125 �M. While at 72 hpfthe heart beat was almost remain the same while at 96 hpf,heart beat also increases significantly (p < 0.05) in the concen-
24 F. Ahmad et al. / Aquatic Toxicology 166 (2015) 21–28
contro l 10 62. 5 12 5 250 50 010
20
30
40
50
60
70
80
90 *****
Hat
chin
g (%
) afte
r 72
hpf
*
Concent ratio n (µM)
Fig. 2. Box plots showing the Hatching delay (%) of zebrafish embryos inducedby Cobalt ferrite (CoFe2O4) nanoparticles for 72 hpf. The results showed a stronginhibition of hatching rate after embryos exposed to Cobalt ferrite (CoFe2O4)nanoparticles. Data are expressed as means ± S.D. n = 3 (*p < 0.05).
48 hours 72 hours 96 hours0
2
4
6
8
10
12
14
16
18
20 *
Hea
rt B
eat p
er 6
0 S
econ
d
Time (hpf)
con tro l 10 (µM) 62 .5 (µM) 125 (µM) 250 (µM) 500 (µM)
**
Fig. 3. Heart Rate of Zebrafish embryo/larvae after incubation with CoFe2O4
nanoparticles from 48–96 hpf time period. The same color of lines with Asterisk (*)shows statistical difference. Data are expressed as means ± S.D. n = 10, (*p < 0.05).
Fig. 4. Optical micrographs (A–X) showing physical abnormalities in comparison with control after incubation with CoFe2O4 NPs for 96 hpf. The malformations wereobserved for 24 hpf (A–D), 48 hpf (E–I), 72 hpf (J–N) and 96 hpf (O–X). (A, E, J and O) represent the normal development of Zebrafish embryos. Multiple types of abnormalitiesoccur in the same Zebrafish: arrested development (B–D), Absence of head and late eyes spot development, finfold abnormality and tail/spinal cord flexure (F–I), cardiacmalformation/edema, yolk sac edema, finfold abnormality, and tail developmental malformation with no tail extension, spinal cord abnormality, pericardial edema/cardiacmalformation, Yolk sac edema (K–N), (c) cardiac malformation/edema, yolk sac edema, finfold abnormality, and tail/spinal cord flexure, and head deformity, and tail/spinalcord flexure, melanocyte delayed development with blood clot (P–X). The malformation of the bending tails was a permanent damage for juvenile Zebrafish.

F. Ahmad et al. / Aquatic Toxicology 166 (2015) 21–28 25
Fig. 5. Effects of CoFe2O4 NPs on apoptosis in embryos after 96 hpf (A–C). (A) Characteristic fluorescence images depicting different regions of apoptosis (head, heart & tail)indicated by red arrows. (B–C) Semiquantitative analysis of apoptosis in the whole body and different regions at 96 hpf. The relative fluorescence intensity was adjusted bysubtracting the auto-fluorescence of the embryos and different regions of the embryo. Each experiment was conducted twice with three replicates and three fish per replicate* er is r
tNdmdamdsvae
3
aNNi
p < 0.05. (For interpretation of the references to color in this figure legend, the read
ration dependent way (Fig. 3). Results manifested that CoFe2O4Ps induce developmental toxicity in the dose and time depen-ent way. Zebrafish embryos also showed the different kinds ofalformations, which also becomes intense in dose and time
ependent way. The most common malformations include therrested development (absence of the head and eyes spot develop-ent), tail/spinal cord flexure, cardiac/yolk sac edema, melanocyte
elayed development with blood clot (Fig 4). The Zebrafish alsohowed reduced movement and sessile at the bottom with reducedisibility as the CoFe2O4 NPs attached to the eyes and body. Thettached NPs induced the membrane disruptions in the Zebrafishmbryos.
