Embed Size (px)
Citation preview

1987 70: 1743-1749
F Dallegri, A Ballestrero, G Frumento and F Patrone vitro from human monocytesAugmentation of neutrophil-mediated erythrocyte lysis by cells derived in
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.20036.the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by
For personal use only. by guest on July 10, 2011. bloodjournal.hematologylibrary.orgFrom

Blood, Vol 70. No 6 (December), 1987: pp 1743-1749 1743
Augmentation of Neutrophil-Mediated Erythrocyte Lysis by Cells Derived
In Vitro From Human Monocytes
By Franco Dallegri, Alberto Ballestrero, Guido Frumento, and Franco Patrone
Neutrophilic polymorphonuclear leukocytes (PMNs) wereincubated with opsonized zymosan and lysed human eryth-
rocytes (RBCs) as measured by a 51Cr release method.Conversely. myeloperoxidase (MPO)-negative hydrogenperoxide (H202)-generating cells. derived in vitro fromhuman monocytes (monocyte-derived cells (MDCs). wereineffective per se but capable of augmenting the lysis byPMNs. The lysis by PMNs and PMNs plus MDCs wasinhibited by catalase. azide. taurine. and alanine. consistent
with the requirement for hypochlorous acid (HOCI). Asdetected under conditions similar to those used for lyticassays. MDCs failed to produce HOCI but augmented theHOCI recovery from the PMN-RBC system. Moreover.when the extent of the lysis was plotted as a function ofthe HOCI recovery. a positive linear relationship was found.
Although the actual size of the H202 extracellular pool
W HEN TRIGGERED by appropriate stimuli, either
soluble (phorbol myristate acetate) or particulate
(opsonized zymosan [OPZ], heat-aggregated IgG), neutro-
phils (neutrophilic polymorphonuclear leukocytes [PMNs])
have been shown to be capable of lysing a wide spectrum of
eukaryotic target cells.’�5 These in vitro systems of cytolysis
have been used to understand the mechanisms by which
PMNs may injure tissue cells at inflammatory sites. In the
present paper, by the use of a previously characterized
system constituted of OPZ-triggered PMNs and human
erythrocytes (RBCs) as targets,6 we provide evidence that
the PMN cytolytic efficiency can be augmented by myelo-
peroxidase (MPO)-negative cells derived in vitro from
human monocytes (MDCs) and per se incapable of mediat-
ing lysis. The results uncover a novel possible role for
MPO-negative mononuclear phagocytes in interaction with
PMNs during inflammatory processes.
MATERIALS AND METHODS
Preparation of PMNs. Heparinized venous blood (heparin, 10
U/mL, Liquemin, Roche, Milan, Italy) was obtained from healthymale volunteers. PMNs were isolated by dextran sedimentation andsubsequent centrifugation on a Ficoll-Hypaque density gradient as
previously described.’ Contaminating erythrocytes were removed byhypotonic lysis.6 The resulting PMNs were washed three times with
Hanks’ balanced salt solution lacking phenol red (HBSS, FlowLaboratories, Ltd, Irvine, Scotland) and resuspended in HBSS
supplemented with 0.1 mg/mL bovine serum albumin (BSA, Cal-
biochem-Behring, San Diego) (BSA-HBSS). Final cell suspensionscontained 97% or more PMNs and more than 98% viable cells asevaluated by the ethidium bromide-fluorescein diacetate test.7
Preparation of MDCs. Venous blood (350 mL) from healthymale volunteers was collected into 53 mL of anticoagulant citrate-phosphate-dextrose-adenine (CDPA) (Baxter, Trieste, Italy). Thebuffy coat, obtained two to three hours after venipuncture, wasdiluted with an equal volume of 0.9% NaCI, and 25 mL of the cellsuspension was layered on 15 mL Ficoll-Hypaque (Nyegaard & Co,
Oslo) in 50-mL polypropylene conical tubes (No. 25330, CorningGlass Works, Corning, NY). After centrifugation (400 g, 30 mm-
utes, 22#{176}C),the upper cell layer (mononuclear cells) was collectedand spun down at 400 g for ten minutes at 22#{176}C.The sedimented
cells were washed three times with HBSS (200 g, ten minutes, 22#{176}C)
could not be measured because of the inexistence of areliable assay to probe our cytolytic model without per-turbing the equilibrium of the system. the results pre-
sented suggest that MDCs enhance the PMN-mediatedlysis by improving the HOCI production. presumably bysupplying extra amounts of H302 to be handled by PMNMPO. In fact. the events mediated by MDCs could bereproduced by using an appropriate H202-generating enzy-matic system (glucose-glucose oxidase). The present
study provides direct evidence for the possibility of cooper-ation between MPO-positive and MPO-negative phago-cytes in exerting functions (HOCI production and. in turn.cytolysis) possibly relevant to the outcome of inflammatoryprocesses.a 1987 by Grune & Stratton, Inc.
and resuspended in Fisher’s medium (Flow) supplemented with 25%filtered horse serum (HS, Flow), 2 mmol/L glutamine, 100 U/mLpenicillin, and 100 ,Lg/mL streptomycin (US-Fisher’s medium8).
