Embed Size (px)
Citation preview

S
tnnmftccqsctssngtotc
a(hm
d
Molecular and Cellular Neuroscience 17, 539–550 (2001)
doi:10.1006/mcne.2000.0946, available online at http://www.idealibrary.com on MCN
A
Autocrine Regulation of NorepinephrineTransporter Expression
Z. G. Ren,* P. Porzgen,† J. M. Zhang,* X. R. Chen,*. G. Amara,†,‡ R. D. Blakely,§ and M. Sieber-Blum* ,1
*Department of Cell Biology, Neurobiology and Anatomy, Medical College of Wisconsin,Milwaukee, Wisconsin 53226; †Vollum Institute and ‡Howard Hughes Medical Institute,Oregon Health Sciences University, Portland, Oregon 97201; and §Department ofPharmacology, Vanderbilt University, Nashville, Tennessee 37232
pnbno
The1 norepinephrine transporter (NET) is a neurotransmit-er scavenger and site of drug action in noradrenergiceurons. The aim of this study was to identify mecha-isms that regulate NET expression during the develop-ent of quail (q) sympathetic neuroblasts, which develop
rom neural crest stem cells. Neurotrophin-3 (NT-3) andransforming growth factor b1 (TGF-b1) cause an in-rease of qNET mRNA levels in neural crest cells. Whenombined, the growth factors are additive in increasingNET mRNA levels. Both NT-3 and TGF-b1 are synthe-ized by neural crest cells. Onset of NET expression pre-edes the onset of neural crest stem cell emigration fromhe neural tube. In older embryos, qNET is expressed byeveral crest-derived and noncrest tissues. The datahow that qNET expression in presumptive sympatheticeurons is initiated early in embryonic development byrowth factors that are produced by neural crest cellshemselves. Moreover, the results support our previousbservations that norepinephrine transport contributes tohe regulation of the differentiation of neural crest stemells into sympathetic neurons.
INTRODUCTION
In adult neurons, NET serves to remove excess trans-mitter from the synaptic cleft (Axelrod, 1965; Iversen,1967; Snyder, 1970) and is a site of action of drugs thatblock norepinephrine transport, such as tricyclic anti-depressants and cocaine, (Axelrod et al., 1961; Whitby etl., 1960). The human (Pacholczyk et al., 1991), mouseFritz et al., 1998), and rat (Bruss et al., 1997) NET genesave been cloned and characterized. In the adult mam-alian organism, the same NET gene is expressed in the
1 To whom correspondence and reprint requests should be ad-ressed. Fax: 414-456-6517. E-mail: [email protected].
1044-7431/01 $35.00Copyright © 2001 by Academic Press
ll rights of reproduction in any form reserved.
eripheral and central nervous systems in sympatheticeurons, the adrenal gland, and the locus ceruleus in therain stem (Amara, 1995). In developing noradrenergiceurons, norepinephrine transport leads to the expressionf tyrosine hydroxylase and dopamine-b-hydroxylase in
differentiating neural crest cells, suggesting that NET isinvolved in promoting the differentiation of neural creststem cells into sympathetic neurons (Sieber-Blum, 1989;Zhang and Sieber-Blum, 1992; Zhang et al., 1997a). Afterleaving the dorsal aspect of the neural tube, presumptivenoradrenergic neural crest cells migrate dorsoventrallybetween the somite and the neural tube and past thenotochord. Ultimately they settle near the dorsal aorta,where they start to synthesize catecholamines and differ-entiate into the primary sympathetic ganglia.
Despite the importance of NET function, the mecha-nisms that regulate NET expression, the time course ofNET expression, and the NET expression patterns in theembryo are not well defined. We have cloned a partialqNET cDNA and used it to determine the function ofcandidate growth factors in NET expression. Our datashow that NT-3 and TGF-b1 cause an increase in NETmRNA levels in an additive manner. Both growth fac-tors are expressed by migratory neural crest cells, thusindicating that they have autocrine or paracrine func-tion. Moreover, NET is expressed in a number of neuralcrest-derived and noncrest embryonic tissues of bothectodermal and mesodermal origin.
RESULTS
Cloning of a Partial Quail NET cDNA
A partial qNET cDNA was cloned from RNA ofcultured quail neural crest cells (Genbank Accession
539

FIG
.1.
Sequ
ence
ofpa
rtia
lqN
ET
clon
e.D
educ
edam
ino
acid
sequ
ence
and
clus
tal
alig
nw
ith
hum
anN
ET
(hN
ET
),m
ouse
NE
T(m
NE
T),
hum
and
opam
ine
tran
spor
ter
(hD
AT
),an
dhu
man
sero
toni
ntr
ansp
orte
r(h
SER
T).
Sequ
ence
mar
ked
blue
,ext
race
llula
rlo
op.S
eque
nce
mar
ked
red
(W12
4–T
373)
was
used
for
insi
tuhy
brid
izat
ion.
Sequ
ence
mar
ked
wit
hye
llow
was
used
asan
imm
unog
ento
rais
ean
tibo
dy
4340
8.
540 Ren et al.

fscTrvcGiepa
s
NtnsT
pwCp
541Autocrine Regulation of NET Expression
No. AF230787). The cloned fragment encodes 515amino acids (aa) and shows 88% aa identity to humanNET (hNET), 88% to mouse NET (mNET), 71% to thehuman dopamine transporter (hDAT), and 52% to thehuman serotonin transporter (hSERT; Fig. 1). The mod-erately conserved large extracellular loop domain of thetransporters shows a high degree of identity between
FIG. 2. RT-PCR amplification of qNET, TGF-b, and NT-3 mRNArom 24-h neural crest cell cultures. (A) qNET RT-PCR. Each panelhows five independent experiments, in which 18–22 neural crest cellultures served as a starting material for the RT-PCR experiments.he band densities were quantified by fluorimaging, expressed asatio NET:GAPDH, and statistically analyzed by 1-factor analysis ofariance. Top panel: In this control the cells were grown in definedulture medium in the absence of added growth factors. Upper bands,APDH; lower bands, qNET. A qNET band was observed, suggest-
ng either an autocrine mechanism of qNET expression or qNETxpression prior to the onset of neural crest cell migration. Secondanel: Cells were grown in defined culture medium in the presence ofnti-NT-3 blocking antibodies (4 mg/ml) and anti-pan TGF-b block-
ing antibodies (4 mg/ml). The qNET band is significantly reduced,uggesting that neural crest cells synthesize NT-3 and TGF-b and that
the growth factors increase qNET mRNA levels by an autocrinemechanism. Third panel: Cells were grown in the presence of FGF-2(2.5 ng/ml) and NT-3 (10 ng/ml). The qNET band is significantlyincreased compared to the control without growth factors and thecontrol with blocking antibodies. Fourth panel: When cells weregrown in the presence of TGF-b1 (1 ng/ml), the qNET band wasincreased. Fifth panel: When the cells were grown in the presence ofall three factors, FGF-2, NT-3, and TGF-b1, the band intensity wasfurther increased, indicating an additive mechanism. All values weresignificantly different from each other (P # 0.006). (B) TGF-b and
T-3 RT-PCR. RNA was isolated from 24-h neural crest cell cultureshat had been grown in defined culture medium in the absence ofoncrest cells and in the absence of added growth factors. The datauggest that early migratory neural crest cells synthesize all threeGF-bs, as well as NT-3.
the qNET and the mammalian NETs, but significantdifferences to hDAT and hSERT, strongly suggesting
sa
that the cloned transporter fragment is part of the quailNET gene. This notion is further supported by a posi-tive in situ hybridization signal in the locus ceruleus(data not shown).