.4. Cell assault measurement
As from the Fig. 5(A), absence of orange fluorescence upon inter-
ction with a mixture of AO and EtBr comfirmed the CoFe2O4Ps induce cell death by the process of apoptosis. CoFe2O4Ps noticeably cause cell death in tail, heart and head regionn a dose dependent way at 96hpf. The maximum cell assault
eferred to the web version of this article.)
was observed in order tail > heart ≥ head is significantly (p < 0.05)higher than the control. The results ascribed adsorption ofaggregates/agglomerates induce the cell death. The whole body flu-orescence also showed a significant (p < 0.05) spike with increasingconcentration.
3.5. Biochemical assay
The results showed, at lower concentration, CoFe2O4 NPs didnot showed interference with GST, CAT and DCF-DA assay at themeasured wavelengths (results not shown). Additional washingsteps and ultracentrifugation proved very effective in minimizinginterferences from NPs. The results (Fig. 6) showed that AP activitysignificantly (p < 0.05) increased at 10 �M and then continuouslydecrease up to 250 �M concentration of CoFe2O4 NPs. But atthe concentration of 500 �M showed the significant (p < 0.05)
increase in the AP activity compared to control and other groups.The GST showed the highest scavenging activity at two lowestconcentrations (10 and 62.5 �M) of CoFe2O4 NPs, then showedthe significant (p < 0.05) lost in the radical scavenging activity with
26 F. Ahmad et al. / Aquatic Toxicology 166 (2015) 21–28
Fig. 6. Expression of biochemical parameters (mean ± S.D) A–E, (A) Acid Phosphatase, (B) Glutathion-S-transferase, (C) Malondialdehyde, (D) Catalase activity and (E), ReactiveO M, 12.s
itoio
xygen Species in Zebrafish (Danio rario) embryo 96hpf exposure to 10 �M, 62.5 �ignificantly different (p < 0.05; ANOVA, Tukey).
ncreasing concentrations. The MDA content directly manifests
he lipid peroxidation and membrane disruptions in response toxidative stress. Our results shed lights on the significant (p < 0.05)ncrease in the MDA content after incubation with CoFe2O4 NPsver a period of 96 hpf elaborating the severe membrane damage.
5 �M, 125 �M, 250 �M and 500 �M CoFe2O4 NPs. Bars with different letters show
The catalase activity was also increased significantly (p < 0.05)
in a dose dependent way. The ROS production also showed theelevated increase with increasing concentration of CoFe2O4 NPsup to 250 �M, then significant (p < 0.05) decrease in the ROS at500 �M.
F. Ahmad et al. / Aquatic Toxico
0
2
4
6
8
10
12
60
80
100
120 *
***
Rel
ativ
e am
ount
of I
nter
naliz
ed io
ns (µ
g/L)
Concentra tion (µM)
Fe Co
10 62.5 125 250 500
*
Ft(
3
oitswCp
4
ttMabfaiosimtbetste
neOcea�A
ig. 7. Intracelullar Concentrations of Fe3+ and Co2+of Zebrafish larvae exposedo CoFe2O4 NPs. Bars with an asterisk (*) are statistically significantly differentp < 0.05).
.6. Internalization of CoFe2O4 NPs and interaction with BSA
Internalization of Fe3+ and Co2+ ions are shown in Fig. 7, amountf internalized Fe3+ ion was significantly (p < 0.05) higher than Co2+
ons. The CD spectrum showed no absorption due to CoFe2O4 NPs inhis range of wavelength. CD spectra of BSA and BSA–CoFe2O4 NPshow decrease in the �-helical character of BSA upon interactionith CoFe2O4 NPs (Supplementary Fig. S8). Furthermore, shape of
D spectra of BSA with and without CoFe2O4 NPs demonstrates theredominant secondary structure.
