Embed Size (px)
Citation preview

Association of Estrogen Receptor-� Gene PolymorphismsWith Coronary Artery Disease in Patients
With Familial HypercholesterolemiaHong Lu, Toshinori Higashikata, Akihiro Inazu, Atsushi Nohara, Wenxin Yu,
Masami Shimizu, Hiroshi Mabuchi
Abstract—To investigate the association of estrogen receptor (ER)-� gene polymorphisms with coronary artery disease(CAD), we studied 197 men and 98 postmenopausal women with heterozygous familial hypercholesterolemia. Weexamined the known polymorphisms, including PvuII, XbaI, TA repeat, and CA repeat, and identified 6 novelpolymorphisms in the ER-� gene. The distributions of �1989T/G (a novel polymorphism in promoter B) and XbaI inintron 1 were associated with CAD in postmenopausal women and in men, with a higher frequency of the G/G genotype(P�0.03) or X1/X1 genotype (P�0.02) in the CAD group. The frequency of alleles of TA repeats �17 was found tobe significantly higher in postmenopausal women with CAD than in those without CAD (P�0.04), but not in men.Logistic regression analysis with all coronary risk factors as covariates showed that the G/G genotype was a higher riskfor CAD (odds ratio 4.5, 95% CI 1.0 to 19.5; P�0.04) but that X1/X1 was not. We conclude that �1989T/G or its linkedpolymorphisms in the ER-� gene may confer risk for CAD and that the G/G genotype may be an independent predictorfor CAD in patients with familial hypercholesterolemia. (Arterioscler Thromb Vasc Biol. 2002;22:821-827.)
Key Words: estrogen receptors � polymorphism � promoters � coronary artery disease � hypercholesterolemia
Estrogens, the effects of which are known to be mediatedprimarily via binding to estrogen receptor (ER)-� and
ER-�, have been reported to provide protection againstcoronary artery disease (CAD) in postmenopausal women.1
ER-� and ER-� have 6 domains (A through F) encoded by 8exons. ER-� has an activation function-1 (AF-1) in the A/Bdomain that is lacking in ER-�, although the DNA- andligand-binding domains are highly conserved in both ERs.2
Five polymorphisms in the ER-� gene have been reported inthe genomic DNA extracted from human breast tumors ornormal human peripheral blood leukocytes.3–11 The firstpolymorphism, PvuII, caused by a C/T transition (P1/P2) inintron 1, is located �0.4 kb upstream from exon 2.3 Thesecond polymorphism, XbaI, caused by a G/A transition(X1/X2), is located �50 bp downstream from the PvuIIpolymorphic site. Recent studies have suggested that the 2polymorphisms might be associated with breast cancer3,4 andpostmenopausal osteoporosis.5,6 The third polymorphism Bvariant, caused by a G/C transversion at codon 87 in exon 1,has been reported to be associated with hypertension.7 An-other 2 polymorphisms are dinucleotide (TA and CA) repeatpolymorphisms. The TA repeat is located at the 5� untrans-lated region, the length of which has associated with post-menopausal osteoporosis8,9 and the severity of CAD in white
men.10 The CA repeat, which was newly defined in intron 5,11
is the marker D6S440.12
Smith et al13 have reported a T/C transition at codon 157 ofboth alleles in exon 2 of the ER-� gene in a 28-year-old man,resulting in a premature stop codon. Subsequently, he hadevidence of early atherosclerosis at the age of 31 years.14 Thisobservation prompted us to test the hypothesis that geneticvariations in the ER-� gene could be a risk factor for CAD.Familial hypercholesterolemia (FH) is an autosomal domi-nant disorder characterized by primary hypercholesterolemiaand a higher incidence of premature CAD. Heterozygotesoccur in the population at a frequency of �1 in 500, placingFH among the most common monogenic diseases in humans.Women with FH suffer clinical CAD later than affected men.In England, Slack15 found the mean age of onset of CAD tobe 43 years for men and 53 years for women. In the UnitedStates, Stone et al16 found that the cumulative probability ofCAD was 52% in male heterozygotes and 33% in femaleheterozygotes aged 60 years. Mabuchi et al17 reported that theincidence of CAD was 43%, and earlier death from CAD wasfound in male Japanese heterozygotes (aged 54 years) com-pared with female heterozygotes (aged 68 years). Interest-ingly, this sex difference, which is also a characteristicfeature of the usual form of CAD in general populations, does
Received August 21, 2001; revision accepted February 13, 2002.From Molecular Genetics of Cardiovascular Disorders, Division of Cardiovascular Medicine, Graduate School of Medical Science, Kanazawa
University, Kanazawa, Japan.Correspondence to Toshinori Higashikata, MD, PhD, Molecular Genetics of Cardiovascular Disorders, Division of Cardiovascular Medicine, Graduate
School of Medical Science, Kanazawa University, Takara-machi 13-1, Kanazawa 920-8641, Japan. E-mail [email protected]© 2002 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol. is available at http://www.atvbaha.org DOI: 10.1161/01.ATV.0000014424.18209.21
817
by guest on September 10, 2016
http://atvb.ahajournals.org/D
ownloaded from
by guest on Septem
ber 10, 2016http://atvb.ahajournals.org/
Dow
nloaded from
by guest on September 10, 2016
http://atvb.ahajournals.org/D
ownloaded from
by guest on Septem
ber 10, 2016http://atvb.ahajournals.org/
Dow
nloaded from
by guest on September 10, 2016
http://atvb.ahajournals.org/D
ownloaded from
by guest on Septem
ber 10, 2016http://atvb.ahajournals.org/
Dow
nloaded from
by guest on September 10, 2016
http://atvb.ahajournals.org/D
ownloaded from
by guest on Septem
ber 10, 2016http://atvb.ahajournals.org/
Dow
nloaded from
by guest on September 10, 2016
http://atvb.ahajournals.org/D
ownloaded from
by guest on Septem
ber 10, 2016http://atvb.ahajournals.org/
Dow
nloaded from
by guest on September 10, 2016
http://atvb.ahajournals.org/D
ownloaded from

not seem to be operative in homozygotes.18 It is a uniqueapproach to investigate the role of ER-� gene polymorphismsin a group with heterozygous FH that has a major coronaryrisk factor and sex difference in the risk of CAD. Therefore,we investigated the association of the known and 6 novelpolymorphisms in the ER-� gene with CAD in patients withheterozygous FH.