Increase in NET mRNA Levels in the Presence ofNT-3 and TGF-b1
Neural crest cell explants were grown in a chemicallydefined culture medium (Sieber-Blum and Chokshi,1985; Sieber-Blum, 1999). RNA from 24-h cultures thatwere grown in the presence and absence of growthfactors was isolated and amplified by RT-PCR withqNET-specific and GAPDH-specific primers. In contrast
FIG. 3. qNET in situ hybridization of cultured neural crest cells. (a)Cells were grown for in the presence of FGF-2 and NT-3 for 48 h (h0–48) to promote proliferation. At 24 h, TGF-b1, which inhibits cell
roliferation, was added as well (h 24–48). At 48 h of culture, the cellsere processed for in situ hybridization with anti-sense probe. (b)ells were grown in the presence of NT-3 blocking antibodies andan-TGF-b blocking antibodies and subsequently hybridized with
anti-sense probe. (c) Control in which cells were grown in the pres-ence of FGF-2, NT-3, and TGF-b1 and hybridization was done with
ense probe. The hybridization signal is significantly reduced in (b)nd (c). Bar, 25 mm.
aqFoambaepw
ieetcp
ilsfwi
542 Ren et al.
to expectation, a qNET band was present in RNA fromcontrol neural crest cell cultures that were grown in theabsence of added growth factors (Fig. 2A; top panel).This indicated either expression of qNET prior to onsetof migration of neural crest cells or autocrine regulationof NET expression. To investigate autocrine regulation,neural crest cells were grown in the presence of anti-NT-3 and anti-pan TGF-b blocking antibodies. The vi-bility of the cells was not affected (not shown), but theNET band was significantly decreased (P 5 0.0001;ig. 2A, second panel), suggesting autocrine regulationf NET expression. Both the FGF-2/NT-3 combinationnd TGF-b1 caused a significant increase in NETRNA levels over the antibody control (P 5 0.0001 for
oth; Fig. 2A, panels 3 and 4). FGF-2 by itself does notffect NE transport but potentiates NT-3 action (Zhangt al., 1997a, b). When the cells were grown in theresence of all three growth factors, NET mRNA levelsere elevated significantly (P 5 0.0001; Fig. 2A, bot-
tom panel) compared to FGF-2 1 NT-3 and to TGF-b1,indicating that FGF-2/NT-3 and TGF-b1 synergize inncreasing NET mRNA levels. Data from all control andxperimental groups were significantly different fromach other (P # 0.006). Equivalent results were ob-ained by in situ hybridization of cultured neural crestells with a NET probe (Fig. 3). Cells were grown in theresence of FGF-2, NT-3, and TGF-b1 (Figs. 3a and 3c)
or in the presence of NT-3 blocking antibodies and panTGF-b blocking antibodies (Fig. 3b) and processed forn situ hybridization. The relative intensity of the alka-ine phosphatase reaction product was high with anti-ense probe in cells grown in the presence of the growthactors (141.3 6 5.7; Fig. 3a), but significantly lower
hen the cells were grown in the presence of the block-ng antibodies (63.3 6 2.5; P , 0.001; Fig. 3b), compa-
rable to the staining intensity with sense probe (nega-tive control; 57.5 6 1.7; P , 0.001; Fig. 3c).
The combination of FGF-2 and NT-3 increases NETfunction as measured by high-affinity uptake of triti-ated NE and subsequent autoradiography in neuralcrest cell colony assays (Zhang et al., 1997a; Sieber-Blum, 1999). Here we show by the same method thatTGF-b1 also promotes NE transport activity and thatthe FGF-2/NT-3 plus TGF-b1 combination is additive(Table 1). In these experiments the cells were grown inthe presence of 4% horse serum and 2.5% chick embryoextract. It is thus possible that under those culture con-ditions additional mechanisms, such as activation of the
transporter, might operate in addition to the NT-3 andTGF-b-mediated increase of NET expression.NT-3 and TGF-b Expression in Neural Crest Cellsand Noncrest Cells in Vitro and in Vivo
The idea of autocrine growth factor production wasconfirmed at the mRNA and protein levels in neuralcrest cell cultures and in the embryo. RNA isolatedfrom 24 h neural crest cell cultures that were grown indefined culture medium was amplified by RT-PCR withgrowth factor-specific primer pairs. RT-PCR productswere obtained for NT-3, TGF-b1, TGF-b2, and TGF-b3(Fig. 2B).
Virtually all neural crest cells in culture and in the3.5-day-old quail embryo were strongly immunoreac-tive for NT-3 as detected by confocal microscopy (Fig.4). During advanced development, the neural tube (seealso Pinco et al., 1993), the dorsal root ganglion, theventral root, the sympathetic ganglion (see also Zhanget al., 1994), the dorsal aorta, and the myotome (station-ary somitic cells adjacent to the neural tube and migra-tory somitic cells on course to the limb bud) wereintensely NT-3 immunoreactive (Fig. 4f). Figure 4gshows HNK-1 stain in the same section as in f. HNK-1immunoreactivity is characteristic for migrating avianneural crest cells. This particular HNK-1 hybridomaclone (VC1.1) recognizes N-CAM and other cell-adhe-sion molecules and therefore also binds to the neuraltube and the notochord (Zaremba et al., 1990). Virtuallyall neural crest cells also expressed TGF-b both in vitro(see also Brauer and Yee, 1993) and in vivo, as deter-mined with anti-pan TGF-b antibodies (Fig. 5). Figure5a shows TGF-b immunoreactivity in cultured neuralcrest cells. Specificity of antibody binding is shown inthe controls in Fig. 5b (no primary antibody) and Fig. 5c(preabsorbed primary antibody). Dorsoventrally mi-grating neural crest cells in the embryo were immuno-reactive for TGF-b (Fig. 5), as were the dorsal rootganglion, neural tube, ventral root, sympathetic gan-glion, and the myotome cells (cells in somites adjacentto the neural tube and cells migrating into the limb bud;Fig. 5d). Figure 5e shows HNK-1 binding in the samesection as in Fig. 5d. Figure 5f shows a higher magni-fication of the area marked in Fig. 5a, in which TGF-bimmunoreactive migrating neural crest cells and cells inthe nascent dorsal root ganglion are visible.
