Embed Size (px)
Citation preview

DEVELOPMENTAL BIOLOGY 184, 61–69 (1997)ARTICLE NO. DB978520
Autocrine Regulation of Neural Crest CellDevelopment by Steel Factor
Chang Sheng Guo,* Bernhard Wehrle-Haller,†John Rossi,* and Gary Ciment*,1
*Department of Cell and Developmental Biology, Oregon Health Sciences University,Portland, Oregon 97201; and †Institute for Neuroscience, University of Oregon,Eugene, Oregon 97403
Steel factor (SLF) and its cognate receptor, c-kit, have been implicated in the generation of melanocytes from migratingneural crest (NC) cells during early vertebrate embryogenesis. However, the source of SLF in the early avian embryo andits precise role in melanogenesis are unclear. We report here that NC cells themselves express and release SLF protein,which in turn acts as an autocrine factor to induce melanogenesis in nearby NC cells. These results indicate that NC cellsubpopulations play an active role in the determination of their cell fate and suggest a different developmental role for theembryonic microenvironment than what has been previously proposed. q 1997 Academic Press
INTRODUCTION nous factors include various peptide growth factors forwhich migrating NC cells express receptors, includingthose for platelet-derived growth factor (PDGF), which hasThe neural crest (NC) is a transient structure observedbeen implicated in craniofacial development (Grunebergduring vertebrate embryogenesis as the neural epitheliumand Truslove, 1960; Morrison-Graham et al., 1992), andfolds to form the neural tube (Le Douarin, 1982; Weston,the neurotrophins, which may be involved in neurogenesis1991). NC cells originate from within the neural tube, but(Sieber-Blum, 1991; Pinco et al., 1993).soon migrate into the periphery of the embryo, traveling
One factor which has been proposed to regulate melano-along two migratory pathways. NC cells in the dorsal–lat-genesis in NC cells is Steel Factor [SLF; also known as c-eral pathway migrate through the space between the ecto-kit ligand, mast cell growth factor, or stem cell factor (Galliderm and dermamyotome of the somites and eventuallyet al., 1994)]. This glycoprotein is expressed in a variety ofgive rise to the melanocytes of the skin. The ventral NCtissues as a transmembrane protein, which is then cleavedmigratory pathway is within the mesenchyme of the form-at a proximal extracellular site, releasing the soluble pro-ing sclerotome, as well as in the space between adjacenttein. Mutations of the SLF gene in mice, or of its cognatesomites (Bronner-Fraser, 1986). NC cells migrating alongreceptor, the c-kit tyrosine kinase, in mice, rats, and hu-the ventral pathway eventually give rise to a wide varietymans, produce animals lacking pigment cells (Williams etof different cell types, including sensory and autonomical., 1992). Injection of c-kit neutralizing antibodies intoneurons, Schwann cells, and adrenal chromaffin cells.pregnant mice has been shown, moreover, to result in theStudies performed in culture and in vivo suggest that thehypopigmentation of offspring (Nishikawa et al., 1991).fate of NC cells is regulated to a large part by the microenvi-
The simple notion of SLF as an environmental factor in-ronment (Anderson, 1989; Weston, 1991; Leblanc and Bron-ducing melanogenesis in NC cells is, however, complicatedner-Fraser, 1992). These studies have led to the hypothesisby two observations. First, the SLF mRNA does not seemthat different exogenous factors (or combinations of factors)to be expressed by cells making up the microenvironmentproduced by different microenvironments are responsiblethrough which melanogenic NC cells migrate in avian em-for the generation of cell type diversity either by influenc-bryos. In situ hybridization studies of chicken embryosing NC cell fate in multipotent cells or by selecting forshow no detectable signal in dorsal aspects of the dermamy-predetermined subpopulations. Candidates for such exoge-otome or ectoderm at the appropriate developmental stages,although there is some hybridization signal in more lateralregions of the ectoderm (Lecoin et al., 1995). Second, mu-1 To whom correspondence and reprint requests should be ad-
dressed. Fax: (503) 494-4253. E-mail: [email protected]. tant mice that express only the soluble form of SLF lack
61
0012-1606/97 $25.00Copyright q 1997 by Academic PressAll rights of reproduction in any form reserved.
AID DB 8520 / 6x1e$$$181 03-20-97 21:06:54 dbal

62 Guo et al.
tioned medium (Developmental Hybridoma Studies Bank)] andpigment cells in the skin (Flanagan et al., 1991), indicatingthen secondary antibodies [fluorescein-labeled goat anti-rabbit IgGthat the full-length, membrane-bound form of SLF is re-(Vector Laboratories) and rhodamine-labeled goat anti-mouse IgMquired for melanogenesis, at least in vivo.(Cappel Laboratories)]. The cultures were then washed briefly withThe cellular mechanism(s) by which SLF induces melano-PBS, and the central portions of each plate were cut out on a smallgenesis in the early embryo is also unclear. On the onelathe and mounted (with the cells facing up) onto a microscope
hand, it has been suggested that SLF acts as a survival factor, slide. The surface of each disk was then covered with a 1:1 mixtureselecting for the survival and/or proliferation of committed, of PBS with 2 mg/ml phenylenediamine and glycerol (to preventlatent melanoblasts in heterogeneous NC cell populations. fluorescence fading) and coverslipped. Cultures were then viewedThis conclusion is supported by culture studies using under fluorescein and rhodamine epifluorescence optics on a Zeiss
Axioplan microscope and photographed using an Olympus photo-mouse NC cells in which removal of SLF from the culturemicroscopy system.medium results in the irreversible loss of pigmentation-
competent cells or cells expressing melanocyte markers(Murphy et al., 1992; Morrison-Graham and Weston, 1993; Western Blot AnalysisReid et al., 1995). Conversely, SLF may influence the com-
Fifty micrograms of protein from homogenates of E6 chickensmitment of multipotent NC cells to a melanocyte fate and/was subjected to electrophoresis in an SDS–polyacrylamide gel andor influence their subsequent differentiation.then transferred electrophoretically onto a nitrocellulose mem-
In this paper, we show that embryonic quail NC cells brane. The membrane was preincubated with 5% nonfat dry milkthemselves express both the SLF mRNA and protein in vivo in PBS with 0.1% Triton X-100 for 1 hr and then treated overnightand in culture and that SLF acts as an autocrine factor regu- with anti-SLF antibodies (1:2000 dilution in PBS with 0.1% Tritonlating cell fate. X-100). After extensive washing, the blot was incubated with horse-
radish peroxidase-conjugated goat anti-rabbit antibodies (1:5000 di-lution in PBS with 0.1% Triton X-100), and peroxidase activitywas visualized using the enhanced chemiluminescence (ECL) kitMATERIALS AND METHODS(Amersham, Arlington Heights, IL).
