Embed Size (px)
Citation preview

AVT is involved in the regulation of ion transport in the intestineof the sea bream (Sparus aurata)
Juan Antonio Martos-Sitcha a,b,c, Silvia Filipa Gregório a, Edison Samir M. Carvalho a,Adelino Vicente M. Canario a, Deborah Mary Power a, Juan Miguel Mancera b,Gonzalo Martínez-Rodríguez c, Juan Fuentes a,!a Centre of Marine Sciences (CCMar), CIMAR – Laboratório Associado, Universidade do Algarve, Campus de Gambelas, 8005-139 Faro, Portugalb Departamento de Biología, Facultad de Ciencias del Mar y Ambientales, Universidad de Cádiz, E-11510 Puerto Real (Cádiz), Spainc Instituto de Ciencias Marinas de Andalucía, Consejo Superior Investigaciones Científicas (ICMAN-CSIC), E-11510 Puerto Real (Cádiz), Spain
a r t i c l e i n f o
Article history:Received 20 April 2013Revised 26 July 2013Accepted 30 July 2013Available online 22 August 2013
Keywords:OsmoregulationArginine vasotocinSea breamSalinityWater absorption
a b s t r a c t
The intestine of marine fish plays a crucial role in ion homeostasis by selective processing of ingestedfluid. Although arginine vasotocin (AVT) is suggested to play a role in ion regulation in fish, its actionin the intestine has not been demonstrated. Thus, the present study investigated in vitro the putative roleof AVT in intestinal ion transport in the sea bream (Sparus aurata). A cDNA encoding part of an AVT recep-tor was isolated and phylogenetic analysis revealed it clustered with the V1a2-type receptor clade. V1a2transcripts were expressed throughout the gastrointestinal tract, from esophagus to rectum, and weremost abundant in the rectum regardless of long-term exposure to external salinities of 12, 35 or55 p.p.t. Basolateral addition of AVT (10!6 M) to the anterior intestine and rectum of sea bream adaptedto 12, 35 or 55 p.p.t. mounted in Ussing chambers produced rapid salinity and region dependentresponses in short circuit current (Isc), always in the absorptive direction. In addition, AVT stimulationof absorptive Isc conformed to a dose–response curve, with significant effects achieved at 10!8 M, whichcorresponds to physiological values of plasma AVT for this species. The effect of AVT on intestinal Isc wasinsensitive to the CFTR selective inhibitor NPPB (200 lM) applied apically, but was completely abolishedin the presence of apical bumetanide (200 lM). We propose a role for AVT in the regulation of ion absorp-tion in the intestine of the sea bream mediated by an absorptive bumetanide-sensitive mechanism, likelyNKCC2.
! 2013 Elsevier Inc. All rights reserved.
1. Introduction
The intestine of marine fish plays a key role in ion regulation.The ionic disequilibrium of marine fish with their surroundingenvironment requires high rates of water ingestion (drinking) aspart of the osmoregulatory process to compensate the dehydratingeffect of seawater in the gills (Evans et al., 2005). In this context,ion assimilation from the ingested fluid is required to drive waterabsorption, making the role of the intestine vital to maintain extra-cellular homeostasis. Water absorption seems to rely on Cl! up-take, which is mediated by an apical Na+/K+/2Cl! co-transporter(Musch et al., 1982) or apical Cl!/HCO3
! anion exchangers (Grosell,2006, 2011), both mechanisms are active in the sea breamintestine (Carvalho et al., 2012; Gregorio et al., 2013). A basolateralNa+,K+-ATPase generates the electrogenic potential to facilitateapical Cl! absorption (Ferlazzo et al., 2012). Accordingly, higherintestinal Na+,K+-ATPase activity is detected in seawater compared
to freshwater trout (Fuentes et al., 1997) and is consistent withhigher ATPase subunit expression in seawater compared to fresh-water eels (Kalujnaia et al., 2007).
Arginine vasotocin (AVT) is a peptide secreted from the neurohy-pophysis of non-mammalian vertebrates, whereas arginine vaso-pressin (AVP) is the structural and functional counterpart inmammals (Acher, 1993). The AVT/AVP family exerts its endocrineaction by binding specific plasma membrane receptors and has di-verse physiological actions ranging from behavior, stress, reproduc-tion, and vascular control to osmoregulation (Balment et al., 2006;Goodson, 2008; Kulczykowska, 2007; Mancera et al., 2008). ThreeAVP receptors (AVPRs) are described in mammals and retain a spe-cific and well-established tissue distribution, e.g. the AVPR typeV1a, is associated with vascular smooth muscle; the AVPR V1b, isin pituitary corticotrophs; and the AVPR V2 is in the kidney andlinked to renal hydrosmotic actions (Mahlmann et al., 1994; Warne,2001). In contrast, AVTRs in teleost fish have a broader tissue distri-bution. Thus, AVTR V1 or V2 subtypes have been described in thecentral nervous system and in tissues such as gill, kidney, gonadsor the gastrointestinal tract (Konno et al., 2009; Lema, 2010).
0016-6480/$ - see front matter ! 2013 Elsevier Inc. All rights reserved.http://dx.doi.org/10.1016/j.ygcen.2013.07.017
! Corresponding author.E-mail address: [email protected] (J. Fuentes).
General and Comparative Endocrinology 193 (2013) 221–228
Contents lists available at ScienceDirect
General and Comparative Endocrinology
journal homepage: www.elsevier .com/locate /ygcen

It seems likely that the prevailing action of the AVT/AVP systemis directed to preservation of blood volume and osmolality (Warneet al., 2002). Thus, AVP stimulates for instance tubular Na+ trans-port by activation of Na+ channels present in the apical membranesof the kidney tubule (Mordasini et al., 2005; Schafer et al., 1990).Yet, in other epithelia, such as the bronchial epithelium, AVP stim-ulates Cl! (and the resulting fluid) secretion via an NPPB-sensitive,likely CFTR dependent mechanism (Bernard et al., 2005). Otherelectrophysiological studies have demonstrated that AVP altersNaCl absorption with K+ requirement in the mouse kidney throughNa-K-2Cl (NKCC) co-transporter (Hebert and Andreoli, 1984; Sunet al., 1991), an action that seems to be mediated by recruitmentof co-transporter to the apical membrane (Molony et al., 1987).
Several studies have demonstrated the importance of AVT in ionand water regulation in fish (Balment et al., 2006; Kulczykowska,1997, 2001; Warne and Balment, 1995). Most of these studies fo-cused on the effects of AVT in ion regulation and demonstrated itparallels AVP actions in mammals. For instance, in trunk kidneypreparations of rainbow trout (Amer and Brown, 1995; Warneet al., 2002) and dogfish, Scyliorhinus canicula (Wells et al., 2002),AVT decreased urine output, an action probably linked to vasculareffects. Additionally, a regulatory action of AVT on Cl! secretion oc-curs in cultured branchial pavement cells from sea bass (Dicentrar-chus labrax) (Avella et al., 1999; Guibbolini and Avella, 2003), andthe effect is DPC-sensitive and likely mediated by CFTR. In fish, thereis also indirect indication for a putative action of AVT in the intes-tine. Thus, intracerebroventricular injections of AVT caused a reduc-tion in water intake in eels kept in seawater (Kozaka et al., 2003).Additionally, the presence of AVT receptors in the gastrointestinaltract of the rainbow trout was inferred from the action of AVT oncontraction of intestinal strips in vitro (Conklin et al., 1999).