MethodsSubjectsTwo hundred ninety-five unrelated Japanese patients with heterozy-gous FH who were dwelling in the Hokuriku area and recruited atKanazawa University Hospital were analyzed: 119 with CAD, eithermyocardial infarction (MI, 48 men and 9 women) or angina pectoris(47 men and 15 women), and 176 without CAD (102 men and 74women). The study was approved by the Ethics Committee ofKanazawa University Medical School. Heterozygous FH was diag-nosed when either of 2 sets of criteria was met: (1) primaryhypercholesterolemia (total cholesterol �5.9 mmol/L and�12.9 mmol/L) with tendon xanthomas or (2) primary hypercholes-terolemia with the definite diagnosis of FH in any first-degreerelative.19 Diagnosis of MI was based on clinical symptoms, appro-priate new onset of ECG changes, and elevated serum creatinephosphokinase levels. All but 3 patients with MI were evaluated bycoronary angiography. Diagnosis of angina pectoris was madeaccording to clinical symptoms and ECG findings, and only thosepatients whose coronary lesions were confirmed by coronary angiog-raphy were eligible for the present study. Those who were diagnosedwith vasospastic angina pectoris, according to clinical symptoms andECG findings with normal coronary angiogram, were excluded. Todetermine the severity of CAD, we used 2 systems: (1) affectedvessel numbers and (2) the coronary stenosis index. In the formersystem, the severity of CAD was expressed simply by affected vesselnumbers (�75% stenosis of lumen size), classifying numbers be-tween 1 and 3. The lesion at left main coronary artery was regardedas 2-vessel disease. In the latter system, the severity of stenoticchanges was assessed by a score assigned to each of 15 segments asdescribed elsewhere.19 For patients with MI, ages at their first eventswere recorded, whereas for patients with angina pectoris, ages atwhich coronary angiography was performed were recorded. Non-CAD patients were characterized by no history of angina and otherheart diseases, a normal resting ECG, and normal exercise ECGstress testing. Half of non-CAD patients (89 of 176) receivedcoronary angiographic examination. No obvious stenosis (0% to50% stenosis of lumen size) was found. Patients who were aged �80years or had thyroid disease, breast cancer, or uterine or ovariantumor were excluded. No individual was receiving or had previouslyreceived hormone replacement therapy. All women were postmeno-pausal, as defined by the absence of menstruation for �6 months orhaving attained an age �60 years. Women with surgical menopausewere excluded. Clinical data, including body mass index, smokinghistory, blood pressure, diabetes status, and lipid profiles, werecollected before introducing lipid-lowering therapy. Hypertensionwas defined as being present if antihypertensive treatment has beeninstituted or if consecutive 2-visit measurements of blood pressurewere �160 mm Hg systolic or 95 mm Hg diastolic. Manifestdiabetes mellitus was defined as present with fasting plasma glucose�140 mg/dL or �200 mg/dL at 120 minutes after 75 g of oralglucose loading.19 Subjects who smoked �10 cigarettes/d wereclassified as current smokers.
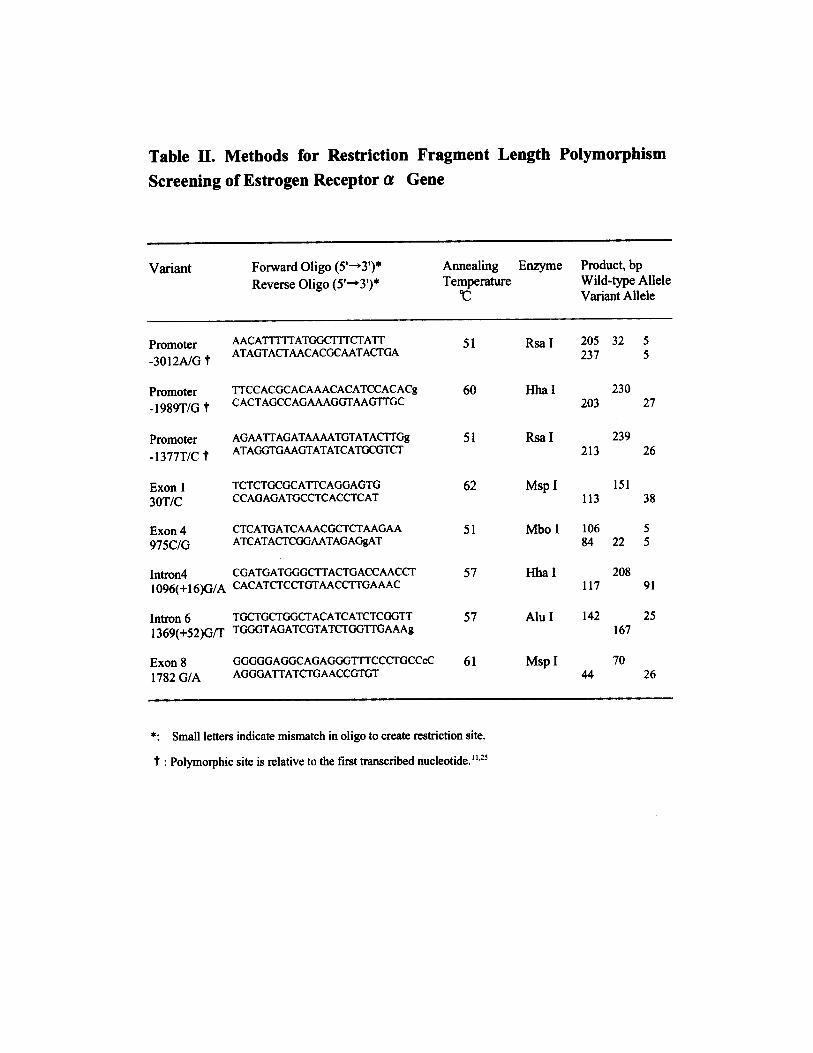
Determination of ER-� GenotypesBlood was obtained from peripheral veins of patients after informedconsent. PvuII, XbaI, and B-variant polymorphisms were analyzedby polymerase chain reaction (PCR) restriction fragment lengthpolymorphism methods. For PvuII and XbaI polymorphisms, a1.3-kb DNA fragment of the ER-� gene that contains the 2polymorphic sites was amplified by PCR,3,4 and for B-variant
polymorphism, a 195-bp DNA fragment was amplified.4 PCRproducts were digested with PvuII or XbaI (Takara Shuzo) for PvuIIor XbaI polymorphism or AccII (Nippon Gene) for B-variantpolymorphism at 37°C overnight, to determine genotypes.
Two dinucleotide repeat polymorphisms of the ER-� gene wereinvestigated by PCR to amplify the TA repeat, which is located �1.2kb upstream from exon 1 (relative to the first transcribed nucleo-tide),9 and the CA repeat, which is located 224 bp downstream fromexon 5.11 The primers were designed to amplify the TA repeat(forward 5�-FITC-TAGACGCATGATATACTTCACCTAT-3�, re-verse 5�-GCAGAATCAAATATCCAGATG-3�) and the CA repeat(forward 5�-GCCTAGTCAAATTCACAGAAAGCTA-3�, reverse5�-HEX-TTGAAGATGGAGTGAGAAAAATACC-3�). The lengthof the 2 polymorphisms was determined by analysis of PCR productson 6% denaturing polyacrylamide gel8 with an ABI 377 DNAsequencer and the use of Genescan software (Applied Biosystems,Perkin-Elmer).
SSCP Analysis and SequencingA total of 33 primer pairs (please see online Table I, which can beaccessed at http://atvb.ahajournals.org) were designed to evaluate thecoding region, promoter A, and partial promoter B regions of theER-� gene. Primers for the coding region flank the exons and allowfor analysis of the adjacent splice junctions. Single-strand confor-mation polymorphism (SSCP) was performed from 60 subjects asdescribed by Spinardi et al.20 Electrophoresis was performed on a10% to 20% gradient polyacrylamide gel (ATTO Corp) and run at100 V constantly for 10 to 15 hours in 0.5� TBE buffer at 3 differentconditions: (1) at 4°C without glycerol and (2) at room temperaturewith 5% glycerol. Fragments that showed a variant by SSCP werereamplified and sequenced with fluorescently labeled dideoxy ter-minators with the use of a Thermo Sequenase II Kit (AmershamPharmacia Biotech Inc) on an ABI 310 genetic analyzer (AppliedBiosystems, Perkin-Elmer).
Statistical AnalysisANOVA was used to evaluate the association between phenotypiccharacteristics and genotype groups, and the Student t test was usedwhen 2 groups were compared. The frequency distribution ofgenotypes was compared by using cross-tabulation and standard �2
tests. Linkage disequilibrium between the different polymorphismswas analyzed by the estimate of a haplotype algorithm (EM) by usingArlequin software.21 To compare observed genotype frequencieswith those expected under the Hardy-Weinberg equilibrium, contin-gency tables were used, with standard �2 tests. Logistic regressionanalysis was used to predict CAD from the genotype of poly-morphisms, with the known risk factors as covariates. Statview 5.0software was used.
ResultsCharacteristics of Study SubjectsThe mean ages for men and women were not significantlydifferent between the CAD and non-CAD groups (Table 1).The CAD group had a higher prevalence of conventionalcoronary risk factors (hypertension and diabetes for men;smoking, hypertension, and diabetes for postmenopausalwomen). Consistent with a previous report involving theJapanese population,22,23 B-variant polymorphism was notdetected in the present study.
