Embed Size (px)
Citation preview

I. PENDAHULUAN
A. Pengertian Ekologi Hutan
Istilah Ekologi diperkenalkan oleh Ernest Haeckel
(1869), yang mana ekologi ini berasal dari bahasa Yunani,
yaitu :
Oikos = Tempat tinggal (rumah)
Logos = ilmu, telaah.
Oleh karena itu, Ekologi adalah ilrnu yang mempelajari
hubungan timbal balik antara mahluk hidup dengan sesamanya
dan dengan lingkungannya.
Hubungan tersebut demikian komplek dan eratnya se-
hingga Odum (1959) menyatakan bahwa ekologi adalah Envi-
romental Ecology.
Hutan adalah masyarakat tumbuh-tumbuhan yang dikuasai
pohon-pohonan dan mempunyai keadaan lingkungan yang berbeda
dengan keadaan di luar hutan. Didalam suatu hutan,
hubungan antara tumbuh-tumbuhan, margasatwa, dan alam
lingkungannya demikian eratnya, sehingga hutan dipandang
sebagai suatu sistem ekologi atau ekosistem.
Ekologi Hutan adalah cabang ekologi yang khusus
mempelajari masyarakat atau ekosistim hutan. B. Bidang Kajian Ekologi Kutan
Didalam Ekologi ada dua bidang kajian, yaitu
1. Autekologi : Ekologi yang mempelajari suatu jenis
organisma yang berinteraksi dengan
1ingkungannya atau ekologi sesuatu

jenis atau bagian ekologi yang
mempelajari pengaruh sesuatu
faktor lingkungan terhadap satu
atau lebih jenis-jenis organisme.
2. Sinekologi : Bagian ekologi yang mempelajari
berbagai kelompok organisme
sebagai satu kesatuan yang saling
berinteraksi antar sesamanya
dan dengan lingkungannya dalam
suatu daerah.
Dalam ekologi hutan, autekologi mempelajari
pengaruh suatu faktor lingkungan terhadap hidup dan
tumbuhnya satu atau lebih jenis-jenis pohon. Jadi,
penyelidikannya mirip fisiologi tumbuh-tumbuhan,
sehingga aspek-aspek tertentu dari autekologi, seperti
penelitian tentang pertumbuhan pohon serir.g disebut
fisioekologi (phisiological ecology). Contoh penelitian
autekologi adalah :
1) Pengaruh intensitas cahaya terhadap
pertumbuhan jenis Shorea leprosula
2) Pengaruh dosis pupuk N terhadap
pertumbuhan jenis sengon.
Sedangkan Sinekologi mempelajari hutan sebagai suatu
ekosistem. Contoh kajian sinekologi adalah pengaruh

keadaan tempat tumbuh terhadap komposisi, struktur dan
produktivitas hutan.
Dalam ekologi hutan baik penge tahuan autekologi
maupun sinekologi bersama-sama diperlukan, karena kita
memerlukan pengetahuan tentang sifat-sifat berbagai jenis
pohon yang membentuk hutan dan pengetahuan tentang hutan
sebagai suatu ekosistem.
C. Sangkut Paut Ekologi Hutan dengan Bidang Ilmu Lain
Berhubung di dalam ekologi hutan yang dipelajari
adalah tumbuh-tumbuhan hutan dan keadaan tempat tumbuhnya,
maka semua bidang ilmu yang mempelajari kedua komponen
ekosistem hutan tersebut sangat diperlukan, yakni:
(1). Taksonomi tumbuh-tumbuhan (terutama Dendrologi).
Bidang ilmu ini sangat diperlukan untuk pengenalan
jenis-jenis tumbuhan di hutan. Untuk pengenalan jenis ini
diperlukan buku-buku pengenalan jenis yang praktis, selain
buku-buku flora yang sudah ada yang bersifat komprehensif.
Cara pengenalan jenis pohon dalam buku-buku itu
dititikberatkan pada sifat-sifat generatif (reproduktif),
yaitu berdasarkan sifat-sifat bunga dan buah. Padahal
menurut pengalaman di lapangan seringkali dijumpai pohon-
pohon yang sedang tidak berbunga atau berbuah, atau sukar
sekali untuk mendapatkan contoh-contoh bunga dan buah.
Karena itu, untuk keperluan di lapangan dibutuhkan
cara pengenalan jenis pohon yang terutama didasarkan pada
sifat-sifat vegetatif, yaitu sifat-sifat batang pohon

(kulit, getah dan kayu) , daun dan kuncup, kemudian baru
sifat-sifat generative. Cara pengenalan ini tidak terikat
pada sistem taksonomi tumbuh-tumbuhan. Di Filipina cara
pengenalan demikian telah dirintis oleh Tamolang (1959),
di Malaysia oleh Kochummen (1963), di Indonesia oleh
Endert (1928, 1956) dan Verteegh (1971) dan di Pantai
Gading, Afrika, oleh den Outer (1972) .
Kepulauan Indonesia, sebagai bagian dari daerah flora
Malesia, terkenal sebagai daerah flora hutan yang kaya.
tetapi pengetahuan kita tentang jenis tumbuh-tumbuhan di
daerah ini masih amat kurang. Banyaknya jenis tumbuh-
tumbuhan di daerah inipun belum diketahui dengan pasti
(Van Steenis, 1948). Menurut taksiran Van Steenis (op.cit)
di daerah Malesia terdapat kira-kira 3000 jenis pohon.
Menurut Lembaga Penelitian Hutan di Indonesia terdapat
lebih kurang 4000 jenis pohon. Dari sekian banyak jenis
itu baru sebagian kecil tercakup dalam buku-buku flora
yang tersedia. Akibatnya, pengenalan masih tergantung pada
jasa para pengenal pohon setempat. Dengan bantuan koleksi
contoh tumbuh-tumbuhan yang kemudian dideterminasi,
dapatlah disusun daftar nama pohon-pohon untuk daerah
tertentu, yang dapat mempermudah inventarisasi hutan.
(2). Geologi dan Geomorfologi
Ilmu-ilmu ini diperlukan dalam ekologi hutan, karena
keadaan geologi dan geomorfologi mempengaruhi pembentukan
dan sifat-sifat tanah serta penyebaran dan hidup tumbuh-

tumbuhan.
Pada keadaan iklim yang sama, jenis-jenis batuan yang
berbeda akan menghasilkan jenis-jenis tanah yang
berlainan. Pada jenis dan keadaan tanah yang khusus,
seperti tanah pasir kuarsa dan tanah serpentin, akan
terbentuk tipe hutan yang khusus pula.
Keadaan topografi juga mempengaruhi komposisi dan
kesuburan tegakan hutan, melalui perbedaan pada kesuburan
dan keadaan air tanah.
Disamping itu, perbedaan letak tinggi mempengaruhi
penyebaran tumbuh-tumbuhan, melalui perbedaan iklim yang
ditimbulkannya.
(3). Ilmu Tanah
Ilmu tanah yang murni seringkali disebut pedologi
tetapi sebagai faktor tempat tumbuh disebut edafologi.
Perbedaan jenis tanah, sifat-sifat serta keadaan tanah
seringkali mempengaruhi penyebaran tumbuh-tumbuhan,
menyebabkan terbentuknya tipe-tipe vegetasi berlainan,
serta mempengaruhi kesuburan dan produktivitas hutan.
(4). Klimatologi
Iklim adalah faktor terpenting yang mempengaruhi
penyebaran tumbuh-tumbuhan. Faktor-faktor iklim seperti
suhu (temperatur), curah hujan, kelembaban, dan defisit
tekanan uap air besar pengaruhnya pada pertumbuhan pohon.
Iklim mikro dari sesuatu ternpat yang dipengaruhi keadaan

topografi dapat mempengaruhi penyebaran dan pertumbuhan
pohon.
(5). Geografi tumbuh-tumbuhan
Pada permulaan perkembangannya ekologi tumbuh-
tumbuhan merupakan cabang dari geografi tumbuh-tumbuan
(phytogeografi) yang membahas pengaruh faktor-faktor
lingkungan terhadap penyebaran tumbuh-tumbuhan. Dari
cabang inilah berkembang sosiologi tumbuh-tumbuhan
(phytososiologi) dan ekologi tumbuh-tumbuhan.
Pada taraf kemajuan sekarang ekologi hutan masih
memerlukan informasi dari geografi tumbuh-tumbuhan untuk
mengerti pola penyebaran berbagai jenis pohon dalam
hubungannya dengan keadaan fisik bumi, terutama iklim dan
geomorfologi atau fisiografi, dan akan sangat membantu
dalam mempelajari susunan serta penyebaran £ormasi-formasi
hutan.
(6). Fisiologi Tumbuh-tumbuhan dan Biokimia
Telah dikemukakan bahwa autekolcgi mempunyai kegiatan
yang mendekati fisiologi tumbuh-tumbuhan. Jadi pada
umumnya informasi dari fisiologi tumbuh-tumbuhan akan
sangat berguna untuk mempelajari proses-proses hidup
tumbuh-tumbuhan, yang mana memerlukan pengetahuan tentang
proses-proses kimia yang berhubungan dengan aktivitas
biologis yang terjadi. Informasi tersebut bisa diperoleh
dari ilmu biokimia. Misalnya, untuk dapat mempelajari

pengaruh faktor-faktor lingkungan terhadap produksi getah
karet atau getah pinus perlu pula pengetahuan tentang
proses pembentukan getah dan proses-proses biokimia
lainnya yang mempengaruhi atau berkaitan dengannya.
(7). Genetika Tumbuh-tumbuhan
Suatu jenis tumbuh-tumbuhan yang penyebarannya luas
seringkali memperlihatkan perbedaan menurut letak geografi
dan keadaan lingkungan-nya. Perbedaan ini bukan hanya
dalam bentuk pertumbuhannya tetapi seringkali pula dalam
hal adaptasi dan persyaratan terhadap keadaan tempat
tumbuhnya, yang berakar pada sifat-sifat genetis, sebagai
akibat dari mutasi dan polyploidy.
Adakalanya apabila daerah penyebaran dari dua jenis
pohon berimpitan pada suatu tempat, maka pada tempat itu
terjadi hybridisasi antara kedua jenis itu, sehingga
timbul jenis pohon baru yang sifat-sifatnya berada
diantara sifat-sifat kedua jenis induknya. Demikianlah,
pada keadaan-keadaan tertentu, untuk mengerti sifat-sifat
ekologis sesuatu jenis atau beberapa jenis pohon
diperlukan pula pengatahuan tentang genetika.
(8). Matematika dan Statistika
Kedua ilmu ini sangatlah penting untuk
memformulasikan dugaan kuantitatif terhadap berbagai
proses ekologis yang terjadi pada ekosistem hutan. Oleh
karena itu, melalui penggunaan kedua bidang ilmu ini

faktor lingkungan yang berperan dan seberapa jauh
peranannya terhadap penelitian kelestarian suatu hutan
dapat diperkirakan.
D. Status Ekologi Hutan dalam Ilmu Pengetahuan Kehutanan
Ekologi Hutan merupakan ilmu dasar yang bersifat
integratif (mengintegrasikan ilmu-ilmu dasar lain) yang
merupakan ilmu dasar penting bagi silvikultur. dalam
terminologi kehutanan, ekologi hutan hampir sama dengan
silvika. Perbedaan ekologi hutan dengan silvika hanyalah
pada lawasan kajiannya, yakni ekologi hutan mempelajari
hutan sebagai ekosistem (jadi lawasannya lebih luas),
sedangkan silvika lebih terarah pada silvikultur dan lebih
mendekati autekologi. Dengan pengetahuan ekologi hutan dan
fisiologi pohon yang tepat bisa ditentukan tindakan
silvikultur yang tepat, sehingga produksi hutan dapat
ditingkatkan baik kualita rnaupun kuantitanya.
E. Aspek-aspek Ekologi Hutan yang renting
Dalam ilmu kehutanan, aspek-aspuk ekologi hutan yang
penting dipelajari adalah :
(1). mempelajari komposisi dan struktur hutan alam
(2). mempelajari hubungan tempat tumbuh denyan:
a. komposisi dan struktur hutan
b. penyebaran jenis-jenis pohon
c. permudaan pohon atau permudaan hutan
d. riap (pertumbuhan) pohon/hutan

e. fenologi pohon (musim berbunga, berbuah, pergantian
daun).
(3). mempelajari syarat-syarat keadaan tempat tumbuh
penanaman atau permudaan alam
(4). mempelajari siklus hara mineral, siklus air, dan
metabolisme.
(5). mempelajari hubungan antara kesuburan tanah, iklim
dan faktor-faktor lain dengan produktivitas hutan
(6). mempelajari suksesi vegetasi hutan secara alam dan
setelah terjadi kerusakan.

II. EKOSISTEM
A. Pengertian
Ekosistem adalah suatu sistem di alam yang
mengandung komponen hayati (organisme} dan komponen non-
hayati (abiotik), dimana antara kedua komponen tersebut
terjadi hubungan timbal balik untuk mempertukarkan zat-zat
yang perlu untuk mempertahankan kehidupan.
Dalam beberapa kepustakaan, istilah biocoenosis,
geocoenosis, dan biogeocoenosis (geobiocoenosis) secara
berurutan digunakan untuk komponen biotik, abiotik dan
ekosistem.
Ekosistem merupakan satuan fungsional dasar ekologi,
karena ekosistem mencakup organisme dan lingkungan abiotik
yang saling berinteraksi.
Pencetus istilah ekosistem adalah A.G. Tarisley pada
tahun 1935, seorang ekolog Inggeris.
B. Komponen Ekosistem
1. Dari Segi "trophic level", ekosistem terdiri atas:
1) Komponen autotrofik, yaitu organisme yang mampu
mensitesis makanannya sendiri yang berupa bahan
organik dari bahan-bahan anorganik sederhana
dengan bantuan sinar matahari dan zat hijau daun.
2) Komponen heterotrofik, yaitu organisme yang sumber
makanannya diperoleh dari bahan-bahan organik yang
dibentuk oleh komponen aututrofik, penyusun
kembali dan menguraikan bahan-bahan organik

kompleks yang telah mati kedalam senyawa
anorganik sederhana.
Dari segi penyusunnya (struktur), komponen
ekosistem terdiri atas :
1) Komponen abiotik yaitu komponen fisik dan kirnia
seperti tanah, air, udara, sinar matahari, dll.'
yang merupakan medium untuk berlangsungnya
kehidupan.
2) Produsen yaitu organisme autotrofik, umumnya
tumbuhan berklorofil, yang mampu mensintesis
makanannya sendiri dari bahan anorganik
3) Konsumen yaitu organisme heterotrofik
4) Pengurai, yaitu organisme heterotrofik yang menguraikan
bahan organik yang berasal dari organisme mati,
menyerap sebagian hasil penguraian tersebut dan
melepas bahan-bahan yang sederhana yang dapat
dipakai oleh produsen.
Untuk tujuan deskripsif, komponen-komponen
ekosistem seyogyanya diperinci sebagai berikut :
1) Bahan-bahan anorganik (C, N, Co2, H20, dll)
2) Senyawa organik (protein, lemak, karbohidrat, dll) 3) Iklim fsuhu, dan faktor fisik lainnya)
4) Produser
5) Konsumer makro ("phagotroph" yaitu organisme
heterotrofik, umumnya hewan) yang memakan
organisme lain atau bahan organik.
6) Konsumer mikro (saprotroph, osmotroph), yaitu

organisme heterotrofik, umumnya jamur dan bakteri,
yang menghancurkan bahan organik mati, menyerap
sebagian hasil perombakannya, dan membebaskan
bahan-bahan anorganik sederhana yang berguna bagi
produser.
Point (1) s/d (3) adalah Komponen abiotik.
Point (4) s/d (6) adalah komponen biotik.
Organisme heterotrofik dapat juga dibedakan kedalam :
1) Biophage, yaitu organisme yang mengkonsumsi organisme
lainnya.
2) Saprophage, yaitu organisme pengurai bahan-bahan
organik yang telah mati. Dari segi fungsional,
suatu ekoisistem sebaiknya dianalisis menurut :
(1). Aliran energi
(2). Rantai pangan
(3). Pola keanekaragaman dalam ruang dan wakcu
(4}. Siklus nutrien
(5). Pengembangan dan evolusi
(6). Kontrol (sibernetik)
Dalam hal konsumer, selain pembagian di atas,
konsumer dapat juga dibedakan kedalam:
1) Konsumer I (konsumer primer) adalah hewan-hewan
herbivora yang makanannya bergantung pada produser
(tumbuhan hLjau), contoh : insekta, rodentia,
kelinci, dll. (ekosistem daratan), moluska,
krustacea, dll (ekosistem akuatik)
2) Konsumer II (konsumer sekunder)adalah karnivora

dan omnivora yang memakan herbivora, contoh:
burung gagak, rubah, kucing, ular, dll.
3) Konsumer III (konsumer tertier) adalah karnivora
dan omnivora, misal singa, hari-mau, dll., disebut
juga Top-Konsumer
4) Parasit, Scavenger dan saprobe
C. Faktor Penyebab Perbedaan Ekosistem
Ekosistem yang satu berbeda dengan ekosistem yang
lain, karena:
1) Perbedaan kondisi iklim (hutan hujan, hutan musim,
hutan savana)
2) Perbedaan letak dari permukaan laut, topografi dan
formasi geologik (zonasi pada pegunungan, lereng
pegunungan yang curam, lembah sungai)
3) Perbedaan kondisi tanah dan air tanah (pasir,
lempung, basah, kering)
D. Macam dan Ukuran Ekosistem
Berdasarkan proses terjadinya ada dua macam
ekosistem, yaitu:
1) Ekosistem alam: laut, sungai, hutan alam, danau
alam, dll.
2) Ekosistem buatan: sawah, kebun, hutan tanaman,
tambak, all.
Ukuran ekosistem bervariasi dari sebetsar kultur
dalam botol di laboratorium, seluas danau, sungai,
lautan sampai biosfir ini.

Secara umum, ada dua tipe ekosistem, yaitu:
1) Ekosistem terestris
− Ekosistem hutan
− Ekosistem padang rumuput
− Ekosistem gurun
− Ekosistem anthropogen (sawah, kebun, dll.}
2) Ekosistem akuatik
(a). Ekosistem air tawar
- Kolam
- Danau
- Sungai
- dll.
(b). Ekosistem lautan
E. Tahap-tahap Dasar Operasi pada Ekosistem
1) Penerimaan energi radiasi
2) Pembuatan bahsn-bahan organik dari bahan
anorganik oleh produser
3) Pemanfaatan produser oleh konsumer dan lebih jauh
lagi pada bahan-bahan terkonsumsi
4) Perombakan bahan-bahan organik dari organisme
yang mati oleh dekomposer kedalam bentuk
anorganik sederhana untuk penggunaan ulang oleh
produser.
F. Ekologi Niche
Niche adalah peranan suatu mahkluk hidup dalam
suatu habitat. Sedangkan habitat adalah tempat hidup
organisme. Dengan demikian ekologi niche adalah peran

total dari suatu species dalam komunitas. Ekologi
niche mencakup species organisme, faktor lingkungan,
areal tempat hidup, spesialisasi dari populasi species
dalam suatu komunicas.
G. Energi dalam Ekosistem
- Energi adalah kemampuan untuk melakukan kerja.
- Bentuk energi yang berperan penting pada mahkluk
hidup adalah energi mekanik, kimia, radiasi dan
panas.
- Perilaku energi di alam mengikuti Hukum
Thermodinamika, yaitu:
Hukum Thermodinamika I:
Energi dapat diubah dari suatu bentuk ke bentuk
lainnya, tetapi energy tak pernah dapat diciptakan
atau dimusnahkan.
Hukum Thermodinamika II:
Setiap terjadi perobahan bentuk energi pasti terjadi
degradasi energi dari bentuk energi yang terpusat
menjadi bentuk energi yang terpencar atau karena
berbagai energi selalu memencar menjadi panas, tidak
ada transformasi secara spontan dari suatu bentuk
energi menjadi energi potensial berlangsung dengan
efisien 100%. Misal, 57% energi surya diserap atmosfir,
dan 35 % disebarkan untuk memanaskan air dan daratan.
Dari sekitar ±3% energi surya yang mengenai permukaan
tumbuhan, 10 - 15% dipantulkan, 5% ditransmit, 80 - 85%

diserap dan ±2% (0.5 -3,5%) dari total energi cahaya
digunakan fotosintesis serta sisanya dirubah menjadi
bentuk panas.
H. Rantai Pangan
Rantai pangan ada1ah pengalihan energi dari
sumberdaya dalam tumbuhan melalui sederetan organisme
yang makan dan yang dimakan. Semakin pendek rantai
pangan semakin besar energi yang dapat disimpan dalam
bentuk tubuh organisme di ujung rantai pangan.
Rantai pangan terdiri atas tiga tipe:
1) Rantai pemangsa, dimulai dari hewan kecil sebagi
mata rantai pertama ke hewan yang lebih besar dan
berakhir pada hewan terbesar dimana landasan
permulaan adalah tumbuhan sebagai produsen.
2) Rantai parasit, berawal dari organisme besar ke
organisme kecil.
3) Rantai saprofit, berawal dari organisme mati ke
mikroorganisme, dikenal juga sebagai rantai pangan
detritus. Dalam suatu ekosistem, rantai-rantai
pangan berkaitan satu sama lain membentuk suatu
jaring-jaring pangan (food web).
- Dalam suatu ekosistem dikenal adanya tingkat
tropik dari suatu kelompok organisme.
- Berbagai organisme yang memperoleh sumber
makanan melalui langkah yang sama dianggap
termasuk pada tingkat tropik yang sama.

- Berdasarkan tingkat tropik :
Tumbuhan hijau : tingkat tropik I
Herbivora : tingkat tropik II
Karnivora : tingkat tropik III
Karnivora sekunder : tingkat tropik IV
I. Struktur Tropik dan Piramida Ekologi
- Ukuran individu menentukan besarnya metabolisms
suatu organisme. Semakin kecil ukuran organisme,
semakin besar rnetabolisrne per gram biomassa. Oleh
karena itu, semakin kecil organisme semakin kecil
biomassa yang dapat ditunjang pada suatu tingkat
tropik dalam ekosistemnya.
- Fenomena interaksi antara rantai-rantai makanan dan
hubungan metabolisme dengan ukuran organisme
menyebabkan berbagai komunitas mempunyai struktur
tropik tertentu.
- Struktur tropik dapat diukur dan dipertelakan,
baik dengan biomassa per satuan luas maupun dengan
banyaknya energi yang ditambat per satuan luas per
satuan waktu pada tingkat tropik yang berurutan.
- Piramida ekologi dapat menggambarkan struktur dan
fungsi tropic: Ada tiga tipe paramida ekologi yaitu :
a) Piramida jumlah individu, yang menggambarkan
jumlah individu dalam produser dan konsumer
suatu ekosistem
b) Piramida biomassa, yang menggambarkan biomassa
dalam setiap tingkat tropik.
c) Piramida energi, yang menggambarkan besarnya

energi pada setiap tingkat tropik. Semakin tinggi
tingkat tropik, semakin efisien dalam
penggunaan energi.
J. Produktivitas
- Produktivitas primer adalah kecepatan
penyimpanan energi potensial oleh organisme
produsen melalui proses fotosintesis dalam bentuk
bahan-bahan organik yang dapat digunakan sebagai
bahan pangan. Unit satuannya:
1) Ash Free Dry Weight Kal./ha/th.
2) Dry Weight Ton/ha/th.
Produktivitas primer dibagi dua macam:
(1). Produktivitas primer kotor:
Kecepatan total fotosintesis, mencakup pula
bahan organic yang dipakai untuk respliasi
selama pengukuran. Istilah ini sama dengan
asimilasi total.
(2). Produktivitas primer bersih:
Kecepatan penyimpanan bahan-bahan organik dalam
jaringan tumbuhan sebagai kelebihan bahan yang
dipakai untuk respirasi oleh tumbuh-tumbuhan
selama pengukuran. Istilah ini sama dengan
asimilasi bersih.
- Produktivitas sekunder adalah kecepatan
penyimpanan energi potensial pada tingkat tropik
konsumen dan pengurai.