The physiological actions of AVT in the intestine of marine fishhave received little attention despite its potential and intrinsicimportance in ion uptake/secretion. Therefore, the present studydetermined AVT function in the gilthead sea bream (Sparus aurata)intestine by establishing: (i) the distribution of AVTR V1a2-type,(ii) the response of AVTR to modified external salinity and; (iii)the effect in vitro of AVT on ion absorption/secretion.
2. Materials and methods
2.1. Peptides and chemicals
Arginine vasotocin ([Arg8]-vasotocin acetate), 5-nitro-2-(3-phenylpropylamino) benzoic acid (NPPB) and bumetanide (Bum)of the highest grade were purchased from Sigma–Aldrich (Madrid,Spain).
2.2. Animals
Sea bream (S. aurata) juveniles were obtained from commercialsources (Cupimar S.A., Cádiz, Spain). Fish were quarantined for60 days in Ramalhete Marine Station (University of Algarve, Faro,Portugal) in 1000 L tanks with running seawater at a density<5 kg/m3 and handfed twice daily to a final ration of 2% bodyweight, with a commercial sea bream diet (Sorgal, Portugal). Fishwere acclimated for at least 1 month before experimentation inflowing seawater (salinity 35 p.p.t.; water temperature 16–20 "C)under natural photoperiod for spring in the Algarve, Portugal. Inall experiments, food was withheld for 36 h before sacrifice and tis-sue collection to ensure the absence of undigested food in theintestine.
For salinity adaptation juvenile sea bream (n = 90; 20–30 g bodyweight) were separated into three equal groups and transferred to250 L tanks in three independent closed water circuits with biolog-
ical filters maintained at final salinities of 12, 35 or 55 p.p.t., tem-perature of 21 "C and a 14:10 L:D photoperiod. Increase in salinitywas achieved by adding Instant Ocean sea-salts to control seawater(35 p.p.t.), and decrease in salinity was achieved by dilution of fullseawater with dechlorinated tap water. Fish were kept at differentsalinities for 2 months before tissue collection and were consid-ered to be fully adapted (Laiz-Carrion et al., 2005). No mortalitywas registered during the trial and fish fed normally.
All animal manipulations were carried out in compliance withthe Guidelines of the European Union Council (86/609/EU) andPortuguese and Spanish legislation for the use of laboratory ani-mals. All animal protocols were performed under a ‘‘Group-I’’ li-cence from the Direcção-Geral de Veterinária, Ministério daAgricultura, do Desenvolvimento Rural e das Pescas (Portugal).
2.3. Cloning of AVT receptor
For cloning of the AVT receptor degenerate primers were de-signed (Table 1) against highly conserved cDNA sequences of pis-cine species: Platichthys flesus: (GenBank accession numberAF184966); Astatotilapia burtoni: (acc. No. AF517936); Catostomuscommersoni: (acc. No. X76321); Cyprinodon variegatus (V1a1; acc.No. GU120189); C. variegatus (V1a2; acc. No. GU120190); Cypryn-odon nevadensis amargosae (V1a1; acc. No. GU014233); Cyprynodonnevadensis amargosae (V1a2; acc. No. GQ981413). Tissues of unfedfish were collected into RNA Later (Sigma–Aldrich, Madrid, Spain)incubated for 24 h at 4 "C and stored at!20 "C until RNA extraction(within two weeks). Total RNA was isolated from brain, liver, kid-ney and gills using RNeasy# Plus MiniKit (Quiagen, Hidlen, Ger-many). Genomic DNA (gDNA) was eliminated with gDNAEliminator spin columns (Quiagen, Hidlen, Germany). After reversetranscription of 500 ng total RNA (Super Script III, Invitrogen™,Paisley, UK) PCR amplifications were carried out with Platinum#
Taq DNA Polymerase (Invitrogen) using the following PCR condi-tions: 94 "C, 1 min and 35 cycles of 94 "C, 30 s; 50 "C, 30 s; 72 "C,1 min and finalized with 72 "C, 10 min. PCR products were clonedinto TA Vectors (TOPO TA Cloning# Kit; Invitrogen™, Paisley, UK),sequenced and identity confirmed by interrogating NCBI databasesusing the blastn protocol (Altschul et al., 1990).
To establish receptor identity/subtype amino acid sequenceswere retrieved from the NCBI protein database (www.ncbi.nlm.nih.gov, accessed in January 2013) and phylogenetic analysis ofAVTR/AVPR translated sequences was performed using MEGA5software (Tamura et al., 2011) with the Close-Neighbor-Inter-change algorithm, based on amino acid differences (p-distances)and pairwise deletions. Reliability of the phylogenetic tree was as-sessed using bootstrap values (1000 replicates).
2.4. AVTR V1a2 in the sea bream intestine
For AVTR V1a2-type expression analysis, specific primer pairswere designed using the software Primer3 (available in http://frod-
Table 1Primers used for molecular identification of partial cDNA sequences of the AVT-R andfor qPCR expression analysis.
Nucleotide sequence
Degenerate primersAVT-RFw 50-AGCGTSCTGCTGGCSATG-30
AVT-RRv 50-GCAKATRAASCCGTAGCA-30
AVT-Rnested-Fw 50-AARCACCTSAGCCTBGCBGA-30
AVT-Rnested-Rv 50-GTBATCCAGGTGATGTASGC-30
qPCR primersAVTRFw 50-TTATCTCAACGTGGATGTGCAG-30
AVTRRv 50-TTACCAGGTGATGTAGGCCTTG-30
18sFw 50-AACCAGACAAATCGCTCCAC-30
18sRv 50-CCTGCGGCTTAATTTGACTC-30
222 J.A. Martos-Sitcha et al. / General and Comparative Endocrinology 193 (2013) 221–228

o.wi.mit.edu/) to amplify a 149 bp of the AVTR V1a2-type (Table 1).Total RNA was isolated using the E.Z.N.A. Total RNA Kit (OMEGABio-tek, Norcross, GA, USA) following the manufacturer’s instruc-tions, and the quantity and quality assessed (Nanodrop 1000, Ther-mo Scientific, Barrington, IL, USA). Total RNA was treated withDNase using an RNAse-free DNase kit (Ambion, Life Technologies,Paisley, UK). Total RNA (500 ng) was reverse transcribed (Revert-Aid™ First Strand cDNA Synthesis Kit, #K1622, Fermentas, ThermoScientific, Barrington, IL, USA).
