Embed Size (px)
Citation preview

www.elsevier.com/locate/brainres
Brain Research 1032
Research report
Avoidance behavior and brain monoamines in fish
Erik Hfglunda, Finn-Arne Weltziena,b, Joachim Schjoldenc, Svante Winbergc,
Holger Ursind, Kjell B. Døvinga,*
aDepartment of Molecular Biosciences, University of Oslo, PO Box 1041 Blindern, 0316 Oslo, NorwaybUSM 0401, UMR 5178 CNRS, Biologie des Organismes Marins et Ecosystemes, Departement des Milieux et Peuplements Aquatiques,
Museum National d’Histoire Naturelle, 7 rue Cuvier, 75231 Paris Cedex 05, FrancecEvolutionary Biology Centre, Department of Comparative Physiology, Uppsala University, Norbyvagen 18 A, SE-75236 Uppsala, Sweden
dDepartment of Biological and Medical Psychology, University of Bergen, Jonas Lies vei 91, 5009 Bergen, Norway
Accepted 30 October 2004
Available online 9 December 2004
Abstract
The crucian carp performs a typical avoidance behavior when exposed to olfactory cues from injured skin of conspecifics. They swim
rapidly to the bottom and hide in available material. This work examines the effects of skin extract exposure and availability of hiding
material on this behavior, and concomitant changes in brain monoaminergic activity in crucian carp.
Individual fish were exposed to skin extract in aquaria with or without hiding material. Exposure to skin extract resulted in the expected
avoidance behavior consisting of rapid movement towards the bottom of the aquarium. This lasted for 1–2 min. Activity then decreased
below the level observed before exposure, suggesting a bfreezingQ type of avoidance behavior. This behavior was independent of availabilityof hiding material.
Brain dopaminergic activity increased in telencephalon and decreased in the brain stem following skin extract exposure, again
independent of availability of hiding material. However, fish kept in aquaria without hiding material showed an elevation of serotonergic
activity in the brain stem and the optic tectum compared to fish with available hiding material. Absence of hiding material increased
serotonergic activity also without exposure to skin extract. In aquaria with hiding material, the fish stirred up a cloud of fine sediments and
showed a more pronounced decrease in locomotor activity in agreement with this being a more efficient freezing or immobile avoidance
behavior. These results show that basic components of avoidance behavior and related brain changes are present in the fish brain, in
accordance with the common phylogenetic roots of avoidance behavior in all vertebrates.
D 2004 Elsevier B.V. All rights reserved.
Theme: Neural basis of behavior
Topic: Monoamines and behavior
Keywords: Fear behavior; Teleost; Monoamine; Predator; Refuge; Olfactory cues
1. Introduction
The crucian carp (Carassius carassius) perform stereo-
typic avoidance behavior when exposed to olfactory cues
from injured skin of conspecifics. They rapidly seek the
bottom (bflightQ), stir up a cloud of fine sediment and hide
0006-8993/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.brainres.2004.10.050
* Corresponding author. Fax: +47 22 85 46 64.
E-mail address: [email protected] (K.B. Døving).
(bfreezeQ) in or on the material available at the bottom [6].
Similar predator-avoidance behavior is elicited by exposure
to olfactory cues from injured skin of conspecifics (i.e. skin
extract) in other fish as well, for instance in the rainbow trout
(Oncorhynchus mykiss) [20]. Direct exposure to predators
involves changes in monoamine signaling as well as behavior
in teleost fish (e.g. Ref. [26]), as it does in mammals [2,14].
Availability of a refuge during predator exposure affects
both the behavior and physiological stress responses in the
long nose killifish (Fundulus majalis) [27]. This implies that
(2005) 104–110

E. Hoglund et al. / Brain Research 1032 (2005) 104–110 105
availability of hiding material interacts with stimuli signal-
ing predators and injury, suggesting an interaction between
stress stimuli and context that in mammals is taken as an
indication of cognitive factors [24]. In this paper, we
examine whether availability of hiding material affects the
behavior as well as the brain monoamine activity also in the
crucian carp. There are two components of the apparently
innate avoidance behavior. Following the nomenclature for
avoidance behavior and limbic mechanisms in mammals
(see Ref. [23]); the active movements (bflightQ) qualify as
active avoidance, and the immobility in the hiding material
(bfreezingQ) as passive avoidance. These innate behaviors
are goal directed behaviors, and, therefore, represent
potential binstrumentalQ behaviors [19]. When instrumental
behavior is well established, the vegetative, endocrine, and
immunological response to the situation diminishes in
mammals (bstress reductionQ or coping [13,23,24]). This is
also documented for by changes in brain monoamines [24].