Produktivitas Primer kotor pada Ekosistem Akuatik
Ho. Ekosistem
Prod . Primer Kotor
Kcal/m2/th
1. Laut terbuka
1.000
2. Pesisir 2.000
3. Upwelling Zone
4. Estuari dan reefs
6.000
20.000
Produktlvitas Primer Kotor pada Ekosistem Terestris
No. Ekosistem
Kcal/m2/th
1. Gurun dan tundra 200
2. Padang rumput
3. Hut an lahan kering
4. Hutan konifer
2.500
2.500
3.000
5. Hutan temperate basah
6. Pertanian
8.000
12.000 7. Hutan tropik dan subtropik 20 .000
K. Siklus Biogeokimia
- Di alam telah diketahui ada ±100 unsur kimia,
tetapi hanya 30 - 40 unsur yang sangat diperlukan
oleh mahkluk hidup.
- Unsur-unsur kimia, termasuk unsur utama dari
protoplasma, cenderung untuk bersirkulasi dalam
biosfir dengan pola tertentu dari 1ingkungannya ke
organisme dan kembali lagi ke lingkungan, siklus
ini disebut siklus biogeokimia. Sedangkan,

pergerakan unsur-unsur dan senyawa-senyawa
anorganik yang penting untuk menunjang kehidupan
disebut siklus hara. Kedua siklus tersebut
masing-masing terdiri atas dua kompartemen atau
dua pool, yaitu :
1) Reservoir poo_l : besar, lambat bergerak,
umumnya bukan komponen ekologi.
2) Exchange atau Cycling pool : kecil, tapi lebih
aktif bertukar dengan cepat antara organisme
dengan lingkungannya.
Dilihat dari sudut biosfir secara keseluruhan,
siklus biogeokimia terdiri atas :
a) Tipe gas, dimana reservoir adalah di atmosfir
atau hidrosfir {lautan), misal siklus Karbon
(CO2) dan siklus Nitrogen (N)
b) Tipe sedimen, dimana reservoir adalah di kerak
bumi, misal siklus Posfor

III.HUTAN SEBAGAI KOMUNITAS TUMBUHAN
Hutan adalah masyarakat tumbuh-tumbuhan yang
dikuasai pohon-pohon yang menempati suatu tempat
dan mempunyai sadaan lingkungan yang berbeda
dengan di luar hutan. Sedangkan satuan masyarakat
hutan adalah tegakan. Karakteristik Komunitas
Tumbuhan
1. Perkembangan Komunitas adalah sejarah
pembentukan dan evolusi komunitas atau tahap-
tahap suksesi.
2. Organisasi Komunitas adalah struktur, komposisi
jenis dan organisasi tropic suatu komunitas.
Struktur Komunitas terdiri atas:
- Struktur vertikal (stratifikasi)
- Struktur horizontal (distribusi spatial jenis)
- Kelimpahan atau "abundance" (kerapatan,
biomasa).
3. Fungsi Komunitas adalah pola metabolisme,
produktivitas serasah dan laju pembusukannya,
siklus hara, aliran energi.
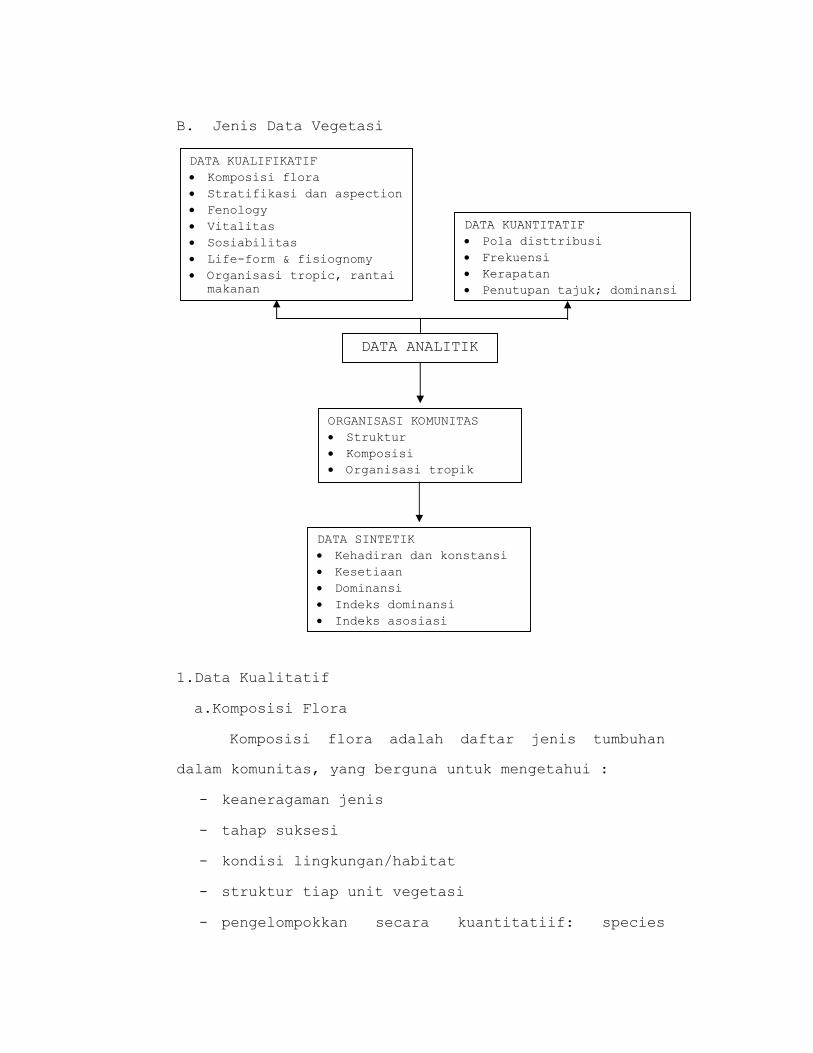
B. Jenis Data Vegetasi
1.Data Kualitatif
a.Komposisi Flora
Komposisi flora adalah daftar jenis tumbuhan
dalam komunitas, yang berguna untuk mengetahui :
- keaneragaman jenis
- tahap suksesi
- kondisi lingkungan/habitat
- struktur tiap unit vegetasi
- pengelompokkan secara kuantitatiif: species
DATA KUALIFIKATIF • Komposisi flora • Stratifikasi dan aspection • Fenology • Vitalitas • Sosiabilitas • Life-form & fisiognomy • Organisasi tropic, rantai
makanan
DATA KUANTITATIF • Pola disttribusi • Frekuensi • Kerapatan • Penutupan tajuk; dominansi
DATA ANALITIK
ORGANISASI KOMUNITAS • Struktur • Komposisi • Organisasi tropik
DATA SINTETIK • Kehadiran dan konstansi • Kesetiaan • Dominansi • Indeks dominansi • Indeks asosiasi

dominan, frequent (daya adaptasi luas), jenis
yang jarang (indikator habitat).
b. Stratifikasi dan "aspection"
Stratifikasi adalah lapisan vertikal komunitas
tumbuhan.
Stratifikasi terdiri :
- pucuk
- akar Manfaat Stratifikasi :
- optimalisasi ruang tumbuh
- peningkatan pemanfaatan energi solar
- optimalisasi pemanfaaCan unsur hara tanah. Aspect
ion adalah perubahan per:ampakan vegetasi dalam
kaitannya dengan musim.
c. Fenologi
Fenologi adalah kalender fase-fase pertumbuhan
yang dilalui oleh suatu tumbuhan selama sejarah
hidupnya, atau studi tentang fase-frase pertumbuhan
penting dalam sejarah hidup suatu tumbuhan, seperti:
saat biji berkecambah, gugur daun, berbunga, berbuah
dan tersebarnya biji.
Tanda proses fenologi
Masa kecambah :/
Masa berbunga :/
Masa berbuah : / /
Masa penyebaran biji : / /

Vitalitas dan Vigor
Vitalitas adalah kondisi dan kapasitas tumbuhan
untuk menyelesaikan siklus hidupnya. Sedangkan vigor
adalah keadaan kesehatan tumbuhan.
Klasifikasi vitalitas :
Klas 1 : Tumbuhan yang berkembang baik dan
dapat menyelesaikan siklus hidupnya.
Klas 2 : Tumbuhan yang tumbuh sehat yang
tersebar secara vegetatif.
Klas 3 : Tumbuhan yang lemah yang tersebar secara
vegetatif dan tak pernah menyelesaikan siklus
hidupnya.
Klas 4 : Tumbuhan yang jarang tumbuh dari biji, tetapi
jumlahnya tak bertambah.
Sosiabilitas
Sosiabiiitas adalah hubungan antara masing-
masing jenis dan menunjukkan cara tumbuhan tersebar.
Sosiabilitas bergantung pada :
- life-form
- vigor
- kondisi habitat
- kemampuan bersaing.
Klas Sosiabilitas (Brown-Blanquet, 1932):
Klas 1 : Hidup menyendiri.
Klas 2 : Agak mengelompok.

Klas 3 : Mengelompok dalam kelompok-kelompok yang
tersebar.
Klas 4 : Mengelompok dalam kelompok yang besar dan
kelompok terputus-putus.
Klas 5 : Membentuk hamparan yang luas dan rapat.
Life-form (bentuk hidup) tumbuhan
- bisa menggunakan klas-klas life-form dari Raunkaier
(1934), Brawn-Blanquet (1951), Backer (1968) :
pohon, semak, liana, epifit, pakuan , herba, lumut,
dll.
- Persentase Life-form adalah Σ species dalam suatu life-form —————————————————————————————— X 100% Σ species dalam semua life-form
- Species dari life-form yang berbeda dapat hidup
berasosiasi, karena mereka memanfaatkan sumberdaya
alam pada waktu/ruang yang berbeda.
Organisasi tropik dan rantai pangan
Rantai pangan ada1ah pengalihan energi dari sumbernya
berupa tumbuhan melalui sederetan organisma yang
memakan dan yang dimakan.
Ada dua tipe rantai makanan :
a) "Grazing food chain" :
Rantai pangan yang dimulai dari tumbuhan, terus
ke herbivora dan karnivora.
b) "Detritus food chain" :

Rantai pangan yang dimulai dari organisme mati ke
mikroorgnisme, detrivor dan predatornya.
Jaring-jaring pangan ("food web") adalah
keterkaitan antara berbagai rantai makanan dalam suatu
komunitas. Species diversity meningkat maka "food
chain" makin panjang. Studi food chain dalam komunitas
sangat berguna untuk mengetahui sistem transfer energi
dalam komunitas.
2. Data Kuantitatif
a. Distribusi Spasial Individu tumbuhan Tiga tipe Pola
Distribusi
1) Random (acak)
Pola ini mencerminkan homogenitas habitat dan/atau
pola behavior yang tidak selektif.
2) Mengelompok ('clumped')
Mencerminkan habitat yang heterogen, mode
reproduktif, behavior berkelompok, dll.
3) Beraturan (reguler, uniform)
Mencerminkan adanya interaksi negatif antara
individu seperti persaingan untuk ruang dan unsur
hara atau cahaya. Faktor yang mempengaruhi pola
sebaran spatial individu:
a) Faktor vektorial dari aksi berbagai tekanan
lingkungan luar (angin, aliran air, intensitas
cahaya).

b) Faktor reproduksi sebagai akibat dari mode
reproduktif organisme (cloning dan regenerasi
progeni).
c) Faktor sosial akibat pembawaan behavior (misal,
behavior teritorial)
4) Faktor koaktif akibat dari interaksi intraspecific
(misal kompetisi).
5) Faktor stokastik akibat dari variasi acak dari
berbagai faktor tersebut di atas, yaitu :
a) faktor intrinsik species (mis., reproduktif,
sosial, koaktif)
b) Faktor extrinsic (vector).
Beberapa indeks penentuan poia Distribusi Spasial
individu
(1). Variance Mean Ratio
V/M = 1 (random)
V/M > 1 (clumped)
V/M < 1 (regular)
Untuk menguj i apakah V/M < 1 atau >
1,digunakan uji X2 dengan derajat bebas (q - 2) ,
dimana q = Σ frekuensi klas, pada tingkat peluang
1%, 5%. Contoh :

Ada 100 petak
Σ Ind. Sp-X dalam masing-
masing kuadrat
0 1 2 3
Frekuensi kehadiran dalam
100 petak
46 34 14 6
0(46)+1(34)+2(14)+3(6) Mean (M)= 100
= 0.8
ΣX2 – (ΣX)2/n Variance = n-1 [12(34)+22(14)+32(6)+02(46)] – (80)2/100 = 100 - 1 = 0.808 V/M = 0.808/0.800 = 1.01 Pengujian V/M = 1? 1). Menghitung banyaknya petak yang mengandung 0,1,2,3
individu Є(0) = (n)p(0) = (100)p(0) = (100)e-0.8 = 44.9 Є(1) = (n)p(1) = (100)(0.8/1)(p(0)) = 100 x 0.8/1 x 0.4493 = 0.3594 Є(1) = m e-m x n = 0.8 x e-0.8 x 100 = 0.8 x 0.4493 x 100 = 0.3594 Є(2) = 0.82/2! x e-m x 100 = 0.64/2 x 0.4493 x 100 0.1438 x 100 = 14.4

Є(3) = 0.83/3! x e-m x 100 = 0.512/6 x e-0.8 x 100 = 0.512/6 x 0.4493 x 100 = 3.8
1 Σ individu/petak 0 1 2 3 2 Σ petak terobservasi 46 34 14 6 3 Σ petak harapan 44.9 35.9 14.4 3.8 4 Perbedaan Σ petaj antara
terobservasi dan harapan 1.1 1.9 0.4 2.2
X2 hitung = (Obs – є)2/є (1.1)2 + (1.9)2 + (2.2)2 = 44.9 35.9 3.8 = 1.4123 X2 tabel(q-2), dimana q = Σ klas frekuensi = 4 X2 (α=0.5,2) = 1.386 = 1.4
Sehingga
X2 hitung = X2 tabel random
2). Indeks Morisita (IS)
( )
( )1
2
1
11−
=∑ −
TTIS
XiXi
Dimana : Xi = jumlah individu species X dalam petak
ke-I (i=1,2,3,………,q)
q = jumlah seluruh petak
T = jumlah total individu dalam semua
petak
Kriteria : IS = 1 (random)
IS > 1 (clumped)
IS < 1 (regular)

Pengujian IS = 1? IS (T-1)+ q – T Fo = Q – 1 Bila Fo ≥ Fαq-1 Clumped
(α = 0.05 atau 0.01)
3). Green’s Index
1
1
−
−⎟⎠⎞
⎜⎝⎛
=n
MeansVariance
GI
GI bervariasi dari: 0 sampai maximum.
0 = random, 1 = clumping.
b. Kerapatan
Adalah jumlah suatu spesies dalam suatu unit
area. Kerapatan menunjukkan kelimpahan suatu spesies
dalam suatu komunitas.
Satuan : ind/m2 (tumbuhan bawah)
Ind/ha (pohon)
Kerapatan relative: persentase kerapatan suatu spesies
terhapdap jumlah kerapatan semua spesies.
c.Frekuensi
Frekuensi adalah derajat penyebaran suatu jenis
di dalam komunitas yang diekspresikan sebagai
perbandingan antara banyaknya petak yang diisi oleh
suatu jenis terhadap jumlah petak contoh seluruhnya.
Frekuensi Relatif : persentase frekuensi suatu
species terhadap jumlah frekuensi semua species.

Frekuensi Klas (Raun kaier, 1934} :
Klas A: species dengan frekuensi 1 - 20%
Klas B: species dengan frekuensi 21 - 40%
Klas C: species dengan frekuensi 41 - 60%
Klas D: species dengan frekuensi 61 - 80%
Klas E: species dengan frekuensi 81 - 100%
"Law of Frequency"
>
A>B>C=D<E
<
(Persentase Frekuensi berdistribusi normal) Jika :
(1) E > D : Komunitas Homogen
(2) E < D : Komunitas terganggu
(3) A, E tinggi : Komunitas buatan
(4) B,C,D tinggi: komunitas heterogen
4.Cover (Penuntupan Tajuk)
Cover adalah proyeksi vertikal tajuk terhadap
permukaan tanah. Tajuk adalah semua bagian tanaman yang
terdapat di atas permukaan tanah. Di dalam hutan, cover
harus ditentukan untuk setiap strata vegetasi, sehingga
cover bisa > 100 %.
Di dalam komunitas rumput, cover digambarkan
dalam "graph paper" dengan bantuan kuadrat (misal, 25 X
25 cm)atau menggunakan plantigraph.
Klas Penutupan Tajuk

Klas A : Species dengan cover 5%
Klas B : Species dengan cover 6 - 25%
Klas C : Species dengan cover 26 - 50%
Klas D : Species dengan cover 51 - 75%
Klas E : Species dengan cover 76 - 100%
Foliage cover meningkat ————-> Intercepting solar
energi meningkat . Naungan meningkat
Pengukuran foliage cover bisa diganti dengan
"basal area" (luas bidang dasar, Ibds).
3. Data Sintetik
Presence
Presence adalah suatu kehadiran species dalam
komunitas.
Klas Kehadiran
- Jarang : 1 - 20 % petak contoh terisi species.
- Kadang terdapat : 21 - 40 petak
contoh terisi species.
- Sering terdapat: 41 – 60% terisi spesies
- Banyak terdapat : 61 - 80 terisi species.
- Selalu ada : 81 - 100 % petak contoh
terisi species.
Constance (Kontansi)
Constance adalah derajat/tingkat kehadiran suatu
species dalam komunitas. Klas Konstansi
Klas 1 : 1 - 20 % Frekuensi

Klas 2 : 21 - 40 % Frekuensi
Klas 3 : 41 - 60 % Frekuensi
Klas 4 : 61 - 80 % Frekuensi
Klas 5 : 81 - 100 % Frekuensi
c. Dominansi Jenis
Jenis dominan adalah jenis yang bei k..i :\sa dan mencirikan
suatu komunitas. Konsep dominansi jenis sebagai petunjuk :
- species tersebut menang dalam persaingan
- species tersebut mempunyai toleransi tinggi
- species tersebut berhasil beradaptasi
- terhadap habitat . Parameter Penentu Dominansi Jenis
- Foliage Cover (penutupan tajuk)
- Kerapatan
- Luas Bidang Dasar
- Biomasa
- Volume
- Indeks Nilai Penting (INP)
INP = Kerapatan Relatif + Frekuensi Relatif + Dominansi
Relatif INP maksimal 300%.
Dominansi adalah luas penutupan tajuk atau luas bidang dasar
suatu species dalam satuan unit area tertentu. Satuannya:
M/ha.
Dominansi Relatif adalah persentase dominansi suatu species
terhadap jumlah dominansi seluruh jenis.

d. Fidelity {Kesetiaan)
Fidelity adalah tingkat kesetiaan suatu species
dalam suatu komunitas. Klas Kesetiaan Jenis:
Klas 1: Ekslusif terhadap suatu jenis komunitas.
Klas 2: Selektif (sering berada pada satu macam
komunitas,tetapi tidak pada komunitas lain).
Klas 3 : Preferensial {berada pada beberapa habitat, tetapi
tumbuh banyak pada beberapa habitat saja).
Klas 4 : Indifferent/masa bodoh (berada secara teratur pada
semua habitat).
Kals 5 : Strange/aneh (jarang dan secara kebetulan berada
dalam komunitas).
e. Indeks of Dominance (ID)
Indeks of dominance adalah indeks untuk memeriksa tingkat
dominansi suatu species dalam komunitas.
Nilai ID tinggi dominansi jenis dipusatkan pada satu
atau beberapa jenis. Nilai ID rendah dominansi jenis
dipusatkan pada banyak jenis.
Simpson (1949)
ID = C = E (ni/N)2
C = indeks of dominance
ni = INP atau kerapatan atau biomasa suatu species.
N = Total INP atau total kerapatan ,atau biomasa dari
semua species.
Hilai C ini bersifat relatif. Nilai C bisa digunakan
apakah suatu komunitas itu asosiasi atau konsosiasi.

f. Interspecific Assosiation
Interspecific assosiation ada1ah suatu
asosiasi/kekariban antara dua species dalam komuninas.
Interspecific Assosiation terjadi bila :
- kedua species tumbuhan pada lingkungan yang
serupa.
- distribusi geografi kedua species ;>erupa dan
keduanya hidup di daerah yang sama.
- kedua jenis berbeda life-form.
- bila salah satu species hidupnya bergan-tung pada
yang lain.
- bila salah satu species menyediakan per lindungan
terhadap yang lain.
Metode mendeteksi interspesif ic asosiasi. ion
(1). Data Kualitatif
(a). 2x2 contingency table, bila datanya kualitatif
(hadir atau tidak).
Spesies A
Hadir +
Tidak 0
+ a b m=a+b
0 c d n=c+d
E a+b=r b+d=r N=a+b+c+d
(ad-bc)2 X N
X2 hit = m X n X r X s
nilai X2 ini bandingkan dengan
X2tab (α = 0.05, db = 1)
Bila X2hit ≥ X2tab ada asosiasi
S P E S I E S B

a = Σ petak dimana 2 spesies ada
b = Σ petak, sp. A ada, sp. B tak ada
c = Σ petak, sp. A tak ada, sp. B ada
d = Σ petak, sp. A dan B tak ada
N = Σ total petak contoh
(b). JACCARD INDEX (JI)
cbaaJI++
=
(2). Data Kuantitatif
Koefisien Korelasi Σ[(X1-X1)(X2-X2)] R hit = √[Σ(X1-X1)2 x (Σ(X2-X2)2]
R hit. ≥ R tab. Untuk p = 0.05 atau p = 0.01
g. Index of Diversity
• keanekaragaman jenis adalah suatu parameter
penting dalam membandingkan dua komunitas,
terutama untuk mempelajari pengaruh gangguan
biotic atau mengetahui tahap suksesi dan
stabilitas komunitas
• pada komunitas klimak, spesies diversity meningkat
food chain meningkat komunitas stabil
Respirasi komunitas • Ecological turnover =
Biomassa komunitas R/B rendah komunitas diversity meningkat

• Metode/cara penentuan spesies diversity
1) Shanon-Weiner Diversity Index
H = -Σ[(ni/N) log (ni/N)]
ni = Nilai kuantitatif suatu spesies
N = jumlah nilai kuantitatif semua spesies
dalam komunitas
Variasi nilai H
0 = satu spesies tak terhingga nilai yang tinggi
(banyak spesies)
2) Simpson’s Diversity Index
∑=
−=s
iNniD
1
2)/(1
S = Σ jenis
Variasi nilai D:
0 = satu spesies tak terhingga
1 – 1/s = diversity spesies max.
h. Koefisien Kesamaan Komunitas (Index of Similarity)
• Index ini sangat berguna untuk membandingkan
kesamaan jenis dua komunitas
• Caranya:
1). Jaccard’s presence-community coefficient
ISJ = [C/(A+B+C)] x 100%
A = Σ jenis di komunitas 1
B = Σ jenis di komunitas 2
C = Σ jenis di dua komunitas
2). Motyka’s Index of Similarity
IS = [2 Mw/(Ma + Mb)] x 100%
Mw = Σ nilai kuantitatif ≤ dari spesies yang
ada di dua komunitas

Ma = Σ nilai kuantitatif semua spesies di
komunitas 1
Mb = Σ nilai kuantitatif semua spesies di
komunitas 2
Nilai IS : 0 – 100
C. Fungsi Komunitas
1. Biomassa
Biomassa adalah jumlah bahan organic yang
diproduksi oleh organism per satuan unit area
pada suatu saat. Satuannya g/m2 atau Kg/ha.
• Biomassa menunjukkan net production
• Biomassa production rate adalah laju akumulasi
biomassa dalam kurun waktu tertentu (Kg/ha/yr)
• Biomassa dinyatakan dalam “dry weight” (berat
kering) oven pada suhu 105o selama 12 jam atau
800C selama 48 jam. Satuan lain adalah berat
kering bebas abu (“ash free dry weight”)
• Biomassa profil menunjukkan jumlah bahan
organic kering pada tingkat yang berbeda dari
komunitas
• Akumulasi biomassa di tropic lebih rendah
daripada di temperate karena laju respirasi di
tropic lebih tinggi
2. Aliran Energi
• Dari sudut energy, komunitas adalah unit
thermodinamika
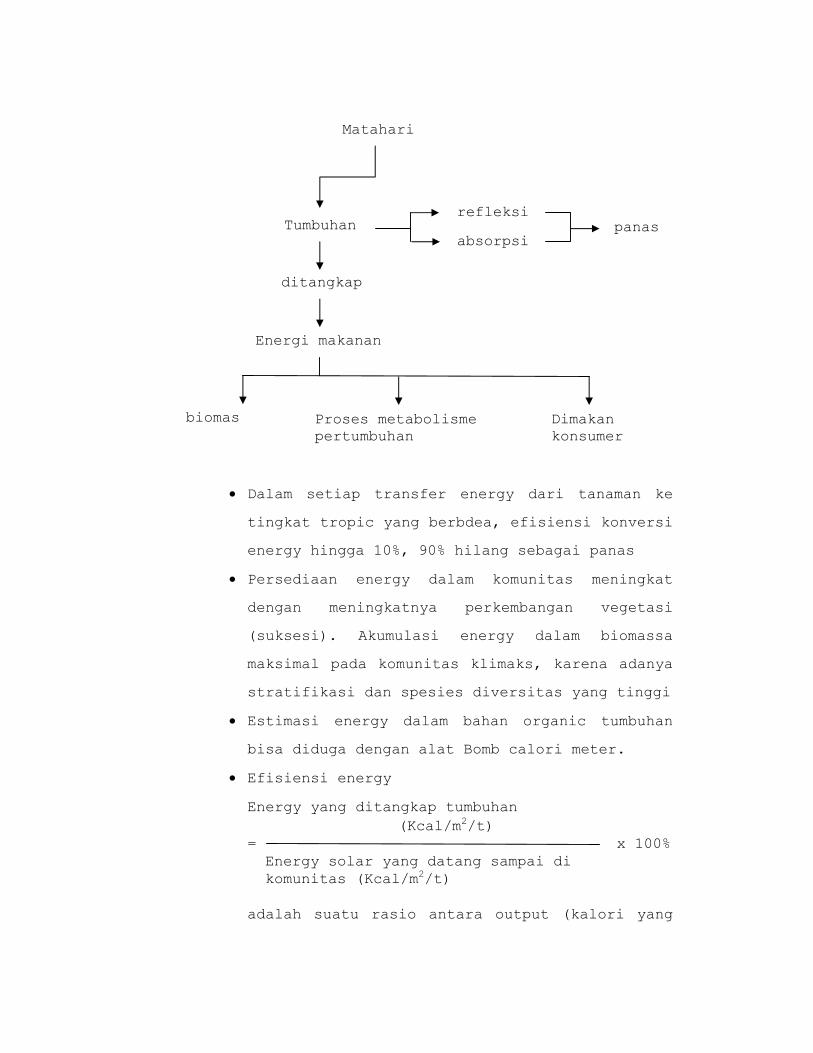
• Dalam setiap transfer energy dari tanaman ke
tingkat tropic yang berbdea, efisiensi konversi
energy hingga 10%, 90% hilang sebagai panas
• Persediaan energy dalam komunitas meningkat
dengan meningkatnya perkembangan vegetasi
(suksesi). Akumulasi energy dalam biomassa
maksimal pada komunitas klimaks, karena adanya
stratifikasi dan spesies diversitas yang tinggi
• Estimasi energy dalam bahan organic tumbuhan
bisa diduga dengan alat Bomb calori meter.
• Efisiensi energy
Energy yang ditangkap tumbuhan (Kcal/m2/t)
= x 100% Energy solar yang datang sampai di komunitas (Kcal/m2/t)
adalah suatu rasio antara output (kalori yang
Matahari
Tumbuhan
ditangkap
Energi makanan
biomas Proses metabolisme pertumbuhan
Dimakan konsumer
refleksi
absorpsipanas

dimanfaatkan tumbuhan) terhadap input (energy
solar sampai di komunitas) dalam suatu unit
area dalam periode waktu tertentu.
• Efisiensi energy adalah rasio antara aliran
energy di setiap titik/tahap yang berbeda
sepanjang rantai makanan, satuannya %.
3. Gross Ecological Effisiency (GEE)
Kalori mangsa yang dikonsumsi pemangsa = x 100% Kalori makanan yang dikonsumsi mangsa
• Siklus hara, Produktivitas dan Dekomposisi
Serasah
• Siklus Biogeokimia, termasuk unsure-unsur utama
dari protoplasma, dari lingkungan ke organism
dan kembali lagi ke lingkungan dalam biosfir
• Siklus hara adalah pergerakan unsur-unsur dan
senyawa-senyawa yang penting bagi kehidupan
Tipe Gas - Siklus N - Siklus CO2
Tipe batuan - Siklus fosfor
Siklus Biogeokimia
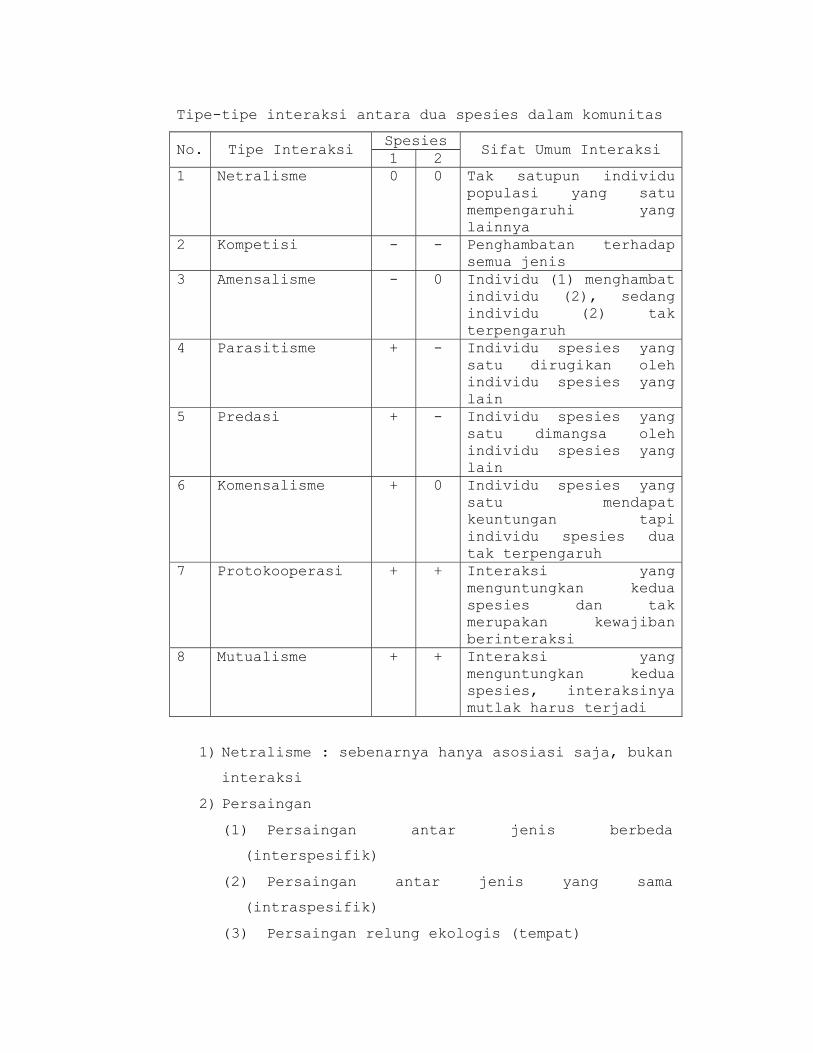
Tipe-tipe interaksi antara dua spesies dalam komunitas
No. Tipe Interaksi Spesies
Sifat Umum Interaksi 1 2
1 Netralisme 0 0 Tak satupun individu populasi yang satu mempengaruhi yang lainnya
2 Kompetisi - - Penghambatan terhadap semua jenis
3 Amensalisme - 0 Individu (1) menghambat individu (2), sedang individu (2) tak terpengaruh
4 Parasitisme + - Individu spesies yang satu dirugikan oleh individu spesies yang lain
5 Predasi + - Individu spesies yang satu dimangsa oleh individu spesies yang lain
6 Komensalisme + 0 Individu spesies yang satu mendapat keuntungan tapi individu spesies dua tak terpengaruh
7 Protokooperasi + + Interaksi yang menguntungkan kedua spesies dan tak merupakan kewajiban berinteraksi
8 Mutualisme + + Interaksi yang menguntungkan kedua spesies, interaksinya mutlak harus terjadi
1) Netralisme : sebenarnya hanya asosiasi saja, bukan
interaksi
2) Persaingan
(1) Persaingan antar jenis berbeda
(interspesifik)
(2) Persaingan antar jenis yang sama
(intraspesifik)
(3) Persaingan relung ekologis (tempat)

(4) Persaingan sumberdaya (makanan)
Akibat persaingan:
- Pertumbuhan tewrganggu
- Produksi berkurang, jumlah biji sedikit
- Menstimulasi serangan hama-penyakit dan
kekurangan unsure hara
- Terjadi stratifikasi dimana jenis tertentu
lebih berkuasa
- Komposisi jenis berubah (Σ jenis, Σ individu,
life-form).
Competitif Ability
Ditentukan secara sederhana dengan rumus:
GA/B = MA/MB atau
GB/A = MB/MA
G = kemampuan pertumbuhan
M = bobot kering tanaman
A,B = spesies A dan B
3) Amensalisme, merupakan persaingan dalam bentuk yang
lemah, adalah hubungan antara individu yang mana
individu yang satu dirugikan (tetapi sesaat) tetapi
individu lain tidak dirugikan (netral). Amensalisme
merupakan persaingan dalam bentuk yang lemah. Contoh
: allelopathy yaitu pengaruh merugikan baik langsung
maupun tak langsung dari suatu tumbuhan terhadap
tumbuhan lain melalui produksi senyawa kimia. Dalam
hal ini, bahan kimia dapat dikategorikan sebagai :
(a). Autotoxic (bahan penghambat) terhadap :
- anakan sendiri
- individu lain sejenis
(b). Antitoxic (bahan penghambat) terhadap individu
lain jenis berbeda.