NET Expression in the Neural Crest and in NeuralCrest-Derivatives
Since NET mRNA is expressed early in neural crestdevelopment (Figs. 2 and 3), we determined by confocal
microscopy the time course of qNET protein expres-sion. In culture, some neural crest cells expressed NET
1fIT
CNT
hfcd
543Autocrine Regulation of NET Expression
at high levels already 6 h after explantation of theneural tube, the earliest time point postexplantation atwhich immunocytochemistry can be performed (Figs.6a and 6b). Staining appeared to be predominantlyintracellular. In 4-day-old cultures that had been grownin the presence of FGF-2 and NT-3, NET was expressedby neuronal cells (Figs. 6c and 6d). Immunoreactivitywas stronger in neurites than in cell bodies.
Similar to cultured neural crest cells, some neuralcrest cells in vivo expressed NET already at the onset oftheir emigration from the neural tube. Figures 6e–6hshows a cross section through the trunk region of a2.5-day-old embryo. Virtually all cells that form theleading edge of migratory neural crest cells are NETimmunoreactive (Fig. 6e, NET fluoresceine fluores-cence; Fig. 6f, HNK-1 rhodamine fluorescence). Thissubset of neural crest cells consists primarily of stemcells (Sieber-Blum, 1989b). Interestingly, the dorsal as-pect of the neural tube, from which neural crest cellsdelaminate, is intensely immunoreactive (Figs. 6e and6g), suggesting that premigratory neural crest cells arealready NET-immunoreactive. In addition, myotomecells are NET-positive (Figs. 6e and 6g). In Figs. 6g and6h, a higher magnification of part of Fig. 6e and 6f isshown. Two neural crest cells that express both NETand HNK-1 are marked by arrows.
In 3.5-day-old embryos (Figs. 6i and 6k), NET wasexpressed by several neural crest-derived tissues, in-cluding the sympathetic ganglion, as expected, but alsothe dorsal root ganglion (drg), and putative Schwanncell precursors in the ventral root (vr). In addition,several noncrest tissues were NET-immunoreactive.The entire cross-section of the neural tube (nt) shows
TABLE 1
Increase in Norepinephrine Transport Activity in the Presence ofNT-3 and TGF-b
Experimental condition
Number ofcolonies per platecontaining grain-
positive cells
Total numberof coloniesper plate
ontrol (no growth factors) 2.0 6 0.4 32.0 6 1.4T-3 4.5 6 0.6* 34.3 6 1.2GF-b1 4.3 6 0.7* 29.8 6 0.6
NT-3 plus TGF-b1 7.0 6 0.7* 35.4 6 1.5
Note. Cells in colony assay were cultured in the presence of 4%orse serum and 2.5% chick embryo extract. The data are averagesrom colony counts from 10 plates in each group. The total number ofolonies per plate was unchanged (P 5 0.38–0.76). *Significantlyifferent from the control, P , 0.01.
variable intensities of NET immunoreactivity. More-over, the myotome (mt) and the lining of the dorsal
aorta (da) are NET-immunoreactive. Controls for im-munohistochemistry included omitting one of the twofirst antibodies and indicated that immunofluorescenewas NET-specific and HNK-1-specific, respectively(data not shown).
DISCUSSION
The main observation in this report is that both theFGF-2/NT-3 combination and TGF-b1 regulate expres-sion of the norepinephrine transporter, as indicated byan increase in steady state qNET mRNA levels. Theiraction is additive. Moreover, the growth factors aresynthesized by neural crest cells, the precursors of sym-pathetic neurons. Therefore they are likely to act asneural crest-autocrine factors. Since the early migratoryneural crest already is a heterogeneous population con-sisting of pluripotent stem cells, committed cells andcells with restricted developmental potentials (Sieber-Blum and Sieber, 1984), it remains to be determinedwhether the neural crest cells that express NET alsoexpress the three growth factors, or whether some or allfactors are supplied by neighboring cells in a paracrinefashion. Moreover, it is possible that the mechanism ofgrowth factor-mediated increase in qNET mRNA levelsis indirect, insofar as NT-3 and/or TGF-b1 could induceanother factor, which in turn promotes NET expression.NT-3 and TGF-b1 activate independent signaling path-ways, which may, however, converge as they sharedownstream mediators. For example, MAP kinase ki-nase-1 (MEKK-1), which is a member of a neurotrophin-activated pathway, can activate Smad2, a signaling me-diator downstream of TGF-b (reviewed by Zhang andDerynck, 1999). Alternatively it is conceivable thatTGF-b promotes expression of NT-3. This possibility,however, is unlikely for two reasons. One, in both neu-rons (Buchman et al., 1994) and Schwann cells (Cai et al.,1999), TGF-b inhibits expression of NT-3. Two, we haveshown that the 10 ng/ml concentration of NT-3 thatwas used in this study elicits a maximal response incultured neural crest cells (Zhang et al., 1997a).
Neural crest cells express the NT-3 receptor, TrkC,when they leave the neural tube (Tessarollo et al., 1993;Henion et al., 1995) and require NT-3 (Farinas et al.,994). NT-3 is unique among neurotrophins of the NGFamily in promoting NE transport (Zhang et al., 1997a).n contrast, it remains to be determined whetherGF-b1 is the sole active physiological member of this
family of growth factors, in particular since TGF-b2 and
TGF-b3 are also produced by neural crest cells. In ad-dition to promoting NET expression, the three growth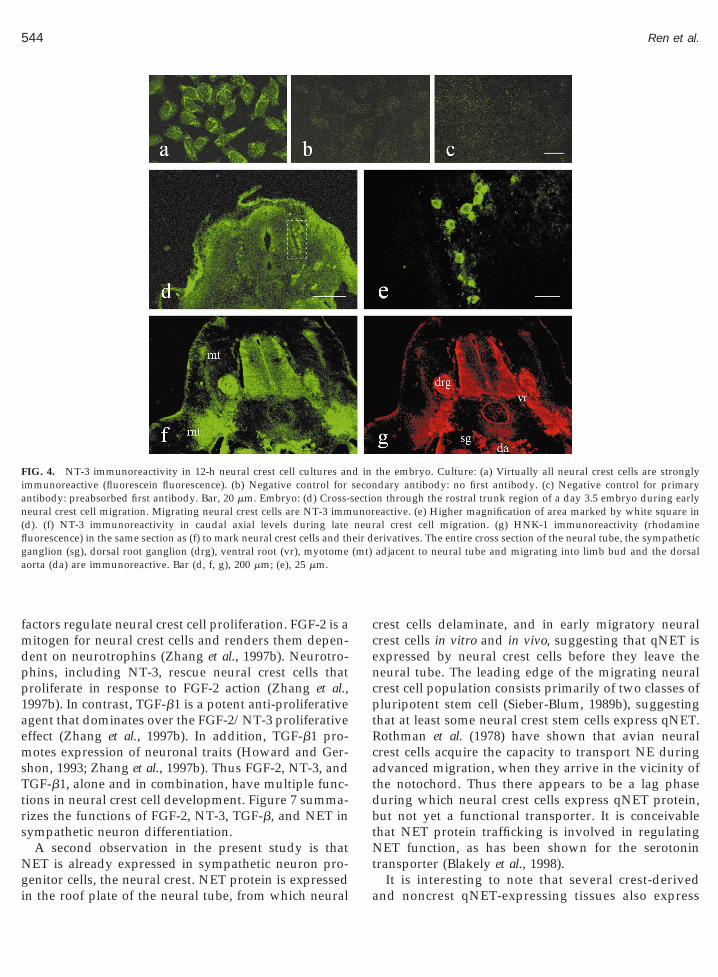