Real-time qPCR amplifications were performed in duplicate in afinal volume of 10 ll with 5 ll SsoFast EvaGreen Supermix (Bio-Rad, Hercules, CA, USA) as the reporter dye, 200 ng cDNA, and0.5 pM of each forward and reverse primers. Amplifications wereperformed in 96-well plates using the One-step Plus sequencedetection system (Applied Biosystems, Foster City, CA, USA) withthe following protocol: denaturation and enzyme activation stepat 95 "C for 2 min, followed by 40 cycles of 95 "C for 5 s and60 "C for 10 s. After the amplification phase, a temperature-deter-mining dissociation step was carried out at 65 "C for 15 s, and95 "C for 15 s. For normalization of cDNA loading, all samples wererun in parallel using 18S ribosomal RNA (18S). To estimate efficien-cies, a standard curve was generated for each primer pair from 10-fold serial dilutions (from 10 to 0.001 ng) of a pool of first-strandedcDNA template from all samples. Standard curves represented thecycle threshold value as a function of the logarithm of the numberof copies generated, defined arbitrarily as one copy for the most di-luted standard. All calibration curves exhibited correlation coeffi-cients R2 > 0.98, and the corresponding real-time PCR efficiencieswere >99%. Following this general method AVTR V1a2-type mRNAdistribution in the gastrointestinal tract was analyzed in esopha-gus, stomach, pyloric caeca, anterior intestine, mid intestine andrectum of sea bream adapted to 35 p.p.t. In addition, the responseof AVTR V1a2-type mRNA to external salinity was analyzed in theanterior intestine and in the rectum in sea bream adapted long-term to 12, 35 and 55 p.p.t. external salinity.
2.5. Short-circuit current (Isc) measurements
The anterior intestine and rectum were collected, isolated andmounted in Ussing chambers as previously described (Gregorioet al., 2013). Briefly, tissue was placed on a tissue holder of0.71 cm2 and positioned between two half- chambers containing2 mL of physiological saline (NaCl 160 mM; MgSO4 1 mM;NaH2PO4 2 mM; CaCl2 1.5 mM; NaHCO3 5 mM; KCl 3 mM; Glucose5.5 mM; HEPES (4-(2-hydroxyethyl)piperazine-1-ethanesulfonicacid, N-(2-hydroxyethyl)piperazine-N0-(2-ethanesulfonic acid))4 mM), at a pH of 7.80. During the experiments the tissue wasbilaterally gassed with 0.3% CO2 + 99.7 O2 and the temperaturemaintained at 22 "C. Short circuit current (Isc, lA/cm2) was moni-tored by clamping of epithelia to 0 mV. Epithelial resistance (Rt,X cm2) was manually calculated (Ohm’s law) using the currentdeflections induced by a 2 mV pulse of 3 s every minute. Voltageclamping and current injections were performed by means of aDVC-1000 voltage clamp amplifier (WPI, Sarasota, USA) or aVCCMC2 (Physiologic Instruments, San Diego, USA). Bioelectricalparameters for each tissue were recorded after the tissue achieveda steady state, which occurred between 30–40 min after mounting.
The response to a single dose of 10!6 M AVT was analyzed inpreparations of anterior intestine and rectum collected from seabream adapted long-term to 12, 35 and 55 p.p.t. external salinity.To test if AVT effects on intestinal Isc conform to typical dose–re-sponse effects, rectal tissue from fish adapted to 55 p.p.t. salinitywere collected and mounted in Ussing chambers as described.After an initial period of Isc stabilization, tissues were treated at45 min intervals with doses of 10!10–10!6 M AVT at increased10-fold intervals covering the range of circulating plasma values
of AVT in the sea bream (Kleszczyska et al., 2006; Mancera et al.,2008).
Apical NPPB (200 lM) or Bumetanide (200 lM) were appliedalone or in combination with 10!6 M AVT in rectal tissue of fishadapted to 35 or 55 p.p.t. to identify the AVT-responsive mecha-nism. Concentration of specific blockers were selected to guarantee100% inhibitory effects on a putative CFTR (NPPB) or a NKCC(Bumetanide).
2.6. Statistics
Results are presented as means ± SEM unless otherwise stated.After normality and homogeneity of variance were checked, com-parison between groups was analyzed as appropriate using theStudent’s t-test or One-way analysis of variance (ANOVA) followedby the post hoc Bonferroni test. All statistical analysis was per-formed with Prism 5.0 (GraphPad Software for Macintosh) andgroups were considered significantly different at p < 0.05.
3. Results
3.1. cDNA for AVT receptor
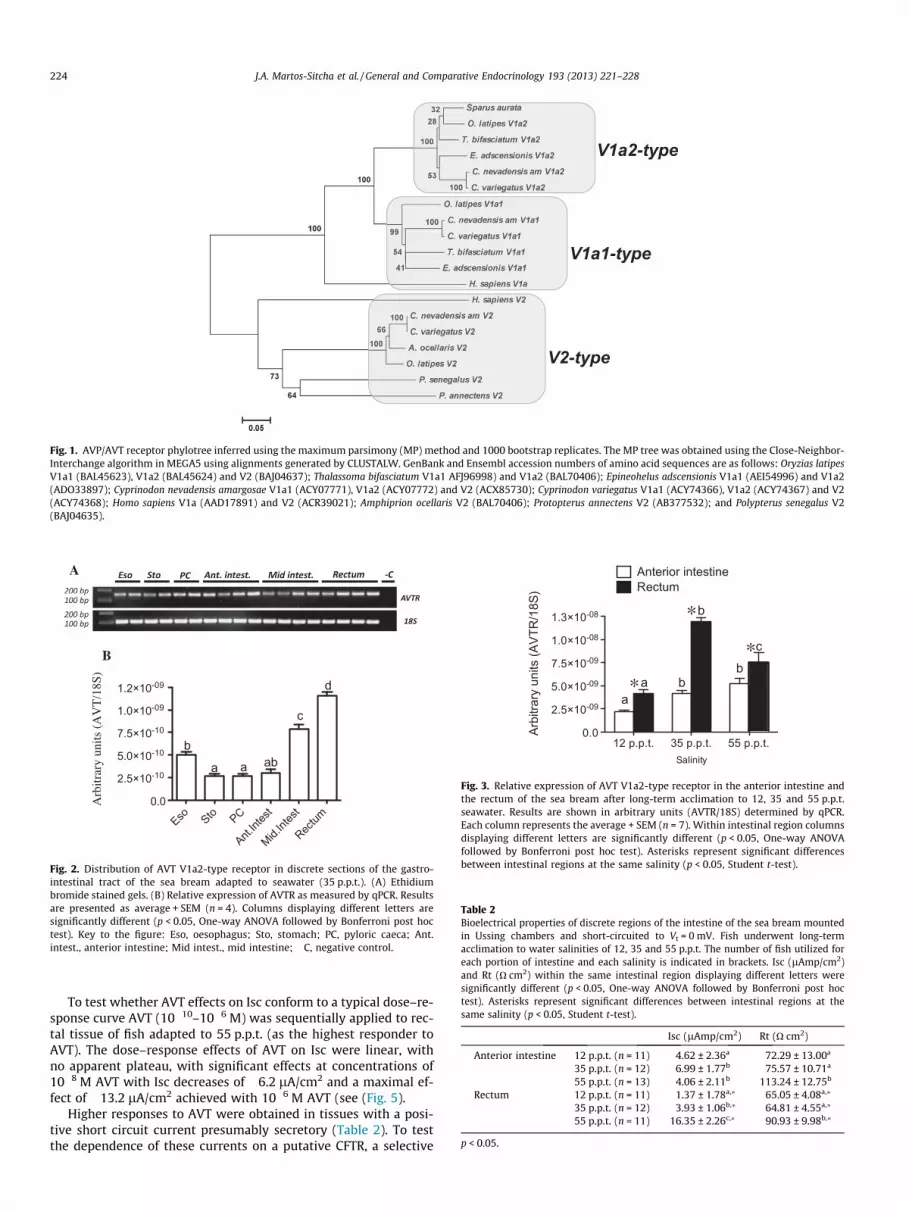
A cDNA for a partial AVT receptor spanning 392 bp of the codingsequence was isolated and clusters with the AVTR V1a2-type asshown by phylogenetic analysis (Fig. 1).