Cara tanaman melepaskan bahan kimia (bahan
allelopati)adalah melalui :
- pencucian daun/batang oleh air hujan
- bahan tanaman yang jatuh sebagai aerasah yang
menjadi humus dalarn tanah.
- gas yang menguap dari permukaan tanaman
- eksudat akar
Media pengeluaran zat alelopatik
jenis tanaman
1.
Daun
Camelina
2. Akar Eucalyptus globulus
3. Setelah mati Apel, sereh
4.
Gas
Reliant bus, Aster
Bahan kimia allelopathic diantaranya adalah
- phenolic, terpeties, alkaloids, nitrit difenol,
- asam benzoat, fenin, sulfida. Pengaruh allelopathy
terhadap pertumbuhan tumbuhan :
- perpanjangan/perbanyakan sel terhambat
- penyerapan hara mineral berkurang
- laju fotosintesa dan respirasi terganggu
- perlambatan perkecambahan biji
- laju pertumbuhan terhambat
- gangguan sistem perakaran
- klorosis
- layu, mati

Parasitisme (+,-)
Suatu organisme untuk hidupnya mengambil makanan
dari organisme lainnya. Interaksi parasitisme
memungkinkan adanya tumbuhan inang (host) dan tumbuhan
parasit.
Host seringkali mengeluarkan antibodi Parasit
yang heterofog lebih bertahan daripada monoloq.
Parasit meliputi parasit akar -» Rafflesia, semipara-sit
{yang tumbuh di cabang-cabang di pohon -> benalu (famili
horuntuceae).
Rafflesia -» bunga liar (famili Rafflesiaceae)
Genus lain : - Rhizanthes
- Mitrastemon Di Sumatera 4 jenis :
- Rafflesia atjehensis
- Rafflesia hasseltii
- Rafflesia arnoldi
- Rafflesia patma. Rafflesia
-» paling khas diantara parasit lain
- besar ukuran bunga
- tidak punya batang, daun dan akar
- hanya punya benang-benang yang tumbuh
di bagian dalam batang dan akar pohon.
Inang (Tetrastigma, famili vitaceae)
- waktu bunga lama, tergantung ukuran R.
arnoldi
- kuncup terbuka mekar (19-21 bulan)0 10 cm

H. 5 bulan 0 15 cm -» 2 bulan 0 25 cm -*
20-30 hari.
(5). Commensalisme ( + , 0)
Interaksi antara individu yang memberikan keuntungan
kepada salah satu individu jenis populasi sementara
yang lain tak memperoleh keuntungan apa-apa (netral).
Merupakan hubungan (+) yang mendasari protokoperasi.
Contoh Epifit: paling banyak terdapat di hutan
hujan tropika (10% pohon hutan hujan tropika ditumbuhi
epifit).
- Anggrek, paku-pakuan, dll.
- Menempel pada batang atau daun (epifit)
Setelah dapat sinar matahari akan menutupi
tajuk.
- liana (tumbuhan merambat suka cahaya =
heliophyta)
- pengaruh negatif liana
1. Menutupi daerah tajuk sehingga
mengurangi proses fotosintesis.
2. Menurunkan kualitas kayu
3. Mengganggu tumbuhan pohon yang dipanjati
4. Berpengaruh negatif terhadap anakan yang suka
cahaya (heliophyta)
- pengaruh positif, diantaranya adalah berpengaruh
baik pada pertumbuhan anakan yang suka naungan
(schyophyta, misalnya jenis-jenis anggota

Dipterocarpaceae)
- sistem silvikultur (tropical shelter-wood system)
penangkaran liana (pembebasan)/tebang penerang
(6). Protocoperasi (+,-)
Kedua jenis individu yang berinteraksi mendapat
keuntungan tetapi bukan merupakan keharusan untuk
saling berhubungan. contoh : Asosiasi lumut dengan
keong air tawar
- Lumut menggunakan zat hara dari keong
- Keong ditumbuhi lumut sebagai perlindungan
Protocoperasi merupakan awal evolusi sebelum
mutualisme.
(7). Mutualisme (+,+)
Memberikan keuntungan kepada masing-masing jenis
yang berinteraksi dan merupakan suatu keharusan untuk
hidupnya, jika dipisahkan akan rugi. Contoh :
- Mikoriza : asosiasi antara jamur dengan akar
tumbuhan. Jamur merubah unsur-unsur sehingga
tersedia dan dapat dihisap oleh akar tumbuhan,
jamur mendapatkan makanan dari hasil fotosintesa
inang.
- Jenis mikoriza adalah:
a) Ektotropik: di luar akar mis: Basidiomycetes
b) Endotropik: di dalam akar mis: Phycomycetes
c) Peritropik: sebagai mantel, contoh: Mikoriza

ekstra material
Ektotropik : Micorhyza di bagian luar sel akar micelia fungi, misal pada Pinus strobus, Dipterocarpaceae, Eucalyptus. Endotropik: Micorhyza di bagian dalam sel akar micelia fungi, yakni hampir semua tanaman kecuali tanaman air. Peritropik: Micorhyza membentuk selubung mantel rongga yang mengelilingi akar, misal pada anakan spruce (Picea pungens).
- Karena tanah Imtan Indonesia relatif miskin hara,
maka banyak pohon-pohon hutan alam yang mengandung
mikoriza.
- Di hutan Cibodas 32% pohon-pohon yang ada mengandung
mikoriza.
- Mikoriza mengeluarkan enzim phosphatase
- Manfaat mikoriza
a) penyerapan unsur hara meningkat terutama Phospor
b) mencegah infeksi perakaran mempertinggi daya
tahan kekeringan akar lebih lama hidup
(memproduksi hormon penumbuh).
- Nodul Akar : gejala pembengkakan akar berupa bintil
akar sebagai akibat sirnbiosis mutualisme antara
bakteri (rhizobium/aktinomisetes) dengan suatu akar
tumbuhan tertentu.

Bakteri rhizobium adalah pengikat N tumbuhan
mendapatkan Nitrogen, rhizobium mendapatkan karbohidrat
berdasarkan jenis tanaman dengan mikroba pembentuk
nodul, maka ada tiga bentuk simbiosa:
1. Legume, (rhizobium)
(Albizia, Akasia, Leucoem -» Leguminosae) tidak
semua legum berasosiasi dengan rhizobium
2. Non Legume, (rhizobium)
(Trema, pnrasponia}
3. Non Legume, (Aktinomisetes) (Frankia)
(Casuarina, Podocarpus)
- Keuntungan adanya nodul akar:
1. Tanaman inang bisa hidup pada tanah miskin N
2. Dapat meningkatkan kesuburan tanah
3. Memungkinkan tanaman tumbuh setelah tanaman
legume
Hewan Hutan, berperan besar dalam pembiakan tanaman,
misal beberapa jenis pohon dalam pembuahan dan
penyerbukan biji/benih tergantung pada hewan tertentu :
serangga, burung, kelelawar, babi hutan, musang, dll.
Tetapi hewan juga bisa merusak tanaman (hama) dan
penular penyakit pada tanaman.
Leguminosae Mimosaceae Caesalpiniaceae Papilionaceae
Jarang

IV. DINAMIKA MASYARAKAT TUMBUH-TUMBUHAN (SUKSESI)
A. Pengertian Suksesi (Sere)
Spurr (1964), mengatakan bahwa suksesi merupakan
proses yang terjadi secara terus-menerus yang ditandai
oleh perubahan vegetasi, tanah dan iklim dimana proses
ini terjadi. Sedangkan Costing (1956), menyatakan bahwa
perubahan-perubahan bertahap atau proses suksesi ini
berlangsung karena habitat tempat tumbuh masyarakat
tumbuhan mengalami modifikasi oleh beberapa daya
kekuatan alam dan aktivitas organisme berupa perubahan-
perubahan terhadap tanah, air, kimia dan lain-lain.
Perubahan masyarakat tumbuhan dimulai dari
tingkat pionir sederhana sampai pada tingkat klimaks,
dalam hal ini tumbuhan pioner merubah habitatnya
sendiri sehingga cocok untuk species baru, keadaaan ini
berlangsung terus hingga tingkat klimak tercapai
(Clements, 1923; halle, 1.97G; Clark, 1954, Ewuse,
1980).
Tentang adanya perubahan habitat, dinyatakan
bahwa komunitas pertama akan merubah keadaan tanah dan
iklim mikro. Dengan demikian memungkinkan masuknya
species kedua yang menjadi dominan dan mengubah keadaan
lingkungan dengan cara mengalahkan species yang pertama
dan hal ini memungkinkan masuknya species yang ketiga,
demikian seterusnya sampai tingkat klimaks tercapai
(Whittaker, 1970; Odurn, 1970; Whitmore,1975)

Secara singkat suksesi adalah suatu proses
perubahan komunitas tumbuh-tumbuhan secara teratur
mulai dari tingkat pionir sampai pada tingkat klimaks
di suatu tempat tertentu
Komunitas klimaks adalah komunitas yang berada
dalam keadaan keseimbangan dinamis dengan
lingkungannya. Sedangkan tingkat sere adalah setiap
tingkat/tahap dari sere, dan komunitas sere adalah
setiap komunitas tumbuhan yang mewakili setiap tingkat
sere.
Species klimak adalah suatu species yang berhasil
beradaptasi terhadap suatu habitat sehingga species
tersebut menjadi dominan di habitat yang bersangkutan.
S. Faktor Penyebab Suksesi
1. Faktor Iklim
- fluktuasi kondisi iklim yang tidak konsisten
- kekeringan
- radiasi yang kuat
- dan lain-lain yang merusak vegetasi sehingga
terjadi suksesi.
2. Faktor Topografi/Edafis
Faktor ini berkaitan dengan perobahan dalam
tanah. Ada 2 faktor penting yang berkaitan dengan tanah
yang membawa perobahan habitat, yaitu:
a. Erosi tanah, yaitu suatu proses hilangnya lapisan

permukaan tanah oleh angin, aliran air dan hujan.
b. Deposisi tanah, yaitu proses pengendapan/
penimbunan tanah oleh angin, longsor, glacier atau
turunya salju di suatu tempat.
3. Faktor biotik penyebab rusaknya vegetasi yang
mengakibatkan suksesi adalah :
- penggembalaan
- penebangan
- deforestasi
- hama dan penyakit
- perladangan
- dan lain-lain
C. Tipe-tipe Suksesi
1. Hidrosere
Hidrosere adalah suksesi tumbuhan yang terjadi di
habitat air atau basah".
2. Halosere
Halosere adalah suksesi tumbuhan yang terjadi di
tanah/air masin.
3. Xerosere
Xerosere adalah suksesi tumbuhan yang terjadi di
habitat kering. Tumbuhan pionirnya berupa lumut
kerak,bakteria,dan ganggang.
4. Psammosere
Psammosere adalah suksesi tumbuhan yang terjadi

di habitat berpasir.
5. Lithosere
Lithosere adalah suksesi tumbuhan yang terjadi di
permukaan batuan.
6. Serule
Serule adalah miniatur suksesi mikroorganisme
bakteri, jamur, dll) pada pohon yang mati, kulit
pohon, dll.
D. Tahab-tahab Suksesi
Shukla dan Chandel (1932) mengemukakan
sembilan macam tahapan dalam proses suksesi,
yaitu:
1. Nudation : terbukanya vegetasi penutup tanah
(terbentuknya tanah kosong).
2. Migrasi : cara-cara dimana tumbuhan sampai
pada daerah tersebut di atas.
Biji-biji tumbuhan sampai pada
daerah tersebut di atas mungkin
terbawa angin, aliran air,
mungkin pula melalui tubuh hewan
tertentu.
3. Ecesis : proses perkecambahan,
pertumbuhan, berkembang biak dan
menetapnya tumbuhan baru
tersebut. Sebagai hasil ecesis
individu-individu species tumbuh

mapan di suatu tempat
(established).
4. Agregation : sebagai hasil dari ecesis,
individu-individu dari suatu
jenis berkembang dan menghasilkan
biji, maka biji-biji tersebut
akan tersebar pada areal yang te
rbuka di sekelilingnya sehingga
tuinbuh berkelompok
(beragregasi).Ecesis dan agregasi
merupakan invasi species
tersebut.
5. Evolution of community relationship : merupakan
suatu proses apabila daerah yang
kosong ditempati species-species
yang berkoloni. Species tersebut
akan berhubungan satu sama lain-
nya. Bentuk hubungan ini kemung-
kinan akan mengikuti salah satu
dari tipe eksploitasi, mutualisme
dan co-existance.
6. Invation : dalam proses koloni, biji
tumbuhan telah beradaptasi dalam
waktu yang relatif panjang pada
tempat tersebut. Biji tumbuh dan
menetap (penguasaan lahan oleh
tumbuh-tumbuhan yang bersifat

agresif dan adaptif).
7. Reaction : terjadinya perubahan habitat
yang disebabkan oleh tumbuhan
tersebut dengan merubah
lingkungannya terutama dengan
cara:
a. Merubah sifat dan reaksi tanah
b. Merubah iklim mikro
Reaksi merupakan proses yang
terus menerus dan menyebabkan
kondisi yang cocok bagi species
yang telah ada dan lebih cocok
pada individu yang baru. Dengan
demikian reaksi memegang peranan
penting dalam pergantian species.
8.Stabilization: kompetisi dan reaksi berlangsung
terus menerus ditandai dengan
perubahan lingkungan yang
mengakibatkan struktur vegetasi
berubah. Dalam jangka waktu lama
akan terbentuk individu yang
dominan dan perubahan yang
terjadipun relatif kecil
disamping iklim mempunyai peranan
penting dalam membatasi proses
ini menjadi stabil. Dengan
perkataan lain, stabilisasi

merupakan suatu proses dimana
individu-individu tumbuhan mantap
tumbuh di suatu habitat tanpa
banyak dipengaruhi oleh
perobahan-perobahan dalam habitat
tersebat.
9. Klimaks :setelah stabilisasi, pada tahap
ini species yang dominan
mempunyai keseimbangan dengan
1ingkungannya, keadaan habitat
dan struktur vegetasi relatif
koristan karena pertumbuhan jenis
dominan telah mencapai batas.
E. Macam Suksesi
Berdasarkan proses terjadinya terdapat dua macam
suksesi;
1. Sukesesi primer (prisere)
Suksesi primer adalah perkembangan vegetasi mulai
dari habitat tak bervegetasi
hingga mencapai masyarakat yang
stabil dan klimaks.
2. Suksesi sekunder (subsere)
Suksesi sekunder terjadi apabila klimaks atau
suksesi yang normal terganggu
atau dirusak, misalnya oleh
kebakaran, perladangan,

penebangan, penggembalaan, dan
kerusakan-kerusakan lainnya.
F. Faham-fahara tentang Klimaks
1. Faham Monoklimaks (Costing, 1956)
Beranggapan bahwa pada suatu daerah iklim hanya
ada satu macam klimaks yaitu suatu formasi yang
paling metaphysic. Jadi klimaks boleh dikatakan
suatu pencerminan keadaan iklim. Disamping itu
iklim sebagai faktor yang paling stabil dan
berpengaruh, terdapat pula faktor-faktor lain
atau profaktor-profaktor, seperti faktor tanah,
biotis dan fisiografi. Profaktor-profaktor ini
menyebab-kan terbentuknya proklimaks-proklimaks
sebagai berikut :
a. Subklimaks terjadi apabila perkembangan
vegetasi terhenti di bawah tingkat terakhir,
dibawah klimaks, sebagai akibat faktor-faktor
bukan iklim, misalnya karena keadaan geografi
seperti keadaan di Pulau Krakatau.
b. Proklimaks Posklimaks, apabila pembentukan
klimaks menyimpang dari tipe yang sewajarnya,
misalnya sebagai akibat dari keadaan
fisiografi. Keadaan yang lebih lembab dan
lebih baik menghasilkan posklimaks, sedangkan
keadaan yang lebih kering dan kurang baik
menghasilkan proklimaks.

c. Disklimaks, terjadi sebagai akibat beberapa
gangguan sekunder yang menyebabkan tak dapat
berkembang lagi ke arah klimaks karena keadaan
tempat tumbuh amat berubah menjadi buruk,
misalnya terhenti pada tingkat semak belukar
2. Faham Polyklimaks(Braun-Blanquet, 1932)
Beranggapan bahwa tidak hanya iklim yang dapat
menumbuhkan klimaks. Bagi penganut faham kedua ini
ada beberapa macam kilmaks: klimaks iklim, klimaks
edafis, klimaks fisiografis, klimaks kebakaran dan
sebagainya.
3. Teori Informasi
Merupakan faham terbaru yang dikembangkan oleh
margalef (1968) dan Odum (1969). Pada tahap
klimaks komunitas tersebut mempunyai informasi
maksimum dan entrophy maksimum. Enthrophy adalah
jumlah energy yang tidak terpakai dalam suatu
sistem ekologi
Menurut faham monoklimaks misalnya dapat dibuat bagan
suksesi primer sebagai berukut:
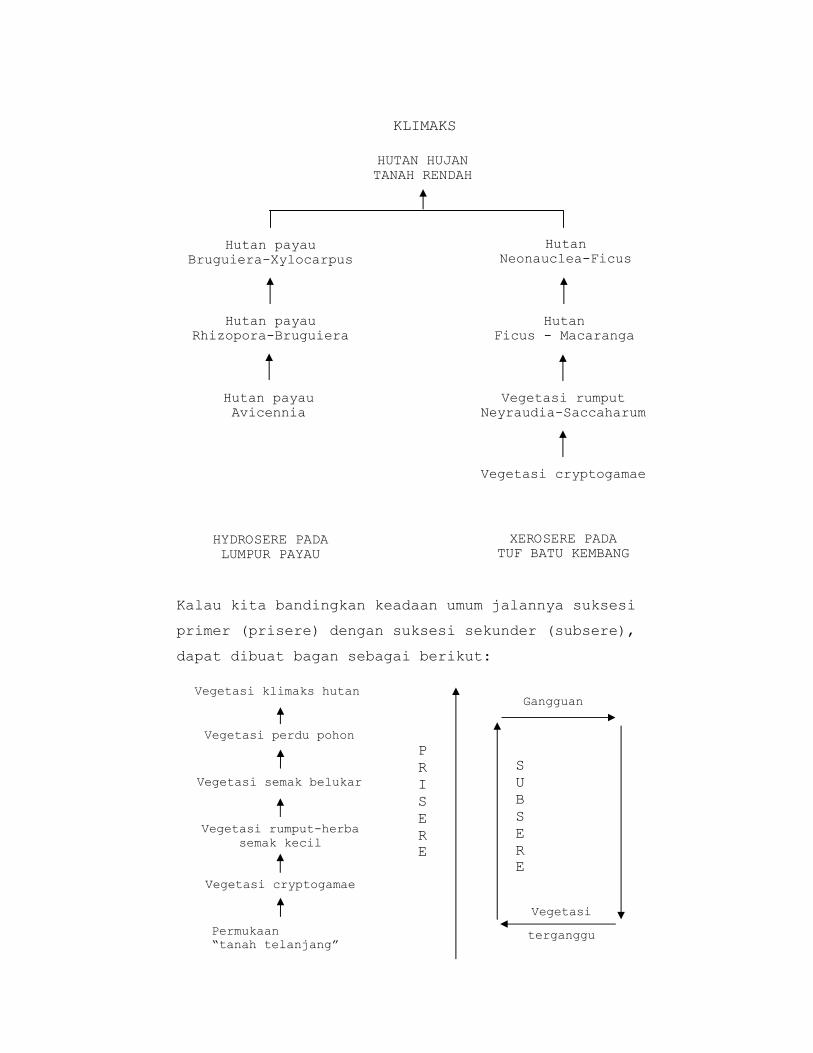
KLIMAKS
Kalau kita bandingkan keadaan umum jalannya suksesi
primer (prisere) dengan suksesi sekunder (subsere),
dapat dibuat bagan sebagai berikut:
HUTAN HUJAN TANAH RENDAH
Hutan payau Bruguiera-Xylocarpus
Hutan Neonauclea-Ficus
Hutan payau Rhizopora-Bruguiera
Hutan Ficus - Macaranga
Hutan payau Avicennia
Vegetasi rumput Neyraudia-Saccaharum
Vegetasi cryptogamae
HYDROSERE PADA LUMPUR PAYAU
XEROSERE PADA TUF BATU KEMBANG
Gangguan
Vegetasi
terganggu
Vegetasi klimaks hutan
Permukaan “tanah telanjang”
Vegetasi cryptogamae
Vegetasi rumput-herba semak kecil
Vegetasi semak belukar
Vegetasi perdu pohonPRISERE
SUBSERE

V. KLASIFIKASI VEGETASI HUTAN
A. Beberapa Pengertian yang Harus Dipahami dalam
Klasifikasi
1. Vegetasi adalah Masyarakat tumbuh-tumbuhan dalam
arti luas.
2. Formasi hutan adalah satuan vegetasi hutan yang
terbesar.
Perbedaan formasi hutan di trcpika disebabkan
oleh:
- Perbedaan iklim.
- Fisiognom.i (struktur) hutan
- Perbedaan habitat
- Suksesinya.
3. Asosiasi adalah satuan-satuan di dalam formasi
hutan yang diberi nama menurut pohon jenis
dominan. Oleh karena itu, Asosiasi adalah satuan
dasar dalam klasifikasi. Asosies adalah istilah
lain untuk asosiasi, dimana satuan ini berada
dalam hutan yang mengalami suksesi sekunder.
4. Asosiasi konkrit adalah bagian dari asosiasi hutan
yang betul-betul diselidiki dan diketahui
komposisi jenis pohonnya.
Asosiasi hutan yang berlainan komposisinya tetapi
memiliki fisiognomi yang sama, digolongkan menjadi
formasi hutan.
5. Subspecies, varietas, ekotype merupakan variasi-
variasi dalam species dalam taksonomi tumbuhan.

6. Varian adalah variasi-variasi di dalam asosiasi
hutan.
7. Asosiasi segregat adalah varian-varian di dalam
hutan campuran yang disebabkan oleh adanya jenis-
jenis pohon yang lebih berkuasa (dominan) daripada
yang lain.
8. Konsosiasi adalah varian yang dikuasai oleh satu
jenis pohon saja. Sedangkan konsosies adalah
varian di dalam suatu hutan yang mengalami sub-
sere/suksesi sekunder.
9. Fasiasi adalah varian yang disebabkan oleh
perbedaan topografi.
10. Losiasi adalah varian yang disebabkan oleh
perbedaan edafis.
11. Ekoton adalah daerah peralihan yang sering
dijumpai apabila ada dua atau lebih type atau
asosiasi vegetasi yang letaknya berbatasan.
3. SISTEM-SISTEM KLASIFIKASI VEGETASI HUTAN TROPIKA
Ada dua cara pendekatan di dalam klasifikasi
vegetasi:
1. Menetapkan dahulu satuan yang besar, kemudian
mengadakan pemisahan berdasarkan sifat-sifat yang
berbeda. Contoh : klasifikasi Schimper(1898) dan
Burtt Davy (1938).
2. Dimulai dengan memisahkan satuan yang kecil,
kemudian menggolongkan ke dalam satuan yang lebih

besar. Contoh : klasifikasi oleh Beard (1944),
dan Richard et. al. (1933) .
Adanya bermacam-macam sistem klasifikasi
disebabkan :
karena perbedaan kriteria yang digunakan, antara lain:
"Sistem Klasifikasi Fisiognomis, Ekologis, Fisiognomis-
Ekologis, Floristis, Fisiognomis-Floristis, Geografis-
Ekologis.
Menurut Aichinger, pada klasifikasi vegetasi,
kriteria pertama yang digunakan adalah fisiognomi,
selanjutnya floristik, geografi tumbuhan, ekologi,
syngenesisi, dan pengaruh manusia.
Menurut Fosberg (1958), klasifikasi vegetasi yang
rasional harus didasarkan kepada kriteria :
(1) Fisiognomi (rupa vegetasi, bentuk umum vegetasi) .
(2) Struktur vegetasi (susunan komponen di dalam
ruang, stratifikasi, jarak, dimcnsi).
(3) Fungsi (sifat-sifat phenothypik yang menyatakan
adaptasi terhadap keadaan lingkungan).
(4) Komposisi susunan floristik
(5) Dinamika suksesi atau perubahan dengan perbedaan
lingkungan.
(6) Riwayat vegetasi.

