Embed Size (px)
Citation preview

Exp. Eye Rtls. (1991) 53, 615-622
Autoradiographic, Electrophoretic, and lmmunocytochemical Studies of Glycoproteins of the Rabbit Iris
ANTONIO HADDAD”, EDUARDO M. LAICINE”, ROSE S. FIFEb*, ANTONIO JORDAO, Jr.” AND GEORGES PELLETIER”
a Departamento de Morfologia, Faculdade de Medicina de Ribeira”o Preto, 74049 Ribeira”o Preto-S.P., Brasil, b Rheumatology Division, Departments of Medicine and Biochemistry and Molecular Biology,
Indiana University School of Medicine, lndianopolis, IN, U.S.A. and c C.H.U.L.-Centre de Recherches en Endocrinologie Mole’culaire, Que’bec, Canada
(Received 4 October 1990 and accepted in revised form 9 January 1991)
L-[3H]fucose was injected either intravitreally or intra-aqueously into adult rabbits which were killed at several time points after injection. SDS-polyacrylamide gel electrophoresis and fluorography of iris extracts revealed that most of the proteins are glycoproteins containing fucose residues. Autoradiography of semi-thin histologic sections demonstrated that glycoprotein synthesis was most prominent in the epithelium of the iris, while little protein synthesis was evident in the stroma of the iris. The results of these experiments indicated that the glycoproteins of the iris undergo renewal. The protein band pattern of the iris extracts was very similar to that of extracts of the ciliary body. The high-molecular-weight cartilage matrix glycoprotein (CMGP), an intrinsic component of the ciliary body, vitreous, and aqueous humor, was detected by immunohistologic studies only in the stroma of the iris. The results of immunohistochemical analyses of the eyes of young rabbits (1-21 days old), in addition to the autoradiographic findings, strongly suggest that CMGP is not an intrinsic glycoprotein of the iris stroma, at least in this species.
Ke;/ words: glycoproteins ; iris : CMGP.
1. Introduction
Glycoprotein biosynthesis has been studied in the ciliary body of rats (Bennett and Haddad, 1986) and rabbits (Haddad et al., 1990) using autoradiography following the administration of [3H]fucose, a specific marker for newly synthesized glycoproteins (Schachter and Roden, 1973). From such investigations it has become evident that most glycoproteins synthesized by both layers of the ciliary epithelium eventually migrate to the plasma membranes. In earlier studies in rabbits, the inner (non-pigmented) layer of the ciliary epithelium was shown to be involved in the secretion of the intrinsic glycoproteins of the vitreous (Haddad et al., 1990). One of these glycoproteins was identified by immunocytochemical studies as a 550 OOO-Da cartilage matrix glycoprotein (CMGP) that has been described previously in the human and bovine vitreous body (Nguyen and Fife, 1986). In other investigations, vitreous glycoproteins have been identified in the aqueous humor as well (Haddad et al., in press).
Studies of the structure and development of the iris and ciliary body demonstrate that these tissues are in continuity (Ozanics and Jakobiec, 1982 ; Streeten, 19 82). The layer of fibroblasts comprising the exterior portion of the anterior surface of the iris does not
* For correspondence at: Rheumatology Division, Indiana Uni- versity School of medicine, 541 Clinical Drive, Indianapolis, IN 46202. IJ.S.A.
00144835/91/110615+08 $03.00/0
appear to be a barrier to macromolecules, since spaces between the fibroblasts are easily identified by electron microscopy (Rodrigues et al., 1982). Therefore, macro- molecular components of the aqueous humor could enter the iris through its anterior surface and reach the deeper regions of its stroma.
To our knowledge there are no reports of the characterization of the glycoproteins of the iris. The present studies were performed to examine the synthesis, migration, and turnover of these proteins. [3H]fucose was injected intravitreally or intra- aqueously into rabbits, and the labeled glycoproteins were studied in tissue sections of the iris by autoradi- ography and in extracts of the iris by electrophoresis and fluorography.
