Embed Size (px)
Citation preview

Antidiabetic Properties of the Histamine H3 ReceptorProtean Agonist Proxyfan
Melanie B. Henry, Shuqin Zheng, Chenxia Duan, Bhuneshwari Patel,Galya Vassileva, Christopher Sondey, Jean Lachowicz, and Joyce J. Hwa
Diabetes/Obesity Research, Merck Research Laboratories, Kenilworth, New Jersey 07033
Proxyfan is a histamine H3 receptor protean agonist that can produce a spectrum of pharmaco-logicaleffects includingagonist, inverseagonist,andantagonist.Wehavediscoveredthatproxyfan(10mg/kg orally) significantly improved glucose excursion after an ip glucose tolerance test in either leanor high-fat/cholesterol diet-induced obese mice. It also reduced plasma glucose levels comparable tothat of metformin (300 mg/kg orally) in a nongenetic type 2 diabetes mouse model. The dose-depen-dent decrease in glucose excursion correlated with inhibition of ex vivo H3 receptor binding in thecerebral cortex. In addition, glucose levels were significantly reduced compared with vehicle-treatedmice after intracerebroventricular administration of proxyfan, suggesting the involvement of centralH3 receptors. Proxyfan-induced reduction of glucose excursion was not observed in the H3 receptorknockout mice, suggesting that proxyfan mediates this effect through H3 receptors. Proxyfan reducedglucose excursion by significantly increasing plasma insulin levels in a glucose-independent manner.However, no difference in insulin sensitivity was observed in proxyfan-treated mice. The H1 receptorantagonist chlorpheniramine and the H2 receptor antagonist zolantidine had modest effects onglucose excursion, and neither inhibited the glucose excursion reduced by proxyfan. The H3receptor antagonist/inverse agonist, thioperamide, had weaker effects on glucose excursioncompared with proxyfan, whereas the H3 receptor agonist imetit did not affect glucose ex-cursion. Inconclusion,thesefindingsdemonstrate,forthefirsttime,thatmanipulationofcentralhistamineH3 receptor by proxyfan can significantly improve glucose excursion by increasing plasma insulinlevels via a glucose-independent mechanism. (Endocrinology 152: 828–835, 2011)
The histamine H3 receptor is one of the four G protein-coupled receptors in the histamine receptor family (1).
The histamine receptor family has produced valuable drugtargets as demonstrated by the successful development ofH1 receptor antagonists for the treatment of allergic re-actions and the H2 receptor antagonists for the treatmentof gastric ulcer. Unlike the H1, H2, and H4 receptors, H3receptors are highly expressed in the central nervous sys-tem and play a regulatory role in the presynaptic release ofhistamine and other neurotransmitters (2). Histamine H3receptors are also highly expressed in the retina, located inthe postsynaptic membrane of ON-bipolar cell tip (3). Al-though the histamine H3 receptor has been implicated inmany important physiological processes like the sleep-
wake cycle, ingestive behavior, and cognition (1), few re-ports mention the direct involvement of the H3 receptor ininsulin secretion.
Mice lacking the histamine H3 receptor (Hrh3�/�)have shown a significant decrease in overall locomotion,wheel-running activity, and body temperature phenotype,leading to decreased energy expenditure and mild obesity inaged mice (2, 4). The mildly obese H3 receptor knockoutmice also demonstrate leptin and insulin resistance with im-pairedglucose tolerance (4);however, it isunknownwhetherthis is secondary to their obese phenotype. It is not clear fromthe literature whether acute manipulation of histamine H3receptors by an H3 receptor agonist or antagonist may haveany direct effect on glucose excursion in mice.
ISSN Print 0013-7227 ISSN Online 1945-7170Printed in U.S.A.Copyright © 2011 by The Endocrine Societydoi: 10.1210/en.2010-0757 Received July 6, 2010. Accepted December 9, 2010.First Published Online January 14, 2011
Abbreviations: AUC, Area under the curve; DIO, diet-induced obese; ES, embryonic stem;GTT, glucose tolerance test; ICV, intracerebroventricular; ITT, insulin tolerance test; MC,methylcellulose; NFG, nonfasting glucose; po, orally; STZ, streptozotocin.
D I A B E T E S - I N S U L I N - G L U C A G O N - G A S T R O I N T E S T I N A L
828 endo.endojournals.org Endocrinology, March 2011, 152(3):828–835
Dow
nloaded from https://academ
ic.oup.com/endo/article/152/3/828/2457356 by guest on 11 M
arch 2022

Proxyfan is a high-affinity ligand to the H3 receptor,with a Ki of 3–5 nM to the mouse and rat H3 receptor (5).Proxyfan is an H3 receptor protean agonist because itshows a spectrum of pharmacological effects from fullagonist, inverse agonist, to full antagonist, depending onthe constitutive activity of the H3 receptors (5). For ex-ample, proxyfan acts on the H3 receptor as a full agonistto enhance consolidation of fear memory in the rat (6); asan inverse agonist to increase wakefulness and decreasesleep in the mouse (5); and as a neutral antagonist to in-hibit H3 agonist-induced water intake in the mouse (7).
The present study demonstrates a direct glucose-low-ering effect of the H3 receptor using the protean agonistproxyfan in lean, diet-induced obese (DIO) mice, andobese type 2 diabetic mice but not in H3 receptor null miceafter oral administration. The data link the H3 receptor toglucose regulation through the release of insulin and in-dicate this receptor as a potential target for the pharma-cological treatment of diabetes.
