Embed Size (px)
Citation preview

Anesthesia induces stress in Atlantic salmon (Salmo salar),Atlantic cod (Gadus morhua) and Atlantic halibut
(Hippoglossus hippoglossus)
Inger Hilde Zahl Æ Anders Kiessling Æ
Ole Bent Samuelsen Æ Rolf Erik Olsen
Received: 11 February 2009 / Accepted: 17 June 2009 / Published online: 13 August 2009
� Springer Science+Business Media B.V. 2009
Abstract Stress in response to anesthesia with
benzocaine, MS-222, metomidate and isoeugenol
was studied in Atlantic salmon (Salmo salar), Atlan-
tic halibut (Hippoglossus hippoglossus), and Atlantic
cod (Gadus morhua) with no concomitant stress from
handling or confinement in association with anesthe-
sia or sampling. All of the anesthetics tested induced
a stress response in all species, displayed by a release
of cortisol to the water. MS-222 anesthesia elicited
the highest cortisol release rates, reaching maximum
levels 0.5 h post-exposure and returning to basal
levels after 3–4 h. Benzocaine anesthesia caused a
bimodal response where the initial peak in cortisol
release rate was followed by a second increase lasting
towards the end of the trial (6 h). This bimodality was
more profound in Atlantic salmon than in Atlantic
halibut and Atlantic cod. Metomidate anesthesia
induced the lowest release of cortisol of the agents
tested in both Atlantic halibut and Atlantic cod, but
resulted in a bimodal response in Atlantic salmon
where the initial increase in cortisol release was
followed by a larger increase peaking at 2–2.5 h post
exposure before returning to basal levels after 5 h.
The stress induced in Atlantic salmon by isoeugenol
anesthesia resembled that of MS-222, but did not
reach the same elevated level. Overall, the cortisol
release was most profound in Atlantic salmon
followed by Atlantic halibut and Atlantic cod.
Keywords Anesthesia � Stress � Cortisol �
MS-222 � Benzocaine � Metomidate �
Isoeugenol
Introduction
During commercial fish farming, the fish are sub-
jected to various practices of handling and confine-
ment. All these situations are associated with acute
stress, which in addition to producing some charac-
teristic stress reactions in the fish all give rise to a
non-specific generalized physiological response,
commonly described by three stages (Wendelaar
I. H. Zahl (&) � O. B. Samuelsen � R. E. Olsen
Institute of Marine Research, P.O. Box 1870, Nordnes,
5817 Bergen, Norway
e-mail: [email protected]
A. Kiessling
Department of Animal and Aquacultural Sciences,
Norwegian University of Life Sciences, P.O. Box 5003,
1432 Aas, Norway
I. H. Zahl � O. B. Samuelsen
Department of Biology, University of Bergen,
P.O. Box 7800, 5020 Bergen, Norway
A. Kiessling
Department of Wildlife, Fish and Environmental Studies,
Swedish University of Agricultural Sciences, 901 83
Umea, Sweden
123
Fish Physiol Biochem (2010) 36:719–730
DOI 10.1007/s10695-009-9346-2

Bonga 1997). The initial reaction is associated with
an activation of neuroendocrine pathways leading to
a massive release of hormones, primarily catechola-
mines and corticosteroids (Donaldson 1981; Mazeaud
and Mazeaud 1981; Mommsen et al. 1999; Reid et al.
1998). This hormonal release results in mobilization
of energy stores to meet the increased energy demand
and stimulates the cardiovascular and respiratory
function, which is important for the fish in overcom-
ing the stressful event and regaining homeostasis.
However, situations of severe stress, and situations of
chronic or recurrent incidents of stress where the fish
are not given time to recover have detrimental effects
which eventually lead to a general impairment of
condition and performance (Barton 2002; Davis
2006; Jentoft et al. 2005; Pickering and Pottinger
1989). These harmful effects from stress are related
to elevated levels of corticosteroids, primarily corti-
sol (Barton and Iwama 1991; Maule et al. 1989;
Mommsen et al. 1999; Pickering and Pottinger 1989).
Cortisol is synthesized and released more slowly than
the catecholamines noradrenaline and adrenaline and
returns more slowly to basal level (Olsen et al. 2002,
2008; Pickering and Pottinger 1989; Sumpter et al.
1986). Cortisol has therefore been widely used as a
parameter in studies of stress in fish and has revealed
that large variations in the magnitude of the response
exist both between and within species (Barton 2000;
Barton and Iwama 1991; Pottinger and Moran 1993;
Pottinger et al. 1994).
In order to facilitate handling, reduce the risk of
injuries, reduce the risk of inflicting pain, and prevent
stress, anesthetic agents are used. While several
studies report that anesthetics are effective in reduc-
ing the stress associated with confinement and
handling (Davis and Griffin 2004; Iversen et al.
2003; Olsen et al. 1995; Sink et al. 2007; Small 2004;
Small and Chatakondi 2005; Thomas and Robertson
1991) some also indicate that the exposure to
anesthetic agents in itself induces a stress response,
measured by increased levels of cortisol (Barton and
Peter 1982; Davidson et al. 2000; Davis and Griffin
2004; Kiessling et al. 2009; Molinero and Gonzalez
1995; Thomas and Robertson 1991). In a preliminary
study carried out at our laboratory, markedly elevated
plasma cortisol levels were found in juvenile Atlantic
salmon (Salmo salar) subjected to stress in the form
of netting although a preventive treatment with
anesthetics had been applied prior to the stressor
(Kiessling, Johansson and Axen, pers. comm.). This,
in combination with work using adult Atlantic salmon
fitted with dorsal aorta cannulae for studying con-
current plasma cortisol and clearance of anesthetic
agents (Kiessling et al. 2009) clearly indicated that
anesthetics could themselves induce a severe stress
reaction, and that the reaction varied consistently
between different agents. However, neither of the
above-mentioned studies was designed to study the
direct causality between exposure to anesthetic
agents and the induction of a stress reaction.
MS-222, benzocaine, metomidate and isoeugenol
are among the most common anesthetic agents used
for fish (Ackerman et al. 2005; Ross and Ross 2008).