2. Materials and Methods
Animals
Male albino rabbits, weighing 2.5-2.9 kg, were injected either intravitreally with 60 pCi of L-( 5,6)- [3H]fucose (specific activity, 56 Ci mmol-I) (New England Nuclear, Boston, MA) in saline or intra- aqueously with 20 ,uCi of the same isotope (Haddad et al., in press). The rabbits were anesthetized with sodium thiopental prior to the ocular injections or decapitation. Pieces of iris were obtained by dissecting the eyes immediately after death. The animals injected intravitreally were killed at 1 and 4 hr, and 1 I 3. 7, 14,
0 199 1 Academic Press Limited
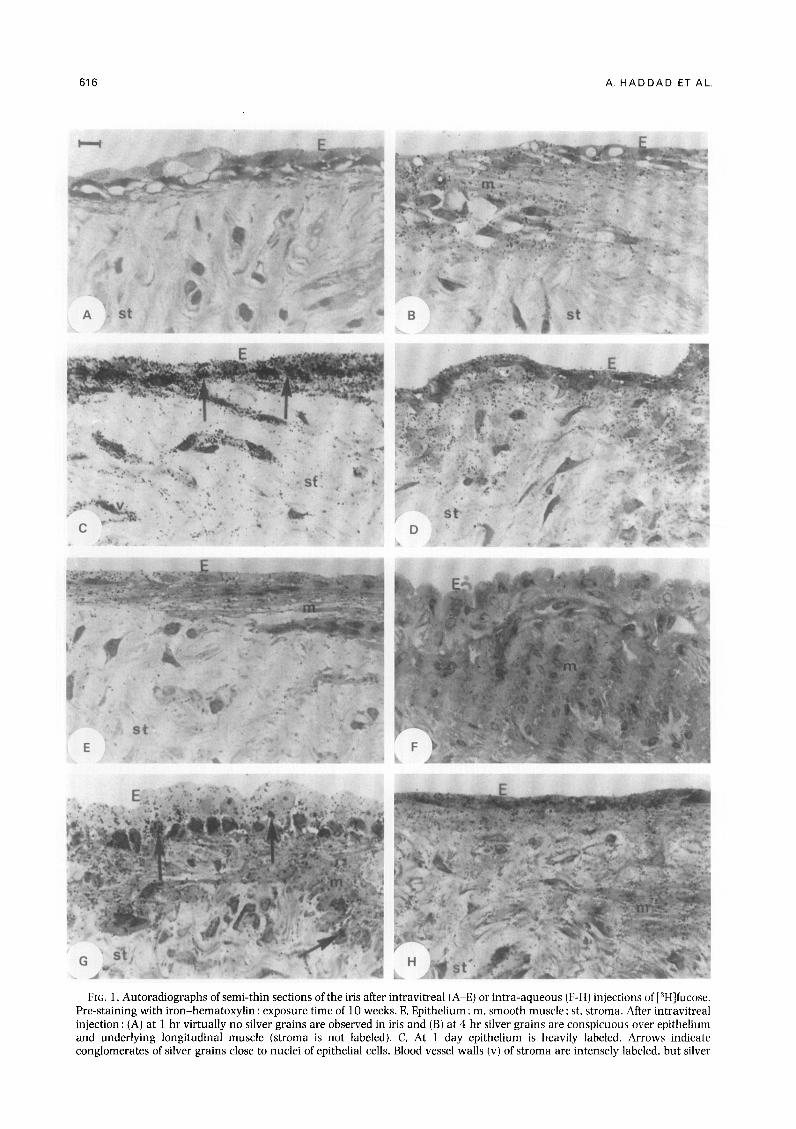
616 A. HADDAD ET AL.
FIG. 1. Autoradiographs of semi-thin sections of the iris after intravitreal (A-E) or intra-aqueous (F-H) injections of [3H]fucose. Pre-staining with iron-hematoxylin : exposure time of 10 weeks. E. Epithelium ; m. smooth muscle : st, stroma. After intravitreal injection: (A) at 1 hr virtually no silver grains are observed in iris and (B) at 4 hr silver grains are conspicuous over epithelium and underlying longitudinal muscle (stroma is not labeled). C, At 1 day epithelium is heavily labeled. Arrows indicate conglomerates of silver grains close to nuclei of epithelial cells. Blood vessel walls (v) of stroma are intensely labeled, but silver

GLYCOPROTEINS OF THE IRIS
21, and 28 days after injection of [3H]fucose. For the experiments involving intra-aqueous injections, rab- bits were killed at 0.5, 1, and 4 hr, and 1, 3, and 7 days after the administration of [3H]fucose.
Animal experiments were conducted in compliance with the ARVO Resolution on the Use of Animals in Research and with the Institutional Committee for Animal Care.
Autoradiographlg
Irises were fixed in 4% glutaraldehyde in 0.1 M
Sorensen’s phosphate buffer for 3 hr, washed over- night in buffer, fixed in 1 (X0 osmium tetroxide in phosphate buffer for 2 hr, dehydrated in graded ethanol, cleared in propylene oxide, and embedded in Epon. The pieces were placed in the lids of BEEM capsules to obtain sections in the anteroposterior orientation. Semi-thin sections (0.75 pm) were stained with iron-hematoxylin, coated with Kodak NTB2 photographic emulsion (Kodak Co., Rochester, NY), exposed for 1, 3, or 10 weeks, developed in D-l 70 (Kodak), fixed in sodium thiosulfate, washed in water, air-dried, and mounted. Counts of silver grains were performed on autoradiographs exposed for 3 weeks, and the silver grain density (mean number of grains per 500 jlrn”) was estimated for the iris epithelium, the iris stroma (connective tissue), and the smooth muscles at several time points after intravitreal or intra-aqueous injection of [3H]fucose.
lmmunocyrochemistry
Young rabbits, aged 1, 7, 14, and 21 days, and adult rabbits were anesthetized and perfused through the left ventricle with 4% formaldehyde (freshly prepared from paraformaldehyde) in 0.1 M phosphate buffer. Irises were processed for paraffin embedding. The eyes of 150-g rats, processed in the same manner, were also studied. Intact whole rat eyes were em- bedded in paraffin.
The immunohistochemical techniques were per- formed as follows: (1) sections were incubated over- night at 4°C with a monoclonal antibody to CMGP (Fife and Brandt. 1989). diluted 1: 500 in 0.05 M
Tris-HCl buffer containing 1% bovine serum albumin ; control sections were incubated with non-immune mouse serum ; (2) after rinsing in buffer, sections were incubated for 4 hr with goat anti-mouse immuno- globulin (Fab’), antiserum conjugated with peroxidase and diluted 1: 500 ; (3) after rinsing in buffer, sections were immersed for 5 min in a solution of 0.0 5 ‘X0 3,3’-
617
diaminobenzidine in Tris containing 0.02 % hydrogen peroxide ; and (4) sections were washed in buffer, dehydrated, cleared, and mounted. Some sections were counterstained with Harris hematoxylin.
