Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Anti-Apoptotic Pro-Survival Effect of Clotrimazole in a Normothermic
Ischemia Reperfusion Injury Animal Model
Antonio Iannelli, M.D., Ph.D.,*,†,‡,1,2 George de Sousa, Ph.D.,§,2 Nathalie Zucchini, Ph.D.,§Marie Christine Saint-Paul, M.D.,* Jean Gugenheim, M.D., Ph.D.,* and Roger Rahmani, Ph.D.§
*Service de Chirurgie Digestive et Transplantation Hepatique, Hopital Archet 2, Nice, France; †Centre Hospitalier Universitaire de Nice,Pole digestif, Nice, France; ‡Universite de Nice-Sophia-Antipolis, Faculte de Medecine, Nice, France; and §Laboratoire de Toxicologie
Moleculaire, Cellulaire et Genomique, Sophia Antipolis, INRA, UMR 1089 France
Submitted for publication November 27, 2009
Background. Increasing evidence suggests that ap-optosis plays a critical role in ischemia reperfusion(IR)-mediated liver injury. Clotrimazole (CTZ) is a po-tent antimycotic drug that also has a free radical scav-enger activity. This study investigated the possibleanti-apoptotic, pro-survival role of CTZ in hepatic IRinjury in rats.
Methods. Male Sprague-Dawley rats were dividedinto three groups: sham, control, and CTZ-treated(n [ 10 each). Control and CTZ-treated animals weresubjected to 60 min of normothermic ischemia of theleft lateral lobe of the liver followed by 6 h of reperfu-sion. Animals were then sacrificed, the liver excised,and blood samples collected.
Results. CTZ induced a significant increase in ex-pression of anti-apoptotic Bcl-xL protein. Serum levelsof aspartate transaminase and alanine transaminasewere significantly lower in CTZ-treated animals thanin controls. Histopathologically, tissue damage in theform of apoptosis was significantly lower in CTZ-treated animals than in controls. Expression of the ac-tivated form of caspase-3 and the cleaved form of itssubstrate, poly-ADP-ribose polymerase, decreased sig-nificantly in the CTZ-treated group compared withcontrols. CTZ increased the expression of phospho-p44/42 ERK1/2 and decreased the phosphorylated formof JNK, without affecting p38 MAPK.
Conclusion. CTZ protects the liver against IR apo-ptosis in rats through overexpression of the anti-apoptotic protein Bcl-xL. Other pro-survival pathwayssuch as phospho-p 44/42 ERK1/2 kinase are also
activated while JNK is down-regulated. Crown
Copyright � 2011 Published by Elsevier Inc. All rights reserved.
Key Words: apoptosis; ischemia; reperfusion; preg-nane X receptor; clotrimazole; free radicals.
INTRODUCTION
Liver ischemia-reperfusion (IR) injury can occur inseveral clinical conditions, including liver trauma,transplantation, and hepatectomy [1]. The inflamma-tory response that follows IR and leads to organdysfunction is caused by hepatocyte necrosis and apo-ptosis [1]. While necrosis has classically been consid-ered to be the sole mode of cell death induced by IR,there is increasing evidence that apoptosis, or pro-grammed cell death, also plays a critical role [1-5].Apoptosis can be triggered by various extracellular(extrinsic apoptosis) or intracellular (intrinsicapoptosis) stimuli, such as binding to cell surfacereceptors or treatment with cytotoxic drugs,respectively [6]. Whatever the stimulus, effective apo-ptotic cell death is often due to caspases. Once initiatorcaspases (caspase-8 and caspase-9 for extrinsic or in-trinsic pathways, respectively) have been activated,they will proteolytically activate effector pro-caspase-3, -6, and -7, responsible for the cleavage of vital cellcomponents, and hence for the execution of cell death.This cell suicide is mainly regulated by the Bcl-2 familyconsisting of anti- (e.g., Bcl-2, Bcl-xL) and pro- (e.g.,Bax, Bak) apoptotic members. These proteins playa key role in apoptotic pathway regulation by decidingwhether a cell lives or dies [7].
Clotrimazole (1-(2-chloro-phenyl) diphenylmethyl-1H-imidazole, CAS-Reg.-No. 23593-75-1) (CTZ) is
1 To whom correspondence and reprint requests should be ad-dressed at Service de Chirurgie Digestive et Transplantation Hepa-tique, Hopital Archet 2, 151 Route Saint Antoine de Ginestiere BP3079, Nice Cedex 3, France. E-mail: [email protected].
2 These authors contributed equally to this study.
0022-4804/$36.00Crown Copyright � 2011 Published by Elsevier Inc. All rights reserved.