MS-222 and benzocaine are local anesthetics that
inhibit neural signal transmission by blocking voltage
sensitive sodium channels (Frazier and Narahashi
1975; Neumcke et al. 1981). Isoeugenol is similar to
eugenol, a widely used analgesic in dentistry which
inhibits sodium, potassium, and calcium channels,
inhibits N-methyl-D-aspartate (NMDA) receptors and
potentiates inhibitory gamma-aminobutyric acid type
A (GABAA) receptors (Aoshima and Hamamoto
1999; Lee et al. 2005; Li et al. 2007; Park et al. 2006;
Wie et al. 1997). Metomidate is a methyl analogue of
etomidate, a non-barbiturate hypnotic that modulates
and activates GABAA receptors (Ashton and Wau-
quier 1985; Grasshoff et al. 2006; Rusch et al. 2004;
Yang and Uchida 1996). Metomidate has been found
to inhibit the cortisol stress response in fish (Olsen
et al. 1995; Thomas and Robertson 1991). The aim of
the current study was to examine whether exposure to
these anesthetic agents would elicit a stress response
in Atlantic salmon, Atlantic cod (Gadus morhua), and
Atlantic halibut (Hippoglossus hippoglossus). The
non-invasive method developed by Ellis et al. (2004)
for assessing the cortisol status of fish by measuring
the level of cortisol released to water was used in
order to obtain relative measurements over time
without disturbing the fish.
Materials and methods
Experimental fish
Atlantic cod and Atlantic halibut were produced at
IMR, Austevoll Research Station, Norway. Upon
reaching an average weight of 400 g (Atlantic cod)
720 Fish Physiol Biochem (2010) 36:719–730
123

and 200 g (Atlantic halibut), 114 Atlantic cod and
180 Atlantic halibut were randomly distributed into
six 600-l tanks to a total biomass of 25 kg m-3 and
20 kg m-3, respectively. The tanks were supplied
with aerated seawater at a flow rate of 20 l min-1
with an average temperature of 9 ± 1�C. The fish
were kept under natural light and were fed daily by
automatic feeders a commercial marine diet (Skrett-
ing, Stavanger, Norway).
Atlantic salmon (Aqua Gen strain) were obtained
from Aqua Gen, Hemne, Norway, and raised to
experimental size at IMR, Matre, Norway. At an
average weight of 420 g the Atlantic salmon were
randomly distributed in three tanks of 1,000 l, 48 fish
per tank, to a total biomass of 20 kg m-3. The tanks
were supplied with a continuous flow of aerated
seawater of 58 ± 6 l min-1 with a temperature of
9 ± 1�C. The fish were kept under natural light and
fed daily with a commercial marine diet (Biomar,
Myre, Norway) by automatic feeders.
The fish were allowed to adapt to the experimental
conditions for 1 month before experiments were
started and were given 1–2 weeks to recover between
each anesthetic exposure.
Experimental procedure
The fish were deprived of diets 24 h prior to the start
of the experiments and triplicates tanks were used
for each treatment. The experiment was initiated by
carefully stopping the water flow. When the valves
were closed the anesthetic was added to each tank
with as little disturbance to the fish as possible. The
water current generated by the newly stopped water
flow caused dispersion of the anesthetic agent
throughout the tanks. Control groups were treated
the same way except that no anesthetic was added.
The water flow was closed for 5 min allowing the
drug to affect the fish and then reopened to the
initial flow washing the anesthetic out. For water
cortisol analysis samples of 1 l were taken from the
inflowing water and from the outlet of each tank
prior to initiation of each experiment and then at
intervals of 0.5 h from 0.5 h to 4 h, and at 5 h and
6 h following administration of the anesthetic. All
samples were collected via the outlet of the tanks
without disturbing the fish and were stored at -20�C
until analyzed.
Anesthetic agents
The following anesthetic agents were tested: benzo-
caine (Benzoak� Vet, A.C.D Pharmaceuticals, Lek-
nes, Norway), tricaine methanesulfonate (MS-222�,
Pharmaq AS, Oslo, Norway), metomidate hydrochlo-
ride (Aquacalm�, Syndel International Inc., Vancou-
ver, Canada) and isoeugenol (Aqui-S�, Scan Aqua
AS, Aarnes, Norway). Manufacturer’s recommenda-
tions of dosages were followed. An overview of the
anesthetics and dosages administrated to each species
is presented in Table 1.
Water sample processing and cortisol analysis
The water samples were processed according to
Greenwood et al. (2001) and Ellis et al. (2004). Using
a peristaltic pump (Watson–Marlow 520S, Watson–
Marlow Bredel Pumps Ltd., Falmouth, UK) set at
25 ml min-1, the tank water samples were first
pumped through a pre-filter (AcroCapTMFilter, pore
size 0.45 lm, Pall Corporation, East Hills, USA) to
remove particles and then through an activated solid-
phase extraction cartridge, SPEC (Sep-pak� PlusC18,
55–105 lm, Waters Corporation, Milford, USA). The
pre-filters were rinsed with 20 ml of de-ionized water
prior to use and the SPECs were conditioned with
10 ml of methanol followed by 10 ml of de-ionized
water. After loading of the samples, the SPECs were
washed with 5 ml of de-ionized water and dried by
aspirating air, and thereafter stored at -20�C until
further processed.
To determine the exact volume of each water
sample, the water bottles were weighed before and
after pumping using a conversion factor of 1. The
Table 1 Anesthetic agents and dosages administrated to
Atlantic cod, Atlantic halibut, and Atlantic salmon
Benzocaine
(mg l-1)
MS-222
(mg l-1)
Metomidate
(mg l-1)
Isoeugenol
(mg l-1)
Atlantic cod 20 55 3 –
Atlantic
halibut
40 80 3 –
Atlantic
salmon
40 55a 3a 5.4a
MS-222, metomidate and isoeugenol were dissolved in water
before being added to the tanks (n = 3)a n = 2
Fish Physiol Biochem (2010) 36:719–730 721
123

SPECs were thawed and processed as described by
Sorensen and Scott (1994) as modified by Ellis et al.
(2004). In brief: the sample was eluted from the
SPECs using 5 ml of ethyl acetate. The eluate was
evaporated to dryness under nitrogen gas at 45�C and
the residue dissolved in 320 ll Calibrator Diluent
RD5-43, a buffered protein base included in the
cortisol immunoassay kit (ParameterTM KGE008,
R&D Systems Europe, Ltd., Abingdon, UK), which
was used in order to determine the cortisol concen-
tration. According to the specification of the immu-
noassay kit, the cross-reactivity with other steroids
was 4.4% with prednisolone, 1.7% with progesterone,
0.2% with cortisone, and\0.1% with corticosterone,
deoxycorticosterone, estradiol, and prednisone. The
intra- and inter-assay precision was 9.2% (n = 20)
and 21.1% (n = 40) (coefficient of variation, CV)
respectively, and the limit of sensitivity (minimum
detectable dose) was 0.071 ng l-1. The analysis was
carried out in compliance with the assay procedure of
the kit.