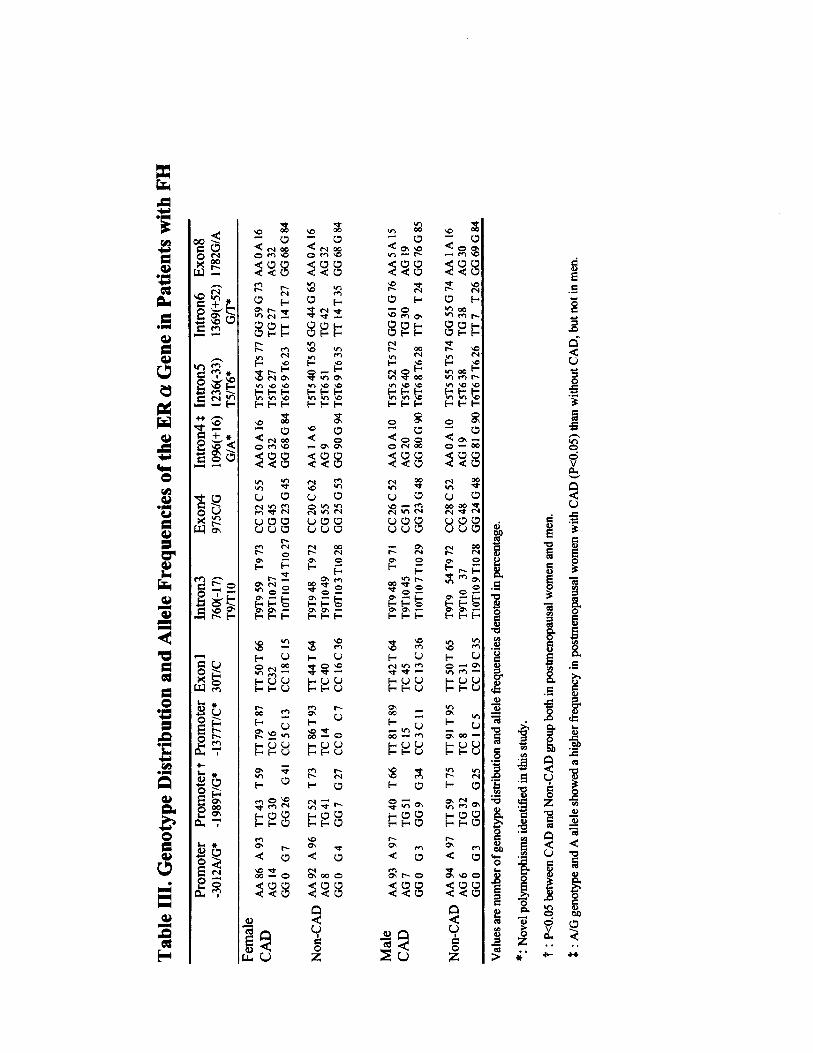
Single Nucleotide Polymorphisms in the ER-�Gene Detected by SSCP Analysis and SequencingWe identified 10 single nucleotide polymorphisms (Figure),including 4 that had been previously described in breastcancer patients,11,24 3 novel in introns, and 3 novel inpromoter B; all exonic single nucleotide polymorphisms were
818 Arterioscler Thromb Vasc Biol. May 2002
by guest on September 10, 2016
http://atvb.ahajournals.org/D
ownloaded from
synonymous. We identified a restriction enzyme whosecleavage pattern was altered by each variant or used amismatch technique allowing restriction fragment lengthpolymorphism analysis (see online Table II, which can beaccessed at http://atvb.ahajournals.org). The distribution ofgenotypes, T9/T10 in intron 3 and T5/T6 in intron 5, wasdetected by PCR-SSCP and sequencing. All the genotypefrequencies (please see online Table III, which can beaccessed at http://atvb.ahajournals.org) followed the Hardy-Weinberg equilibrium. A T/G transversion in promoter B(�1989 relative to the first transcribed nucleotide in ahomozygous for 14 TA repeats)9 was newly detected in thepresent study. The polymorphism of �1989T/G was in tightlinkage disequilibrium with the TA repeat, PvuII and XbaI(please see online Table IV, which can be accessed athttp://atvb.ahajournals.org), and it showed a significant asso-ciation with CAD in men (P�0.02) and in postmenopausalwomen (P�0.05), with a higher frequency of G/G genotypein the CAD group (P�0.03). There was no significantdifference in the severity of CAD or serum lipid levels,although HDL cholesterol was lower in subjects with the G/Ggenotype than in those with the T/T genotype (0.99�0.25versus 1.14�0.32 mmol/L, P�0.05). Of a novel polymor-phism (G/A in intron 4), G/A genotype and A allele fre-quency were significantly higher in postmenopausal womenwith CAD (P�0.03 and P�0.04, respectively) but not in men(please see online Table III). A stepwise logistic regressionanalysis with age, sex, body mass index, HDL cholesterol,and LDL cholesterol as covariates showed that the G/Ggenotype of �1989T/G (odds ratio [OR] 4.5, 95% CI 1.0 to19.5; P�0.04), hypertension (OR 2.4, 95% CI 1.2 to 7.7;P�0.03), and diabetes (OR 3.0, 95% CI 1.6 to 10.0; P�0.02)
were independent risks for CAD but that smoking and theX1/X1 genotype were not. When all the above confoundingfactors but HDL cholesterol and LDL cholesterol wereforced, the G/G genotype weakened the statistical signifi-cance (OR 3.2, 95% CI 0.8 to 12.3; P�0.09), whereas theX1/X1 genotype showed significant risk for CAD (OR 5.0,95% CI 1.2 to 21.4; P�0.03).