101
Journal of Surgical Research 171, 101–107 (2011)doi:10.1016/j.jss.2010.03.035

Author's personal copy
a common pharmaceutical in human and veterinarymedicine for the treatment of fungal infections. It hasalso been discussed for the treatment of sickle cell dis-ease, rheumatoid arthritis, malaria [8], and tuberculo-sis [9]; derivatives of clotrimazole are investigated asimmunosuppressants [10]. CTZ belongs to the largeand structurally diverse group of sazole fungicidesthat block sterol synthesis by inhibiting cytochromeP450-dependent sterol 14-demethylases [11]. Previousin vitro studies from our laboratory showed that CTZprotects hepatocytes against staurosporine-inducedapoptosis [12–14]. Here we describe the effect of CTZin an in vivo model of liver apoptosis (normothermicIR of rat liver).
This report demonstrates that CTZ is capable of pro-tecting the rat liver against normothermic IR injury.The mechanism involves inhibition of caspase-like ac-tivities, up-regulation of the anti-apoptotic proteinBclxL, together with activation of the MAPK kinaseERK1/2 pathway and concomitant inhibition of thec-jun N-terminal kinase (JNK) pathway.
MATERIALS AND METHODS
Animals
All experiments were performed according to the institutionalguidelines for the care and use of laboratory animals. Age-matchedmale Sprague Dawley rats weighting between 250 and 300 g (CharlesLaboratories, L’Arbresle, France) were used. Animals were housed inindividual Plexiglas cages with free access to water and food (before,during and after the I/R procedure). Light (from 8 am to 7 pm) androom temperature (21 6 2 �C) were kept constant.
Experimental Design and Surgical Procedure
Animals were divided into three groups (10 animals/group). Ani-mals in the CTZ-treated group were injected intraperitoneally with50 mg/kg CTZ daily (Sigma, l’Isle d’Abeau Chesnes, France) dilutedin 1 mL of corn oil (Sigma) for 3 d before the I/R procedure. Animalsin the control group (OIL) were injected intraperitoneally with a dailydose of 1 mL corn oil alone for 3 d before the IR procedure. Sham an-imals (SHAM) received no treatment. Animals, under light ether an-esthesia, were placed on an ad hoc operating table and the abdomenwas prepared with iodine-povidone solution (Betadine; Astra Medica,Merignac, France). A midline laparotomy was made and the abdomi-nal cavity was explored to eliminate any complications related to pre-vious intraperitoneal injections. The vascular and biliary pedicle tothe left lobe of the liver (representing 30% of the whole liver) wascross-clamped with a nontraumatic vascular clamp (Moria, ref 201/A; Antony, France). The abdominal wall was then closed with onelayer of running sutures. After 1 h of ischemia, the abdominal sutureswere removed, and the clamp released to allow reperfusion of the is-chemic left liver lobe. Six hours later, animals were sacrificed by ex-sanguination and blood samples collected from the inferior venacava for serum transaminase determination. The liver was retrievedand washed in phosphate-buffered saline (PBS). The left liver lobewas separated and samples snap frozen in liquid nitrogen. Sham an-imals were laparotomized for 5 min without vascular clamping.
Western Blotting
Liver samples (100 mg) were homogenized mechanically in 1 mL ofhypotonic buffer (HEPES pH 7.5: 25 mM MgCl2, 5 mM DTT, 5 mMPMSF, 2 mM pepstatin A, 10 mg/mL leupeptin, 5 mM EDTA, 10 mg/mL aprotinin, anti-phosphatases; Roche, Meylan, France). Protein con-centration was determined with a BCA protein assay kit according tothe manufacturer’s instructions (Pierce, Brebieres, France). Sampleswere resolved by 12% SDS-PAGE and transferred to a polyvinylfluoridemembrane (Millipore, Molsheim, France). Membranes were blockedwith 5% non-fat skimmed milk in TBS-T (10 mM Tris pH 7.5, 140mM NaCl, 0.1% Tween 20) for 1 h at 37 �C, washed and incubatedwith the corresponding primary antibody in TBS-T containing 3% bo-vine serum albumin. Membranes were then incubated for 12 h withthe appropriate antibody diluted 1:1000 in TBS:rabbit anti-Bcl-xL(Cell Signaling Technology, Danvers, MA); rabbit anti P6 ERK1/2(Cell Signaling Technology); rabbit anti-CYP3A (Oxford, USA); mousemonoclonal anti-PARP (Becton-Dickinson, Le pont de Claix, France);rabbit anti-caspase-3 p17-19 (Cell Signaling Technology), rabbit antiP-p38 (Cell Signaling Technology) and rabbit anti-JNK (Cell SignalingTechnology). The membranes were washed extensively in TBS. Detec-tion was carried out using horseradish peroxide-conjugated anti-rabbitor anti-mouse antibodies and enhanced chemiluminescence reagents;the latter were used according to the manufacturer’s instructions(Immobilon Western; Millipore). The signal was recorded with a CCDcamera (Chemimager 2; Syngene, Cambridge, UK), and analyzedwith Gene Tools software (Syngene).