C. Berbagai Macam Sistem Klasifikasi Vegetasi Hutan
C.I. Klasifikasi Ekosistem Menurut Van Steenis
Van Steenis (1957) dalam Soerianegara dan
Indrawan (1934), telah mengemukakan dan membahas tipe-
tipe vegetasi yang dijumpai di Kepulauan Indonesia
dan wilayah sekitarnya.
Cara penetapan dan pembagian formasi-formasi
hutan di dalam sistem ini, yang disebut sistem
alami, didasarkan atas perbedaan iklim basah dan
bermusim, perbedaan edafis, dan perbedaan altitudinal.
Forrnasi-formasi hutan yang ditentukan dalam
sistem ini adalah :
I. IKLIM BASAH
Kadang-kadang selalu tergenang
Air asin (laut), dipengaruhi pasang surut :
........................ 1. Mangrove
Air tawar (hujan, sungai) , diam :
Eutrofik ............... 2. Hutan rawa
oligotropik ............ 3. Hutan gambut
Air tawar (tepi sungai), deras:
........................ 4.Vegetasi Rheofit
Tanah Kering
Pantai
........................ 5. Vegetasi pantai

Pedalarnan hingga batas pohon (timber line)
Tanah podsol kuarsa, dataran rendah :
.................. 6 . Vegetasi tanah
kuarsa
Tanah kapur, dataran rendah :
.................. 7 . Vegetasi tanah
kapur
Jenis- jenis tanah lain
Elevasi 2 - 1000 m .... 8. Hutan Hujan Tropika
Elevasi 1000-2400 m ... 9. Hutan Hujan Pegunungan
Elevasi 2400-4150 m .. 10. Hutan Hujan Sub-
alpin
II. IKLIM BERMUSIM
Elevasi di bawah 1000 m
................. 11 . Hutan Musim
(monsoon) Dataran
Rendah
Elevasi di atas 1000 m
................. 12 . Hutan Musim
Pegunungan .
C.2. Klasifikasi Vegetasi Dunia Menurut Unesco
Unesco (1973), telah melakukan klasifikasi dan
pembuatan peta vegetasi secara menyeluruh. Kategori
klasifikasi adalah unit-unit vegetasi, termasuk formasi zonal
dan azonal serta formasi-formasi yang telah berubah lainnya.
Dasar umum klasifikasi vegetasi dunia ini memakai sistem

floristik, klasifikasi selanjutnya didasarkan terutama pada
sifat-sifat fisiognomi struktural dan sifat-sifat ekologi yang
digabungkan dengan vegetasi natural dan semi natural sebagai
tambahan.
Menurut klasifikasi ini, vegetasi dunia dibedakan
menjadi enam tingkatan, dari tingkatan tertinggi sampai kelas
terendah, yaitu : Kelas Formasi (Formation Class), Sub-kelas
Formasi (Formation Subclass), Kelompok Formasi (Group
Formation), Formasi (Formation), dan Subdivisi (Subdivisions).
Kelas Formasi sebagai tingkatan tertinggi, membagi
vegetasi menjadi lima bagian. Pembagian ini berdasarkan kepada
struktur tegakan, dalam hal ini penutupan kanopi tegakan
(tajuk-tajuk pohon), tingkatan vegetasi (pohon atau semak
belukar); dan habitus veqetasi (berkayu atau herba).
Kelas formasi pertama adalah Closed forest (hutan
tertutup) adalah hutan-hutan yang mempunyai kanopi
tertutup, dimana tajuk-tajuk pohon saling mengisi. Tinggi
pohon paling rendah 5 m, kecuali untuk pohon, yang belum
dewasa atau masa reproduksi kurang dari 5 m.
Kelas Formasi Woodland (tegakan terbuka) terdiri dari
pohon-pohon dengan ketinggian paling rendah 5 m, penutupan
tajuk paling rendah 40%. Penutupan tajuk dikatakan 40% jika
jarak antara dua tajuk pohon sama dengan jari-jari sebuah
tajuk pohon.
Kelas Formasi Scrub (semak belukar) kebanyakan dari
jenis-jenis phanerophytes berkayu, tinggi antara 0.5 m sampai
5 m. Dibedakan atas semak individu-individunya tidak saling

bertautan, misalnya rumput-rumputan, sedangkan belukar saling
bertautan.
Kelas Formasi dwart-scrub dan Related Communities
(semak-semak kecil dan komunitas kerabat lainnya), sering
disebut formasi rumput-rumputan, tinggi jarang yang melebihi
50 cm. Berdasarkan kepadatannya dibedakan atas Dwart Shrub
thicket (cabang-cabangnya saling bertautan), Dwart Shrubland
(individu-individu saling terpisah atau dalam rumpun-rumpun),
dan Formasi Cryptogamic dengan semak-semak kecil.
Kelas Formasi terakhir adalah Herbaceous vegetation
(vegetasi herba). Ada dua tipe besar dari vegetasi ini, yaitu
graminoid dan forbs. Termasuk graininoid adalah semua rumput
herba dan tanaman rumput-rumputan seperti Carex sejenis alang-
alang), Juncus (sejenis tebu) dan sebagainya. Forbs adalah
tanaman herba daun lebar seperti Helianthus (bunga matahari),
Trifolium dan sebagainya.
Dasar pembagian kelas formasi menjadi subkelas formasi
adalah keadaan daun (evergreen, decidous, dan xeromorphic),
ukuran vegetasi, dan tempat hidup (habitat). Pengertian
evergreen adalah kanopi hutan tidak pernah tanpa daun hijau
(selalu hijau), walaupun ada pohon-pohon secara individu
mungkin menggugurkan daun. Kebalikan dari evergreen, pohon-
pohon decidous menggugurkan daun secara simultan apabila
berhubungan dengan musim yang tidak menguntungkan. sedangkan
xeromorphic adalah vegetasi yang khas daerah kering, seperti
phanerophyties, hemicytophties, geophyties. dengan daun atau
batang kadang-kadang sukulen.

Selanjutnya, Subkelas Formasi dibagi menjadi kelompok-
kelompok formasi (Group Formation) berdasarkan antara lain :
tempat atau garis 1intang (tropik, sub-tropik, temperate,
subpolar, dan lain-lain), keadaan daun (evergreen, decidous,
semi decidous), bentuk daun (daun jarum atau lebar), dan
kombinasi sifat-sifat di atas. Sedangkan formasi-formasi hutan
dibentuk berdasarkan antara lain : ketinggian tempat (lowland
dan montane), jenis tanah (alluvia), keadaan habitat (swamp,
bog, desert), bentuk tajuk, bentuk daun, dan sebagainya.
Di bawah ini diberikan bagan klasifikasi vegetasi
menurut Unesco (1973) secara global.
I. CLOSED FOREST
A. EVERGREEN
1. Tropical Ombrophilous Forest (Tropical Rain
Forest)
2. Tropical and Subtropical Evergreen Seasonal
Forest
3. Tropical and Subtropical Semi Decidous Purest
4. Subtropical Ombrophilous Forest
5. Mangrove Forest
6. Temperate and Subpolar Evergreen Ombropuilous
Forest
7. Temperate Evergreen Seasonal Broad heaved
Forest
8. Winter-Rain Evergreen Sclerophykous Forest
9. Tropical and Subtropical Evergreen Needle-Leaved
Forest

10.Temperate and Subpolar Evergreen Needle-Leaved
Forest
B. DECIDUOUS
1. Tropical and Subtropical Drought-Deciduous
Forest
2. Cold-Deciduous Forest with Evergreen Trees (or
Shrubs) Admixed
3. Cold-Deciduous Forest without Evergreen Trees
C. XEROMORPHIC
1. Sclerophyllous-Dominated Extremely Xero-morphic
Forest
2. Thorn-Forest
3 . Mainly Succulent Forest
II. WOODLAND
A. EVERGREEN
1. Evergreen Broad-Leaved Woodland
2. Evergreen Needle-Leaved Woodland
3. Cold-Deciduous Woodland without Evergreen
Trees
B. XEROMORPHIC
1. Sclerophyllous-Dominated Extremely Xeromorphic
Woodland
2. Thorn-Woodland

3. Mainly Succulent Woodland
III. SCRUB
A. EVERGREEN
1. Evergreen Broad-Leaved Shrubland
2. Evergreen Needle-Leaved and Microphylous
Shrubland
B. DECIDUOUS
1. Drought-Deciduous Scrub with Evergreen Woody
Plants Admixed
2. Drought-Deciduous Scrub eithout Evergreen
Woody Plant Admixed
3. Cold-Deciduous Scrub
C. XEROMORPHIC
1. Mainly Evergreen Subdesert Shrubland
2. Deciduous Subdesert Shrubland
IV. DWARF-SCRUB AND RELATED COMMUNITIES
A. EVERGREEN
1. Evergreen Dwarf-Shurb Thicket
2. Evergreen Dwarf-Shrubland
3. Mixed Evergreen Dwarf-Shrub and Herbaceous
Formation.

B. DECIDUOUS
1. Facultatively Drought-Deciduous Dwarf-Thicket
2. Obligatory, Drought-Deciduous Dwarf-Thicket
3. Cold-Deciduous Dwarf-Thicket
C. XEROMORPHIC
1. Mainly Evergreen Subdesert Dwart-scrub
2. Deciduous Subdesert Dwarf-Scrub
D. TUNDRA
1. Mainly Bryophyte Tundra
2. Mainly Lichen Tundra
E. MOSSY BOG FORMATIONS WITH DWARF-SHRUB
1. Raised Bog
2. Non-Raised Bog
V. HERBACEOUS VEGETATION
A. TAIL GRAMINOID VEGETATION
B. MEDIUM TALL GRASSLAND
C. SHORT GRASSLAND
D. FORB VEGETATION
E. HYDROMORPHIC FRESH-WATER GRASSLAND
C.3. Klasifikasi Ekosistem Menurut Kartawinata
Kartawinata telah membuat bagan unit-unit
ekosistem atau tipe-tipe ekosistem darat dan rawa yang

ada di Indonesia. Tipe ekosistem dianggap unit-unit
yang paling kecil dan dibentuk berdasarkan fisiognomi
(kenampakan) struktur dan takson (unit taksonomi) yang
khas atau dominan dari vegetasi yang dikombinasikan
dengan faktor-faktor iklim dan ketinggian dari
permukaan laut serta tanah. Faktor-faktor tidak
dimasukkan karena datanya kurang, lagipula perincian
ekositem dengan ciri-ciri vegetasi dan lingkungan dapat
dianggap cukup. Berdasarkan komposisi jenis masing-
masing tipe ekosistem dapat saja terdiri dari unit-unit
yang lebih kecil. Ekosistem hutan kerangas, misalnya,
mungkin tersusun dari unit komunitas Combretocarpus-
Dactylocladus dan Tristania-Cratoxylum.
Menurut Klasifikasi Kartawinata (1976) ini, ada
tiga tingkatan klasifikasi, yaitu : Bioma, Subbioma,
dan Tipe Ekosistem. Bioma dapat pula disebut sebuah
ekosistem yang merupakan unit komunitas terbesar yang
mudah dikenal dan terdiri atas forrnasi vegetasi dan
hewan serta mahluk hidup lainnya, baik yang sudah
mencapai fase klimaks maupun yang masin dalam fase
perkembangan. Di Indonesia dapat dikenal beberapa
bioma, yaitu : (a) Hutan Hujan, (b) Hutan Musim, (c)
Savana, (d) Padang Rumput. Unit-unit ekosistem ini
masih terlalu besar untuk digunakan dengan maksud-
maksud khusus, sehingga memerlukan pembagian yang lebih
kecil lagi.
Pembagian Bioma menjadi Subbioma didasarkan kepada

keadaan iklim, misalnya, untuk Hutan Hujan dibedakan
antara Hutan Hujan Tanah Kering dan Hutan Hujan Tanah
Rawa (permanen atau musiman). Sedangkan pembagian
Tipe-tipe Ekosistem sebagai unit yang paling kecil
dibentuk berdasarkan struktur fisiognomi, faktor-faktor
iklim, ketinggian dari permukaan laut, dan jenis tanah.
Klasifikasi Ekosistem menurut Kartawinata tertera dalam
Tabel 1, berikut.
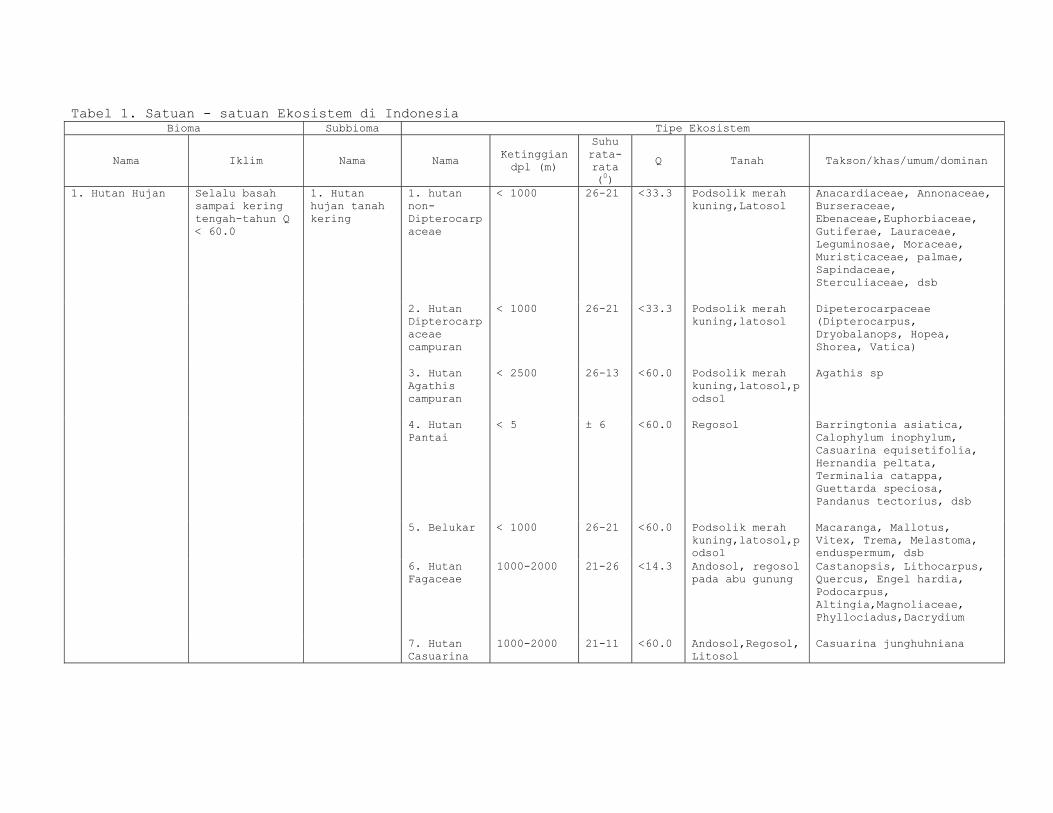
Tabel 1. Satuan - satuan Ekosistem di Indonesia Bioma Subbioma Tipe Ekosistem
Nama Iklim Nama Nama Ketinggian dpl (m)
Suhu rata-rata (0)
Q Tanah Takson/khas/umum/dominan
1. Hutan Hujan Selalu basah sampai kering tengah-tahun Q < 60.0
1. Hutan hujan tanah kering
1. hutan non-Dipterocarpaceae
< 1000 26-21 <33.3 Podsolik merah kuning,Latosol
Anacardiaceae, Annonaceae, Burseraceae, Ebenaceae,Euphorbiaceae, Gutiferae, Lauraceae, Leguminosae, Moraceae, Muristicaceae, palmae, Sapindaceae, Sterculiaceae, dsb
2. Hutan Dipterocarpaceae campuran
< 1000 26-21 <33.3 Podsolik merah kuning,latosol
Dipeterocarpaceae (Dipterocarpus, Dryobalanops, Hopea, Shorea, Vatica)
3. Hutan Agathis campuran
< 2500 26-13 <60.0 Podsolik merah kuning,latosol,podsol
Agathis sp
4. Hutan Pantai
< 5 ± 6 <60.0 Regosol Barringtonia asiatica, Calophylum inophylum, Casuarina equisetifolia, Hernandia peltata, Terminalia catappa, Guettarda speciosa, Pandanus tectorius, dsb
5. Belukar < 1000 26-21 <60.0 Podsolik merah kuning,latosol,podsol
Macaranga, Mallotus, Vitex, Trema, Melastoma, enduspermum, dsb
6. Hutan Fagaceae
1000-2000 21-26 <14.3 Andosol, regosol pada abu gunung
Castanopsis, Lithocarpus, Quercus, Engel hardia, Podocarpus, Altingia,Magnoliaceae, Phyllociadus,Dacrydium
7. Hutan Casuarina
1000-2000 21-11 <60.0 Andosol,Regosol, Litosol
Casuarina junghuhniana
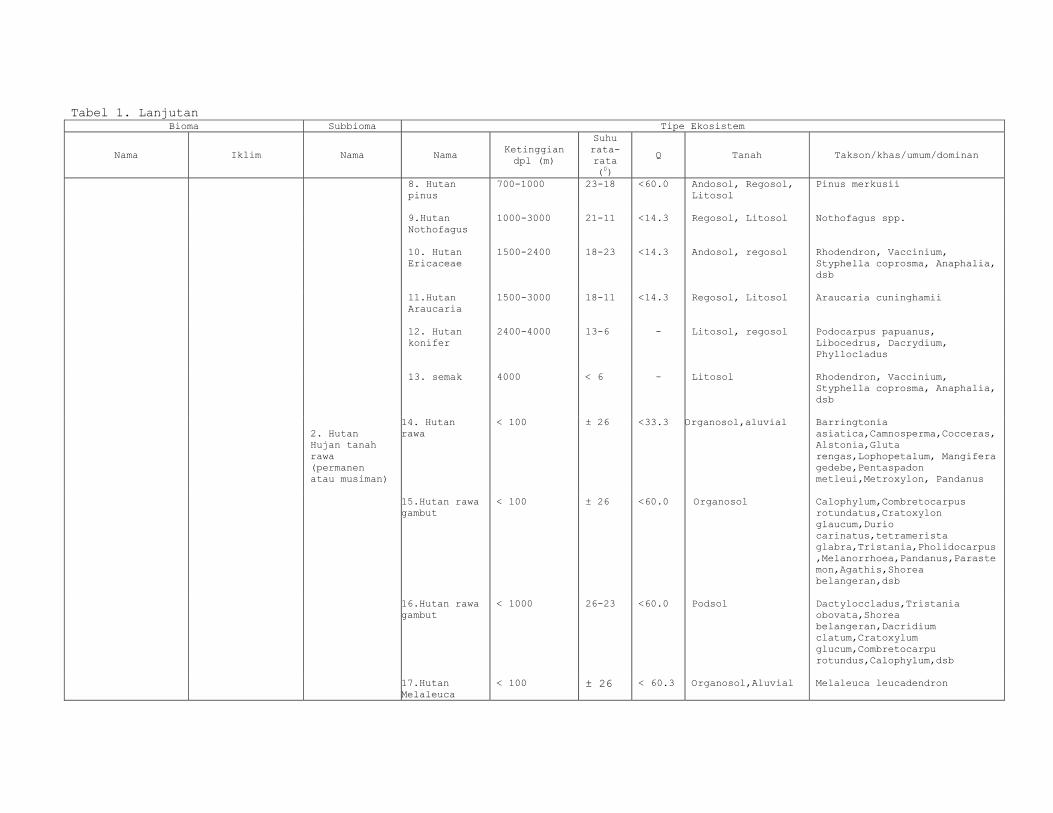
Tabel 1. Lanjutan Bioma Subbioma Tipe Ekosistem
Nama Iklim Nama Nama Ketinggian dpl (m)
Suhu rata-rata (0)
Q Tanah Takson/khas/umum/dominan
2. Hutan Hujan tanah rawa (permanen atau musiman)
8. Hutan pinus
700-1000 23-18 <60.0 Andosol, Regosol, Litosol
Pinus merkusii
9.Hutan Nothofagus
1000-3000 21-11 <14.3 Regosol, Litosol Nothofagus spp.
10. Hutan Ericaceae
1500-2400 18-23 <14.3 Andosol, regosol
Rhodendron, Vaccinium, Styphella coprosma, Anaphalia, dsb
11.Hutan Araucaria
1500-3000 18-11 <14.3 Regosol, Litosol Araucaria cuninghamii
12. Hutan konifer
2400-4000 13-6 - Litosol, regosol Podocarpus papuanus, Libocedrus, Dacrydium, Phyllocladus
13. semak 4000 < 6 - Litosol Rhodendron, Vaccinium, Styphella coprosma, Anaphalia, dsb
14. Hutan rawa
< 100 ± 26 <33.3 Organosol,aluvial Barringtonia asiatica,Camnosperma,Cocceras,Alstonia,Gluta rengas,Lophopetalum, Mangifera gedebe,Pentaspadon metleui,Metroxylon, Pandanus
15.Hutan rawa gambut
< 100 ± 26 <60.0 Organosol Calophylum,Combretocarpus rotundatus,Cratoxylon glaucum,Durio carinatus,tetramerista glabra,Tristania,Pholidocarpus,Melanorrhoea,Pandanus,Parastemon,Agathis,Shorea belangeran,dsb
16.Hutan rawa gambut
< 1000 26-23 <60.0 Podsol Dactyloccladus,Tristania obovata,Shorea belangeran,Dacridium clatum,Cratoxylum glucum,Combretocarpu rotundus,Calophylum,dsb
17.Hutan Melaleuca
< 100 ± 26 < 60.3 Organosol,Aluvial Melaleuca leucadendron
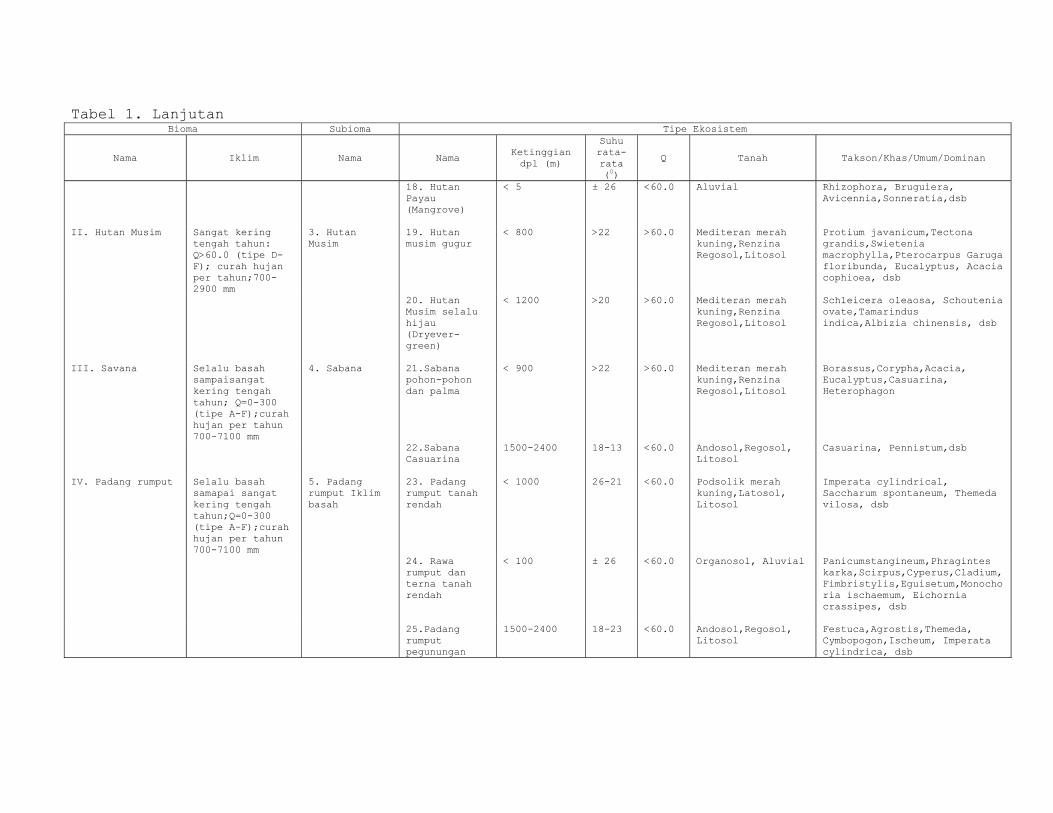
Tabel 1. Lanjutan Bioma Subioma Tipe Ekosistem
Nama Iklim Nama Nama Ketinggian dpl (m)
Suhu rata-rata (0)
Q Tanah Takson/Khas/Umum/Dominan
18. Hutan Payau (Mangrove)
< 5 ± 26 <60.0 Aluvial Rhizophora, Bruguiera, Avicennia,Sonneratia,dsb
II. Hutan Musim Sangat kering tengah tahun: Q>60.0 (tipe D-F); curah hujan per tahun;700-2900 mm
3. Hutan Musim
19. Hutan musim gugur
< 800 >22 >60.0 Mediteran merah kuning,Renzina Regosol,Litosol
Protium javanicum,Tectona grandis,Swietenia macrophylla,Pterocarpus Garuga floribunda, Eucalyptus, Acacia cophioea, dsb
20. Hutan Musim selalu hijau (Dryever-green)
< 1200 >20 >60.0 Mediteran merah kuning,Renzina Regosol,Litosol
Schleicera oleaosa, Schoutenia ovate,Tamarindus indica,Albizia chinensis, dsb
III. Savana Selalu basah sampaisangat kering tengah tahun; Q=0-300 (tipe A-F);curah hujan per tahun 700-7100 mm
4. Sabana 21.Sabana pohon-pohon dan palma
< 900 >22 >60.0 Mediteran merah kuning,Renzina Regosol,Litosol
Borassus,Corypha,Acacia, Eucalyptus,Casuarina, Heterophagon
22.Sabana Casuarina
1500-2400 18-13 <60.0 Andosol,Regosol, Litosol
Casuarina, Pennistum,dsb
IV. Padang rumput Selalu basah samapai sangat kering tengah tahun;Q=0-300 (tipe A-F);curah hujan per tahun 700-7100 mm
5. Padang rumput Iklim basah
23. Padang rumput tanah rendah
< 1000 26-21 <60.0 Podsolik merah kuning,Latosol, Litosol
Imperata cylindrical, Saccharum spontaneum, Themeda vilosa, dsb
24. Rawa rumput dan terna tanah rendah
< 100 ± 26 <60.0 Organosol, Aluvial Panicumstangineum,Phragintes karka,Scirpus,Cyperus,Cladium,Fimbristylis,Eguisetum,Monochoria ischaemum, Eichornia crassipes, dsb
25.Padang rumput pegunungan
1500-2400 18-23 <60.0 Andosol,Regosol, Litosol
Festuca,Agrostis,Themeda, Cymbopogon,Ischeum, Imperata cylindrica, dsb
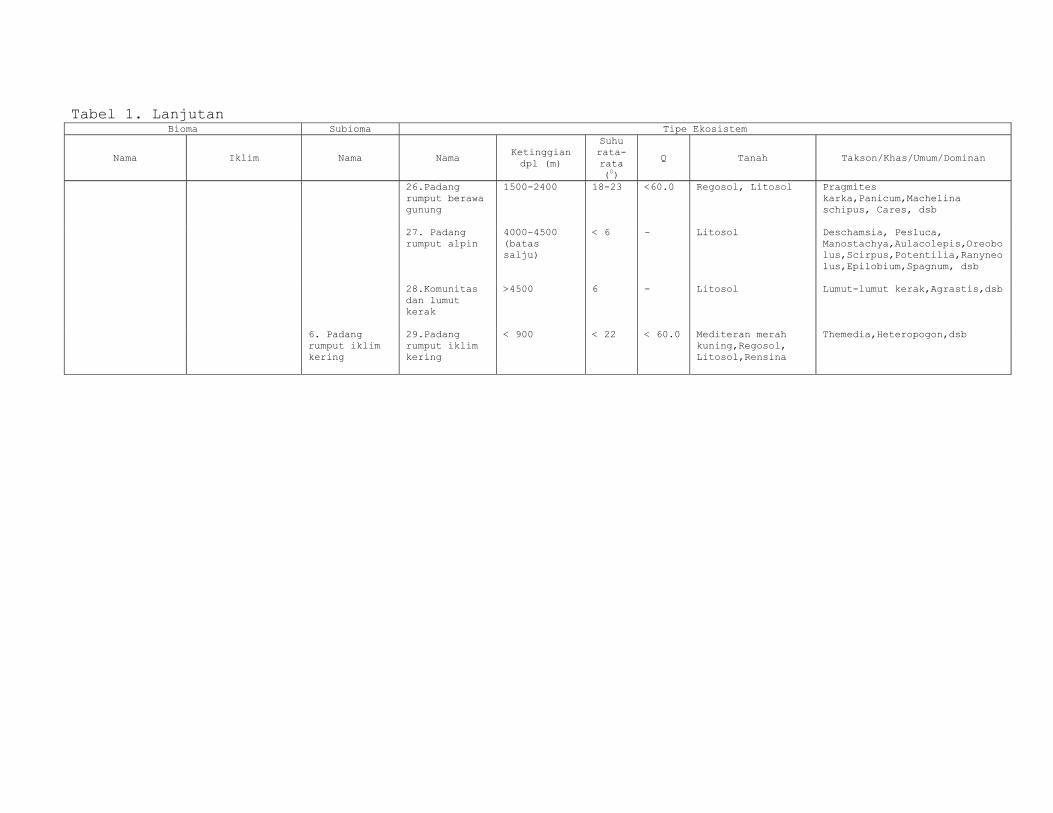
Tabel 1. Lanjutan Bioma Subioma Tipe Ekosistem
Nama Iklim Nama Nama Ketinggian dpl (m)
Suhu rata-rata (0)
Q Tanah Takson/Khas/Umum/Dominan
26.Padang rumput berawa gunung
1500-2400 18-23 <60.0 Regosol, Litosol Pragmites karka,Panicum,Machelina schipus, Cares, dsb
27. Padang rumput alpin
4000-4500 (batas salju)
< 6 - Litosol Deschamsia, Pesluca, Manostachya,Aulacolepis,Oreobolus,Scirpus,Potentilia,Ranyneolus,Epilobium,Spagnum, dsb
28.Komunitas dan lumut kerak
>4500 6 - Litosol Lumut-lumut kerak,Agrastis,dsb
6. Padang rumput iklim kering
29.Padang rumput iklim kering
< 900 < 22 < 60.0 Mediteran merah kuning,Regosol, Litosol,Rensina
Themedia,Heteropogon,dsb