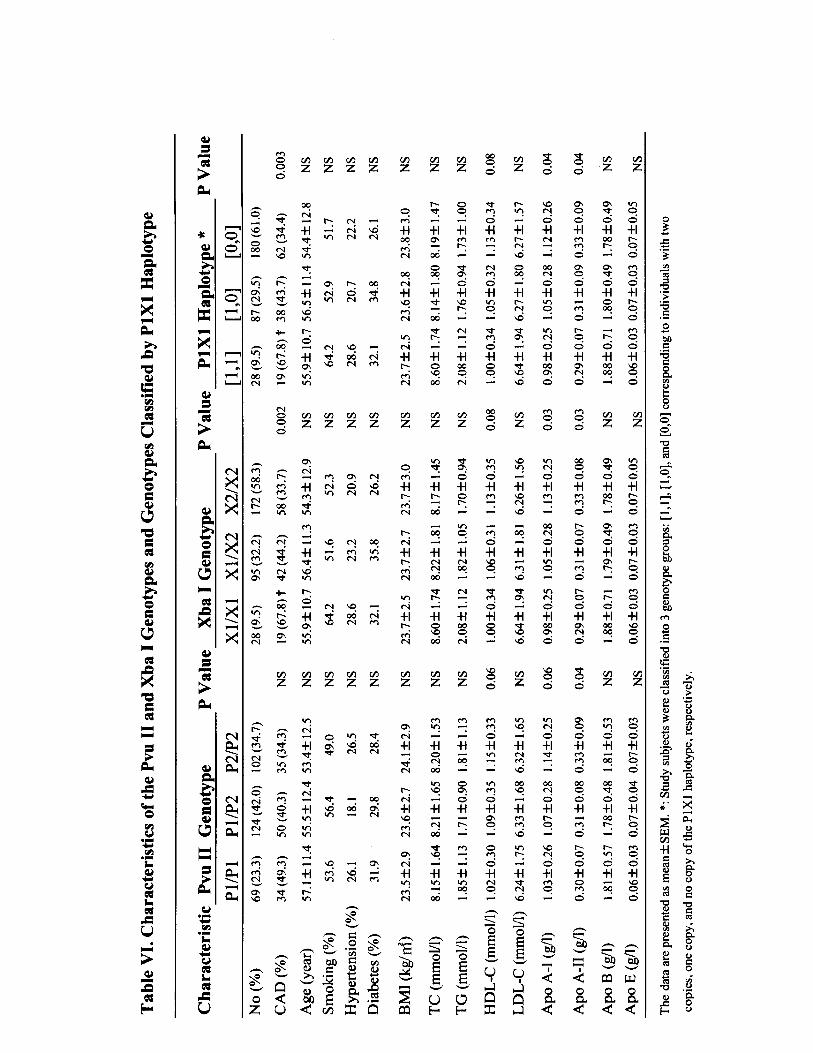
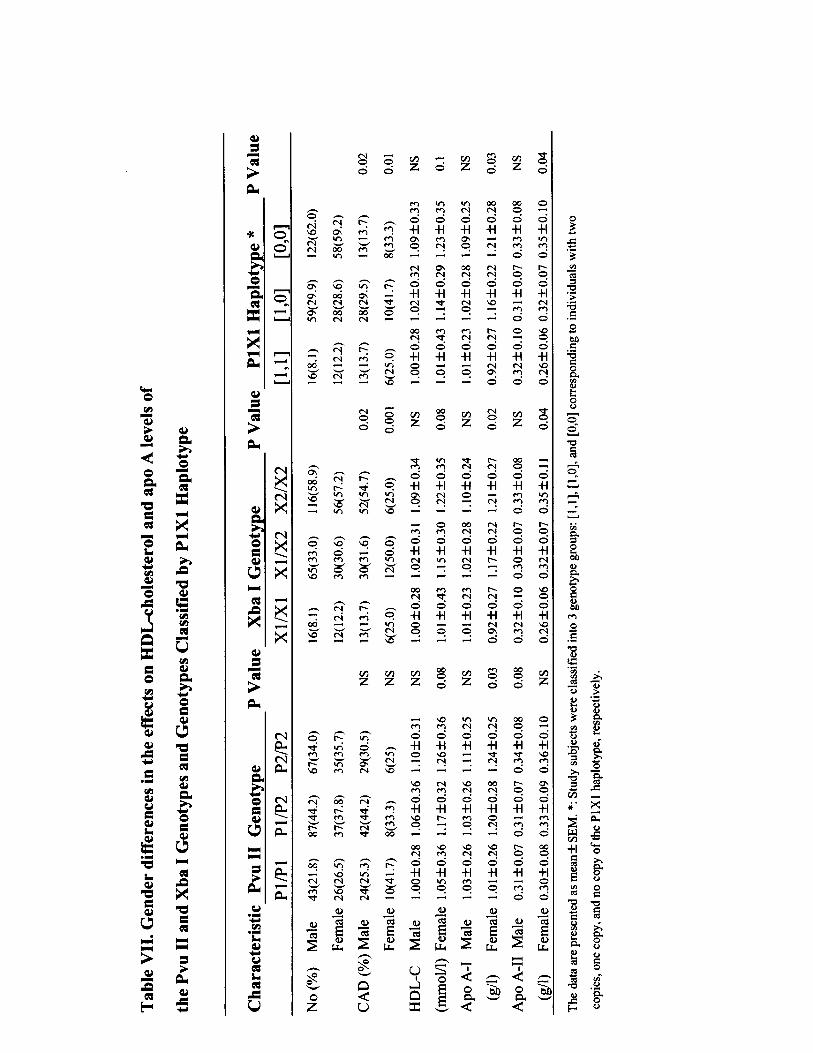
Effect of PvuII and XbaI Polymorphisms on CADThe distribution of genotypes was in Hardy-Weinberg equi-librium, and as previously reported,5,6,9,22 PvuII and XbaIwere tightly linked, so that a higher than expected number ofP1/P1 individuals were X1/X1 homozygotes and a higherthan expected number of P2/P2 individuals were X2/X2homozygotes (df 4, �2�160.33; P�0.0001). XbaI genotypedistributions were significantly different between the CADand non-CAD groups in postmenopausal women (P�0.001)and in men (P�0.02), with a higher frequency of X1/X1genotype in the CAD group (P�0.02). Serum HDL choles-terol, apoA-I, and apoA-II levels were significantly lower inpatients with the P1/P1 genotype than in those with the P2/P2genotype (P�0.04, P�0.03, and P�0.02, respectively) andin patients with the X1/X1 genotype compared with thosewith the X2/X2 genotype (P�0.05, P�0.02, and P�0.03,respectively). When genotypes were analyzed separately bysex, there was still a significant difference in HDL cholesterolas well as apoA-I for both polymorphisms and a significantdifference between XbaI genotypes and apoA-II (P�0.04) inpostmenopausal women but not in men. In addition, there wasa significant difference in the mean age at onset of menopauseamong the genotypes of PvuII and XbaI in postmenopausalwomen (please see online Table V, which can be accessed athttp://atvb.ahajournals.org). When we combined the 2 poly-morphisms, 3 major haplotype alleles were recognized: P2X2(54.0%), P1X1 (23.9%), and P1X2 (20.4%), thus demonstrat-ing that 2 copies of the P1X1 haplotype are present morefrequently than 1 or no copy of the P1X1 haplotype inpostmenopausal women and men with CAD (P�0.01 andP�0.03, respectively). Serum apoA-I and apoA-II levelswere significantly lower in patients with 2 copies of the P1X1haplotype than in those with 1 or no copy of the P1X1haplotype (P�0.02 and P�0.04, respectively), whereas HDLcholesterol showed a tendency to be lower in patients with 2copies of the P1X1 haplotype (P�0.07). When the P1X1haplotype was analyzed separately by sex, there was still asignificant difference for apoA-I and apoA-II in postmeno-pausal women (P�0.02 and P�0.04, respectively) but not inmen. No significant difference between P1X1 haplotype andHDL cholesterol was found for men or postmenopausalwomen (please see online Tables VI and VII, which can beaccessed at http://atvb.ahajournals.org).