Cell CultureMorphometric AnalysesNeural tubes were isolated as described (Loring et al., 1982;
Stocker et al., 1991) from stage 12 and 13 quail embryos (Coturnix Morphometric analysis of the numbers of cell processes and theircoturnix japonica) (Hamburger and Hamilton, 1951) and cultured lengths was performed using an eyepiece reticle containing a serieson either 35-mm or 24-well Primaria tissue culture plates (Falcon of calibrated, concentric circles. Cells were then picked at randomPlastics, Ventura, CA). In each of the 24-well plates, one neural and moved to the center of the reticle. The length of processes wastube (corresponding to a 10-somite length) was plated as the source determined by calculating the shortest distance from the proximalof NC cells; in each of the 35-mm dishes, four to six neural tubes to the distal ends of each process. Only those cell processes of onewere used. The culture medium was CCM-5 (Hyclone, Logan, UT), cell diameter or longer were included in these calculations. Ofwhich is a serum- and embryo extract-free defined culture medium. course, this approach does not take into consideration the absoluteIt contains insulin-like growth factor type I, transferrin, and bovine length of those processes that do not travel along a straight lineserum albumin as the only proteins, but lacks phorbol esters, reti- and therefore tends to underestimate process lengths.noids, and other agents known to influence neural crest cell fate.
In Situ HybridizationImmunocytochemistry In situ hybridization was carried out as described previously
(Wehrle-Haller and Weston, 1995). Briefly, embryos were first pro-To generate anti-SLF antibodies, recombinant chicken SLF pro-cessed for whole-mount in situ hybridization using digoxygenin-tein was made by subcloning a cDNA corresponding to the extracel-labeled antisense RNA probes generated by in vitro transcriptionlular domain (amino acids 16–181) into the pTrcHis prokaryoticof a cDNA corresponding to the extracellular domain of chickenexpression system (Invitrogen, San Diego, CA) and then purifyingSLF. Hybridization signal was then detected by immunohisto-the recombinant protein over a nickel column. Rabbits were in-chemistry using rabbit anti-digoxygenin antibodies conjugatedjected a total of six times with the recombinant protein over awith alkaline phosphatase. Embryos were then embedded in paraf-period of 6 weeks. The first injection was a mixture of 150 mgfin and sectioned at 10 mm on a microtome. Sense RNA probesprotein in complete Freund’s adjuvant injected subcutaneously; theserved as negative controls. Attempts to perform colabel the samesecond was similar to the first, but in incomplete Freund’s adju-sections by in situ hybridization for SLF and immunostaining forvant. The remaining injections were performed using recombinantthe HNK-1 epitope were unsuccessful due to the lability of theSLF (300–600 mg protein) dialyzed against phosphate-buffered sa-HNK-1 epitope to the alcohol extraction steps necessary for in situline and injected into the ear vein.hybridization.NC cell cultures were fixed for 30 min with 4% paraformalde-
hyde in Hanks’ balanced salt solution supplemented with 25 mMHepes buffer (pH 7.6). After extensive washing, the cultures were Analysis of SLF mRNA Using Reversepreincubated overnight with ‘‘antibody dilution buffer’’ consisting Transcription/Polymerase Chain Reactionof phosphate-buffered (10 mM, pH 7.4) isotonic saline (PBS) con- (RT/PCR) and Southern Blotstaining 0.1% Triton X-100 and 10% goat serum. Cultures werethen incubated with primary antibody [1:300 dilution of rabbit anti- RNA was isolated from NC cell cultures using a kit (Qiagen,
Chatsworth, CA) and transcribed using Superscript reverse tran-chicken SLF; 1:50 dilution of mouse anti-HNK-1 hybridoma condi-
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8520 / 6x1e$$$181 03-20-97 21:06:54 dbal

63Expression of Steel Factor in Neural Crest Cells
scriptase (Gibco-BRL, Gaithersburg, MD), following standard meth- liver, brain, heart, kidney, and lung. This molecular sizeods (Kawasaki, 1990). The cDNA was then subjected to polymerase corresponds to that of the soluble form of SLF as predictedchain reaction (PCR) amplification (30 cycles with an annealing by the sequence of the chicken SLF cDNA (Zhou et al.,temperature of 507C) using primers corresponding to the published 1993), and these tissues correspond to locations in whichsequence of chicken SLF (Zhou et al., 1993). These primers were SLF has been observed in mouse (Matsui et al., 1990). This5*-GCCTGCCTAATCACTGTTGGTTG-3* and 5*-AAGCCAAG-
immunoreactivity was not present when the antibodiesTGCCTCTTTGTTACTG-3*, which generated a 466-base-pair re-were preincubated with recombinant SLF (data not shown).action product corresponding to amino acid residues 64 through
To determine whether this immunoreactivity in NC cul-218. The PCR product was transferred to Genescreen Plus filterstures was bon fide SLF or a related gene product, RNA was(NEN, Boston, MA) and the blot was probed with a nick-translated
32P-labeled cDNA (Prime-It, Stratagene, La Jolla, CA) corresponding prepared from similar cultures and processed for RT/PCRto a 501-base-pair HindIII/PstI restriction fragment of the pTrcHis- amplification using primers corresponding to SLF. Figure 3SLF plasmid. Finally, autoradiography was performed using Dupont is a Southern blot of this amplification reaction probed forReflection film and processed by an X-OMAT X-ray film processor. SLF and shows a band at the appropriate size in the lane
For DNA sequence analysis, the PCR product was subcloned prepared from NC cell cultures (lane 4). This band in theinto the pTA cloning vector (Invitrogen) and automated sequence NC cell lanes was not present when the reverse tran-analysis was performed on an Applied Biosystems 373 sequencer.