AVTR V1a2 is expressed throughout the intestinal tract: esoph-agus, stomach, pyloric caeca, anterior intestine, mid intestine andrectum (Fig. 2A). Furthermore expression increased in more distalportions of the intestinal tract, with highest expression in the rec-tum when compared to all other regions (Fig. 2B).
After acclimation of sea bream to external salinities of 12, 35and 55 p.p.t., AVTR V1a2 transcript abundance in the anteriorintestine paralleled decreases/increases of external salinity withsignificantly lower expression levels at 12 p.p.pt. (Fig. 3). In addi-tion, the transcript abundance of AVTR V1a2 was significantlyhigher in the rectum compared to the anterior intestine at allsalinities.
3.2. AVT intestinal electrophysiology
Basal values of short circuit current (Isc, lA/cm2) and tissueresistance (Rt, O cm2) are shown in Table 2. Positive Isc recordedshow secretory currents while absorptive currents are shown bynegative values. Control preparations sustained constant Isc andRt for the duration of the experimental periods (up to 3.5 h). Ingeneral the effects of salinity followed the pattern previously de-scribed for the sea bream intestine (Gregorio et al., 2013). In theanterior intestine a small absorptive current was observed in fishat 35 and 55 p.p.t., while at 12 p.p.t. a secretory current was re-corded (Table 2). In contrast the rectum of fish acclimated to 35and 55 p.p.t. showed a secretory current, which in low salinityadapted fish was highly variable but averaged a small absorptivecurrent.
The basolateral application of 10!6 M AVT to preparations ofanterior intestine and rectum from fish at 55, 35 and 12 p.p.t didnot affect tissue resistance (Rt, data not shown). In contrast, AVTevoked a rapid stimulatory action on Isc, (Fig. 4) in the absorptivedirection in all intestinal regions (the current became either morenegative or less positive). The effects of AVT on Isc were rapid, withan onset between 3 and 5 min of application and reached maximaleffects within 25–30 min after application (Fig. 4A). Interestingly,the effect of AVT in the anterior intestine was higher in fishadapted to 12 p.p.t., while in the rectum it was higher in fishadapted to 55 p.p.t.
J.A. Martos-Sitcha et al. / General and Comparative Endocrinology 193 (2013) 221–228 223
To test whether AVT effects on Isc conform to a typical dose–re-sponse curve AVT (10!10–10!6 M) was sequentially applied to rec-tal tissue of fish adapted to 55 p.p.t. (as the highest responder toAVT). The dose–response effects of AVT on Isc were linear, withno apparent plateau, with significant effects at concentrations of10!8 M AVT with Isc decreases of !6.2 lA/cm2 and a maximal ef-fect of !13.2 lA/cm2 achieved with 10!6 M AVT (see (Fig. 5).
Higher responses to AVT were obtained in tissues with a posi-tive short circuit current presumably secretory (Table 2). To testthe dependence of these currents on a putative CFTR, a selective
Table 2Bioelectrical properties of discrete regions of the intestine of the sea bream mountedin Ussing chambers and short-circuited to Vt = 0 mV. Fish underwent long-termacclimation to water salinities of 12, 35 and 55 p.p.t. The number of fish utilized foreach portion of intestine and each salinity is indicated in brackets. Isc (lAmp/cm2)and Rt (O cm2) within the same intestinal region displaying different letters weresignificantly different (p < 0.05, One-way ANOVA followed by Bonferroni post hoctest). Asterisks represent significant differences between intestinal regions at thesame salinity (p < 0.05, Student t-test).
Isc (lAmp/cm2) Rt (O cm2)
Anterior intestine 12 p.p.t. (n = 11) 4.62 ± 2.36a 72.29 ± 13.00a
35 p.p.t. (n = 12) !6.99 ± 1.77b 75.57 ± 10.71a
55 p.p.t. (n = 13) !4.06 ± 2.11b 113.24 ± 12.75b
Rectum 12 p.p.t. (n = 11) !1.37 ± 1.78a,⁄ 65.05 ± 4.08a,⁄
35 p.p.t. (n = 12) 3.93 ± 1.06b,⁄ 64.81 ± 4.55a,⁄
55 p.p.t. (n = 11) 16.35 ± 2.26c,⁄ 90.93 ± 9.98b,⁄
p < 0.05.
V1a2-type
V1a1-type
V2-type
Fig. 1. AVP/AVT receptor phylotree inferred using the maximum parsimony (MP) method and 1000 bootstrap replicates. The MP tree was obtained using the Close-Neighbor-Interchange algorithm in MEGA5 using alignments generated by CLUSTALW. GenBank and Ensembl accession numbers of amino acid sequences are as follows: Oryzias latipesV1a1 (BAL45623), V1a2 (BAL45624) and V2 (BAJ04637); Thalassoma bifasciatum V1a1 AFJ96998) and V1a2 (BAL70406); Epineohelus adscensionis V1a1 (AEI54996) and V1a2(ADO33897); Cyprinodon nevadensis amargosae V1a1 (ACY07771), V1a2 (ACY07772) and V2 (ACX85730); Cyprinodon variegatus V1a1 (ACY74366), V1a2 (ACY74367) and V2(ACY74368); Homo sapiens V1a (AAD17891) and V2 (ACR39021); Amphiprion ocellaris V2 (BAL70406); Protopterus annectens V2 (AB377532); and Polypterus senegalus V2(BAJ04635).
B
Eso Sto PC
Ant.Int
est
Mid.Int
est
Rectum
0.0
2.5!10-10
5.0!10-10
7.5!10-10
1.0!10-09
1.2!10-09
Arbit
rary u
nits (
AVT/
18S)
a ab
d
c
ab
A
Fig. 2. Distribution of AVT V1a2-type receptor in discrete sections of the gastro-intestinal tract of the sea bream adapted to seawater (35 p.p.t.). (A) Ethidiumbromide stained gels. (B) Relative expression of AVTR as measured by qPCR. Resultsare presented as average + SEM (n = 4). Columns displaying different letters aresignificantly different (p < 0.05, One-way ANOVA followed by Bonferroni post hoctest). Key to the figure: Eso, oesophagus; Sto, stomach; PC, pyloric caeca; Ant.intest., anterior intestine; Mid intest., mid intestine; !C, negative control.