Electrophoresis and Fluorography
For these experiments adult rabbits were injected either intravitreally or intra-aqueously with [3H]fucose. Irises were cut into small pieces, washed in ice-cold saline, and immersed for 3-S hr in ice-cold 70% ethanol with frequent changes and shaking. The pieces were dissolved either in a 4% solution of SDS in water or in the sample solvent used for polyacrylamide gel electrophoresis (Laemmli, 1970) at a proportion of 1 mg of wet mass per 20 ~1 of either solution. Complete dissolution of tissues was achieved after heating for a few minutes at 50% followed by treatment with ultrasound, shaking, and centrifugation. The con- centration of iris proteins was determined for the SDS solutions using the method of Lowry et al. (19 51). Radioactivity was estimated by counting aliquots of iris solutions in a Beckman LS-6800 liquid scintillation counter.
Electrophoresis was performed in l-mm thick SDS-polyacrylamide slab gels [9.7X T (total poly- acrylamide polymer), 10% C (cross-linked)], using 30 ,ug of iris protein dissolved in sample solvent in each well (Laemmli, 1970). After electrophoresis, gels were stained with Coomassie blue (Bio-Rad Lab- oratories, Richmond, CA), photographed, and pro- cessed for fluorography (Waterborg and Matthews, 1984). In other gels, samples of ciliary body and iris from the same eyes were electrophoresed in adjacent lanes.
3. Results
Autoradiography
Silver grains, indicating the presence of newly synthesized glycoproteins labeled with [3H]fucose, were observed in the different tissue components of the iris between 4 hr and 14 days after intravitreal injection [Fig. l(A)-(E)]. At 1 hr [Fig. l(A)] and at 21 and 28 days, the number of silver grains in the iris was negligible. Except for the paranuclear accumulations of silver grains found in the epithelial cells 1 day after injection [Fig. l(C)], the silver grains were seen primarily over the epithelium, smooth muscles, and stroma. The epithelium was the most heavily labeled component of the iris at all time points, and its silver grain density reached a peak 1 day after injection. decreasing continuously thereafter (Fig. 2). The curve
grain density over remainder of stroma is low. D. At 3 days silver grain density over the iris is less than in (C). E. At 14 days some silver grains are visualized over epithelium and smooth muscle but are less conspicuous over stroma. After intra-aqueous injection: (F) at 30 min the autoradiographic reaction over the iris is virtually absent; (G) at 1 hr silver grains are numerous over the entire iris. Note conglomerates of silver grains (arrows) near nuclei. H, At 1 day [compare with (A)] silver grain density over epithelium and adjacent smooth muscles is much lower than in (C). Bar = 10 jtm.
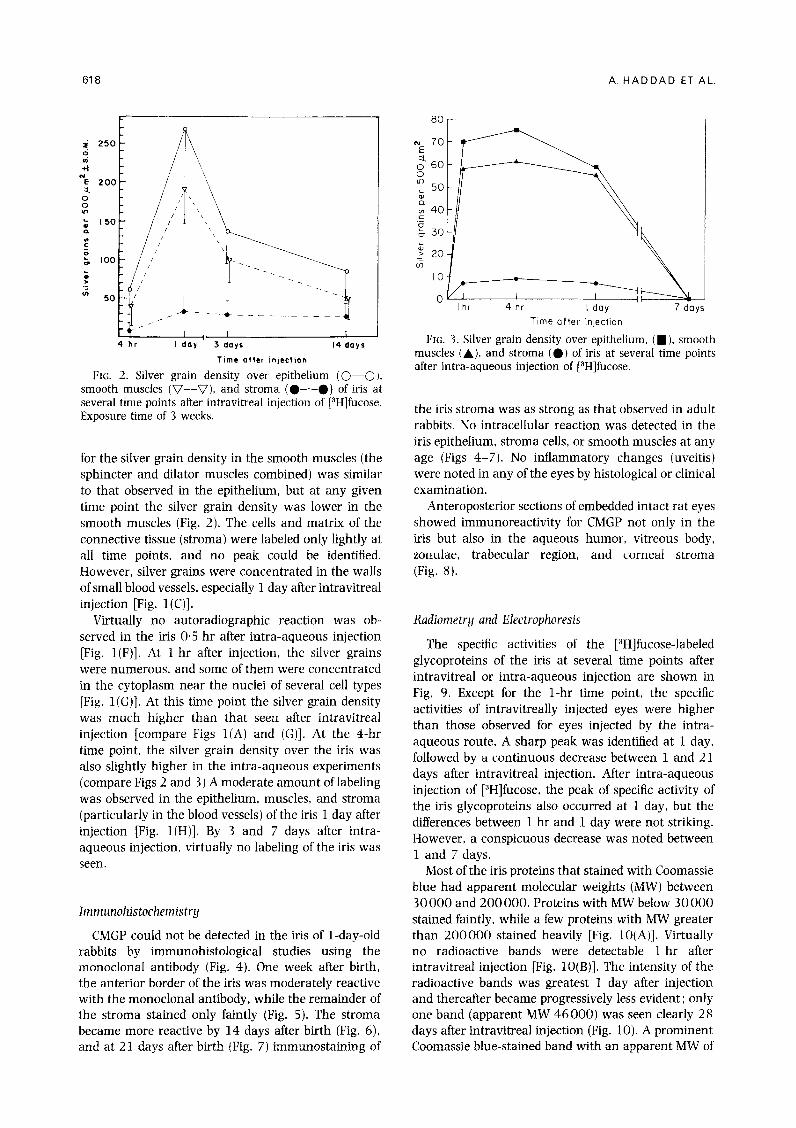
618 A. HADDAD ET AL.
4 hr ” I uoy 3 cloys 14 uays
Time offer injection
FK. 2. Silver grain density over epithelium (O-O), smooth muscles (O--V), and stroma (e-.-e) of iris at several time points after intravitreal injection of [3H]fucose. Exposure time of 3 weeks.
for the silver grain density in the smooth muscles (the sphincter and dilator muscles combined) was similar to that observed in the epithelium, but at any given time point the silver grain density was lower in the smooth muscles (Fig. 2). The cells and matrix of the connective tissue (stroma) were labeled only lightly at all time points, and no peak could be identified. However, silver grains were concentrated in the walls of small blood vessels, especially 1 day after intravitreal injection [Fig. 1 (C)l.
