Embed Size (px)
Citation preview

Astrocytic and Microglia Cells ReactivityInduced by Neonatal Administration ofGlutamate in Cerebral Cortex of theAdult Rats
A. Martınez-Contreras,1,2 M. Huerta,4 S. Lopez-Perez,1,3 J. Garcıa-Estrada,5,6
S. Luquın,6 and C. Beas Zarate1,3*1Lab de Neuroquımica, Div de Neurociencias, C.I.B.O., Guadalajara, Jal. Mexico2HGZ-10, Colima, IMSS, Colima, Col., Mexico3Depto. de Biol. Cel. y Molec. C.U.C.B.A., Guadalajara, Jal. Mexico4C.U.I.B., U. de Colima, Guadalajara, Jal. Mexico5Lab. de Neuroendocrinologıa celular, Div. Neurociencias, C.I.B.O., IMSS, Guadalajara, Jal. Mexico6Depto. De Neurociencias, C.U.C.S., U de G., Guadalajara, Jal. Mexico
Recent studies confirm that astrocytes and neurons areassociated with the synaptic transmission, particularly withthe regulation of glutamate (Glu) levels. Therefore, they havethe capacity to modulate the Glu released from neuronsinto the extracellular space. It has also been demonstratedan intense astrocytic and microglia response to physical orchemical lesions of the central nervous system. However,the persistence of the response of the glial cells in adultbrain had not been previously reported, after the excitotoxicdamage caused by neonatal dosage of monosodium glu-tamate (MSG) to newborn rats. In this study, 4 mg/g bodyweight of MSG were administered to newborn rats at 1, 3,5, and 7 days after birth, at the age of 60 days the astro-cytes and the microglia cells were analyzed with immuno-histochemical methods in the fronto-parietal cortex. Doublelabeling to glial fibrillary acidic protein (GFAP) and BrdU, orisolectin-B4 and BrdU identified astrocytes or microgliacells that proliferated; immunoblotting and immunoreactiv-ity to vimentin served for assess immaturity of astrocyticintermediate filaments. The results show that the neonataladministration of MSG-induced reactivity of astrocytes andmicroglia cells in the fronto-parietal cortex, which was char-acterized by hyperplasia; an increased number of astro-cytes and microglia cells that proliferated, hypertrophy; in-creased complexity of the cytoplasm extension of both glialcells and expression of RNAm to vimentin, with the pres-ence of vimentin-positive astrocytes. This glial response toneuroexcitotoxic stimulus of Glu on the immature brain,which persisted to adulthood, suggests that the neuro-transmitter Glu could trigger neuro-degenerative illnesses.© 2002 Wiley-Liss, Inc.
Key words: glutamate; neuroexcitotoxicity; astrocytes;microglia; cerebral cortex
The synaptic excitatory transmission in the centralnervous system (CNS) of mammals is mainly mediated by
glutamate (Glu) (Curtis and Johnson, 1974). The release oflarge amounts of this neurotransmitter and the excessiveactivation of its receptors induce excitotoxicity (Olney,1971). When Glu is administered systemically during theneonatal period, it produces excitotoxic damage in variousbrain regions of rats (Olney, 1980a; Lorden and Sims,1987). Young animals are more susceptible to the toxiceffects of Glu than are adults (Olney, 1978, 1980b).
The cerebral cortex and hippocampus contain manyN-methyl-D-aspartate (NMDA) receptors (Young et al.,1991), and both regions may be more vulnerable to neu-rotoxicity induced by L-Glu; this is manifested by abnor-malities in different neurotransmission parameters, mainlyof the GABAergic type (Beas-Zarate et al., 1998).
The astroglia has been observed to be important insome physiological and pathological processes in the CNS(Bignami and Dahl, 1995) and, under normal or patho-logical conditions, the astrocytes may be immunohisto-chemically detectable with GFAP (Amenta et al., 1998).Several distinctive characteristics are shown by astrocytesin the neurological damage, such as an intense proliferativeresponse, enlargement, and greater complexity of cyto-plasmic branching; additionally, molecules that normallyare not expressed are resynthesized (Del Rıo-Ortega,1927; Penfield, 1932; Eddlesston and Mucke, 1993). Mostof the information regarding astrocyte types and their
Contract grant sponsor: CONACyT; Contract grant number: 30901-M;Contract grant sponsor: FOFOI; Contract grant sponsor: 0038/204; Con-tract grant sponsor: SIMORELOS; Contract grant number: 19990302034.
*Correspondence to: Dr. Carlos Beas Zarate, Lab. of Neurochemistry,Division of Neuroscience, C.I.B.O., IMSS, APDO Postal 4-160, Guadala-jara Jal., 44421, Mexico.E-mail: [email protected] or [email protected]
Received 6 June 2001; Revised 10 June 2001; Accepted 24 October 2001
Published online 13 December 2001
Journal of Neuroscience Research 67:200–210 (2002)
© 2002 Wiley-Liss, Inc.DOI 10.1002/jnr.10093

properties, however, come from in vitro studies using cellcultures (Hansson et al., 2000).
In the synaptic transmission of Glu, both astrocytes andneurons participate together (Hansson et al., 1997; Laming etal., 1998; Araque et al., 1999). Because the astroglia expressionotropic and metabotropic receptors to Glu (Steinhauserand Gallo, 1996), astrocytes possess a high affinity transportsystem to take up the Glu released into the intersynaptic space(Brown, 1999). Astrocytes appear to be basic elements for themaintenance of physiological concentrations of Glu throughintracytoplasmic metabolism of this neurotransmitter; thesecells also exert neuroprotective functions mediated by differ-ent neurotrophic factors such as IGF-I (Venters et al., 2000).In addition, the neuroexcitotoxicity induced in vitro by Gluproduces astrocytic edema (Schneider et al., 1992; Bender etal., 1998). The increased volume of astrocytes reduces theextracellular space and, consequently, alters the concentrationof some neuroactive substances so that neuronal excitability isaffected (Hansson et al., 2000). Furthermore, a deficientastrocytic Glu uptake from the extracellular space is a cause ofexcitotoxicity (Norenberg, 1998). An intense reactive astro-cytosis associated with neuronal degeneration has been ob-served in some neurodegenerative illnesses. This finding istypical of this type of pathological process (Lafarga et al.,1993). Immature brains, however, have been shown to beless reactive against injury stimulus, possibly due to absence ofinflammatory cytokines that contribute to the gliosis devel-opment (Balasingam et al., 1994).
Other glial cells that can rapidly respond to neuronaldamage are the microglia; they may release neurotrophicor neurotoxic factors in response to specific neuronalsignals. Therefore, from microglia cell cultures high con-centrations of Glu (Ramırez-Exposito and Martınez-Martos, 1998) can be released. This could mean thatmicroglia cells are involved in the neuronal death process,which is mediated by the activation of NMDA type re-ceptors, in a similar manner to the processes in ischemia,cerebral damage, or trauma (Meldrum and Garthwaite,1990; Ramırez-Exposito and Martınez-Martos, 1998).Despite numerous in vitro studies of the neuroexcitotox-icity of Glu, there are few studies examining the responseof glia against their neuroexcitotoxic effects on in vivosystems. The glial response resulting from early exposureto high Glu concentrations, particularly in brain regionsthat possess an important glutamatergic innervation, suchas the fronto-parietal cortex (Young et al., 1991), have notbeen described previously regarding its persistent effects inadulthood.
Previous findings using this model of study showedthat neonatal exposure to monosodium glutamate induceda reduced cellular number, and shorter and less ramifieddendritic processes in fronto-parietal pyramidal neurons ofadult rats (Gonzalez-Burgos et al., 2001). Thus, the aim ofthis study was to evaluate the response of the microglia andastrocytic cells in cerebral cortex of rats sacrificed 60 daysafter the neonatal subcutaneous administration of Glu (4mg/g of body weight). A quantitative analysis of thenumber and morphological transformation of the astro-
cytes and microglia cells was carried out. The proliferativeresponse of these cells against the early excitotoxic stimuliinduced by Glu was also studied. These results will permitus to develop an in vivo model for the study of themechanisms of neuroexcitotoxicity implied in some neu-rodegenerative illnesses (Spokes, 1981; Delacourte, 1990;Banati and Beyreuther, 1995; Monzon et al., 2000).