. Discussion
Factors such as size, shape, coating material, agglomera-ion/aggregation, zeta potential, solubility and impurities of NPs arehe main contributing factors in nanotoxicology (Choi et al., 2013;
aurer-Jones et al., 2013). Drasler et al., 2014 reported that citriccid coated CoFe2O4 NPs are more injurious to biological mem-ranes compared to bare ones. Amine coating is considered safeor biological application as animals have the ability to metabolizemine or amine derivatives into less toxic chemicals and can eas-ly eliminate such toxic chemicals from their bodies. Irrespectivef round shape and amine coating, CoFe2O4 NPs showed exten-ively high toxicity even at lower concentration manifests severenherited toxic potential due to aggregation/agglomeration and
echanical damage by NPs. Larger agglomerates of Fe2O3 are moreoxic to the lipid membranes because of higher affinity for mem-ranes and higher cytotoxicity (Drasler et al., 2014; Mahmoudit al., 2012). In our study, rate of aggregation also increase withime and concentration, hence the toxicity. Adhesion of aggregates,edimentation, internalization of NPs and ions were also main con-ributing factors in inducing developmental toxicity in Zebrafishmbryos (Zhu et al., 2012).
Presence of metals (e.g., Cu, Co & Fe) and NPs can induce sig-ificant oxidative stress and apoptosis in biological systems (Pintot al., 2003) by catalyzing the formation of highly toxic ROS (OH•,2
•− and H2O2) species (Li et al., 2012). Nanoparticles of differentategories are reported to associate quickly with BSA (Lundqvist
t al., 2011). The decreased intensity of negative bands of �-helixt 208 and 222 nm position (Supplementary fig. S8) depicts BSA-helical content decreases while retaining the actual structure.ctually, these weak interactions between CoFe2O4 NPs with BSAlogy 166 (2015) 21–28 27
were governed by the surface charges of NPs and amino/carboxylicgroups of BSA (Vidic et al., 2014).
According to the results, CoFe2O4 NPs are very devastatingtoward Zebrafish embryo. Fig. 6 shows depletion of ROS at higherconcentrations and ROS bursts at lower concentrations. This indi-cates that CoFe2O4 NPs at higher concentration lead to apoptosisor necrosis (Fig. 5) while at a lower concentration initiates theuncontrolled proliferation of cells (Orel et al., 2015). Higher valueof Lipid peroxidation with increasing concentration of CoFe2O4 NPsis found to be due to the cumulative effect of internalization (Fig. 2)and aggregates/agglomerates CoFe2O4 NPs and of Fe3+ & Co2+ ions.NPs and ions (Fe3+ & Co2+) favored the pore formation by magne-tolysis (Sincai et al., 2001) and lipid peroxidation, making it morepermeable and less selective (Zhou et al., 2014). CoFe2O4 NPs acti-vates antioxidants by ROS production (Donaldson and Stone, 2003).A significant (p < 0.05) increase in the CAT activity after incubationwith CoFe2O4 NPs shows higher production of H2O2. Depletion ofGSH can be estimated by an increase in GST concentration (Lee et al.,2012), GST acts as an antioxidant and Phase II biotransformation,assisting in the elimination of xenobiotics. Higher activities of GSTand AP are also indicative of increased metabolic rates (Clementeet al., 2014) and vice versa. Activation of the antioxidant defensemechanisms containing CAT, AP and GST are possibly very com-plex (Mittler, 2002; Dewez et al., 2005), as their principle functionis to neutralize the ROS species no matter in which part of the cellthey are. While the toxicity of CoFe2O4 NPs by ROS production canbe attributed to CoFe2O4 NPs or Fe3+ and Co2+ accumulation andspecific functions in different cellular compartments. There wasan increase in protein content due to oxidative breakdown of longchains of amino acids (Herman et al., 2007).