C.4.Klasifikasi Tipe-tipe Hutan di Indonesia oleh
Departemen Kehutanan
Departemen Kehutanan dalam Vademecum (1976) telah
mengklasifikasikan hutan di Indonesia berdasarkan
keadaan iklim, edafis, dan komposisi tegakan. Faktor
iklim menurut pembagian F.H. Schimidt dan J.H. Ferguson
yang didasarkan pada nilai Q, yaitu persentase perban-
dingan antara jumlah bulan kering dan jumlah bulan
basah, sehingga diperoleh tipe-tipe iklim A, B, C, D
dan seterusnya berturut-turut dari nilai Q yang
terkecil sampai terbesar. Faktor iklim yang
mempengaruhi pernbentukan vegetasi adalah temperatur,
kelembaban, intensitas cahaya dan kecepatan angin.
Tipe hutan yang pembentukannya sangat dipengaruhi
oleh faktor iklim disebut Formasi Klimatis (Klimatic
Formation). Termasuk kedalamnya, yaitu : Hutan Hujan
(Tropical Rain Forest), Hutan Musim (Monsoon Forest),
dan Hutan Gambut (Peat Forest).
Hutan Nipa [Nipa Formation) dianggap sebagai
suatu konsosiasi dari Hutan Payau atau Hutan Rawa
tergantung kepada faktor edafis yang ada.
Hutan Palma tanah rawa (Palm swamp forest.)
dimana banyak terdapat jenis-jenis Phoenix atau
Oncosperma dianggap sebagai suatu konsosiasi.
Berikut diberikan bagan klasifikasi tipe-tipe
hutan di Indonesia menurut Departemen Kehutanan :
I. FORMASI KLIMATIS
1. Hutan Hujan (Tropical Rain Forest)
Ciri-ciri : iklim A atau B; jenis tanah latosol,
aluvial, dan regosol; drainase baik,jauh dari
pantai; dan tegakan selalu hijau.
a. Hutan Hujan Bawah (0-1000 m dpl)
Jenis pohon yanq dominan : famini

Dipterocarpaceae (Kalimantan dan Sumatera);
Agathis, Ficus, Castanopsis (Jawa dan Husa
Tenggara);Palaquium spp., Pometia pinnata,
Diospyros spp. (Indonesia Timur)
b. Hutan Hujan Tengah (1000-3300 m dpl)
Quercus, Castanopsis, Nothofagus, dan jenis-
jenis dari famili Magnoliaceae; Pinus merkusii
(Aceh); Albizia montana, Casuarina (Jawa);
Trema, Podocarpus imbricatus (Indonesia Timur)
c. Hutan Hujan Atas (3300-4100 m dpl)
Merupakan kelompok-kelompok yang terpisah-
pisah oleh padang rumput atau belukar. Jenis-
jenis pohon: Dacrydium, Podocarpus,
Phyllocladus (Irian Jaya),Eugenia, dan
Calophyl1um.
2. Hutan Musim (Monsoon Forest)
Ciri-ciri : iklim C atau D; gugur daun musim
kemarau; terdapat 2 lapisan tajuk yang berbeda;
dan banyak herba dan tumbuhan bawah.
a. Hutan Musim Bawah (0-1000 m dpl)
Jenis-jenis pohon : Tectona grandis, Acacia
leucophloea, Albizia chinensis (Jawa); Euca-
lyptus, Santalum album (Nusa Tenggara).
b. Hutan Musim Tengah-Atas (1000-4100 m dpl)
Casuarina junghuhniana (Jawa Tengah dan
Timur); Eucalyptus (Indonesia Timur), Pinus
merkusii (Sumatera).
3. Hutan Gambut (Peat Forest)
Ciri-ciri: iklim A atau B; tanah organosol;
terletak anatar hutan hujan dan hutan rawa;
selalu hijau dan banyak lapisan tajuk.
Jenis-jenis : Alstonia spp., Palaquium spp.,
Dactylocladus; Eugenia spp., Gonystylus spp.

(khusus di Kalimantan dan beberapa daerah di
Sumatera).
II. FORMASI EDAFIS
1. Hutan Rawa (Swamp Forest)
Ciri-ciri: tidak terpengaruh iklim; selalu
tergenang air tawar; terletak di belakang hutan
payau; jenis tanah aluvial, selalu hijau; dan
banyak lapisan tajuk.
Jenis-jenis pohon: Xylopia spp., Palaquium
leiocarpu/n, Campnoserma macrophylla, Garcinia
spp., Canarium spp., Koompassia spp., dan
Calophyllum spp..
2. Hutan Payau (Mangrove Forest)
Ciri-ciri : daerah pantai dan selalu tergenang
air laut; terpengaruh pasang surut; tidak
terpengaruh iklim; tanah pasir, lumpur, dan
lumpur berpasir; hanya satu stratum tajuk. Jenis-
jenis Avicennia spp., Sonneratia spp., Rhizophora
spp., Bruguiera spp., Xylocarpus spp., Lumnitzera
3. Hutan Pantai (Littoral Forest)
Ciri-ciri : di daerah kering pantai; tidak ter-
pengaruh iklim; tanah pasir dan berbatu; terletak
pada garis pasang tertinggi; dan banyak epifit.
Jenis-jenis : Baringtonia speciosa, Terminalia
catappa, Calophyllum inophyllum, Hibiscus tilia-
ceua, Casuarina equisetifolia, Pisonia grandis.
Disamping ini banyak terdapat Pandanus tectorius.
Banyak terdapat epifit terutama paku-pakuan dan
anggrek. Jenis-jenis pioner pada pantai berpasir
diantaranya adalah Ipomea pescaprae dan
Coccoloba.

C.5. Klasifikasi Ekosistem Makro di Sumatera Menurut
Djamhuri
Djamhuri et al. (1988) mengklasifikasikan
ekosistem makro di Sumatera bersumber pada empat jenis
data hasdl penelitian oleh pihak lain, yaitu :
1. Peta Vegetasi di Sumatera berskala 1 : 1.000.000
oleh Laumunier, Purnadjaja, dan Setiabudi (198S).
2. Peta Tanah Eksploitasi Pulau Sumatera skala 1:
2.500.000 oleh Lembaga Penelitian Tanah Bogor (1979)
3. Peta Geologi Pulau Sumatera skala 1 : 1.000.000 oleh
Direktorat Geologi Bandung (1965).
4. Peta Agroklimat Pulau Sumatera skala 1 : 1.000.000
oleh Oldeman (1973).
Dengan demikian Djamhuri et al (1988)
mengklasifikasikan ekosistem makro ini berdasarkan
beberapa parameter, yaitu : keadaan habitat, arah
fisiografi, ketinggian tanah, geologi (batu-batuan),
iklim dan keadaan vegetasi. Hasil klasifikasi ekosistem
makro ini disajikan dalam tiga buah peta tipe ekosistem
makro di Sumatera berskala 1 : 1.000.000, yaitu Peta Tipe
Ekosistem Makro Sumatera Bagian Utara, Peta Tipe
Ekosistem Makro Sumatera Tengah, dan Peta Tipe Ekosistem
Makro Sumatera Bagian Selatan.
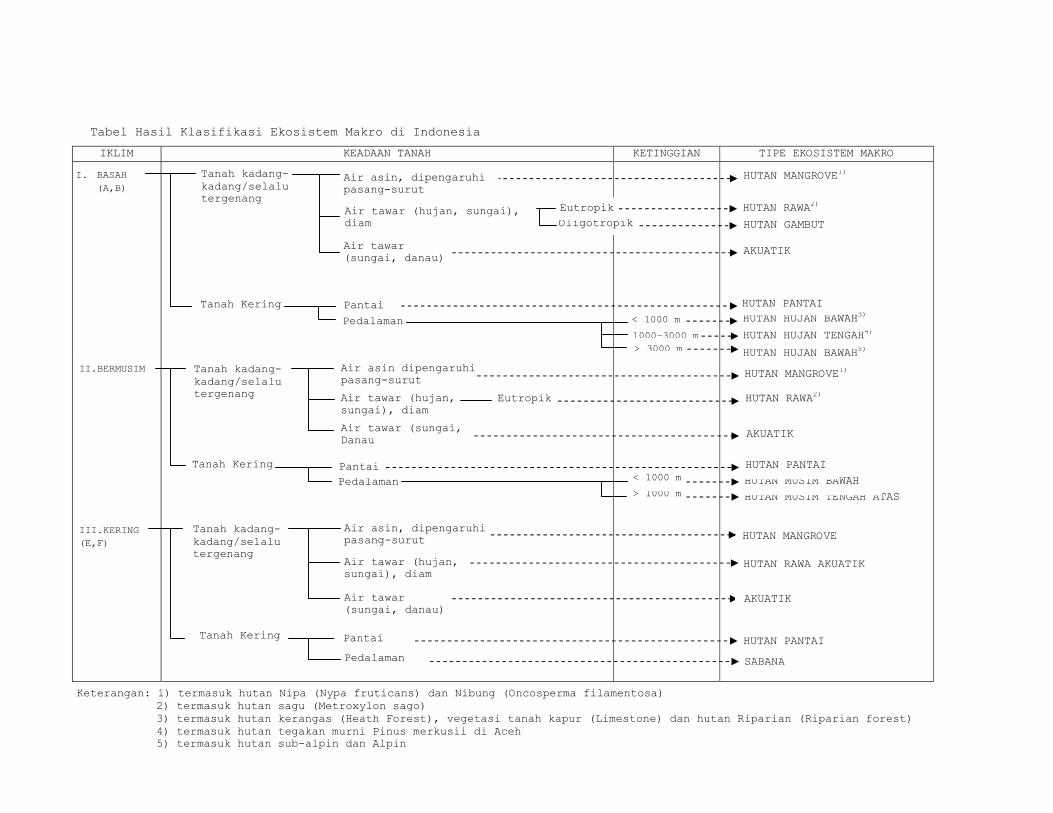
Tabel Hasil Klasifikasi Ekosistem Makro di Indonesia
IKLIM KEADAAN TANAH KETINGGIAN TIPE EKOSISTEM MAKRO
I. BASAH (A,B)
II.BERMUSIM
III.KERING (E,F)
Keterangan: 1) termasuk hutan Nipa (Nypa fruticans) dan Nibung (Oncosperma filamentosa) 2) termasuk hutan sagu (Metroxylon sago) 3) termasuk hutan kerangas (Heath Forest), vegetasi tanah kapur (Limestone) dan hutan Riparian (Riparian forest) 4) termasuk hutan tegakan murni Pinus merkusii di Aceh 5) termasuk hutan sub-alpin dan Alpin
Pantai
HUTAN HUJAN BAWAH5)HUTAN HUJAN TENGAH4)HUTAN HUJAN BAWAH3)
> 3000 m1000–3000 m
< 1000 m
HUTAN PANTAI
AKUATIK
HUTAN GAMBUT
HUTAN RAWA2)
HUTAN MANGROVE1)
Oligotropik Eutropik
Tanah Kering
Tanah kadang-kadang/selalu tergenang
Air tawar (sungai, danau)
Air tawar (hujan, sungai), diam
Air asin, dipengaruhi pasang-surut
PedalamanPantai
HUTAN MUSIM TENGAH ATAS HUTAN MUSIM BAWAH HUTAN PANTAI
> 1000 m
< 1000 m
AKUATIK
HUTAN RAWA2)
HUTAN MANGROVE1)
Eutropik
Air tawar (sungai, Danau
Air tawar (hujan, sungai), diam
Air asin dipengaruhi pasang-surut
Tanah kadang-kadang/selalu tergenang
Tanah Kering
Pedalaman Pantai
SABANA
HUTAN PANTAI
HUTAN RAWA AKUATIK
HUTAN MANGROVE
Pedalaman
Air tawar (sungai, danau)
Air tawar (hujan, sungai), diam
Air asin, dipengaruhi pasang-surut
Tanah kadang-kadang/selalu tergenang
Tanah Kering Pantai
AKUATIK

VI. TEKNIK ANALISIS VEGETASI RUANG LINGKUP
Dalam analisis vegetasi ada beberapa hal yang
harus diperhatikan oleh seorang surveyor agar survey
vegetasi yang dilakukan dapat memberikan data/informasi
yang teliti dan dapat: dipertanggung jawabkan. Hal-hal
tersebut adalah ukuran, jumlah dan bentuk petak contoh
yang akan dipilih, cara meletakkan petak contoh, obyek
yang akan diamati, parameter vegetasi yang akan diukur,
dan akhirnya teknik analisis vegetasi yang akan
digunakan.
PETAK CONTOH VEGETASI
Untuk mempelajari komposisi jenis dan struktur
komunitas tumbuhan umumnya dilakukan dengan sampling.
Dalam hal ini ada tiga hal yang harus diperhatikan,
yaitu ukuran bentuk dan jumlah petak contoh, cara
meletakkan petak, dan teknik analisa vegetasi yang
harus digunakan.
A. Ukuran, Jumlah dan Bentuk Petak
Ukuran petak bergantung pada ukuran tumbuhan
(semai, pancang, tiang, pohon), kerapatan tumbuhan dan
keragaman jenis serta keheterogenan life-formnya. Dalam
penentuan ukuran petak prinsipnya adalah bahwa petak
harus cukup besar agar individu species yang ada dalam
contoh dapat mewakili komunitas, tetapi harus cukup
kecil agar individu yang ada dapat dipisahkan, dihitung
dan diukur tanpa duplikasi atau pengabaian.
Salah satu cara/metoda untuk menentukan
ukuran/jumlah petak contoh adalah menggunakan kurva
species area. Cara membuat kurva ini adalah sebagai
berikut:

(1) Buat sebuah petak contoh (pc) berukuran 1 X 1 m
atau sebuah lingkaran beradius 0.56 m.
(2) Catat jumlah jenis dalam pc tersebut.
(3) Buat pc kedua yang besarnya dua kali lipat pc
pertama. Catat jumlah jenis pada pc kedua.
(4) Buat pc ketiga dan seterusnya yang ukurannya
masing-masing dua kali lipat pc sebelumnya.
Catat jumlah jenis masing-masing pc tersebut.
(5) Pembuatan pc dihentikan kalau penambahan jumlah
jenis sekitar 10%.
(6) Buat sumbu-X (luas petak contoh) dan sumbu-Y
(jumlah jenis).
(7) Buat suatu garis (misal garis m) yang melewati
titik 0 (0,0) dan titik A dengan koordinat (10%
luas petak contoh, 10% jumlah jenis).
(8) Buat suatu garis (misal garis n) yang sejajar m
yang menyinggung kurva species-area. Titik
persinggungan tersebut diproyeksikan pada
surnbu-X, sehingga didapat luas minimum petak
contoh.
Untuk menentukan jumlah petak contoh minimal,
prose durnya sama dengan di atas, tetapi sebagai sumbu-
X (absis) adalah jumlah petak contoh.
Bentuk petak contoh sangat penting dalam
memudahkan letak petak dan efisiensi sampling. Ada tiga
bentuk petak contoh yaitu lingkaran, bujur sangkar, dan
empat persegi panjang. Diantara bentuk-bentuk petak
tersebut, bentuk lingkaran mempunyai ketelitian yang
cukup tinggi dalam proses pernbuatannya. Petak bentuk
lingkaran akan praktis kalau digunakan untuk komunitas
rumput, herba dan semak-belukar. Sedangkan petak
berbentuk persegi panjang akan lebih efisien daripada

petak berbentuk bujur sangkar dalam jumlah dan luasan
yang sama, bila sumbu panjang petak sejajar perubahan
gradient lingkungan.
B. Cara Meletakkan Petak Contoh
Pada dasarnya ada dua cara peletakan petak
contoh, yaitu cara acak (random sampling) dan cara
sistematik (systematic sampling). Dari segi floristis-
ekologis, random sampling hanya mungkin digunakan
apabila lapangan dan vegetasinya homogen, misalnya
hutan tanaman dan padang rumput. Sedangkan untuk
keperluan survey vegetasi yang lebih teliti sistematik
sampling dianjurkan, karena mudah dalam pelaksanaannya
dan data yang dihasilkan akan dapat lebih bersifat
representative. Bahkan dalam keadaan tertentu yang
terkait dengan keterbatasan biaya, tenaga dan waktu,
purposive sampling pun dapat digunakan dalam analisis
vegetasi.
C. Kriteria Stadium Pertumbuhan
Secara ekologis cukup penting untuk membeda-
bedakan tumbuhan ke dalam stadium pertumbuhan semai,
pancang, tiang dan pohon, bahkan tumbuhan bawah. Untuk
keperluan ini kriteria yang dapat digunakan adalah
sebagai berikut :
a) Semai : Permudaan mulai dari kecambah sampai
anakan setinggi kurang dari 1,5 m.
b) Pancang : Permudaan dengan tinggi 1,5 m sampai
anakan berdiameter kurang dari 10 cm.
c) Tiang : Pohon muda berdiameter 10 cm sampai
kurang dari 20 cm.
d) Pohon : Pohon dewasa berdiameter 20 cm dan
lebih.

e) Tumbuhan bawah : Tumbuhan selain permudaan pohon,
misal rumput, herba, dan sernak belukar.
Khusus untuk mangrove stadium tiang biasanya
ditiadakan, sehingga stadium pohon meliputi pohon
berdiameter 10 cm ke atas. Selain itu, diameter pohon
diukur pada ketinggian 20 cm di atas akar tunjang
(Rhizophora. spp.) dan keting¬gian 20 cm di atas banir
untuk jenis non-Rhizophora spp. Bagi pohon-pohon tidak
berakar tunjang dan berbanir, pengukuran diameter pohon
dilakukan pada ketinggian 1,3 m di atas permukaan tanah
(DBH, diameter at breast-height).
D. Parameter Vegetasi yang Diukur di Lapangan
Dalam analisis vegetasi ada beberapa parameter
vegetasi yang diukur secara langsung di lapangan, yaitu
:
a) Nama species (lokal dan ilmiah).
b) Jumlah individu untuk menghitung kerapatan.
c) Penutupan tajuk (covering) untuk mengetahui
prosentase penutupan vegetasi terhadap lahan.
d) Diameter batarig untuk mengetahui luas bidang dasar
yang diantaranya sangat berguna untuk memprediksi
volume pohon dan tegakan.
e) Tinggi pohon baik tinggi pohon bebas cabang maupun
tinggi pohon total. Tinggi pohon ini cukup penting
untuk mengetahui stratifikasi dan menduga volume
pohon serta volume tegakan.
f) Pemetaaan lokasi individu pohon untuk mendeteksi
spatial distribution pattern pada berbagai luasan
areal yang berbeda.
Dalam prakteknya, hampir semua kegiatan survey
vegetasi mengadakan pengukuran terhadap jumlah individu

per jenis, diameter batang, dan tinggi pohon serta
tentu saja identifikasi jenis. Walaupun demikian
parameter vegetasi yang akan diobservasi tergantung
pada informasi yang diinginkan oleh surveyor/peneliti.
E. Ukuran Sub-plot untuk Berbagai Stadium Pertumbuhan
Untuk keperluan risalah tumbuhan bawah, permudaan
dan pohon di dalam petak contoh seyogyanya dilakukan di
dalam subplot-subplot contoh agar memudahkan dalam
risalahnya dan tidak terjadi duplikasi penghitungannya.
Teknik pembuatan sub-plot-sub-plot tersebut biasanya
dilakukan secara nested sampling, yaitu sub-plot yang
berukuran lebih besar mengandung sub-plot yang berukuran
lebih kecil. Dalam hal ini ukuran sub-plot untuk berbagai
stadium pertumbuhan adalah :
a. Semai dan tumbuhan bawah : 2 X 2 m atau 1 X 1 m atau 2 X 5 m.
b. Pancang : 5 X 5 m
c. Tiang : 10 X 10 m
d. Pohon : 20 x 20 m atau 20 X 50 m.
F. Metoda Analisis Vegetasi F.I. Metoda dengan petak F.I.I. Metode kuadrat
(1) . Petak tunggal
Di dalam metoda ini dibuat satu petak sampling
dengan ukuran tertentu yang mewakili suatu tegakan
hutan, Ukuran petak ini dapat ditentukan dengan kurva
species-area. Untuk lebih jelasnya suatu contoh petak
tunggal dapat dilihat gambar 1.
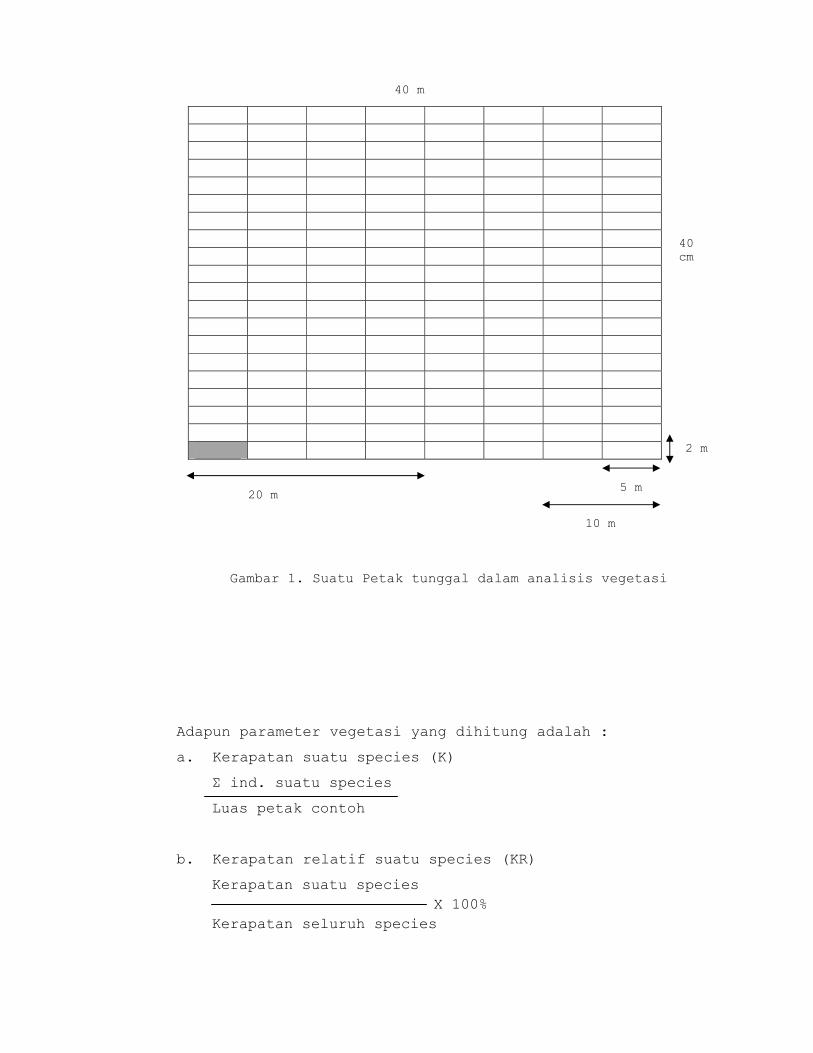
Adapun parameter vegetasi yang dihitung adalah :
a. Kerapatan suatu species (K)
Σ ind. suatu species
Luas petak contoh
b. Kerapatan relatif suatu species (KR)
Kerapatan suatu species X 100%
Kerapatan seluruh species
40 cm
20 m
2 m
5 m
10 m
40 m
Gambar 1. Suatu Petak tunggal dalam analisis vegetasi

c. Frekuensi suatu species (F)
Σ Sub-petak ditemukan suatu sp.
Σ Seluruh sub-petak contoh
d. Dominansi suatu species (D)
d.l. Pohon, Tiang, Pancang
Luas bidang dasar suatu species
Luas petak contoh
d.2. Semai, Tumbuhan bawah
Luas penutupan tajuk
Luas petak contoh
Kadang-kadang untuk semai dominansi tidak
dihitung.
e. Dominansi relatif suatu species (DR)
Dominansi suatu species X 100
Dominansi seluruh species
f. Frekuensi relatif suatu species (FR)
Frekuensi suatu species X 100
Frekuensi seluruh species g. Indeks Nilai Penting (INP)
INP = KR + FR + DR
Kadang-kadang untuk semai
INP = KR + FR

(2) . Petak ganda
Di dalam metoda ini pengambilan contoh vegetasi
dilakukan dengan menggunakan banyak petak contoh
letaknya tersebar merata. Peletakan petak contoh
sebaiknya secara sistematis. Untuk menentukan banyaknya
petak contoh dapat digunakan kurva species-area.
Sebagai ilustrasi pada gambar 2 disajikan cara
peletakan petak contoh pada metoda petak ganda.
Cara menghitung besarnya nilai
kuantitatif parameter vegetasi sama dengan metoda
petak tunggal.
Sistematik Random
Gambar 2. Desain Petak Ganda di Lapangan