ae
n(flg (mt)a
544 Ren et al.
factors regulate neural crest cell proliferation. FGF-2 is amitogen for neural crest cells and renders them depen-dent on neurotrophins (Zhang et al., 1997b). Neurotro-phins, including NT-3, rescue neural crest cells thatproliferate in response to FGF-2 action (Zhang et al.,1997b). In contrast, TGF-b1 is a potent anti-proliferativegent that dominates over the FGF-2/NT-3 proliferativeffect (Zhang et al., 1997b). In addition, TGF-b1 pro-
motes expression of neuronal traits (Howard and Ger-shon, 1993; Zhang et al., 1997b). Thus FGF-2, NT-3, andTGF-b1, alone and in combination, have multiple func-tions in neural crest cell development. Figure 7 summa-rizes the functions of FGF-2, NT-3, TGF-b, and NET insympathetic neuron differentiation.
A second observation in the present study is thatNET is already expressed in sympathetic neuron pro-
FIG. 4. NT-3 immunoreactivity in 12-h neural crest cell cultures animmunoreactive (fluorescein fluorescence). (b) Negative control forantibody: preabsorbed first antibody. Bar, 20 mm. Embryo: (d) Cross-
eural crest cell migration. Migrating neural crest cells are NT-3 immd). (f) NT-3 immunoreactivity in caudal axial levels during lateuorescence) in the same section as (f) to mark neural crest cells and thanglion (sg), dorsal root ganglion (drg), ventral root (vr), myotomeorta (da) are immunoreactive. Bar (d, f, g), 200 mm; (e), 25 mm.
genitor cells, the neural crest. NET protein is expressedin the roof plate of the neural tube, from which neural
crest cells delaminate, and in early migratory neuralcrest cells in vitro and in vivo, suggesting that qNET isexpressed by neural crest cells before they leave theneural tube. The leading edge of the migrating neuralcrest cell population consists primarily of two classes ofpluripotent stem cell (Sieber-Blum, 1989b), suggestingthat at least some neural crest stem cells express qNET.Rothman et al. (1978) have shown that avian neuralcrest cells acquire the capacity to transport NE duringadvanced migration, when they arrive in the vicinity ofthe notochord. Thus there appears to be a lag phaseduring which neural crest cells express qNET protein,but not yet a functional transporter. It is conceivablethat NET protein trafficking is involved in regulatingNET function, as has been shown for the serotonintransporter (Blakely et al., 1998).
the embryo. Culture: (a) Virtually all neural crest cells are stronglydary antibody: no first antibody. (c) Negative control for primaryn through the rostral trunk region of a day 3.5 embryo during earlyeactive. (e) Higher magnification of area marked by white square inal crest cell migration. (g) HNK-1 immunoreactivity (rhodamineerivatives. The entire cross section of the neural tube, the sympatheticadjacent to neural tube and migrating into limb bud and the dorsal
d inseconsectiounorneureir d
It is interesting to note that several crest-derivedand noncrest qNET-expressing tissues also express

gtcsscststae
CAstwaG
pr me (e tives.m nglio
545Autocrine Regulation of NET Expression
both NT-3 and TGF-b. They include the dorsal rootanglion, the sympathetic ganglion, the ventral root,he neural tube, and the somitic myotome. It is thusonceivable that autocrine regulation of NET expres-ion is not limited to the neural crest, but occurs ineveral or all NET-expressing tissues. The signifi-ance of qNET expression in tissues other than theympathetic ganglion remains to be determined. Inhis context it is interesting, however, that both sen-ory neuroblasts in the dorsal root ganglion and en-eric neuroblasts express NET and become transientlydrenergic during embryonic development (Gershont al., 1984; Xue et al., 1985).
In summary, our data indicate that the quail NET ishighly homologous to mammalian NETs and that itsexpression is mediated by autocrine FGF-2, NT-3, andTGF-b. They further suggest that NET is expressedalready in some premigratory neural crest cells, mostlikely in stem cells.
EXPERIMENTAL PROCEDURES
Cloning of a Partial Quail NET
Quail neural crest cells were grown for 4 days inculture and total RNA was isolated using the RNASTAT-60 kit (TEL-TEST Inc., Friendswood, TX). A
FIG. 5. TGF-b immunoreactivity in 12 h neural crest cell cultures anbind anti-pan TGF-b antibodies. (b) Negative control for second a
reabsorbed primary antibody. Bar, 20 mm. Embryo: (d) pan-TGF-b imoot ganglion (drg), ventral root (vr), neural tube (nt), and myotombryonic section to identify neural crest cells and neural crest derivaigrating neural crest cells and the dorsal part of the dorsal root ga
cDNA pool was generated using hexamers and oligodT12-18mer as primers (cDNA Synthesis System, Life
Technologies), followed by touch-down RT-PCR (RouxKH, 1995) with degenerated NET-specific oligos: NET-EL2s (59TGG ACC ASY CCM AAY TGY ACN GAC
C39) and NET-IL4as (59GTG ATR ACH GCY TCCTR CCW CCC AT39). A 750-bp PCR products was
ubcloned into pCRII (Invitrogen) and sequenced. Fromhis fragment we designed two outward facing primers,
hich were subsequently used together with degener-ted NET primers from TM1 (59GRG AGM MYT GGGCA AGR ARA TYG A39) and TM12 (59TAG RYR GGB
ACC AGR ABC ATG GAK GA39) to clone 3 overlap-ping fragments that encode 515aa of the qNET gene. Forthe in situ hybridization experiments we used the initial750-bp PCR product, cloned in both orientations intopCRII, reverse-transcribed from the vector’s T7 pro-moter.