MATERIALS AND METHODS
Preparation of Animals
Newborn male and female Swiss-Wistar rats from ourin-house colony were injected s.c. with 4 mg/g body weight ofmonosodium glutamate (MSG) at 1, 3, 5, and 7 days after birth.Throughout the experiment, the animals were kept in a 12:12 hrlight:dark cycle at 22 � 2°C and 55% relative ambient humidity,with free access to balanced food (Ralston rations, Ralston,Purina, USA) and potable water. The rats were housed inpolycarbonate cages in groups of four or five, all had reached60 days of age when sacrificed.
When the rats were 14 days old, one group was slightlyanesthetized with anhydrous ether inhalation, to be implanted inthe dorsal subcutaneous region with 1 mL of 0.1 M Tris-HCI,pH 7.6, containing 1.25% agar and 28 mg 5�-bromo-2�-deoxyuridine (BrdU), corresponding to a daily doses of 50 mg/kgof body weight. This total dose was calculated to guaranteecontinuous delivery of BrdU for 14 days. BrdU is analogous tothymidine that incorporates to a recently synthesized DNAchain. It is completely metabolized during 4 hr through hepaticactivity; thereby it has been widely used as an exogenous markerto cell proliferation (Del Rio and Soriano, 1989).
Intravascular Perfusion to Blood Elimination and BrainFixation
At 60 days of age, all the control and experimental ratswere sacrificed by perfusion. The animals were deeply anesthe-tized with a lethal dose of intraperitoneal sodium pentobarbital(50 mg/kg) and were perfused with 180 mL of 0.9% NaCl with10 U/mL of heparin and 0.01% procaine (v/v) for approxi-mately 2 min at body temperature. This solution was followedby perfusion with 280 mL of 4% paraformaldehyde in 0.1 Mphosphate buffer, pH 7.4, for 10 min. The brain was removedby craniotomy and was additionally fixed by immersion for12 hr at 4°C in the same fixative solution. The brain was thenimmersed for 48 hr at 4°C in a cryoprotective solution of 30%sucrose, 1% arabic gum in 0.9% NaCl (until the brain hadbecome completely sunken). Coronal slices were cut in 40 �mthick sections on a freezing microtome and deposited in 0.1 Mphosphate buffer saline (PBS, 0.9% NaCl in 0.1 M phosphates,pH 7.4) to be washed before being immersed in an antifreezepreserving solution (30% dextrose, 60% ethyleneglycol, and 10%polyvinylpyrrolidone). The tissues were preserved in this solu-tion until the beginning of immunohistochemical procedures.
For quantitative morphological analyses of cerebral cortex,coronal slices were cut at ca. �0.3–4.30 mm from the bregma,according to the atlas of Paxinos and Watson (1998). Chosentissues were immunohistochemically processed by the free-floating procedure. To avoid technical variations samples ofcontrol and experimental animals were processed in parallel.
Non-Neuronal Cell Reactivity Induced by Glutamate 201

Unmasking treatments to reduce background for unspe-cific reactivity of the tissues was accomplished with an electro-chemical chamber (18 cm long � 15 cm wide � 13 cm high),with two cylindrical mineral coal electrodes (11 cm long �8 mm diameter), placed in contralateral position. Tissues wereimmersed in 500 mL of 0.1 M PBS, pH 7.3, at room temper-ature, then a 110 V, 6 A alternating current was applied for15 min, until the temperature reached 60°C. The same processwas repeated again when the PBS temperature had lowered. Inour experience, this procedure obviated prolonged incubationswith blocking solutions, which could result in damage of tissues.
Glial Fibrillary Acidic Protein Immunohistochemistryto Astrocytes
The sections were incubated for 15 min with 10% meth-anol containing 3% hydrogen peroxide to inhibit the endoge-nous peroxidase activity. Sections were rinsed twice in 0.1 MPBS for 15 min with gentle shaking, and incubated for 2 hr atroom temperature in a blocking solution of 0.1 M PBS con-taining 3% normal swine serum, 0.2 M glycine, 0.2 M DL-lysine,0.2% gelatin, 0.02% sodium azide, and 0.2% Triton X-100. Thesections were then incubated overnight in polyclonal rabbitantibodies (ab) raised against bovine spinal cord glial fibrillaryacidic protein (GFAP) (1st ab Dako Z0334) diluted 1:1,000 at4°C floating in glass vials.
The next day, sections were rinsed three times for 15 minin 0.1 M PBS with 0.2% Triton X-100, and then the sectionswere incubated at room temperature in swine IgG raised againstrabbit IgG diluted 1:250 (2nd ab Dako Z0196) dissolved in0.1 M PBS containing 0.2% gelatin, 0.2% Triton X-100, and 3%normal swine serum for 2.5 hr. After rinsing three times for15 min in 0.1 M PBS, slices were incubated in darkness at roomtemperature for 2 hr in the rabbit monoclonal peroxidase–antiperoxidase complex (PAP Dako Z-113) diluted 1:250 todetect the first ab. After the slices were rinsed twice in 0.1 MPBS with 0.2% Triton X-100 for 5 min, and then in 0.1 M PBSfor 10 min, bound peroxidase was visualized with 0.07% dia-minobenzidine-tetrahydrochloride as the chromogen (DAB),0.01% hydrogen peroxide in 0.1 M phosphate buffer (pH 7.3)for 15–20 min. The reaction was stopped when a brown colorhad developed. Immunostaining was absent when the first an-tiserum was omitted. Tissues were briefly rinsed in 0.1 M PBS,pH 7.4, (three times) and the samples were mounted in Entellanresin (Merck, Darmstadt, Germany) for microscopic analysiswith an automatic image analyzer (Leica 500IW Imaging work-station, Leica Inc., Deerfield, IL).
Microglia Identification
Slices were unmasked in the electrochemical chamber andendogenous peroxidase activity was inhibited by incubating thetissues for 15 min with 10% methanol containing 3% hydrogenperoxide. Sections were rinsed in 0.1 M PBS (2 � 15 min) andincubated for 10 min in a cationic solution containing 0.1 mMCaCl2, 0.1 mm MgCl2, 0.1 mM MnCl dissolved in 0.1 M PBS,pH 7.4). After being rinsed in 0.1 M PBS (2 � 15 min), thetissues were incubated overnight at 4°C in isolectin-B4 (Griffoniasimplicifolia) peroxidase labeled (Sigma, St. Louis, MO; L-5391),dissolved in 0.1 M PBS, pH 7.3, at a 1:100 dilution. After threewashes in buffer, peroxidase activity was visualized with 0.07%
3,3�-diaminobenzidine (Sigma) in 0.1 M phosphate buffer (pH7.3) with 0.01% hydrogen peroxide, for 10–15 min. In a givenexperiment, tissues for all the groups were incubated in parallel.
Labeling the Mitotic Cells With Bromodeoxyuridine
To investigate the distribution of mitotic cells, one groupof the animals were subcutaneously implanted with BrdU dis-solved in agar on Day 14 of postnatal age, (1 mL of 0.1 MTris-HCI, pH 7.6, containing 1.25% agar and 28 mg BrdU).
Under deep pentobarbital anesthesia (50 mg/kg, i.p.), therats were perfused via the left ventricle and the brain wasremoved by craniotomy and additionally fixed by immersion inthe same fixative solution (4% paraformaldehyde in 0.1 Mphosphate buffer, pH 7.4) for 12 hr at 4°C. The brains wererinsed three times in 0.1 M PBS for 30 min at room tempera-ture, then they were cryoprotected, and 40-�m coronal sliceswere prepared in a freezing microtome, deposited in 0.1 M PBSto be gently washed for GFAP or isolectin-B4 immunohisto-chemistry.
BrdU Immunohistochemical Staining
The immunohistochemical detection of BrdU was initi-ated with inhibition of the peroxidase activity because of theprevious GFAP or isolectin-B4 immunostaining, as describedabove. After three rinses in 0.1 M PBS for 20 min, the sectionswere immersed in 0.1 N HCl in distilled water, to be passedthrough ice-cold 0.1 N HCl for 10 min at 4°C, followed byimmersion in 2 N HCl in 0.05 M PBS at 37°C for 10–15 minto complete the DNA denaturation, which was monitored bystaining some control sections with 0.01% acridine orange indistilled water and examining them in a microscope equippedwith an epifluorescence module. When viewed with a blue filter(excitation wavelength 450–495 nm), double-stranded DNAfluoresced green, whereas denatured single-stranded DNA ap-peared red.