The cells were then plated in 75-cm2 tissue culture flasks (Sterilin,Ltd. Teddington, England) in thin suspension layers (2 x l07/mL, 5
mL). After incubation for two hours at 37#{176}Cin a humidifiedatmosphere of 95% air and 5% C02, nonadherent cells were removed
by washing five times with prewarmed (37#{176}C)HBSS and theadherent cells overlaid with 8 mL HS-Fisher’s medium. The flaskswere then returned to the CO2 incubator. Three days later, the
medium containing detached cells was decanted, and adherent cells
were rinsed three times with prewarmed (37#{176}C)HBSS. Adherentcells were then removed by pipetting after incubation (20 minutes,
4#{176}C)with phosphate-buffered saline (PBS, pH 7.4) containing 0.2%
EDTA (Sigma Chemical Co. St. Louis) and 5% heat-inactivated (30minutes, 56#{176}C)fetal calf serum (Flow).9 The resulting cells were
centrifuged (200 g, ten minutes, 4#{176}C),washed three times with cold
(4#{176}C)HBSS, and resuspended in cold (4#{176}C)BSA-HBSS. Only cellsuspensions containing >90% a-naphthyl acetate esterase-positivecells, as determined by the method of Li et al,’#{176}were used. Thesecells, referred to as MDCs, were >97% viable as judged by theethidium bromide-fluorescein diacetate test.7
Preparation of OPZ. OPZ was prepared by incubating a sus-pension containing 10 mg/mL boiled zymosan (Sigma) in PBS (pH7.4) with an equal volume of fresh human serum for 30 minutes at
37#{176}C.The particles were then washed three times with PBS andresuspended in BSA-HBSS at the appropriate concentration.
Measurement of RBC lysis. The RBC lysis was studied by a
From the First Medical Clinic, University of Genoa Medical
School, Italy.
Submitted February 17. 1987; accepted July 29. 1987.
Supported by the Italian Consiglio Nazionale delle RicercheFinalized Project “Oncology” No. 86.00518.44/115.11547.
Address reprint requests to Franco Dallegri. MD, ISMI-Clinica
Medica I, Universita’ degli Studi, Viale Benedetto XV, n. 6. 1-16132Genova, Italy.
The publication costs ofthis article were defrayed in part by page
charge payment. This article must therefore be hereby marked“advertisement” in accordance with /8 U.S.C. §1 734 solely to
indicate this fact.© I 987 by Grune & Stratton, Inc.
0006-4971/87/7006-0006$3.00/0
For personal use only. by guest on July 10, 2011. bloodjournal.hematologylibrary.orgFrom

1744 DALLEGRI ET AL
�‘Cr release method as previously described.6 Briefly, RBCs were
washed four times with HBSS and labeled (60 minutes, 37#{176}C)with100 to 200 MCi Na2[5’Cr104 (Radiochemical Centre, Amersham,
England). The cells were then washed three times with HBSS and
resuspended in BSA-HBSS. When required, RBCs were pretreated
(60 minutes, 37#{176}C)with 20 mmol/L 3-amino-l,2,4-triazole (AT,Sigma) plus 2 mmol/L L-ascorbic acid (Sigma) (RBCAT) to inhibit
catalase activity.” A quantity of l0� RBCs were mixed with PMNs,
MDCs, or PMNs plus MDCs at appropriate concentrations. In someexperiments, MDCs were replaced by glucose oxidase (GO, type II,Lot I I 1F-9605, Sigma) dissolved in HBSS (prepared without glu-cose) and mixed with the other components of the reaction to reach afinal concentration of 5 mU/mL (incubation medium, BSA-HBSSwith 1 mg/mL glucose). Incubations were initiated by the addition
of OPZ (0.4 mg/mL) and followed by centrifugation of the samples
at 50 g (five minutes, 22#{176}C).Tests were performed in triplicate and
in a final volume of 200 ILL in round-bottomed microplates (Steril-
in). After incubation for four hours at 37#{176}Cin a humidified
atmosphere of 95% air and 5% CO2. the �‘Cr release was determined
in the cell-free supernatants. The percentage of cytolysis (percent
5’Cr release) was calculated according to the formula (E - S)/(T - 5) x 100, where E is the counts per minute released in the
presence of effector cells, T is the counts per minute released after
lysing target cells with 5% Triton X-lOO (Sigma), S is the counts per
minute spontaneously released by target cells in the absence of
effector cells (in each case, <10%).
Measurement of H202 release. The hydrogen peroxide (H2O2)release was studied by a modification of the homovanillic acid(HVA)-horseradish peroxidase (HRP) method’2 as previouslydescribed.” Tests were performed in triplicate under experimental
conditions similar to those used for cytolytic assays except that
incubations were done in Falcon plastic tubes ( I 2 x 75 mm, Falcon
Labware, Oxnard, CA). Briefly, 5 x iO� PMNs were incubated
(37#{176}C)with and without 0.4 mg/mL OPZ in the presence of 200
Mmol/L HVA (Sigma) plus 2 U/mL HRP (Sigma). When required,2 x 10’ RBC targets were added. Similar experiments were carried
out with an appropriate number of MDCs both in presence and inabsence of PMNs. The cells were sedimented by centrifugation (50g, five minutes) at the beginning of the assay. The reactions,
performed in a final volume of 2 mL, were stopped after 20 minutes
with 0.25 mL of0.l mol/L glycine-NaOH buffer (pH 12) contain-
ing 25 mmol/L EDTA. The amount of H202 in the cell-free
supernatants was determined fluorimetrically (X,�, = 3 1 2 nm,
Acm 420 nm) by using appropriate standards of H2O2. Preliminary
experiments showed that the amount of H202 recovered fromOPZ-triggered neutrophils incubated (20 minutes) as a pellet equalsthat recovered from OPZ-triggered neutrophils incubated as a pellet
during the first five minutes of the assay period (20 minutes) and
then resuspended and maintained in suspension.