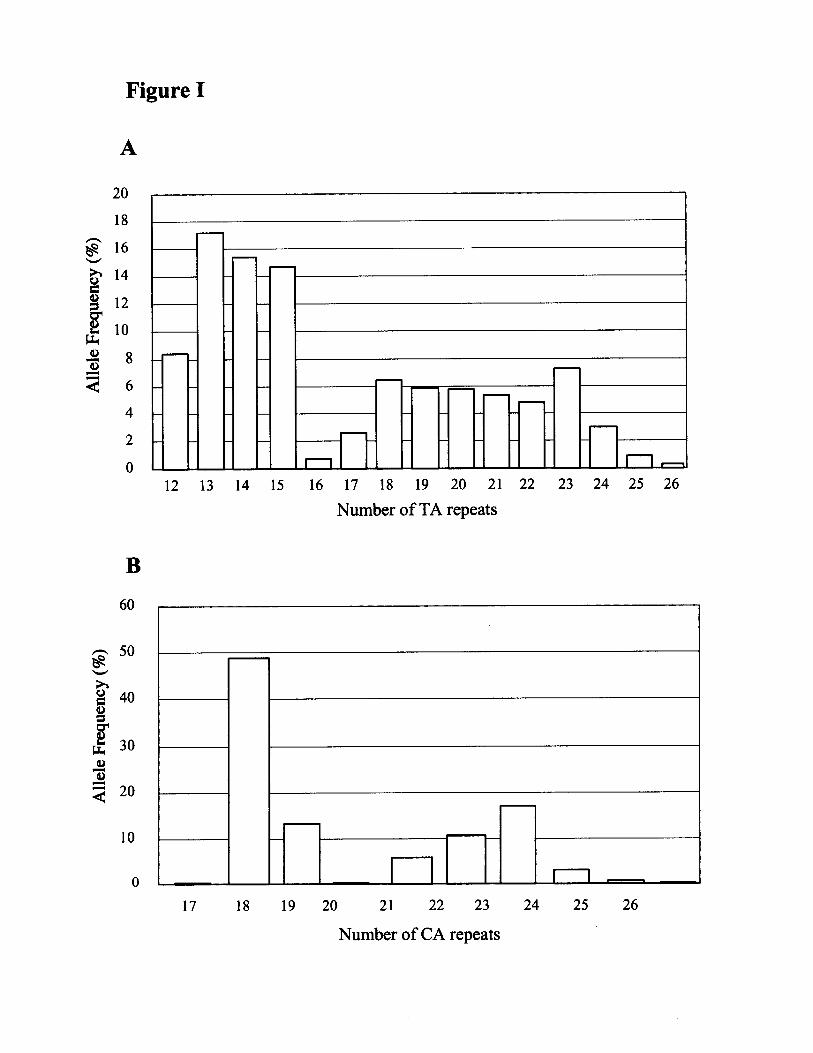
Effect of Dinucleotide Repeat Polymorphismson CADAccording to the bimodal distribution pattern of TA alleleswith a low distribution of 16 or 17 repeat alleles (please seeonline Figure IA, which can be accessed at http://atvb.ahajournals.org), subjects were classified into 3 groups: (1)
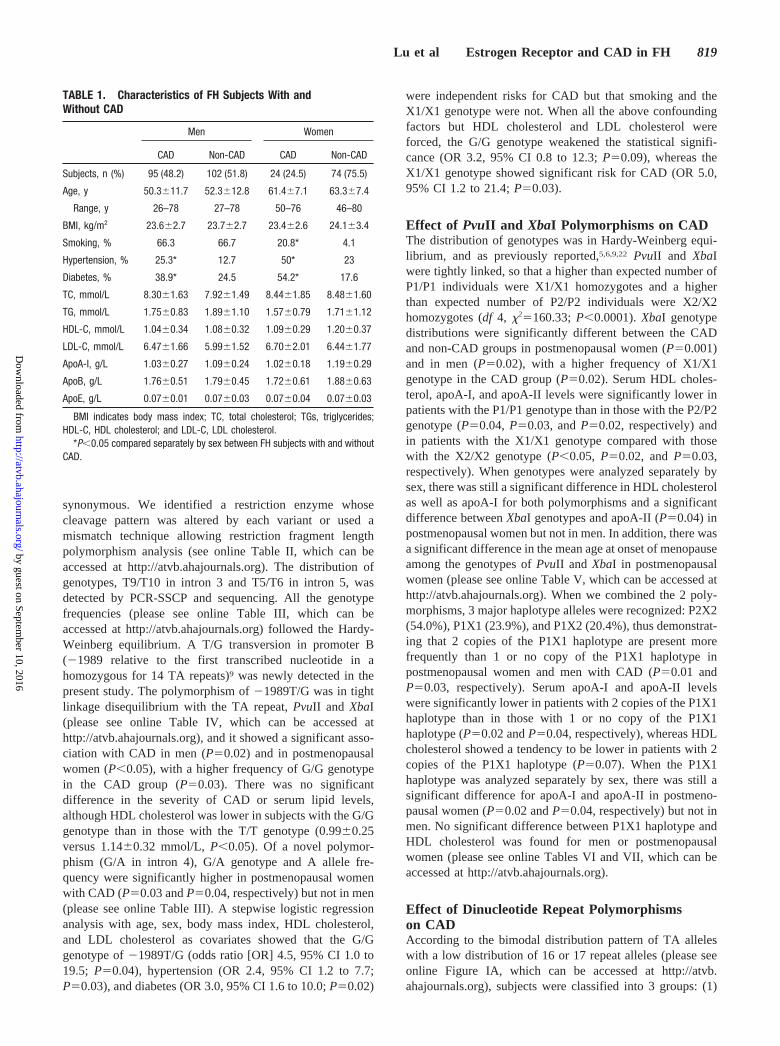
TABLE 1. Characteristics of FH Subjects With andWithout CAD
Men Women
CAD Non-CAD CAD Non-CAD
Subjects, n (%) 95 (48.2) 102 (51.8) 24 (24.5) 74 (75.5)
Age, y 50.3�11.7 52.3�12.8 61.4�7.1 63.3�7.4
Range, y 26–78 27–78 50–76 46–80
BMI, kg/m2 23.6�2.7 23.7�2.7 23.4�2.6 24.1�3.4
Smoking, % 66.3 66.7 20.8* 4.1
Hypertension, % 25.3* 12.7 50* 23
Diabetes, % 38.9* 24.5 54.2* 17.6
TC, mmol/L 8.30�1.63 7.92�1.49 8.44�1.85 8.48�1.60
TG, mmol/L 1.75�0.83 1.89�1.10 1.57�0.79 1.71�1.12
HDL-C, mmol/L 1.04�0.34 1.08�0.32 1.09�0.29 1.20�0.37
LDL-C, mmol/L 6.47�1.66 5.99�1.52 6.70�2.01 6.44�1.77
ApoA-I, g/L 1.03�0.27 1.09�0.24 1.02�0.18 1.19�0.29
ApoB, g/L 1.76�0.51 1.79�0.45 1.72�0.61 1.88�0.63
ApoE, g/L 0.07�0.01 0.07�0.03 0.07�0.04 0.07�0.03
BMI indicates body mass index; TC, total cholesterol; TGs, triglycerides;HDL-C, HDL cholesterol; and LDL-C, LDL cholesterol.
*P�0.05 compared separately by sex between FH subjects with and withoutCAD.
Lu et al Estrogen Receptor and CAD in FH 819
by guest on September 10, 2016
http://atvb.ahajournals.org/D
ownloaded from
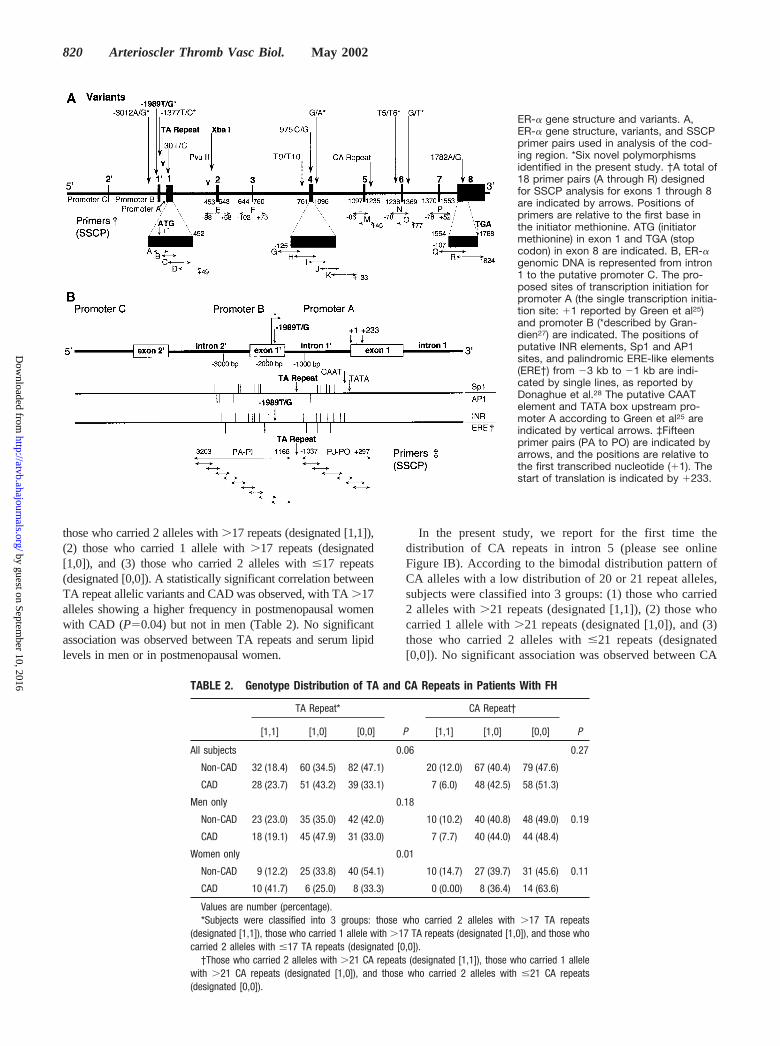
those who carried 2 alleles with �17 repeats (designated [1,1]),(2) those who carried 1 allele with �17 repeats (designated[1,0]), and (3) those who carried 2 alleles with �17 repeats(designated [0,0]). A statistically significant correlation betweenTA repeat allelic variants and CAD was observed, with TA �17alleles showing a higher frequency in postmenopausal womenwith CAD (P�0.04) but not in men (Table 2). No significantassociation was observed between TA repeats and serum lipidlevels in men or in postmenopausal women.