After the denaturation was completed, the preparationswere rinsed for 10 min in 0.1 M borate buffer (pH 8.5) toneutralize HCl. The sections were additionally rinsed for 10 minin 0.1 M PBS (pH 7.2) and incubated for 2 hr at room tem-perature in a blocking solution containing 0.2 M glycine, 0.2 Mlysine, 10% normal horse serum, 0.2% gelatin, 0.05% TritonX-100 and 0.02% sodium azide in 0.1 M PBS. After rinsing for15 min in 0.1 M PBS (pH 7.2) with 0.05% Triton X-100 atroom temperature, the sections were incubated overnight inmoist chambers at 4°C with mouse monoclonal anti-BrdUantiserum (Dako M 744) diluted 1:50 with 0.2% gelatin, 0.05%Triton X-100, 0.02% sodium azide and 5% normal horse serumin 0.1 M PBS.
The next day the samples were rinsed three times for10 min in 0.1 M PBS (pH 7.2) with 0.2% Triton X-100, beforeincubation in a horse anti-mouse IgG biotinylated antiserum(Vector Laboratories, Burlingame, CA, BA-2000) diluted 1:200in 0.1 M PBS (pH 7.2) with 0.2% Triton X-100, 0.2% gelatin,0.02% sodium azide and 5% normal horse serum for 2 hr at 18°Cin moist chambers. After three rinses in 0.1 M PBS (pH 7.2)with 0.5% Triton X-100 for 10 min, tissues were stained by theavidin-biotin-peroxidase complex (ABC) method (Vector PK-4000). The sections were incubated for 2.5 hr at room temper-ature in ABC (Vector) as indicated by the manufacturer in 0.1 M
202 Contreras et al.
PBS (pH 7.2), with 0.2% Triton X-100, 0.2% gelatin, 5%normal horse serum, and 5% dehydrated skim milk. After rinsingwith 0.1 M PBS (pH 7.2) for 30 min followed by rinsing in0.1 M buffer Tris-HCl (pH 7.6) the tissue bound peroxidase wasvisualized by incubation in 0.03% diaminobenzidine-tetrahydrochloride (DAB), 0.01% hydrogen peroxide, and 0.1%nickel ammonium sulfate (DAB-Ni) in 0.1 M Tris-buffer (pH8.2) for 10–15 min at room temperature. The sections wererinsed in 0.1 M Tris-HCl (pH 8.5) for 10 min and dehydrated,passed through xylene and mounted in Entellan (Merck). Slideswere viewed in a 100� oil-immersion objective.
As a result of revealed peroxidase activity with DAB, orDAB using nickel ammonium sulfate to obtain a dark bluepurple color, examination of proliferative double-labeled cells(astrocytes or microglia) showed brown stain in the cytoplasmand their extensions (GFAP or Isolectin-B4), and dark blue todark purple stain in the cell nucleus (BrdU-immunoreactivity).The number of cells with simple label and the number ofdouble-labeled cells were recorded separately. Labeling index wascalculated as number of BrdU-labeled GFAP-immunoreactiveastrocytes/total number of GFAP immunoreactive astrocytes �100. Immunostaining was absent when the first antisera wereomitted. Sets of sections for all the experimental groups wereincubated in parallel at the same time.
Morphometric Analysis
Immunostained sections of the fronto-parietal cortex wereanalyzed with light microscopy to quantify the number of GFAPlabeled astrocytes and Isolectin-B4 stained microglia, or double-labeled; GFAP � BrdU and Isolectin-B4 � BrdU cells. Incoronal slices of cerebral cortex of each intact rat and thoseinjected with MSG, 200 microscopic fields randomly wereexamined at 40� enlargement (0.78 mm2 of tissue), through thecomplete cortical thickness. Only those cells showing theirnucleus in the same plane of focus of the section were recorded.The morphological transformation of astrocytes GFAP-immunopositive and microglia was assessed with the aid of astereological graticule placed on the cellular soma and theircytoplasmic visible prolongations (Sterio et al., 1984). The grati-cule was superimposed on the image of the GFAP or isolectinB4-immunoreactive cells by using a microscope fitted with acamera lucida attachment. This graticule consisted of sevenconcentric circles with 28 �m of distance between each circle,the cellular nucleus was positioned in the center of the graticule,to register the total number of sites in which the cellular pro-longations intersected with the concentric lines. Additionally; toassess the 3D cytoplasmic arrangement of the microglia cells,drawings were elaborated at 100�, varying the focal planes forentire reconstruction of these cells, which is not possible in afixed focal plane.
Statistical Analysis
All the material was analyzed blind and were compared todata obtained from the control rats with those from animalstreated with MSG. Simple statistical comparisons were carriedout with a one-way ANOVA. The Student’s t-test was used forthe post-hoc comparison of individual mean values with P �
0.05 considered significant. Data shown represents mean �SEM.
Characterization of Astrocytic Immature Phenotype
Western blot for vimentin analysis. Total proteinextracts were prepared by homogenization of cerebral cortex
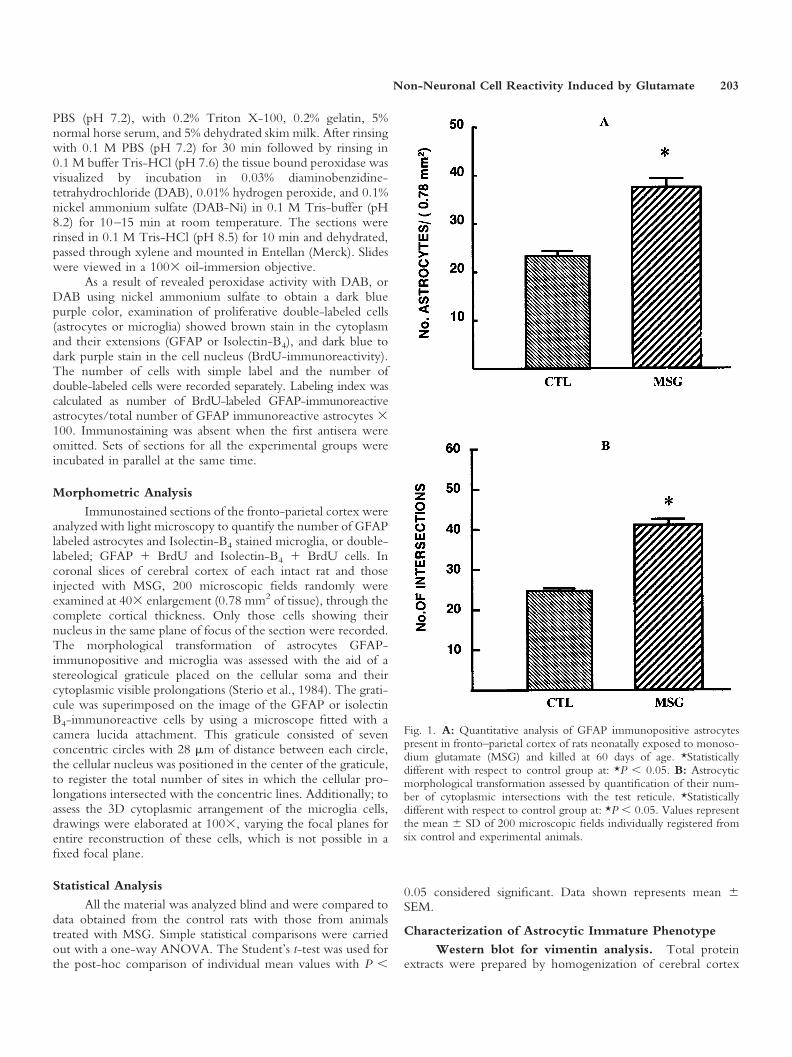
Fig. 1. A: Quantitative analysis of GFAP immunopositive astrocytespresent in fronto–parietal cortex of rats neonatally exposed to monoso-dium glutamate (MSG) and killed at 60 days of age. *Statisticallydifferent with respect to control group at: *P � 0.05. B: Astrocyticmorphological transformation assessed by quantification of their num-ber of cytoplasmic intersections with the test reticule. *Statisticallydifferent with respect to control group at: *P � 0.05. Values representthe mean � SD of 200 microscopic fields individually registered fromsix control and experimental animals.