Calculation of cortisol release rates
The concentration of cortisol in each water sample
was calculated from the standard curve. Background
cortisol concentration of the inflowing water sampled
prior to initiation of each experiment (0.07 ± 0.01
ng l-1) was subtracted. The cortisol release rate
(ng g-1 h-1) of the fish in each tank was then
calculated from the fish biomass of the tank, the tank
water volume and water flow, according to Ellis et al.
(2004) and Folkedal et al. (in prep).
Statistics
The data were analyzed by the main factorial model
(general linear model, Statistical Analysis System
(SAS) for PC (ver.8.2), ANOVA for unbalanced data).
Included in the model as main factors were sample
time (categorical), anesthetics (categorical), tank as
block (categorical). Groups were compared by the ad
hoc variance test (F-test) using the least-squares
means procedure when significant effects were found
in the main model. The level of statistical significance
was set at P\ 0.05. All data were tested for normality
by a normal probability plot (proc univariate).
Results
Atlantic cod
A significant increase in the release of cortisol in
response to anesthesia was seen in all treatment
groups, with the highest measured release rate at 0.5 h
(Fig. 1; Table 2). MS-222 elicited the highest increase
in cortisol with a peak value at 0.5 h of 0.28 ±
0.03 ng g-1 h-1 followed by benzocaine (0.23 ±
0.02 ng g-1 h-1) and metomidate (0.05 ± 0.02 ng
Fig. 1 Cortisol release
rates into water by Atlantic
cod after exposure to
different anesthetic agents,
and control (no anesthetic).
Values are mean ± SE
(n = 3)
722 Fish Physiol Biochem (2010) 36:719–730
123
g-1 h-1). The cortisol release then decreased in a
linear manner returning to the initial basal level 2 h
post-exposure. Although the increase in cortisol
release following benzocaine anesthesia was slightly
lower compared to MS-222 at 0.5 h, the release of
cortisol was maintained at a higher level throughout
the experiment returning to basal levels only after 6 h
post-exposure. The small increase in cortisol secretion
after exposure to metomidate returned quickly to basal
level within 1.5 h of exposure. The temporal halt in
water supply did not seem to elicit any stress response
in the fish as no increase in cortisol release was seen in
the untreated control group during the 4-h period.
Atlantic halibut
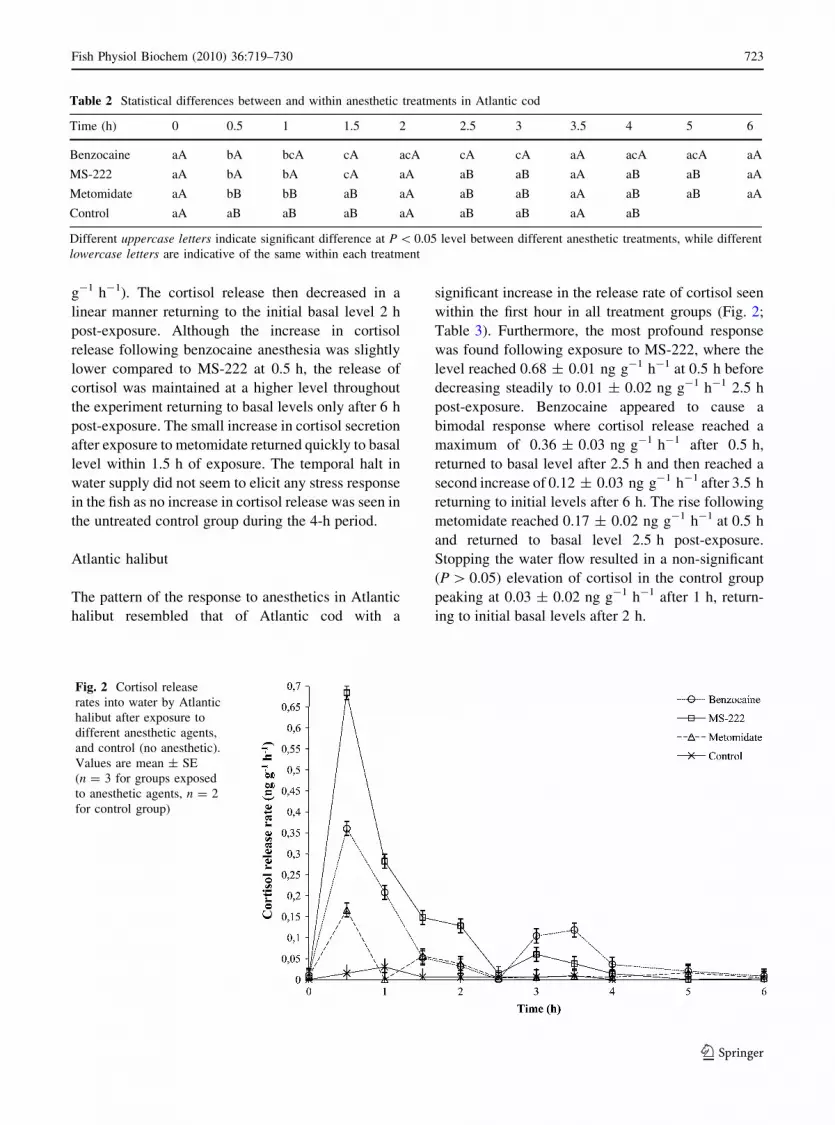
The pattern of the response to anesthetics in Atlantic
halibut resembled that of Atlantic cod with a
significant increase in the release rate of cortisol seen
within the first hour in all treatment groups (Fig. 2;
Table 3). Furthermore, the most profound response
was found following exposure to MS-222, where the
level reached 0.68 ± 0.01 ng g-1 h-1 at 0.5 h before
decreasing steadily to 0.01 ± 0.02 ng g-1 h-1 2.5 h
post-exposure. Benzocaine appeared to cause a
bimodal response where cortisol release reached a
maximum of 0.36 ± 0.03 ng g-1 h-1 after 0.5 h,
returned to basal level after 2.5 h and then reached a
second increase of 0.12 ± 0.03 ng g-1 h-1 after 3.5 h
returning to initial levels after 6 h. The rise following
metomidate reached 0.17 ± 0.02 ng g-1 h-1 at 0.5 h
and returned to basal level 2.5 h post-exposure.