In the present study, we report for the first time thedistribution of CA repeats in intron 5 (please see onlineFigure IB). According to the bimodal distribution pattern ofCA alleles with a low distribution of 20 or 21 repeat alleles,subjects were classified into 3 groups: (1) those who carried2 alleles with �21 repeats (designated [1,1]), (2) those whocarried 1 allele with �21 repeats (designated [1,0]), and (3)those who carried 2 alleles with �21 repeats (designated[0,0]). No significant association was observed between CA
ER-� gene structure and variants. A,ER-� gene structure, variants, and SSCPprimer pairs used in analysis of the cod-ing region. *Six novel polymorphismsidentified in the present study. †A total of18 primer pairs (A through R) designedfor SSCP analysis for exons 1 through 8are indicated by arrows. Positions ofprimers are relative to the first base inthe initiator methionine. ATG (initiatormethionine) in exon 1 and TGA (stopcodon) in exon 8 are indicated. B, ER-�genomic DNA is represented from intron1 to the putative promoter C. The pro-posed sites of transcription initiation forpromoter A (the single transcription initia-tion site: 1 reported by Green et al25)and promoter B (*described by Gran-dien27) are indicated. The positions ofputative INR elements, Sp1 and AP1sites, and palindromic ERE-like elements(ERE†) from �3 kb to �1 kb are indi-cated by single lines, as reported byDonaghue et al.28 The putative CAATelement and TATA box upstream pro-moter A according to Green et al25 areindicated by vertical arrows. ‡Fifteenprimer pairs (PA to PO) are indicated byarrows, and the positions are relative tothe first transcribed nucleotide (1). Thestart of translation is indicated by 233.
TABLE 2. Genotype Distribution of TA and CA Repeats in Patients With FH
TA Repeat*
P
CA Repeat†
P[1,1] [1,0] [0,0] [1,1] [1,0] [0,0]
All subjects 0.06 0.27
Non-CAD 32 (18.4) 60 (34.5) 82 (47.1) 20 (12.0) 67 (40.4) 79 (47.6)
CAD 28 (23.7) 51 (43.2) 39 (33.1) 7 (6.0) 48 (42.5) 58 (51.3)
Men only 0.18
Non-CAD 23 (23.0) 35 (35.0) 42 (42.0) 10 (10.2) 40 (40.8) 48 (49.0) 0.19
CAD 18 (19.1) 45 (47.9) 31 (33.0) 7 (7.7) 40 (44.0) 44 (48.4)
Women only 0.01
Non-CAD 9 (12.2) 25 (33.8) 40 (54.1) 10 (14.7) 27 (39.7) 31 (45.6) 0.11
CAD 10 (41.7) 6 (25.0) 8 (33.3) 0 (0.00) 8 (36.4) 14 (63.6)
Values are number (percentage).*Subjects were classified into 3 groups: those who carried 2 alleles with �17 TA repeats
(designated [1,1]), those who carried 1 allele with �17 TA repeats (designated [1,0]), and those whocarried 2 alleles with �17 TA repeats (designated [0,0]).
†Those who carried 2 alleles with �21 CA repeats (designated [1,1]), those who carried 1 allelewith �21 CA repeats (designated [1,0]), and those who carried 2 alleles with �21 CA repeats(designated [0,0]).
820 Arterioscler Thromb Vasc Biol. May 2002
by guest on September 10, 2016
http://atvb.ahajournals.org/D
ownloaded from

repeats and CAD (Table 2) or serum lipid levels in men or inpostmenopausal women.
DiscussionIn the present study, we reported a polymorphism analysis ofthe ER-� gene in the coding region, promoter A region, andpartial promoter B regions in 295 patients with heterozygousFH, providing evidence that common genetic polymorphismswithin the ER-� gene are associated with the risk of CAD inpostmenopausal women and in men. The polymorphisms of�1989T/G in promoter B and XbaI in intron 1 are associatedwith an increasing risk of CAD in postmenopausal womenand in men with heterozygous FH. Our results suggest thatthe TA repeat might be associated with CAD in postmeno-pausal women. The present study suggests that commonallelic variants of the ER-� gene, existing in FH patients, maycause differential responsiveness to estrogen, although furtherexperiments will be required to define the mechanism.
We first reported 3 novel polymorphisms in promoter Band found a high degree of linkage disequilibrium between�1989T/G and the variable length of TA repeats,�3012A/G, �1377T/C, PvuII, XbaI, 30T/C, and T9/T10.Subjects with the G/G genotype showed a significantly highermean number of TA repeats and higher frequencies of�3012A/G, �1377C/C, P1/P1, X1/X1, 30C/C, and T10/T10genotypes (P�0.0001). No comparable data are actuallyreported for other populations, although the linkage disequi-librium between the TA repeat and the polymorphisms ofPvuII and XbaI was previously reported in white postmeno-pausal women.6,9 The discovery of such a high degree oflinkage disequilibrium in a group of 295 heterozygous FHindividuals may have important implications and may partlyexplain previous discrepancies among ER-� polymorphismstudies.5,6,9,10,22 The results from the present study indicatethat there is a relationship between �1989T/G or its linkedpolymorphisms at the ER-� gene locus and CAD. However,the molecular mechanism by which �1989T/G or its linkedpolymorphisms is associated with CAD remains unclear. Atleast 3 different promoters have been identified in thisgene.9,25–28 The first characterized promoter at the 5� end ofexon 1 was termed promoter A. It contains a TATA box anda CAAT element and possesses a single site of transcriptioninitiation at 1.25 Subsequently, sequencing of upstreamgenomic DNA revealed a region at position �1.9 kb with anadditional exon, denoted exon 1�, and an additional promoter,denoted promoter B.26 Several sites of transcription initiationare identified in this promoter.26,27 There are few putativeTATA box elements and no putative CAAT elements locatednear the proposed initiation sites, whereas a number ofinitiation response (INR) elements are present.28 Therefore, itis likely that transcription initiation is positioned via INRelements at this promoter. There are INR elements either at orjust upstream from 3 of the 4 transcription initiation sitesproposed by Grandien.27 There are several specificity protein1 sites, 1 activator protein 1 site, and several estrogenresponse element (ERE)-like elements in the vicinity of exon1�, which may augment transcription from promoter B.28
Promoter C, located �21 kb upstream from promoter A, hasbeen postulated recently.28 Transcription of the ER-� gene
from these promoters yields 3 different mRNA isoforms withunique 5� untranslated regions but identical coding regions.Several studies have identified that promoter A and B areused in some breast cancer cell lines and endometrium but notin liver.27,28 Expression of promoter C mRNA has beenpredominantly detected in liver, whereas only the promoter BmRNA expression has been detected in bone cells.27,28 Thefactors controlling the level of expression of ER-� are notwell characterized; however, these studies suggest that cell-and tissue-specific expression may be regulated by differen-tial promoter usage. Recent studies have demonstrated thathuman vascular smooth muscle cells (VSMCs) express ER-�mRNA and protein and that ER-� in human VSMCs iscapable of estrogen-dependent gene activation.29,30 However,no study has ever investigated the expression of differentpromoter-mRNA isoforms in VSMCs. As detected in thepresent study, �1989T/G is located 2 bp downstream from 1of the 4 identified transcription initiation sites in the promoterB reported by Grandien,27 which has several INR elementsand Sp1 sites, 1 AP1 site, and 1 palindromic ERE-likeelement just upstream from this polymorphism.27,28 More-over, the TA repeat between promoter A and B regions,which was reported to be a predictor of the risk in postmeno-pausal osteoporosis,9 is �0.8 kb downstream from�1989T/G, and a high degree of linkage disequilibrium wasobserved between the 2 polymorphisms. We speculate that�1989T/G, or its linked TA repeats or perhaps yet uniden-tified polymorphisms within the linkage disequilibrium re-gion, may directly or indirectly, by affecting promoter usagein VSMCs, influence the expression of the ER-� genethrough transcriptional and translational regulation. Furtherstudy is required to identify whether or not the differentmodulation of ER-� gene expression that occurs at thetranscription or translation level in human VSMCs associateswith a polymorphism of �1989T/G or its linkedpolymorphisms.