Virtually no autoradiographic reaction was ob- served in the iris 0.5 hr after intra-aqueous injection [Fig. l(F)]. At 1 hr after injection, the silver grains were numerous, and some of them were concentrated in the cytoplasm near the nuclei of several cell types [Fig. l(G)]. At this time point the silver grain density was much higher than that seen after intravitreal injection [compare Figs l(A) and (G)]. At the 4-hr time point, the silver grain density over the iris was also slightly higher in the intra-aqueous experiments (compare Figs 2 and 3 ) A moderate amount of labeling was observed in the epithelium, muscles, and stroma (particularly in the blood vessels) of the iris 1 day after injection [Fig. l(H)]. By 3 and 7 days after intra- aqueous injection, virtually no labeling of the iris was seen.
lrnmunohistochemistry
CMGP could not be detected in the iris of l-day-old rabbits by immunohistological studies using the monoclonal antibody (Fig. 4). One week after birth, the anterior border of the iris was moderately reactive with the monoclonal antibody, while the remainder of the stroma stained only faintly (Fig. 5). The stroma became more reactive by 14 days after birth (Fig. 6), and at 21 days after birth (Fig. 7) immunostaining of
80
“E 70
2 60
p 50
,” 40 .c 0 & 30
cii ’ 20
CT5 IO
I 0 L L
I hr 4 hr I day 7 days
Time after injection
FIG. 3. Silver grain density over epithelium. (H), smooth muscles (A), and stroma (0) of iris at several time points after intra-aqueous injection of [3H]fucose.
the iris stroma was as strong as that observed in adult rabbits. No intracellular reaction was detected in the iris epithelium, stroma cells, or smooth muscles at any age (Figs 4-7). No inflammatory changes (uveitis) were noted in any of the eyes by histological or clinical examination.
Anteroposterior sections of embedded intact rat eyes showed immunoreactivity for CMGP not only in the iris but also in the aqueous humor, vitreous body, zonulae, trabecular region, and cornea1 stroma (Fig. 8).
Radiometry and Electrophoresis