Neural Crest Primary Cultures
Neural crest cell cultures were prepared and main-tained in chemically defined culture medium as de-scribed previously (Sieber-Blum and Cohen, 1980;Sieber-Blum, 1999). Thirty-five-millimeter culture plateswere coated with collagen, laminin (28 mg per plate),and fibronectin (100 mg plasma fibronectin from horseserum). The last six segments of Hamburger-Hamilton(1951) stage 14 quail embryos (49 h of incubation) were
he embryo. Culture: (a) Virtually all neural crest cells in 12-h culturesdy: no primary antibody. (c) Negative control for first antibody:oreactivity in day 3.5 embryo. The sympathetic ganglion (sg), dorsal
m) are immunoreactive. (e) HNK-1 immunoreactivity in the same(f) Higher magnification of area marked by white square in (e) shows
n. Bars: (d, e) 200 mm; (f) 25 mm.
d in tntibomun
excised and isolated. At 24 h postexplantation, the neu-ral tubes were removed, leaving the emigrated neural

O
w
1
4fcia4(
dFempnsqcs
lh1
546 Ren et al.
crest cells on the substratum. The culture medium wasexchanged every day. The cultures were incubated at37°C in a humidified atmosphere of 5% CO2 and 10%
2. The defined culture medium consisted of MCDB201 and was supplemented with transport factors, hor-mones, vitamins, and small molecular nutrients as de-scribed (Sieber-Blum and Chokshi, 1985; Sieber-Blum,1999). It did not contain any serum or embryo extractexcept where indicated (Table 1).
FIG. 7. Summary of FGF-2, NT-3, TGF-b1, and NET action in theifferentiation of quail neural crest cells into sympathetic neuroblasts.GF-2 is synthesized by early migratory neural crest cells, whichxpress FGF receptor (Heuer et al., 1990; Murphy et al., 1994). FGF-2 isitogenic for all neural crest cells (Zhang et al., 1997b). However,
roliferating presumptive neuronal cells do not survive unless aeurotrophin (NGF, BDNF, or NT-3) is present as well. NT-3 isynthesized by neural crest cells (Fig. 3). NT-3 causes an increase inNET mRNA levels (Fig. 2). TGF-b1 causes cessation of neural crestell proliferation (Zhang et al., 1997) and an increase in qNET expres-ion (Fig. 2). TGF-bs are synthesized by neural crest cells (Fig. 3) and
may thus also act as autocrine factors. The avian notochord synthe-sizes NE (Strudel et al., 1977). As NET-expressing neural crest cellsmigrate past the notochord, they are likely to encounter and transportNE. NE transport causes an increase in Ca21 transients and eventuallyeads to the expression of the NE biosynthetic enzymes, tyrosineydroxylase (TH) and dopamine-b-hydroxylase (DBH; Zhang et al.,997b).
TGF b1 (Upstate Biotechnology, Lake Placid, NY)as used at 1 ng/ml (Zhang et al., 1997b), FGF-2 (Up-
fiw
state Biotechnology) at 2.5 ng/ml, and hrNT-3 (gift ofGenentech Inc., South San Francisco, CA and from Pro-mega, Madison, WI) at 10 ng/ml.
Where indicated anti-NT-3 blocking antibody (mousemonoclonal IgG, 2 mg/ml; gift of Y.-A. Barde and I.Bartke; Gaese et al. 1994) were added to the cultures at:500 and pan-TGF-b blocking antibodies (rabbit IgG, 1
mg/ml, Research Biochemicals International, Natick,MA) at 1:250.
Neural Crest Colony Assay
Neural crest cell colony assays were performed asdescribed (e.g., Sieber-Blum and Cohen, 1980; Zhang etal., 1997a; Sieber-Blum, 1999). Briefly, cells from 18–24 hprimary explants were resuspended by trypsinizationand the cell density adjusted to 500 cells per milliliter.One-milliliter aliquots were then placed into each of 10collagen- and fibronectin-coated plates per experimen-tal group. The cells attached to the substratum withinhalf an hour. One hour after plating, a second milliliterof culture medium, which in experimental groups con-tained a 23 concentration of growth factor(s), wasadded to each plate. The culture medium and growthfactors were replaced every other day. At day 10, thecultures were processed for norepinephrine uptake asdetailed below.
Norepinephrine Uptake and Autoradiography
Neural crest cells with a functional norepinephrinetransporter were identified in situ in colony assays ex-actly as described previously (Zhang and Sieber-Blum,1992). Briefly, the cultures were rinsed with Hanks’balanced salt solution (HBSS) containing 1% bovineserum albumin (BSA). They were then incubated for 2 hat 37°C with 0.5 ml of 0.5 mM [3H]norepinephrine (spact, 40.8 Ci/mmol) in HBSS that also contained 1 mMascorbic acid and 0.1 mM monoamine oxidase inhibitor,pargyline (Sigma). Subsequently, uptake of radioactivenorepinephrine was terminated by rinsing the culturesthree times with HBSS that contained 24 mM nonradio-active norepinephrine (d,l-arterenol; Sigma), fixed with% paraformaldehyde in calcium-magnesium-free PBSor 20 min at room temperature, and rinsed again. Theultures were then dried in a stream of cold air, coatedn the dark with NTB2 emulsion (Kodak) for 2.5 min,nd air-dried in the dark. After 10 days of exposure at°C, autoradiograms were developed with D-19Kodak) for 2.5 min, rinsed with 1% acetic acid, and
xed with Rapid Fix (Kodak). Plates were mountedith mineral oil and a coverslip. The total number of
Gl
547Autocrine Regulation of NET Expression
colonies per plate and the number of colonies that con-tained grain-positive cells were scored and the datastatistically analyzed by 1-factor analysis of variance.
RT-PCR
Primers were synthesized by Operon Technologies,Inc. (Alameda, CA). The following primer pairs wereused. TGF-b1; CAAGCTGAGCGTGCACTGT and TC-CTTGCGGAAGTCGATGT (product length, 281 bp; Ja-kowlew et al., 1988a). TGF-b2; AGGAATGTGCAG-
ATAATT and ATTTTGGGTGTTTTGCCAA (productength, 269 bp; Burt and Paton, 1991). TGF-b3; GAG-
CAGAGTTCCGGGTGCT and GTGCAGAAGCCACT-CACGC (product length, 200 bp; Jakowlew et al.,1988b). NT-3; CATATCTTCGTGGCATTCAG andCCAGTGGTGTGTTGTCACTT (product length, 306bp; Maisonpierre et al., 1992). qNET; GGATTGATGC-GGCTACTCAGA and GCTTCCATGCCACCCATAGA(product length, 358 bp). Glyceraldehyde-3-phosphatedehydrogenase (GAPDH); ACG CCA TCA CTA TCTTCC AG and TCAGCTCAGGGATGACTTTC (productlength, 458 bp; Panabieres et al., 1984).