scriptase step was omitted (lane 5), indicating that the origi-nal template was RNA and not contaminating genomicDNA. Sequence analysis of these bands confirmed that they
RESULTS were highly homologous to the published sequence of quailSLF (GenBank accession No. U43078), with only two con-servative amino acid substitutions, which probably corre-Although the product of the SLF gene has clearly been
implicated in melanogenesis during development (Galli et spond to polymorphisms present in this outbred species.To determine whether NC cells express SLF at the rele-al., 1994), the cellular source of SLF in the embryo is still
unclear. Although some SLF mRNA signal has been re- vant ages in vivo, in situ hybridization studies were per-formed. Figure 4 shows the earliest observed expression ofported in the lateral ectoderm of later stage embryos by in
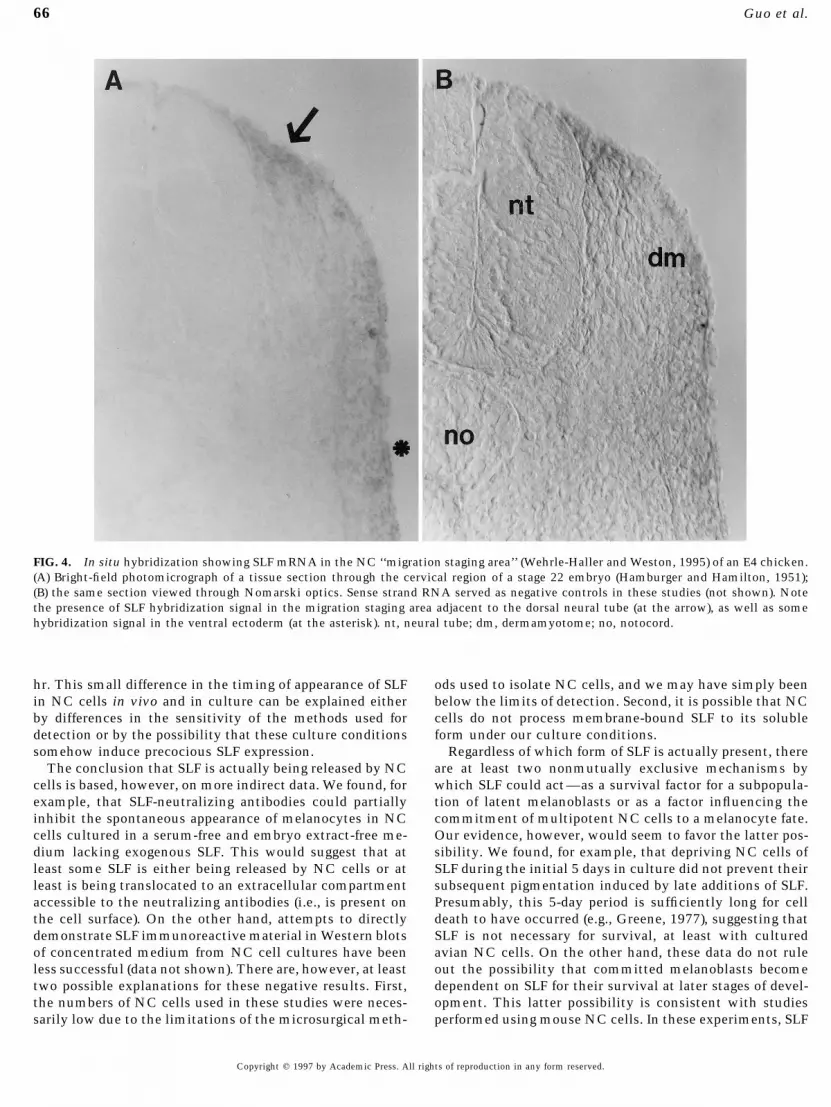
situ hybridization (Matsui et al., 1990; Keshet et al., 1991; SLF hybridization signal in the dorsal NC migratory spaceof the upper cervical regions of an E4 chicken (at the arrow),Lecoin et al., 1995; Wehrle-Haller and Weston, 1995), these
sites and stages are different from those where melanoblasts corresponding to the region described as the migration‘‘staging area’’ (Wehrle-Haller and Weston, 1995). This re-first appear. This suggests that other sources of SLF may
also be present in the embryo. One formal possibility, of gion contains NC cells which remain in close physical asso-ciation for a period of 24–36 hr until the space betweencourse, is that NC cells themselves express SLF and that
this growth factor acts in an autocrine fashion to influence the dermamyotome and ectoderm opens to allow migrationunder the presumptive skin (Weston et al., 1978). Figure 4melanogenesis. To address this possibility, we immuno-
stained cultured NC cells with monospecific antibodies also shows SLF hybridization signal in the surface ectodermat ventral locations, as had been shown earlier (Lecoin etgenerated against the extracellular portion of this growth
factor. Figure 1 shows that the vast majority of quail NC al., 1995), but not in the neural tube nor in the sclerotome.Hybridization signal was not observed in the dorsal NCcells (approximately 98%) cultured for 48 hr in a serum-
and embryo extract-free defined medium expressed SLF-im- migratory space at more caudal levels of these embryos, norat any level in younger embryos (data not shown). Thesemunoreactivity (see also Table 1). The immunostaining pat-
tern showed a punctate appearance within the cytoplasm, results suggest that NC cells localized within the stagingarea express the SLF gene product in vivo and are consistentas well as some cell surface staining. Similar staining pat-
terns and proportions of SLF/ cells were also observed in with observations that NC cells emigrating from the stagingarea have become committed to a melanogenic fate by theseNC cultures grown for periods as short as 12 hr or in the
presence of 10% fetal bovine serum (data not shown). Stud- developmental stages (Vogel and Weston, 1988; Erickson etal., 1992; Rogers et al., 1992).ies with antisera against the chicken c-kit gene product
showed a similar pattern of staining to that of SLF, with One important remaining question, however, is whetherSLF acts by supporting the survival of melanocyte precursorsabout 96% of HNK-1/ NC cells expressing c-kit immunore-
active material (not shown). Although we were unable to or by influencing the commitment of multipotent NC cells.To distinguish between these possibilities, we performed aimmunostain cultures simultaneously for both c-kit and
SLF (since both antisera were generated in rabbits), the high series of culture experiments. In the first set of studies, NCcells were grown in serum- and embryo extract-free definedproportions of SLF/ or c-kit/ cells indicate that most NC
cells in these cultures express both the growth factor and medium in the presence of either SLF or bFGF, as a positivecontrol for melanogenesis (Stocker et al., 1991). Figure 5Aits receptor.
As a control for antibody specificity, Western blot experi- shows that SLF was more potent than bFGF at inducingpigmentation in these cultures and that antibodies againstments were performed. Figure 2 is a protein immunoblot
showing that this anti-recombinant SLF antibody recog- SLF partially neutralized the effects of SLF. Similar propor-tions of pigmented cells were also observed when SLF andnizes only a single band at around 23 kDa in homogenates