The specific activities of the [3H]fucose-labeled glycoproteins of the iris at several time points after intravitreal or intra-aqueous injection are shown in Fig. 9. Except for the l-hr time point, the specific activities of intravitreally injected eyes were higher than those observed for eyes injected by the intra- aqueous route. A sharp peak was identified at 1 day, followed by a continuous decrease between 1 and 21 days after intravitreal injection. After intra-aqueous injection of [3H]fucose. the peak of specific activity of the iris glycoproteins also occurred at 1 day, but the differences between 1 hr and 1 day were not striking. However, a conspicuous decrease was noted between 1 and 7 days.
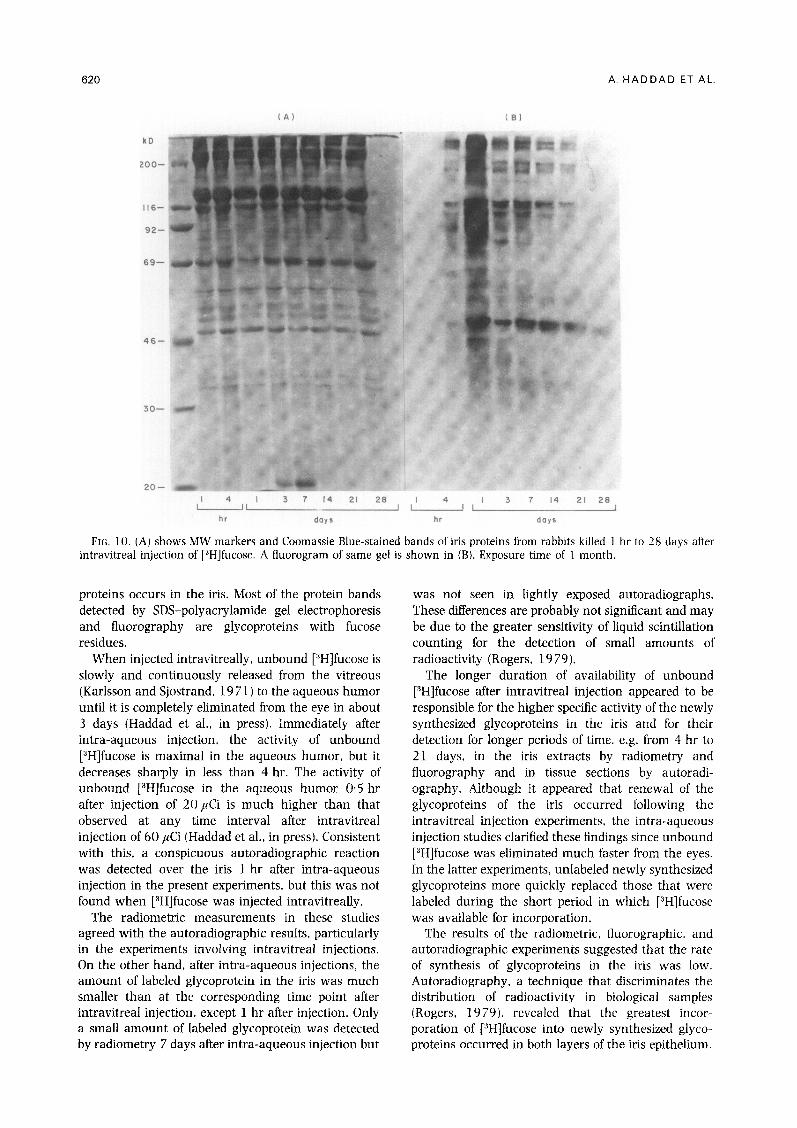
Most of the iris proteins that stained with Coomassie blue had apparent molecular weights (MW) between 30 000 and 200 000. Proteins with MW below 30 000 stained faintly, while a few proteins with MW greater than 200000 stained heavily [Fig. 10(A)]. Virtually no radioactive bands were detectable 1 hr after intravitreal injection [Fig. 10(B)]. The intensity of the radioactive bands was greatest 1 day after injection and thereafter became progressively less evident ; only one band (apparent MW 46000) was seen clearly 28 days after intravitreal injection (Fig. 10). A prominent Coomassie blue-stained band with an apparent MW of
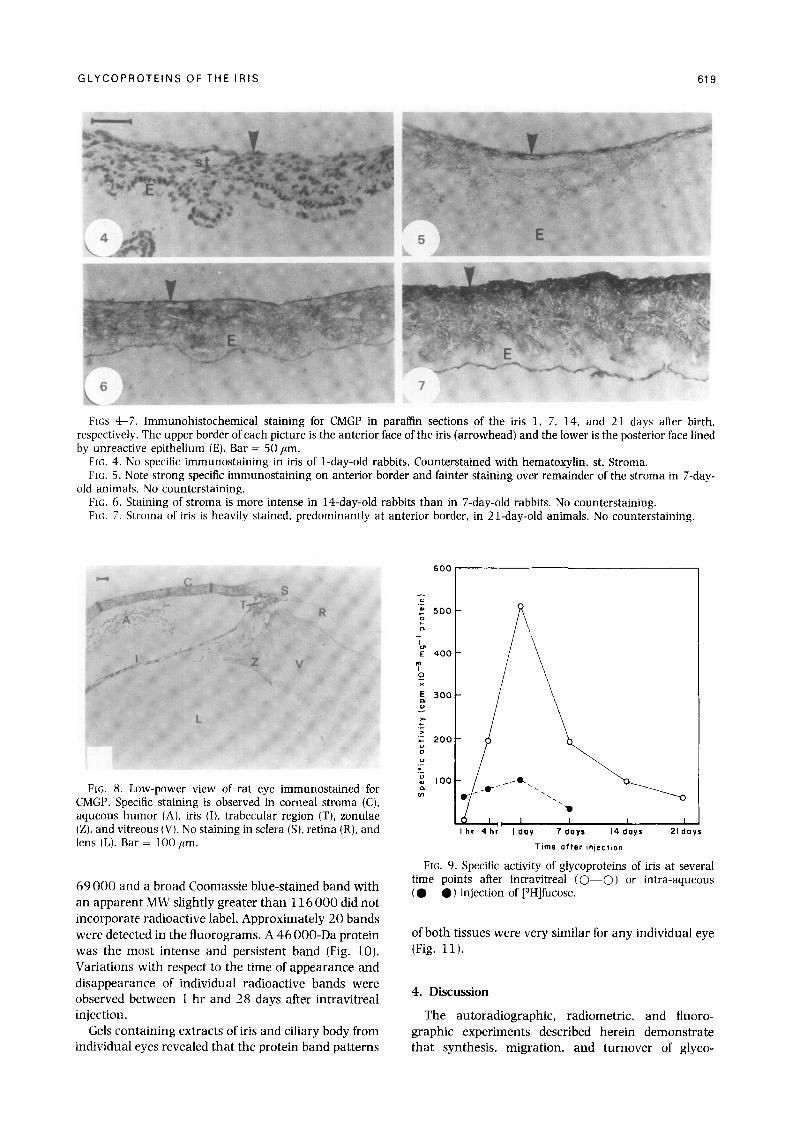
FKS 4-7. Immunohistochemical staining for CMGP in partin sections of the iris 1, 7, 14, and 21 days after birth. respectively. The upper border of each picture is the anterior face of the iris (arrowhead) and the lower is the posterior face lined by unreactive epithelium (E). Bar = 50 ,um.
Frc,. 4. No specific immunostaining in iris of l-day-old rabbits. Counterstained with hematoxylin. st. Stroma. FIG. 5. Note strong specific immunostaining on anterior border and fainter staining over remainder of the stroma in 7-day-
old animals. No counterstaining. FK. 6. Staining of stroma is more intense in l4-day-old rabbits than in 7-day-old rabbits. No counterstaining. FK;. 7. Stroma of iris is heavily stained. predominantly at anterior border, in 21-day-old animals. No counterstaining.
FIG. 8. Low-power view of rat eye immunostained for CMGP. Specific staining is observed in cornea1 stroma (C), aqueous humor (A), iris (I), trabecular region (T), zonulae (Z), and vitreous (V). No staining in sclera (S), retina (R), and lens (L). Bar = 100 /lrn.