5. Conclusion
CoFe2O4 NPs induce acute developmental toxicity in dose andtime dependent way by arresting cell cycle, apoptosis, decreasedmetabolism, hatching delay, instable heart beat, cardiac/yolk sacedema, tail and spinal cord flexture. Our findings also suggest thatCoFe2O4 NPs also cause behavioral changes in Zebrafish embryosmaking them sessile at the bottom with reduced visibility. CoFe2O4NPs rapidly envelop the chorion of the embryo and induce physi-cal stress resulting reduced movement of embryos with hatchingdelay. CoFe2O4 NPs insulted the membranes by ROS induced apo-ptosis in the head, heart and tail region. Interference of CoFe2O4 NPswas effectively reduced by additional washing and ultracentrifuga-tion steps. These results evidenced the potential risk of CoFe2O4 NPsto the environment and suggest complete risk assessment for priorbiological applications.
Acknowledgements
The authors are very grateful to the Project of Science andTechnology, Department of Zhejiang Province (2012C37058) andthe Key Innovation Team of Science and Technology in ZhejiangProvince (2010R50018) for financial support.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.aquatox.2015.07.003
References
Azaria, L.H., Baldi, G., Beno, D., Bonacchi, D., Golla-Schindle, U., Kirkpatrick, J.C.,Kolle, S., Landsiede, R., Maimon, O., Marche, P.N., Ponti, J., Romano, R., Rossi, F.,Sommer, D., Uboldi, C., Unger, R.E., Villiers, C., Korenstein, R., 2013. Predictive

2 Toxico
A
B
B
C
C
C
C
D
D
D
D
F
G
H
H
H
H
I
I
K
K
K
L
L
L
8 F. Ahmad et al. / Aquatic
toxicology of cobalt ferrite nanoparticles: comparative in-vitro study ofdifferent cellular models using methods of knowledge discovery from data.Part. Fibre Toxicol. 10, 32.
zaria, L.H., Kirkpatrick, C.J., Korenstein, R., Marche, P.N., Maimon, O., Ponti, J.,Romano, R., Rossi, F., Schindler, U.G., Sommer, D., Uboldi, C., Unger, R.E.,Villiers, 2011. Predictive toxicology of cobalt nanoparticles and ions:comparative in vitro study of different cellular models using methods ofknowledge discovery from data. Toxicol. Sci. 122, 489–501.
arhoumi, L., Dewez, D., 2013. Toxicity of superparamagnetic iron oxidenanoparticles on green alga Chlorella vulgaris. BioMed. Res. Int., 1, Article ID647974.
raunbeck, T., Böttcher, M., Hollert, H., Kosmehl, T., Lammer, E., Leist, E., Rudolf, M.,Seitz, N., 2005. Towards an alternative for the acute fish LC50 test in chemicalassessment: the fish embryo toxicity test goes multi-species—an update.ALTEX 22, 87–102.
hen, C.Y., Li, Y.F., Qu, Y., Chaibc, Z.F., Zhao, Y.L., 2013. Advanced nuclear analyticaland related techniques for the growing challenges in nanotoxicology. Chem.Soc. Rev. 42, 8266–8303.
hoi, S.J., Lee, J.K., Jeong, J., Choy, J.H., 2013. Toxicity evaluation of inorganicnanoparticles: considerations and challenges. Mol. Cell. Toxicol. 9, 205–210.
lemente, Z., Castro, V.L.S.S., Moura, M.A.M., Jonsson, C.M., Fraceto, L.F., 2014.Toxicity assessment of TiO2 nanoparticles in zebrafish embryos underdifferent exposure conditions. Aquat. Toxicol. 147, 129–139.
olognato, R., Bonelli, A., Bonacchi, D., Baldi, G., Migliore, L., 2007. Analysis ofcobalt ferrite nanoparticles induced genotoxicity on human peripherallymphocytes: comparison of size and organic grafting-dependent effect.Nanotoxicology 1, 301–308.
evasagayam, T.P., Tilak, J.C., Boloor, K.K., Sane, K.S., Ghaskadbi, S.S., Lele, R.D.,2004. Free radicals and antioxidants in human health: current status andfuture prospects. J. Assoc. Physician India 52, 794–804.