F.I.2. Metoda jalur
Metoda ini paling efektif untuk mempelajari
perubahan keadaan vegetasi menurut kondisi
tanah, topografi dan elevasi. Jalur-jalur contoh
ini harus dibuat memotong garis-garis
topografi, misal tegak lurus garis pantai,
memotong sungai, dan menaik atau menurun
lereng gunung.
Untuk lebih jelasnya, contoh petak sampling
berbentuk jalur ini dapat dilihat Gambar 3.
Arah rintis
Gambar 3. Desain jalur contoh di lapangan
Perhitungan besarnya nilai kuantitatif parameter
vegetasi sama dengan metoda petak tunggal.
F.I.3. Metoda garis berpetak
Metoda ini dapat dianggap sebagai modifikasi
C
B
D
A

metoda petak ganda atau metoda jalur, yakni dengan cara
melompati satu atau lebih petak-petak dalam jalur
sehingga sepanjang rintis terdapat petak-petak pada
jarak tertentu yang sama. Gambar 4 memperlihatkan
pelaksanaan metoda garis berpetak di lapangan.
Perhitungan bersama nilai kuntitatif parameter
vegetatif sama dengan metoda petak tunggal.
F.I.4. Metoda Kombinasi antara metoda jalur dan metoda
garis berpetak
Di dalam metoda ini risalah pohon dilakukan
dengan metoda jalur dan permudaan dengan metoda garis
berpetak. Untuk lebih jelasnya desain metoda ini dapat
dilihat Gambar 5.
Jarak tertentu sama
Arah rintis
B
C
D
A
Gambar 4. Desain metoda garis berpetak

Perhitungan besarnya nilai kuantitatif parameter
vegetasi sama dengan metode petak tunggal.
F.2. Metoda tanpa petak
Di dalam metoda ini terlebih dahulu dibuat garis-
garis rintis dengan arah azimuth tertentu. Dengan jarak
tertentu (secara sistematis atau acak) di sepanjang
gatis tersebut dibuat titik pengukuran di mana
dilakukan pendaftaran dan pengukuran pohon.
F.2.1. Metoda Bitterlich
Di dalam metoda ini pengukuran dilakukan dengan
Tongkat Bitterlich (tongkat sepanjang 66 cm yang
ujungnya dipasangi alat seng berbentuk bujur sangkar
berukuran 2 X 2 cm). Dengan mengangkat tongkat setinggi
mata, plot seng diarahkan ke pohon-pohon yang ada di
sekelilingnya.
20 m
10 m
5 m 2 m
Gambar 5. Desain Kombinasi Metoda Jalur dan Metoda garis Berpetak
Arah rintis
B
C
D
A

Pohon yang tampak berdiameter lebih besar dan
sama dengan plot seng didaftar namanya dan diukur.
Sedangkan pohon yang tampak berdiameter lebih kecil
dari sisi plot seng tidak masuk hitungan.
Untuk setiap jenis ditentukan luas bidang
dasarnya dengan rumus:
N B = X 2,3 m2/ha
n
dimana : N = banyaknya pohon dari jenis yang
bersangkutan.
n = banyaknya titik-titik pengamatan
dimana jenis itu ditemukan.
2.3 = faktor bidang dasar untuk alat.
F.2.2. Metoda titik guadran (point quarter method)
Di dalam metoda ini di setiap titik pengukuran
dibuat garis absis dan ordinat khayalan, sehingga di
setiap titik pengukuran terdapat 4 buah quadran.
Pilih satu pohon di setiap kuadran yang letaknya
paling dekat dengan titik pengukuran dan ukur jarak
dari masing-masing pohon tersebut ke titik
pengukuran. Pengukuran dimensi pohon hanya dilakukan
terhadap keempat pohon yang terpilih.
Gambar 6 memperlihatkan pelaksanaan metoda ini
di lapangan.
Gambar 6. Desain Point quarter method di lapangan

Perhitungan besarnya nilai kuantitatif
parameter vegetasi adalah sebagai berikut :
a. Jarak rata-rata individu pohon ke titik pengukuran
dl + d2 + .......... + dn d =
n
dimana: d = jarak ind. pohon ke titik pengukuran di
setiap kuadran
n = banyaknya pohon
b. Kerapatan total semua jenis
Unit area
(d)2
(d)2 adalah rata-rata unit area/ind., yaitu rata-
rata luasan permukaan tanah yang diokupasi oleh satu
ind. tumbuhan.
c. Kerapatan relatif suatu jenis
Jumlah individu suatu jenis —————————————————————————- X 100% Jumlah individu semua jenis
d. Kerapatan suatu jenis
Kerapatan relatif suatu jenis ——————————————————————————— X
100
e. Dominansi suatu jenis
Kerapatan suatu jenis X dominansi rata-rata per
jenis
Kerapatan total semua jenis

f. Dominansi relatif suatu jenis
Dominansi suatu jenis X 100%
Dominansi seluruh jenis
g. Frekuensi Jumlah titik ditemukannya suatu jenis Suatu jenis =
Jumlah semua titik pengukuran
h. Frekuensi relative
Frekuensi suatu jenis X 100%
Frekuensi semua jenis
i . INP = KR + FR + DR
F.2.3. Metoda berpasangan acak (random pair method)
Di dalam metoda ini di setiap titik pengukuran
pilih-lah salah satu pohon yang terdekat dengan titik
pengamatan tersebut. Kemudian hubungkan pohon tersebut
dengan sebuah garis ke titik pengukuran. Buat sebuah
garis yang tegak lurus garis pertma dan pilihlah sebuah
pohon yang terdekat dengan pohon pertama tapi letaknya
di dalam sektor lain yang dibatasi oleh garis yang
ditarik tadi. Setelah jarak antara pohon pertama dan
kedua dicatat. Untuk lebih jelasnya pelaksanaan metoda
ini di lapangan dapat dilihat Gambar 7.
900 900
Gambar 7. Ilustrasi metoda berpasangan acak dalam analisis vegetasi

Besarnya nilai parameter vegetasi dihitung dengan
rumus-rumus sebagai berikut:
a. Kerapatan seluruh jenis
Unit area (Luas)
0.8 X jarak pohon rata-rata
b. Rumus lainnya sama dengan cara kuadran.
F.2.4. Metoda titik intersept (point intercept method)
Metoda ini cocok untuk komunitas tumbuhan bawah
seperti rumput, herba dan semak.
Dalam pelaksanaannya di lapangan dapat digunakan alat
pembantu seperti terlihat pada Gambar 8.

Dengan mengangkat dan menyentuhkan pin
yang terbuat dari kawat yang maka kita catat jenis
apa yang tersentuh sehingga dominansi dari jenis
tersebut dapat dihitung dengan rumus :
a. Dominansi suatu jenis
Σ sentuhan suatu jenis
Σ seluruh sentuhan
b. Dominansi relatif suatu jenis
Dominansi suatu jenis X100%
Dominansi seluruh jenis
c. Rumus-rumus lainnya sama dengan metoda dengan petak.
Hal yang sama dapat dilakukan dengan alat b dengan
cara memindahkan alat tersebut pada plot contoh tiap
10 cm, sehingga didapatkan dominansi dari jenis-
jenis yang tersentuh.
F.2.5. Metoda garis intersep (line intercept method)
Cara ini digunakan untuk komunitas padang rumput,
semak/belukar.
Prosedure pelaksanaan di lapangan:
- salah satu sisi areal dibuat garis dasar
- garis dasar tersebut menjadi tempat titik tolak
garis intersep.
- letakan garis-garis intersep secara random atau
sitema-tik pada areal yang akan diteliti.
Garis intersep sebaiknya berupa :
- pita ukur dengan panjang 50 - 100 kaki (1 kaki =
30.48 cm) .
- tambang, tali

Alat tersebut dibagi ke dalam interval-interval
jarak tertentu.
Hanya tumbuh-tumbuhan yang tersentuh, di atas atau di
bawah garis intersep yang diinventarisir. Jenis data yang
diinventarisir adalah :
(1) panjang garis yang tersentuh oleh setiap individu
tumbuhan.
(2) panjang segmen garis yang berupa tanah kosong.
(3) jumlah interval yang diisi oleh setiap species.
(4) lebar maksimum turnbuhan yang disentuh garis intersep.
Sebaiknya, kalau komunitas tumbuhan terdiri atas
beberapa strata, penarikan contoh dilaksanakan secara
terpisah-pisah untuk setiap strata.
Besaran/parameter vegetasi yang dihitung adalah :
(1) jumlah individu setiap jenis (N).
(2) Total panjang intersep setiap jenis (I).
(3) Jumlah interval transek/garis ditemukannya suatu jenis (G).
(4) Total dari kebalikan dari lebar tumbuhan maksimum (X 1/m).
(5) Kerapatan suatu jenis
unit area (E 1/m) = (——————————————————————————)
total panjang garis intersep
(6) Kerapatan Relatif suatu jenis kerapatan suatu jenis
X 100% kerapatan seluruh jenis

(7). Dominansi suatu jenis
Total panjang garis intersep suatu jenis X100%
Total panjang garis intersep
(8). Dominansi Relatif suatu jenis
Total panjang garis intersep suatu jenis X100%
Total panjang garis intersep semua jenis
(9). Frekuensi suatu jenis
Σ interval ditemukannya suatu jenis
Σ semua interval transek
(10).Frekuensi Relatif suatu jenis
Frekuensi yang dipertimbangkan untuk suatu jenis
X 100% Total frekuensi yang dipertimbangkan untuk semua jenis Frekuensi yang dipertimbangkan =
Σ 1/m F =
N
(ll). INP = KR + FR + DR.

VII. HUBUNGAN MASYARAKAT TUMBUHAN DENGAN LINGKUNGAN
A. Pengertian Lingkungan
Lingkungan adalah suatu sistem yang kompleks di
mana berbagai factor berpengaruh timbale balik satu
sama lain dan dengan komunitas organism hidup.
Satu/beberapa factor lingkungan dikatakan penting bila
berada pada taraf minimal, maksimal dan optimal menurut
batas toleransi dari tumbuh-tumbuhan sehingga factor-
faktor tersebut sangat mempengaruhi tumbih dan hidupnya
tumbuh-tumbuhan. Satu atau beberapa organism/tumbuhan.
Factor penghambat adalah setiap keadaan jumlah
sesuatu zat atau derajat sesuatu factor fisik yang
berada dekat atau melampaui batas-batas toleransi.
Kisaran toleransi organism terhadap lingkungan ada dua
macam, yaitu:
1). Steno (sempit)
2). Eury (lebar)
Setiap organism kemungkinan hidupnya dibatasi oleh:
1. jumlah dan variabilitas zat-zat tertentu yang ada,
kebutuhan minimum, dan factor-faktor fisik yang
kritis
2. batas-batas toleransi dari masing-masing organism
terhadap factor-faktor itu dan factor-faktor lainnya
B. Tujuan Pengamatan Faktor Lingkungan
Tujuan pengamatan/analisa factor lingkungan di
dalam kajian ekologis adalah:
1. merumuskan factor-faktor mana yang operasionil
penting
2. menentukan bagaimana pengaruh factor-faktor itu
terhadap individu, populasi dan komunitas tumbuhan
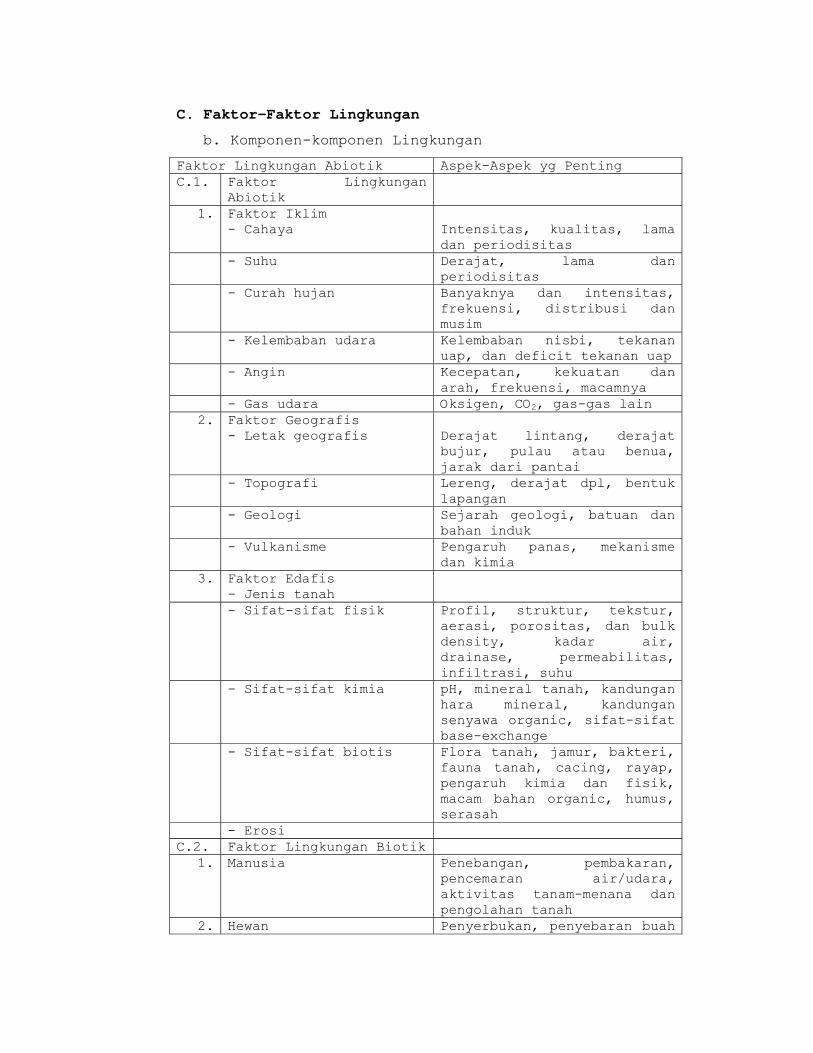
C. Faktor-Faktor Lingkungan
b. Komponen-komponen Lingkungan
Faktor Lingkungan Abiotik Aspek-Aspek yg Penting C.1. Faktor Lingkungan
Abiotik
1. Faktor Iklim - Cahaya
Intensitas, kualitas, lama dan periodisitas
- Suhu Derajat, lama dan periodisitas
- Curah hujan Banyaknya dan intensitas, frekuensi, distribusi dan musim
- Kelembaban udara Kelembaban nisbi, tekanan uap, dan deficit tekanan uap
- Angin Kecepatan, kekuatan dan arah, frekuensi, macamnya
- Gas udara Oksigen, CO2, gas-gas lain 2. Faktor Geografis
- Letak geografis Derajat lintang, derajat bujur, pulau atau benua, jarak dari pantai
- Topografi Lereng, derajat dpl, bentuk lapangan
- Geologi Sejarah geologi, batuan dan bahan induk
- Vulkanisme Pengaruh panas, mekanisme dan kimia
3. Faktor Edafis - Jenis tanah
- Sifat-sifat fisik Profil, struktur, tekstur, aerasi, porositas, dan bulk density, kadar air, drainase, permeabilitas, infiltrasi, suhu
- Sifat-sifat kimia pH, mineral tanah, kandungan hara mineral, kandungan senyawa organic, sifat-sifat base-exchange
- Sifat-sifat biotis Flora tanah, jamur, bakteri, fauna tanah, cacing, rayap, pengaruh kimia dan fisik, macam bahan organic, humus, serasah
- Erosi C.2. Faktor Lingkungan Biotik
1. Manusia Penebangan, pembakaran, pencemaran air/udara, aktivitas tanam-menana dan pengolahan tanah
2. Hewan Penyerbukan, penyebaran buah

dan biji, pengaruh kotoran, memakan dan merusak bagian-bagian tumbuh-tumbuhan, transmisi penyakit
3. Tumbuh-tumbuhan Persaingan, parasitisme, simbiosis, pengaruh toksis
D. Iklim
1. Pengertian iklim dapat dikategorikan ke dalam:
a. Iklim mikro : iklim yang nilai-nilainya berlaku
untuk tempat/ruang yang terbatas (habitat mikro)
b. Iklim makro : iklim yang nilai-nilainya berlaku
untuk daerah yang luas. Nilai-nilai iklim makro
dipergunakan untuk menetapkan tipe iklim, zona
iklim, zona vegetasi
2. Klasifikasi iklim yang paling banyak dipergunakan
di Indonesia adalah klasifikasi tipe hujan oleh
Schmidt dan Fergusson (1951). Dimana:
Jumlah rata-rata bulan kering (< 60 mm) Q =
Jumlah rata-rata bulan kering (> 100 mm)
Berdasarkan nilai Q, setiap tipe iklim mempunyai
tipe-tipe hujan sebagai berikut:
Tipe Iklim Nilai Q Keadaan Ikllim A 0 – 0.143 Tanpa musim kering, hutan hujan
tropika selalu hijau B 0.143 – 0.333 Tanpa musim kering, hutan hujan
tropika selalu hijau C 0.333 – 0.600 Musim kering nyata, peralihan
hutan hujan tropika ke hutan musim
D 0.600 – 1.000 Musim kering agak keras, hutan musim yang pohon-pohonnya gugur daun
E 1.000 – 1.670 Musim kering keras, hutan savanna
F 1.670 – 3.000 Musim kering keras, hutan savanna
G 3.000 – 7.000 Daerah kering, padang pasir H > 7.000 Daerah kering, padang pasir

Umumnya di hutan hujan, jumlah curah hujan
berkisar antara 1600-4000 mm/tahun. Di daerah
dengan curah hujan < 1000 mm/tahun ditemukan
komunitas savanna, sedangkan daerah gurun curah
hujannya ≤ 300 mm/tahun.
3. Cahaya
a. Fotosintesis
- Cahaya matahari merupakan sumber utama energy
yang diperlukan bagi kehidupan. Bagi tumbuhan,
energy cahaya melalui butir-butir daun diserap
dan dirubah menjadi energy kimia dalam bentuk
mineral molekul gula yang sederhana.
Cahaya CO2 + 2 H20 CH20 + H20 + 02 + Energi
Klorofil
Selain itu, bagai tanaman cahaya merangsang
proses diferensiasi jaringan dan sel-sel
tanaman. Untuk pertumbuhan yang normal,
minimal harus ada keseimbangan antara
fotosintesis dan respirasi. Intensitas cahaya,
dimana masukan energy tumbuhan melalui
fotosintesis dapat mengimbangi penggunaan
energy tersebut oleh respirasi disebut titik
kompensasi
- Radiasi matahari yang diterima permukaan bumi
berpanjang gelombang 300-10000 mu. Sedangkan
sekitar 40% dari radiasi total merupakan
cahaya kasat mata dengan panjang gelombang
400-750 mu. Dari energy cahaya kasat mata
tersebut hanya 1-4% yang digunakan untuk
fotosintesis. Dari spectrum cahaya kasat mata
yang terutama diabsorbsi daun untuk

fotosintesis adalah sinar biru (panjang
gelombang 450 mu) dan sinar merah (670 mu)
- Hanya sebagian kecil saja energy yang tersedia
digunakan untuk fotosintesis, selebihnya
dipantulkan, dan lalu 60-90% diabsorbsi lalu
manjadi energy panas yang sebagian besar
digunakan untuk transpirasi. Menurut Galston
(1961), dari energy matahari yang diterima
permukaan bumi, sebesar 100 000 cal/cm2/th
tersedia untuk fotosintesis daun.
b. Transpirasi
- Besarnya transpirasi bergantung pada:
a. keadaan tempat tumbuh terutama iklim.
Penguapan daun semakin banyak bila kadar
air tanah semakin tinggi
b. kesuburan komunitas tumbuhan.
Tegakan yang lebih subur di areal beriklim
basah mempunyai transpirasi yang lebih
tinggi daripada tegakan yang kurang subur.
c. Kedudukan tumbuh-tumbuhan dalam komunitas.
Di dalam huutan, strata pohon penguapannya
lebih tinggi dari strata semak
- Transpirasi berbagai jenis tumbuhan menurut
Coster (1937):
a. Tumbuhan yang penguapannya kuat (> 2000
mm/th). Albizia falcataria, Acacia
villaosa, Leucaena galuca, Eupatorium
palescens, Bambu, Samanea samans, Lantana
camara, dll. Tipe vegetasi yang
penguapannya kuat: belukar.
b. Tumbuhan yang penguapannya sedang (1000-
2000 mm/th). Tectona grandis, Hevea

brasiliensis, Imperata cylindrical,
Artocarpus integra, Coffea robusta,
Eucalyptus alba, dll.
c. Tumbuhan yang penguapannya lemah (<1000
mm/th). Thea sinensis, Cocoa nucifera,
Casuarina equisetifolia, Ficus elastic,
Garcinia mangostana, Pinus merkusii,
Agathis alba, Lagerstroemia speciosa, dll.
Tipe vegetasi yang penguapannya lemah:
hutan pegunungan.
- Selain cahaya kasat mata, ada cahaya infra
merah yang tidak dapat dilihat. Secara
ekologis, cahaya ini penting karena memberikan
efek pemanasan dan berpengaruh pada proses
perkecambahan dan pertumbuhan batang. Selain
itu ada juga sinar ultra violet yang sebagian
besar diserap oleh lapisan ozon di luar
atmosfir. Sinar ini dapat mematikan
protoplasma.
- Factor cahaya yang penting diketahui adalah:
1). Intensitas cahaya atau jumlah radiasi per
satuan luas per satuan waktu, 2). Kualitas
atau komposisi panjang gelombang, dan 3)
lamanya penyinaran dalam sehari.
- Berdasarkan adaptasi tumbuhan terhadap
perbedaan intensitas cahaya, tumbuh-tumbuhan
dikategorikan ke dalam:
a. Tumbuhan toleran (shade-adapted plant)
yaitu tumbuhan yang tahan hidup di bawah
naungan (scyophyt), missal: Altingia
excelsa, Schima noronhae, Swietenia
mahagoni, Dypterocarpus spp., Dryobalanops
aromatic, Shorea accuminatisima, Vatica

rassak, Eusyderoxylon zwageri.
b. Tumbuhan intoleran (sun-adapted atau light
demanding plant) yaitu tumbuhan yang untuk
hidupnya memerlukan banyak cahaya matahari
(Heliophyt), missal: Acacia mangium,
Paraserianthes falcataria, Eucalyptus alba,
Gmelina arborea, Pinus merkusii, Tectona
grandis, Pterospermum javanicum
- Warna cahaya mempengaruhi kecepatan
fotosintesis. Dalam hal ini warna biru dan
merah lebih berpengaruh. Lamanya penyinaran
atau periodisitas cahaya sangat berpengaruh
terhadap fase generative (pembungaan).
Berdasarkan reaksi tanaman terhadap panjang
hari, tumbuhan dibagi menjadi tiga vegetasi,
yaitu:
a. Tumbuhan hari pendek, yaitu tumbuhan yang
berbunga bila panjang hari < 12 jam, missal
ubi jalar, arbei, aster, semai, dll.
b. Tumbuhan berhaari panjang, yaitu tumbuhan
yang berbunga bila panjang hari > 12 jam,
misal: kentang, lobak, dll
c. Tanaman netral, yaitu tanaman yang tidak
dipengaruhi oleh panjang hari, misal:
tomat, nenas, kapas, ubi kayu.
Bayangan hutan merupakan bayangan merah karena
cahaya yang telah melalui daun-daun hijau tersaring dan
chaya yang diteruskan banyak mengandung sinar-sinar
hijau, merah dan infra merah. Bila hutan terlau rapat,
ada kemungkinan bayangan hutan terlalu banyak
mengandung sinar infra merah yang tidak baik bagi
pertumbuhan jenis-jenis pohon tertentu. Karenanya

permudaan banyak terdapat di tempat-tempat terbuka
dalam hutan sehingga di hutan yang rapat perlu ada
tindakan seeding cutting.
D. ADAPTASI TUMBUHAN
Warming (1895) mengklasifikasikan tumbuhan kedalam
bebera-pa grup ekologi berdasarkan persaratan tumbuh
terhadap air dan macam subtrat/tanah dimana tumbuhan
hidup.
A. Grup Ekologi Tumbuhan berdasarkan Jenis Tanah
sebagai Media Tumbuh :
1. Tumbuhan tanah asam (Oxylophytes)
2. Tumbuhan tanah basa (Halophytes)
3. Tumbuhan tanah pasir (Psammophytes)
4. Tumbuhan pada permukaan batu-batuan (Lithophytes).
Epifit tidak termasuk, karena tidak mempunyai
hubungan yang permanen dengan tanah.
B. Grup Ekologi Tumbuhan berdasarkan kebutuhannya
terhadap air :
1. Hydrophytes : tumbuhan yang hidup didalam atau
dekat air.
2. Xerophytes : tumbuhan yang hidup di habitat kering
(kurang suplai air).
3. Mesophytes : tumbuhan yang hidup di habitat yang
tidak kering juga tidak basah (lembab).
HYDROPHYTES
A. Asal Kata
Bahasa Yunani : Hudor = air
Phyton = tumbuhan

B. Pengertian
Hydrophytes : tumbuhan yang hidup ditempat basah
atau dalam air baik sebagian maupun
seluruh tubuhnya terendam
Contoh : Hydrilla, Utricularia, Eichornia, dll
C. Klasifikasi Hydrophytes
Berdasarkan hubungan dengan air dan udara,
hydrophytes dibagi kedalam:
1. Submerged hydrophytes
Tumbuhan yang tumbuh di bawah permukaan air
(tanpa adanya kontak dengan atmosfir).
Contoh : Hydrilla, Vallisneria, Nitella, dll.
2. Floating Hydrophytes
Tumbuhan yang terapung di permukaan atau sedikit
di bawah permukaan air. Tumbuhan ini ada kontak
baik dengan air maupun udara, serta tumbuhan
tersebut mungkin berakar atau tidak dalam tanah.
Floating hydrophytes dibagi kedalam dua grup,
yaitu :
a. Free floating hydrophytes
Tumbuhan yang terapung bebas di permukaan air
tanpa berakar di lumpur.
Contoh: Wolffia arhiza
Wolffia microscopic Azolla
Eichornia crassipes
b. Floating but rooted hydrophytes
Tumbuhan yang berakar dalam lumpur di dasar
perairan, tapi daun dan bunganya terapung di
atau di atas permukaan air.
Contoh : Victoria regia (water lily)
Nymphaea
Ceratopteris thaiictroides

3. Amphibious hydrophytes
Tumbuhan yang beradaptasi terhadap lingkungan
perairan dan daratan. Tumbuhan ini bisa tumbuh di
saluran air atau di subtrat lumpur. "Amphibious
plants" yang hidup di tempat pasang-
surut/payau digolongkan halophytes. Akar dan
bebernpa bagian batang terendam oleh air atau
terkubur oleh lumpur, tapi sebagian pucuk daun,
cabang, dan bunga di atas permukaan air atau
bahkan tersebar di daratan.
Bagian aerial dari tumbuhan tersebut
memperlihat-kan penampilan mesophytik atau
xerophytic, tapi bagian yang terendam
berkarakter seperti hydrophytic.
Contoh : Oryza sativa (padi)
Enhydra flutuars
Marsilea Sagittaria
Phragmites Scirpus
Alisma
Jussiaea
Neptunia
Commelina
Polygonum
Typha
Ranunculus aquatilis, dll.
Faktor-faktor yang mempengaruhi tumbuhan di lingkungan
perairan
a. Suhu air
Tumbuhan air jarang dipengaruhi oleh suhu yang
ekstrim, karena air merupakan konduktor panas
yang buruk.

b. Konsentrasi osmotik dan toksisitas air
Keadaan ini bergantung pada jumlah dan komposisi
bahan kimia yang terlarut dalam air. Fisiologis
tumbuhan air sangat dipengaruhi oleh perubahan
konsentrasi osmotik air.
C. Adaptasi Hydrophytic
Beberapa penampilan adaptif dari tumbuhan air dalam:
a. Akar
- Tumbuhan air mempunyai akar yang berkembang kurang
baik.
- Bagian akar yang berhubungan langsung dengan air
berfungsi sebagai perraukaan penyerap air dan
mineral.
- Akar pada tumbuhan air yang terapunq miskin akan
rambut akar.
- Beberapa hydrophytes berakar seperti Hydrilla
memperoleh makanan dari air melalui
permukaan tubuhnya, tetapi sebagian tergantung
pada akarnya dalara tanah dalam hal unsur
mineral.
- Akar kadang-kadang tidak ditemukan dalam
tumbuhan air seperti: Azolla, Salvinia,
Ceratophyylurn, Utricularia.
- Dalam Jussiaea repens berkembang dua macam akar
bila tumbuh dipermukaan air, yaitu akar normal
dan akar "negatively geotrophic" yang mempunyai
struktur sponge.
- Akar terapung membantu tumbuhan air dalam
kestabilan posisi terapung.
b. Batang
- Batang bersifat lunak, warna hijau/kuning.

c. Daun
- Umumnya berbulu, panjang, bulat, warna hijau
pucat/gelap, permukaan atas berhubungan bebas
dengan atmosfir, tapi bagian dibawahnya
bersentuhan dengan air.
2. Heterophylly
- Heterophyly adalah suatu penomena dimana beberapa
tumbuhan air mengembangkan dua tipe daun yang
berbeda, yaitu daun yang terendam air dan daun di
atau di atas permukaan air. Contoh : Sagittaria,
Azolla, Salvinnia.
- Daun yang terendam umumnya berbentuk pita linier
atau sedikit terpotong, dan daun yang terapung
atau di atas permukaan air berbentuk bulat atau
seperti telinga.
- Heterophylly berkaitan dengan karakteristik
fisiologi tumbuhan air sbb:
a. Pengurangan jumlah transpirasi
b. Daun lebar di/diatas permukaan air menaungi
daun yang terendam beradaptasi terhadap
intensitas cahaya yang rendah.
c. Tumbuhan air kurang menunjukkan respon terhadap
kekeringan, karena kekurangan air dapat
dikompensasi oleh daun yang terendam dalam
air.
d. Variasi dalam life-form dan habitat,
e. Daun yang lebar dipermukaan air bertranspirasi
secara aktif dan mengatur tekanan hidrostatik di
dalam tubuh tanaman.
3. Daun tumbuhan air yang terapung bebas adalah
bersifat halus dan sering dilindungi oleh "wax".