69 000 and a broad Coomassie blue-stained band with an apparent MW slightly greater than 116 000 did not incorporate radioactive label. Approximately 20 bands were detected in the fluorograms. A 46 OOO-Da protein was the most intense and persistent band (Fig. 10). Variations with respect to the time of appearance and disappearance of individual radioactive bands were observed between 1 hr and 28 days after intravitreal injection.
Gels containing extracts of iris and ciliary body from individual eyes revealed that the protein band patterns
I hr 4hr I dOY 7 eayr 14 days 21 day.9
Time after injection
FIG. 9. Specific activity of glycoproteins of iris at several time points after intravitreal (0-O) or intra-aqueous (e--O) injection of [3H]fucose.
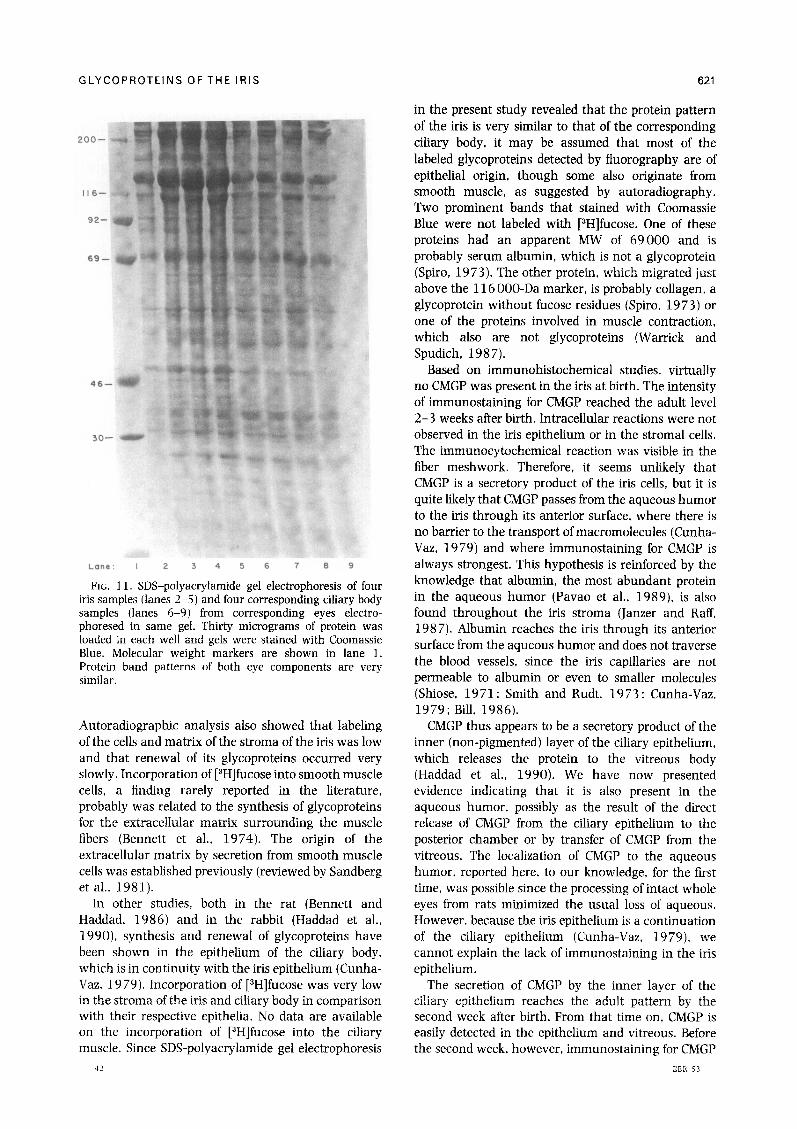
of both tissues were very similar for any individual eye (Fig. 11).
4. Discussion
The autoradiographic, radiometric, and fluoro- graphic experiments described herein demonstrate that synthesis, migration, and turnover of glyco-
620
kD
116-
92-
69-
46-
30-
A. HADDAD ET AL.
20-
,I 4 ,, I 3 7 14 21 28 , LI ll2213
hr days hr days
FIG. 10. (A) shows MW markers and Coomassie Blue-stained bands of iris proteins from rabbits killed 1 hr to 28 days after intravitreal injection of [3H]fucose. A fluorogram of same gel is shown in (B). Exposure time of 1 month.
proteins occurs in the iris. Most of the protein bands detected by SDS-polyacrylamide gel electrophoresis and fluorography are glycoproteins with fucose residues.
When injected intravitreally, unbound [“Hlfucose is slowly and continuously released from the vitreous (Karlsson and Sjostrand, 19 7 1) to the aqueous humor until it is completely eliminated from the eye in about 3 days (Haddad et al., in press). Immediately after intra-aqueous injection, the activity of unbound [3H]fucose is maximal in the aqueous humor, but it decreases sharply in less than 4 hr. The activity of unbound [3H]fucose in the aqueous humor 0 5 hr after injection of 20 ,&i is much higher than that observed at any time interval after intravitreal injection of 60 ,uCi (Haddad et al., in press). Consistent with this, a conspicuous autoradiographic reaction was detected over the iris 1 hr after intra-aqueous injection in the present experiments, but this was not found when [3H]fucose was injected intravitreally.
The radiometric measurements in these studies agreed with the autoradiographic results, particularly in the experiments involving intravitreal injections. On the other hand, after intra-aqueous injections, the amount of labeled glycoprotein in the iris was much smaller than at the corresponding time point after intravitreal injection. except 1 hr after injection. Only a small amount of labeled glycoprotein was detected by radiometry 7 days after intra-aqueous injection but
was not seen in lightly exposed autoradiographs. These differences are probably not significant and may be due to the greater sensitivity of liquid scintillation counting for the detection of small amounts of radioactivity (Rogers, 1979).