Histopathologic Assessment of Liver Apoptosis
Formalin-fixed hepatic tissue samples were embedded in paraffinand 3 mm sections cut. Serial sections were stained with hematoxylinand eosin (H and E) to reveal histologic evidence of apoptosis: cellshrinkage, chromatin condensation (nuclear condensation and chro-matin fragmentation), and cell disintegration into small apoptoticbodies. Results are expressed as percentages and represent themean number of apoptotic or steatosic hepatocytes per high powerfield (hpf) (400 cells/hpf; 10 hpf/examined section). Histologic exami-nation was performed in a blinded fashion by the same pathologist.
Measurement of Plasma Alanine Aminotransferase (ALT) and
Aspartate Aminotransferase (AST)
Blood sample were centrifuged at 3000 rpm for 15 min at a roomtemperature. ALT and AST plasma concentrations were measuredwith an Olympus AU5400 (Olympus France, Rungis, France) auto-matic analyzer. Both values were expressed as U/l.
Statistics
Results are presented as means 6 standard deviation. The non-parametric Mann-Whitney U test was used to compare mean values.P < 0.05 was considered to be statistically significant.
RESULTS
CTZ Protection Against Normothermic IR-Induced Apoptosis
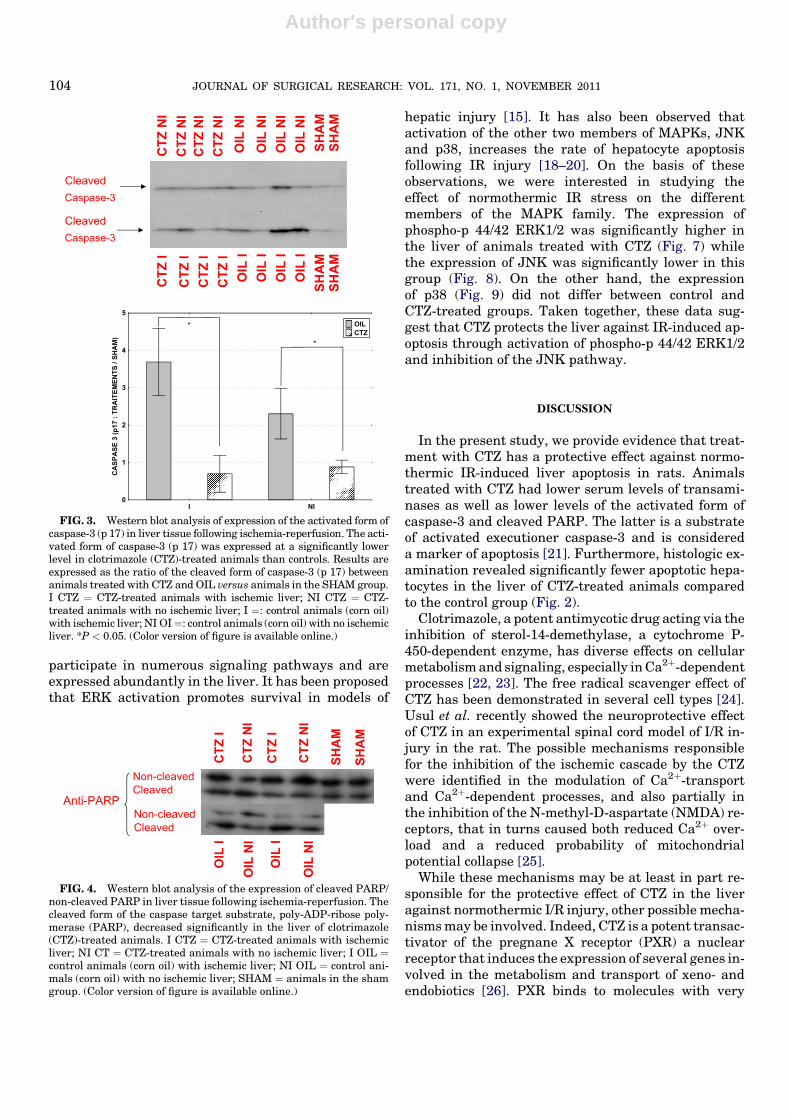
The group of animals treated with CTZ displayedsignificantly lower serum levels of AST and ALT(Fig. 1) and a significantly lower number of apoptotichepatocytes, compared with control animals (Fig. 2).CTZ treatment was associated with decreased expres-sion of the activated form of caspase-3 (Fig. 3). Accord-ingly, expression of the cleaved form of the caspase
JOURNAL OF SURGICAL RESEARCH: VOL. 171, NO. 1, NOVEMBER 2011102

Author's personal copy
target substrate, poly-ADP-ribose polymerase (PARP),an important regulator of apoptosis, was significantlydecreased in the liver of CTZ-treated animals (Fig. 4).