Measurement of HOCI production. The generation of hypo-chlorous acid (HOCI) by OPZ-triggered PMNs was quantitated bythe taurine trapping technique’3 as previously described.” Theconditions of the assay were identical to those used for the H202
release except that the reactions were carried out in a final volume ofI mL. Briefly, an appropriate number of PMNs was incubated (20
minutes, 37#{176}C)with 0.4 mg/mL OPZ in the presence of 20 mmol/Ltaurine (Sigma). Tests were performed in the presence and in theabsence of 2 x 106 RBC targets or 300 units catalase (bovine liver,
Sigma; amount of enzyme chosen on the basis of preliminary
experiments). Similar experiments were carried out with MDCsboth in presence and in absence of PMNs. The cells were sedimentedby centrifugation (50 g, five minutes) at the beginning of the assay.At the end of the incubation period (20 minutes), catalase (600
units, Sigma) was added to reduce any residual H2O2. The sampleswere then incubated (22#{176}C,five minutes) with 100 nmol of5-thio-2-nitrobenzoic acid (Nbs) prepared by reducing 5-5’-dithiobis
(2-nitrobenzoic acid) (Sigma) as described by Aune and Thomas.’4The amounts of Nbs oxidized were determined by measuring theabsorbance at 412 nm of the cell-free supernatants (� = 1.36 x i04mol/L/cm). The quantities of HOCI trapped by taurine (yelding
taurine chloramine) were calculated by taking into account that 1mol taurine chloramine oxidizes 2 mol Nbs.’3 As for the H202 assay,the method of HOC1 detection maintained its efficiency also when
tests were carried out with cells incubated as a pellet for 20 minutes.
Under the conditions described earlier, the uptake of taurine chlo-ramine by RBCs, via the 4,4’-diisothiocyano-2,2’-disulfonic acid
stilbene (DIDS)-inhibitable anion transport system,’5 did not affectthe taurine chloramine recovery (the amounts of taurine chloraminerecovered in the presence of RBCs equalled those recovered in the
presence of RBCs pulsed (30 minutes) with 0.1 mmol/L DIDS(Sigma).”
Enzyme assays. The total cell content of MPO was measuredafter lysing the cells by sonication followed by freezing-thawing andsolubilization with 0.5% hexadecyltrimethylammonium bromide
(HTAB, Sigma).’6 The MPO activity released by cells incubated
(one hour, 37#{176}C)with 0.4 mg/mL OPZ was determined in HTAB-untreated cell-free supernatants. The MPO assay was done by an
O-dianisidine method as previously described.” One unit of enzyme
activity was defined as that oxidizing I izmol of O-dianisidine/min at
25#{176}C(optical density, 550, t - 1 1.3 mmol/L/cm).
For determination of RBC catalase activity, the cells were lysed asdescribed by Beutler,’7 and the enzyme activity was determined by
recording the rate of decomposition of H202 at 230 nm (t - 0.071mmol/L/cm).’7 The activity of “reagent” catalase (bovine liver,Sigma) was measured similarly. The results were expressed asnanomoles H202/min at 25#{176}Cor units (1 unit of enzyme activity was
defined as that catalyzing the breakdown of I �mol H202/min at
25#{176}C).Special materials. Superoxide dismutase (SOD, type 1, bovine
blood, Sigma), stored at - 20#{176}Cat a concentration of 5 mg/mL in
distilled water, was diluted in medium before use. Catalase (bovine
liver, 40,000 U/mg protein), taurine, L-alanine, mannitol, andbenzoate were purchased from Sigma. Sodium azide (NaN3) and