Lapisan "wax" ini berfungsi sebagai pelindung daun
dari luka kemis-fisis dan menghalangi penyumbatan
stomata oleh air.
4. Dalam beberapa tumbuhan air, petiol (daun penumpu)
menj adi bengkak dan berkembang dengan susunan
sponga relatif sedikit yang berperan sebagai
"bouyancy" dari tanaman tersebut.
5. Daun yang terendam umumnya kecil dan sempit
panjang. Hal ini berguna untuk melawan/beradaptasi
dengan aliran air. Dengan demikian, tumbuhan
tersebut dipengaruhi sedikit tekanan mekanik air.
6. Dalam "amphibious plants", daun yang berada
dipermukaan air memperlihatkan penampilan
mesophytic. Daun-daun tersebut lebih keras
daripada daun dari grup hydrophytes lainnya.
- penyerbukan dan dipersal buah dan biji
dilakukan oleh media air termasuk organisme
yang hidup di air). Buah dan biji ringan,
sehingga mudah terapung di air.
- Tumbuhan air umumnya bereproduksi secara
vegetatif. Pada Alga, reproduksi dilakukan o]eh
Zoospore dan motile/non-motile spora.
D. Modifikasi Anatomi
Dalam tumbuhan hydrofit, modifikasi anatomi berperan
sebagai :
1. Pengurangan struktur pelindung
a. Cuticle
- tidak ditemukan dibagian tanaman terendam
air.
- beberapa lapisan tipis di permukaan tubuh
tumbuhan di permukaan air.

b. Epidermis
- berfungsi sebagai alat penyerap air, hara
mineral, dan gas secara langsung dari
lingkungan air.
c. Sel epidermal mengandung kloroplas, sehingga
sel tersebut berfungsi sebagai alat
fotosintesis, terutama daun dan batang sangat
tipis, misal Hydrilla.
d. Hypodermis
- kurang berkembang
- beberapa sel yang berdinding sangat tipis.
2. Peningkatan Aerasi
a. Stomata
- tidak dijumpai pada tanaman yang terendam
air.
- pada tumbuhan terapung stomata berkembang
dengan jumlah yang terbatas pada permukaan
sebelah atas.
- pada tumbuhan amphibia stomata tersebar di
seluruh bagian tumbuhan yang kontak dengan
atmosfir dalam jumlah yang lebih besar
daripada tumbuhan terapung.
b. Lubang udara ("air chamber")
- Aerenchyma dalam daun yang terendam dan
batang berkembang dengan baik.
- “Air Chamber” diisi oleh gas dan air
- CO2 dalam “Air Chamber” digunakan dalam
fotosintesis dan O2 yang dihasilkan oleh
fotosintesis dan O2 dalam “Air Chamber”

digunakan dalam proses respirasi
- “Air chamber” mengembangkan “septa-cross”
perforasi yang halus: disphragma, yang berguna
bagi peningkatan aerasi dan mengontrol ekses
air.
- Aeranchyma berperan untuk buoyancy dan
penunjang mekanik tumbuhan air
3. Pengurangan jaringan penunjang mekanik
a. Jaringan mekanik sangat minim/tidak ada pada
tumbuhan yang terendam air, karena gaya bouyant
air memungkinkan tumbuhan terlindung dari luka
fisik. Jaringan ini berkembang dalam korteks
tumbuhan amfibi terutama di bagian yang
berhubungan dengan atmosfir
b. Dalam teratai dan beberapa tanaman lainnya,
asterosclereid (sel berlignin berbentuk bintang)
berkembang untuk menunjang tubuh tanaman.
4. Pengurangan Jaringan Pembuluh
- Karena absorbs air dan hara mineral dilakukan
oleh seluruh permukaan tubuh tumbuhan yang
terendam air, makan jaringan pembuluh tidak
begitu diperlukan
- Dalam tumbuhan air berjaringan pembuluh, xylem
tidak berkembang dengan baik. Beberapa tumbuhan
air inemperlihatkan lacuna dipusat tempat xylem
terbentuk. Lacuna berperan sebagai "air
chamber" .
- Jaringan phloem umumnya tidak berkembang dengan
baik, tetapi dalam beberapa kasus phloem
berkembang dengan baik, tapi endodermis kadang-
kadang tak dapat dibedakan secara jelas.

- Kumpulan pembuluh umurnnya berkelompok ke arah
pusat. Sedangkan pertumbuhan sekunder tidak
terjadi dalam tumbuhan air (batang dan akar).
XEROPHYTES
A. Pengertian
Tumbuhan yang tumbuh di habitat kering atau xeric.
Habitat xeric adalah habitat dimana ketersediaan,
air sangat terbatas.
B. Tipe-tipe Habitat Xeric
1. Habitat yang secara fisik kering, dimana
kkapasitas menahan air dari tanah adalah rendah
dan beriklim kering, misal gurun, permukaan
batuan, lahan kritis, dll.
2. Habitat yang secara fisiologi kering (daerah
yang banyak kelebihan air, tapi air tersebut
sukar diserap oleh tumbuhan). Habitat tersebut
mungkin terlalu masin, terlalu dingin atau
terlalu asam.
3. Habitat yang secara fisik dan fisiologis kerng,
misal lereng gunung.
- Xerophytes adalah tumbuhan karakteristik gurun
dan semigurun, tapi tumbuhan tersebut dapat
tumbuh dikondisi mesophytic dimana air yang
tersedia jumlahnya sedikit.
- Xerophytes dapat beradaptasi dengan kondisi
yang ekstrem kering, kelembaban rendah dan
suhu tinggi.
- Xerophytes yang tumbuh pada kondisi yang
kurang sesuai, maka tumbuhan tersebut
mengembangkan suatu sifat karakteristik

fisiologi dan struktur khusus yang berfungsi
untuk :
a. Mengabsorpsi air sebanyak mungkin dari
lingkungannya.
b. Menahan air dalam organ untuk periode waktu
yang lama.
c. Mengurangi transpirasi serninimal mungkin.
d. Mengontrol penggunaan konsumsi air.
C. Klasifikasi Xerophytic Berdasarkan Ketahanannya
terhadap Kekeringan.
1. Tumbuhan terhindar dari kekeringan
- Bersiklus hidup pendek
- Selama periode kering yang ekstrirn, tumbuhan
berada dalam fase buah dan biji dengan kulit
biji dan pericarp yang keras.
- Dalam kondisi yang memungkinkan, biji
berkecambah yang bersiklus hidup pendek
(beberapa minggu).
- Biji masak sebelum mendekati musim kering,
sehingga tanaman selamat dari kondisi kering
yang ekstrim. Tumbuhan demikian disebut
EPHEMERAL.
- Tumbuhan tersebut umum tumbuh di semiarid,
dimana curah hujan periodenya pendek.
- Contoh : Astragalus, Artemesia, Boaginaceae,
rumput.
2. Tumbuhan yang menderita kekeringan
- Tumbuhan berukuran kecil yang berkapasitas
untuk mentolerir atau menderita kekeringan.
3. Tumbuhan yang tahan kekeringan
- Tumbuhan ini membentuk organ adaptif untuk

bertahan terhadap kondisi kekeringan yang
ekstrim.
Xerophytes tumbuh di habitat yang berbeda :
- Tanah berbatu (Lithophytes)
- Gurun
- Pasir dan kerikil (Psamrnophytes)
- Tanah marginal (Eremophytes)
Beberapa tumbuhan habitat kering mempunyai organ
penyimpanan air. Dalam hal ini, xerophytes
dibagi dua kelompok :
a. Succulent xerophytes :
Adalah tumbuhan yang mempunyai organ-organ
yang bengkak dan berdaging akibat secara
aktif mengakumulasi air dalam organ tersebut.
Air yang disimpan dalam organ tersebut
dikonsumsi tumbuhan pada saat musim kering
yang ekstrim.
b. Non-succulent atau xerophytes sejati.
D. Adaptasi Xerophytes
1. Karakter xeromorphic
- Karakter xerofitik yang dapat diwariskan
(bersi-fat genetik) disebut xeromorhic.
Contoh : halophytic mangrove dan beberapa pchon
selalu-hijau selalu memperlihatkan
xeromorphic characters.
2. Karakter xeroplastic
- Karakter xeroplastik : karakter yang disebabkan
oleh kekeringan dan selalu berasosiasi dengan
kondisi kering. Karakter ini tidak menurun, dan
akan hilang kalau faktor lingkungan
memungkinkannya

- Penampilan xerophytic yang penting adalah
sebagai berikut :
a. Adaptasi morfologi
(1). Akar
Xerophyte mempunyai sistem perakaran
yang berkembang dengan baik. Banyak
tumbuhan dari golongan ini mempunyai
perakaran yang dalam untuk mencapai
lapisan tanah yang banyak air.
(2). Batang
a) Batang dari beberapa xerophytic
menjadi keras dan berkayu baik
batang yang aerial maupun sub-terranean
b) Batang banyak diselimuti oleh
lapisan lilin yang tebal (misal
Equisetum). Selain itu, ada juga batang
yang diselimuti bulu-bulu yang rapat
(misal Calotropis)
c) Dalam beberapa xerophytic, batang
mungkin termodifikasi menjadi
duri, misal Duranta, Ulex, dll.
d) Dalam batang yang berdaging, batang
utama sering menjadi umbi dan
berdaging dan nampaknya daun pada
tanaman tersebut muncul secara
langsung dari bagian atas akar.
e) Batang dalam beberapa xerophytic
termodifikasi menjadi daun yang
datar, hijau dan berdaging,
yang terkenal dengan istilah
phylloclades. Contohnya: kaktus
dan kokoloba (Maehlenbeckia).

(3). Daun
a) Dalam beberapa xerophytes, daun
sering "caducous" (gugur cepat),
tetapi mayoritas daun umumnya
tereduksi menjadi seperti sisik
rnisal daun Casuarina, Ruacus,
Asparagus, dll.
b) Beberapa xerophytes mempunyai daun
seperti jarum, misal Pinus.
c) Dalam daun berdaging yang berfungsi
sebagai penyimpan kelebihan air dan
getah, umumnya mempunyai batang
yang tereduksi. Contoh: Sedum acre,
Aloe spinossissima, dll.
d) Umumnya xerophytes mempunyai daun
yang tereduksi (ukurannya mengecil)
dan terlapisi oleh silika lilin dan
mengandung cuticle yang tebal.
Kadang-kadang daunnya tereduksi
menjadi duri seperti pada Opuntia
(kaktus)
e) Umumnya daun xerophytic mempunyai
daun yang kecil dengan semacam
jarum/duri dan berurat yang rapat.
f) Trichophylly
Beberapa xerophytic yang tumbuh di
daerah yang berangin kencang,
permukaan bawah daun untuk melin-
dungi stomata. Xerophytic yang daun
dan batangnya tertutupi oleh bulu
disebut "trichophy1lous plants",
misal Nerium.
g) Daun tergulung (Rolling of leaves).

Dalam hal ini stomata hanya
tersebar dipermukaan atas; dan
karena daun tergulung ke arah atas
dengan sendirinya stomata tertutup
dari atraosfir. Cara ini merupakan
modifikasi yang efektif untuk
mengurangi hilangnya air bagi
tanaman. Contoh: rumput di gurun
pasir.
(4). Bunga, buah dan biji
Bunga selalu berkembang pada
kondisi lingkungan yang
memungkinkan. Buah dan biji
terlindungi oleh kulit yang cukup
keras.
b. Adaptasi Anatomi
Bagi xerophytes, adaptasi anatomi ditujukan
untuk mengefisienkan penggunaan air.
1) Adanya deposisi lilin, lignifikasi, dan
kutinisasi pada perraukaan epidermis dan
bahkan di hipodermis.
2) Epidermis
Sel pada xerophytes adalah kecil dan
kompak. Umumnya epidermis terdiri atas
dua atau tiga lapisan memanjang secara
radial. Sel epidermis ini diselimuti
lilin, tanin, resin, selulose, dll,
membentuk suatu saringan melawan
intensitas cahaya yang tinggi.
3) Rambut
Rambut-rambut ini berperan untuk
rnelindungi stomata dan mencegah
kehilangan air yang berlebihan. Rambut

tersebut bisa sederhana atau kompleks,
uni atau multiseluler.
4) Stomata
Dalam xerophytes, reduksi transpirasi
adalah sangat penting. Hal ini dapat
terjadi bila jumlah stomata/unit area
berkurang atau stomata termodifikasi
strukturnya. Umumnya stomata pada
xerophytes bertipe cekung tenggelam.
Stomata ini bisa berada di atas permukaan
daun (rolled leave) dan di bawah
permukaan daun (dorsiventral leave).
5) Hypodermis.
Dalam xerophytes, langsung di bawah
epidermis terdapat satu atau beberapa
lapisan kompak grup sel berdinding tebal
yang membentuk hydrodermis. Hypodermis
dapat berasal dari epidermis atau korteks
(batang) atau mesophyll (daun). Kadang-
kadang hipodermis diisi oleh tanin dan
mucilage (lendir).
6) Ground tissue
a. Dalam batang, sebagian besar bagian
tubuh dibentuk oleh sklerenkim. Dalam
kasus dimana daun tereduksi (ukurannya
kecil) atau daun gugur cepat,
fotosintesis dilakukan oleh kortek
klorenkimatis terluar yang dihubungkan
oleh stomata dengan atmosfir.
Pertukaran gas secara teratur
dilakukan oleh batang.
b. Dalam batang dan daun berdaging,
ground tissue terisi oleh jaringan

parenkimatis berdinding yang menyimpan
kelebihan air, lendir, lateks, dll.
Hal ini membuat batang bengkak dan
berdaging.
c. Dalam daun, mesofil adalah sangat
kompak dan ruang antar sel semakin
sempit. Ada beberapa xerophyte yang
mesofilnya dikelilingi oleh lembaran
hipodermal yang tebal dari sklerenkim
dari semua sisi kecuali dari bawah.
Lembaran ini membentuk diafragma untuk
melawan intensitas cahaya. Jenis
xerophyte yang skhlerenkimanya
berkembang ektensif disebut tumbuhan
sklerofilous. Dalam daun berdaging,
parenkim berkembang ekstensif sebagai
penyimpan air.
d. Ruang antar sel semakin sempit. Sel-
sel dalam tubuh xerophytes adalah
sangat kecil, berdinding tebal, dan
kompak. Sel tersebut mungkin sperikal,
melingkar, atau bentuk kuboid.
e. Conducting tissue, yaitu xylem dan
phloem berkembang baik dalam tubuh
xerophytes.
c. Adaptasi Fisiologi
Semula para ahli berasumsi bahwa adaptasi
struktural dalam tubuh xerophytes sangat
berguna dalam pengurangan transpirasi.
Tetapi saat ini hasil-hasil penelitian
menunjukkan bahwa kecuali sukulen, xerophyt
sejati menunjukkan laju transpirasi yang

tinggi. Dalam kondisi yang serupa, laju
transpirasi per unit area dalam xerofit
lebih besar dari mesofit. Begitu pula
jumlah stomata/unit area daun dalam xerofit
lebih besar dari mesofit.
(1) Sukulen mengandung polisakarida,
pentosan dan sejumlah masam yang
berperan sebagai penahan panas.
Modifikasi struktural dari sukulen
xerofit diatur oleh proses fisiologi.
Sukulen terbentuk dengan baik, karena
konversi polisakarida menjadi pentosan
menyebabkan akumulasi jumlah air
berlebihan dalam sel.
(2) Pada tanaman sukulen, stomata terbuka
dimalam hari dan tertutup disiang hari.
Pada malam hari, tumbuhan berespirasi
dan menghasilkan masam. Akumulasi masam
dalam sel pelindung akan meningkatkan
konsentrasi osmotik yang menyebabkan
aliran air dalam sel pelindung
tersebut. Saat sel pelindung
membengkak, stomata terbuka. Di siang
hari, masam-masam terurai menghasilkan
C02 yang digunakan dalam fotosintesis
dan sebagai akibat dari
tekanan/konsentrasi osmotik sel, serupa
(getah) menurun yang menyebabkan
stomata tertutup.
(3) Dalam xerofit, komposisi kimia dari
cairan sel secara aktif dikonversi
kedalam dinding sel. Misalnya,
perobahan polisakarida terhadap bentuk

anhidruous seperti selulosa.
(4) Beberapa enzim seperti eatalases,
peroxidases adalah lebih aktif dalam
xerofit daripada mesofit. Dalam
xerophytes, enzim amilase
menghidrolisis pati secara aktif.
(5) Kapasitas xerophyte untuk bertahan
hidup dalam kondisi kering dalam waktu
yang lama, tidak hanya disebabkan oleh
penampilan khusus struktural, tetapi
juga oleh resistensi protoplasma
terhadap panas dan kekeringan.
(6) Pengaturan transpirasi
Adanya cuticle, permukaan yang
licin, sel yang kompak dan
stomata yang cekung dilindungi
oleh rarnbut stomata untuk
mengatur transpirasi.
(7) Cairan sel bertekanan osmotik tinggi
Fenomena ini akan meningkatkan
pembengkakan sel yang menekan dinding
sel. Cairan sel yang bertekanan osmotik
tinggi juga akan mempengaruhi absorbsi
air.
MESOPHYTES
Mesofit adalah tumbuhan daratan yang tumbuh
dalam kondisi tidak terlalu basah dan tidak terlalu
kering. Tumbuhan ini tidak dapat tumbuh dalam tanah
jenuh air dan tanah kering. Contoh: vegetasi hutan
hujan, meadow, dll. Mesofit yang sederhana terdiri
atas rumput dan herba, sedangkan mesofit yang lebih
kaya terdiri atas herba dan semak, dan mesofit terkaya

terdiri atas pohon-pohonan. Mesofit dapat dibagi ke
dalam dua kelompok tumbuhan, yaitu:
1. Kornunitas rumput dan herba
2. Komunitas tumbuhan berkayu
Komunitas Rumput dan Herba
Komunitas ini meliputi rumput dan herba semusim atau
tahunan. Umumnya komunitas rumput/herba berada pada
daerah dengan curah hujan tahunan antara 10" sampai
30".
Tipe-tipe komunitas rumput dan herba adalah :
1. Komunitas rumput dan herba arctic dan alpine.
Komunitas tumbuhan ini berada di daerah kutub
(arctic) dan puncak pegunungan (alpine). Tumbuhan
berupa semak yang lembut dan berukuran kecil.
Kadang-kadang tumbuhan tersebut bercampur dengan
lumut, tetapi lumut kerak umumnya tidak ada.
Komunitas tumbuhan ini terdiri atas dua kategori,
yaitu :
a. Komunitas rumput
b. Komunitas herba (herba dikotiledon sepertii
Saxi-fraga, Delphirium, dll)
2. Meadow
Meadow ini dianggap sebagai penghubung antara
mesofit dan hidrofit sebab mereka tumbuh di tanah
jika kadar air 60 - 80 %. Tumbuhnya berupa herba
tahunan dengan batang yang panjang dan umumnya
berizoma (berakar rimpang). Daun berpenampilan
mesofitik yaitu tipis, lebar, datar dan globrous.
3. Komunitas tumbuhan berkayu (semak-belukar dan
hutan)

Komunitas tumbuhan ini diklasifikasikan sebagai
berikut :
a. Semak-belukar mesofitik.
Komnuitas mesofitik ini timbul kalau kondisi
lingkungan tidak sesuai untuk pertumbuhan
hutan, tetapi sangat sesuai untuk pertumbuhan
vegetasi herba. Dalam banyak tempat, semak
xerofitik dan mesofitik sering bercampur
seperti Salix, Arabis, Lathyrus, Vicea, dll.
b. Hutan bergugur daun ("deciduous forest")
- Hutan ini tumbuh pada daerah dengan tipe
hujan cukup tinggi (30" sampai 60" per
tahun), dimana temperatur moderat.
- Hutan ini terdiri atas pohon-pohon yang
menggugurkan daun. Di daerah tropika, daun
gugur pada musim panas/kering.
- Tanah hutan banyak mengandung mikroflora dan
pada akar pohon banyak dijumpai mikoriza.
- Umumnya pohon diselimuti oleh lumut dan
penyer-bukannya dilakukan oleh angin.
- Tropohyte (tumbuhan yang berobah) dapat
dikelom-pokkan dalam grup mesofitik ini.
Tropofit dijumpai di daerah tropik yang
mempunyai musim kering dan basah yang jelas,
dimana selama musim hujan tropofit akan
berperan sebagai xerofit. Sedangkan daun
mulai gugur pada permulaan musim dingin atau
panas. Jenis adaptasi yang diperil-hatkan
oleh tropofit adalah :
(1) Perlindungan tunas musim dingin yang
lebih baik.
(2) Lapisan kulit pohon yang tebal.
(3) Formasi batang dalam tanah yang

melindungi tunas tahunan dari kekeringan
dan kedinginan. Contoh : Conifer.
c. Hutan yang selalu hijau daun ("evergreen
forest")
- Hutan ini ditemukan di daerah tropika,
subtropi-ka, dan daerah temperate dari
hemisfir sebelah Selatan.
- Pohon pada hutan ini selalu hijau daun,
yaitu pohon tersebut berdaun lebih dari satu
tahun sampai daun baru muncul.
- Hutan "evergreen" dibagi kedalam tiga tips :
(1). Hutan Antartika
Hutan ini tumbuh di Wew Zealand dan
negara-negara dimana suhu tahunan
berkisar antara 5°C - 70°C dan curah
hujan cukup banyak sepanjang tahun.
Tumbuh-tumbuhan penting dalam hutan ini
adalah Konifer, Myrtaceae,
Hymenophyllaceae, lumut.
(2). Hutan Sub-tropika
Hutan ini tumbuh di daerah yang
mempunyai curah hujan tinggi, tapi
perbedaan suhu antara musim dingin dan
musim panas tidak terlalu besar. Hujan
urnurnriya turun pada musim panas.
Tumbuhan pada hutan ini adalah Caks,
Magnolias, Tamarindus dan lumut. Hutan
ini dijumpai dibagian timur USA, Brazil
Selatan, Afrika Selatan, Australia
Timur, bagian Selatan Cina, dan Jepang.
(3) Hutan hujan tropika
- Hutan ini tumbuh di daerah tropika
(sekitar garis ekuator), dengan curah

hujan tahunan sekitar 1800 mm dan
suhu di atas 24°C.
- Keadaan iklim dari hutan ini
dicirikan oleh:
a) Kelembaban tinggi (jenuh udara,
95% kelembaban)
b) Suhu yang tinggi
c) Hampir hujan setiap hari
d) Tidak ada musim kering yang
berarti
e) Tanah sangat kaya akan humus,
warna gelap, dan porous.
- Hutan dihuni oleh banyak jenis
tumbuhan dan terdiri atas beberapa
strata.
- Akar pohon banyak mengandung rnikoriza
dan saprofit/parasit (Rafflesia,
Balanophora, Monotropa, dll) .
- Epifit dan liana di hutan ini umum
dijumpai .
- Tumbuhan yang umum dijumpai di hutan
ini adalah dari anggota Leguminosae,
Lauraceae, Myrtaceae, Moraceae , dll .
- Hutan ini terdapat di bagian tengah dan
selatan Amerika, Afrika Tengah, Pulau
Pasifik, Indonesia, Malaysia, Brazil,
dan lain-lain daerah tropika.
- Hutan ini sangat penting bagi
pembangunan, karena bernilai ekonomis
tinggi.

EPIPHYTE
A. Arti Kata
Epiphyte berasal dari kata Epi (di atas),
dan phyton (tumbuhan]. Secara harfiah, epifit
adalah tumbuhan yang hidup di atas tumbuhan lain.
Secara umum, epifit adalah tumbuhan yang tumbuh
pada permukaan tumbuhan tempat bertumpu dan secara
permanen tidak berakar di tanah.
Epifit menyerap air dari atmosfir, dan
menyerap unsur hara mineral dari kulit yang busuk
dari pohon tempat bertumpu. Karena epifit ini
merupakan tumbuhan yang bersifat autotropik,
epifit mensintesis makananannya (karbohidrat)
sendiri dari air dan C02 dari atmosfir dengan
bantuan sinar matahari.
Epifit berbeda dari parasit, karena epifit
tidak memperoleh unsur hara dan air dari tumbuhan
tempat bertumpu. Begitu pula epifit berbeda dari
liat:a, karena epifit tidak berakar di tanah.
Epifit disebut juga Aerophyte atau tumbuhan
yang hidup di udara.
B. Distribusi
Epifit hidup diberbagai macam habitat yaitu :
- Permukaan tumbuhan air yang terendam.
- Permukaan batang pohon.
- Percabangan pohon.
- Permukaan daun, batu-batuan, dsb.
Beberapa epifit memilih tempat bertumpu yang

spesifik, misal Tortula pagorum (epifit
lumut)adalah lumut epifit yang spesifik tumbuh
pada batang-batang pohon di perbatasan perkotaan.
Jenis lumut ini tumbuh di atmosfir perkotaan,
karena epifit ini memerlukan suhu tinggi dan udara
berkabut untuk pertumbuhannya yang normal.
Daerah yang dingin dan lembab biasanya kaya
akan lumut (epifit). Tetapi epifit jarang terdapat
di daerah yang kering-dingin.
C. Struktural Adaptasi Epifit
Karena epifit kebutuhan airnya bergantung pada
hujan, embun dan kadar air di udara, epifit mempunyai
suatu struktural adaptasi untuk menyimpan air dan
mengurangi kekurangan (kehilangan) air berlebihan.
Adaptasi struktural yang penting bagi epifit
adalah sebagai berikut:
1. Penampilan Perakaran
a. Sistem perakaran
(1). Normal Absorbing Root.
Akar yang mengabsorbsi air, mineral dan
nutrien organik dari celah-celah yang
lembab dari kulit tumbuhan tempat
bertumpu yang membusuk.
(2). Clinging Root
Akar yang berperan untuk menjaga agar
epifit tetap melekat di permukaan

tempatnya bertumpu serta menyerap
nutrien dari humus dan debu yang
terakumu1asi di permukaan kulit tumbuhan
inang.
(3). Aerial Root
Akar yang berwarna hijau dan berspon
yang posisinya menggantung di atmosfir
dan berperan untuk menyerap air dari
udara. Akar ini dapat melakukan
fotosintesis karena mengandung
kloroplast.
b. Batang
Batang epifit yang berpembuluh bisa atau
tidak bisa berkembang dengan baik. Beberapa
epifit berbatang sukulen dan berkembang
menjadi pseudobulbous atau tuberous.
c. Daun
Umumnya epifit mempunyai daun yang jumlahnya
terbatas. Beberapa anggrek hanya mempunyai
satu daun. Kadang-kadang daun berdaging dan
berkulit. Dalam jenis Dischidia nummularia,
Platyceriutn dan Asplenium nidus daunnya
berobah ke dalam pitcher,
d. Buah, biji dan penyebarannya
Umumnya buah dan biji epifit disebarkan cieh
angin, serangga dan burung.