In mice [1] and rats [14], limbic monoaminergic signal-
ing is affected by direct exposure to a predator (cat and
ferret). Direct predator exposure affects monoaminergic
signaling also in teleost fish like the bicolor damselfish
(Pomacentrus parties) [26], rainbow trout [12,16], and
Arctic charr (Salvelinus alpinus) [8,9].
If the presence or absence of hiding materials that offer
avoidance possibilities affects the behavior of the crucian
carp, it may also affect the brain monoamines. Our
hypothesis, therefore, is that availability of hiding material
may affect avoidance behavior as well as monoamine
signaling in the brain much in the same way as in other
vertebrates including mammals.
2. Materials and methods
2.1. Animals
Crucian carp (C. carassius) were caught in a small lake
outside of Oslo, Norway (608N), in June 2002. The fish
were kept indoors at the Department of Molecular Bio-
sciences, University of Oslo, in 700 L tanks continuously
supplied with aerated water at 7–9 8C, and with a photo
cycle of 12/12-h (light/dark). The fish were fed once a day
with TetraR pond sticks (Tetra, Germany). The experiment
was performed in February–March 2003 with fish (37.5%
males and 62.5% females) with a mean body weight of
29F7.5 g (meanFstandard deviation).
2.2. Preparation of skin extract
For preparation of the skin extract used in the experi-
ments, crucian carp were killed by decapitation and skin was
taken from the sides of the fish. Approximately 2 g of skin
was homogenized in 100 mL distilled water with pestle and
mortar. The homogenate was centrifuged at 2400 rpm for 5
min at 4 8C. The supernatant was frozen immediately and a
working concentration of 1:10 in pond water was made
fresh before use.
2.3. Experimental protocol
Two days before the behavioral observations, six fish
were transferred to six visually separated 40 L observation
aquaria. In three of the observation aquaria, about half of the
bottom floor was covered by an approximately 4 cm thick
layer of fine sediments taken from the lake where the fish
were caught. In the other three observation aquaria the
bottom floor consisted of uncovered glass.
After 2 days of acclimatization, each individual fish was
video filmed for 30 min, during which they received two 5
mL injections of either skin extract or aquarium water
(vehicle control). The first injection was given after 10 min,
followed by a second addition after 20 min to ensure
exposure of the fish to stable levels of alarm substance. To
minimize disturbance during the behavioral studies, the fish
were filmed through a hole in a black curtain covering the
front of the aquaria, and skin extract was delivered to the
aquarium water through a plastic tube from outside the
covering curtain. The video filmed fish were analyzed for
locomotor activity and position in the water tank. Locomo-
tor activity was quantified throughout the 30-min observa-
tional period as the distance the fish had moved during 15 s
periods. The position in the water tank was measured every
15th second, throughout the 30-min observational period, as
the distance from the bottom floor in the aquaria. A total of
14 fish were exposed to skin extract, 7 with hiding material
and 7 without. Ten fish, five with hiding material and five
without, received aquarium water instead of skin extract and
served as controls. All experiments were conducted at room
temperature in the light period of the 12/12-h photo cycle.
2.4. Brain tissue sampling and analysis of brain
monoaminergic activity
Directly after the 30-min observation period, the fish
were netted and rapidly killed by severing the spinal cord
just behind the head of the fish. The brain was then removed
and dissected into four regions: brain stem (including
medulla and part of the spinal cord but excluding
cerebellum and the vagal lobes), optic tectum, hypothal-
amus (di- and mesencephalon except optic tectum), and
telencephalon (excluding the olfactory bulbs). Each brain
part was immediately wrapped in aluminum foil, frozen in
liquid nitrogen, and stored at �80 8C.Upon HPLC analyses, the frozen brain parts were
homogenized in 4% (w/v) ice-cold perchloric acid contain-
ing 0.2% EDTA and 40 ng/mL epinine (deoxyadrenaline;
used as internal standard in the HPLC analyses). Homog-
enization was performed using either a Potter-Elvehjem
homogenizer (optic tectum and brain stem), or a MSE 100 W
ultrasonic disintegrator (telencephalon and hypothalamus).