Non-Neuronal Cell Reactivity Induced by Glutamate 203
obtained from control and MSG-treated animals in a sodiumdodecyl sulfate (SDS) buffer (20% glycerol, 4.6% SDS, 125 mMTris-HCl), pH 6.8. Protein concentrations were determinedaccording to Lowry et al. (1951). The standard consisted ofbovine serum albumin. Samples were incubated (3 min, 100°C)with 5% 2-mercaptoethanol, 50 �g of protein were deposited toeach lane in a 10% polyacrylamide gel. Electrophoresis wascarried out at 20 mA. Proteins in the gel were electroblottedonto nitrocellulose membranes (Hybond™-C pure, AmershamPharmacia Biotech, Piscataway, NJ). The membranes were in-cubated for 1 hr at room temperature with a blocking solution(TTBS) containing 10 mM Tris (pH 7.4), 150 mM NaCl, 0.01%
Tween 20, 2% BSA and 2% skim milk. They were then incu-bated overnight at 4°C with a monoclonal antiserum anti-vimentin (Dako Immunochemicals, Carpinteria, CA; M0725)diluted 1:2,000, followed by incubation with the secondaryantibody; horse anti-mouse IgG-biotinylated, diluted 1:4,000(Vector). The immunoblot density was a protein 57 kDa visu-alized and analyzed with a personal computer using a Kodakdigital Science 1D software (Eastman Kodak, Rochester, NY).Three control and three experimental animals were analyzed.Brain determinations were made five times for triplicate. Re-sultant data were normalized to be expressed as percentage forthe comparison between both groups.
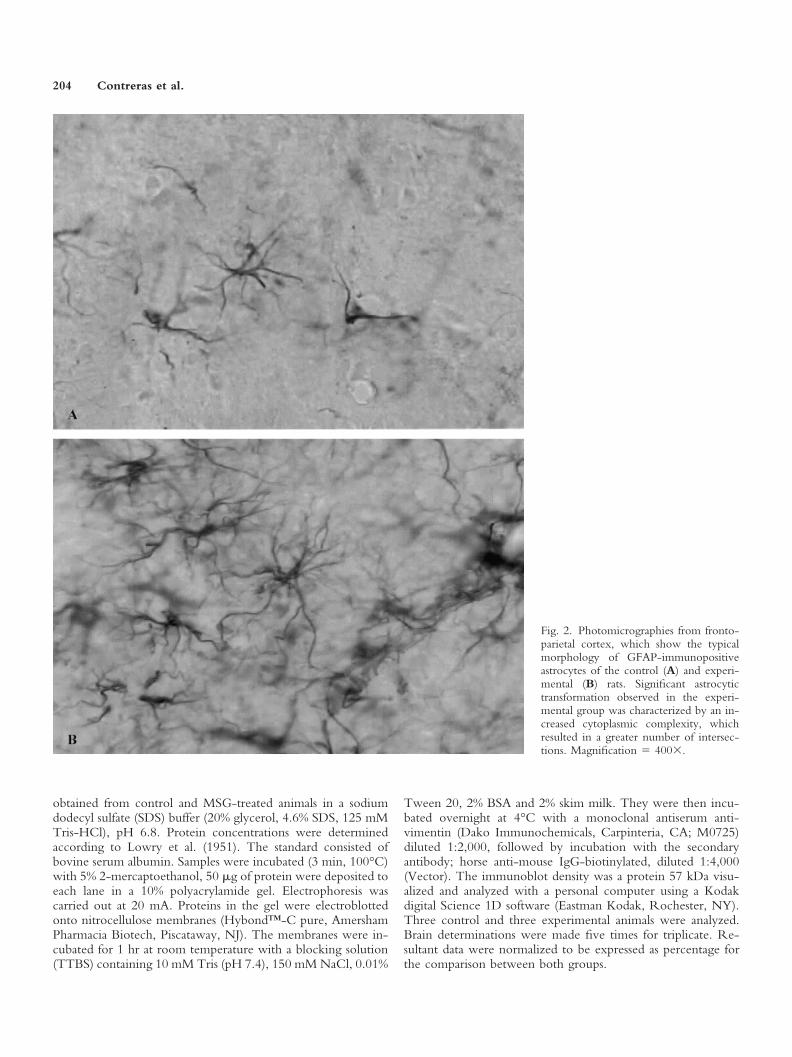
Fig. 2. Photomicrographies from fronto-parietal cortex, which show the typicalmorphology of GFAP-immunopositiveastrocytes of the control (A) and experi-mental (B) rats. Significant astrocytictransformation observed in the experi-mental group was characterized by an in-creased cytoplasmic complexity, whichresulted in a greater number of intersec-tions. Magnification � 400�.
204 Contreras et al.
Vimentin immunohistochemistry. The immuno-histochemical procedure to identify immature astrocytes withtheir intermediate filaments immunoreactive to vimentin(Kindy et al., 1992), was essentially followed as described above.Tissues were first incubated with an anti-vimentin monoclonal
antiserum (Dako; M0725) diluted 1:100, followed by the linkantiserum it consisted of goat IgG generated against mouseIgG with fluorescein coupled (Sigma), diluted 1:200. Thetissues were examined in a microscope equipped with epi-fluorescence.
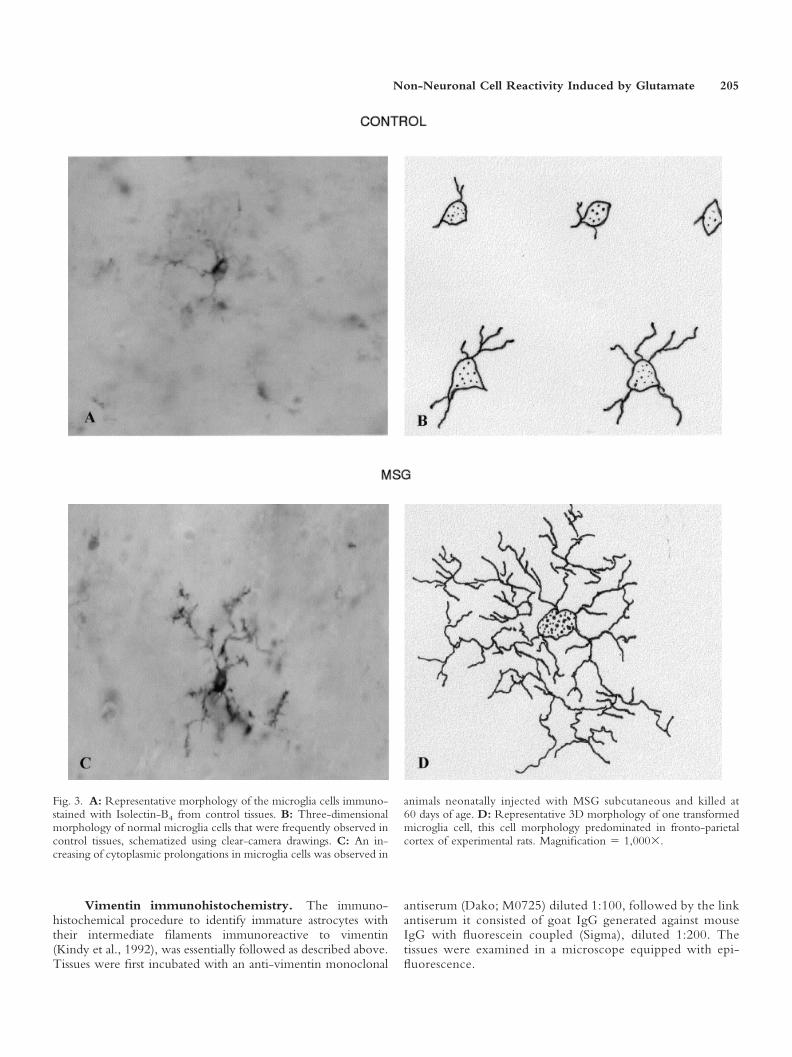
Fig. 3. A: Representative morphology of the microglia cells immuno-stained with Isolectin-B4 from control tissues. B: Three-dimensionalmorphology of normal microglia cells that were frequently observed incontrol tissues, schematized using clear-camera drawings. C: An in-creasing of cytoplasmic prolongations in microglia cells was observed in
animals neonatally injected with MSG subcutaneous and killed at60 days of age. D: Representative 3D morphology of one transformedmicroglia cell, this cell morphology predominated in fronto-parietalcortex of experimental rats. Magnification � 1,000�.