Stopping the water flow resulted in a non-significant
(P[ 0.05) elevation of cortisol in the control group
peaking at 0.03 ± 0.02 ng g-1 h-1 after 1 h, return-
ing to initial basal levels after 2 h.
Table 2 Statistical differences between and within anesthetic treatments in Atlantic cod
Time (h) 0 0.5 1 1.5 2 2.5 3 3.5 4 5 6
Benzocaine aA bA bcA cA acA cA cA aA acA acA aA
MS-222 aA bA bA cA aA aB aB aA aB aB aA
Metomidate aA bB bB aB aA aB aB aA aB aB aA
Control aA aB aB aB aA aB aB aA aB
Different uppercase letters indicate significant difference at P\ 0.05 level between different anesthetic treatments, while different
lowercase letters are indicative of the same within each treatment
Fig. 2 Cortisol release
rates into water by Atlantic
halibut after exposure to
different anesthetic agents,
and control (no anesthetic).
Values are mean ± SE
(n = 3 for groups exposed
to anesthetic agents, n = 2
for control group)
Fish Physiol Biochem (2010) 36:719–730 723
123

Atlantic salmon
All treatments resulted in a significant elevation of the
cortisol (Fig. 3; Table 4). MS-222 caused a sharp
increase peaking at 0.47 ± 0.04 ng g-1 h-1 after
0.5 h, which was significantly higher than the other
anesthetics tested. The level then dropped rapidly
until 2.5 h (0.02 ± 0.01 ng g-1 h-1) and remained in
the region of the initial basal level throughout the
experiment. As seen with Atlantic cod and Atlantic
halibut, the increase in cortisol release of the benzo-
caine-treated group was lower than that of MS-222,
and appeared later, peaking at 0.29 ± 0.04 ng
g-1 h-1 after 1.5 h. A reduction in the release seen
after 2 h was immediately followed by a second
increase peaking after 2.5–3 h before slowly declining
Table 3 Statistical differences between and within anesthetic treatments in Atlantic halibut
Time (h) 0 0.5 1 1.5 2 2.5 3 3.5 4 5 6
Benzocaine aA bA bA aA aA aA cA cA aA aA aA
MS-222 aA bB cA dB dB aA eA aeB aeA aA aA
Metomidate aA bC aB aB aB aA aB aB aA aA aA
Control aA aD aB aB aB aA aB aB aA
Different uppercase letters indicate significant difference at P\ 0.05 level between different anesthetic treatments, while different
lowercase letters are indicative of the same within each treatment
Fig. 3 Cortisol release
rates into water by Atlantic
salmon after exposure to
different anesthetic agents,
and control (no anesthetic).
Values are means ± SE
(n = 3 for benzocaine and
control group, n = 2 for
metomidate, MS-222, and
isoeugenol)
Table 4 Statistical differences between and within anesthetic treatments in Atlantic salmon
Time (h) 0 0.5 1 1.5 2 2.5 3 3.5 4 5 6
Benzocaine aA bA bA bA cAB bA bA bcA cA cA cA
MS-222 aA bB cAB dB adA aB aB aB aB aB aB
Metomidate aA bBC cB cB bB bA acB acB acAB aB aB
Isoeugenol aA bC cAB bcB bA abB aB aB aB aB aB
Control aA aD aC aC aC aB aB aB aB
Different uppercase letters indicate significant difference at P\ 0.05 level between different anesthetic treatments, while different
lowercase letters are indicative of the same within each treatment
724 Fish Physiol Biochem (2010) 36:719–730
123

towards the last sampling at 6 h, but without reaching
basal levels. Metomidate appeared to cause a bimodal
response in cortisol secretion, where a small initial
increase was followed by a larger increase reaching
0.31 ± 0.05 ng g-1 h-1 between 1.5 and 3 h after
exposure. The release rate returned to the initial basal
level after 5 h. Isoeugenol was only tested in Atlantic
salmon. The exposure induced the lowest increase of
cortisol of the anesthetics tested. But as for the others,
there was a significant rise within the first 1.5 h, with a
maximum level of 0.23 ± 0.07 ng g-1 h-1 measured
1 h post-exposure returning to initial basal level after
5 h. Only small, and from zero non-significant,
fluctuations in cortisol release were seen in the control
group.
Discussion
Traditionally, the main incentive to use anesthesia for
fish has been to facilitate various forms of handling.
In more recent times, the perspective of fish welfare
has received increasing attention, including the
potential of using anesthetics to reduce or prevent
stress. The anesthetic agents are administered through
the water, either by being added directly to the
holding tanks without subjecting the fish to any
handling or confinement or to separate tanks in which
the fish are subsequently transferred. Subjecting the
fish to handling and confinement prior to anesthesia
would probably elicit a stress response (Olsen et al.
1995; Sumpter et al. 1986). Administration of
anesthetics directly to the tank without any distur-
bance may provide time for detecting the agents
through taste and smell. The agents may also act as
irritants to the skin by causing damage to the mucous
layer. Furthermore, as the anesthetic starts to take its
effect, the initial struggle to avoid losing balance may
possibly also induce stress. The mode of action of the
various agents will probably also affect the stress
response. Anesthetic agents may affect the endocrine
system and themselves induce elevations in plasma
cortisol concentrations (Oyama 1973; Oyama and
Wakayama 1988).
With reference to these variables, it is not
surprising that literature is contradictory with regard
to the amount of stress a given anesthetic may cause.
Stress in response to anesthesia, assessed by
increased levels of cortisol, has been reported for
several anesthetic agents in various fish species.
Exposure to MS-222 has been found to induce a
cortisol response in a range of species, including
Atlantic salmon, rainbow trout (Oncorhynchus
mykiss), channel catfish (Ictalurus punctatus), striped
bass (Morone saxatilis), hybrid striped bass (Morone
chrysops 9 Morone saxatilis), gilthead sea bream
(Sparus aurata), and red drum (Sciaenops ocellatus)
(Barton and Peter 1982; Davis and Griffin 2004;
Davis et al. 1982; Kiessling et al. 2009; Molinero and
Gonzalez 1995; Small 2003; Thomas and Robertson
1991). However, when administered in connection
with handling MS-222 has been found to suppress the
cortisol response in channel catfish, red drum and
black sea bass (Centropristis striata) (King et al.