The longer duration of availability of unbound [3H]fucose after intravitreal injection appeared to be responsible for the higher specific activity of the newly synthesized glycoproteins in the iris and for their detection for longer periods of time. e.g. from 4 hr to 21 days, in the iris extracts by radiometry and fluorography and in tissue sections by autoradi- ography. Although it appeared that renewal of the glycoproteins of the iris occurred following the intravitreal injection experiments, the intra-aqueous injection studies clarified these findings since unbound [3H]fucose was eliminated much faster from the eyes. In the latter experiments, unlabeled newly synthesized glycoproteins more quickly replaced those that were labeled during the short period in which [“Hlfucose was available for incorporation.
The results of the radiometric, fluorographic. and autoradiographic experiments suggested that the rate of synthesis of glycoproteins in the iris was low. Autoradiography, a technique that discriminates the distribution of radioactivity in biological samples (Rogers, 1979). revealed that the greatest incor- poration of [“Hlfucose into newly synthesized glyco- proteins occurred in both layers of the iris epithelium.
GLYCOPROTEINS OF THE IRIS 621
I l6-
92-
69-
46.
30.
:;i’
Lane: I 2 3 4 5 6 7 6 9
FIG. Il. SDS-polyacrylamide gel electrophoresis of four iris samples (lanes 2-5) and four corresponding ciliary body samples (lanes 6-9) from corresponding eyes electro- phoresed in same gel. Thirty micrograms of protein was loaded in each well and gels were stained with Coomassie Blue. Molecular weight markers are shown in lane 1. Protein band patterns of both eye components are very similar.
Autoradiographic analysis also showed that labeling of the cells and matrix of the stroma of the iris was low and that renewal of its glycoproteins occurred very slowly. Incorporation of [3H]fucose into smooth muscle cells, a linding rarely reported in the literature, probably was related to the synthesis of glycoproteins for the extracellular matrix surrounding the muscle fibers (Bennett et al., 19 74). The origin of the extracellular matrix by secretion from smooth muscle cells was established previously (reviewed by Sandberg et al., 1981).
In other studies, both in the rat (Bennett and Haddad, 1986) and in the rabbit (Haddad et al., lVVO), synthesis and renewal of glycoproteins have been shown in the epithelium of the ciliary body, which is in continuity with the iris epithelium (Cunha- Vaz, 1979). Incorporation of [3H]fucose was very low in the stroma of the iris and ciliary body in comparison with their respective epithelia. No data are available on the incorporation of [3H]fucose into the ciliary muscle. Since SDS-polyacrylamide gel electrophoresis
in the present study revealed that the protein pattern of the iris is very similar to that of the corresponding ciliary body, it may be assumed that most of the labeled glycoproteins detected by fluorography are of epithelial origin, though some also originate from smooth muscle, as suggested by autoradiography. Two prominent bands that stained with Coomassie Blue were not labeled with [3H]fucose. One of these proteins had an apparent MW of 69000 and is probably serum albumin, which is not a glycoprotein (Spiro, 1973). The other protein, which migrated just above the 116 OOO-Da marker, is probably collagen, a glycoprotein without fucose residues (Spiro, 19 73) or one of the proteins involved in muscle contraction, which also are not glycoproteins (Warrick and Spudich, 1987).
Based on immunohistochemical studies, virtually no CMGP was present in the iris at birth. The intensity of immunostaining for CMGP reached the adult level 2-3 weeks after birth. Intracellular reactions were not observed in the iris epithelium or in the stromal cells. The immunocytochemical reaction was visible in the fiber meshwork. Therefore, it seems unlikely that CMGP is a secretory product of the iris cells, but it is quite likely that CMGP passes from the aqueous humor to the iris through its anterior surface, where there is no barrier to the transport of macromolecules (Cunha- Vaz, 1979) and where immunostaining for CMGP is always strongest. This hypothesis is reinforced by the knowledge that albumin, the most abundant protein in the aqueous humor (Pavao et al., 1989), is also found throughout the iris stroma (Janzer and Raff, 1987). Albumin reaches the iris through its anterior surface from the aqueous humor and does not traverse the blood vessels, since the iris capillaries are not permeable to albumin or even to smaller molecules (Shiose, 1971; Smith and Rudt, 1973 : Cunha-Vaz, 1979; Bill, 1986).
CMGP thus appears to be a secretory product of the inner (non-pigmented) layer of the ciliary epithelium, which releases the protein to the vitreous body (Haddad et al., 1990). We have now presented evidence indicating that it is also present in the aqueous humor, possibly as the result of the direct release of CMGP from the ciliary epithelium to the posterior chamber or by transfer of CMGP from the vitreous. The localization of CMGP to the aqueous humor, reported here, to our knowledge, for the first time, was possible since the processing of intact whole eyes from rats minimized the usual loss of aqueous. However, because the iris epithelium is a continuation of the ciliary epithelium (Cunha-Vaz, 1979), we cannot explain the lack of immunostaining in the iris epithelium.
The secretion of CMGP by the inner layer of the ciliary epithelium reaches the adult pattern by the second week after birth. From that time on, CMGP is easily detected in the epithelium and vitreous. Before the second week, however, immunostaining for CMGP
EER 53

622 A. HADDAD ET AL.
is minimal in all of the eye components that are usually heavily stained in adult animals (Andre, Haddad and Fife, unpubl. res.). Since staining of the iris begins at the anterior surface and progressively reaches the posterior regions during the first 3 weeks of post-natal life and since no immunostaining for CMGP is detected intracellularly in any of the cells of the iris, we conclude that CMGP is not an intrinsic iris component.