Non-Neuronal Cell Reactivity Induced by Glutamate 205
RESULTS
Quantification of Cells Number andMorphological Transformation
Astrocytes. To evaluate the reactivity of GFAPimmunopositive astrocytes against the neuroexcitotoxiceffects induced by the neonatal subcutaneous administra-tion of MSG, the number and cytoplasmic complexity ofthe astrocytes were analyzed in the fronto-parietal cortexof rats injected with MSG and sacrificed at 60 days of age.The results show that MSG induced a significant increasein the number of astrocytes, compared to the control rats(Fig. 1A). Similarly, morphological analysis of cytoplasmicarborizations indicate that the early exposure to MSGproduced greater complexity, which was manifested by anincreased number of cytoplasmic intersections, than thecontrol group (Figs. 1B, 2A,B). Both results indicate apersistent reactive astrocytic response, which is character-istic of a neuroexcitotoxic lesion of the CNS.
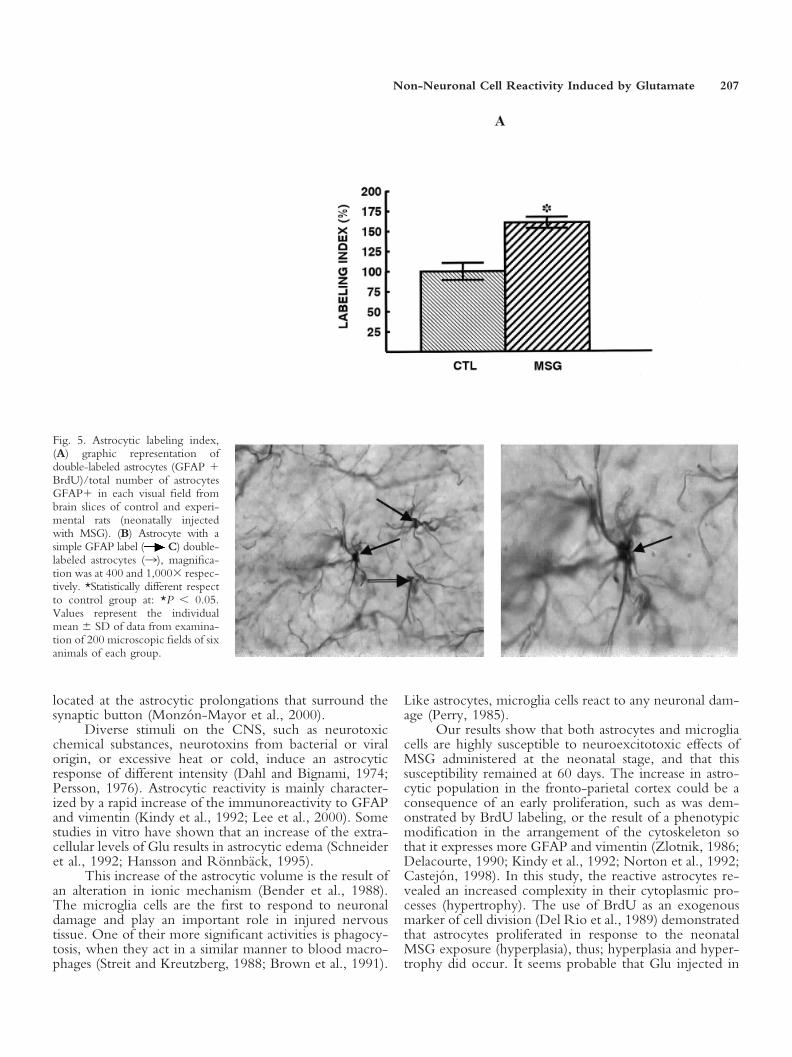
Astrocyte labeling index. To determine if theincrease in the number of glial cells induced by the neo-natal administration of MSG could be due to a prolifera-tive response against this excitotoxic stimuli, implants forlong-lasting delivery of BrdU were placed in a group ofrats when they were 14 days old. A labeling index wasobtained from this double immunolabeling study(GFAP � BrdU). The results showed that the neonataltreatment with MSG increased the labeling-index of as-trocytes in the fronto-parietal cortex. This is indicative ofreactive proliferation (hyperplasia) (Fig. 5A–C). The pres-ence of double-labeled astrocytes in the fronto-parietalcortex, identified by their cytoplasmic brown stain(GFAP) and nuclear dark purple stain (BrdU) demon-strated their proliferative origin provoked by neonatalexposure to MSG.
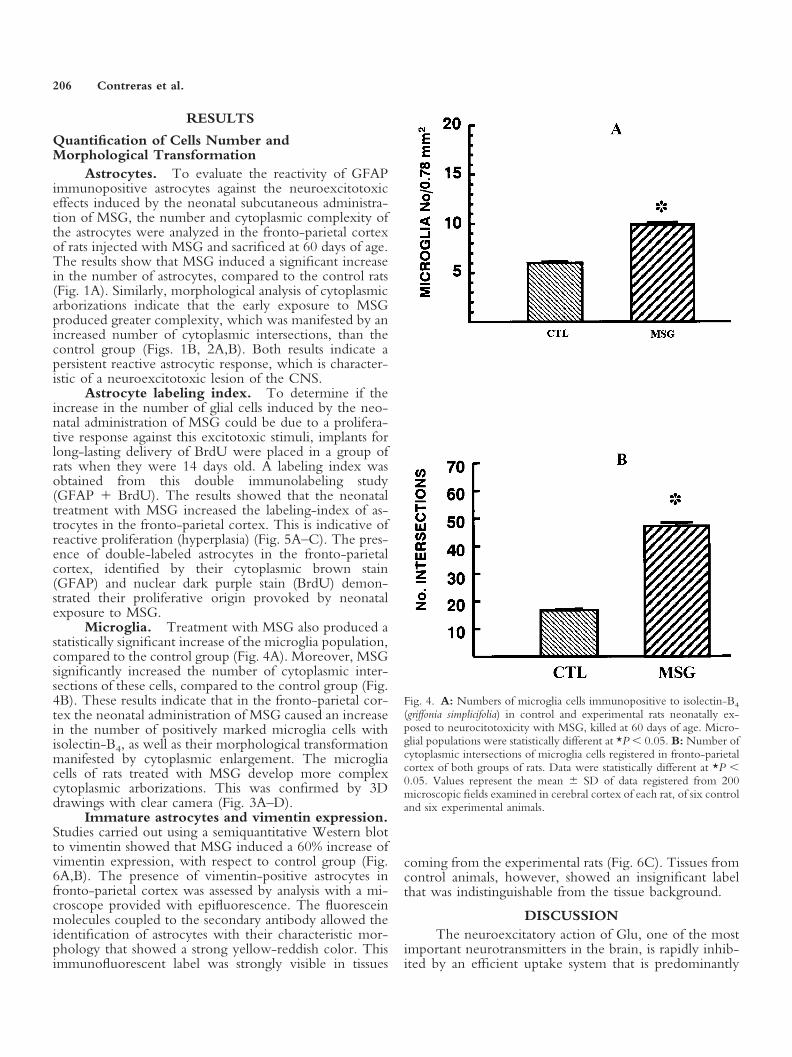
Microglia. Treatment with MSG also produced astatistically significant increase of the microglia population,compared to the control group (Fig. 4A). Moreover, MSGsignificantly increased the number of cytoplasmic inter-sections of these cells, compared to the control group (Fig.4B). These results indicate that in the fronto-parietal cor-tex the neonatal administration of MSG caused an increasein the number of positively marked microglia cells withisolectin-B4, as well as their morphological transformationmanifested by cytoplasmic enlargement. The microgliacells of rats treated with MSG develop more complexcytoplasmic arborizations. This was confirmed by 3Ddrawings with clear camera (Fig. 3A–D).