The atheroprotective effects of estrogens were attributed todirect effects on the vessel wall and indirect effects onlipoprotein metabolism.31,32 It is believed that estrogen leadsto an increase in the mRNA for the LDL receptor.33 Manystudies, including 1 large, randomized, controlled trial,32 havedocumented that estrogens decrease LDL cholesterol levelsand increase HDL cholesterol levels. And it is reported thatestrogens increase apoA-I production in hepatic cells byincreasing the transcription of the apoA-I gene.34 A currentstudy in 102 healthy Japanese school children has suggestedthat XbaI polymorphism might be related to LDL metabo-lism.35 Our results have shown that XbaI or its linkedpolymorphisms are associated with HDL and its principalapolipoproteins (apoA-I and apoA-II) in patients with het-erozygous FH, although no significant association with LDLwas found. This finding indicates that the ER-� gene mightplay a role in the transcriptional regulation of HDL metabo-lism in this special group of patients, whose primary defect isa mutation in the LDL receptor gene. There may be a trueeffect of XbaI or its linked polymorphisms on CAD, partiallythrough an indirect effect via HDL metabolism and through adirect effect on coronary arteries. ER-� gene polymorphisms
Lu et al Estrogen Receptor and CAD in FH 821
by guest on September 10, 2016
http://atvb.ahajournals.org/D
ownloaded from

in HDL metabolism appear to be sex specific, but the numberof subjects studied was too small for a definitive conclusion.
Our findings contradict those reported by Matsubara et al,22
who found no association between CAD and either of the 2polymorphisms (PvuII and XbaI). However, in the presentstudy, the X1 allele (25% versus 17%) and the X1/X1genotype (9% versus 2%) were more frequent than thoseinvestigated in their study. The controversy may be partiallyexplained by differences in the populations (FH patientsversus the general population) or in the mean ages of CADonset (50 versus 58 in men, and 61 versus 65 in postmeno-pausal women). It was reported that polymorphisms were notassociated with serum lipid levels, including HDL cholesterollevels in their study. However, their study did not investigateapoA-I. Weel et al36 reported that homozygous P1/P1 womenhad a significantly earlier onset of menopause and a higherrisk of surgical menopause than did homozygous P2/P2women. We also found an earlier onset of menopause forwomen who carried the P1/P1 genotype or the X1/X1genotype. Thus, the P1 or X1 allele appeared to be associatedwith an estrogen deficiency phenotype. We then speculatedthat the XbaI polymorphism might be a genetic risk factor forthe age of CAD onset in women. However, in the study ofMatsubara et al, the relationship between the 2 polymor-phisms and the onset of menopause was not investigated.Sample size in the present study was calculated a priori,considering an � error �0.05 and a � error �0.1. Thissuggests that our sample size was large enough to minimizetype I error and to assume a standard type II error for the XbaIpolymorphism but not for PvuII. The statistical power withonly 181 subjects in their study might have been too low toconclude that there was no association of XbaI or PvuIIpolymorphisms with CAD.
Interestingly, the studies in the Japanese populationshowed a striking difference in the genotype distribution fromwhite populations. B-variant polymorphism was not detectedamong the Japanese population, although its allele frequencyin white women has been reported to be 10% to 20%.23 Thedistribution of the X1/X1 genotype, which is likely associatedwith the estrogen deficiency phenotype in the Japanesepopulation, was significantly lower than that observed in thewhite populations.5,6,9,22 The relationship between these poly-morphisms and CAD has not been widely studied in largersamples, and conflicting results have been reported in similarstudies10,22 and in studies of osteoporosis.5,6,9 The results weobtained in the present analysis clearly indicate the need foranalyzing larger population samples in non-FH populationsbefore reaching final conclusions.
In conclusion, the present study suggests that the poly-morphisms in the ER-� gene, �1989T/G or its linkedpolymorphism XbaI, may confer risk for CAD in men and inpostmenopausal women with FH. Carriers of the G/G geno-type or its linked polymorphisms may benefit less from thecardiovascular protective effect of ER-� by its effect on HDLmetabolism as well as by direct effects on blood vessels.Further studies in a non-FH population and the discovery ofa functional molecular mechanism are necessary before�1989T/G might prove useful in the prediction of CAD.
AcknowledgmentsThis work has been supported by a scientific research grant from theMinistry of Education, Science, and Culture of Japan to H. Mabuchi(No. 09307010). We also thank Sachio Yamamoto and MihokoMizuno for technical assistance.
References1. Barrett-Connor E. Sex differences in coronary heart disease: why are
women so superior?: the 1995 Ancel Keys Lecture. Circulation. 1997;95:252–264.
2. Cowley SM, Parker MG. A comparison of transcriptional activation byER alpha and ER beta. J Steroid Biochem Mol Biol. 1999;69:165–175.
3. Yaich L, Dupont WD, Cavener DR, Parl FF. Analysis of the Pvu IIrestriction fragment-length polymorphism and exon structure of theestrogen receptor gene in the breast cancer and peripheral blood. CancerRes. 1992;52:77–83.
4. Andersen TI, Heimdal KR, Skrede M, Tveit K, Berg K, Borresen AL.Oestrogen receptor (ESR) polymorphisms and breast cancer suscepti-bility. Hum Genet. 1994;94:665–670.
5. Kobayashi S, Inoue S, Hosoi T, Ouchi Y, Shiraki M, Orimo H. Asso-ciation of bone mineral density with polymorphism of the estrogenreceptor gene. J Bone Miner Res. 1996;11:306–311.
6. Albagha OM, McGuigan FE, Reid DM, Ralston SH. Estrogen receptor�gene polymorphisms and bone mineral density: haplotype analysis inwomen from the United Kingdom. J Bone Miner Res. 2001;16:128–134.
7. Lehrer S, Rabin J, Kalir T, Schachter BS. Estrogen receptor variant andhypertension in woman. Hypertension. 1993;21:439–441.
8. Sano M, Inoue S, Hosoi T, Ouchi Y, Emi M, Shiraki M, Orimo H.Association of estrogen receptor dinucleotide repeat polymorphism withosteoporosis. Biochem Biophys Res Commun. 1995;217:378–383.
9. Becherini L, Gennari L, Masi L, Mansani R, Massart F, Morelli A,Falchetti A, Gonnelli S, Fiorelli G, Tanini A, Brandi ML. Evidence of alinkage disequilibrium between polymorphisms in the human estrogenreceptor� gene and their relationship to bone mass variation in postmeno-pausal Italian women. Hum Mol Genet. 2000;9:2043–2050.