Acknowledgements
The authors would like to thank Mara S. A. Costa, Vani M. A. Correa, Cirlei A. Vanni, and Kelly D. King for their technical assistance and Roberta Fehrman for her secretarial assistance. This work was supported in part by Funda@o de Amparo & Pesquisa do Estado de SBo Paulo (A.H.). the BID/LISP Project (A. H.), CNPq (A.H. and E.M.L.). the Medical Research Council of Canada (G.P.), NIH grant AR 39250 (R.S.F.), and by a Biomedical Research Grant from the Arthritis Foundation (R. S.F.).
References
Bennett, G. and Haddad, A. (1986). Synthesis and migration of 3H-fucose-labeled glycoproteins in the ciliary epi- thelium of the eye: effects of microtubule-disrupting drugs. Am. I. Anat. 177. 441-55.
Bennett, G., Leblond, C. P. and Haddad. A. (1974). Migration of glycoprotein from the Golgi apparatus to the surface of various cell types as shown by radioautography after labeled fucose injection into rats. J. Cell Biol. 60, 258-84.
Bill, A. (1986). The blood-aqueous barrier. Trans. Ophthalmol. Sot. U.K. 105. 149-55.
Cunha-Vaz, J. (1979). The blood+cular barriers. Surv. Ophthalmol. 23, 279-96.
Fife, R. S. and Brandt, K. D. (1989). Cartilage matrix glycoprotein is present in the serum of dogs with experimental osteoarthritis. 1. CJin. Invest. 84. 1432-9.
Haddad, A., de Almeida, J. C., Laicine, E. M.. Fife, R. S. and Pelletier, G. (1990). The origin of the intrinsic glyco- proteins of the rabbit vitreous body: an immunohisto- chemical and autoradiographic study. Exp. Eye Res. SO. 555-61.
Haddad. A., Laicine, E. M. and de Almeida. J. C. ( 199 1). Origin and renewal of the intrinsic glycoproteins of the aqueous humor. Graefes Arch. Clin. Exp. Ophthalmol. (in press).
Janzer, R. C. and Raff, M. C. (1987). Astrocytes induce blood-brain barrier properties in endothelial cells. Wature 325, 253-7.
Karlsson, J.-O. and Sjostrand, J. (1971). Rapid intracellular transport of fucose-containing glycoproteins in retinal ganglion cells. J. Neurochem. 18. 2209-16.
Laemmli, U. K. (1970). Cleavage of the structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-5.
Lowry, 0. H., Rosebrough. N. J., Farr, A. L. and Randall, K. J. ( 19 5 1). Protein measurements with the Folin phenol reagent. 1. Biol. Chem. 193, 265-75.
Nguyen, B. Q. and Fife, R. S. (1986). Vitreous contains a cartilage-related protein. Exp. Eye Res. 43, 3 75-82.
Ozanics, V. and Jakobiec. F. A. (1982). Prenatal development of the eye and its adnexa. In Ocular Anatomy. Em- bryology and Teratology. (Ed. Jakobiec. F. A.) Pp. 11 -Y 6. Harper and Row : Philadelphia.
Pavao, A. F.. Lee, D. A., Ethier. C. R.. Johnson. M. C’.. Anderson, P. J. and Epstein, D. I,. (1989). Two- dimensional gel electrophoresis of calf aqueous humor. serum, and filter-bound proteins. Invest. Ophthabnol. Vis. Sci. 30, 731-8.
Rodrigues. M. M., Hackett. J. and Donohoo. P. (1982). In Ocular Anatomy, Embryology and Teratology. (Ed. Jakobiec, F. A.) Pp. 285-302. Harper and Row: Philadelphia.
Rogers, A. W. (1979). Techniques of Autorudiogruphy. 3rd edn. Elsevier/North Holland Biomedical Press : Amster- dam.
Sandberg, L. B.. Soskel. N. T. and Leslie, J. G. ( 1 Y 8 1 ). Elastin structure. biosynthesis, and relation to disease states. New England J. Med. 304, 566-79.
Schachter. H. and Roden. L. (1973). The biosynthesis of animal glycoproteins. In Metnbolir Conjugation and Metabolic Hydrolysis. (Ed. Fishman. W. H.i Pp. l-149. Academic Press: New York.
Shiose, Y. (1971). Morphological study on permeability of the blood-aqueous barrier. Jpn. /. Ophthalmol. 15. 17-26.
Smith, R. S. and Rudt, L. A. (1975). Ocular vascular and epithelial barriers to microperoxidase. Invest. Ophthnlmol. 14, 556-W.
Spiro, R. G. ( 1973 ). Glycoproteins. Adv. J'rotrin Chem. 27. 349-467.
Streeten, B. W. (1982). Ciliary body. In Oculur Anatomy. Embryology and Teratology. (Ed. Jakobiec. F. A.) Pp. 303-30. Harper and Row: Philadelphia.
Warrick, H. M. and Spudich, J. A. (1987). Myosin structure and function in cell motility. Ann. Rev. CeJI. Biol. 3, 379-421.
Waterborg, J, H. and Matthews. H. R. (1984). Fluorography of polyacrylamide gels containing tritium. In Methods in Molecular Biology. Vol. 7 : Proteins. (Ed. Walker. J. M.l Pp. 147-52. Humana Press: Clifton, NJ.





