of various tissues dissected from E14 chickens, including bFGF were added to the medium 24 hr after the neural tubes
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8520 / 6x1e$$$181 03-20-97 21:06:54 dbal

64 Guo et al.
FIG. 1. Presence of SLF immunoreactivity in embryonic quail NC cells in culture. NC cells were isolated as described (Loring et al.,1982; Stocker et al., 1991) from stage 12 and 13 quail embryos (Hamburger and Hamilton, 1951) and cultured in serum- and embryoextract-free defined medium (CCM-5; Hyclone, Logan, UT). After 2 days, the cultures were fixed and double-immunostained for SLF andHNK-1 as a marker of NC cells (Bronner-Fraser, 1986). (A) SLF immunostaining of NC cells; (B) the same field of cells as in A immunostainedfor HNK-1; (C) preimmune serum substituted for anti-SLF serum; (D) the same field of cells as in C immunostained for HNK-1. The barat the upper right in B corresponds to 10 mm.
were placed in culture. This suggests that the effects of these with fresh medium. Presumably, if SLF was a survival fac-tor, then the lack of SLF during the initial 5-day periodgrowth factors were not due to initial differences in the types
of NC cells that migrated onto the culture surface. should result in the irreversible loss of most SLF-dependentcells. Figure 5B shows, however, that cultures lacking SLFInterestingly, the morphology of the pigmented cells in-
duced by SLF or bFGF also differed in our cultures. Figures during the initial 5 days (‘‘ø:SLF’’) showed only a 2-day delayin the onset of pigmentation, compared with cultures5C and 5D show that NC cells cultured for 14 days in the
presence of SLF display a more mature melanocyte mor- treated with SLF for the entire 14-day period (‘‘SLF:SLF’’),and that the final extents of pigmentation were similar.phology with a larger somata and more extensive cell pro-
cesses, compared with bFGF-treated cells (Table 1). This These results indicate that SLF probably does not act as asurvival factor for NC-derived melanocyte precursors, butdifference in morphology may indicate that SLF induces a
more complete stage of cytodifferentiation than does bFGF. instead acts to induce their differentiation into melano-cytes. However, these data do not exclude the possibilityIn any case, the finding that SLF increased the numbers of
pigmented cells in defined medium is consistent with those that low levels of endogenous SLF may be sufficient for thesurvival of melanocyte precursors. Such low levels of SLFof Lahav et al. (1994) using serum- and embryo extract-
containing culture medium. produced by NC cells during the initial 5 days may, in fact,explain why pigmented cells appeared in the untreated cul-To test whether this effect of SLF was due to the survival
of melanocyte precursors, highly enriched cultures of NC tures (Fig. 5A) and in the ø:SLF cultures only 1 day followinggrowth factor addition (Fig. 5 B). In any case, we also foundcells were grown in defined medium in the presence or
absence of SLF for 5 days, and then the medium was changed that NC cultures treated with SLF-neutralizing antibody
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8520 / 6x1e$$$181 03-20-97 21:06:54 dbal

65Expression of Steel Factor in Neural Crest Cells
TABLE 1Effects of SLF and bFGF on the Morphology and Differentiation of Cultured Neural Crest Cells
Proportion ofNumber of processes per cell Mean length of each process pigmented NC cells
Control cells 0.45 { 0.12 15.6 { 5.8 mm 0.3 { 0.7bFGF-treated cells 0.54 { 0.19 (ns) 18.2 { 2.6 mm (ns) 14.0 { 5.4 (P õ 0.001)SLF-treated cells 2.69 { 0.25 (P õ 0.02) 122.4 { 27.1 mm (P õ 0.02) 95.2 { 6.9 (P õ 0.001)
Note. Neural crest cells were cultured in serum-free and embryo extract-free CCM-5 culture medium for 14 days and then fixed andprocessed for microscopic examination. Morphometric analysis was performed on 300 cells in each of 3 control, 3 bFGF-treated, and 3SFL-treated cultures. The percentage of pigmented cells was determined by counting pigmented and unpigmented cells in 12 control, 12bFGF-treated, and 12 SLF-treated cultures. Statistical comparisons between the growth factor-treated cultures and the control cultureswere made using ANOVA tests. Note that SLF induced statistically significant differences in the proportions of pigmented cells, thenumbers of processes per cell, and the mean length of each process. ns, not significant (i.e., P ú 0.05).