Serotonin (5-HT), 5-hydroxyindoleacetic acid (5-HIAA, the
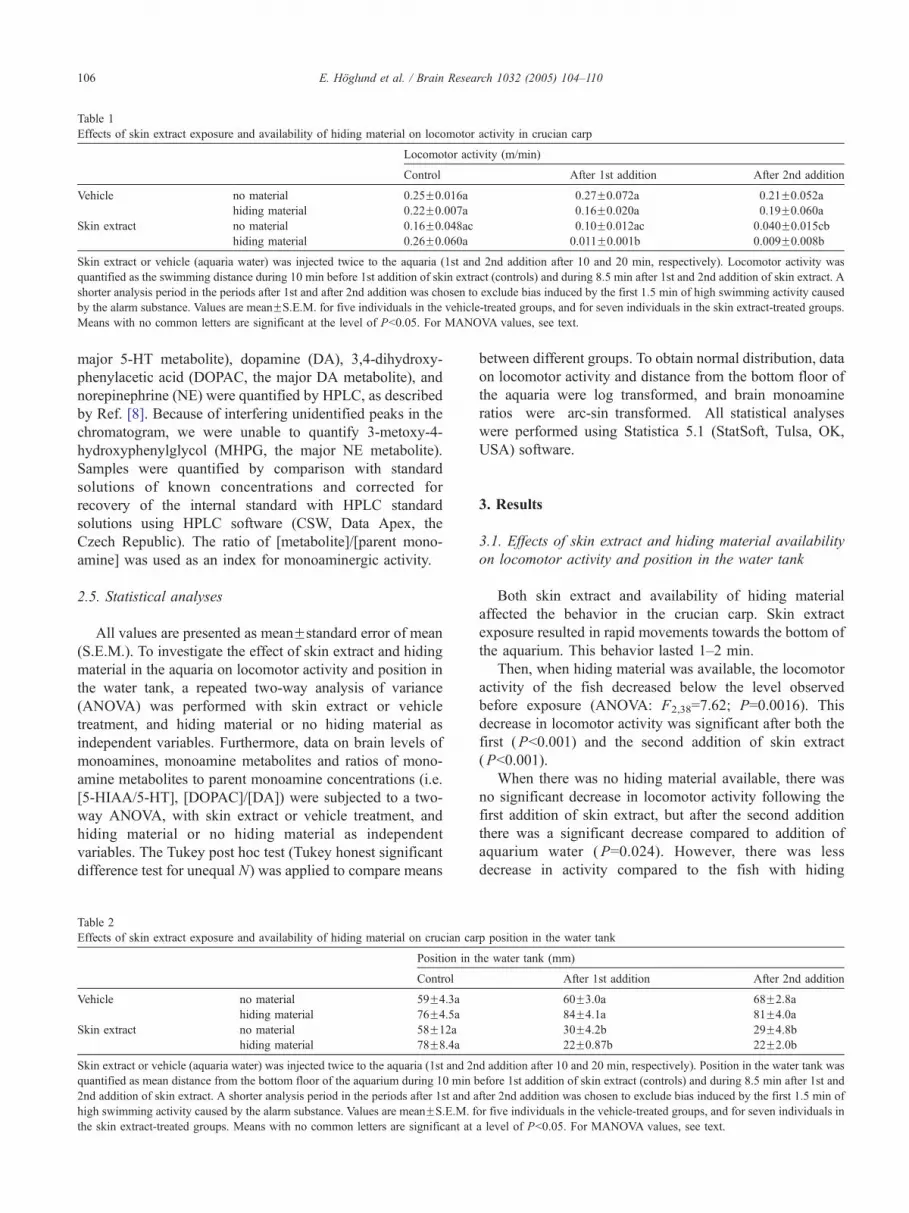
Table 1
Effects of skin extract exposure and availability of hiding material on locomotor activity in crucian carp
Locomotor activity (m/min)
Control After 1st addition After 2nd addition
Vehicle no material 0.25F0.016a 0.27F0.072a 0.21F0.052a
hiding material 0.22F0.007a 0.16F0.020a 0.19F0.060a
Skin extract no material 0.16F0.048ac 0.10F0.012ac 0.040F0.015cb
hiding material 0.26F0.060a 0.011F0.001b 0.009F0.008b
Skin extract or vehicle (aquaria water) was injected twice to the aquaria (1st and 2nd addition after 10 and 20 min, respectively). Locomotor activity was
quantified as the swimming distance during 10 min before 1st addition of skin extract (controls) and during 8.5 min after 1st and 2nd addition of skin extract. A
shorter analysis period in the periods after 1st and after 2nd addition was chosen to exclude bias induced by the first 1.5 min of high swimming activity caused
by the alarm substance. Values are meanFS.E.M. for five individuals in the vehicle-treated groups, and for seven individuals in the skin extract-treated groups
Means with no common letters are significant at the level of Pb0.05. For MANOVA values, see text.
E. Hoglund et al. / Brain Research 1032 (2005) 104–110106
major 5-HT metabolite), dopamine (DA), 3,4-dihydroxy-
phenylacetic acid (DOPAC, the major DA metabolite), and
norepinephrine (NE) were quantified by HPLC, as described
by Ref. [8]. Because of interfering unidentified peaks in the
chromatogram, we were unable to quantify 3-metoxy-4-
hydroxyphenylglycol (MHPG, the major NE metabolite).
Samples were quantified by comparison with standard
solutions of known concentrations and corrected for
recovery of the internal standard with HPLC standard
solutions using HPLC software (CSW, Data Apex, the
Czech Republic). The ratio of [metabolite]/[parent mono-
amine] was used as an index for monoaminergic activity.