PXR Transactivation and Bcl-xL Expression
In order to assess PXR transactivation in the mecha-nism of hepatic protection, we investigated the expres-sion of CYP3A1, a known PXR target gene. CYP3A1was expressed in animals treated with CTZ, but notin control animals (Fig. 5). This result indicates thatCTZ induced PXR transactivation. Along with PXRtransactivation, CTZ also induced a significant increasein the expression of anti-apoptotic Bcl-xL protein(Fig. 6).
MAPKs (Extracellular Signal-Regulated Kinases) Phospho-p
44/42 ERK1/2, p38, and JNK
It has been reported that IR might modulate mitogen-activated protein kinase (MAPK) activity in rat liverand hepatocyte cell cultures after IR injury [15–17].Extracellular signal-regulated kinases (ERK1/2)
SHAM
TREATMENT
0
200
400
600
800
1000
1200
1400
1600
1800
2000
U/L
ALT
AST
CTZOIL
FIG. 1. Serum levels of AST and ALT following ischemia-reperfusion of the left lobe of theliver. Animals in the CTZ-treatedgroup showed significantly lower serum levels of AST and ALT.AS ¼ aspartate aminotransferase; ALT ¼ alanine aminotransferase;CTZ ¼ clotrimazole-treated animals; OI ¼ control animals (corn oil);SHAM ¼ animals in the sham group.
I CTZ NI CTZ I OIL NI OIL SHAM
TREATMENTS
0
2
4
6
8
10
12
14
16
18
% o
f A
po
pto
sis
*
* * BA
DC
FIG. 2. Histopathologic findings following ischemia-reperfusion of the left lobe of the liver (H and E). (A) Significantly fewer apoptotic he-patocytes were observed in clotrimazole (CTZ)-treated animals than in controls. Results are expressed in percentages as the mean number ofapoptotic cells per high power field (hpf) (400 cells/hpf; five hpf/examined section). I CTZ: CTZ-treated animals with ischemic liver; NI CTZ:CTZ-treated animals with no ischemic liver; I OIL ¼ control animals (corn oil) with ischemic liver; NI OIL ¼ control animals (corn oil) withno ischemic liver; SHAM¼ animals in the sham group. *P< 0.05. (B) (H and E) I CTZ. (C) (H and E) I OIL. (D) (H and E) SHAM. Arrows indicateapoptotic hepatocytes. (Color version of figure is available online.)
IANNELLI ET AL.: CLOTRIMAZOLE PROTECTS THE LIVER AGAINST ISCHEMIA-REPERFUSION INJURY 103
Author's personal copy
participate in numerous signaling pathways and areexpressed abundantly in the liver. It has been proposedthat ERK activation promotes survival in models of
hepatic injury [15]. It has also been observed thatactivation of the other two members of MAPKs, JNKand p38, increases the rate of hepatocyte apoptosisfollowing IR injury [18–20]. On the basis of theseobservations, we were interested in studying theeffect of normothermic IR stress on the differentmembers of the MAPK family. The expression ofphospho-p 44/42 ERK1/2 was significantly higher inthe liver of animals treated with CTZ (Fig. 7) whilethe expression of JNK was significantly lower in thisgroup (Fig. 8). On the other hand, the expressionof p38 (Fig. 9) did not differ between control andCTZ-treated groups. Taken together, these data sug-gest that CTZ protects the liver against IR-induced ap-optosis through activation of phospho-p 44/42 ERK1/2and inhibition of the JNK pathway.
DISCUSSION
In the present study, we provide evidence that treat-ment with CTZ has a protective effect against normo-thermic IR-induced liver apoptosis in rats. Animalstreated with CTZ had lower serum levels of transami-nases as well as lower levels of the activated form ofcaspase-3 and cleaved PARP. The latter is a substrateof activated executioner caspase-3 and is considereda marker of apoptosis [21]. Furthermore, histologic ex-amination revealed significantly fewer apoptotic hepa-tocytes in the liver of CTZ-treated animals comparedto the control group (Fig. 2).
Clotrimazole, a potent antimycotic drug acting via theinhibition of sterol-14-demethylase, a cytochrome P-450-dependent enzyme, has diverse effects on cellularmetabolism and signaling, especially in Ca2þ-dependentprocesses [22, 23]. The free radical scavenger effect ofCTZ has been demonstrated in several cell types [24].Usul et al. recently showed the neuroprotective effectof CTZ in an experimental spinal cord model of I/R in-jury in the rat. The possible mechanisms responsiblefor the inhibition of the ischemic cascade by the CTZwere identified in the modulation of Ca2þ-transportand Ca2þ-dependent processes, and also partially inthe inhibition of the N-methyl-D-aspartate (NMDA) re-ceptors, that in turns caused both reduced Ca2þ over-load and a reduced probability of mitochondrialpotential collapse [25].