ewez, D., Geoffroy, L., Vernet, G., Popovic, R., 2005. Determination ofphotosynthetic and enzymatic biomarkers sensitivity used to evaluate toxiceffects of copper and fludioxonil in alga Scenedesmus obliquus. Aquat. Toxicol.74, 150–159.
onaldson, K., Stone, V., 2003. Current hypotheses on the mechanisms of toxicityof ultrafine particles. Ann. Ist Super Sanità 39, 405–410.
rasler, B., Drobne, D., Novak, S., Valant, J., Boljte, S., Otrin, L., Rappolt, M., Artori, B.,Iglic, A., Iglic, V.K., Sustar, V., Makovec, D., Gyergyek, S., Hocevar, M., Godec, M.,Zupanc, J., 2014. effects of magnetic cobalt ferrite nanoparticles on biologicaland artifiial lipid membranes. Int. J. Nanomed. 9, 1559–1581.
ako, V.E., Furgeson, D.Y., 2009. Zebrafish as a correlative and predictive model forassessing biomaterial nanotoxicity. Adv. Drug Deliv. Rev. 61, 478–486.
uglielmo, C.D., López, D.R., Lapuente De, J., Llobet Mallafre, J.M., Suàrez, M.B.,2010. Embryotoxicity of cobalt ferrite and gold nanoparticles: a first in vitroapproach. Reprod. Toxicol. 30, 271–276.
erman, N., Cos, P., Maes, L., BruyneT De, D., Berghe, V., Vlietinck, A.J., Pieters, L.,2007. Challenges and pitfalls in antioxidant research. Curr. Med. Chem. 14,417–430.
ill, A.J., Teraoka, H., Heideman, W., Peterson, R.E., 2005. Zebrafish as a modelvertebrate for investigating chemical toxicity. Toxicol. Sci. 86, 6–19.
ong, N.H., Raghavender, A.T., Ciftja, P.M.H., Stojak, K.S.H., Zhang, Y.H., 2013.Ferrite nanoparticles for future heart diagnostics. Appl. Phys. A 112,323–327.
wang, D.W., Lee, D.S., Kim, S.H., 2012. Gene expression profiles for genotoxiceffects of silica-free and silica-coated cobalt ferrite nanoparticles. J. Nucl. Med.53, 1–7.
SO, 2007. Water quality—Determination of the acute toxicity of waste water tozebrafish eggs (Danio rerio). ISO 15088:2007 (E).
wase, T., Tajima, A., Sugimoto, S., Okuda, K., Hironaka Ippei, Kamata Yuko, Takada,K., Mizunoe, Y., 2013. A simple assay for measuring catalase activity: a visualapproach. Sci. Rep. 3, 3081, http://dx.doi.org/10.1038/srep03081
ane, D.A., Hammerschmidt, M., Mullins, M.C., Maischein, H.M., Brand, M., vanEeden, F.J., Furutani-Seiki, M., Granato, M., Haffter, P., Heisenberg, C.P., Jiang,Y.J., Kelsh, R.N., Odenthal, J., Warga, R.M., Nusslein-Volhard, C., 1996. Thezebrafish epiboly mutants. Development 123, 47–55.
apilevich, L.V., D’yakova Yu, E., Nosarev, A.V., Zaitseva, T.N., Petlina, Z.R.,Ogorodova, L.M., Ageev, B.G., Magaeva, A.A., Itin, V.I., Terekhova, O.G., 2010.Effect of nanodisperse ferrite cobalt (CoFe2O4) particles on contractilereactions in guinea pigs airways. Bull. Exp. Biol. Med. 149, 77–79.
rug, H.F., 2014. Nanosafety research—are we on the right track? Angew. Chem.53, 12304–12319.
ammer, E., Kamp, H.G., Hisgen, V., Koch, M., Reinhard, D., Salinas, E.R., Wendler, K.,Zok, S., Braunbeck, T., 2009. Development of a flow-through system for the fishembryo toxicity test (FET) with the zebrafish (Danio rerio). Toxicol. In Vitro 23,1436–1442.
ee, B., Duong, C.N., Cho, J., Lee, J., Kim, K., Seo, Y., Kim, P., Choi, K., Yoon, J., 2012.