2005; Small and Chatakondi 2005; Thomas and
Robertson 1991) but not in striped bass and hybrid
striped bass (Davis and Griffin 2004; Davis et al.
1982). Opposing results in rainbow trout indicate that
the stress-reducing potential of MS-222 during han-
dling depends on the nature of the handling (Barton
and Peter 1982; Sink et al. 2007). Metomidate has
been tested during various handling practices and has
been found to reduce the cortisol response in Chinook
salmon (Oncorhynchus tshawytscha), red drum,
channel catfish, black sea bass, hybrid striped bass
and Atlantic salmon (Davis and Griffin 2004; Iversen
et al. 2003; King et al. 2005; Kreiberg and Powell
1991; Olsen et al. 1995; Small 2003, 2004; Thomas
and Robertson 1991). The dosage of metomidate
required for preventing the stress response seems
however to vary both between and within species
(Iversen et al. 2003; Kreiberg and Powell 1991; Olsen
et al. 1995; Sandodden et al. 2001). Isoeugenol has
also been studied with respect to stress-reducing
potential in situations of handling and has been found
to reduce the cortisol response in channel catfish and
Atlantic salmon (Iversen et al. 2003; Small 2004;
Small and Chatakondi 2005). Isoeugenol was how-
ever not found to reduce the stress response to
handling in rainbow trout and hybrid striped bass, and
merely the exposure itself was sufficient for inducing
a cortisol response in these species (Davidson et al.
2000; Davis and Griffin 2004).
In the present study, all anesthetics induced a stress
response in the fish, displayed as a release of cortisol
into the water. Considering that the sampling was not
continuous, it is not possible to determine the exact
time for the cortisol peak. Maximum levels of cortisol
Fish Physiol Biochem (2010) 36:719–730 725
123

were measured at the first sampling point 0.5-h post-
exposure for all agents in Atlantic cod and Atlantic
halibut, whereas in Atlantic salmon the occurrence of
the cortisol peak varied between the anesthetic agents.
Regardless of species, MS-222 appeared to cause the
strongest stress response with cortisol peaking at 0.5 h
before returning to initial basal levels after 3–4 h. A
similar pattern of response has also been reported for
adult Atlantic salmon and Chinook salmon following
MS-222 exposure (Cho and Heath 2000; Kiessling
et al. 2009). The preliminary study with juvenile
Atlantic salmon, mentioned in the introduction,
showed the same trend as found here with MS-222
eliciting the highest increase in plasma cortisol
concentration followed by benzocaine and isoeuge-
nol, respectively, (Kiessling, Johansson and Axen,
pers. comm.). This further underlines that the stress
response induced by these anesthetic agents is
consistent also between species and life stages.
The purpose of administrating the anesthetics
directly to the water of the holding tanks and
collecting water samples from the outlet of the tanks
was to minimize the risk of inducing a stress reaction
due to handling or any other external stimuli that
would mask the effect of the anesthetic agents. The
success of this strategy was supported by the very low
release of cortisol from fish in the control group. The
absence of external stressors may explain the more
rapid clearance of cortisol seen in the present study
compared to earlier studies as these often includes
additional stress from handling in connection to
anesthesia or sample collection in the form of netting
and transfer into and out of anesthesia baths.
As benzocaine and MS-222 have the same mode of
action, it is interesting to note that the stress response
differed when fish were exposed to the two com-
pounds. Firstly, the rapid increase in cortisol release
upon exposure did not reach the same magnitude for
benzocaine as for MS-222. Secondly, the cortisol
release following benzocaine exposure was notably
bimodal with a second peak lasting towards the end of
trial (6 h) a pattern never seen for MS-222. This
bimodality was more profound in Atlantic salmon
than in Atlantic halibut and Atlantic cod. The
bimodality is consistent with findings in Atlantic
salmon anesthetized with benzocaine, given artificial
ventilation during recovery (Kiessling et al. 2009).
Bimodality in the stress response assessed by plasma
cortisol levels has also been observed in rainbow trout
(16 h post-anesthesia) and hybrid striped bass (mea-
sured 6 and 24 h post-anesthesia) anesthetized with
isoeugenol and quinaldine sulphate, respectively
(Davidson et al. 2000; Davis and Griffin 2004). The
dosage of MS-222 used in the current investigation
was higher than of benzocaine, for Atlantic halibut
twice as high. This may possibly contribute to the
difference in response between the two substances
observed immediately following exposure. The effect
of an anesthetic agent is closely related to pharmaco-
kinetic properties, i.e., absorption, distribution, clear-
ance, and elimination. In Atlantic salmon, clearance
rates of 3.1 l kg-1 h-1 and 0.35 l kg-1 h-1 have been
found for MS-222 and benzocaine respectively (Kies-
sling et al. 2009) indicating that the fish may need
longer time to recover from benzocaine than from
MS-222. Benzocaine has higher lipid solubility than
MS-222, and as anesthetics tend to accumulate in
tissues with a high content of fat (Rang et al. 2003)
this may lead to a build-up of benzocaine in fat-rich
tissues, possibly prolonging the effect. A substance
with higher lipid solubility may also more easily pass
through the blood–brain barrier and result in a more
profound central effect, depressing the centers con-
trolling respiration and circulation leading to insuffi-
cient respiration and gill circulation, which may delay
the elimination of the anesthetic, as the major pathway
for drug elimination in fish is over the gills (Hunn and
Allen 1974). The second peak of cortisol could
possibly relate to secondary metabolites formed due
to a delayed elimination or a more pronounced
hypoxia during the initial phase of the anesthesia.