2.5. Statistical analyses
All values are presented as meanFstandard error of mean
(S.E.M.). To investigate the effect of skin extract and hiding
material in the aquaria on locomotor activity and position in
the water tank, a repeated two-way analysis of variance
(ANOVA) was performed with skin extract or vehicle
treatment, and hiding material or no hiding material as
independent variables. Furthermore, data on brain levels of
monoamines, monoamine metabolites and ratios of mono-
amine metabolites to parent monoamine concentrations (i.e.
[5-HIAA/5-HT], [DOPAC]/[DA]) were subjected to a two-
way ANOVA, with skin extract or vehicle treatment, and
hiding material or no hiding material as independent
variables. The Tukey post hoc test (Tukey honest significant
difference test for unequal N) was applied to compare means
Table 2
Effects of skin extract exposure and availability of hiding material on crucian carp position in the water tank
Position in the water tank (mm)
Control After 1st addition After 2nd addition
Vehicle no material 59F4.3a 60F3.0a 68F2.8a
hiding material 76F4.5a 84F4.1a 81F4.0a
Skin extract no material 58F12a 30F4.2b 29F4.8b
hiding material 78F8.4a 22F0.87b 22F2.0b
Skin extract or vehicle (aquaria water) was injected twice to the aquaria (1st and 2nd addition after 10 and 20 min, respectively). Position in the water tank was
quantified as mean distance from the bottom floor of the aquarium during 10 min before 1st addition of skin extract (controls) and during 8.5 min after 1st and
2nd addition of skin extract. A shorter analysis period in the periods after 1st and after 2nd addition was chosen to exclude bias induced by the first 1.5 min o
high swimming activity caused by the alarm substance. Values are meanFS.E.M. for five individuals in the vehicle-treated groups, and for seven individuals in
the skin extract-treated groups. Means with no common letters are significant at a level of Pb0.05. For MANOVA values, see text.
.
between different groups. To obtain normal distribution, data
on locomotor activity and distance from the bottom floor of
the aquaria were log transformed, and brain monoamine
ratios were arc-sin transformed. All statistical analyses
were performed using Statistica 5.1 (StatSoft, Tulsa, OK,
USA) software.
3. Results
3.1. Effects of skin extract and hiding material availability
on locomotor activity and position in the water tank
Both skin extract and availability of hiding material
affected the behavior in the crucian carp. Skin extract
exposure resulted in rapid movements towards the bottom of
the aquarium. This behavior lasted 1–2 min.
Then, when hiding material was available, the locomotor
activity of the fish decreased below the level observed
before exposure (ANOVA: F2,38=7.62; P=0.0016). This
decrease in locomotor activity was significant after both the
first (Pb0.001) and the second addition of skin extract
(Pb0.001).
When there was no hiding material available, there was
no significant decrease in locomotor activity following the
first addition of skin extract, but after the second addition
there was a significant decrease compared to addition of
aquarium water (P=0.024). However, there was less
decrease in activity compared to the fish with hiding
f

Fig. 1. Effects of skin extract exposure on [DOPAC]/[DA] in the telencephalon (A), brain stem (B), hypothalamus (C) and optic tectum (D) of crucian carp. The
values are presented as meanFS.E.M. * indicates significant difference at a level of Pb0.05. See Table 3 for ANOVA values.
E. Hoglund et al. / Brain Research 1032 (2005) 104–110 107
material available (first addition, P=0.0015; Table 1).
Availability of hiding material did not affect locomotor
activity before addition of skin extract (or water) in any of
the two groups (Table 1).
The two-way repeated measurement MANOVA indi-
cated a combined effect of hiding material and skin extract
on position in the water tank (ANOVA: F2,38=3.40,
Fig. 2. Effects of hiding material availability on [5-HIAA]/[5-HT] in the telenceph
carp. The values are presented as meanFS.E.M. * indicates significant difference
P=0.043). The post hoc test, however, failed to show any
significant effects. Skin extract exposure resulted in a
position lower in the water tank, both after first (hiding
material, Pb0.001; no hiding material, P=0.009) and second
addition of skin extract (hiding material, Pb0.001; no hiding
material, P=0.004), with no significant difference in water
tank position between fish with or without available hiding
alon (A), brain stem (B), hypothalamus (C) and optic tectum (D) of crucian
at a level of Pb0.05. See Table 3 for ANOVA values.