Toxicity of citrate-capped silver nanoparticles in common carp (Cyprinuscarpio). J. Biomed. Biotechnol., 262–670.i, Y., Zhang, W., Niu, J., Chen, Y., 2012. Mechanism of photogenerated reactiveoxygen species and correlation with the antibacterial properties of engineeredmetal-oxide nanoparticles. ACS Nano 6, 5164–5173.
logy 166 (2015) 21–28
Lundqvist, M., Stigler, J., Cedervall, T., Berggard, T., Flanagan, M.B., Lynch, I., Elia, G.,Dawson, K., 2011. The evolution of the protein corona around nanoparticles: atest study. ACS Nano 5, 7503–7509.
Luo, C., Li, Y. Yang, L. Wang, X. Long, J. Liu, J., 2014. Superparamagnetic iron oxidenanoparticles exacerbate the risks of reactive oxygen species-mediatedexternal stresses. Arch. Toxicol., http://dx.doi.org/10.1007/s00204-014-1267-x
Luyts, K., Napiersk, D., Nemery, B., Hoet, P.H.M., 2013. How physico-chemicalcharacteristics of nanoparticles cause their toxicity: complex and unresolvedinterrelation. Environ. Sci. Process. Impacts 1, 5–23.
Maccormack, T.J., Clark, R.J., Dang, M.K.M., Kelly, J.A., Veinot, J.G.C., Goss, G.G., 2012.Inhibition of enzyme activity by nanomaterials: potential mechanisms andimplications for nanotoxicity testing. Nanotoxicology 6, 514–525.
Mahmoudi, M., Hofmann, H., Rutishauser, B.R., Fink, A.P., 2012. Assessing thein vitro and in vivo toxicity of superparam agnetic iron oxide nanoparticles.Chem. Rev. 112, 2323–2338.
Marmorato, P., Ceccone, G., Gianoncelli, A., Pascolo, L., Ponti, J., Rossia, F., Saloméc,M., Kaulich, B., Kiskinova, M., 2011. Cellular distribution and degradation ofcobalt ferrite nanoparticles in Balb/3T3 mouse fibroblasts. Toxicol. Lett. 207,128–136.
Maurer-Jones, M.A., Gunsolus, I.L., Murphy, C.J., Haynes, C.L., 2013. Toxicity ofengineered nanoparticles in the environment. Anal. Chem. 85, 3036–3049.
McGahon, A.J., Martin, S.J., Bissonnette, R.P., Mahboubi, A., Shi, Y.F., Mogil, R.J.,Nishioka, W.K., Green, D.R., 1995. The end of the (cell) line-methods for thestudy of apoptosis in vitro. Methods Cell. Biol. 46, 153–185.
Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends PlantSci. 7, 405–410.
Nicanuzia dos Prazeres, J., Ferreira, C.V., Aoyama, H., 2004. Acid phosphataseactivities during the germination of glycine max seeds. Plant Phys. Biochem.42, 15–20.
Novak, S., Drobne, D., Golobic, M., Zupanc, J., Romih, T., Gianoncelli, A., Kiskinova,M., Kaulich, B., Pelicon, P., Vavpetic l, J.L., Ogrinc, N., Makovec, D., 2013. Cellularinternalization of dissolved cobalt ions from ingested CoFe2O4 nanoparticles:in vivo experimental evidence. Environ. Sci. Technol. 47, 5400–5408.
OECD, 2012. Series on Testing and Assessment No 179 – validation report (phase 2)for the zebrafish embryo toxicity test; Series on Testing and Assessment No171 – Fish toxicity testing framework.