While these mechanisms may be at least in part re-sponsible for the protective effect of CTZ in the liveragainst normothermic I/R injury, other possible mecha-nisms may be involved. Indeed, CTZ is a potent transac-tivator of the pregnane X receptor (PXR) a nuclearreceptor that induces the expression of several genes in-volved in the metabolism and transport of xeno- andendobiotics [26]. PXR binds to molecules with very
Cleaved
CT
Z I
OIL
I
Anti-PARP
SH
AM
Non-cleaved
CleavedNon-cleaved
CT
Z I
CT
Z N
I
CT
Z N
I
OIL
I
OIL
N
I
OIL
N
I
SH
AM
FIG. 4. Western blot analysis of the expression of cleaved PARP/non-cleaved PARP in liver tissue following ischemia-reperfusion. Thecleaved form of the caspase target substrate, poly-ADP-ribose poly-merase (PARP), decreased significantly in the liver of clotrimazole(CTZ)-treated animals. I CTZ ¼ CTZ-treated animals with ischemicliver; NI CT ¼ CTZ-treated animals with no ischemic liver; I OIL ¼control animals (corn oil) with ischemic liver; NI OIL ¼ control ani-mals (corn oil) with no ischemic liver; SHAM ¼ animals in the shamgroup. (Color version of figure is available online.)
INI
0
1
2
3
4
5
CA
SP
AS
E 3 (p
17 : T
RA
IT
EM
EN
TS
/ S
HA
M)
OIL
CTZ
*
*
CT
Z N
I
OIL
N
I
OIL
I
CleavedCaspase-3
SH
AM
CleavedCaspase-3
CT
Z N
I
CT
Z N
I
CT
Z N
I
OIL
N
I
OIL
N
I
OIL
N
I
SH
AM
SH
AM
SH
AM
OIL
I
OIL
I
OIL
I
CT
Z I
CT
Z I
CT
Z I
CT
Z I
FIG. 3. Western blot analysis of expression of the activated form ofcaspase-3 (p 17) in liver tissue following ischemia-reperfusion. The acti-vated form of caspase-3 (p 17) was expressed at a significantly lowerlevel in clotrimazole (CTZ)-treated animals than controls. Results areexpressed as the ratio of the cleaved form of caspase-3 (p 17) betweenanimals treated with CTZ and OIL versus animals in the SHAM group.I CTZ ¼ CTZ-treated animals with ischemic liver; NI CTZ ¼ CTZ-treated animals with no ischemic liver; I ¼: control animals (corn oil)with ischemic liver; NI OI¼: control animals (corn oil) with no ischemicliver. *P < 0.05. (Color version of figure is available online.)
JOURNAL OF SURGICAL RESEARCH: VOL. 171, NO. 1, NOVEMBER 2011104

Author's personal copy
different molecular structures and is currently consid-ered as a xenosensor, playing a pivotal role in the pro-tection of cells against xeno- and endobiotics [26].Previous in vitro studies by our group have provided ev-idence that the transactivation of the PXR is also asso-
ciated with the regulation of apoptosis. In fact, PXRtransactivation diminishes the rate of spontaneous orprovoked apoptosis by inducing the anti-apoptotic pro-teins Bcl-2 and Bcl-xL in human and rat hepatocytes,respectively [12–14]. Protection of hepatocytes,through the modulation of apoptosis, may be part ofa defense strategy orchestrated by PXR against celldeath induced by endo- and xenobiotics.
In the present study, CTZ treatment effectively acti-vated PXR, as shown by the increased expression ofCYP3A, a known target gene of PXR and an indicatorof PXR transactivation [27]. PXR transactivation wasassociated with a significant increase in the expressionof the anti-apoptotic protein BclxL. These data concurwith previous results from our laboratory indicatingthat PXR transactivation induces a transient increase(lasting 24 h) in expression of the Bcl-xL gene and, inparallel, an increase in level of protein expression[14]. Once transactivated with CTZ in vivo, PXR pro-tects hepatocytes exposed to a pro-apoptotic stimulus(such as normothermic IR) through the induction ofBcl-xL protein expression.
CYP3A1
CT
Z I
OIL
N
IC
TZ
N
I
CT
Z N
I
CT
Z N
I
CT
Z N
I
CT
Z I
CT
Z I
*C
TZ
I
*C
TZ
I
OIL
I
OIL
N
I
OIL
N
I
OIL
N
I
OIL
N
I
OIL
I
OIL
I
CYP3A1
FIG. 5. Western blot analysis of CYP3A1 expression in liver tis-sue following ischemia reperfusion of the left lobe. Clotrimazole(CTZ) induced the expression of CYP3A1, a known PXR target gene,in the liver, while no expression of CYP3A1 was detected in the liverof control animals. I CTZ ¼ CTZ-treated animals with ischemic liver;NI CT¼CTZ-treated animals with no ischemic liver; I O¼ control an-imals (corn oil) with ischemic liver; NI OIL¼ control animals (corn oil)with no ischemic liver. (Color version of figure is available online.)