12 p.p.t. 35 p.p.t. 55 p.p.t.0.0
2.5!10-09
5.0!10-09
7.5!10-09
1.0!10-08
1.3!10-08
Salinity
Arbi
trary
uni
ts (A
VTR
/18S
) RectumAnterior intestine
aa b
b
b
c
*
**
Fig. 3. Relative expression of AVT V1a2-type receptor in the anterior intestine andthe rectum of the sea bream after long-term acclimation to 12, 35 and 55 p.p.t.seawater. Results are shown in arbitrary units (AVTR/18S) determined by qPCR.Each column represents the average + SEM (n = 7). Within intestinal region columnsdisplaying different letters are significantly different (p < 0.05, One-way ANOVAfollowed by Bonferroni post hoc test). Asterisks represent significant differencesbetween intestinal regions at the same salinity (p < 0.05, Student t-test).
224 J.A. Martos-Sitcha et al. / General and Comparative Endocrinology 193 (2013) 221–228

inhibitor NPPB (200 lM) was applied apically to preparations ofrectum collected from fish adapted to 35 and 55 p.p.t. NPPB pro-duced a significant fall in the Isc recorded i.e. !4 lA/cm2 in the rec-tum of 35 p.p.t. acclimated fish and !14 lA/cm2 in the rectum of55 p.p.t. acclimated fish (Fig. 6).
To test whether the effects of AVT on Isc were mediated by reg-ulation of the secretory or absorptive pathways, rectal tissues offish acclimated to 35 and 55 p.p.t. were tested in Ussing chamberin the presence or absence of apical NPPB (200 lM) or bumetanide(200 lM). NPPB did not modify the response to 10!6 M AVT of rec-tal tissue from fish acclimated to 55 or 35 p.p.t seawater (Fig. 7). Incontrast, apical bumetanide (200 lM) completely abolished theepithelial response to AVT (Fig. 7) in 35 p.p.t. fish (no fish accli-mated to 55 p.p.t. were available for testing).
4. Discussion
In keeping with the proposed role for AVT in ion and water bal-ance in teleosts (Balment et al., 2006; Kulczykowska, 1997, 2001;
Warne and Balment, 1995), the present study established theintestine of marine teleosts as a target for a physiological role forthe vasotocinergic system. We show the heterogeneous distribu-tion and salinity dependence of the expression of a V1a2-typeAVT receptor in the gastrointestinal tract and characterized theinvolvement of AVT on ion movements in Ussing chambers. The ef-fect of AVT on ion transport in the intestine seems to be mediatedby a bumetanide sensitive mechanism, likely a NKCC co-transporter.
The sea bream cope with large changes in environmental salin-ity ranging from 5 to 60 p.p.t. (Laiz-Carrión et al., 2005), but areunable to withstand freshwater (Fuentes et al., 2010a). Acclimationto salinity occurs through water absorption and ion balance
0 10 20 30 40 50 60-30-20-10
010203040
Time (min)
Isc
( µAm
p/cm
2 )
AVT (10-6 M)
A
Anterior intestine Rectum
-15
-10
-5
Isc
(µAm
p/cm
2 )
12 ppt35 ppt55 ppt
aab b
b
a a
B
Fig. 4. (A) Original trace of short-circuit current (Isc, lA/cm2) recorded afterbasolateral application of AVT 10!6 M to the rectum from sea bream acclimated to55 p.p.t. Vertical current deflections are generated by ±1 mV pulses to calculate Rt.(B) Changes in the effects of basolateral AVT 10!6 M in the short circuit (DIsc,lAmp/cm2) in anterior intestine and rectum of the sea bream. Fish underwent long-term acclimation to changing external salinity 12, 35 and 55 p.p.t. Each columnrepresents the average + SEM (n = 6–8). Within intestinal region columns displayingdifferent letters are significantly different (p < 0.05, One-way ANOVA followed byBonferroni post hoc test).
-10 -9 -8 -7 -6
-20
-15
-10
-5
0
AVT log [M]
!Isc
(µAm
p/cm
2 )
*
**
Fig. 5. Changes in short-circuit current (DIsc, lAmp/cm2) measured in Ussingchambers in response to consecutive basolateral addition of AVT (10!10–10!6 M) torectum from sea bream acclimated to 55 p.p.t. Each point represents theaverage + SEM (n = 3). Asterisks represent significant differences from controls(p < 0.05, One-way ANOVA followed by Bonferroni post hoc test).
-2
0
2
4
6
*
Rectum: 35ppt
Basal +NPPB0
5
10
15
20
*
Rectum: 55ppt
Isc
(µAm
p/cm
2 )
Fig. 6. NPPB-sensitive short-circuit current (Isc, lAmp/cm2) in the rectum of seabream adapted to 35 or 55 p.p.t. Basal values of Isc are shown for epithelia after astabilization period followed by apical addition of NPPB (200 lM). Results areshown as mean + SEM (n = 5–6). Asterisks represent significant differences frombasal values (p < 0.05, Student’s t-test).
AVT +NPPB +Bum
-5
-4
-3
-2
-1
0
*
Rectum: 35ppt
-20
-15
-10
-5
0
Rectum: 55ppt
ND!Isc
(µAm
p/cm
2 )
Fig. 7. AVT-dependent short circuit current (DIsc, lAmp/cm2) in the rectum ofjuvenile sea bream acclimated to 35 or 55 p.p.t. meassured in Using chambers. AVTwas tested alone (AVT; 10!6 M) or in combination with apical NPPB (+NPPB,200 lM) or bumetanide (+Bum, 200 lM). ND, Not determined. Results are shown asmean + SEM (n = 6). Asterisks represent significant differences from AVT alone(p < 0.05, One-way ANOVA followed by Bonferroni post hoc test).