Ohkawa, H., Ohishi, N., Yagi, K., 1979. Assay for lipid peroxides in animal tissues bythiobarbituric acid reaction. Anal. Biochem. 95, 35l–358.
Ong, K.J., MacCormack, T.J., Clark, R.J., Ede, J.D., Ortega, V.A., et al., 2014. Widespreadnanoparticle-assay interference: implications for nanotoxicity testing. PLoSOne 9 (3), e90650, http://dx.doi.org/10.1371/journal.pone.0090650
Orel, V., Shevchenko, A., Romanov, A., Tselepi, M., Mitrelias, T., Barnes, C.H.W.,Burlaka, A., Lukin, S., Shchepotin, I., 2015. Magnetic properties and antitumoreffect of nanocomplexes of iron oxide, and doxorubicin. Nanomed.: NBM 11,47–55.
Papis, E., Gornati, R., Prati, M., Ponti, J., Sabbioni, E., Bernardini, G., 2007. Geneexpression in nanotoxicology research: analysis by differential display inBALB3T3 fibroblasts exposed to cobalt particles and ions. Toxicol. Lett. 170,185–192.
Pinto, E., Sigaud-Kutner, T.C.S., Leitao, M.A.S., Okamoto, O.K., Morse, D., Colepicolo,P., 2003. Heavy metal-induced oxidative stress in algae. J. Phycol. 39,1008–1018.
Ponti, J., Sabbioni, E., Munaro, B., Broggi, F., Marmorato, P., Franchini, F., Colognato,R., Rossi, F., 2009. Genotoxicity and morphological transformation induced bycobalt nanoparticles and cobalt chloride: an in vitro study in Balb/3T3 mousefibroblasts. Mutagen 24, 439–445.
Sanpo, N., Berndt, C.C., Wen, C., Wang, J., 2013. Transition metal-substituted cobaltferrite nanoparticles for biomedical applications. Acta Biomater. 9, 5830–5837.
Sincai, M., Ganga, D., Bica, D., Vekas, L., 2001. The antitumor effect of locoregionalmagnetic cobalt ferrite in dog mammary adenocarcinoma. J. Magn. Magn.Mater. 225, 235–240.
Sun, Z.Z., Yathindranath, V., Worden, M., Thliveris, J.A., Chu, S., Parkinson, F.E.,Hegmann, T., Miller, D.W., 2013. Characterization of cellular uptake andtoxicity of aminosilane-coated iron oxide nanoparticles with different chargesin central nervous system relevant cell culture models. Int. J. Nanomed. 8,961–970.
Vidic, J., Haque, F., Guigne, J.M., Vidy, A., Chevalier, C., Stankic, S., 2014. Effects ofwater and cell culture media on the physicochemical properties of ZnMgOnanoparticles and their toxicity toward mammalian cells. Langmuir 30,11366–11374.
Wang, Y., Seebald, J.L., Szeto, D.P., Irudayaraj, J., 2010. Biocompatibility andbiodistribution of surface-enhanced Raman scattering nanoprobes in zebrafishembryos: in vivo and multiplex imaging. ACS Nano 4 (7), 4039–4053.
Westerfield, M., 1995. The Zebrafish Book: A Guide for the Laboratory Use ofZebrafish (Brachydanio rerio). University of Oregon Press, Eugene, pp. 267–272.
Zhou, H., Wang, X., Zhou, Y., Yao, H., Ahmad, F., 2014. Evaluation of the toxicity of
ZnO nanoparticles to Chlorella vulgaris by use of the chiral perturbationapproach. Anal. Bioanal. Chem. 406, 3689–3695.Zhu, X., Tian, S., Cai, Z., 2012. Toxicity assessment of iron oxide nanoparticles inzebrafish (Danio rerio) early life stages. PLoS One 7 (9), e46286, http://dx.doi.org/10.1371/journal.pone.0046286.





