Table 3
Effects of skin extract exposure and availability of hiding material on the concentration of monoamines and monoamine metabolites, and on the ratio bet een monoamine metabolite to parent monoamine in
crucian carp
Effect Skin extract Substrate Skin extract and substrate
ANOVA ANOVA ANOVA No substrate Substrate
Skin extract ehicle Skin extract Vehicle
Hypothalamus
[DOPAC] F(1,20)=0.174, P=0.681 F(1,20)=1.68, P=0.209 F(1,20)=4.09, P=0.057 8.80F2.10 5.43F2.58 7.28F1.68 12.3F2.27
[DA] F(1,20)=0.077, P=0.783 F(1,20)=0.026, P=0.871 F(1,20)=0.181, P=0.675 2390F244 165F469 2203F286 2260F460
10�3�[DOPAC]/[DA] F(1,20)=0.001, P=0.969 F(1,20)=2.18, P=0.155 F(1,20)=1.46, P=0.240 3.79F0.96 2.27F0.74 4.12F1.51 5.73F1.19
[5-HIAA] F(1,20)=0.587, P=0.452 F(1,20)=1.13, P=0.300 F(1,20)=0.230, P=0.636 166F34.9 206F59.9 166F6.77 156F25.3
[5-HT] F(1,20)=0.021, P=0.884 F(1,20)=0.0003, P=0.986 F(1,20)=0.046, P=0.831 2910F244 790F552 2840F260 2859F238
10�3�[5-HIAA]/[5-HT] F(1,20)=0.738, P=0.400 F(1,20)=0.848, P=0.368 F(1,20)=1.03, P=0.321 54.7F5.92 70.9F14.3 55.6F3.36 54.2F6.16
[NA] F(1,18)=1.40, P=0.250 F(1,20)=0.658, P=0.658 F(1,20)=0.0126, P=0.912 3530F412 090F547 3390F547 2860F534
Optic tectum
[DOPAC] F(1,19)=0.191, P=0.667 F(1,19)=0.0578, P=0.813 F(1,19)=2.53, P=0.128 4.26F0.681 .41F0.91 3.60F0.83 5.22F0.735
[DA] F(1,19)=0.833, P=0.372 F(1,19)=1.13, P=0.813 F(1,19)=0.366, P=0.850 124F33.8 154F46.0 155F46.0 105F8.5
10�3�[DOPAC]/[DA] F(1,19)=0.0001, P=0.989 F(1,19)=1.06, P=0.316 F(1,19)=2.47, P=0.133 42.5F7.72 0.4F9.61 38.3F6.21 50.2F6.70
[5-HIAA] F(1,19)=0.120, P=0.733 F(1,19)=3.46, P=0.0714 F(1,19)=0.305, P=0.862 36.1F3.90 7.0F4.54 27.4F4.52 26.3F2.09
[5-HT] F(1,19)=0.0279, P=0.869 F(1,19)=0.551, P=0.467 F(1,19)=0.120, P=0.733 234F51.1 220F51.2 203F43.6 190F12.1
10�3�[5-HIAA]/[5-HT] F(1,19)=0.0876, P=0.770 F(1,19)=6.77, P=0.0175 F(1,19)=0.398, P=0.535 167F13.4 178F15.4 142F10.8 138F7.10
[NA] F(1,19)=0.224, P=0.642 F(1,19)=0.215, P=0.647 F(1,19)=0.0836, P=0.775 791F62.7 856F189 778F130 793F79.3
Brain stem
[DOPAC] F(1,18)=6.13, P=0.0234 F(1,18)=2.22, P=0.15 F(1,18)=1.01, P=0.326 8.20F2.79 0.0F2.27 3.92F0.54 14.5F5.52
[DA] F(1,18)=1.65, P=0.214 F(1,18)=0.922, P=0.349 F(1,18)=1.08, P=0.310 270F46.5 178F35.0 181F17.0 279F75.5
10�3�[DOPAC]/[DA] F(1,18)=5.07, P=0.0370 F(1,18)=1.65, P=0.214 F(1,18)=2.07, P=0.167 27.7F4.25 3.4F26.5 21.4F2.14 40.6F9.83
[5-HIAA] F(1,19)=0.0937, P=0.763 F(1,19)=3.65, P=0.0711 F(1,19)=0.0150, P=0.904 73.9F12.9 1.1F11.5 46.4F4.38 50.5F15.5
[5-HT] F(1,19)=0.103, P=0.75 F(1,19)=0.819, P=0.379 F(1,19)=4.12, P=0.0567 437F68.2 455F72.9 318F33.0 404F92.0
10�3�[5-HIAA]/[5-HT] F(1,19)=2.44, P=0.134 F(1,19)=4.47, P=0.0479 F(1,19)=0.593, P=0.464 168F7.86 147F6.40 155F20.2 95.7F22.9
[NA] nd nd nd nd
Telencephalon
[DOPAC] F(1,19)=13.3, P=0.0017 F(1,19)=0.0604, P=0.808 F(1,19)=0.0663, P=0.799 23.4F4.61 1.74F7.65 24.5F6.34 6.77F1.98
[DA] F(1,19)=1.67, P=0.211 F(1,19)=0.172, P=0.682 F(1,19)=0.0198, P=0.682 674F120 569F113 637F100 459F70.0
10�3�[DOPAC]/[DA] F(1,19)=4.48, P=0.0402 F(1,19)=00009, P=0.975 F(1,19)=0114, P=0.916 41.0F11.5 18.9F12.1 41.8F9.66 17.4F7.40
[5-HIAA] F(1,19)=1.06, P=0.314 F(1,19)=0.691, P=0.416 F(1,19)=0.0229, P=0.881 342F110 272F96.2 213F25.2 152F24.7
[5-HT] F(1,19)=0.936, P=0.345 F(1,19)=0.0375, P=0.848 F(1,19)=0.772, P=0.129 2540F848 2430F800 2210F284 1460F146
10�3�[5-HIAA]/[5-HT] F(1,19)=2.19, P=0.154 F(1,19)=0.201, P=0.658 F(1,19)=0.382, P=0.554 136F28.8 118F13.2 99.3F8.09 102F10.3
[NA] F(1,19)=1.87, P=0.187 F(1,19)=0.0238, P=0.879 F(1,19)=0.115, P=0.738 6200F1010 5630F1320 6375F942 4650F587
Five milliliters of either skin extract or vehicle (aquaria water) was injected twice to the aquaria (after 10 and 20 min, respectively; see Materials and methods r details). Fourteen fish were exposed to skin extract,