NI CTZI CTZNI OILI OIL
TREATMENT
0
1E5
2E5
3E5
4E5
5E5
CH
EM
IL
UM
IN
ES
CE
NT
IN
TE
NS
IT
Y
(arb
itrary u
nits)
*
OIL
N
I
Bcl-xL
Bcl-xL
SHAM
OIL
I
OIL
N
I
OIL
N
I
OIL
N
I
OIL
I
OIL
I
OIL
I
CT
Z I
CT
Z I
CT
Z I
CT
Z I
SHAM
SHAM
SHAM
CT
Z N
I
CT
Z N
I
CT
Z N
I
CT
Z N
I
FIG. 6. Western blot analysis of Bcl-xL expression in liver tissuefollowing ischemia reperfusion of the left lobe. Clotrimazole (CTZ)treatment was associated with a significant increase in expressionof antiapoptotic pro-survival Bcl-xL protein compared with thecontrol groups. Results are expressed as the ratio of Bcl-xL betweenanimals treated with CTZ and OIL versus animals in the SHAMgroup. I CTZ ¼ CTZ-treated animals with ischemic liver; NI CTZ ¼CTZ-treated¼ control animals (corn oil) with no ischemic liver. (Colorversion of figure is available online.)
SHAM I CTZ NI CTZ I OIL NI OIL
TREATMENT
0
20000
40000
60000
80000
1E5
1,2E5
1,4E5
1,6E5
1,8E5
2E5
2,2E5
2,4E5
CH
EM
IL
UM
IN
ES
CE
NT
IN
TE
NS
IT
Y
(a
rb
itra
ry
u
nits
)
SH
AM
CT
Z I
P-ERK 1/2
P-ERK 1/2
OIL
I
OIL
I
OIL
I
OIL
I
OIL
N
I
OIL
N
I
OIL
N
I
OIL
N
I
CT
Z I
CT
Z I
CT
Z I
CT
Z N
I
CT
Z N
I
CT
Z N
I
CT
Z N
I
SH
AM
FIG. 7. Western blot analysis of ischemia-reperfusion (IR)-induced phosphorylation of P- 42/44 ERK in liver tissue. Clotrimazole(CTZ)-treated animals showed a significant increase in expression ofthe antiapoptotic pro-survival P-42/44 ERK compared with controlanimals. I CTZ ¼ CTZ-treated animals with ischemic liver; NI C ¼CTZ-treated animals with no ischemic liver; I OIL ¼ control animals(corn oil) with ischemic liver; NI OIL¼ control animals (corn oil) withno ischemic liver; SHAM ¼ animals in the sham group. *P < 0.05.(Color version of figure is available online.)
IANNELLI ET AL.: CLOTRIMAZOLE PROTECTS THE LIVER AGAINST ISCHEMIA-REPERFUSION INJURY 105

Author's personal copy
We also investigated the effect of CTZ on the regula-tion of MAPKs (p-38, JNK, ERK1/2), which are all acti-vated during IR of the liver [15–20]. Activation of p-38and JNK has been associated with a pro-apoptotic ef-fect. Kobayashi et al. showed the protective effect of in-hibition of p-38 on IR-induced liver apoptosis in the rat[18]. Yoshinari observed activation of JNK and p-38 inthe liver following IR and demonstrated the protectiveeffect of inhibition of p-38 following addition ofFR167653 to the University of Wisconsin solution[19]. In the present study, there was no difference inthe expression phosphorylation of p-38 between CTZ-treated animals and controls. On the other hand,a significant increase in expression phosphorylation ofP-ERK1/2 and inhibition of activation of JNK wereobserved following CTZ treatment. Cursio et al. ob-served that the ERK1/2 pathway is activated duringliver ischemia, remains activated during reperfusionfor 3 and 6 h, and is associated with an antiapoptoticpro-survival effect against IR-induced liver apoptosis
[20]. In an experimental model of reduced size livertransplantation, Liang et al. also found that activationof the ERK1/2 pathway is associated with significantlyless apoptosis in the liver [28]. ERK1/2 activation in-hibits apoptosis through a conformational change inBax required for its translocation to the mitochondria[29] and caspase-3 activation [30]. JNK is known to bestimulated in vivo after rat liver transplantation or par-tial hepatectomy [15, 16]. Crenesse et al. demonstratedthat warm hypoxia-reoxygenation is able to stimulateJNK in cultured rat hepatocytes and that this kinaseis involved in apoptosis [17]. Cursio et al. demonstratedthe pro-apoptotic effect of JNK in IR-induced liver apo-ptosis in the rat [20]. Taken together, these results sug-gest a broad role for PXR in protection of the cell, notonly through modulation of metabolism and transportof xeno- and endobiotics, but also by protecting thecell against apoptosis. In conclusion, overexpression ofthe anti-apoptotic protein Bcl-xL following transactiva-tion of PXR in hepatocytes in vitro can also be observedin the liver in vivo. Other pro-survival pathways suchas ERK1/2 kinase are activated by the inductionof PXR.