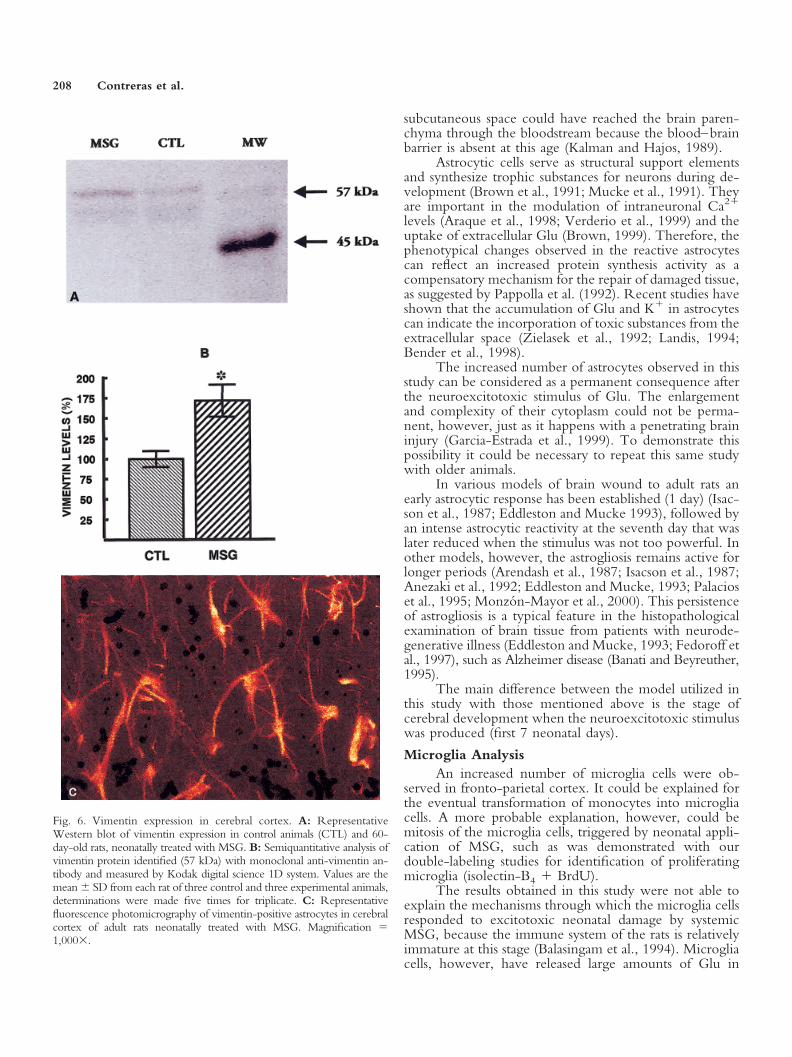
Immature astrocytes and vimentin expression.Studies carried out using a semiquantitative Western blotto vimentin showed that MSG induced a 60% increase ofvimentin expression, with respect to control group (Fig.6A,B). The presence of vimentin-positive astrocytes infronto-parietal cortex was assessed by analysis with a mi-croscope provided with epifluorescence. The fluoresceinmolecules coupled to the secondary antibody allowed theidentification of astrocytes with their characteristic mor-phology that showed a strong yellow-reddish color. Thisimmunofluorescent label was strongly visible in tissues
coming from the experimental rats (Fig. 6C). Tissues fromcontrol animals, however, showed an insignificant labelthat was indistinguishable from the tissue background.
DISCUSSIONThe neuroexcitatory action of Glu, one of the most
important neurotransmitters in the brain, is rapidly inhib-ited by an efficient uptake system that is predominantly
Fig. 4. A: Numbers of microglia cells immunopositive to isolectin-B4
(griffonia simplicifolia) in control and experimental rats neonatally ex-posed to neurocitotoxicity with MSG, killed at 60 days of age. Micro-glial populations were statistically different at *P � 0.05. B: Number ofcytoplasmic intersections of microglia cells registered in fronto-parietalcortex of both groups of rats. Data were statistically different at *P �0.05. Values represent the mean � SD of data registered from 200microscopic fields examined in cerebral cortex of each rat, of six controland six experimental animals.
206 Contreras et al.
located at the astrocytic prolongations that surround thesynaptic button (Monzon-Mayor et al., 2000).
Diverse stimuli on the CNS, such as neurotoxicchemical substances, neurotoxins from bacterial or viralorigin, or excessive heat or cold, induce an astrocyticresponse of different intensity (Dahl and Bignami, 1974;Persson, 1976). Astrocytic reactivity is mainly character-ized by a rapid increase of the immunoreactivity to GFAPand vimentin (Kindy et al., 1992; Lee et al., 2000). Somestudies in vitro have shown that an increase of the extra-cellular levels of Glu results in astrocytic edema (Schneideret al., 1992; Hansson and Ronnback, 1995).
This increase of the astrocytic volume is the result ofan alteration in ionic mechanism (Bender et al., 1988).The microglia cells are the first to respond to neuronaldamage and play an important role in injured nervoustissue. One of their more significant activities is phagocy-tosis, when they act in a similar manner to blood macro-phages (Streit and Kreutzberg, 1988; Brown et al., 1991).
Like astrocytes, microglia cells react to any neuronal dam-age (Perry, 1985).
Our results show that both astrocytes and microgliacells are highly susceptible to neuroexcitotoxic effects ofMSG administered at the neonatal stage, and that thissusceptibility remained at 60 days. The increase in astro-cytic population in the fronto-parietal cortex could be aconsequence of an early proliferation, such as was dem-onstrated by BrdU labeling, or the result of a phenotypicmodification in the arrangement of the cytoskeleton sothat it expresses more GFAP and vimentin (Zlotnik, 1986;Delacourte, 1990; Kindy et al., 1992; Norton et al., 1992;Castejon, 1998). In this study, the reactive astrocytes re-vealed an increased complexity in their cytoplasmic pro-cesses (hypertrophy). The use of BrdU as an exogenousmarker of cell division (Del Rio et al., 1989) demonstratedthat astrocytes proliferated in response to the neonatalMSG exposure (hyperplasia), thus; hyperplasia and hyper-trophy did occur. It seems probable that Glu injected in
Fig. 5. Astrocytic labeling index,(A) graphic representation ofdouble-labeled astrocytes (GFAP �BrdU)/total number of astrocytesGFAP� in each visual field frombrain slices of control and experi-mental rats (neonatally injectedwith MSG). (B) Astrocyte with asimple GFAP label ( C) double-labeled astrocytes (3), magnifica-tion was at 400 and 1,000� respec-tively. *Statistically different respectto control group at: *P � 0.05.Values represent the individualmean � SD of data from examina-tion of 200 microscopic fields of sixanimals of each group.
Non-Neuronal Cell Reactivity Induced by Glutamate 207
subcutaneous space could have reached the brain paren-chyma through the bloodstream because the blood–brainbarrier is absent at this age (Kalman and Hajos, 1989).
Astrocytic cells serve as structural support elementsand synthesize trophic substances for neurons during de-velopment (Brown et al., 1991; Mucke et al., 1991). Theyare important in the modulation of intraneuronal Ca2�
levels (Araque et al., 1998; Verderio et al., 1999) and theuptake of extracellular Glu (Brown, 1999). Therefore, thephenotypical changes observed in the reactive astrocytescan reflect an increased protein synthesis activity as acompensatory mechanism for the repair of damaged tissue,as suggested by Pappolla et al. (1992). Recent studies haveshown that the accumulation of Glu and K� in astrocytescan indicate the incorporation of toxic substances from theextracellular space (Zielasek et al., 1992; Landis, 1994;Bender et al., 1998).
The increased number of astrocytes observed in thisstudy can be considered as a permanent consequence afterthe neuroexcitotoxic stimulus of Glu. The enlargementand complexity of their cytoplasm could not be perma-nent, however, just as it happens with a penetrating braininjury (Garcia-Estrada et al., 1999). To demonstrate thispossibility it could be necessary to repeat this same studywith older animals.