J.A. Martos-Sitcha et al. / General and Comparative Endocrinology 193 (2013) 221–228 225

mechanisms in the gills and intestine (Fuentes et al., 2006, 2010b;Gregorio et al., 2013; Raldúa et al., 2008). Furthermore, the gastro-intestinal tract of the sea bream is a target for endocrine regulationof ion transport (Ferlazzo et al., 2012; Fuentes et al., 2006,2010a,b). Previous studies in vertebrates have demonstrated thewidespread distribution of receptors for AVP/AVT in the intestineof birds (Tan et al., 2000), amphibians (Hasunuma et al., 2010)and mammals (Lolait et al., 1992; Morel et al., 1993). In fish, theintestinal expression of AVT receptor V1a-type and V2-type havebeen demonstrated in the African lungfish, Protopterus annectens(Konno et al., 2009). In addition, in the amargosa river pupfish(Cyprinodon nevadensis amargosae) three different types of AVTreceptors i.e. V1a1-type, V1a2-type and V2-type, are described(Lema, 2010). Our analysis of the sea bream partial AVT receptorindicates that it corresponds to an AVTR V1a2-type, one of the sub-types of the AVTR V1a described in teleosts (Fig. 1), although thesubtype V1a of the AVTR/AVPR has been associated with vascularsmooth muscle (V1a1/V1) or preferentially described in the pitui-tary corticotrophe cells (V1a2/V1b) (Mahlmann et al., 1994;Verbalis, 2002). The expression of the AVTR V1a2-type receptorthroughout the gastrointestinal tract of the sea bream highlightsa potential role for the vasotocinergic system in the intestine. Fur-thermore, expression of AVT V1a2 is high in the esophagus, andthere is an apparent antero-posterior increase from pyloric caecato rectum, where it is expressed at the highest level (Fig. 2). Addi-tionally, salinity seems to act as regulatory trigger for AVT receptorexpression in the amargosa river pupfish (Lema, 2010), although,the only receptor that seems to sustain higher branchial expressionin response to salinity is the V1a2-subtype, at least at 20 h postchallenge (Lema, 2010). In the sea bream, we show that along-term challenge with varying external salinity impacts theexpression of V1a2-type AVT receptor in the intestine (Fig. 3). Itis tempting to suggest that the regionalization and transcriptionalresponse to salinity of the V1a2-type AVT receptor are both relatedto functional specificity in ion regulation of discrete regions of theintestine in sea bream as has previously been described (Gregorioet al., 2013).
The intestine of fish maintains both absorptive and secretorypathways. Cl! uptake via NKCC co-transporters (Musch et al.,1982) or apical anion Cl!/HCO3
! exchangers (Grosell, 2011; Grosellet al., 2005) act to produce the driving force for water absorption.Additionally, the intestinal epithelium of Fundulus heteroclitusmounted in Ussing chambers can switch from net Cl! absorptionto net Cl! secretion as shown by directional changes of the tissueshort circuit current (Marshall et al., 2002). The sea bream intestinehas preferential Cl! absorption or secretion depending on externalsalinity and intestinal region (Carvalho et al., 2012; Gregorio et al.,2013). In the present study we describe how changes in externalsalinity and target tissue condition the response to a single doseof AVT in the anterior intestine and the rectum of the sea bream(Fig. 4A). A consistent feature of the responses to AVT by the seabream intestine in vitro is the consistency of the effect, always inthe absorptive direction. AVT makes Isc less positive (or more neg-ative) in all cases either by decreasing secretion or by increasingabsorptive currents. However, the magnitude of absorption stimu-lation is different in different regions of the sea bream intestine.Interestingly, preparations from tissues with positive currents i.e.secretory (Isc > 0) have the highest response to AVT and this isthe case of the anterior intestine from fish maintained at lowsalinity (12 p.p.t.) and the rectum of fish kept in hyperosmotic con-ditions i.e. 35 and 55 p.p.t. (Fig. 4B). In the latter we show that api-cal application of NPPB, a potent chloride channel blocker specificto CFTR (McCarty, 2000), results in decreases of the basal current of!3.5 lA/cm2 in fish at 35 p.p.t. and !14 lA/cm2, in fish at 55 p.p.t.demonstrating the presence of a functional CFTR.
AVT has a stimulatory action on Cl! secretion in cultured pave-ment cells of the sea bass gill via a DPC-sensitive mechanism, likelyCFTR (Avella et al., 1999; Guibbolini and Avella, 2003). Surpris-ingly, the effect of AVT on Isc in the intestine of the sea bream par-alleled in direction and magnitude the inhibitory effect of NPPB onbasal currents specifically in the rectum, that could be the reflec-tion of a pharmacological effect of the high concentration of AVT(10!6 M) in the Ussing chamber. However, the effects of AVT fol-lowed a linear dose–response in the range of 10!10–10!6 M andthe effect of AVT has no apparent plateau up to the highest concen-tration of AVT used (10!6 M). This is in keeping with the previousreport of heterologous expression of P. flesus AVT receptor in Xeno-pus oocytes where the response to AVT did not reach a plateau atconcentrations in a range of 10!14–10!10 M (Warne, 2001). Fur-thermore, regardless of the concentration, AVT actions are notmediated by regulation of the secretory pathway via CFTR. Onthe contrary a consistent single stimulatory effect on the absorp-tive pathway was observed. This is also supported by the un-changed stimulatory effect of AVT on the absorptive currentrecorded in the presence or absence of apical NPPB (Fig. 7).Remarkably, the hormone level at which a significant response toAVT occurred was 10!8 M, which falls within circulating plasmalevels (2–30 nM) of AVT in the sea bream (Kleszczyska et al.,2006; Mancera et al., 2008). The present results in the sea breamcontrast with previous reports in tilapia where AVT (2 lg/mL)was without effect on water, sodium and chloride transport inthe anterior intestine either in freshwater- or seawater-adaptedfish (Mainoya, 1985). However, our results support an importantphysiological role for AVT in the regulation of ion transport inthe marine fish intestine in vivo. This role for AVT is likely moreimportant during hyperosmotic adaptation and is probably linkedwith the functional specialization of the sea bream intestine, spe-cially at high salinities (Gregorio et al., 2013). Interestingly, highsalinity challenge results in coordinated increases/decreases ofplasma AVT/pituitary storage, respectively (Balment et al., 2006;Kulczykowska, 1997, 2001; Warne and Balment, 1995). Therefore,the linear response of the sea bream intestine to increasing dosesof AVT indicates that the receptor is not a limiting factor of the bio-logical action in ion transport.
In rats AVP regulates the expression of secretory-type Na-K-2Cl co-transporter (NKCC1) mRNA and protein in a dose-depen-dent manner in the outer medullary-collecting duct (OMCD),and therefore promotes the secretory pathway (Wakamatsuet al., 2009). However, AVP also controls NKCC2, the absorptiveform of the Na-K-2Cl co-transporter (Ares et al., 2011). It seemsthat the action of AVT in the intestine of the sea bream isachieved solely by its stimulatory effect on the absorptive path-way, which is preferentially mediated by the Na-K-2Cl co-trans-porter (Musch et al., 1982). The loop diuretic bumetanide is aspecific inhibitor of the NKCC (Haas, 1994) and when applied api-cally to sea bream rectum preparations, abolished almost com-pletely the stimulatory effect of AVT in the absorptive Isc(Fig. 7), and indicates there is functional association, likely similarto that described in the thick ascending limb of the loop of Henlein terrestrial vertebrates (Ares et al., 2011). However, it remainsto be established if these effects are achieved solely via theAVT V1a2-receptor type.
In conclusion, the present study provides strong evidencethat AVT is involved in the regulation of ion transport in theintestine of the sea bream. At least one AVT receptor, whichis distributed throughout the gastrointestinal tract, is expressedin response to salinity challenge in different intestinal regions.The intestine of the sea bream in vitro responds to stimulationwith AVT, an effect that is region and salinity dependent.Lowest effective doses for significant effects of AVT at
226 J.A. Martos-Sitcha et al. / General and Comparative Endocrinology 193 (2013) 221–228

concentrations similar to circulating plasma levels of the hor-mone support the physiological relevance of this regulation,which is likely achieved via apical NKCC2.