Materials and Methods
Animals, chemicals, and dietsMale ICR mice (5 wk old) were purchased from Taconic
Farms (Albany, NY) and singly housed in a 12-h light, 12-h darkcycle with ad libitum access to water and chow (8604 TekladRodent Diet; Harlan Laboratories, Madison, WI). In some stud-ies, mice were fed semipurified nutritionally complete diet withhigh-fat and moderate-cholesterol content [45% kcal from fatand 0.12% (wt/wt) from cholesterol; catalog no. D04012801from Research Diets, New Brunswick, NJ] for at least 3 wkbefore testing to generate DIO mice. Male ICR mice (9 wk old)with intracerebroventricular (ICV) cannula were also purchasedfrom Taconic Farms and housed as described above. Briefly,mice were anesthetized by inhalation of isoflurane and ip injec-tion of buprenorphine. Using aseptic technique, the guide can-nula was stereotaxically placed with (coordinates: anteroposte-rior � �1.0 mm, mediolateral � �1.0 mm, dorsoventral � �2.0mm) into a hole in the skull and anchored with cyanoacrylateglue and cranioplastic cement to anchor screws. The guide can-nula was then sealed with dummy cannula. Proxyfan hydrochlo-ride was custom synthesized by Tocris Bioscience (Ellisville,MO). Thioperamide, imetit dihybrobromide, zolantidine dimal-eate, and proxyfan maleate were also purchased from TocrisBioscience. Chlorpheniramine (SCH190) was synthesized inhouse. Doses for thioperamide (8), imetit (9), zolantidine (9), andchlorpheniramine (10) were based on literature findings of invivo receptor selectivity, in-house verification of plasma expo-sure, and, in some cases, dose titration of ip glucose tolerance test(ipGTT) studies in ICR mice. All H3 receptor ligands were for-mulated in 0.4% methylcellulose (MC) for oral administration.A recombinant mouse leptin ELISA kit was purchased fromR&D Systems, Inc. (Minneapolis, MN). Compounds for ip orICV administration were dissolved in physiological buffer solu-tion (Dulbecco’s PBS, Invitrogen Corp., Carlsbad, CA). NaOHor HCl was used to titrate the PBS to a neutral solution for ip orICV dosing. All studies were conducted in a facility accredited by
the American Association for Accreditation of Laboratory An-imal Care following protocols approved by the Animal Care andUse Committee of Merck Research Laboratories.
ipGTT or ip insulin tolerance test (ipITT)Mice were deprived of food overnight (�18 h), and groups
were balanced based on fasting glucose levels before oral ad-ministration of either vehicle (0.4% MC) or test compound 1 hbefore glucose (1 g/kg, ip) or insulin challenge (Humulin; Lilly,Indianapolis, IN; 1 U/kg ip). Tail blood was collected at 0, 20, 40,60, and 120 min after ip glucose or insulin challenge. Bloodglucose was measured at each time point using the glucose ox-idase method (Ascensia Breeze 2; Bayer Corp., Tarrytown, NY).Area under the curve (AUC) of blood glucose concentration timewas analyzed by trapezoidal integration. Change in blood glu-cose AUC was based on time zero subtracted from 0–120 minAUC. Comparison of hrh3 (�/� or �/�) mice treated with eitherproxyfan or vehicle was conducted using a crossover study de-sign in the ipGTT experiments.
Plasma insulin and glucagon analysisMice were dosed orally with either vehicle (0.4% MC) or test
compound 1 h before glucose (1 g/kg, ip) after an overnight fast.Tail blood was collected at 0, 20, and 40 min after ip glucoseadministration. Each animal was used only at one time point.Plasma insulin and glucagon were measured by Luminex-beadimmunoassay kit (Millipore Corp./Linco, Billerica, MA). Forcalculation of plasma insulin to glucagon ratio, the plasma in-sulin concentration was divided by the plasma glucagon con-centration at the 0-, 20-, and 40-min time points.
Diabetes induction with streptozotocin (STZ)Mice were placed on a high-fat/cholesterol diet for at least 3
wk before injecting once with low-dose STZ (75 mg/kg ip). Threeweeks after STZ injection, blood glucose was measured in micefrom tail blood using a glucose oxidase method. Mice were stud-ied in a nonfasting state when dosed orally with either vehicle(0.4% MC) or proxyfan at 1000 h. Nonfasting glucose (NFG)was measured by tail bleed at 2, 4, and 6 h after dosing.
Generation of Hrh3 knockout miceHrh3 knockout mice (Hrh3�/�) were custom generated by
Caliper Life Sciences, Inc. (Hopkinton, MA). Briefly, a targetingvector was designed to delete a 4.9-kb fragment correspondingto the entire gene-coding and noncoding sequence (GenBankaccession no. NM_133849), and electroporated into C57BL/6-derived embryonic stem (ES) cells. G418-resistant colonies werescreened for homologous recombination (Supplemental Fig. 1Apublished on The Endocrine Society’s Journals Online web siteat http://endo.endojournals.org), and two targeted ES cell cloneswere selected for Flp electroporation. Two ES cell clones withconfirmed neo excision (Supplemental Fig. 1B) were injected intomouse blastocysts to generate chimeric mice that were thenmated to C57BL/6Tac females. Hrh3 heterozygous (Hrh3�/�)offspring were identified by a PCR-based screening strategy(Supplemental Fig. 1C) and then interbred to generate Hrh3�/�
mice in the expected Mendelian ratio of 1:2:1, demonstratingthat Hrh3 expression is not essential for embryonic develop-ment. Disruption of Hrh3 expression was confirmed by real-timequantitative PCR analysis (Supplemental Fig. 1D).
Endocrinology, March 2011, 152(3):828–835 endo.endojournals.org 829
Dow
nloaded from https://academ
ic.oup.com/endo/article/152/3/828/2457356 by guest on 11 M
arch 2022
mRNA expression analysismRNA from various tissues was extracted using the Ultraspec