Total RNA was isolated using the RNA STAT-60 kit(TEL-TEST Inc.). For RT-PCR, the “Access RT-PCR Sys-tem” (Promega) was used. Template RNA was reversetranscribed with AMV RT for 45 min at 48°C, followedby 2 min of inactivation of RT and denaturation at 94°C.The conditions for PCR reactions are described below.
Denaturation Annealing Extension Cycles
NET 94°C, 30 s 60°C, 1 min 68°C, 2 min 36GAPDH 94°C, 30 s 60°C, 1 min 68°C, 2 min 36TGF-b1 94°C, 1 min 55°C, 1 min 72°C, 2 min 40TGF-b2 94°C, 1 min 55°C, 1 min 72°C, 2 min 40TGF-b3 94°C, 1 min 50°C, 2 min 72°C, 3 min 40NT-3 94°C, 30 s 60°C, 1 min 68°C, 2 min 40
PCR products were analyzed by agarose gel (2%) elec-trophoresis, sequenced, and found to be correct. Forcontrols, the PCR reaction was carried out without atemplate and without reverse transcriptase. The PCRconditions were optimized in pilot experiments to as-sure linear amplification.
Immunocytochemistry and Immunohistochemistry
Antibodies. NET rabbit antiserum 43408 (used at
1:2000; Schroeter et al., 2000) is an anti-peptide antibodythat was raised against an external epitope of humanNET (TKYSKYKFTPAAEFY), which is highly con-served in qNET (identical except for F 3 L in qNET).Anti-NT-3 antibodies used for immunocytochemistryand immunohistochemistry (rabbit antiserum, 1:400;Pepro Tech Inc., Rocky Hill, NJ). Anti-panTGF b anti-bodies; rabbit IgG (used at 1:400; AB-100-NA; R&DSystems Inc., Minneapolis, MN). HNK-1; mouse mono-clonal IgM (used at 1:1000; N-202RBI; Research Bio-chemicals International); Rhodamine-conjugated goatantibody to mouse (used at 1:100; Cappel 55540, Or-ganon Teknika Corp., West Chester, PA). Fluorescein-conjugated goat antibody to rabbit (used at 1:80; Jack-son Immunoresearch Laboratories, Inc., West Grove,PA).
Method. The cell culture plates were rinsed withPBS, incubated with 5% normal goat serum in PBS for30 min, and subsequently incubated overnight at 10°Cwith primary antibodies in PBS that contained 5% goatserum and 0.1% Triton-3100. For multiple stains, theprimary antibodies were pooled. The sections/cultureswere rinsed three times for 10 min each with PBS. Thesecondary antibodies were diluted in 5% normal goatserum in PBS, added to the sections/cultures, and in-cubated in the dark for 1 h at room temperature. Formultiple stains the secondary antibodies were pooled.After three more rinses at 10 min each, the slides/culture plates were mounted with 50% glycerol [con-taining 1 mg/ml paraphenylene diamine (PPD) in PBS,pH 8.5] and a cover slip and were observed with theconfocal microscope.
In Situ Hybridization
In situ hybridization was performed as described byTessarollo and Parada (1995), except for the followingmodifications: The embryos/cultures were fixed with4% paraformaldehyde 4–12 h, incubated with 0.2% gly-cine in DEPC-PBS for 2 min, and permeabilized with0.2% Triton X-100 for 30 min. For probe preparation,qNET was linearized with BamHI. The Genius 4 RNALabeling kit (Boehringer-Mannheim) was used to syn-thesize digoxigenin-labeled RNA according to manu-facturer instruction. For hybridization, the cultureswere prehybridized with 50% formamide, 43 SSC, 10%Dextran sulfate, 13 Denhardts’ solution, 0.25% mg/mlyeast RNA, and 10 mg/ml sperm DNA (without probe)for 2 h at 50°C. The probe was used at 0.5–1 mg/ml. Forvisualization, we used alkaline phosphatase-conjugated
anti-digoxigenin-antibodies and BM purple accordingto manufacturer instruction. The tissue was then rinsed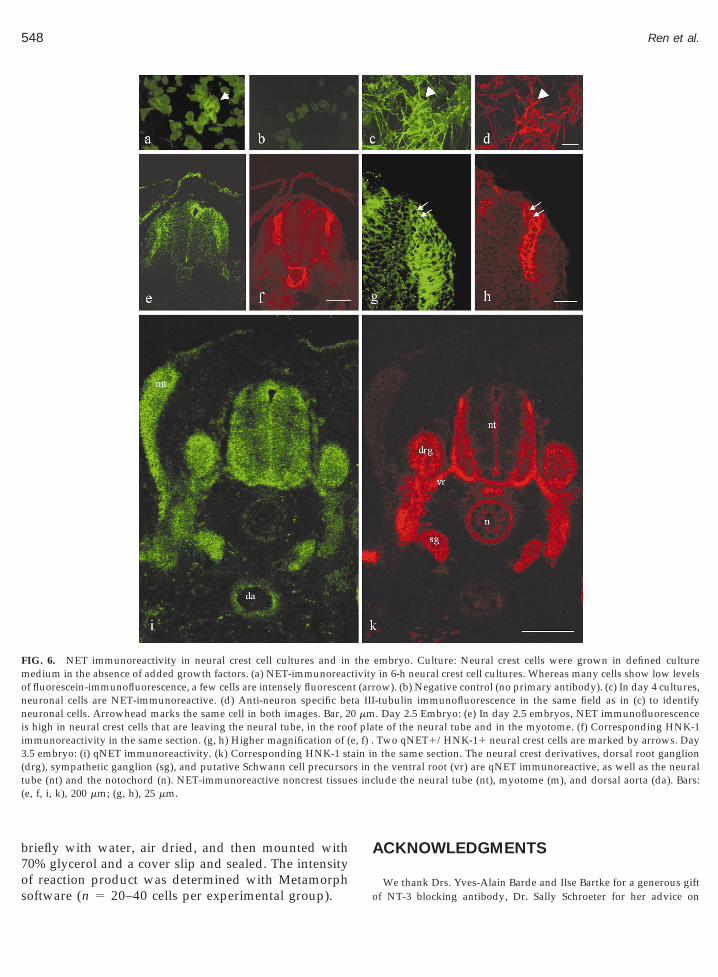
3(t s inc(
548 Ren et al.
briefly with water, air dried, and then mounted with70% glycerol and a cover slip and sealed. The intensity
FIG. 6. NET immunoreactivity in neural crest cell cultures and inmedium in the absence of added growth factors. (a) NET-immunoreacof fluorescein-immunofluorescence, a few cells are intensely fluorescenneuronal cells are NET-immunoreactive. (d) Anti-neuron specific beneuronal cells. Arrowhead marks the same cell in both images. Bar, 2is high in neural crest cells that are leaving the neural tube, in the rooimmunoreactivity in the same section. (g, h) Higher magnification of (.5 embryo: (i) qNET immunoreactivity. (k) Corresponding HNK-1 stdrg), sympathetic ganglion (sg), and putative Schwann cell precursoube (nt) and the notochord (n). NET-immunoreactive noncrest tissuee, f, i, k), 200 mm; (g, h), 25 mm.
of reaction product was determined with Metamorphsoftware (n 5 20–40 cells per experimental group).