Metomidate induced the lowest increase in cortisol
release of the agents examined here. In Atlantic cod
and Atlantic halibut, the small increase returned to
basal levels within 2 h. In Atlantic salmon, the initial
increase in release rate was followed by a larger
increase peaking at 2–2.5 h post-anesthesia, thus
resulting in a bimodal response not seen in Atlantic
cod or Atlantic halibut. As opposed to the sustained
elevation of cortisol characterizing the bimodal
response following benzocaine anesthesia, the corti-
sol levels returned to basal levels after 5 h. The
increased cortisol release following exposure to the
current dosage of metomidate (3 mg l-1) was rather
unexpected as metomidate has been reported to
prevent the stress response by inhibiting the synthesis
of cortisol (Olsen et al. 1995; Thomas and Robertson
1991). No increase in cortisol level was observed in
726 Fish Physiol Biochem (2010) 36:719–730
123

Atlantic salmon exposed to metomidate following
netting when a dosage of 2 mg l-1 was used (Iversen
et al. 2003), and a dosage of 3 mg l-1 was found to
prevent the cortisol response to handling stress (Olsen
et al. 1995). One can only speculate as to why
exposure to metomidate induced a stress response in
the current study. Since the fish were not subjected to
any stress from handling prior to exposure or in
connection to sampling a slow dispersion, introduc-
ing the fish to low concentrations of the drug might
have elicited a stress response. Low dosages of
metomidate added to the tank (0.5–1 mg l-1) have
previously been found insufficient for inhibiting the
stress response in Atlantic salmon (Eliason et al.
2007; Olsen et al. 1995) and have also been
inadequate for blocking the stress response during
transport (Sandodden et al. 2001). Low dosages of
metomidate (1.5 mg l-1) have on the other hand been
found to inhibit the cortisol response to confinement
stress in both channel catfish and hybrid striped bass
(Davis and Griffin 2004; Small 2004). Metomidate
added to the tanks did not elicit a stress response in
any of these species. There appears thus to be species
differences in the potency of metomidate in reducing
the stress response, supporting the data from the
current investigation. The fact that no further eleva-
tion of cortisol was seen in response to metomidate in
Atlantic cod and Atlantic halibut may indicate that
metomidate was not perceived as a strong stressor by
these species or that the concentration of metomidate
was sufficient for inhibiting any further elevation of
cortisol. The underlying rationale for this apparent
species difference in response may be a result of
behavior. Atlantic salmon is an active swimmer
normally responding to perceived reduction in O2
availability by a strong escape reaction. Metomidate
causes sedation and hypnosis by affecting GABAA
receptors. One of the side effects linked to agents that
act via these receptors is respiratory depression
(Grasshoff et al. 2006; Zeller et al. 2005). If lower
dosages inhibit the respiratory capacity but not the
perception of blood gases indicating a respiratory
insufficiency the fish might respond by stress.
Isoeugenol was only examined in Atlantic salmon.
The peak in cortisol release rate was lower following
exposure to isoeugenol than MS-222 and was mea-
sured 0.5 h later, but paralleled MS-222 in returning
to basal levels 4 h post-exposure. Stress in response to
isoeugenol exposure is consistent with previous
findings in Atlantic salmon (Iversen et al. 2003;
Kiessling et al. 2009, and Kiessling, Johansson and
Axen, pers. comm.). In these studies, however, the fish
were subjected to handling in connection with anes-
thesia. The dosage used in the current study is lower
than used by Kiessling et al. (2009) and Iversen et al.
(2003) but was chosen according to the manufac-
turer’s recommendation in order to obtain sedated and
handleable fish (5.4 mg l-1 isoeugenol corresponds to
10 ml Aqui-S� per 1,000 l).
Atlantic salmon subjected to acute stress in the
form of crowding, netting, and 1.5 min of air exposure
display an immediate release of cortisol, reaching a
maximum level of about 2.8 ng g-1 h-1 approxi-
mately 3 h post-stress (Ellis et al. 2007). In rainbow
trout subjected to a similar stressor, and in European
sea bass (Dicentrarchus labrax) chased for 5 min
prior to 1.5 min of air exposure cortisol release rates
have been found to peak at 1.3 ng g-1 h-1 after 1 h,
returning to basal levels 3 hours post-stress (Ellis et al.
2004; Fanouraki et al. 2008). In these studies the
cortisol release rate and plasma cortisol concentra-
tions were found to correspond. Plasma levels of
cortisol in Atlantic salmon subjected to acute stress
(30 s in air in dip net or chased in the tank for 15 min)
have previously been found to peak 1 hour post-stress,
reaching maximum levels of 110–215 ng ml-1 (Olsen
et al. 2002; Olsen et al. 1995). In Atlantic cod
subjected to the same stressors, increased levels of
plasma cortisol of 80–95 ng ml-1 have been found
0.5–1 h post-stress (King et al. 2006; Olsen et al.
2008). These reports show that identical stressors
elicit a higher cortisol production in Atlantic salmon
than in Atlantic cod, which support the findings in the
current investigation where the magnitude of the
cortisol release in response to the anesthetic agents
was more profound in Atlantic salmon than in Atlantic
cod. Furthermore, the temporal dynamics of the
cortisol response found here, with a maximum peak
in cortisol levels between 0.5 and 1.5 h is in line with
previous studies, both of plasma and water measure-
ments (Barton 2002; Barton and Iwama 1991; Scott
and Ellis 2007; Scott et al. 2008).
Conclusions
This study shows that exposure to anesthetic agents
elicits a stress response in Atlantic salmon, Atlantic
Fish Physiol Biochem (2010) 36:719–730 727
123

halibut, and Atlantic cod. MS-222 induces the most
rapid response in all species while benzocaine
induces a prolonged bimodal response, more pro-
found in Atlantic salmon and Atlantic cod than in
Atlantic halibut. The amount of cortisol released in
response to anesthesia is low compared to what is
reported following netting and air exposure, which
are considered to be strong stressors in fish, but may
represent an extra load during otherwise stressful
circumstances.
Acknowledgments The authors would like to thank the staff
of the Institute of Marine Research, and in particular Ivar
Helge Matre and Rina Helen Skoglund for their valuable
technical assistance, and Grethe Thorsheim for skilful help and
support with water sample processing and analysis. Ole
Folkedal and Thomas Torgersen are highly acknowledged for
proficient support in calculating cortisol release rates. Financial
support was given by the Norwegian Research Council, grant
no. 152898/120.
References
Ackerman PA, Morgan JD, Iwama GK (2005) Anesthetics.