RNA isolation kit from Biotecx (Houston, TX) following spec-ifications from the manufacturer. cDNA was generated by re-verse transcription using random hexamers (Promega Corp.,Madison, WI), and oligo-dT primers (Life Technologies, Gaith-ersburg, MD). Quantitative real-time PCR analysis was per-
formed on an ABI 7700 sequence-detection instrument follow-ing manufacturer’s instructions. For the analysis, 25 ng cDNAwere used together with primers at 0.9 �M final concentration,and 6-carboxyfluorescein-labeled diagnostic probe at a finalconcentration of 0.25 �M. rRNA primers and probe (PE AppliedBiosystems, Foster City, CA) were used as an internal control.Plasmidcontaining thehrh3genewasusedasa standard, ranging
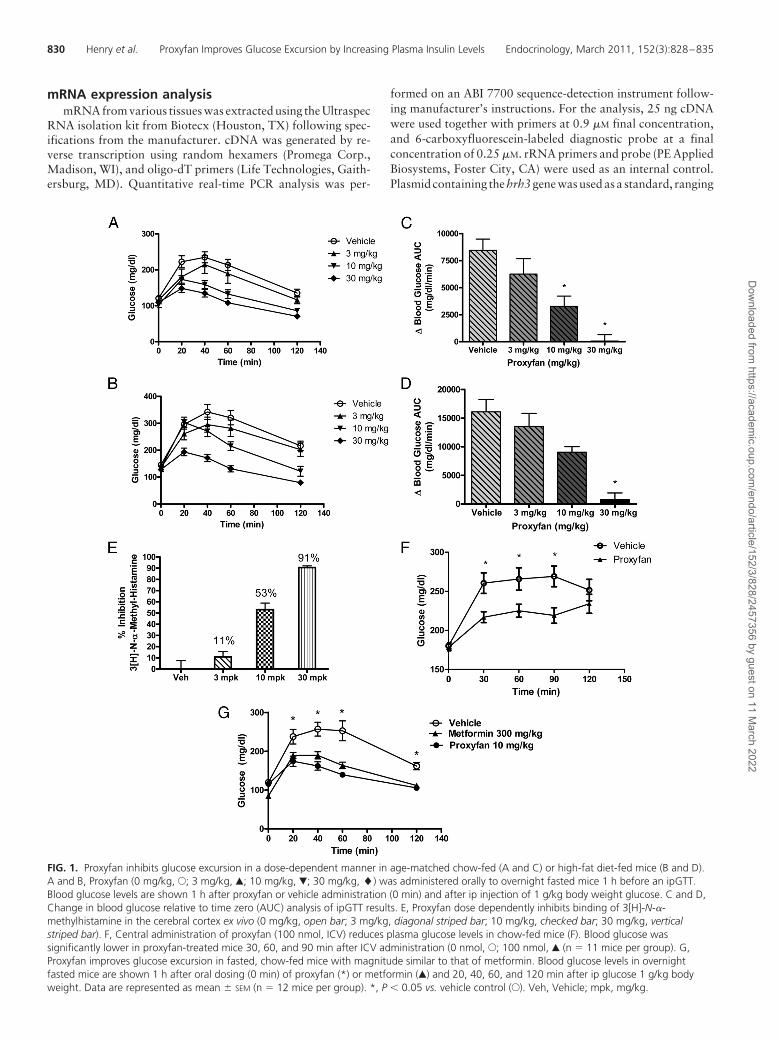
FIG. 1. Proxyfan inhibits glucose excursion in a dose-dependent manner in age-matched chow-fed (A and C) or high-fat diet-fed mice (B and D).A and B, Proxyfan (0 mg/kg, E; 3 mg/kg, Œ; 10 mg/kg, �; 30 mg/kg, �) was administered orally to overnight fasted mice 1 h before an ipGTT.Blood glucose levels are shown 1 h after proxyfan or vehicle administration (0 min) and after ip injection of 1 g/kg body weight glucose. C and D,Change in blood glucose relative to time zero (AUC) analysis of ipGTT results. E, Proxyfan dose dependently inhibits binding of 3[H]-N-�-methylhistamine in the cerebral cortex ex vivo (0 mg/kg, open bar; 3 mg/kg, diagonal striped bar; 10 mg/kg, checked bar; 30 mg/kg, verticalstriped bar). F, Central administration of proxyfan (100 nmol, ICV) reduces plasma glucose levels in chow-fed mice (F). Blood glucose wassignificantly lower in proxyfan-treated mice 30, 60, and 90 min after ICV administration (0 nmol, E; 100 nmol, Œ (n � 11 mice per group). G,Proxyfan improves glucose excursion in fasted, chow-fed mice with magnitude similar to that of metformin. Blood glucose levels in overnightfasted mice are shown 1 h after oral dosing (0 min) of proxyfan (*) or metformin (Œ) and 20, 40, 60, and 120 min after ip glucose 1 g/kg bodyweight. Data are represented as mean � SEM (n � 12 mice per group). *, P � 0.05 vs. vehicle control (E). Veh, Vehicle; mpk, mg/kg.
830 Henry et al. Proxyfan Improves Glucose Excursion by Increasing Plasma Insulin Levels Endocrinology, March 2011, 152(3):828–835
Dow
nloaded from https://academ
ic.oup.com/endo/article/152/3/828/2457356 by guest on 11 M
arch 2022
from 1 ng to 1 fg. Data were analyzed using Sequence DetectionSystems software version 1.7 (Supplemental Fig. 2).
Ex vivo binding assayAfter the final time point of the proxyfan dose response in the
ipGTT study (120 min), mice were euthanized by CO2 gas, andbrains were removed. Individual mouse cortexes were dissectedand homogenized in ice-cold assay buffer, 50 mM Na2HPO4-KH2PO4 buffer (pH 6.8). Samples were then frozen at �80 C forat least12h.Protein concentrationwasdeterminedbyBCAProteinAssay.Homogenizedcortexsamplecontaining140�g/assay,0.1%BSA, 1 nM 3H-N-�-methylhistamine, and 10 �M thioperamide wasincubated for 30 min at room temperature for nonspecific bindingor total binding. Incubations were performed in quadruplicate andstopped by rapid filtration on a Brandel Harvester (Brandel, Inc.,Plantation, FL) using Unifilter-96, GF/B plates presoaked in 0.3%polyethyleneimine for 30 min. The remaining radioactivity wasmeasured on a Packard TopCount-NTX (Packard Instruments,Meriden, CT). Specific binding was calculated by subtracting thenonspecific binding from the total.
ICV infusionICV cannulated mice (12 wk old) were deprived of food for 2 h,
and groups were balanced based on fasting glucose levels beforeICVadministrationofeithervehicle (sterile saline)orproxyfanmal-eate (100 nmol) while in their home cage. Tail blood was collectedat30,60,90, and120minafter infusionofproxyfan into the lateralventricle using a BAS Bee pump (Bioanalytical Systems, WestLafeyette, IN) at a rate of 1 �l/min for 1 min. Blood glucose wasmeasuredateachtimepointusingtheAscensiaBreeze2glucometer.
Statistical analysisResults are given as means � SE. Statistically significant
differences between vehicle and proxyfan or test compound-treated mice were determined using a one-way ANOVA andDunnett, Tukey, or Bonferri multiple comparisons posttest.When SD values were significantly different between groups, anonparametric ANOVA (Kruskal-Wallis test) was used formultiple comparisons. All statistical tests were performed us-ing GraphPad InStat version 3.01 for Windows 95 (GraphPadSoftware, San Diego CA; www.graphpad.com). Values of P �0.05 were considered statistically significant.
Results
Proxyfan improves ip glucose tolerance throughactivation of central H3 receptors
Oral administration of proxyfan decreased glucose ex-cursion after an ipGTT in age-matched 8-wk-old chow-fedand DIO ICR mice in a dose-dependent manner (Fig. 1,A–D). At the lowest dose tested (3 mg/kg) proxyfan showedmodest glucose-lowering effects in both groups, whereas 10mg/kg produced significant inhibition of glucose excursionin the lean, chow-fed mice. DIO mice were less sensitive toproxyfan at the 10 mg/kg dose. The highest dose elicitedprofound glucose lowering in both lean and DIO mice. Inaccord with the in vivo results, proxyfan inhibited ex vivo
binding, in a dose-dependent manner, of 3[H]-N-�-methyl-histamine in the cerebral cortex of these mice (Fig. 1E). Cen-tral administration of proxyfan (100 nmol) significantlyreduced the plasma glucose levels compared with vehicle-treated mice at 30, 60, and 90 min post-ICV dosing in 2-hfasted, chow-fed mice (Fig. 1F). Proxyfan (10 mg/kg) im-proved glucose excursion to the same levels as that of met-formin (300 mg/kg, Fig. 1G) in lean, chow-fed mice.