during the entire 14-day period (i.e., ‘‘Ab:Ab’’) gave rise to time during early stages of NC cell emigration from theneural tube. In vivo, for example, we first observed SLF infewer pigmented cells than those in defined medium alone
(i.e., ‘‘ø:ø’’), indicating that the spontaneous pigmentation situ hybridization signal in the dorsal NC migratory spaceof the upper cervical spinal cord at E4, which is at least 24was due to the presence of SLF and not some other NC-
produced protein. hr after NC cell emigration has begun at this axial level(Bronner-Fraser, 1986). In culture, on the other hand, SLF-immunoreactive NC cells were observed as early as NC cellsfirst began to emigrate onto the culture surface at about 12DISCUSSION
In this paper, we show that NC cells express both the c-kit receptor and its ligand, the SLF growth factor. In vivo,SLF in situ hybridization signal was found in cells proximalto the neural tube at a development stage when NC cellsare known to be migrating in this location (Bronner-Fraser,1986). In culture, we found that NC cells express an mRNAwhose sequence is highly homologous to the published se-quence of the chicken and quail SLF gene products and thatapproximately 98% of the cells expressed SLF immunoreac-tivity. The onset of SLF expression is likely to occur some-
FIG. 3. Southern blot of an RT-PCR reaction using SLF primersindicating expression of a 466-base-pair SLF reaction product fromRNA extracted from cultured embryonic quail NC cells (lane 4).Positive control for the Southern blot involved electrophoresis ofa restriction digest of the SLF plasmid (lane 1). Positive controlsfor the RT-PCR involved substituting total quail brain RNA forFIG. 2. Protein immunoblot showing SLF-immunoreactivity in
various tissues dissected from E14 chickens. Note the single immu- NC RNA (lane 2). Negative controls involved omitting the reversetranscriptase steps in reactions with total brain RNA and NC RNAnoreactive band at 23 kDa (at the arrow). Lane 1, brain; lane 2,
heart; lane 3, kidney; lane 4, liver; lane 5, muscle. (lanes 3 and 5, respectively).
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8520 / 6x1e$$$181 03-20-97 21:06:54 dbal
66 Guo et al.
FIG. 4. In situ hybridization showing SLF mRNA in the NC ‘‘migration staging area’’ (Wehrle-Haller and Weston, 1995) of an E4 chicken.(A) Bright-field photomicrograph of a tissue section through the cervical region of a stage 22 embryo (Hamburger and Hamilton, 1951);(B) the same section viewed through Nomarski optics. Sense strand RNA served as negative controls in these studies (not shown). Notethe presence of SLF hybridization signal in the migration staging area adjacent to the dorsal neural tube (at the arrow), as well as somehybridization signal in the ventral ectoderm (at the asterisk). nt, neural tube; dm, dermamyotome; no, notocord.