Acknowledgments
JAM-S is funded by Ministry of Education (Spain) through theprogram ‘‘Formación de Profesorado Universitario’’ (Ref: AP2008-01194). This work was partially supported by Ministry of Scienceand Education, Spain by Project AGL2010-14876 to JMM and bythe Ministry of Science and Higher Education and European SocialFunds through the Portuguese National Science Foundation by Pro-ject PTDC/MAR/104008/2008 to JF.
References
Acher, R., 1993. Neurohypophysial peptide systems: processing machinery, hydro-osmotic regulation, adaptation and evolution. Regul. Pept. 45, 1–13.
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990. Basic localalignment search tool. J. Mol. Biol. 215, 403–410.
Amer, S., Brown, J.A., 1995. Glomerular actions of arginine vasotocin in the in situperfused trout kidney. Am. J. Physiol. 269, R775–780.
Ares, G.R., Caceres, P.S., Ortiz, P.A., 2011. Molecular regulation of NKCC2 in the thickascending limb. Am. J. Physiol. Renal Physiol. 301, F1143–1159.
Avella, M., Part, P., Ehrenfeld, J., 1999. Regulation of Cl- secretion in seawater fish(Dicentrarchus labrax) gill respiratory cells in primary culture. J. Physiol. 516,353–363.
Balment, R.J., Lu, W., Weybourne, E., Warne, J.M., 2006. Arginine vasotocin a keyhormone in fish physiology and behaviour: a review with insights frommammalian models. Gen. Comp. Endocrinol. 147, 9–16.
Bernard, K., Bogliolo, S., Ehrenfeld, J., 2005. Vasotocin and vasopressin stimulation ofthe chloride secretion in the human bronchial epithelial cell line, 16HBE14o. Br.J. Pharmacol. 144, 1037–1050.
Carvalho, E.S., Gregorio, S.F., Power, D.M., Canario, A.V., Fuentes, J., 2012. Waterabsorption and bicarbonate secretion in the intestine of the sea bream areregulated by transmembrane and soluble adenylyl cyclase stimulation. J. Comp.Physiol. B 182, 1069–1080.
Conklin, D.J., Smith, M.P., Olson, K.R., 1999. Pharmacological characterization ofarginine vasotocin vascular smooth muscle receptors in the trout (Oncorhynchusmykiss) in vitro. Gen. Comp. Endocrinol. 114, 36–46.
Evans, D.H., Piermarini, P.M., Choe, K.P., 2005. The multifunctional fish gill:dominant site of gas exchange, osmoregulation, acid-base regulation, andexcretion of nitrogenous waste. Physiol. Rev. 85, 97–177.
Ferlazzo, A., Carvalho, E.S., Gregorio, S.F., Power, D.M., Canario, A.V., Trischitta, F.,Fuentes, J., 2012. Prolactin regulates luminal bicarbonate secretion in theintestine of the sea bream (Sparus aurata L.). J. Exp. Biol. 215, 3836–3844.
Fuentes, J., Soengas, J.L., Rey, P., Rebolledo, E., 1997. Progressive transfer to seawaterenhances intestinal and branchial Na+-K+-ATPase activity in nonanadromousrainbow trout. Aquacult. Int. 5, 217–227.
Fuentes, J., Figueiredo, J., Power, D.M., Canário, A.V.M., 2006. Parathyroid hormone-related protein regulates intestinal calcium transport in sea bream (Sparusauratus). Am. J. Physiol 291, R1499–R1506.
Fuentes, J., Brinca, L., Guerreiro, P.M., Power, D.M., 2010a. PRL and GH synthesis andrelease from the sea bream (Sparus auratus L.) pituitary gland in vitro inresponse to osmotic challenge. Gen. Comp. Endocrinol. 168, 95–102.
Fuentes, J., Power, D.M., Canário, A.V.M., 2010b. Parathyroid hormone-relatedprotein-stanniocalcin antagonism in regulation of bicarbonate secretion andcalcium precipitation in a marine fish intestine. Am. J. Physiol. 299, R150–R158.
Goodson, J.L., 2008. Nonapeptides and the evolutionary patterning of sociality. Prog.Brain Res. 170, 3–15.
Gregorio, S.F., Carvalho, E.S., Encarnacao, S., Wilson, J.M., Power, D.M., Canario, A.V.,Fuentes, J., 2013. Adaptation to different salinities exposes functionalspecialization in the intestine of the sea bream (Sparus aurata L.). J. Exp. Biol.216, 470–479.
Grosell, M., 2006. Intestinal anion exchange in marine fish osmoregulation. J. Exp.Biol. 209, 2813–2827.
Grosell, M., 2011. Intestinal anion exchange in marine teleosts is involved inosmoregulation and contributes to the oceanic inorganic carbon cycle. ActaPhysiol. (Oxf.) 202, 421–434.
Grosell, M., Wood, C.M., Wilson, R.W., Bury, N.R., Hogstrand, C., Rankin, C., Jensen,F.B., 2005. Bicarbonate secretion plays a role in chloride and water absorption ofthe European flounder intestine. Am. J. Physiol. Regul. Integr. Comp. Physiol.288, R936–946.
Guibbolini, M.E., Avella, M., 2003. Neurohypophysial hormone regulation of Cl!
secretion: physiological evidence for V1-type receptors in sea bass gillrespiratory cells in culture. J. Endocrinol. 176, 111–119.
Haas, M., 1994. The Na-K-Cl cotransporters. Am. J. Physiol. 267, C869–885.Hasunuma, I., Toyoda, F., Kadono, Y., Yamamoto, K., Namiki, H., Kikuyama, S., 2010.
Localization of three types of arginine vasotocin receptors in the brain andpituitary of the newt Cynops pyrrhogaster. Cell Tissue Res. 342, 437–457.
Hebert, S.C., Andreoli, T.E., 1984. Control of NaCl transport in the thick ascendinglimb. Am. J. Physiol. 246, F745–756.
Kalujnaia, S., McWilliam, I.S., Zaguinaiko, V.A., Feilen, A.L., Nicholson, J., Hazon, N.,Cutler, C.P., Cramb, G., 2007. Transcriptomic approach to the study ofosmoregulation in the European eel Anguilla anguilla. Physiol. Genomics 31,385–401.
Kleszczyska, A., Vargas-Chacoff, L., Gozdowska, M., Kalamarz, H., Martínez-Rodríguez, G., Mancera, J.M., Kulczykowska, E., 2006. Arginine vasotocin,isotocin and melatonin responses following acclimation of gilthead sea bream(Sparus aurata) to different environmental salinities. Comp. Biochem. Physiol.145, 268–273.
Konno, N., Hyodo, S., Yamaguchi, Y., Kaiya, H., Miyazato, M., Matsuda, K., Uchiyama,M., 2009. African lungfish, Protopterus annectens, possess an arginine vasotocinreceptor homologous to the tetrapod V2-type receptor. J. Exp. Biol. 212, 2183–2193.
Kozaka, T., Fujii, Y., Ando, M., 2003. Central effects of various ligands on drinkingbehavior in eels acclimated to seawater. J. Exp. Biol. 206, 687–692.