(NH4)2S04 were from Merck, Darmstadt, FRG. MPO, as a super-natant from the PMN granule fraction (MPO 4,800 mU/mL, 24.9mg/mL protein), was prepared by using normal human PMNs andstored at - 20#{176}C.’8Other reagent-grade compounds were used as
obtained from commercial suppliers. In some experiments, BSA-
HBSS medium was replaced with a Cl-free, 0.1 mol/L phosphate
buffer (pH 7.4) with I .5 mmol/L MgSO4 supplemented with Img/mL glucose (Merck) and 0.1 mg/mL BSA (Calbiochem-
Behring)(Cl-free medium).
RESULTS
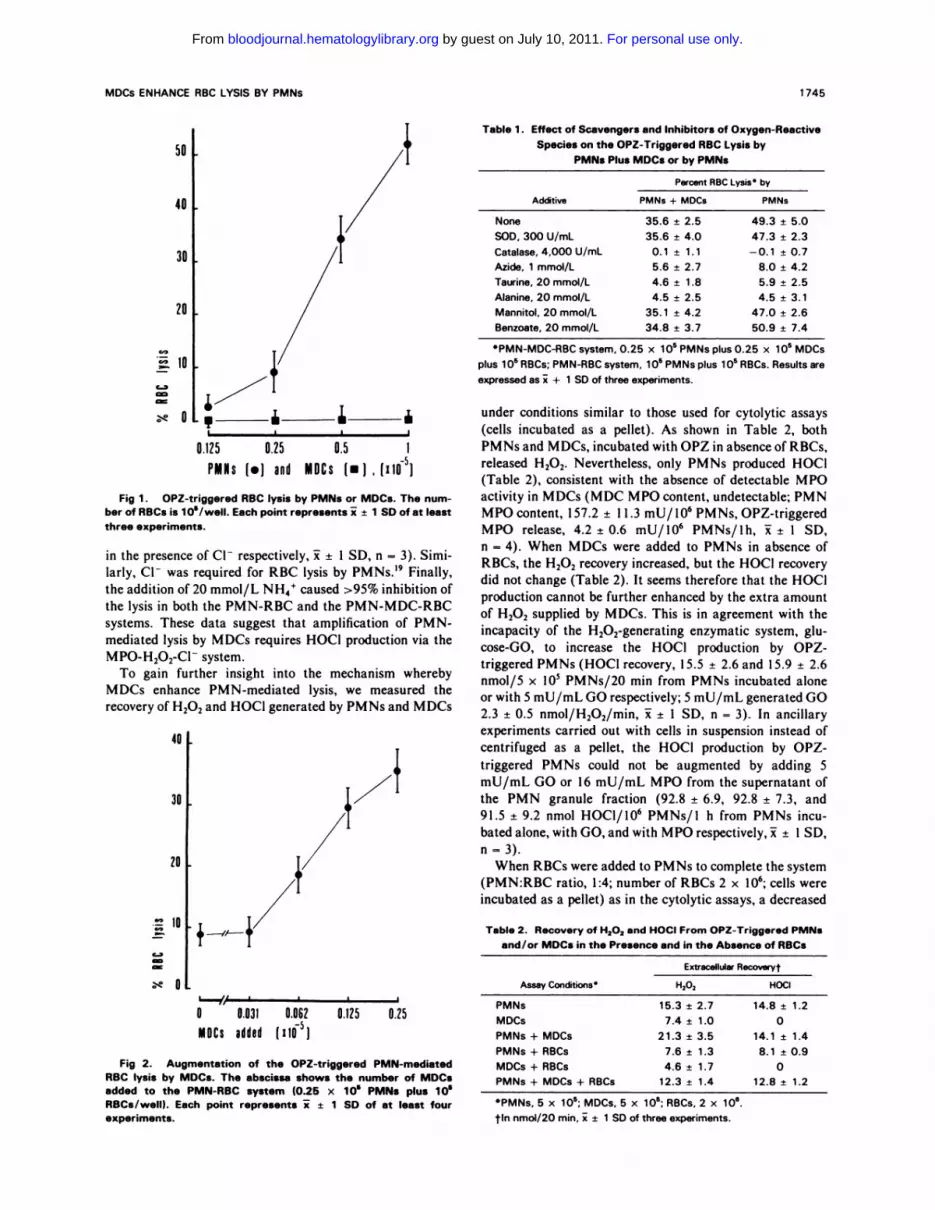
As depicted in Fig 1 , OPZ-triggered PMNs lysed RBC
targets, whereas MDCs were ineffective. When added to the
PMN-RBC system (0.25 x l0� PMNs plus l0� RBCs,
PMN:RBC ratio, I :4), MDCs enhanced the lysis in a
dose-dependent manner (Fig 2). As detailed in Table 1, the
lysis in the complete PMN-MDC-RBC system (0.25 x l0�
PMNs plus 0.25 x l0� MDCs plus I0� RBCs) was inhibited
by adding catalase to destroy H202 (heat-inactivated cata-
lase was ineffective), azide to inhibit MPO, and amino acids
(alanine, taurine) to scavenge HOCI. Scavengers of superox-
ide anion (SOD) and hydroxyl radicals (mannitol, benzoate)
were ineffective (Table 1). Similar results were obtained in
the PMN-RBC system (Table I), in agreement with our
previous findings.6” Moreover, the absence of Cl corn-
pletely prevented lysis in the PMN-MDC-RBC system (per-
cent 51Cr release, 0. 1 ± 0.8 and 36.0 ± 2.3 in the absence and
For personal use only. by guest on July 10, 2011. bloodjournal.hematologylibrary.orgFrom
‘450
40
30
20
.�.10
C-,
�! � �
40 I-
30
20
10
01
MDCs
0.031 0.062 0.125 0.25
added tiio��j
Fig 2. Augmentation of the OPZ-triggered PMN-mediatedRBC Iysis by MDCs. The abscissa shows the number of MDCsadded to the PMN-RBC system (0.25 x 10’ PMNs plus i0RBCs/weIl). Each point represents i ± 1 SD of at least fourexperiments.
PMN5, 5 x 10’; MDCs, 5 x io�; RBCs, 2 x 106.
tIn nmol/2O mm, i ± 1 SD of three experiments.
MDCS ENHANCE RBC LYSIS BY PMNs 1745
0.125 0.25 0.5 1
PMNs (.1 and MOCs .1 , [x1O�)
Fig 1 . OPZ-triggered RBC lysis by PMNs or MDCs. The num-ber of RBCs is 1 08/weIl. Each point represents i ± 1 SD of at leastthree experiments.
in the presence of Cl respectively, ic ± 1 SD, n = 3). Simi-
larly, Cl was required for RBC lysis by PMNs.’9 Finally,
the addition of 20 rnmol/L NH4� caused >95% inhibition of
the lysis in both the PMN-RBC and the PMN-MDC-RBC
systems. These data suggest that amplification of PMN-
mediated lysis by MDCs requires HOC1 production via the
MPO-H2O2-C1 system.
To gain further insight into the mechanism whereby
MDCs enhance PMN-mediated lysis, we measured the
recovery of H2O2 and HOCI generated by PMNs and MDCs
Table 1 . Effect of Scavengers and Inhibitors of Oxygen-ReactiveSpecies on the OPZ-Triggered RBC Lysis by
PMNs Plus MDCs or by PMNs
Percent RBC Lysis by
Additive PMNs + MDCs PMNs
None 35.6 ± 2.5 49.3 ± 5.0
SOD, 300 U/mL 35.6 ± 4.0 47.3 ± 2.3
Catalase,4,000U/mL 0.1 ± 1.1 -0.1 ±0.7
Azide, 1 mmol/L 5.6 ± 2.7 8.0 ± 4.2
Taurine, 20 mmol/L 4.6 ± 1.8 5.9 ± 2.5
Alanine, 20 mmol/L 4.5 ± 2.5 4.5 ± 3.1
Mannitol, 20 mmol/L 35. 1 ± 4.2 47.0 ± 2.6
Benzoate, 20 mmol/L 34.8 ± 3.7 50.9 ± 7.4
PMN-MDC-RBC system, 0.25 x i0� PMNs plus 0.25 x 10� MDCs
plus i05 RBCs; PMN-RBC system, 10� PMNs plus � RBCs. Results are
expressed as i + 1 SD of three experiments.