ACKNOWLEDGMENTS
embryo. Culture: Neural crest cells were grown in defined culturein 6-h neural crest cell cultures. Whereas many cells show low levels
ow). (b) Negative control (no primary antibody). (c) In day 4 cultures,-tubulin immunofluorescence in the same field as in (c) to identify. Day 2.5 Embryo: (e) In day 2.5 embryos, NET immunofluorescencete of the neural tube and in the myotome. (f) Corresponding HNK-1Two qNET1/HNK-11 neural crest cells are marked by arrows. Daythe same section. The neural crest derivatives, dorsal root ganglion
the ventral root (vr) are qNET immunoreactive, as well as the neurallude the neural tube (nt), myotome (m), and dorsal aorta (da). Bars:
thetivityt (arrta III0 mmf plae, f) .ain inrs in
We thank Drs. Yves-Alain Barde and Ilse Bartke for a generous giftof NT-3 blocking antibody, Dr. Sally Schroeter for her advice on

pCINIfw
B
B
B
B
C
F
F
G
G
H
H
H
549Autocrine Regulation of NET Expression
NET-immunohistochemistry and Dr. Stephen Duncan for his advicewith in situ hybridization. MSB thanks Dr. Joe Besharse for his sup-
ort. The work was supported by a research grant from the Medicalollege of Wisconsin (MSB) and USPHS grants from the National
nstitute of Neurological Disease and Stroke (NS38281, MSB), theational Institute of Drug Abuse (DA07595, SGA) and the National
nstitute for Mental Health (MH58921, RDB). PP received fundingrom the Deutsche Forschungsgemeinschaft. SGA is an investigator
ith the Howard Hughes Medical Institute.
REFERENCES
Amara, S. G. (1995). Monoamine transporters: Basic biology withclinical implications. Neuroscientist 1: 259–267.
Axelrod, J. (1965). The metabolism, storage, and release of cat-echolamines. Recent Progr. Hormone Res. 21: 597–619.
Axelrod, J., Weil-Malherbe, H., and Tomchick, R. (1959). The physi-ological disposition of H3-epinphrine and its metabolite metaneph-rine. J. Pharmacol. Exp. Ther. 127: 251–256.
lakely, R. D., Ramamoorthy, S., Schroeter, S., Qian, Y., Apar-sundaram, S., Galli, A., and DeFelice, L. J. (1998). Regulated phos-phorylation and trafficking of antidepressant-sensitive serotonintransporter proteins. Biol. Psychiatry 44: 169–178.
uchman, V. L., Sporn, M., and Davies, A. M. (1994). Role of trans-forming growth factor-beta in regulating the expression of nervegrowth factor and neurotrophin-3 mRNA levels in embryonic cu-taneous cells at different stages of development. Development 120:1621–1629.
rauer, P. R., and Yee, J. A. (1993). Cranial neural crest cells synthesizeand secrete a latent form of transforming growth factor b that canbe activated by neural crest cell proteolysis. Dev. Biol. 155: 281–285.
russ, M., Porzgen, P., Bryan-Lluka, L. J., and Bonisch, H. (1997). Therat norepinephrine transporter: Molecular cloning from PC12 cellsand functional expression. Brain Res. Mol. Brain Res. 52: 257–262.
Burt, D. W., and Paton, I. R. (1991). Molecular cloning and primarystructure of the chicken transforming growth factor-b2 gene. DNACell Biol. 10: 723–734.
ai, F., Campana W. M., Tomlinson, D. R., and Fernyhough, P. (1999).Transforming growth factor-beta1 and glial growth factor 2 reduceneurotrophin-3 mRNA expression in cultured Schwann cells via acAMP-dependent pathway. Brain Res. Mol. Brain Res. 71: 256–264.
arinas, I., Jones, K. R., Mackus, C., and Reichardt, L. F. (1994). Severesensory and sympathetic deficits in mice lacking neurotrophin-3.Nature 369: 658–661.
ritz, J. D., Jayanthi, L. D., Thoreson, M. A., and Blakely, R. D. (1998).Cloning and chromosomal mapping of the murine norepinephrinetransporter. J. Neurochem. 70: 2241–2251.aese, F., Kolbeck, R., and Barde, Y.-A. (1994). Sensory ganglia re-quire neurotrophin-3 early in development. Development 120: 1613–1619.ershon, M. D., Rothman, T. P., Joh, T. H., and Teitelman, G. N.(1984). Transient and differential expression of aspects of the cat-echolaminergic phenotype during development of the fetal bowelof rats and mice. J. Neurosci. 4: 2269–2280.amburger, V., and Hamilton, H. (1951). A series of normal stages inthe development of the chick embryo. J. Morphol. 88: 49–92.enion, P. D., Garner, A. S., Large, T. H., and Weston, J. A. (1995).TrkC-mediated NT-3 signaling is required for the early develop-
ment of a subpopulation of neurogenic neural crest cells. Dev. Biol.172: 602–613.euer, J. G., von Bartheld, C. S., Kinoshita, Y., Evers, P., and Bothwell,M. (1990). Alternating phases of FGF receptor and NGF receptorexpression in the developing chicken nervous system. Neuron 5:283–296.
Howard, M. J., and Gershon, M. D. (1993). Role of growth factors incatecholaminergic expression by neural crest cells: In vitro effects oftransforming growth factor beta 1. Dev. Dynam. 196: 1–10.
Iversen, L. L. (1967). The Uptake and Storage of Noradrenaline in Sym-pathetic Nerves. Cambridge Univ. Press, New York.
Jakowlew, S. B., Dillard, P. J., Sporn, M. B., and Roberts, A. B. (1988a).Complementary deoxyribonucleic acid cloning of a messenger ri-bonucleic acid encoding transforming growth factor b 4 fromchicken embryo chondrocytes. Mol. Endocrinol. 2: 1186–1195.
Jakowlew, S. B., Dillard, P. J., Kondaiah, P., Sporn, M. B., and Roberts,A. B. (1988b). Complementary deoxyribonucleic acid cloning of anovel transforming growth factor-b messenger ribonucleic acidfrom chicken embryo chondrocytes. Mol. Endocrinol. 2: 747–755.