Kulczykowska, E., 1997. Response of circulating arginine vasotocin and isotocin torapid osmotic challenge in rainbow trout. Comp. Biochem. Physiol. 118, 773–778.
Kulczykowska, E., 2001. Responses of circulating arginine vasotocin, isotocin, andmelatonin to osmotic and disturbance stress in rainbow trout (Oncorhynchusmykiss). Fish Physiol. Biochem. 24, 201–206.
Kulczykowska, E., 2007. Arginine vasotocin and isotocin towards their role in fishosmoregulation. In: Baldisserotto, J.M., Mancera Romero, Kapoor, B.G. (Eds.),Fish Osmoregulation B. Science Publisher, Enfield (NH), pp. 151–176.
Laiz-Carrión, R., Guerreiro, P.M., Fuentes, J., Canário, A.V.M., Martín Del Río, M.P.,Mancera, J.M., 2005. Branchial osmoregulatory response to salinity in thegilthead sea bream, Sparus auratus. J. Exp. Zool. Part A 303, 563–576.
Laiz-Carrion, R., Sangiao-Alvarellos, S., Guzman, J.M., del Rio, M.P.M., Soengas, J.L.,Mancera, J.M., 2005. Growth performance of gilthead sea bream Sparus auratain different osmotic conditions: implications for osmoregulation and energymetabolism. Aquaculture 250, 849–861.
Lema, S.C., 2010. Identification of multiple vasotocin receptor cDNAs in teleost fish:sequences, phylogenetic analysis, sites of expression, and regulation in thehypothalamus and gill in response to hyperosmotic challenge. Mol. CellEndocrinol. 321, 215–230.
Lolait, S.J., O’Carroll, A.M., McBride, O.W., Konig, M., Morel, A., Brownstein, M.J.,1992. Cloning and characterization of a vasopressin V2 receptor and possiblelink to nephrogenic diabetes insipidus. Nature 357, 336–339.
Mahlmann, S., Meyerhof, W., Hausmann, H., Heierhorst, J., Schönrock, C., Zwiers, H.,Lederis, K., Richter, D., 1994. Structure, function, and phylogeny of[Arg8]vasotocin receptors from teleost fish and toad. Proc. Natl. Acad. Sci.USA 91, 1342–1345.
Mainoya, J.R., 1985. Effect of peptide hormones on intestinal transport in fish. In:Lofts, B., Holmes, W.N. (Eds.), Current Trends in Comparative Endocrinology,vol. 2. Hong Kong University Press, Hong Kong, pp. 910–913, ISBN 962-209-116-4.
Mancera, J.M., Vargas-Chacoff, L., Garcia-Lopez, A., Kleszczynska, A., Kalamarz, H.,Martinez-Rodriguez, G., Kulczykowska, E., 2008. High density and fooddeprivation affect arginine vasotocin, isotocin and melatonin in gilthead seabream (Sparus auratus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 149, 92–97.
Marshall, W.S., Howard, J.A., Cozzi, R.R.F., Lynch, E.M., 2002. NaCl and fluid secretionby the intestine of the teleost Fundulus heteroclitus: involvement of CFTR. J.Exp. Biol. 205, 745–758.
McCarty, N.A., 2000. Permeation through the CFTR chloride channel. J. Exp. Biol.203, 1947–1962.
Molony, D.A., Reeves, W.B., Hebert, S.C., Andreoli, T.E., 1987. ADH increases apicalNa+, K+, 2Cl! entry in mouse medullary thick ascending limbs of Henle. Am. J.Physiol. 252, F177–187.
Mordasini, D., Bustamante, M., Rousselot, M., Martin, P.Y., Hasler, U., Féraille, E.,2005. Stimulation of Na+ transport by AVP is independent of PKAphosphorylation of the Na-K-ATPase in collecting duct principal cells. Am. J.Physiol. 289, F1031–F1039.
Morel, A., Lolait, S.J., Brownstein, M.J., 1993. Molecular cloning and expression of ratV1a and V2 arginine vasopressin receptors. Regul. Pept. 45, 53–59.
Musch, M.W., Orellana, S.A., Kimberg, L.S., Field, M., Halm, D.R., Krasny Jr., E.J.,Frizzell, R.A., 1982. Na+-K+-Cl- co-transport in the intestine of a marine teleost.Nature 300, 351–353.
Raldúa, D., Otero, D., Fabra, M., Cerdá, J., 2008. Differential localization andregulation of two aquaporin-1 homologs in the intestinal epithelia of themarine teleost Sparus aurata. Am. J. Physiol. 294, R993–R1003.
Schafer, J.A., Troutman, S.L., Schlatter, E., 1990. Vasopressin and mineralocorticoidincrease apical membrane driving force for K+ secretion in rat CCD. Am. J.Physiol. 258, F199–F210.
Sun, A., Grossman, E.B., Lombardi, M., Hebert, S.C., 1991. Vasopressin alters themechanism of apical Cl! entry from Na+:Cl! to Na+:K+:2Cl! cotransport inmouse medullary thick ascending limb. J. Membr. Biol. 120, 83–94.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., Kumar, S., 2011. MEGA5:molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28,2731–2739.
Tan, F.L., Lolait, S.J., Brownstein, M.J., Saito, N., MacLeod, V., Baeyens, D.A., Mayeux,P.R., Jones, S.M., Cornett, L.E., 2000. Molecular cloning and functional
J.A. Martos-Sitcha et al. / General and Comparative Endocrinology 193 (2013) 221–228 227

characterization of a vasotocin receptor subtype that is expressed in the shellgland and brain of the domestic chicken. Biol. Reprod. 62, 8–15.
Verbalis, J.G., 2002. Vasopressin V2 receptor antagonists. J. Mol. Endocrinol. 29, 1–9.Wakamatsu, S., Nonoguchi, H., Ikebe, M., Machida, K., Izumi, Y., Memetimin, H.,
Nakayama, Y., Nakanishi, T., Kohda, Y., Tomita, K., 2009. Vasopressin andhyperosmolality regulate NKCC1 expression in rat OMCD. Hypertens. Res. 32,481–487.
Warne, J.M., 2001. Cloning and characterization of an arginine vasotocin receptorfrom the euryhaline flounder Platichthys flesus. Gen. Comp. Endocrinol. 122,312–319.
Warne, J.M., Balment, R.J., 1995. Effect of acute manipulation of blood volume andosmolality on plasma [AVT] in seawater flounder. Am. J. Physiol. 269, R1107–R1112.
Warne, J.M., Harding, K.E., Balment, R.J., 2002. Neurohypophysial hormonesand renal function in fish and mammals. Comp. Biochem. Physiol. 132, 231–237.
Wells, A., Anderson, W.G., Hazon, N., 2002. Development of an in situ perfusedkidney preparation for elasmobranch fish: Action of arginine vasotocin. Am. J.Physiol. 282, R1636–R1642.
228 J.A. Martos-Sitcha et al. / General and Comparative Endocrinology 193 (2013) 221–228





