In various models of brain wound to adult rats anearly astrocytic response has been established (1 day) (Isac-son et al., 1987; Eddleston and Mucke 1993), followed byan intense astrocytic reactivity at the seventh day that waslater reduced when the stimulus was not too powerful. Inother models, however, the astrogliosis remains active forlonger periods (Arendash et al., 1987; Isacson et al., 1987;Anezaki et al., 1992; Eddleston and Mucke, 1993; Palacioset al., 1995; Monzon-Mayor et al., 2000). This persistenceof astrogliosis is a typical feature in the histopathologicalexamination of brain tissue from patients with neurode-generative illness (Eddleston and Mucke, 1993; Fedoroff etal., 1997), such as Alzheimer disease (Banati and Beyreuther,1995).
The main difference between the model utilized inthis study with those mentioned above is the stage ofcerebral development when the neuroexcitotoxic stimuluswas produced (first 7 neonatal days).
Microglia AnalysisAn increased number of microglia cells were ob-
served in fronto-parietal cortex. It could be explained forthe eventual transformation of monocytes into microgliacells. A more probable explanation, however, could bemitosis of the microglia cells, triggered by neonatal appli-cation of MSG, such as was demonstrated with ourdouble-labeling studies for identification of proliferatingmicroglia (isolectin-B4 � BrdU).
The results obtained in this study were not able toexplain the mechanisms through which the microglia cellsresponded to excitotoxic neonatal damage by systemicMSG, because the immune system of the rats is relativelyimmature at this stage (Balasingam et al., 1994). Microgliacells, however, have released large amounts of Glu in
Fig. 6. Vimentin expression in cerebral cortex. A: RepresentativeWestern blot of vimentin expression in control animals (CTL) and 60-day-old rats, neonatally treated with MSG. B: Semiquantitative analysis ofvimentin protein identified (57 kDa) with monoclonal anti-vimentin an-tibody and measured by Kodak digital science 1D system. Values are themean � SD from each rat of three control and three experimental animals,determinations were made five times for triplicate. C: Representativefluorescence photomicrography of vimentin-positive astrocytes in cerebralcortex of adult rats neonatally treated with MSG. Magnification �1,000�.
208 Contreras et al.

culture, in response to variations in the ionic concentra-tion of culture medium (Ramırez-Exposito and Martınez-Martos, 1998). This fact could be relevant because micro-glia may have a role in neuronal death mediated by theglutamatergic stimulation that takes place in some mech-anisms of cerebral damage (Meldrum and Garthwaite,1990; Ramırez-Exposito and Martınez-Martos, 1998),neurological disorders (Schwarcz et al., 1983), and viraldementia (Giulian et al., 1993). Moreover, the release ofexcitatory amino acids is associated with an increase inextracellular K� concentration, which the microglia cellsare particularly sensitive to because of the characteristics oftheir ionic channels (Pellegrini-Giampetro, 1990; Ketten-mann et al., 1990; Zielasek et al., 1992). Therefore, in thismodel of neuroexcitotoxic damage to immature brain, it isunknown if the microglia cells could have an importantcontribution to the presumed neuronal damage in fronto-parietal cerebral cortex.
This study did not quantify the neuronal populationin the cerebral cortex, however, a reduction in the num-ber of neurons has been demonstrated in other experi-ments with rats also subjected to systemic administration ofMSG at birth (Gonzalez-Burgos et al., 2001). Moreover, ahistopathological examination of cerebral cortex slicesstained by Nissl’s technique showed evidence of neuronalmorphological abnormalities in dendritic spines, whichcan be attributed to Glu neurotoxicity (Gonzalez-Burgoset al., 2001). Delayed effects of early MSG neurotoxicityalso results in learning disorders (Ali et al., 2000; Laming etal., 2000).
ACKNOWLEDGMENTSThis study was partially supported by CONACyT
grant No. 30901-M to C.B.Z., by FOFOI grant No.0038/204 to J.G.E. and SIMORELOS 19990302034 toS.L.
REFERENCESAli MM, Bawari M, Misra UK, Babu GN. 2000. Locomotor and learning
deficits in adult rats exposed to monosodium-L-glutamate during earlylife. Neurosci Lett 284:57–60.
Amenta F, Bronzetti E, Sabbatini M, Vega JA. 1998. Astrocyte changes inaging cerebral cortex and hippocampus: a quantitative immunohisto-chemical study. Micros Res Tech 43:29–33.
Anezaki T, Yanagisawa K, Yakahashi H, Nakajima T, Miyashita K, Ish-ikawa A, Ikuta F, Miyatake T. 1992. Remote astrocytic response offronto-parietal cortex is caused by the lesions in the nucleus basalis ofMeynert, but not in the ventral tegmental area. Brain Res 574:63–69.
Araque A, Parpura V, Sanzigiri RP, Haydon PG. 1998. Glutamate-dependent astrocyte modulation of synaptic transmission between cul-tured hippocampal neurons. Eur J Neurosci 10:2129–2142.
Araque A, Parpura V, Sanzigiri RP, Haydon PG. 1999. Tripartite synapsesglia, the unacknowledged partner. Trends Neurosci 22:208–215.
Arendash GW, Millard WJ, Dunn AJ, Meyer EM. 1987. Long termneuropathological and neurochemical effects of nucleus basalis lesions inthe rat. Science 238:952–956.
Balasingam V, Tejada-Berges T, Wright E, Bouckova R Yong VW. 1994.Reactive astrogliosis in the neonatal mouse brain and its modulation bycytokines. J Neurosci 14:846–856.
Banati RB, Beyreuther K. 1995. Alzheimer disease. In: Kettenmann H,
Ransom BR, editors. Neuroglia. New York: Oxford University Press. p937–1043.
Beas-Zarate C, Sanchez-Ruız MY, Urena Guerrero M, Feria Velasco A.1998. Effect of neonatal exposure to monosodium L-glutamate on re-gional GABA release during postnatal development. Neurochem Int33:217–232.
Bender AS, Schousboe A, Reichelt W, Norenberg M. 1998. Ionic mech-anisms in glutamate-induced astrocyte swelling: role of K� influx. J Neu-rosci Res 52:307–321.
Bignami A, Dahl D, 1995. Gliosis. In: Kettenman H, Ransom BR, editors.Neuroglia. New York: Oxford University Press. p 843–858.
Brown DR. 1999. Neurons depend on astrocytes in a coculture system forprotection from glutamate toxicity. Mol Cel Neurosci 13:379–389.
Brown MC, Perry VH, Lunn ER, Gordon S, Heumann R.1991. Macrophage dependence of peripheral sensory nerve regeneration:possible involvement of nerve growth factor. Neuron 6:359–370.
Castejon OJ. 1998. Morphological astrocytic changes in complicated hu-man brain trauma. A light and electron microscopic study. Brain Inj12:409–427.
Curtis DR, Johnson GA. 1974. Amino acid transmitters in the mammaliancentral nervous system. Ergeb Physiologie 69:98–1889.
Dahl D, Bignami A. 1974. Heterogeneity of the glial fibrillary acidic proteinin gliosed human brains. J Neurol Sci 23:551–563.
Delacourte A. 1990. General and dramatic glial reaction in Alzheimerbrains. Neurology 40:33–37.
Del Rio JA, Soriano E. 1989. Immunohistochemical detection of 5�-bromo-deoxyuridine incorporation in the central nervous system of themouse. Brain Res Dev Brain Res 49:311–317.
Del Rıo Ortega P, Penfield W. 1927. Cerebral cicatrix. The reaction ofneuroglia and microglia to brain wounds. Bull Johns Hopkins Hosp41:278–303.
Eddleston M, Mucke L. 1993. Molecular profile of reactive astrocytesimplications for their role in neurologic disease. Neuroscience 54:5–36.
Fedoroff S, Berezovskaya O, Maysinger D. 1997. Role of colony stimu-lating factor-1 in brain damage caused by ischemia. Neurosci BiobehavRev 21:187–191.
Garcia-Estrada J, Luquin S, Fernandez AM, Garcia-Segura LM, 1999.Dehydroepiandrosterone, pregnenolone and sex steroids down-regulatereactive astroglia in the male rat brain after a penetrating brain injury. IntJ Dev Neurosci 17:145–152.
Giulian D, Vaca K, Corpuz M. 1993. Brain glia release factors withopposing actions upon neuronal survival. J Neurosci 13:29–37.
Gonzalez-Burgos I, Perez-Vega MI, Beas-Zarate C. 2001. Cell death anddendritic hypotrophy induced by neonatal exposure to monosodiumglutamate, on prefrontocortical pyramidal neurons of rats. Neurosci Lett297:69–72.