hr. This small difference in the timing of appearance of SLF ods used to isolate NC cells, and we may have simply beenbelow the limits of detection. Second, it is possible that NCin NC cells in vivo and in culture can be explained either
by differences in the sensitivity of the methods used for cells do not process membrane-bound SLF to its solubleform under our culture conditions.detection or by the possibility that these culture conditions
somehow induce precocious SLF expression. Regardless of which form of SLF is actually present, thereare at least two nonmutually exclusive mechanisms byThe conclusion that SLF is actually being released by NC
cells is based, however, on more indirect data. We found, for which SLF could act—as a survival factor for a subpopula-tion of latent melanoblasts or as a factor influencing theexample, that SLF-neutralizing antibodies could partially
inhibit the spontaneous appearance of melanocytes in NC commitment of multipotent NC cells to a melanocyte fate.Our evidence, however, would seem to favor the latter pos-cells cultured in a serum-free and embryo extract-free me-
dium lacking exogenous SLF. This would suggest that at sibility. We found, for example, that depriving NC cells ofSLF during the initial 5 days in culture did not prevent theirleast some SLF is either being released by NC cells or at
least is being translocated to an extracellular compartment subsequent pigmentation induced by late additions of SLF.Presumably, this 5-day period is sufficiently long for cellaccessible to the neutralizing antibodies (i.e., is present on
the cell surface). On the other hand, attempts to directly death to have occurred (e.g., Greene, 1977), suggesting thatSLF is not necessary for survival, at least with cultureddemonstrate SLF immunoreactive material in Western blots
of concentrated medium from NC cell cultures have been avian NC cells. On the other hand, these data do not ruleout the possibility that committed melanoblasts becomeless successful (data not shown). There are, however, at least
two possible explanations for these negative results. First, dependent on SLF for their survival at later stages of devel-opment. This latter possibility is consistent with studiesthe numbers of NC cells used in these studies were neces-
sarily low due to the limitations of the microsurgical meth- performed using mouse NC cells. In these experiments, SLF
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8520 / 6x1e$$$181 03-20-97 21:06:54 dbal

67Expression of Steel Factor in Neural Crest Cells
FIG. 5. Effects of chicken SLF on melanogenesis (A) and survival (B) of cultured embryonic quail NC cells. (A) NC cells were culturedin CCM-5 defined medium in the presence or absence of SLF (100 ng/ml), basic fibroblast growth factor (25 ng/ml), SLF plus bFGF, orSLF plus SLF-neutralizing antiserum (1:100 dilution). Cultures were observed daily for the presence of pigmented cells, and the proportionsof cultures containing such cells were calculated. These experiments were performed three times with qualitatively similar results, andthese data represent the average of these studies. Note that SLF induced robust pigmentation (ú90%) by 8 days and that the presence ofSLF-neutralizing antibodies reduced this by more than half. The small decreases in the percentages of cultures containing pigmented cellsat later times are probably due to some cell death beginning around 8–10 days in culture. However, these decreases were more apparentin those cultures treated with bFGF, rather than SLF. (B) Medium-switch experiments in which NC cells were cultured in the presenceor absence of SLF or SLF-neutralizing antiserum for 5 days, and then the medium was changed for the remaining 9 days. Again, theseexperiments were performed three times with qualitatively similar results, and the results here represent the average of these studies.‘‘ø:ø’’ refers to cultures treated with defined medium for the entire 14 days; ‘‘ø:SLF’’ refers to cultures treated with defined medium alonefor 5 days followed by 9 days in SLF-containing medium; ‘‘SLF:SLF’’ refers to cultures treated with SLF for the entire 14 days; ‘‘Ab:Ab’’refers to cultures treated with antibody for the entire 14 days. (C and D) Morphology of NC-derived cells cultured for 14 days in thepresence of 100 ng/ml SLF (C) or 25 ng/ml bFGF (D). Note that SLF induced a more mature melanocyte morphology in NC cells than didbFGF, with cells displaying significantly longer processes. The bar in the upper right corner of D corresponds to 10 mm.
was found to increase the numbers of c-kit positive cells in plex culture medium containing serum, embryo extract,and phorbol esters, complicating the interpretation ofNC cultures, suggesting that SLF was acting as either a
survival or mitogenic signal (Murphy et al., 1992). It should which effects of SLF were direct and which involved combi-nations of factors. It is also possible, however, that the NCbe noted, however, that these studies involved the use com-
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8520 / 6x1e$$$181 03-20-97 21:06:54 dbal

68 Guo et al.
cells of mice and birds utilize SLF in different ways for staging area specifies a melanogenic fate for those cells thatsubsequently enter this pathway.inducing the appearance of melanocytes. Perhaps mice do
In contrast to the dorsal migratory pathway, NC cellsindeed require SLF as a survival factor (and perhaps a differ-that migrate along the ventral pathway soon become mixedentiation factor as well), whereas birds utilize SLF at anwith non-NC cells in a loose mesenchyme (Bronner-Fraser,earlier step in the process of melanogenesis to influence cell1986), which may not be permissive for melanogenesis. If,fate decisions among pluripotent cells.for example, SLF requires a relatively high concentrationIn any case, the possibility that SLF acts to influenceand/or direct contact between responding cells, then thesuch cell fate decisions is also consistent with earlier obser-mixed mesenchyme of the ventral pathway would not bevations by Weston and colleagues. They showed that theexpected to produce melanocytes. In addition, it is possibleability of cultured avian NC cells to give rise to melano-that there are other factors acting in a redundant fashion tocytes depended on the period of time in which the cellsinhibit ectopic melanogenesis along the ventral NC migra-were in close physical apposition to each other (Glimeliustory pathway. One such factor may be transforming growthand Weston, 1981; Vogel and Weston, 1988). Whereas NCfactor type-b (TGFb), which has been shown to inhibit mel-cells that formed tight clusters for periods of at least 36 hranogenesis by NC cells in response to other peptide growthgave rise to numerous pigmented cells, NC cells that werefactors, as well as factors present in the substratum (Stockereither prevented from forming clusters or in which the clus-et al., 1991; Rogers et al., 1992). Various TGFb isoformsters were dissociated within the 36-hr period contained sig-have been shown to be expressed, moreover, by cells adja-nificantly fewer melanocytes. Although these studies didcent to the ventral NC migratory space of chicken embryosnot identify the factor(s) involved, they are consistent withat the appropriate developmental stages in vivo (Jakolew etthe view that some endogenous factor influences NC cellal., 1992, 1994). It is interesting to note, therefore, thatfate, possibly including SLF. Interestingly, these effects ofcranial neural crest cells, which undergo melanogenesis toprolonged association had the opposite effect on the appear-a far lesser extent than those of the trunk (Leblanc et al.,ance of NC-derived cells expressing a neuronal marker (Vo-1995), have been shown to express and process TGFbgel and Weston, 1988). This suggests that the endogenous(Brauer and Yee, 1993).factor or factors may actually change the fate of multipo-
In any case, the observations that NC cells express growthtent NC cells from a neuronal to a melanocyte fate. Al-factors that are capable of influencing their own develop-though our data are consistent with the possibility thatment would suggest that NC cells play a more active rolethis endogenous factor is SLF, it remains to be determinedin the determination of their cell fate than has previouslydirectly whether SLF actually suppresses neurogenesis inbeen hypothesized.multipotent NC cells.