Lo, L, Tiveron, M.-C., and Anderson, D. J. (1998). Mash 1 activatesexpression of the paired homeodomain transcription factor Phox2a,and couples pan-neuronal and subtype-specific components of au-tonomic neuronal identity. Development 125: 609–620.
Maisonpierre, P. C., Belluscio, L., Conover, J. C., and Yancopoulos,G. D. (1992). Gene sequences of chicken BDNF and NT-3. DNA Seq.3(1): 49–54.
Murphy, M., Reid, K., Ford, M., Furness, J. B., and Bartlett, P. F. (1994).FGF2 regulates proliferation of neural crest cells, with subsequentneuronal differentiation regulated by LIF or related factors. Devel-opment 120: 3519–3528.
Pacholczyk, T., Blakely, R. D., and Amara, S. G. (1991). Expressioncloning of a cocaine- and antidepressant-sensitive human nor-adrenaline transporter. Nature 350: 350–353.
Panabieres, F., Piechaczyk, M., Rainer, B., Dani, C., Fort, P., Riaad, S.,Marti, L., Imbach, J. L., Jeanteur, P., and Blanchard, J.-M. M. (1984).Complete nucelotide sequence of the messenger RNA coding forchicken muscle glyceraldehyde-3-phosphate dehydrogenase. Bio-chem. Biophys. Res. Commun. 118: 767–773.
Pinco, O., Carmeli, C., Rosenthal, A., and Kalcheim, C. (1993). Neu-rotrophin-3 affects proliferation and differentiation of distinct neu-ral crest cells and is present in the early neural tube of avianembryos. J. Neurobiol. 24: 1626–1641.
Rothman, T. P., Gershon, M. D., and Holtzer, H. (1978). The relation-ship of cell division to the acquisition of adrenergic characteristicsby developing sympathetic ganglion cell precursors. Dev. Biol. 65:322–341.
Roux, K. H. (1995). Optimization and troubleshooting in PCR. PCRMethods Appl. 4(5): S185–S194.
Schroeter, S., Apparsundaram, S., Wiley, R. G., Miner, L. A. H.,Sesack, S. R., and Blakely, R. D. (2000). Immunolocalization of thecocaine- and antidepressant-sensitive l-norepinephrine transporter.J. Comp. Neurol. 420: 211–232.
Sieber-Blum, M. (1989a). Inhibition of the adrenergic phenotype incultured neural crest cells by norepinephrine uptake inhibitors.Dev. Biol. 136: 372–380.
Sieber-Blum, M. (1989b). Commitment of neural crest cells to thesensory neuron lineage. Science 243: 1608–1611.
Sieber-Blum, M. (1999). The neural crest colony assay: Assessingmolecular influences on development in culture. In The Neuron inTissue Culture, IBRO, pp 5–22. Wiley.
Sieber-Blum, M., and Chokshi, H. R. (1985). In vitro proliferation andterminal differentiation of quail neural crest cells in a defined
culture medium. Exp. Cell Res. 158: 267–272.Sieber-Blum, M., and Cohen, A. M. (1980). Clonal analysis of quail

S
S
S
T
T
W
X
Z
Z
Z
Z
Z
Z
550 Ren et al.
neural crest cells: They are pluripotent and differentiate in vitro inthe absence of non-crest cells. Dev. Biol. 80: 96–106.
ieber-Blum, M., and Sieber, F. (1984). Heterogeneity among earlyquail neural crest cells. Dev. Brain Res. 14: 241–246.
nyder, S. H. (1970). Putative neurotransmitters in the brain: Selectiveneuronal uptake, subcellular localization, and interactions withcentrally acting drugs. Biol. Psychiatry 2: 367–389.
trudel, G., Recasens, M., and Mandel, P. (1997). Identification decatecholamines et de serotonine dans les chordes d’embryons depoulet. C.R. Acad. Sci. Paris 284: 967–969.
essarollo, L., Tsoulfas, P., Martin-Zanca, D., Gilbert, D. J., Jenkins,N. A., Copeland, N. G., and Parada, L. F. (1993). TrkC, a receptor forneurotrophin-3 is widely expressed in the developing nervoussystem. Development 118: 463–475.
essarollo, L., and Parada, L. F. (1995). In situ hybridization. MethodsEnzymol. 254: 419–430.hitby, L. G., Hertting, G., and Axelrod, J. (1960). Effect of cocaine onthe disposition of noradrenaline labelled with tritium. Nature 187:604–605.
ue, Z.-G., Smith, J., and Le Douarin, N. M. (1985). Differentiation of
catecholaminergic cells in cultures of embryonic avian sensoryganglia. Proc. Natl. Acad. Sci. USA 82: 8800–8804.aremba, S., Naegele, J. R., Barnstable, C. J., and Hockfield, S. (1990).Neuronal subsets express multiple high-molecular-weight cell-sur-face glycoconjugates defined by monoclonal antibodies Cat-301 andVD1.1. J. Neurosci. 10: 2985–2995.
hang, Y., and Derynck, R. (1999). Regulation of Smad signaling byprotein associations and signaling crosstalk. Trends Cell Biol. 9:274–279.
hang, J.-M., and Sieber-Blum, M. (1992). Characterization of thenorepinephrine uptake system and the role of norepinephrine inthe expression of the adrenergic phenotype by quail neural crestcells in clonal culture. Brain Res. 570: 251–258.
hang, J.-M., Dix, J., Langtimm-Sedlak, C. J., Trusk, T., Schroeder, B.,Hoffmann, R., Strosberg, A. D., Winslow, J. W., and Sieber-Blum,M. (1997a). Neurotrophin-3 and Norepinephrine-mediated adren-ergic differentiation and the inhibitory action of desipramine andcocaine. J. Neurobiol. 32: 262–280.
hang, J. M., Hoffmann, R., and M. Sieber-Blum, M. (1997b). Mito-genic and anti-proliferative signals for neural crest cells and theneurogenic action of TGF-b1. Dev. Biol. 208: 375–386.
hang, D. Yao, L., and Bernd, P. (1994). Expression of trk and neu-
rotrophin RNA in dorsal root and sympathetic ganglia of the quailduring development. J. Neurobiol. 25: 1517–1532.Received September 11, 2000Revised November 20, 2000Accepted December 4, 2000















![The norepinephrine transporter (NET) radioligand (S,S)-[ 18F]FMeNER-D 2 shows significant decreases in NET density in the human brain in Alzheimer's disease: A post-mortem autoradiographic](https://img.pdfslide.net/doc/110x75/635cac5beab478482e01741c/the-norepinephrine-transporter-net-radioligand-ss-18ffmener-d-2-shows-significant.jpg)












![Glucose transporter proteins in brain. [Review]](https://img.pdfslide.net/doc/110x75/634931b8de40dd034d099a07/glucose-transporter-proteins-in-brain-review.jpg)