CCAC guidelines on: the care and use of fish in research,
teaching and testing. Canadian Council on Animal Care,
Ottawa. Available at: http://www.ccac.ca/en/CCAC_
Programs/Guidelines_Policies/GDLINES/Fish/Fish%20
Anesthetics%20-%20ENG.pdf Accessed 10 Feb 2009
Aoshima H, Hamamoto K (1999) Potentiation of GABAA
receptors expressed in Xenopus oocytes by perfume and
phytoncid. Biosci Biotechnol Biochem 63:743–748
Ashton D, Wauquier A (1985) Modulation of a GABA-ergic
inhibitory circuit in the in vitro hippocampus by etomidate
isomers. Anesth Analg 64:975–980
Barton BA (2000) Salmonid fishes differ in their cortisol and
glucose responses to handling and transport stress. N Am J
Aquac 62:12–18
Barton BA (2002) Stress in fishes: a diversity of responses with
particular reference to changes in circulating corticoste-
roids. Integ Comp Biol 42:517–525
Barton BA, Iwama GK (1991) Physiological changes in fish
from stress in aquaculture with emphasis on the response
and effects of corticosteroids. Annu Rev Fish Dis 1:3–26
Barton BA, Peter RE (1982) Plasma cortisol stress response in
fingerling rainbow trout, Salmo gairdneri Richardson, to
various transport conditions, anaesthesia, and cold shock.
J Fish Biol 20:39–51
Cho GK, Heath DD (2000) Comparison of tricaine methane-
sulphonate (MS222) and clove oil anaesthesia effects on
the physiology of juvenile Chinook salmon Oncorhynchus
tshawytscha (Walbaum). Aquac Res 31:537–546
Davidson GW, Davie PS, Young G, Fowler RT (2000) Phys-
iological responses of rainbow trout Oncorhynchus mykiss
to crowding and anesthesia with AQUI-STM. J World
Aquac Soc 31:105–114
Davis KB (2006) Management of physiological stress in finfish
aquaculture. N Am J Aquac 68:116–121
Davis KB, Griffin BR (2004) Physiological responses of hybrid
striped bass under sedation by several anesthetics. Aqua-
culture 233:531–548
Davis KB, Parker NC, Suttle MA (1982) Plasma corticoste-
roids and chlorides in striped bass exposed to tricaine
methanesulfonate, quinaldine, etomidate, and salt. Prog
Fish-Cult 44:205–207
Donaldson EM (1981) The pituitary-interrenal axis as an
indicator of stress in fish. In: Pickering AD (ed) Stress and
Fish. Academic, London, pp 11–47
Eliason EJ, Kiessling A, Karlsson A, Djordjevic B, Farrell AP
(2007) Validation of the hepatic portal vein cannulation
technique using Atlantic salmon Salmo salar L. J Fish
Biol 71:290–297
Ellis T, James JD, Stewart C, Scott AP (2004) A non-invasive
stress assay based upon measurement of free cortisol
released into the water by rainbow trout. J Fish Biol
65:1233–1252
Ellis T, James JD, Sundh H, Fridell F, Sundell K, Scott AP
(2007) Non-invasive measurement of cortisol and mela-
tonin in tanks stocked with seawater Atlantic salmon.
Aquaculture 272:698–706
Fanouraki E, Papandroulakis N, Ellis T, Mylonas CC, Scott
AP, Pavlidis M (2008) Water cortisol is a reliable indi-
cator of stress in European sea bass, Dicentrarchus lab-
rax. Behaviour 145:1267–1281
Frazier DT, Narahashi T (1975) Tricaine (MS-222): effects on
ionic conductances of squid axon membranes. Eur J
Pharmacol 33:313–317
Grasshoff C, Drexler B, Rudolph U, Antkowiak B (2006)
Anaesthetic drugs: linking molecular actions to clinical
effects. Curr Pharm Des 12:3665–3679
Greenwood LN, Scott AP, Vermerissen ELM, Mylonas CC,
Pavlidis M (2001) Plasma steroids in mature common
dentex (Dentex dentex) stimulated with a gonadotropin-
releasing hormone agonist. Gen Comp Endocrinol 123:
1–12
Hunn JB, Allen JL (1974) Movement of drugs across gills of
fishes. Annu Rev Pharmacol Toxicol 14:47–55
Iversen M, Finstad B, McKinley RS, Eliassen RA (2003) The
efficacy of metomidate, clove oil, Aqui-STM and Benz-
oak� as anaesthetics in Atlantic salmon (Salmo salar L.)
smolts, and their potential stress-reducing capacity.
Aquaculture 221:549–566
Jentoft S, Aastveit AH, Torjesen PA, Andersen O (2005)
Effects of stress on growth, cortisol and glucose levels in
non-domesticated Eurasian perch (Perca fluviatilis) and
domesticated rainbow trout (Oncorhynchus mykiss).
Comp Biochem Physiol A 141:353–358
Kiessling A, Johansson D, Zahl IH, Samuelsen OB (2009)
Pharmacokinetics, plasma cortisol and effectiveness of
benzocaine, MS-222 and isoeugenol measured in indi-
vidual dorsal aorta-cannulated Atlantic salmon (Salmo
salar) following bath administration. Aquaculture
286:301–308
King WV, Hooper B, Hillsgrove S, Benton C, Berlinsky DL
(2005) The use of clove oil, metomidate, tricaine metha-
nesulphonate and 2-phenoxyethanol for inducing anaes-
thesia and their effect on the cortisol stress response in
728 Fish Physiol Biochem (2010) 36:719–730
123

black sea bass (Centropristis striata L.). Aquac Res
36:1442–1449
King WV, Buckley LJ, Berlinsky DL (2006) Effect of accli-
mation temperature on the acute stress response in juve-
nile Atlantic cod, Gadus morhua L., and haddock,
Melanogrammus aeglefinus L. Aquac Res 37:1685–1693
Kreiberg H, Powell J (1991) Metomidate sedation reduces
handling stress in chinook salmon. World Aquac 22:58–
59
Lee MH, Yeon KY, Park CK, Li HY, Fang Z, Kim MS, Choi
SY, Lee SJ, Lee S, Park K, Lee JH, Kim JS, Oh SB (2005)
Eugenol inhibits calcium currents in dental afferent neu-
rons. J Dent Res 84:848–851
Li HY, Park CK, Jung SJ, Choi SY, Lee SJ, Park K, Kim JS,
Oh SB (2007) Eugenol inhibits K? currents in trigeminal
ganglion neurons. J Dent Res 86:898–902
Maule AG, Tripp RA, Kaattari SL, Schreck CB (1989) Stress
alters immune function and disease resistance in chinook
salmon (Oncorhynchus tshawytscha). J Endocrinol
120:135–142
Mazeaud MM, Mazeaud F (1981) Adrenergic responses to
stress in fish. In: Pickering AD (ed) Stress and Fish.