If the SLF/c-kit axis is indeed involved in melanogenesis,then our finding that most NC cells express both the SLF ACKNOWLEDGMENTSand c-kit receptor proteins in culture might have predictedthat melanocytes should comprise the largest population of
We thank Dr. A. Johnson (Rutger’s University, New Brunswick,NC-derived cells. This is, however, not the case in vivo. NJ), Dr. J. Carnahan (Amgen, Inc., Thousand Oaks, CA), and Dr.Lineage-tracing studies, in which individual NC cells were M. Sakurai (National Institute of Animal Health, Japan) for theirinjected in vivo with a marker and then the fates of their generous gifts of chicken SLF cDNA, recombinant chicken SLFprogeny monitored, clearly show that only a small subpopu- protein, and chicken c-kit cDNA, respectively. We also thank Drs.
Gabrielle Leblanc and Steven Matsumoto for their helpful com-lation of NC cells actually give rise to melanocytes (Fraserments on the manuscript. This work was supported by a grant fromand Bronner-Fraser, 1991). Together with the studies re-the NIH to G.C.ported here, this may indicate that negative factors or condi-
tions play a major role in the determination of cell fate.One likely condition acting in such a negative fashion may
REFERENCESbe the extent to which NC cells are dispersed during earlystages of their migration in vivo. NC cells that form melano- Anderson, D. J. (1989). The neural crest cell lineage problem: Neu-cytes of the skin, for example, are initially tightly clustered ropoiesis? Neuron 3, 1–12.in a space adjacent to the neural tube for a period of about Brauer, P. R., and Yee, J. A. (1993). Cranial neural crest cells synthe-36 hr, until the space between the dermamyotome and ecto- size and secrete a latent form of transforming growth factor b
that can be activated by neural crest cell proteolysis. Dev. Biol.derm opens up allowing dispersion of these cells. This clus-155, 281–285.tering of NC cells has been termed the ‘‘migration staging
Bronner-Fraser, M. (1986). Analysis of the early stages of trunkarea’’ and has been suggested to play a critical role in theneural crest migration in avian embryos using monoclonal anti-determination of melanogenesis by NC cells (Weston,body HNK-1. Dev. Biol. 115, 44–55.1991). Consistent with this notion are observations that
Erickson, C. A., and Goins, T. L. (1995). Avian neural crest cellsNC cells migrating along the dorsal–lateral pathway seem can migrate in the dorsolateral path only if they are specified asalready to be committed to a melanocyte fate (Erickson and melanocytes. Development 121, 915–924.Goins, 1995). Perhaps the relatively high concentrations of Erickson, C. A., Duong, T. D., and Tosney, K. W. (1992). Descrip-
tive and experimental analysis of dispersion of neural crest cellsSLF expressed by neighboring NC cells in the migration
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8520 / 6x1e$$$181 03-20-97 21:06:54 dbal

69Expression of Steel Factor in Neural Crest Cells
along the dorsolateral path and their entry into ectoderm in the Matsui, Y., Zsebo, K. M., and Hogan, B. L. M. (1990). Embryonicexpression of a haematopoietic growth factor encoded by the Slchicken embryo. Dev. Biol. 151, 251–272.locus and the ligand for c-kit. Nature 347, 667–669.Flanagan, J. G., Chan, D. C., and Leder, P. (1991). Transmembrane
Morrison-Graham, K., and Weston, J. A. (1993). Transient steel fac-form of the kit ligand growth factor is determined by alternativetor dependence by neural crest-derived melanocyte precursors.splicing and is missing in the Sld mutant. Cell 64, 1025–1035.Dev. Biol. 159, 346–352.Fraser, S. E., and Bronner-Fraser, M. (1991). Migrating neural crest
Morrison-Graham, K., Schatteman, G. C., Bork, T., Bowen-Pope,cells in the trunk of the avian embryo are multipotent. Develop-D. F., and Weston, J. A. (1992). A PDGF receptor mutation in thement 112, 913–920.mouse (Patch) perturbs the development of a non-neuronal subsetGalli, S. J., Zsebo, K. M., and Geissler, E. N. (1994). The kit ligand,of neural crest-derived cells. Development 115, 133–142.stem cell factor. Adv. Immunol. 55, 1–96.
Murphy, M., Reid, K., Williams, D. E., Lyman, S. D., and Bartlett,Glimelius, B., and Weston, J. A. (1981). Analysis of developmentallyP. F. (1992). Steel factor is required for maintenance, but nothomogeneous neural crest cell populations in vitro. III. Role ofdifferentiation, of melanocyte precursors in the neural crest. Dev.culture environment in cluster formation and differentiation.Biol. 153, 396–401.Cell Differ. 10, 57–67.
Nishikawa, S., Kusakabe, M., Yoshinaga, K., Ogawa, M., Hayashi,Greene, L. A. (1977). Quantitative in vitro studies on the nerveS., Kunisade, T., Era, T., Sakakura, T., and Nishikawa, S. (1991).growth factor (NGF) requirement of neurons. I. Sympathetic neu-In utero manipulation of coat color formation by monoclonalrons. Dev. Biol. 58, 96–105.anti-c-kit antibody: Two distinct waves of c-kit-dependency dur-Gruneberg, H., and Truslove, G. M. (1960). Two closely linkeding melanocyte development. EMBO J. 10, 2111–2118.genes in the mouse. Genet. Res. 1, 69–90.