under conditions similar to those used for cytolytic assays
(cells incubated as a pellet). As shown in Table 2, both
PMNs and MDCs, incubated with OPZ in absence of RBCs,
released H2O2. Nevertheless, only PMNs produced HOCI
(Table 2), consistent with the absence of detectable MPO
activity in MDCs (MDC MPO content, undetectable; PMN
MPO content, 157.2 ± 1 1.3 mU/106 PMNs, OPZ-triggered
MPO release, 4.2 ± 0.6 mU/l06 PMNs/lh, � ± I SD,
n = 4). When MDCs were added to PMNs in absence of
RBCs, the H202 recovery increased, but the HOCI recovery
did not change (Table 2). It seems therefore that the HOCI
production cannot be further enhanced by the extra amount
of H202 supplied by MDCs. This is in agreement with the
incapacity of the H202-generating enzymatic system, glu-
cose-GO, to increase the HOC1 production by OPZ-
triggered PMNs (HOCI recovery, 1 5.5 ± 2.6 and 1 5.9 ± 2.6
nmol/5 x l0� PMNs/20 mm from PMNs incubated alone
or with 5 mU/mL GO respectively; 5 mU/mL generated GO
2.3 ± 0.5 nmol/H202/min, � ± 1 SD, n = 3). In ancillary
experiments carried out with cells in suspension instead of
centrifuged as a pellet, the HOC1 production by OPZ-
triggered PMNs could not be augmented by adding 5
rnU/mL GO or 16 mU/mL MPO from the supernatant of
the PMN granule fraction (92.8 ± 6.9, 92.8 ± 7.3, and
91.5 ± 9.2 nmol HOC1/106 PMNs/l h from PMNs incu-
bated alone, with GO, and with MPO respectively, � ± I SD,
n=3).
When RBCs were added to PMNs to complete the system
(PMN:RBC ratio, 1:4; number of RBCs 2 x 106; cells were
incubated as a pellet) as in the cytolytic assays, a decreased
Table 2. Recovery of H202 and HOCI From OPZ-Triggered PMNs
and/or MDCs in the Presence and in the Absence of RBCs
Assay Conditions
Extracellular Recovery�
H202 HOCI
PMNs 15.3 ± 2.7 14.8 ± 1.2
MDCs 7.4±1.0 0
PMNs + MDCs 21.3 ± 3.5 14.1 ± 1.4
PMNs + RBCs 7.6 ± 1.3 8.1 ± 0.9
MDCs + RBCs 4.6 ± 1.7 0
PMNs + MDCs + RBCs 12.3 ± 1.4 12.8 ± 1.2
For personal use only. by guest on July 10, 2011. bloodjournal.hematologylibrary.orgFrom

Co
50
40
30
20
�l0
6 8 10
HOCI recovery I I) mel
12 14 16
20
�1oC.,
r’ir’iA B C 0
1746 DALLEGRI ET AL
H2O2 and HOCI recovery (-50%) was found (Table 2). This
reduction in HOC1 generation results in a relative impair-
ment of the HOCI-dependent PMN lytic efficiency.6 As
summarized in Table 2, MDCs increased the H2O2 and
HOC1 recovery from the PMN-RBC system. Moreover,
when the extent ofthe lysis by PMNs and PMNs plus MDCs
(Figs 1 and 2) was plotted as a function of the HOCI
recovery detected under conditions similar to those used for
cytolysis (identical cell ratios but with 2 x 106 RBCs instead
of l0� RBCs used in lytic assays), a positive linear relation-
ship was found (Fig 3). Together, these results suggest the
possibility that MDCs amplify the lysis primarily by improv-
ing the HOCI production in the PMN-RBC system, possibly
through an effective restoration of the PMN H202 extracel-
lular pool depleted by RBC targets. Three additional find-
ings are consistent with this possibility. First, the replace-
ment of RBC targets with RBCAT (RBCAI had a >94%
reduction in catalase activity; catalase activity of native
RBCs 4,258 ± I 12 nmol H2O2/min/lO6cells, � ± 1 SD,
n = 6), incapable of efficiently consuming PMN-derived
H202,6 completely abrogated the MDC capacity to augment
the PMN-mediated HOCI-dependent (not shown) lysis (per-
cent increment, 1.1 ± 3.7, i ± 1 SD, n = 3) and the HOC1
recovery from the system (percent increment, -0.02 ± 8.2,
� ± I SD, n = 4). Second, enhancement of the HOCI pro-
duction by MDCs in the PMN-RBC system (Table 2) could
be reproduced by substituting catalase for RBCs in the
system (Table 3) and choosing an amount of enzyme (300
units in I mL) that results in a reduction of the HOC1
recovery from PMNs similar in magnitude to that observed
in the presence of 2 x 106 RBCs (Tables 2 and 3). The
requirement for 300 units catalase, ie, � 35 times the amount
of catalase in 2 x 106 RBCs, is conceivably due to the fact
that the enzyme is distributed throughout the assay volume
(I mL) whereas RBC catalase is confined to the cell pellet
(the HOC1 assays were carried out with cells as a pellet to
Table 3. Recovery of HOCI From OPZ-Triggered PMNs and/orMDCs in the Presence and in the Absence of Catalase
Assay Con�tions HOCI Recoveryt
PMNs
MDCs
PMNs + MDCs
PMNs + catalase
MDCs + catalase
PMNs + MDCs + catalase
15.6 ± 1.9
0
15.7 ± 1.3
8.8 ± 1.4
0
14.3 ± 1.0
PMNs, 5 x i0�; MDCs, 5 x iOn; catalase, (bovine liver, Sigma).
300 units.
tIn nmol/20 mm, � ± 1 SD of three experiments.
simulate the conditions used for measuring the lysis). Third,
the PMN-mediated RBC lysis was augmented by 5 mU/mL
GO, per se virtually ineffective (Fig 4). The lysis was
prevented by the MPO inhibitor azide as well as by the
HOC1-trapping agent taurine (Fig 4). In addition, 5 mU/mL
GO enhanced the recovery of HOC1 from the PMN-RBC
system by 46.9% + 5.2% (� ± 1 SD, n = 3), against an
expected value of 55% (value calculated on the basis of the
regression line from Fig 3 by using the extent of the RBC
lysis by PMNs plus GO as known variables).
DISCUSSION
In our model system as reported elsewhere6 and herein
confirmed by probing cytolysis with chemical agents, the
RBC lysis by PMNs requires HOC1 production via the
MPO-catalyzed oxidation of chloride ions by H202.