Academic, London, pp 49–75
Molinero A, Gonzalez J (1995) Comparative effects of MS 222
and 2-phenoxyethanol on gilthead sea bream (Sparus
aurata L) during confinement. Comp Biochem Physiol A
111:405–414
Mommsen TP, Vijayan MM, Moon TW (1999) Cortisol in
teleosts: dynamics, mechanisms of action, and metabolic
regulation. Rev Fish Biol Fish 9:211–268
Neumcke B, Schwarz W, Stampfli R (1981) Block of Na
channels in the membrane of myelinated nerve by ben-
zocaine. Pflugers Arch 390:230–236
Olsen YA, Einarsdottir IE, Nilssen KJ (1995) Metomidate
anesthesia in Atlantic salmon, Salmo salar, prevents
plasma cortisol increase during stress. Aquaculture
134:155–168
Olsen RE, Sundell K, Hansen T, Hemre GI, Myklebust R,
Mayhew TM, Ringo E (2002) Acute stress alters the
intestinal lining of Atlantic salmon, Salmo salar L.: an
electron microscopical study. Fish Physiol Biochem
26:211–221
Olsen RE, Sundell K, Ringo E, Myklebust R, Hemre GI,
Hansen T, Karlsen O (2008) The acute stress response in
fed and food deprived Atlantic cod, Gadus morhua L.
Aquaculture 280:232–241
Oyama T (1973) Endocrine responses to anesthetic agents. Br J
Anaesth 45:276–281
Oyama T, Wakayama S (1988) The endocrine responses to
general anesthesia. Int Anesthesiol Clin 26:176–181
Park CK, Li HY, Yeon KY, Jung SJ, Choi SY, Lee SJ, Lee S,
Park K, Kim JS, Oh SB (2006) Eugenol inhibits sodium
currents in dental afferent neurons. J Dent Res 85:900–904
Pickering AD, Pottinger TG (1989) Stress responses and disease
resistance in salmonid fish - effects of chronic elevation of
plasma cortisol. Fish Physiol Biochem 7:253–258
Pottinger TG, Moran TA (1993) Differences in plasma cortisol
and cortisone dynamics during stress in two strains of
rainbow trout (Oncorhynchus mykiss). J Fish Biol 43:
121–130
Pottinger TG, Moran TA, Morgan JAW (1994) Primary and
secondary indices of stress in the progeny of rainbow trout
(Oncorhynchus mykiss) selected for high and low
responsiveness to stress. J Fish Biol 44:149–163
Rang HP, Dale MM, Ritter JM, Moore PK (2003) Pharma-
cology. Churchill Livingstone, London
Reid SG, Bernier NJ, Perry SF (1998) The adrenergic stress
response in fish: control of catecholamine storage and
release. Comp Biochem Physiol C 120:1–27
Ross LG, Ross B (2008) Anaesthetic and sedative techniques
for aquatic animals. Blackwell Publishing, Oxford
Rusch D, Zhong HJ, Forman SA (2004) Gating allosterism at a
single class of etomidate sites on a1b2c2L GABAA
receptors accounts for both direct activation and agonist
modulation. J Biol Chem 279:20982–20992
Sandodden R, Finstad B, Iversen M (2001) Transport stress in
Atlantic salmon (Salmo salar L.): anaesthesia and recov-
ery. Aquac Res 32:87–90
Scott AP, Ellis T (2007) Measurement of fish steroids in
water—a review. Gen Comp Endocrinol 153:392–400
Scott AP, Hirschenhauser K, Bender N, Oliveira R, Earley RL,
Sebire M, Ellis T, Pavlidis M, Hubbard PC, Huertas M,
Canario A (2008) Non-invasive measurement of steroids
in fish-holding water: important considerations when
applying the procedure to behaviour studies. Behaviour
145:1307–1328
Sink TD, Strange RJ, Sawyers RE (2007) Clove oil used at
lower concentrations is less effective than MS-222 at
reducing cortisol stress responses in anesthetized rainbow
trout. N Am J Fish Manage 27:156–161
Small BC (2003) Anesthetic efficacy of metomidate and
comparison of plasma cortisol responses to tricaine
methanesulfonate, quinaldine and clove oil anesthetized
channel catfish Ictalurus punctatus. Aquaculture 218:
177–185
Small BC (2004) Effect of isoeugenol sedation on plasma
cortisol, glucose, and lactate dynamics in channel catfish
Ictalurus punctatus exposed to three stressors. Aquacul-
ture 238:469–481
Small BC, Chatakondi N (2005) Routine measures of stress are
reduced in mature channel catfish during and after AQUI-
S anesthesia and recovery. N Am J Aquac 67:72–78
Sorensen PW, Scott AP (1994) The evolution of hormonal sex-
pheromones in teleost fish–poor correlation between the
pattern of steroid release by goldfish and olfactory sen-
sitivity suggests that these cues evolved as a result of
chemical spying rather than signal specialization. Acta
Physiol Scand 152:191–205
Sumpter JP, Dye HM, Benfey TJ (1986) The effects of stress
on plasma ACTH, a-MSH, and cortisol levels in salmonid
fishes. Gen Comp Endocrinol 62:377–385
Thomas P, Robertson L (1991) Plasma-cortisol and glucose
stress responses of red drum (Sciaenops ocellatus) to
handling and shallow-water stressors and anesthesia with
MS-222, quinaldine sulfate and metomidate. Aquaculture
96:69–86
Wendelaar Bonga SE (1997) The stress response in fish.
Physiol Rev 77:591–625
Wie MB, Won MH, Lee KH, Shin JH, Lee JC, Suh HW, Song
DK, Kim YH (1997) Eugenol protects neuronal cells from
Fish Physiol Biochem (2010) 36:719–730 729
123

excitotoxic and oxidative injury in primary cortical cul-
tures. Neurosci Lett 225:93–96
Yang J, Uchida I (1996) Mechanisms of etomidate potentiation
of GABAA receptor-gated currents in cultured postnatal
hippocampal neurons. Neuroscience 73:69–78
Zeller A, Arras M, Lazaris A, Jurd R, Rudolph U (2005)
Distinct molecular targets for the central respiratory and
cardiac actions of the general anesthetics etomidate and
propofol. FASEB J 19:1677–1679
730 Fish Physiol Biochem (2010) 36:719–730
123





