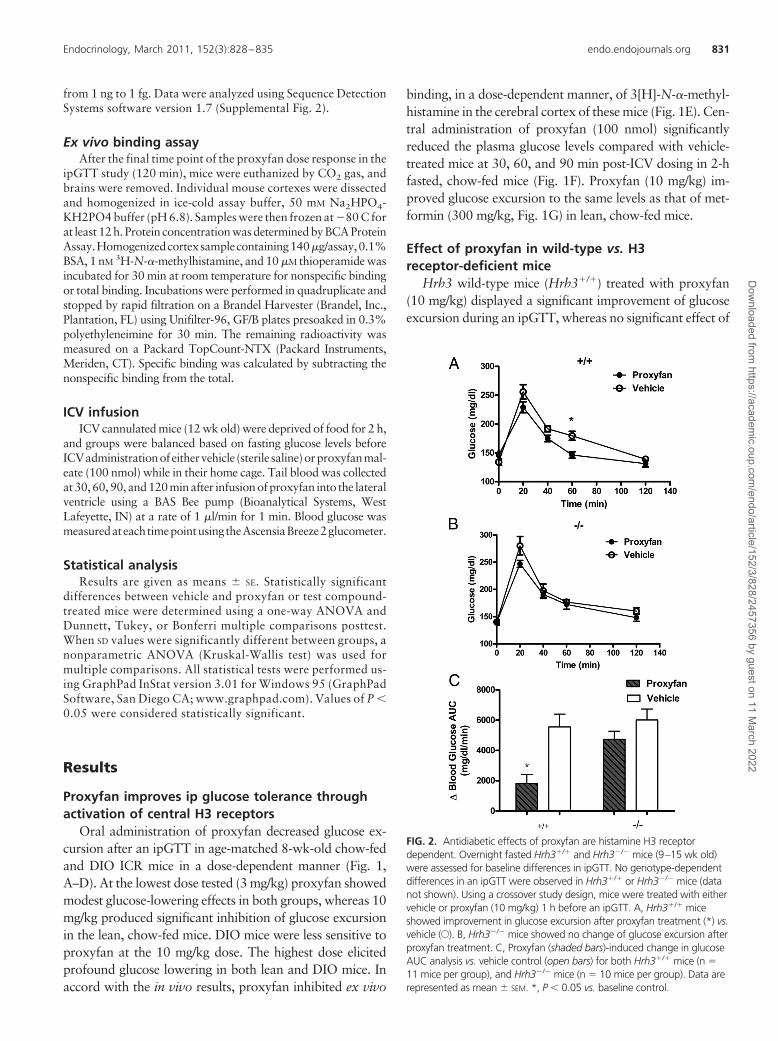
Effect of proxyfan in wild-type vs. H3receptor-deficient mice
Hrh3 wild-type mice (Hrh3�/�) treated with proxyfan(10 mg/kg) displayed a significant improvement of glucoseexcursion during an ipGTT, whereas no significant effect of
FIG. 2. Antidiabetic effects of proxyfan are histamine H3 receptordependent. Overnight fasted Hrh3�/� and Hrh3�/� mice (9–15 wk old)were assessed for baseline differences in ipGTT. No genotype-dependentdifferences in an ipGTT were observed in Hrh3�/� or Hrh3�/� mice (datanot shown). Using a crossover study design, mice were treated with eithervehicle or proxyfan (10 mg/kg) 1 h before an ipGTT. A, Hrh3�/� miceshowed improvement in glucose excursion after proxyfan treatment (*) vs.vehicle (E). B, Hrh3�/� mice showed no change of glucose excursion afterproxyfan treatment. C, Proxyfan (shaded bars)-induced change in glucoseAUC analysis vs. vehicle control (open bars) for both Hrh3�/� mice (n �11 mice per group), and Hrh3�/� mice (n � 10 mice per group). Data arerepresented as mean � SEM. *, P � 0.05 vs. baseline control.
Endocrinology, March 2011, 152(3):828–835 endo.endojournals.org 831
Dow
nloaded from https://academ
ic.oup.com/endo/article/152/3/828/2457356 by guest on 11 M
arch 2022
proxyfan treatment was observed in the Hrh3�/� mice (Fig.2, A–C). Although the effects of 10 mg/kg proxyfan were H3receptor dependent, both genotypes showed inhibition ofglucose excursion at 30 mg/kg (data not shown), suggestingtheactivityofproxyfanat thishighdosemay involveamech-anism distinct from H3 receptor interaction. There were nosignificant differences between wild-type and H3 receptorknockout mice treated with vehicle in 0-, 20-, 40-, or 60-mintime points of GTTs. The blood glucose in the wild-type micehad come down to baseline at 120-min time point, whereas theH3 receptor knockout mice had significantly higher blood glu-cose level than the baseline. Therefore, the endogenous H3 re-ceptorsignalingmayplaysomerole inpreventing impairedglu-cose tolerance. However, the overall glucose excursionanalyzed by change of glucose AUC was not significantly dif-ferent between wild-type and H3 receptor knockout mice.