10. Kunnas TA, Laippala P, Penttila A, Lehtimaki T, Karhunen PJ. Asso-ciation of polymorphism of human � oestrogen receptor gene with cor-onary artery disease in men: a necropsy study. BMJ. 2000;321:273–274.
11. Schubert EL, Lee MK, Newman B, King MC. Single nucleotide poly-morphisms (SNPs) in the estrogen receptor gene and breast cancer sus-ceptibility. J Steroid Biochem Mol Biol. 1999;71:21–27.
12. Dib C, Faure S, Fizames C, Samson D, Drouot N, Vignal A, MillasseauP, Marc S, Hazan J, Seboun E, Lathrop M, Gyapay G, Morissette J,Weissenbach J. A comprehensive genetic map of the human genomebased on 5264 microsatellites. Nature. 1996;380:152–154.
13. Smith EP, Boyd J, Frank GR, Takahashi H, Cohen RM, Specker B,Williams TC, Lubahn DB, Korach KS. Estrogen resistance caused by amutation in the estrogen-receptor gene in a man. N Engl J Med. 1994;331:1056–1061.
14. Sudhir K, Chou TM, Chatterjee K, Smith EP, Williams TC, Kane JP,Malloy MJ, Korach KS, Rubanyi GM. Premature coronary artery diseaseassociated with a disruptive mutation in the estrogen receptor gene in aman. Circulation. 1997;96:3774–3777.
15. Slack J. Risks of ischaemic heart disease in familial hyperlipopro-teinaemic states. Lancet. 1969;2:1380–1382.
16. Stone NJ, Levy RI, Fredrickson DS, Verter J. Coronary artery disease in116 kindreds with familial type II hyperlipoproteinemia. Circulation.1974;49:476–488.
17. Mabuchi H, Miyamoto S, Ueda K, Oota M, Takegoshi T, Wakasugi T,Takeda R. Causes of death in patients with familial hypercholesterolemia.Atherosclerosis. 1986;61:1–6.
18. Goldstein JL, Hobbs HH, Brown MS. Familial hypercholesterolemia. In:Scriver CR, Beaudet AL, Sly WS, Valle D, eds. The Metabolic andMolecular Bases of Inherited Diseases. 8th ed. New York, NY:McGraw-Hill; 2001:2863–2913.
19. Mabuchi H, Koizumi J, Shimizu M, Takeda R. Development of coronaryheart disease in familial hypercholesterolemia. Circulation. 1989;79:225–232.
20. Spinardi L, Mazars R, Theillet C. Protocols for an improved detection ofpoint mutations by SSCP. Nucleic Acids Res. 1991;19:4009.
21. Schneider S, Kueffer JM, Roessli D, Excoffier L. Arlequin Version 2.0:A Software for Population Genetics Data Analysis. Geneva, Switzerland:Genetics and Biometry Laboratory, University of Geneva; 2000.
822 Arterioscler Thromb Vasc Biol. May 2002
by guest on September 10, 2016
http://atvb.ahajournals.org/D
ownloaded from

22. Matsubara Y, Murata M, Kawano K, Zama T, Aoki N, Yoshino H,Watanabe G, Ishikawa K, Ikeda Y. Genotype distribution of estrogenreceptor polymorphisms in men and postmenopausal women fromhealthy and coronary populations and its relation to serum lipid levels.Arterioscler Thromb Vasc Biol. 1997;17:3006–3012.
23. Fujimoto J, Hirose R, Ichigo S, Sakaguchi H, Tamaya T. DNA poly-morphism in B-domain of the estrogen receptor-� among Japanesewomen. Steroids. 1998;63:146–148.
24. Zhang QX, Borg A, Wolf DM, Oesterreich S, Fuqua SA. An estrogenreceptor mutant with strong hormone-independent activity from a meta-static breast cancer. Cancer Res. 1997;57:1244–1249.
25. Green S, Walter P, Kumar V, Krust A, Bornert JM, Argos P, Chambon P.Human oestrogen receptor cDNA: sequence, expression and homology tov-erb-A. Nature. 1986;320:134–139.
26. Piva R, Gambari R, Zorzato F, Kumar L, del Senno L. Analysis ofupstream sequences of the human estrogen receptor gene. BiochemBiophys Res Commun. 1992;183:996–1002.
27. Grandien K. Determination of transcription start sites in the human estrogenreceptor gene and identification of a novel, tissue-specific, estrogenreceptor-mRNA isoform. Mol Cell Endocrinol. 1996;116:207–212.
28. Donaghue C, Westley BR, May FE. Selective promoter usage of thehuman estrogen receptor-� gene and its regulation by estrogen. MolEndocrinol. 1999;13:1934–1950.
29. Karas RH, Patterson BL, Mendelsohn ME. Human vascular smooth muscle cellscontain functional estrogen receptor. Circulation. 1994;89:1943–1950.
30. Losordo DW, Kearney M, Kim EA, Jekanowski J, Isner JM. Variableexpression of the estrogen receptor in normal and atheroscleroticcoronary arteries of premenopausal women. Circulation. 1994;89:1501–1510.
31. Farhat MY, Lavigne MC, Ramwell PW. The vascular protective effects ofestrogen. FASEB J. 1996;10:615–624.
32. The Writing Group for the PEPI Trial. Effects of estrogen or estrogen/progestin regimens on heart disease risk factors in postmenopausalwomen: the Postmenopausal Estrogen/Progestin Interventions (PEPI)Trial. JAMA. 1995;273:199–208.
33. Patrick TS, Yamamoto T, Goldstein JL, Brown MS. Increased mRNA forlow density lipoprotein receptor in livers of rabbits treated with 17-ethinylestradiol. Proc Natl Acad Sci U S A. 1986;83:792–796.
34. Lamon-Fava S, Ordovas JM, Schaefer EJ. Estrogen increases apoli-poprotein (apo) A-I secretion in Hep G2 Cells by modulating tran-scription of the apo A-I gene promoter. Arterioscler Thromb Vasc Biol.1999;19:2960–2965.
35. Kikuchi T, Hashimoto N, Kawasaki T, Uchiyama M. Association ofserum low-density lipoprotein metabolism with oestrogen receptor genepolymorphisms in healthy children. Acta Paediatr. 2000;89:42–45.
36. Weel AE, Uitterlinden AG, Westendorp IC, Burger H, Schuit SC,Hofman A, Helmerhorst TJ, van Leeuwen JP, Pols HA. Estrogen receptorpolymorphism predicts the onset of natural and surgical menopause.J Clin Endocrinol Metab. 1999;84:3146–3150.
Lu et al Estrogen Receptor and CAD in FH 823
by guest on September 10, 2016
http://atvb.ahajournals.org/D
ownloaded from

and Hiroshi MabuchiHong Lu, Toshinori Higashikata, Akihiro Inazu, Atsushi Nohara, Wenxin Yu, Masami Shimizu
in Patients With Familial Hypercholesterolemia Gene Polymorphisms With Coronary Artery DiseaseαAssociation of Estrogen Receptor-
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2002 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/01.ATV.0000014424.18209.212002;22:817-823; originally published online March 7, 2002;Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/22/5/817World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org/content/suppl/2002/06/27/22.5.817.DC1.htmlData Supplement (unedited) at:
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. AnswerPermissions and Rights Question andunder Services. Further information about this process is available in the
permission is being requested is located, click Request Permissions in the middle column of the Web page whichCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on September 10, 2016
http://atvb.ahajournals.org/D
ownloaded from















![[Comparison of three different methods of axial rotation assessment in idiopathic scoliosis]](https://img.pdfslide.net/doc/110x75/634e5d324f29425739097d6d/comparison-of-three-different-methods-of-axial-rotation-assessment-in-idiopathic.jpg)

