seven with and seven without hiding material available. Ten fish, five with and five without hiding material available, received aquaria water and served as v icle treated controls. Values are meanFS.E.M. F, df
and P values are results of ANOVA (significant results are indicated in bold). DA, Dopamine; DOPAC, 3,4-dihydroxyphenylacetic acid; NE, norepinephrine; -HT, serotonin; 5-HIAA, 5-hydroxyindoleacetic acid.
E.Hoglundet
al./Brain
Resea
rch1032(2005)104–110
108
w
V
2
2
3
3
3
3
1
7
7
1
fo
eh
5

E. Hoglund et al. / Brain Research 1032 (2005) 104–110 109
material after the first (P=0.78) or the second addition
(P=0.88; Table 2). There were no significant effects of
available hiding material on position in the water tank in
vehicle treated individuals, or before the experimental
additions in any of the groups, see Table 2.
3.2. Effects of skin extract and availability of hiding
material on brain monoaminergic activity
Skin extract exposure significantly affected the ratio of
[DOPAC]/[DA] and resulted in an increased ratio in the
telencephalon (ANOVA: F1,19=4.48, P=0.040) and a
decreased ratio in the brain stem (ANOVA: F1,19=5.07,
P=0.037), compared to vehicle-treated controls (Fig. 1).
Availability of hiding material affected the [5-HIAA]/[5-
HT] ratio. Fish with no available hiding material had
significantly elevated ratios in the brain stem (ANOVA:
F1,19=4.47, P=0.048) and the optic tectum (ANOVA:
F1,19=6.77, P=0.023). A trend towards elevated ratios was
observed also in the hypothalamus and the telencephalon of
fish with no available hiding material, although they were
not significantly different from control (Fig. 2).
However, there were no combined effects of hiding
material and skin extract exposure on brain monoaminergic
activity, see Table 3 for ANOVA values.
4. Discussion
Exposures to skin extract from conspecifics resulted in a
typical predator avoidance behavior in crucian carp. This
avoidance behavior consists of an initial bactive avoidanceQphase (1–2 min of intense swimming against the bottom
floor), followed by a bpassive avoidanceQ phase (decreased
locomotor activity and a position lower in the water tank).
Active and passive avoidance of predators and of other
types of threat have been described in a wide range of
species. Even if both active and passive avoidance are
regarded as fear behaviors, the brain mechanisms seem
distinct and separate, in rats [25], cats [22], and possibly
also in primates [7].
Exposure to skin extract affected the brain DAergic
activity, it increased in the telencephalon and decreased in
the brain stem. This was independent of availability of
hiding material. Exposure to skin extract did not change the
brain 5-HT activity.
Lack of hiding material led to an elevation of 5-HTergic
activity in the brain stem and the optic tectum compared to
levels in fish with available hiding material. Major 5-
HTergic systems in hypothalamic and pretectal areas were
not significantly affected [11]. The most remarkable finding
was that this was independent of exposure to skin extract.
Absence of hiding material, therefore, influenced brain stem
5-HT activity even when no threat was present. The fish
were captured from their natural habitat (a lake) and
introduced to a totally new environment. Availability of
hiding material was enough to reduce the brain stem 5-HT
activity. This is a response antedating any avoidance
behavior, and, therefore, may qualify for bexpectancyQ,referred to as bcognitiveQ activity in mammals [24]. An
alternative explanation may be that the presence of hiding
material makes the environment less novel. Under any
circumstance the brain biochemistry of carps that register
familiarity and /or potential escape routes differs from that
of carps that do not register this.