Proxyfan increases plasma insulin levels but doesnot affect insulin sensitivity
Animals were administered proxyfan (10 mg/kg) or ve-hicle (0.4% MC) 1 h before an ip glucose challenge (1
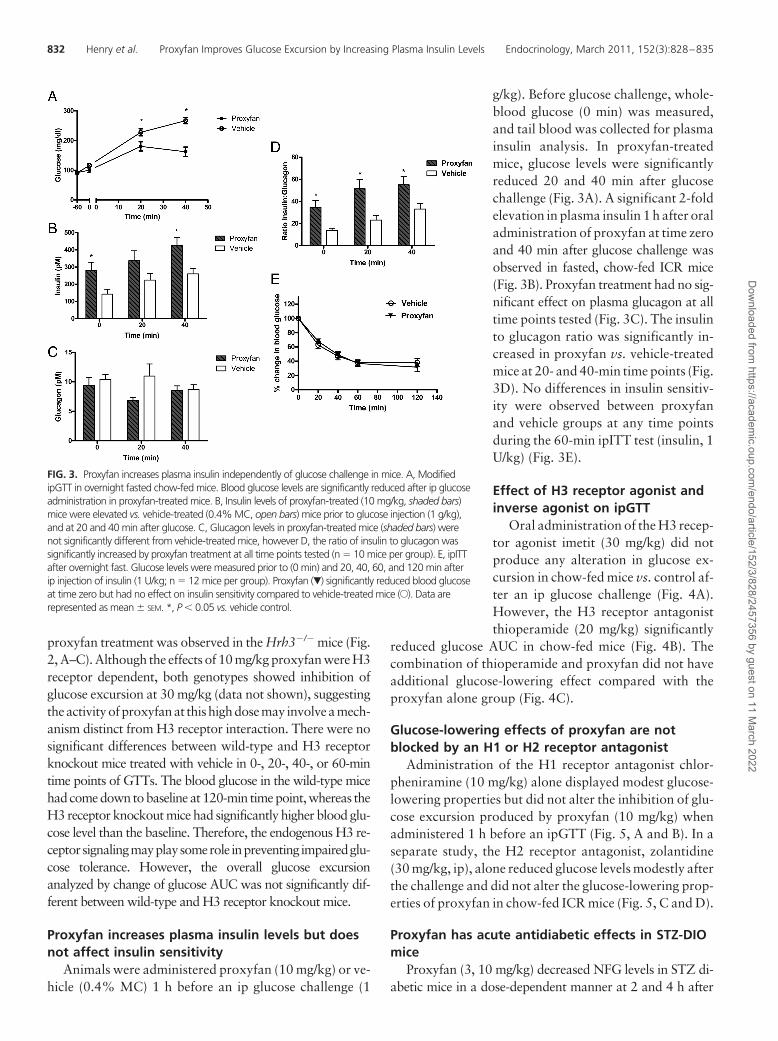
g/kg). Before glucose challenge, whole-blood glucose (0 min) was measured,and tail blood was collected for plasmainsulin analysis. In proxyfan-treatedmice, glucose levels were significantlyreduced 20 and 40 min after glucosechallenge (Fig. 3A). A significant 2-foldelevation in plasma insulin 1 h after oraladministration of proxyfan at time zeroand 40 min after glucose challenge wasobserved in fasted, chow-fed ICR mice(Fig. 3B). Proxyfan treatment had no sig-nificant effect on plasma glucagon at alltime points tested (Fig. 3C). The insulinto glucagon ratio was significantly in-creased in proxyfan vs. vehicle-treatedmice at 20- and 40-min time points (Fig.3D). No differences in insulin sensitiv-ity were observed between proxyfanand vehicle groups at any time pointsduring the 60-min ipITT test (insulin, 1U/kg) (Fig. 3E).
Effect of H3 receptor agonist andinverse agonist on ipGTT
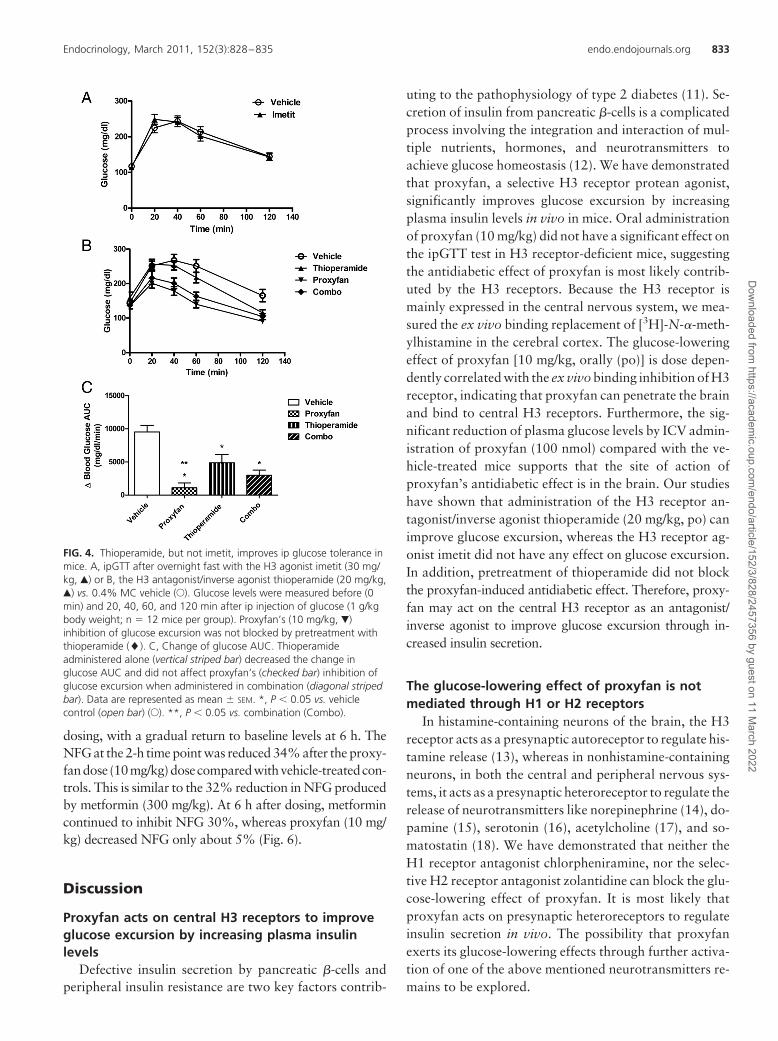
Oral administration of the H3 recep-tor agonist imetit (30 mg/kg) did notproduce any alteration in glucose ex-cursion in chow-fed mice vs. control af-ter an ip glucose challenge (Fig. 4A).However, the H3 receptor antagonistthioperamide (20 mg/kg) significantly
reduced glucose AUC in chow-fed mice (Fig. 4B). Thecombination of thioperamide and proxyfan did not haveadditional glucose-lowering effect compared with theproxyfan alone group (Fig. 4C).
Glucose-lowering effects of proxyfan are notblocked by an H1 or H2 receptor antagonist
Administration of the H1 receptor antagonist chlor-pheniramine (10 mg/kg) alone displayed modest glucose-lowering properties but did not alter the inhibition of glu-cose excursion produced by proxyfan (10 mg/kg) whenadministered 1 h before an ipGTT (Fig. 5, A and B). In aseparate study, the H2 receptor antagonist, zolantidine(30 mg/kg, ip), alone reduced glucose levels modestly afterthe challenge and did not alter the glucose-lowering prop-erties of proxyfan in chow-fed ICR mice (Fig. 5, C and D).