CT
Z N
IO
IL
N
I
SH
AM
SHAM I CTZ NI CTZ I OIL NI OIL
TREATMENT
0
20000
40000
60000
80000
1E5
CH
EM
IL
UM
IN
ES
CE
NT
IN
TE
NS
IT
Y
(a
rb
itra
ry
u
nits
)
* *
JNK
JNK
OIL
N
I
OIL
N
I
OIL
N
I
OIL
I
OIL
I
OIL
I
CT
Z N
I
CT
Z N
I
CT
Z N
I
CT
Z I
CT
Z I
CT
Z I
SH
AM
FIG. 8. Western blot analysis of ischemia-reperfusion (IR)–induced phosphorylation of c-jun N-terminal kinase (JNK) in livertissue. Clotrimazole (CTZ)-treated animals showed a significant de-crease in expression of the phosphorylated form of pro-apoptotic JNKcompared with control animals. I CTZ ¼ CTZ-treated animals with is-chemic liver; NI CTZ ¼ CTZ-treated animals with no ischemic liver; IOIL ¼ control animals (corn oil) with ischemic liver; NI OIL ¼ controlanimals (corn oil) with no ischemic liver; SHAM¼ animals in the shamgroup. *P < 0.05. (Color version of figure is available online.)
SHAM I CTZ NI CTZ I OIL NI OIL
TREATMENT
0
10000
20000
30000
40000
50000
60000
70000
80000
CH
EM
IL
UM
IN
ES
CE
NT
IN
TE
NS
IT
Y
(arb
itrary u
nits)
CT
Z I
SH
AM
p38
p38
CT
Z I
CT
Z I
CT
Z I
CT
Z N
I
CT
Z N
I
CT
Z N
I
CT
Z N
I
OIL
I
OIL
I
OIL
I
OIL
I
OIL
N
I
OIL
N
I
OIL
N
I
OIL
N
I
SH
AM
FIG. 9. Western blot analysis of ischemia-reperfusion (IR)-induced phosphorylation of p38 MAPK in liver tissue. There was nosignificant difference in the expression of the phosphorylated pro-apoptotic form of p38 MAPK between CTZ-treated animals and con-trol animals. I CTZ ¼ CTZ-treated animals with ischemic liver; NICTZ ¼ CTZ-treated animals with no ischemic liver; I OIL ¼ controlanimals (corn oil) with ischemic liver; NI OIL ¼ control animals(corn oil) with no ischemic liver; SHAM¼ animals in the sham group.*P < 0.05. (Color version of figure is available online.)
JOURNAL OF SURGICAL RESEARCH: VOL. 171, NO. 1, NOVEMBER 2011106

Author's personal copy
ACKNOWLEDGMENTS
The authors thank Dr. Floch and 193 Newmed Publishing Servicesfor the correction of the proofs and O. Migliore for his assistance.
REFERENCES
1. Montalvo-Jave EE, Escalante-Tattersfield T, et al. Factors in thepathophysiology of the liver ischemia-reperfusion injury. J SurgRes 2008;147:153.
2. Cursio R, Gugenheim J, Ricci JE, et al. A caspase inhibitor fullyprotects rats against lethal normothermic liver ischemia by inhi-bition of liver apoptosis. FASEB J 1999;13:253.
3. Honda K, Tohyama T, Kotegawa H, et al. Protective effect ofadeno-mediated human Bcl-xL gene transfer to the mouse liverin a partial ischemia/reperfusion model. J Surg Res 2009;157:e107.
4. Baskin-Bey ES, Washburn K, Feng S, et al. J. Clinical trial of thepan-caspase inhibitor, IDN-6556, in human liver preservationinjury. Am J Transplant 2007;7:218.
5. Harada N, Hatano E, Koizumi N, et al. Akt activation protects ratliver from ischemia/reperfusion injury. J Surg Res 2004;121:159.
6. Sprick MR, Walczak H. The interplay between the Bcl-2 familyand death receptor-mediated apoptosis. Biochim Biophys Acta2004;1644:125.
7. Korsmeyer SJ. BCL-2 gene family and the regulation of pro-grammed cell death. Cancer Res 1999;59:1693s.
8. Tiffert T, Ginsburg H, Krugliak M, et al. Potent antimalarial ac-tivity of clotrimazole in in vitro cultures of Plasmodium falcipa-rum. Proc Nat Acad Sci USA 2000;97:331.
9. McLean KJ, Marshall KR, Richmond A, et al. Azole antifungalsare potent inhibitors of cytochrome P450 mono-oxygenases andbacterial growth in mycobacteria and streptomycetes. Microbiol-ogy (Reading, England) 2002;148:2937.