Although the cytolysin or cytolysins actually operative
remain to be identified,”�#{176}’2’ the extent of the lysis strictly
correlates with the amount of HOC1 generated by PMNs, at
least under our experimental conditions (the present paper
and Dallegri et al6). Consistent with this view, MPO-
negative MDCs incapable of generating HOC1 did not cause
lysis unless supplemented with MPO activity from a super-
40
30
Fig 3. Positive linear relationship (P < .01) between the RBClysis mediated by PMNs (#{149})and PMNs plus MDCs (�) (data fromFigs 1 and 2) and the HOCI recovery (nmol/20 mm) from PMNs (#{149})and PMNs plus MDCs (U) in presence of RBC targets (as for thelytic assays. the number of RBCs was kept constant. 2 x 10). Eachpoint represents � ± 1 SD of at least three experiments. Regres-sion line, y - 5.436 x -31.266.
Fig 4. Effect of GO on the RBC lysis by OPZ-triggered PMNs.The ordinate shows the percent lysis by (A) 0.25 x 10’ PMNs. (B) 5mU/mi GO. (C) 0.25 x i0� PMNs plus 5 mU/mL GO. (D) 0.25 x 10’PMNs plus 5 mU/mi GO plus 1 mmol/L azide. and (E) 0.25 x 10’PMNs plus 5 mU/mi GO plus 20 mmol/L taurine. The number ofRBCs is 10’/welI. Results are expressed as � ± I SD of threeexperiments.
For personal use only. by guest on July 10, 2011. bloodjournal.hematologylibrary.orgFrom

MDCS ENHANCE RBC LYSIS BY PMNs 1747
natant of the PMN granule fraction (unpublished observa-
tions). Nevertheless, they efficiently augmented both HOC1
recovery from the PMN-RBC lytic system and the HOC1-
dependent lysis.
A series of interconnected events underlies the expression
of the observed MDC “helper” activity. After the interaction
between the PMN surface and OPZ, a plasmalemma-
associated NADPH oxidase system is activated in the area of
PMN-OPZ contact,22 with consequent H202 production. The
generated H202 is released directly into the extracellular
fluid until the OPZ particle is completely engulfed. Part of
this H2O2 can probably undergo backdiffusion into PMNs
themselves.23 At the luminal surface of the phagosomal
membrane, which is derived from the plasmalemma, H202
production continues,24 and the generated H202 can diffuse
into PMN cytoplasm and outside the cell.25 The net amount
of H202 released extracellularly (extracellular H202 pool)
represents that fraction of H202 synthesized and not used by
PMN catabolic pathways in different subcellular compart-
ments, including MPO within phagosomes5’2’ and the cata-
lase and glutathione cycle within the cytosol.2�28 Although
discrepancies exist among different authors,’3’29 in our set-
ting the extracellular H2O2 pool is likely to be transformed
entirely into HOCI by the MPO released into the medium or
absorbed to the PMN surface. In fact, HOCI production by
OPZ-triggered PMNs could not be augmented by adding an
appropriate surplus of MPO activity, which suggests that the
H2O2 available for the chlorinating reaction is entirely used
by the released MPO. Moreover, HOC1 production could not
be augmented by biochemical (GO) or cellular (MDCs)
extra sources of H2O2, which suggests that the amount of
MPO released functions at a maximum rate without a
potential reserve of activity. Thus, it appears that OPZ-
triggered PMNs produce a definite amount of HOC1 by
releasing MPO and H2O2 via a process so balanced that
excesses of MPO and H202 are virtually absent or, if present,
trivial at least under our conditions (pH, C I concentration,
temperature). Although the H202 released and transformed
into HOCI represents a relatively small fraction of the total
amount of H202 produced (Nauseef et al,26 Test and Weiss,27
and unpublished observations) and is generally assumed to
be in dynamic equilibrium with the remainder (primarily
dissipated by the PMN catabolic pathways mentioned ear-
her), it can be expected that the addition of extra H202-
consuming pathways to the PMN microenvironment can
perturb the equilibrium of the system so much that subse-
quent HOC1 production is impaired. RBCs added to OPZ-
triggered PMNs to make up a complete cytolytic system
reduce the HOCI production by consuming H202 primarily
via catalase (Dallegri Ct al,6 Test and Weiss,27 and the
present paper). Such a consumption of H202, which results in
a relative impairment of HOC1-dependent PMN lytic effi-
ciency,6 is likely to take place within RBC targets before
lysis. In fact, the PMN-delivered lethal hit and the RBC
catalase-mediated impairment of the lysis occur during the
first 45 minutes of incubation when target cells are still
“viable” (changes in the final extent of the lysis could not be
detected when exogenous catalase was added to block the
lytic pathway 45 minutes after the start of the incubation
period, lysis at this time being virtually absent; unpublished
observations). This is consistent with results obtained by
other authors in a different system�#{176}and with the fact that
H202, having a relatively low reactivity and no charge, can
diffuse away from its sites of formation and across cell
membranes easily.3’ Although attempts to directly measure
the actual size of the extracellular H202 pool in our cytolytic
system with cells as a pellet (without adding extra H202-
consuming pathways, for instance, HRP) are presently ham-
pered by the nonexistence of a reliable H202 assay, the
results herein reported provide inferential evidence that the
reduction of the PMN extracellular H202 pool by RBC
catalase and the reconstitution of this pool by MDCs are
crucial to MDC-mediated augmentation of PMN HOC1
production and, in turn, lysis. This is suggested by different
findings. First, neither HOC1 recovery nor the extent of the
lysis could be augmented by MDCs after inhibition of the
RBC H202-consuming capacity, ie, using RBCAT as targets.