Pinco, O., Carmeli, C., Rosenthal, A., and Kalcheim, C. (1993).Hamburger, V., and Hamilton, J. L. (1951). A series of normal stagesNeurotrophin-3 affects proliferation and differentiation of dis-in the development of the chick embryo. J. Morphol. 88, 49–92.tinct neural crest cells and is present in the early neural tube ofJakowlew, S. B., Ciment, G., Tuan, R. S., Sporn, M. B., and Roberts,avian embryos. J. Neurobiol. 24, 1626–1641.
A. B. (1992). Pattern of expression of transforming growth factor-Reid, K., Nishikawa, S.-I., Bartlett, P. F., and Murphy, M. (1995).
b4 mRNA and protein in the developing chicken embryo. Dev.Steel factor directs melanocyte development in vitro through
Dyn. 195, 276–289. selective regulation of the number of c-kit/ progenitors. Dev.Jakowlew, S. B., Ciment, G., Tuan, R. S., Sporn, M. B., and Roberts, Biol. 169, 568–579.
A. B. (1994). Expression of transforming growth factor-b2 and b3 Rogers, S. L., Gegick, P. J., Alexander, S. M., and McGuire, P. G.mRNAs and proteins in the developing chicken embryo. Differ- (1992). Transforming growth factor-b alters differentiation in cul-entiation 55, 105–118. tures of avian neural crest-derived cells: Effects on cell morphol-
Kawasaki, E. S. (1990). Amplification of RNA. In ‘‘PCR Protocols’’ ogy, proliferation, fibronectin expression, and melanogenesis.(M. A. Innis, D. H. Gelfand, J. J. Sninsky, and T. J. White, Eds.), Dev. Biol. 151, 192–203.pp. 21–27. Academic Press, San Diego. Sieber-Blum, M. (1991). Role of the neurotrophic factors BDNF and
Keshet, E., Lyman, S. D., Williams, D. E., Anderson, D. M., Jenkins, NGF in the commitment of pluripotent neural crest cells. Neu-N. A., Copeland, N. G., and Parada, L. F. (1991). Embryonic RNA ron 6, 949–955.expression patterns of the c-kit receptor and its cognate ligand Stocker, K. M., Sherman, L., Rees, S., and Ciment, G. (1991). Basicsuggest multiple functional roles in mouse development. EMBO FGF and TGF-b1 influence commitment to melanogenesis inJ. 10, 2425–2435. neural crest-derived cells of avian embryos. Development 111,
Lahav, R., Lecoin, L., Ziller, C., Nataf, V., Carnahan, J. F., Martin, 635–645.F. H., and Le Douarin, N. M. (1994). Effect of the steel gene prod- Vogel, K. S., and Weston, J. A. (1988). A subpopulation of cultureduct on melanogenesis in avian neural crest cell cultures. Differen- avian neural crest cells has transient neurogenic potential. Neu-tiation 58, 133–139. ron 1, 569–577.
Le Douarin, N. M. (1982). ‘‘The Neural Crest.’’ Cambridge Univ. Wehrle-Haller, B., and Weston, J. A. (1995). Soluble and cell-boundforms of steel factor activity play distinct roles in melanocytePress, Cambridge, UK.precursor dispersal and survival on the lateral neural crest migra-Leblanc, G. G., and Bronner-Fraser, M. E. (1992). Neural crest celltion pathway. Development 121, 731–742.differentiation. In ‘‘Development, Regeneration, and Plasticity
Weston, J. A. (1991). Sequential segregation and fate of developmen-of the Autonomic Nervous System’’ (I. A. Hendry and C. E. Hill,tally restricted intermediate cell populations in the neural crestEds.), pp. 95–137. Harwood Academic, Chur, Switzerland.lineage. Curr. Top. Dev. Biol. 25, 133–153.Leblanc, G. G., Holbert, T. E., and Darland, T. (1995). Role of the
Weston, J. A., Derby, M. A., and Pintar, J. E. (1978). Changes intransforming growth factor-beta family in the expression of cra-the extracellular environment of neural crest cells during theirnial neural crest-specific phenotypes. J. Neurobiol. 26, 497–510.migration. Zoon 6, 103–113.Lecoin, L., Lahav, R., Martin, F. H., Teillet, M.-A., and Le Douarin,
Williams, D. E., de Vries, P., Namen, A. E., Widmer, M. B., andN. M. (1995). Steel and c-kit in the development of avian melano-Lyman, S. D. (1992). The Steel factor. Dev. Biol. 151, 368–376.cytes: A study of normally pigmented birds and of the hyperpig-
Zhou, J.-H., Ohtaki, M., and Sakurai, M. (1993). Sequence of amented mutant Silky Fowl. Dev. Dyn. 203, 106–118.cDNA encoding chicken stem cell factor. Gene 127, 269–270.Loring, J., Glimelius, B., and Weston, J. A. (1982). Extracellular ma-
trix materials influence quail neural crest cell differentiation in Received for publication October 2, 1996Accepted January 17, 1997vitro. Dev. Biol. 90, 165–174.
Copyright q 1997 by Academic Press. All rights of reproduction in any form reserved.
AID DB 8520 / 6x1e$$$181 03-20-97 21:06:54 dbal





