Proxyfan has acute antidiabetic effects in STZ-DIOmice
Proxyfan (3, 10 mg/kg) decreased NFG levels in STZ di-abetic mice in a dose-dependent manner at 2 and 4 h after
FIG. 3. Proxyfan increases plasma insulin independently of glucose challenge in mice. A, ModifiedipGTT in overnight fasted chow-fed mice. Blood glucose levels are significantly reduced after ip glucoseadministration in proxyfan-treated mice. B, Insulin levels of proxyfan-treated (10 mg/kg, shaded bars)mice were elevated vs. vehicle-treated (0.4% MC, open bars) mice prior to glucose injection (1 g/kg),and at 20 and 40 min after glucose. C, Glucagon levels in proxyfan-treated mice (shaded bars) werenot significantly different from vehicle-treated mice, however D, the ratio of insulin to glucagon wassignificantly increased by proxyfan treatment at all time points tested (n � 10 mice per group). E, ipITTafter overnight fast. Glucose levels were measured prior to (0 min) and 20, 40, 60, and 120 min afterip injection of insulin (1 U/kg; n � 12 mice per group). Proxyfan (�) significantly reduced blood glucoseat time zero but had no effect on insulin sensitivity compared to vehicle-treated mice (E). Data arerepresented as mean � SEM. *, P � 0.05 vs. vehicle control.
832 Henry et al. Proxyfan Improves Glucose Excursion by Increasing Plasma Insulin Levels Endocrinology, March 2011, 152(3):828–835
Dow
nloaded from https://academ
ic.oup.com/endo/article/152/3/828/2457356 by guest on 11 M
arch 2022
dosing, with a gradual return to baseline levels at 6 h. TheNFG at the 2-h time point was reduced 34% after the proxy-fandose (10mg/kg)dosecomparedwithvehicle-treatedcon-trols. This is similar to the 32% reduction in NFG producedby metformin (300 mg/kg). At 6 h after dosing, metformincontinued to inhibit NFG 30%, whereas proxyfan (10 mg/kg) decreased NFG only about 5% (Fig. 6).
Discussion
Proxyfan acts on central H3 receptors to improveglucose excursion by increasing plasma insulinlevels
Defective insulin secretion by pancreatic �-cells andperipheral insulin resistance are two key factors contrib-
uting to the pathophysiology of type 2 diabetes (11). Se-cretion of insulin from pancreatic �-cells is a complicatedprocess involving the integration and interaction of mul-tiple nutrients, hormones, and neurotransmitters toachieve glucose homeostasis (12). We have demonstratedthat proxyfan, a selective H3 receptor protean agonist,significantly improves glucose excursion by increasingplasma insulin levels in vivo in mice. Oral administrationof proxyfan (10 mg/kg) did not have a significant effect onthe ipGTT test in H3 receptor-deficient mice, suggestingthe antidiabetic effect of proxyfan is most likely contrib-uted by the H3 receptors. Because the H3 receptor ismainly expressed in the central nervous system, we mea-sured the ex vivo binding replacement of [3H]-N-�-meth-ylhistamine in the cerebral cortex. The glucose-loweringeffect of proxyfan [10 mg/kg, orally (po)] is dose depen-dently correlated with the ex vivo binding inhibition of H3receptor, indicating that proxyfan can penetrate the brainand bind to central H3 receptors. Furthermore, the sig-nificant reduction of plasma glucose levels by ICV admin-istration of proxyfan (100 nmol) compared with the ve-hicle-treated mice supports that the site of action ofproxyfan’s antidiabetic effect is in the brain. Our studieshave shown that administration of the H3 receptor an-tagonist/inverse agonist thioperamide (20 mg/kg, po) canimprove glucose excursion, whereas the H3 receptor ag-onist imetit did not have any effect on glucose excursion.In addition, pretreatment of thioperamide did not blockthe proxyfan-induced antidiabetic effect. Therefore, proxy-fan may act on the central H3 receptor as an antagonist/inverse agonist to improve glucose excursion through in-creased insulin secretion.
The glucose-lowering effect of proxyfan is notmediated through H1 or H2 receptors
In histamine-containing neurons of the brain, the H3receptor acts as a presynaptic autoreceptor to regulate his-tamine release (13), whereas in nonhistamine-containingneurons, in both the central and peripheral nervous sys-tems, it acts as a presynaptic heteroreceptor to regulate therelease of neurotransmitters like norepinephrine (14), do-pamine (15), serotonin (16), acetylcholine (17), and so-matostatin (18). We have demonstrated that neither theH1 receptor antagonist chlorpheniramine, nor the selec-tive H2 receptor antagonist zolantidine can block the glu-cose-lowering effect of proxyfan. It is most likely thatproxyfan acts on presynaptic heteroreceptors to regulateinsulin secretion in vivo. The possibility that proxyfanexerts its glucose-lowering effects through further activa-tion of one of the above mentioned neurotransmitters re-mains to be explored.
FIG. 4. Thioperamide, but not imetit, improves ip glucose tolerance inmice. A, ipGTT after overnight fast with the H3 agonist imetit (30 mg/kg, Œ) or B, the H3 antagonist/inverse agonist thioperamide (20 mg/kg,Œ) vs. 0.4% MC vehicle (E). Glucose levels were measured before (0min) and 20, 40, 60, and 120 min after ip injection of glucose (1 g/kgbody weight; n � 12 mice per group). Proxyfan’s (10 mg/kg, �)inhibition of glucose excursion was not blocked by pretreatment withthioperamide (�). C, Change of glucose AUC. Thioperamideadministered alone (vertical striped bar) decreased the change inglucose AUC and did not affect proxyfan’s (checked bar) inhibition ofglucose excursion when administered in combination (diagonal stripedbar). Data are represented as mean � SEM. *, P � 0.05 vs. vehiclecontrol (open bar) (E). **, P � 0.05 vs. combination (Combo).