Second, enhancement of HOC1 production by MDCs in the
PMN-RBC system could be reproduced by substituting an
appropriate amount of reagent catalase for RBCs in the
system. Third, MDC-dependent enhancement of both HOCI
recovery from the PMN-RBC system and PMN-mediated
lysis could be reproduced by replacing MDCs with an
appropriate H202-generating enzymatic system (GO, Fig 4)
or H202-generating, granule-depleted PMN cytoplasts
obtained as described by Roos et al32 and unpublished
observations). Fourth, the “final” events occurring in our
system in the presence and in the absence of MDCs (amount
of HOC1 produced and extent of the lysis) strictly correlate
(Fig 3). Thus, on the basis of these observations coupled with
the evidence that MDCs are per se cytolytically ineffective
and with the results obtained by probing the lytic systems
with chemical agents, we conclude that (a) MDCs amplify
the lysis by improving HOC1 production, presumably via an
effective restoration of the extracellular H202 pool generated
by PMNs and depleted by RBCs; and (b) if present, the
intervention of nonoxidative cytolytic factors, released by
MDCs and capable of contributing to the lysis,33 would be
trivial at least under the present conditions. In addition, the
involvement of PMN-directed cytokines released by MDCs
and capable of increasing the HOC1 production can be ruled
out because MDCs did not affect the HOCI recovery from
PMNs in absence of RBCs. Similarly, the possibility that
MDCs may act by releasing NH4�, which favors NH2CI
formation (NH4� + HOC1 -b NH2CI + H20 + � can
be ruled out because exogenous NH4� inhibited the lysis in
the PMN-RBC system thus suggesting that lytic levels of
NH2C1 cannot be reached in our conditions. Finally, because
MDCs had no effect when added to the PMN�RBCAI
system, it is unlikely that they act by sensitizing RBC targets
to the PMN-delivered, HOC1-mediated attack.
In conclusion, it has been reported that macrophages
ingesting effete PMNs or granules from PMNs become
capable of mediating MPO-dependent iodination.35 Further,
macrophages can use eosinophil peroxidase (EPO) to
enhance their microbicidal or cytolytic activity?�38 These
results have suggested a way by which MPO-negative and
MPO-positive phagocytes can interact synergistically in
For personal use only. by guest on July 10, 2011. bloodjournal.hematologylibrary.orgFrom

REFERENCES
1748 DALLEGRI ET AL
exerting certain effector functions. Nevertheless, the afore-
mentioned experiments were performed with cellular debris
or purified cellular components (EPO, MPO). In the present
study using two complete viable cellular systems, ie, PMNs
and MPO-negative MDCs, we attained direct evidence for
cooperation between two distinct populations of phagocytes
in mediating cytolysis. Although speculative, the present use
of MDCs as a model raises the possibility that macrophages
at inflammatory sites may interact with neutrophils to
increase the local HOC1 production and, in turn, the injury of
I . Klebanoff Si: Oxygen-dependent cytotoxic mechanisms of
phagocytes, in Gallin JI, Fauci AS (eds): Advances in Host DefenseMechanisms, vol 1 . New York, Raven, 1982, p 1 1 1
2. Weiss Si, LoBuglio AF: Phagocyte-generated oxygen metabo-lites and cellular injury. Lab Invest 47:5, 1982
3. Fantone iC, Ward PA: Role of oxygen-derived free radicalsand metabolites in leukocyte-dependent inflammatory reactions.
Am i Pathol 107:397, 19824. Clark RA: Extracellular effects of the myeloperoxidase-
hydrogen peroxide-halide system, in Weissmann G (ed): Advancesin Inflammation Research, vol 5. New York, Raven, 1983, p 146
5. Hamers MN, Roos D: Oxidative stress in human neutrophilicgranulocytes: Host defense and self-defense, in Sies H (ed): Oxida-
tive Stress. Orlando, FL, Academic, 1985, p 3516. Dallegri F, Ballestrero A, Frumento G, Patrone F: Interfer-
ence of target cell catalase with an early step of the neutrophilcytolytic pathway. Clin Exp Immunol 65:664, 1986
7. Dankberg F, Persidsky MA: A test of granulocyte membraneintegrity and phagocyte function. Cryobiology 13:430, 1976
8. Packard 85, Tavassoli M, Dale GL, Beutler E: A method for
the establishment and long-term maintenance of in vitro monocyticcultures with functional and morphological homogeneity. Blood
60:623, 19829. Kumagai K, Itoh K, Hinuma 5, Tada M: Pre-treatment of
plastic Petri dishes with fetal calf serum. A simple method formacrophage isolation. i Immunol Methods 29:17, 1979
10. Li CY, Lam KW, Yam LT: Esterases in human leukocytes. JHistochem Cytochem 21 :1, 1973
1 1 . Dallegri F, Patrone F, Ballestrero A, Frumento G, SacchettiC: Inhibition of neutrophil cytolysin production by target cells.B1ood67:1265, 1986
12. Ruch W, Cooper PH, Baggiolini, M: Assay of Hf)2 produc-
tion by macrophages and neutrophils with homovanillic acid andhorse-radish peroxidase. i Immunol Methods 63:347, 1983
I 3. Weiss Si, Klein R, Slivka A, Wei M: Chlorination of taurineby human neutrophils. Evidence for hypochlorous acid generation. J
Clin Invest 70:598, 198214. Aune TM, Thomas EL: Accumulation ofhypothiocyanite ion
during peroxidase-catalyzed oxidation of thiocyanate ion. Eur i
Biochem 80:209, 197715. Thomas EL, Grisham MB, Melton DF, Jefferson MM:
Evidence for a role of taurine in the in vitro oxidative toxicity of
neutrophils toward erythrocytes. i Biol Chem 260:3321, 198516. Bradley PP, Christensen RD. Rothstein G: Cellular and