10. Wulff H, Miller MJ, Hansel W, et al. Design of a potent and se-lective inhibitor of the intermediate-conductance Ca2þ-acti-vated Kþ channel, IKCa1: A potential immunosuppressant.Proc Nat Acad Sci USA 2000;97:8151.
11. Sheehan DJ, Hitchcock CA, Sibley CM. Current and emergingazole antifungal agents. Clin Microbiol Rev 1999;12:40.
12. Bailly-Maitre B, de Sousa G, Boulukos K, et al. Dexamethasoneinhibits spontaneous apoptosis in primary cultures of humanand rat hepatocytes via Bcl-2 and Bcl-xL induction. Cell DeathDiffer 2001;8:279.
13. Bailly-Maitre B, de Sousa G, Zucchini N, et al. Spontaneous ap-optosis in primary cultures of human and rat hepatocytes: Mo-lecular mechanisms and regulation by dexamethasone. CellDeath Differ 2002;9:945.
14. Zucchini N, de Sousa G, Bailly-Maitre B, et al. R. Regulation ofBcl-2 and Bcl-xL anti-apoptotic protein expression by nuclear re-ceptor PXR in primary cultures of human and rat hepatocytes.Biochim Biophys Acta 2005;1745:48.
15. Bradham CA, Stachlewitz RF, Gao W, et al. Reperfusion afterliver transplantation in rats differentially activates themitogen-activated protein kinases. Hepatology (Baltimore,MD) 1997;25:1128.
16. Westwick JK, Weitzel C, Leffert HL, et al. Activation of Jun ki-nase is an early event in hepatic regeneration. J Clin Invest1995;95:803.
17. Crenesse D, Schmid-Alliana A, Laurens M, et al. JNK(1)/SAPK(1) involvement in hypoxia-reoxygenation-induced apo-ptosis in rat hepatocytes. Transplant Proc 2001;33:260.
18. Kobayashi M, Takeyoshi I, Yoshinari D, et al. P38 mitogen-activated protein kinase inhibition attenuates ischemia-reperfusion injury of the rat liver. Surgery 2002;131:344.
19. Yoshinari D, Takeyoshi I, Kobayashi M, et al. Effects of a p38mitogen-activated protein kinase inhibitor as an additive to Uni-versity of Wisconsin solution on reperfusion injury in liver trans-plantation. Transplantation 2001;72:22.
20. Cursio R, Filippa N, Miele C, et al. Involvement of protein kinaseB and mitogen-activated protein kinases in experimental normo-thermic liver ischemia-reperfusion injury. Br J Surg 2006;93:752.
21. Lazebnik YA, Kaufmann SH, Desnoyers S, et al. Cleavage ofpoly(ADP-ribose) polymerase by a proteinase with propertieslike ICE. Nature 1994;371:346.
22. Burdon RH, Rice-Evans C. Free radicals and the regulation ofmammalian cell proliferation. Free Rad Res Commun 1989;6:345.
23. Yoshida Y, Aoyama Y. Interaction of azole antifungal agentswith cytochrome P-45014DM purified from Saccharomyces cere-visiae microsomes. Biochem Pharmacol 1987;36:229.
24. Isaev NK, Stelmashook EV, Dirnagl U, et al. Neuroprotective ef-fects of the antifungal drug clotrimazole. Neuroscience 2002;113:47.
25. Usul H, Arslan E, Cansever T, et al. Effects of clotrimazole on ex-perimental spinal cord ischemia/reperfusion injury in rats.Spine 2008;33:2863.
26. Kakizaki S, Yamazaki Y, Takizawa D, et al. New insights on thexenobiotic-sensing nuclear receptors in liver diseases—CAR andPXR. Curr Drug Metab 2008;9:614.
27. Bertilsson G, Heidrich J, Svensson K, et al. Identification of a hu-man nuclear receptor defines a new signaling pathway forCYP3A induction. Proc Nat Acad Sci USA 1998;95:12208.
28. Liang T, Xu S, Yu J, et al. Activation pattern of mitogen-activated protein kinases in early phase of different size liver iso-grafts in rats. Liver Transpl 2005;11:1527.
29. Weston CR, Balmanno K, Chalmers C, et al. Activation of ERK1/2 by deltaRaf-1:ER* represses Bim expression independently ofthe JNK or PI3K pathways. Oncogene 2003;22:1281.
30. Terada K, Kaziro Y, Satoh T. Analysis of Ras-dependent signalsthat prevent caspase-3 activation and apoptosis induced by cyto-kine deprivation in hematopoietic cells. Biochem Biophys ResCommun 2000;267:449.
IANNELLI ET AL.: CLOTRIMAZOLE PROTECTS THE LIVER AGAINST ISCHEMIA-REPERFUSION INJURY 107





