Endocrinology, March 2011, 152(3):828–835 endo.endojournals.org 833
Dow
nloaded from https://academ
ic.oup.com/endo/article/152/3/828/2457356 by guest on 11 M
arch 2022
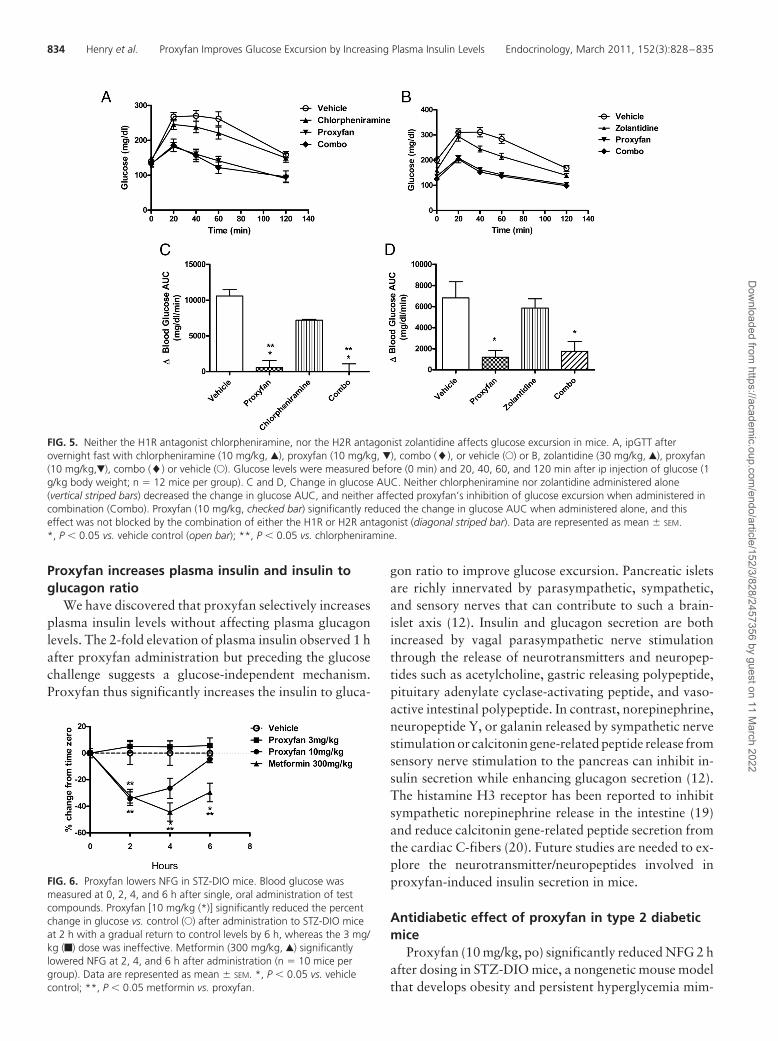
Proxyfan increases plasma insulin and insulin toglucagon ratio
We have discovered that proxyfan selectively increasesplasma insulin levels without affecting plasma glucagonlevels. The 2-fold elevation of plasma insulin observed 1 hafter proxyfan administration but preceding the glucosechallenge suggests a glucose-independent mechanism.Proxyfan thus significantly increases the insulin to gluca-
gon ratio to improve glucose excursion. Pancreatic isletsare richly innervated by parasympathetic, sympathetic,and sensory nerves that can contribute to such a brain-islet axis (12). Insulin and glucagon secretion are bothincreased by vagal parasympathetic nerve stimulationthrough the release of neurotransmitters and neuropep-tides such as acetylcholine, gastric releasing polypeptide,pituitary adenylate cyclase-activating peptide, and vaso-active intestinal polypeptide. In contrast, norepinephrine,neuropeptide Y, or galanin released by sympathetic nervestimulation or calcitonin gene-related peptide release fromsensory nerve stimulation to the pancreas can inhibit in-sulin secretion while enhancing glucagon secretion (12).The histamine H3 receptor has been reported to inhibitsympathetic norepinephrine release in the intestine (19)and reduce calcitonin gene-related peptide secretion fromthe cardiac C-fibers (20). Future studies are needed to ex-plore the neurotransmitter/neuropeptides involved inproxyfan-induced insulin secretion in mice.
Antidiabetic effect of proxyfan in type 2 diabeticmice
Proxyfan (10 mg/kg, po) significantly reduced NFG 2 hafter dosing in STZ-DIO mice, a nongenetic mouse modelthat develops obesity and persistent hyperglycemia mim-
FIG. 5. Neither the H1R antagonist chlorpheniramine, nor the H2R antagonist zolantidine affects glucose excursion in mice. A, ipGTT afterovernight fast with chlorpheniramine (10 mg/kg, Œ), proxyfan (10 mg/kg, �), combo (�), or vehicle (E) or B, zolantidine (30 mg/kg, Œ), proxyfan(10 mg/kg,�), combo (�) or vehicle (E). Glucose levels were measured before (0 min) and 20, 40, 60, and 120 min after ip injection of glucose (1g/kg body weight; n � 12 mice per group). C and D, Change in glucose AUC. Neither chlorpheniramine nor zolantidine administered alone(vertical striped bars) decreased the change in glucose AUC, and neither affected proxyfan’s inhibition of glucose excursion when administered incombination (Combo). Proxyfan (10 mg/kg, checked bar) significantly reduced the change in glucose AUC when administered alone, and thiseffect was not blocked by the combination of either the H1R or H2R antagonist (diagonal striped bar). Data are represented as mean � SEM.*, P � 0.05 vs. vehicle control (open bar); **, P � 0.05 vs. chlorpheniramine.
FIG. 6. Proxyfan lowers NFG in STZ-DIO mice. Blood glucose wasmeasured at 0, 2, 4, and 6 h after single, oral administration of testcompounds. Proxyfan [10 mg/kg (*)] significantly reduced the percentchange in glucose vs. control (E) after administration to STZ-DIO miceat 2 h with a gradual return to control levels by 6 h, whereas the 3 mg/kg (f) dose was ineffective. Metformin (300 mg/kg, Œ) significantlylowered NFG at 2, 4, and 6 h after administration (n � 10 mice pergroup). Data are represented as mean � SEM. *, P � 0.05 vs. vehiclecontrol; **, P � 0.05 metformin vs. proxyfan.
834 Henry et al. Proxyfan Improves Glucose Excursion by Increasing Plasma Insulin Levels Endocrinology, March 2011, 152(3):828–835
Dow
nloaded from https://academ
ic.oup.com/endo/article/152/3/828/2457356 by guest on 11 M
arch 2022

icking human type 2 diabetes. This antidiabetic effect wascomparable to that of metformin (300 mg/kg, po) at 2 hafter dosing. Metformin demonstrated a sustained antidi-abetic effect for 6 h, whereas the effect of proxyfan lastedonly for 4 h after dosing. Metformin, a biguanide thatexerts its effects by reducing endogenous hepatic glucoseproduction, is the first-line drug of choice for the treatmentof type 2 diabetes, particularly in overweight and obesepeople. It lowers HbA1c by 1.3% without the weight gainside effect of the thiazolidinediones or sulfonylureas (21).Despite the gastrointestinal side effects associated withmetformin treatment due to the gram quantities requiredfor efficacy, it is widely prescribed because hypoglycemiacomplication rates are low. Although proxyfan has similarantidiabetic efficacy as metformin at early time points,proxyfan showed shorter duration of action in the mousemodel. Because proxyfan-induced insulin secretion is glu-cose independent, the potential for hypoglycemia maylimit the utility of this class of compound.