To what extent do the innate avoidance responses in
crucian carp represent instrumental behavior with the same
neurochemical concomitants as learned behavior in mam-
mals, including humans? Is the crucian carp brain capable of
registering the potential escape possibilities to the extent
that this reduces the 5-HT activation induced by a novel or
threatening environment?
Presence or absence of hiding material also affected the
avoidance behavior, even if the 5-HT change was inde-
pendent of whether there was any adequate stimulus for
avoidance. When hiding material was available, the fish
stirred up a cloud of fine sediments and showed a more
pronounced decrease in locomotor activity in agreement
with this being a more efficient freezing or immobile
avoidance behavior. Lack of hiding material led to less
bfreezingQ (passive avoidance) behavior. In mammals,
established avoidance behavior is accompanied by low
levels of arousal, measured by behavior or plasma levels of
corticosterone [4,13,23]. If, however, the behavior is made
difficult, like in forced extinction of avoidance behavior,
arousal reappears. In our case, lack of hiding material may
be postulated to lead to more arousal and less avoidance
behavior. In mammals including humans, it has been
postulated that what the brain stores when avoidance is
acquired is a positive response outcome expectancy [3],
which leads to a low arousal level (the cognitive activation
theory of stress, CATS [24]). Similar expectancy functions
may be present in fish.
Such mechanisms appear to be present also in other
species of fish. In the long nose killifish submerged aquatic
vegetation as a refuge affected behavior and decreased
plasma cortisol content during visual predator exposure
[27]. Arctic charr exposed to a white background showed an
elevated NE activity in the optic tectum and the brainstem,
and a tendency for elevated 5-HT activity in the optic
tectum, compared to fish that were exposed to a black
background [10]. Absence of hiding material or avoidance
possibility elicits a high arousal also in other species, for
instance the rat. In rats, a typical response to threat by a
shock prod is to bury the shock prod, if burying material is
available. This leads to a low level of arousal. However, if
there is no burying material, the rats show a high level of
arousal [5].
DAergic systems in the fish brain are located in
telencephalon, in hypothalamic and pretectal areas, and in
the medulla oblongata [11]. The alarm response in our fish
involves increased DAergic activity in the telencephalon

E. Hoglund et al. / Brain Research 1032 (2005) 104–110110
and decreased DAergic activity in the brain stem, while
major DAergic systems in hypothalamic areas remained
unchanged. It is well established that the teleost tel-
encephalon is involved in the regulation of fear behavior
and the learning of avoidance responses, in particular with
the production and reinforcement mechanisms for avoidance
responses [18]. The telencephalon is involved in, but not
necessary for, both passive and active avoidance [17],
possibly through inhibitory mechanisms necessary for
freezing and passive avoidance. In particular, it seems to
be the telencephalic DAergic systems that are important for
fear behavior [15]. Our results confirm this position. The
active and the passive avoidance responses (flight and
freezing) observed in our experiment appear to be related to
the increase in telencephalic DAergic activity. However, our
study does not permit conclusions as to the possible
differential role of different structures in the telencephalon,
as has been demonstrated in other vertebrates [21].
Acknowledgments
Supported by the Norwegian Research Council. We
thank Trude Haug and ayvind averli for valuable com-
ments on earlier versions of this paper.
References
[1] C. Belzung, W. El Hage, N. Moindrot, G. Griebel, Behavioral and
neurochemical changes following predatory stress in mice, Neuro-
pharmacology 41 (2001) 400–408.
[2] D.C. Blanchard, G. Griebel, R.J. Rodgers, R.J. Blanchard, Benzodia-
zepine and serotonergic modulation of antipredator and conspecific
defense, Neurosci. Biobehav. Rev. 22 (1998) 597–612.
[3] R.C. Bolles, Reinforcement, expectancy and learning, Psychol. Rev.
79 (1972) 394–409.
[4] G.D. Coover, H. Ursin, S. Levine, Plasma-corticosterone levels during
active-avoidance learning in rats, J. Comp. Physiol. Psychol. 82
(1973) 170–174.
[5] S.F. De Boer, J.L Slangen, J. Van der Gugten, Plasma catecholamine
and corticosterone levels during active and passive shock-prod
avoidance behavior in rats: effects of chlordiazepoxide, Physiol.
Behav. 47 (1990) 1089–1098.
[6] K.B. Dbving, E.H. Hamdani, E. Hfglund, A. Kasumyan, A.
Tuvikene, A review on the chemical and physiological basis of alarm
reactions in cyprinids, in: A.S. Kapoor, K. Reuter (Eds.), Fish
Chemosenses, Science Publishers, Plymouth, 2004, ISBN: 1-57808-
319-2, (in press).