Hansson E, Muyderman H, Leonova J, Allanson L, Sinclair J, BlomstrandF, Thorlin T, Nilsson M, Ronnback L. 2000. Astroglia and glutamate inphysiology and pathology: aspects on glutamate transport, glutamate-induced cell swelling and gap-junction communication. Neurochem Int37:317–329.
Hansson E, Olsson T, Ronnback L. 1997. On astrocytes and glutamateneurotransmission: new waves in brain information processing. Berlin:Springer. 191 p.
Hansson E, Ronnback L. 1995. Astrocytes in glutamate neurotransmission.FASEB J 9:343–350.
Isacson O, Fischer W, Wictorin K, Dawbarn D, Bjourklund A. 1987.Astroglial response in the excitotoxically lesioned neostriatum and itsprojection areas in the rat. Neuroscience 20:1043–1056.
Kalman M, Hajos F. 1989. Distribution of glial fibrillary acidic proteinGFAP-immunoreactive astrocytes in the rat brain. I. Forebrain. Exp BrainRes 78:147–163.
Non-Neuronal Cell Reactivity Induced by Glutamate 209

Kindy MS, Bhat AN, Bhat NR. 1992.Transient ischemia stimulates glialfibrillary acid protein and vimentin gene expression in the gerbil neocor-tex, striatum and hippocampus. Brain Res Mol Brain Res 3:199–206.
Kettenmann H, Hoppe D, Gottmann K, Banati R, Kreutzberg GW. 1990.Cultured microglia cells have a distinct pattern of membrane channelsdifferent from peritoneal macrophages. J Neurosci Res 26:278–287.
Lafarga M, Berciano MT, Suarez I, Andres MA, Berciano J. 1993. Reactiveastroglia-neuron relationships in the human cerebellar cortex: a quantita-tive, morphological and immunohistochemical study in Creutzfeldt-Jakobdisease. Int J Dev Neurosci 11:199–213.
Laming PR, Kimelberg H, Robinson S, Salm A, Hawrylak N, Muller C,Roots B, Ng K. 2000. Neuronal-glial interactions and behavior. NeurosciBiobehav Rev 24:295–340.
Laming PR, Sykova E, Reichenbach A, Hatton GI, Bauer H. 1998. Glialcells: their role in behavior. Cambridge: Cambridge University Press. p1–424.
Landis DMD. 1994. The early reactions of non-neuronal cells to braininjury. Ann Rev Neurosci 17:133–151.
Lee YB, Du S, Rhim H, Lee EB, Markelonis GJ, Oh TH. 2000. Rapidincrease in immunoreactivity to GFAP in astrocytes in vitro induced byacidic pH is mediated by calcium influx and calpain I. Brain Res 864:220–229.
Lorden JF, Sims, JS. 1987. Monosodium L-glutamate lesions reduce sus-ceptibility to hypoglycemic feeding and convulsions. Behav Brain Res24:139–146.
Lowry O, Rosebrough N, Farr A, Randall R. 1951. Protein measurementwith the Folin phenol reagent. J Biol Chem 193:265–275.
Meldrum B, Garthwaite J. 1990. Excitatory amino acids neurotoxicity andneurodegenerative diseases. Topics Pharm Sci 11:379–387.
Monzon-Mayor M, Alvarez MI, Arbelo-Galvan JF, Romero-Aleman MM,Yanes C, Plaza ML, Rodrıguez JR, Rodrıguez JJ, Toledano A. 2000.Long-term evolution of local, proximal and remote astrocyte responsesafter diverse nucleus basalis lesioning an experimental Alzheimer model:GFAP immunohistochemical study. Brain Res 865:245–258.
Mucke L, Oldstone MB, Morris JC, Nerenberg MI. 1991. Rapid activationof astrocyte-specific expression of GFAP-lacZ transgene by focal injury.New Biol 3:465–474.
Norenberg MD. 1998. Astroglial dysfunction in hepatic encephalopathy.Metab Brain Dis 13:319–335.
Norton WT, Aquino DA, Hozumi I, Chiu FC, Brosnan CF. 1992.Quantitative aspects of reactive gliosis: a review. Neurochem Res 17:877–885.
Olney JW. 1971. Glutamate induce neuronal necrosis in the infant mousehypothalamus. J Neuropath Exp Neurol 30:75–90.
Olney JW. 1978. Neurotoxicity of excitatory amino acids. In: McGeer E,Olney J, McGeer P, editors. Kainic acid as tool in neurobiology. NewYork: Raven Press. p 95–121.
Olney JW. 1980a. Excitotoxic mechanisms of neurotoxicity. In: SpencerPS, Schaumburg HH, editors. Experimental and clinical neurotoxicity.Baltimore: William Wilkins. p 272–294.
Olney JW. 1980b. Excitatory neurotoxins as food additives: an evaluationof risk. Neurotoxicology 2:153–192.
Palacios G, Mengod G, Tortosa A, Ferrer I, Palacios JM. 1995. Increased-amyloid precursor protein expression in astrocytes in the gerbil hip-pocampus following ischemia: association with proliferation of astrocytes.Eur J Neurosci 7:501–510.
Pappolla MA, Omar RA, Kim KS, Robakis NK. 1992. Immunohisto-chemical evidence of antioxidant stress in Alzheimer disease. Am J Pathol140:621–628.
Paxinos G, Watson, C. 1998. The rat brain in stereotaxic coordinates, 4thed. San Diego, London: Academic Press. 270 p.
Pellegrini-Giampetro DE, Cherici G, Alesiani M, Carla V, Moroni F. 1990.Excitatory amino acid release and free radical formation may cooperate inthe genesis of ischemia-induced neuronal damage. J Neurosci 10:1035–1041.
Penfield W. 1932. Neuroglia: normal and pathological. In: Penfield W,editor. Cytology and cellular pathology of the nervous system. NewYork: PB Horber. p 423–479.
Perry VH. 1985. Macrophages, microglia and cell death in the developingmouse brain. Pontific Acad Sci Scripta Varia 59:281–295.
Persson L. 1976. Cellular reactions to small cerebral stab wounds in the ratfrontal lobe. Virchows Arch B Cell Path 22:21–37.
Ramırez-Exposito MJ, Martınez-Martos JM. 1998. Estructura y funcionesde la microglia en el sistema nervioso central. Respuesta a procesosdegenerativos. Rev Neurol 26:600–611.
Schneider GH, Baethmann A, Kempski O. 1992. Mechanism of glialswelling induced by glutamate. Can J Physiol Pharmacol S334–S343.
Schwarcz R, Whetsell WO, Mangano RM. 1983. Quinolinic acid: endog-enous metabolite that produces axon-sparing lesions in rat brain. Science219:316–318.
Spokes EG. 198l. The neurochemistry of Huntington chorea. TINS 4:115–118.
Steinhauser C, Gallo V. 1996. News on glutamate receptors in glial cells.TINS 19:339–345.
Sterio DC. 1984, The unbiased estimation of number and sizes of arbitraryparticles using the dissector. J Microsc 134:127–136.
Streit WJ, Kreutzberg GW. 1988. Response of endogenous glial cells tomotor neuron, degeneration induced by toxic ricin. J Comp Neurol268:248–263.
Venters HD, Dantzer R, Kelly KW. 2000. A new concept in neurodegen-eration: TNF is a silencer of survival signals. TINS 23:175–180.
Verderio C, Bacci A, Coco S, Pravettoni E, Fumagalli G, Matteoli M.1999. Astrocytes are required for the oscillatory activity in culturedhippocampal neurons. Eur J Neurosci 8:2793–2800.
Young AB, Sakurai SY, Albin RL, Makowiec R, Penney JB. 1991.Excitatory amino acid receptor distribution, In: Thompson AM, editor.Excitatory amino acids and synaptic transmission. New York: AcademicPress. 19–31 p.
Zielasek J, Tauscher M, Toyka KV, Hartung P. 1992. Production of nitriteby neonatal microglial cells/brain macrophages. Cell Immunol 141:111–120.
Zlotnik I. 1986. The reaction of astrocytes to acute virus infections of thecentral nervous system. Br J Exp Pathol 49:555–564.
210 Contreras et al.





