extracellular myeloperoxidase in pyogenic inflammation. Blood
60:618, 1982
17. Beutler E: Red Cell Metabolism. A Manual of BiochemicalMethods. Orlando, FL, Grune & Stratton, 1984, p 105
I 8. Dallegri F, Ballestrero A, Frumento G, Patrone F: Erythro-cyte lysis by PMA-triggered neutrophil polymorphonuclears: Evi-
dence for an hypochlorous acid-dependent process. Immunology
55:639, 1985
surrounding tissue cells. This might be only one of the events
amplified by macrophages and relevant to the outcome of the
inflammatory process. In fact, aside from the role in promot-
ing cytolysis, phagocyte-derived HOCI and/or its chlori-
nated derivatives have recently been recognized to mediate a
variety of biologic effects, either protective or harmful to the
host,21’39 ranging from the destruction of microorganisms�#{176}
and modulation of lymphocyte function4’ to the inhibition of
antiproteinase activities42 and inactivation of chemotactic
factors.43
19. Dallegri F, Ballestrero A, Frumento G, Patrone F: Role ofhypochlorous acid and chloramines in the extracellular cytolysis by
neutrophil polymorphonuclear leukocytes. J Clin Lab Immunol
20:37, 198620. Grisham MB, Jefferson MM, Melton DF, Thomas EL:
Chlorination of endogenous amines by isolated neutrophils. Ammo-nia-dependent bactericidal, cytotoxic and cytolytic activities of the
chloramines. J Biol Chem 259:10404, 1984
21. Test ST. Weiss Si: The generation and utilization of chlori-
nated oxidants by human neutrophils. Adv Free Radical Biol Med
2:91, 198622. Whitin JC, Ryan DH, Cohen Hi: Graded responses of human
neutrophils induced by serum-treated zymosan. Blood 66: 1 182,
198523. Ohno Y, Gallin JI: Diffusion ofextracellular hydrogen perox-
ide into intracellular compartments of human neutrophils. Studiesutilizing the inactivation of myeloperoxidase by hydrogen peroxideand azide. i Biol Chem 260:8438, 1985
24. Ohno Y, Hirai KI, Kanoh T, Uchino H, Ogawa K: Subcellu-lar localization of H202 production in human neutrophils stimulatedwith particles and effect ofcytochalasin B on the cells. Blood 60:253,
198225. Root RK, Metcalf JA: H202 release from human granulo-
cytes during phagocytosis. Relationship to superoxide anion forma-tion and cellular catabolism of H2O2: Studies with normal andcytochalasin B-treated cells. i Clin Invest 60: 1266, 1977
26. Nauseef WM, Metcalf iA, Root RK: Role of myeloperoxi-dase in the respiratory burst of human neutrophils. Blood 61:483,
198327. Test ST. Weiss Si: Quantitative and temporal characteriza-
tion of the extracellular H202 pool generated by human neutrophils.J Biol Chem 259:399, 1984
28. Cohen Hi, Tape EH, Novak i, Chovaniec ME, Liegey P,Whitin JC: The role of glutathione reductase in maintaining human
granulocyte function and sensitivity to exogenous H2O2. Blood69:493, 1987
29. Thomas EL, Grisham MB, iefferson MM: Myeloperoxidase-dependent effect of amines on functions of isolated neutrophils. iClin Invest 72:441, 1983
30. Weiss Si, Slivka A: Monocyte and granulocyte-mediatedtumor cell destruction. A role for the hydrogen peroxide-myelo-peroxidase-chloride system. i Clin Invest 69:255, 1982
31. Halliwell B, Gutteridge MC: Oxygen toxicity, oxygen radi-
cals, transition metals and disease. Biochem J 219: 1 , 1984
32. Roos D, Voetmann AA, Meerhof Li: Functional activity ofenucleated human polymorphonuclear leukocytes. J Cell Biol
97:368, 198333. Nathan CF: Secretory products of macrophages. J Clin
Invest 79:319, 1987
34. Grisham MB, Jefferson MM, Thomas EL: Role of monochlo-ramine in the oxidation of erythrocyte hemoglobin by stimulated
neutrophils. J Biol Chem 259:6757, 1984
For personal use only. by guest on July 10, 2011. bloodjournal.hematologylibrary.orgFrom

MDCS ENHANCE RBC LYSIS BY PMNs 1749
35. Heifets L, Imai K, Goren MB: Expression of peroxidase-dependent iodination by macrophages ingesting neutrophil debris. JReticuloendothel Soc 28:391 , I 980
36. Locksley RM, Wilson CB, KlebanoffSJ: Role for endogenous
and acquired peroxidase in the toxo-plasmacidal activity of murineand human mononuclear phagocytes. J Clin Invest 69:1099, 1982
37. Nathan CF, Klebanoff Si: Augmentation of spontaneous
macrophage-mediated cytolysis by eosinophil peroxidase. J Exp
Med 155:1291, 1982
38. Ramsey PG, Martin T, Chi E, Klebanoff SJ: Arming ofmononuclear phagocytes by eosinophil peroxidase bound to Staphy-
lococcus aureus. J Immunol 128:415, 198239. Babior BM: Oxidants from phagocytes: Agents of defense
and destruction. Blood 64:959, 1984
40. Passo SA, Weiss Si: Oxidative mechanisms utilized by
human neutrophils to destroy Escherichia coli. Blood 63:1361,1984
41. El-Hag A, Lipsky PE, Bennett M, Clark RA: Immunomodu-lation by neutrophil myeloperoxidase and hydrogen peroxide: Differ-
ential susceptibility of human lymphocyte functions. J Immunol136:3420, 1986
42. Weiss Si, Regiani 5: Neutrophils degrade subendothelialmatrices in the presence of alpha-l-proteinase inhibitor. Cooperativeuse of lysosomal proteinases and oxygen metabolites. J Clin Invest
73:1297, 1984
43. Clark RA, Szot 5, Venkatasubramanian K, Schiffmann E:Chemotactic factor inactivation by myeloperoxidase-mediated oxi-dation of methionine. J Immunol 124:2020, 1980
For personal use only. by guest on July 10, 2011. bloodjournal.hematologylibrary.orgFrom





