[7] J.A. Gray, The Neuropsychology of Anxiety: an Enquiry into the
Functions of the Septo-Hippocampal System, Oxford University
Press, Oxford, 1981.
[8] E. Hfglund, P.H. Balm, S. Winberg, Skin darkening, a potential social
signal in subordinate Arctic charr (Salvelinus alpinus): the regulatory
role of brain monoamines and pro-opiomelanocortin-derived peptides,
J. Exp. Biol. 203 (2000) 1711–1721.
[9] E. Hfglund, N. Kolm, S. Winberg, Stress-induced changes in brain
serotonergic activity, plasma cortisol and aggressive behavior in
Arctic charr (Salvelinus alpinus) is counteracted by l-DOPA, Physiol.
Behav. 74 (2001) 381–389.
[10] E. Hfglund, P.H. Balm, S. Winberg, Behavioural and neuroendocrine
effects of environmental background colour and social interaction in
Arctic charr (Salvelinus alpinus), J. Exp. Biol. 205 (2002) 2535–2543.
[11] J. Kaslin, P. Panula, Comparative anatomy of the histaminergic and
other aminergic systems in zebrafish (Danio rerio), J. Comp. Neurol.
440 (2001) 342–377.
[12] O. Lepage, a. averli, E. Petersson, T. Jarvi, S. Winberg, Differential
stress coping in wild and domesticated sea trout, Brain Behav. Evol.
56 (2000) 259–268.
[13] S. Levine, H. Ursin, What is stress? in: M.R. Brown, G.F. Koob, C.
Rivier (Eds.), Stress: Neurobiology and Neuroendocrinology, Marcel
Decker, New York, 1991, pp. 3–21.
[14] D.C. McIntyre, P. Kent, S. Hayley, Z. Merali, H. Anisman, Influence
of psychogenic and neurogenic stressors on neuroendocrine and
central monoamine activity in fast and slow kindling rats, Brain Res.
840 (1999) 65–74.
[15] E.Y. Mok, A.D. Munro, Effects of dopaminergic drugs on locomotor
activity in teleost fish of the genus Oreochromis (Cichlidae): involve-
ment of the telencephalon, Physiol. Behav. 64 (1998) 227–234.
[16] a. averli, C.A. Harris, S. Winberg, Short-term effects of fights for
social dominance and the establishment of dominant–subordinate
relationships on brain monoamines and cortisol in rainbow trout,
Brain Behav. Evol. 54 (1999) 263–275.
[17] J.B. Overmier, N.B. Flood, Passive avoidance in forebrain ablated
teleost fish, Carassius auratus, Physiol. Behav. 4 (1969) 791–794.
[18] J.B. Overmier, M.R. Papini, Factors modulating the effects of
teleost telencephalon ablation on retention, relearning, and
extinction of instrumental avoidance behavior, Behav. Neurosci.
100 (1986) 190–199.
[19] R.A. Rescorla, R.L. Solomon, Two process learning theory: relation-
ships between pavlovian conditioning and instrumental learning,
Psychol. Rev. 74 (1967) 151–182.
[20] G.R. Scott, K.A. Sloman, C. Rouleau, C.M. Wood, Cadmium disrupts
behavioural and physiological responses to alarm substance in
juvenile rainbow trout (Oncorhynchus mykiss), J. Exp. Biol. 206
(2003) 1779–1790.
[21] C.H. Summers, T.R. Summers, M.C. Moore, W.J. Korzan, S.K.
Woodley, P.J. Ronan, E. Hfglund, M.J. Watt, N. Greenberg, Temporal
patterns of limbic monoamine and plasma corticosterone response
during social stress, Neuroscience 116 (2003) 553–563.
[22] H. Ursin, The effect of amygdaloid lesions on flight and defense
behavior in cats, Exp. Neurol. 11 (1965) 61–79.
[23] H. Ursin, The psychology in psychoneuroendocrinology, Psychoneur-
oendocrinology 23 (1998) 555–570.
[24] H. Ursin, H.R. Eriksen, The cognitive activation theory of stress,
Psychoneuroendocrinology 290 (2004) 567–592.
[25] H. Ursin, P. Linck, R.A. McCleary, Spatial differentiation of
avoidance deficit following septal and cingulated lesions, J. Comp.
Physiol. Psychol. 68 (1969) 74–79.
[26] S. Winberg, A.A. Myrberg, G.E. Nilsson, Predator exposure alters
brain serotonin metabolism in bicolour damselfish, NeuroReport 4
(1993) 399–402.
[27] C.M. Woodley, M.S. Peterson, Measuring responses to simulated
predation threat using behavioral and physiological metrics: the role of
aquatic vegetation, Oecologia 136 (2003) 155–160.





