Conclusion
Type 2 diabetes mellitus is a multifactorial metabolic dis-ease characterized by insulin secretion deficiency and mul-tiple organ insulin resistance, which lead to hyperglycemiawith macro- and/or microvascular complications. Ourstudies provide evidence that modulation of the central H3receptor by proxyfan can improve glucose excursion byincreasing plasma insulin levels in a glucose-independentmanner. The effect of proxyfan cannot be blocked by H1,H2, or H3 receptor antagonists. Although the antidiabeticefficacy of proxyfan in STZ-DIO mice was similar to met-formin, the duration of action of proxyfan was shorterthan metformin. Whether modulation of the central H3receptor has a role in type 2 diabetes pathophysiologyand/or treatment remains to be established.
Acknowledgments
We thank Dr. Timothy Kowalski for critical review of this manu-script. We also thank Yun-Sheng Hsieh and Christine Duncanfor Drug Metabolism/Pharmacokinetic support and LorraineHarmon, David Lauer, George Nartey, and Tamaira Roberson(Laboratory Animal Resources Department) for exceptional an-imal care provided throughout these studies (Merck ResearchLabs, Kenilworth, NJ).
Address all correspondence and requests for reprints to: JoyceJ. Hwa, Ph.D., In vivo Pharmacology, Merck Research Labora-tories, 126 East Lincoln Avenue, Rahway, New Jersey 07065.E-mail: [email protected].
Disclosure Summary: All authors are employees of MerckResearch Laboratories.
References
1. Tiligada E, Zampeli E, Sander K, Stark H 2009 Histamine H3 andH4 receptors as novel drug targets. Expert Opin Investig Drugs 18:1519–1531
2. Toyota H, Dugovic C, Koehl M, Laposky AD, Weber C, Ngo K, WuY, Lee DH, Yanai K, Sakurai E, Watanabe T, Liu C, Chen J, BarbierAJ, Turek FW, Fung-Leung WP, Lovenberg TW 2002 Behavioralcharacterization of mice lacking histamine H(3) receptors. MolPharmacol 62:389–397
3. Gastinger MJ, Barber AJ, Vardi N, Marshak DW 2006 Histaminereceptors in mammalian retinas. J Comp Neurol 495:658–667
4. Takahashi K, Suwa H, Ishikawa T, Kotani H 2002 Targeted dis-ruption of H3 receptors results in changes in brain histamine toneleading to an obese phenotype. J Clin Invest 110:1791–1799
5. Gbahou F, Rouleau A, Morisset S, Parmentier R, Crochet S, Lin JS,Ligneau X, Tardivel-Lacombe J, Stark H, Schunack W, Ganellin CR,Schwartz JC, Arrang JM 2003 Protean agonism at histamine H3 re-ceptors invitroand invivo.ProcNatlAcadSciUSA100:11086–11091
6. Baldi E, Bucherelli C, Schunack W, Cenni G, Blandina P, Passani MB2005 The H3 receptor protean agonist proxyfan enhances the expres-sion of fear memory in the rat. Neuropharmacology 48:246–251
7. Fox GB, Pan JB, Esbenshade TA, Bitner RS, Nikkel AL, Miller T,Kang CH, Bennani YL, Black LA, Faghih R, Hancock AA, DeckerMW 2002 Differential in vivo effects of H3 receptor ligands in a newmouse dipsogenia model. Pharmacol Biochem Behav 72:741–750
8. Mollet A, Lutz TA, Meier S, Riediger T, Rushing PA, Scharrer E2001 Histamine H1 receptors mediate the anorectic action of thepancreatic hormone amylin. Am J Physiol Regul Integr CompPhysiol 281:R1442–R1448
9. Yokota E, Kuyama S, Sugimoto Y, Ogawa M, Kamei C 2008 Par-ticipation of histamine H3 receptors in experimental allergic rhinitisof mice. J Pharmacol Sci 108:206–211
10. Rahman MA, Inoue T, Ishikawa T, Yatsuzuka R, Ohtsu H, KameiC 2007 Involvement of chemical mediators in nasal allergic re-sponses of HDC-KO mice. Eur J Pharmacol 567:245–251
11. Abdul-Ghani MA, Tripathy D, DeFronzo RA 2006 Contributionsof �-cell dysfunction and insulin resistance to the pathogenesis ofimpaired glucose tolerance and impaired fasting glucose. DiabetesCare 29:1130–1139
12. Ahren B 2000 Autonomic regulation of islet hormone secretion—implications for health and disease. Diabetologia 43:393–410
13. Malinowska B, Godlewski G, Schlicker E 1998 Histamine H3 re-ceptors—general characterization and their function in the cardio-vascular system. J Physiol Pharmacol 49:191–211
14. Molderings GJ, Weissenborn G, Schlicker E, Likungu J, Gothert M1992 Inhibition of noradrenaline release from the sympatheticnerves of the human saphenous vein by presynaptic histamine H3receptors. Naunyn Schmiedebergs Arch Pharmacol 346:46–50
15. Nowak JZ 1985 Depolarisation-evoked release of dopamine andhistamine from brain tissue and studies on presynaptic dopamine-histamine interaction. Pol J Pharmacol Pharm 37:359–381
16. Racke K, Schworer H 1991 Regulation of serotonin release from theintestinal mucosa. Pharmacol Res 23:13–25
17. Stocking EM, Letavic MA 2008 Histamine H3 antagonists as wake-promoting and pro-cognitive agents. Curr Top Med Chem 8:988–1002
18. Barocelli E, Ballabeni V 2003 Histamine in the control of gastric acidsecretion: a topic review. Pharmacol Res 47:299–304
19. Blandizzi C, Tognetti M, Colucci R, Del Tacca M 2000 HistamineH(3) receptors mediate inhibition of noradrenaline release from in-testinal sympathetic nerves. Br J Pharmacol 129:1387–1396
20. Imamura M, Smith NC, Garbarg M, Levi R 1996 Histamine H3-receptor-mediated inhibition of calcitonin gene-related peptide re-lease from cardiac C fibers. A regulatory negative-feedback loop.Circ Res 78:863–869
21. Bosi E 2009 Metformin—the gold standard in type 2 diabetes: whatdoes the evidence tell us? Diabetes Obes Metab 11(Suppl 2):3–8
Endocrinology, March 2011, 152(3):828–835 endo.endojournals.org 835
Dow
nloaded from https://academ
ic.oup.com/endo/article/152/3/828/2457356 by guest on 11 M
arch 2022





