2. Penampilan Anatomi
a. Terbentuknya cuticle yang tebal dan stomata
yang cekung terbenam berperan untuk
mengurangi kehilangan air dari tumbuhan.
b. Epifit yang berbatang sukulen, jaringan
parenkim berkembang baik.
c. Aerial root" dari beberapa epifit dari
famili Artaceae dan Orchidaceae membentuk
suatu jaringan masif berdinding tipis
berwarna putih kehijauan yang disebut
velamen. Velamen ini merupakan suatu
jaringan higroskopis yang menyerap air
secara cepat dari atmosfir yang jenuh uap
air. Di dalam velamen terdapat exodertnis.
Sel exodermis terdiri atas :
(1). Sel berdinding tebal yang berlignin.
(2). Sel berdinding tebal yang permeable
terhadap air.
d. Struktur lain yang serupa dengan mesofit.
D. Tipe-tipe Epifit
Terdapat empat tipe epifit, yaitu :
1. Protoepifit
Epifit yang memperoleh makanan dari permukaan
tempatnya bertumpu dan atmosfir. Epifit ini
tidak membentuk struktur adaptasi khusus,
kecuali "aerial root" dengan vilamen. Contoh:
Peperomia, Dischidia, dll.
2. Hemiepifit
Epifit yang semula tumbuh dipermukaan tumbuhan
tempatnya bertumpu, tetapi kemudian epifit ini
berhubungan dengan tanah melalui akarnya.

Contoh: Scindapus officinalis. Beberapa
tumbuhan yang batangnya pemanjat tumbuh dalam
tanah, tapi secara berangsur batang bagian
bawah mati dan ujungnya hidup secara bebas
seperti hemiepifit; tumbuhan demikian disebut
Pseudoepifit.
3. Nest Epiphyte
Epifit yang mempunyai kemampuan untuk
mengumpulkan humus dan air dalam jumlah yang
cukup besar untuk keperluannya sendiri. Contoh :
anggrek.
4. Tank Epiphyte
Epifit yang rnembentuk akar pancer yang fibrous
yang berkembang baik, dimana akar tersebut tidak
berperan di dalam penyerapan air. Daun berperan
sebagai penyerap air dan pembuat makanan.
Contoh: Nidularium, Tillandsia.

VIII. FORMASI-FORMASI HUTAN DI INDONESIA
A. Zone Vegetasi di Indonesia
Letak geografis Indonesia adalah diantara dua
benua (Asia dan Australia) dan di sekitar katulistiwa.
Kondisi seperti ini mengakibatkan terjadinya zone-zone
vegetasi dan tipe-tipe hutan di Indonesia.
Zone vegetasi hutan di Indonesia adalah:
a. Zone Barat, dibawah pengaruh vegetasi Asia,
meliputi: Sumatera, Kalimantan, dimana jenis
dominan adalah Dipterocarpaceae
b. Zone Timur, dibawah pengaruh Vegetasi Australia
meliputi: Maluku, Nusa Tenggara, Irian Jaya, dimana
jenis dominan adalah Artiucariaceae dan Myrtaceae
c. Zone Peralihan, dibawah pengaruh Asia dan
Australia, meliputi: Jawa dan Sulawesi, dimana
jenis dominan adalah Araucariaceae, Myrtaceaedan,
Verbenaceae
B. Formasi Hutan utama di Indonesia
a. Hutan Payau (Mangrove)
Ciri-ciri
1. Tidak terpengaruh iklim
2. Terpengaruh pasang surut
3. Tanah tergenang air laut, lumpur, pasir atau
tanah liat
4. Tanah rendah pantai
5. Tidak ada stratum tajuk
6. Tinggi pohon mencapai 30 meter
7. Jenis pohon: dari laut ke darat: Avicennia,
Sonneratia, Rhizophora, Xylocarpus,
Lumnitzera, Bruguiera.

8. Tumbuhan bawah : Acanthus ebracteatus,
Acanthus ilicifolius, Acrostichum aureum.
9. Terdapat di pantai terlindung, teluk/tanjung,
dan muara sungai.
b. Hutan Rawa (Swamp Forest)
Ciri-ciri:
1. Tidak terpengaruh iklim
2. Tanah tergenang air tawar
3. Lokasi: di belakang hutan payau
4. Tanah rendah
5. Tajuk terdiri atas beberapa strata
6. Tinggi pohon dapat mencapai 50-60m.
7. Jenis pohon, antara lain : Baringtonia
spicata, Campnospernta sp, Dillenia sp, Dyera
sp, Gluta renghas, Shorea belangeran,
Pandanus spp
8. Terutama terdapat di Sumatera dan Kalimantan
c. Hutan Pantai (Beach Forest)
Ciri-ciri :
1. Tidak terpengaruh iklim
2. Tanah kering (tanah pasir, lempung, berbatu
karang)
3. Di pantai (tanah rendah pantai)
4. Pohon penuh epifit : Paku-pakuan & anggrek
5. Jenis pohon, antara lain: Baringtonia
asiatica, Calophyllum inophyllum, Hibiscus
tiliaceus, Casuarina equisetifolia., Cocos
nucifera, Terminalia catappa, Manilkara kauki
6. Terutama di pantai Selatan Jawa, pantai Barat
Daya Sumatera, pantai Sulawesi

d. Hutan Gambut (Peat Swamp Forest)
Ciri-ciri :
1. Iklim selalu basah
2. Tanah tergenang air gambut, lapisan gambut
±1-20 m
3. Tanah rendah rata
4. Jenis pohon al: Alstonia, Ammora, Anisoptera,
Campnosperma, Cratoxylon, Eugenia,
Dryobalanops spp, Durio carinatus, Litsea,
Koompassia malaccensis, Shorea spp., Payena,
Palaquim, Tristania, dll.
5. Terdapat di Kalbar, Kalteng, Jambi dan Sumsel
e. Hutan Kerangas (Heath Forest)
Ciri-ciri :
1. Iklim selalu basah
2. Tanah pasir podsol
3. Tanah rendah rata
4. Jenis pohon: Shorea spp, Gonystylus, Agathis
borneensis, Casuarina sumatrana, Dacridium,
Calophyllum.
5. Terdapat di Kalimantan Tengah
f. Hutan Hujan Tropika (Tropical Rain Forest)
Ciri-ciri :
1. Iklim selalu basah
2. Tanah kering dan macam-macam jenisnya
3. Tersebar di pedalaman pada tanah rendah
rata/berbukit (< 1.000 m dpi) dan pada tanah
tinggi (s/d 4.000 m dpl)
4. Berdasarkan ketinggian, ada 3 zone;
- Hutan Hujan Bawah : 2 - 1.000 m dpl

- Hutan Hujan Tengah : 1.000-3.000 m dpl
- Hutan Hujan Atas :11.000-3.000 m dpl
5. Jenis pohon pada Hutan Hujan Bawah :
suku Dipterocarpaceae meliputi genera:
Shorea, Hopea, Dipterocarpus, Vatica,
Dryobalanops.
Genus lainnya: Agathis, Altingia, Dialium,
Duabanga, Dyera, Koompassia, Octomeles
Jenis pohon pada Hutan Hujan Tengah: suku
Lauraceae, Fagaceae (Quercus, castanea,
Nothofagus), Magnoliaceae, Hammamelidaceae,
Ericaceae
Jenis pohon pada Hutan Hujan Atas: suku
Conifer (Araucaria, Dacrydium, Podocarpus),
Ericaceae, Loptospermum, Clearia, Quercus.
Hutan Hujan Tropika terutama terdapat di
Sumatera, Kalimantan, Jawa, Sulawesi,
Ma]uku dan Irian Jaya.
g. Hutan Musim (Monsoon Forest)
Ciri-ciri :
1. Iklim musim
2. Pada tanah rendah rata atau berbukit dan pada
tanah tinggi
3. Tanah kering dan macam-macam jenis tanah
4. Berdasarkan ketinggian, ada 2 zone :
- Hutan Musim Bawah : 2 - 1.000 m dpl
- Hutan Musim Tengah-Atas: 1.000-3.000 m dpl
Hutan Musim Bawah, ciri-cirinya :
- Satu stratum, campuran, rapat, unmmnya jenis
luruh daun.
- Jenis pohon : Acacia leucophloea,

Caesalpinia digyna, Dalbergia latifolia,
Tetratneles, Tamarindus indica, Tectona
grandis
Hutan Musim Tengah - Atas :
- Jenis pohonnya: Casuarina junghuhniana,
Eucalyptus spp, Pinus merkusii, Pinus
insularis
Hutan musim terdapat secara mozaik diantara
hutan hujan di: Karawang, Cirebon (Jabar),
Jateng, Jatim, Nusa Tenggara.

IX. GANGGUAN HUTAN
A. Jenis Gangguan Hutan
1. Gangguan Alam
- longsor
- hama/penyakit
- gempa bumi
- kebakaran yang terjadi secara alami
- gelombang pasang
2. Gangguan yang disebabkan oleh aktivitas manusia
(gangguan buatan).
- kebakaran tidak terkontrol
- penebangan
- perladangan
- pemukiman
- industri
- pencemaran
- dll
B. Intensitas Gangguan Hutan
Bergantung pada:
- waktu gangguan terjadi
- jenis gangguan
- luasan akbat gangguan
- pemulihan (lama waktu) dampak gangguan
- jenis komponen lingkungan yang terganggu
C. Jenis dan Akibat Gangguan pada Formasi Hutan
1. Hutan Mangrove
Jenis gangguan
- abrasi

- sedimentasi tidak terkontrol
- pemukiman
- industri/pertambangan
- pencemaran
- tambak
- pertanian
- rekreasi
- kolam penggaraman
Akibat Gangguan
- Invasi Acrosticum aureuin, Acanthus sp. dan
Derris sp.
- penggunaan komunitas tumbuhan mangrove yang
sesuai dengan kondisi habitat
- interusi air masim ke daratan
- abrasi pantai meningkat
- potensi perikanan menurun
- kehidupan satwa liar terganggu
2. Hutan Rawa
Jenis gangguan
- kebakaran
- pertanian (terutama kebun kelapa)
- pemukiman
- penebangan
- industri
Akibat Gangguan Hutan
- Invasi rumput rawa (Paspalum sp, Fibrystilis
sp, dll), Melastoma sp (harendong)
- hutan rawa sekunder
(hutan melaleuca, hutan macaranga, hutan
Ploiarium alternifolium, hutan Campnosperma
coriaceum) dengan jenis-jenis pionir lainnya

seperti Alstonia sp., ratoxylum
cochinchinense, Scleria spp., Stenochlaena
palustris, dll.
3. Hutan Pantai
Jenis Gangguan
- abrasi pantai
- gelombang pasang (tsunami)
- penebangan
- pemukiman
- industri
- rekreasi
Akibat Gangguan
- umumnya berupa perkebunan kelapa
- invasi rumput-rumputan (Ischaemum muticum,
Fhuarea involuta, Zoysia matrella) dan terna
(Nephrolopis biserrata)
- interusi air masim ke daratari/pedalaman
- abrasi pantai meningkat
4. Hutan Kerangas
Jenis Gangguan
- penebangan
- kebakaran Akibat Gangguan
- areal hutan menjadi savana
- invasi herba insectivora di tempat terbuka
seperti Drosera, Nepenthes, dan Urticularia.
Selain itu terdapat juga Myrmecodia (epifit).
- tanah tandus berpasir Hutan Musim
Jenis Gangguan
- penebangan
- kebakaran

- penggembalaan
- pertanian
Akibat Gangguan
- hutan Casuarina junghuhniana (akibat kebakaran
- hutan jati (tahan terhadap kebakaran)
- hutan campuran Dodonaea viscosa, Enggelhardtia
spicata, Homalanthus gigantheus, Vernonia
arborea, Weinmannia blumei.

X. PEMILIHAN JENIS POHON
A. Beberapa Istilah
1. Hutan Tanaman adalah tegakan pohon/hutan yang
dibina dengan penebaran biji/benih atau dengan
penanaman bibit/anakan
2. Afforestation adalah penanaman jenis-jenis pohon
hutan pada lahan yang belum pernah ditumbuhi
pohon-pohon hutan
3. Reforestation (Bhs. Perancis : Reboisation) adalah
pembinaan dengan penanaman pada lahan yang semula
berhutan dengan menggunakan jenis-jenis yang
berbeda dengan jenis semula
4. Artificial Regeneration pembinaan hutan dengan
penanaman pada lahan yang semula berhutan dengan
menggunakan jenis-jenis yang sama dengan jenis
semula
5. Tree planting (termasuk regreening/penghijauan)
penanaman tegakan pohon/hutan di luar kawasan
hutan
6. Beberapa istilah lain untuk hutan tanaman, yaitu
a. Man-made forest
b. Forest plantation
c. Timber Estate
d. Hutan Tanaman Industri (HTI)
B. Keadaan Hutan di Indonesia
Luas kawasan hutan di Indonesia meliputi lebih
dari 74 % dari luas daratan. Jika luas daratan

Indonesia 193,6 juta ha maka luas kawasan hutannya 143
juta ha. Luas hutan tersebut menurut fungsinya dibagi
menjadi hutan hutan lindung, hutan konservasi alam dan
hutan wisata, hutan produksi (produksi terbatas dan
produksi tetap) dan hutan produksi yang dapat
dikonversi seperti terlihat pada label 1.
FAO (1985) memperkirakan bahwa luas hutan di
Indonesia adalah 157 juta ha atau 1/2 dari total luas
hutan negara-negara se Asia Tenggara termasuk. Hutan
produksi, menurut Departemen Kehutanan seluas 64 Juta
ha, sedangkan FAO menilai lebih luas yaitu 73 juta ha
tahun 1980 dan 67,7 juta ha pada tahun 1985.
Tabel 1. Luas hutan Indonesia dalam juta Ha (Departemen
Kehutanan, 1985)
1 Total luas daratan 193.6 2 Total luas hutan 143.0 3 Status huutan menurut fungsi:
a. hutan lindung b. konservasi alam dan hutan wisata c. Hutan produksi:
c.1. Produksi terbatas c.2. Produksi tetap
d. Hutan produksi yg dapat dikonvsersi
30.3 19.0 64.0 30.0 34.0 30.0
4 Luas hutan untuk konsesi HPH atau dalam proses konsesi a. konsesi b. Forestry agreements
65.4
52.2 13.2
5 Luas hutan menurut tipenya a. Total area hutan hujan tropis b. Total area hutan payau/rawa c. Total area hutan sekunder d. Total area hutan lainnya
143.0 82.2 12.0 14.6 34.2
Karena berbagai kegiatan manusia, hutan di
Indonesia telah banyak mengalami kerusakan
(deforestasi) sejak tahun 1960, terutama hutan hujan

tropika di Kalimantan yang telah banyak mengalami
penyusutan. FAO (1985) telah melaporkan bahwa laju
deforestasi di Indonesia diperkirakan 550.000 ha/tahun
sampai tahun 1980, kemudian meningkat menjadi 700.000
ha/tahun sampai tahun 1985 jauh di atas rata-rata laju
deforestaai di Asia Tenggara. Diperkirakan bahwa dari
tahun 1950 1985 telah terjadi deforestasi seluas 39
Juta ha atau 1.1 juta ha/tahun.
Data lain menyebutkan bahwa dari luas hutan alam
di Indonesia yang sebesar 143 juta ha, saat ini (1990)
yang masih utuh adalah 119,7 juta ha, sisanya seluas
23,3 juta ha menjadi areal semak belukar dan areal
bekas perladangan. Deforestasi karena perladangan
berpindah diperkirakan sebesar 400.000 ha/tahunn. Land
clearing karena program transmigrasi 200.000 - 300.000
ha/tahun, sedangkan karena pengaruh penebangan seluss
40.000 - 80.000 ha/tahun. Kerusakan akibat pengumpulan
kayu bakar yang ternyata tidak begitu serius bila
dibandingkan dengan total deforestasi, kecuali di pulau
Jawa.
Data dari Statistik Kehutanan (1987) menyebutkan
bahwa luas lahan kritis pada tahun 1986/1987 di
Indonesia di dalam kawasan hutan 4.316,540 ha dan
diluar kawasan hutan 6.080.060 ha. Sedangkan realisasi
reboisasi (reforestation) pada tahun 1985/1986 hanya
seluas 59.910 ha dan penghijauan (afforestation) hanya
seluas 200.462 ha. Tampak bahwa laju kerusakan hutan
jauh lebih tinggi daripada laju pemulihannya.
Dalam rangka mengurangi tekanan terhadap
kelestarian hutan alam Pemerintah, dalam hal ini
Departemen Kehutanan pada tahun 1984 telah mencanangkan
pembangunan Hutan Tanaman Industri (Timber Estate)
seluas 6,2 Juta ha yang ditargetkan selesai dalam masa

15-20 tahun. Dari luas tersebut telah ada hutan tanaman.
terutama di Jawa seluas 1,8 juta sehingga luas HTI yang
hams dibangun seluas 4,4 juta ha dan diharapkan pada
waktunya akan mampu menghasilkan kayu sebanyak 90 iuta m3
setiap tahunnya, baik untuk kebutuhan kayu pertukangan,
kayu energi dan keperluan pulp dan kertas.
Salah satu masalah utama dalam pelaksanaan kegiatan
penanaman, balk untuk HTI ataupun kegiatan reboisasi dan
penghijauan adalah pemilihan jenis pohon yang akan
ditanam.
C. Dasar Pemikiran Dalam Pemilihan Jenis
Pemilihan Jenis merupakan salah satu kegiatan utama
dalam penanaman. Kegiatan ini sesungguhnya termasuk dalam
praktek silvikultur, namun mengingat banyak aspek
lingkungan yang terlibat, balk lingkungan lahan yang akan
ditanami dan persyaratan lingkungan setiap jenis yang
akan ditanam maka aspek ekologi sangat menentukan
keberhasilannya.
Prinsip umum yang menjadi pertimbangan dalam
pemilihan jenis pohon yang akan ditanam harus inemenuhi
tiga prinsip kelayakan, yaitu kelayakan ekologis-ekonomis
dan sosial. Prinsip umum tersebut dapab dijabarkan
menjadi lebih rinci yang menjadi pertimbangan dalam
pemilihan jenis pohon adalah:
1. Sasaran penanaman, yang menyangkut kondisi maupun
fungsi/status areal yang akan ditanam, misalnya hutan
produksi, hutan lindung, hutan konservasi atau areal
di luar kawasan hutan dengan kondisi bekas tebangan,
padang alang-alang, tanah kosong atau lahan kritis
lainnya.
2. Tujuan penanaman, yang menyangkut tujuan akhir dari
penanaman misalnya untuk tujuan konservasi tanah dan

air, untuk produksi kayu atau untuk meningkatkan
pendapatan dan memenuhi kebutuhan masyarakat seki-
tarnya. Untuk tujuan produksi kayu terutama untuk
memenuhi kebutuhan bahan-baku induotri. yaitu : kayu
pertukangan, kayu energi dan kebutuhan pulp dan ker-
tas.
3. Kesesuaian ekologis, yaitu kecocokan antara persya-
ratan ekologis jenis terpilih dengan fakt.or-i iktor
ekologis lahan yang akan ditanami.
Apabila penanaman akan dilakukan di hutan
produksi dengan maksud untuk memproduksi kayu (misalnya
untuk pembangunan HTI) maka kriteria jenis pohon yang
alcan ditanam adalah :
1. Kayu yang bernilai tinggi dengan prospek pemasaran
yang baik
2. Kesesuaian tempat tumbuh
3. Jenis-jenis yang khas Indonesia dengan keunggulan-
keunggulan tertentu
4. Riap yang tinggi, misalnya dengan riap :
- Kayu pertukangan : 15 m3/ha/tahun
- Kayu serat : 25 m3/ha/tahuri
- Kayu energi : 35 m3/ha/tahun
5. Daur yang (relatif) rendah, misalnya :
- Kayu pertukangan : 10-30 tahun
- Kayu serat : S - 20 tahun
- Kayu energi : ± 5 tahun
6. Kualitas kayu dan bentuk batang yang sesuai dengan
persyaratan bahan untuk jenis industri yang
bersangkutan
Dalam reboisasi di hutan lindung tujuan penanaman
dititikberatkan pada aspek pengawetan tanah dan air

(hiroorologi). Untuk itu pemllihan jenis pohon
diutamakan yang mempunyai persyaratan :
1. Pohon-pohon yang perakaran utamanya tumbuh cepat
kedalam tanah dan mempunyai susunan akar permukaan
yang berkembang dengan kuat dan intensif,
2. Pohon-pohon yang cepat tumbuh sehingga secepat mung-
kin menutup tanah dan mengurangi bahaya banjir dan
erosi. Akan lebih balk apabila merupakan jenis cam-
puran antara tanaman cepat tumbuh (fast growing spe-
cies) dan jenis lambat tumbuh (slow growing species)
atau campuran antara jenis toleran dan setengtil.
toleran terhadap naungan.
3. Hutan lindung yang terdapat di daerah yang curah
hujannya tinggi dipilih jenis-jenis yang
penguapannya tinggi sebaliknya di daerah yang curah
hujannya rendah dipilih jenis pohon yang
penguapannya kecil.
Untuk lahan terbuka aeperti padang alang-alang
dan tanah-tanah kritis persyaratan jenis yang akan
ditanam adalah sebagai berikut :
1. Mampu tumbuh di tempat terbuka di bawah sinar
matahari penuh, Jadi termasuk jenis-jenis pohon
intoleran dan pionir.
2. Mampu bersaing dengan alang-alang dan gulma lainnya.
Jadi dipilih yang cepat tumbuh tingginya dan
agresif.
3. Mudah bertunas kembali, bila terbakar atau dipangkas
4. Sesuai dengan keadaan tanah yang kurus dan mi skin
hara, serta tahan kekeringan.
5. Biji atau bagian vegetatif untuk pembiakannya mudah
diperoleh dan mudah disimpan.
6. Untuk daerah yang aering terbakar harus dilengkapi

dengan jalur isolasi (misalnya jaur hijau/sekat
bakar) dengan jenis-jenis yang tahan api atau
jenis yang mudah tumbuh setelah terbakar.
7. Khusus untuk penghijauan, ditambah lagi dengan
syarat harus disenangi oleh rakyat/masyarakat,
sehingga merangsang mereka untuk menanam dan
memeliharanya, karena bermanfaat.
Pada areal semak belukar (hutan sekunder muda)
dipilih jenis-jenis yang waktu muda memerlukan naungan
(setengah toleran). Apabila di daerah ini banyak
permudaan jenis pohon komersial dapat dilakukan
tindakan penanaman perkayaan (enrichment planting) dan
pembebasan permudaan dari tumbuhan pengganggu (gulma).
Pada areal bekas tebangan sesuai dengan pedoman
TPTI (1993) dilaksanakan penanaman perkayaan
(enrichment planting) atau rehabilitasi pada areal-
areal yang permudaan alamnya kurang yang bertujuan
untuk memper-baiki komposisi Jenis dan penyebaran
permudaan Jenis niagawi. Jenis pohon yang dipilih
adalah jenis-jenis pohon komersial yang dominan dan
bersifat toleran sedangkan untuk penanaman di areal
terbuka dipilih jenis komersial yang suka cahaya atau
jenis pioner. Areal penanaman adalah areal yang kosong
akan permudaan yang luasnya mengelompok tidak kurang
dari 1 ha.
D. Persyaratan Ekologis
Faktor yang tidak kalah pentingnya didalam
menunjang keberhasilan pembangunan hutan tanaman,
seperti yang telah disinggung di atas, adalah
persyaratan ekologis. Persyaratan ekologis pada

dasarnya adalah mengkombinasinakan antara persyaratan
ekologis jenis terpilih dengan faktor-faktor ekologis
lahan yang akan ditanami. Jawaban yang paling tepat
untuk hal itu bfialah jenis asli, karena jenis-jenis
asli setempat adalah jenis terbaik yang sudah
beradaptasi dalam waktu cukup lama dan sudah teruji
kemampuaannya menghsdapi gangguan dan hambatan tumbuh
dari alam.
Apabila terpaksa didatangkan jenis dari luar atau
jenis asing (jenis-jenis exot) baik dari luar daerah
atau luar pulau jenis-jenis tersebut harus mempunyai
persyaratan tempat tumbuh yang sama dengan tempat
tumbuh lahan yang akan ditaiiami atau mampu
menyesuaikan diri iklim dan lingkungan hidup yang baru.
Oleh kareria itu, Jenis-jenis terpilih pertama-
tama harus sesuai dengan keadaan ekologi tempat tumbuh
yang baru atau jenis yang berasal dfiri tempat yang
keadaan iklim dan tanahnya serupa atau mendekati sama
dengan di tempat yang baru. Jika mendatangkan jenis
asing sebaiknya yang berasal dari daerah tropik,
seperti dari Afrika, Amerika, dan Asia tropik.
Komponen utama yang harua diperhatikan dan
menjadi tolok ukur faktor lingkungan (keadaan ekologis)
dalam pemelihan jenis adalah : (1) iklim (curah hujan,
suhu), (2) tanah (sifat fisik, kimia, keasaman), (3)
ketinggian dari permukaan laut, dan (4) lingkungan
biotik (jasad renik seperti mikoriza, kebakaran,
gangguan binatang dan sebagainya).
1. Iklim
Setiap jenis pohon mempunyai persyaratan tumbuh
sesuai dengan kondisi iklimnya. Unsur iklim yang
menentukan pertumbuhan pohon adalah curah hujan.

Berdasarkan jumlah curah hujan serta jumlah bulan-bulan
basah dan bulan-bulan kering ini pula Schmidt dan
ferguson telah membuat klasisifikasi tipe iklim dari
tipe iklim basah (tipe iklim A) sampai daerah
kering/padang pasir (H).
2. Tanah
Setiap jenis pohon memerlukan tingkat kesuburan
tanah yang berbeda-beda mulai dari tanah subur sampai
tahan terhadap tanah yang kurus. Kadang-kadang ada
jenis yang memerlukan unsur hara tertentu agar dapat
tumbuh dengan balk. Seperti Jati (Tectona grandis)
dapat tumbuh baik pada tanah berkapur yang bersifat
alkalis, sementara Ramin (Gonystylus bancanus) justru
tumbuh dominan pada tanah gambut yang kesuburannya
sangat rendah dan mempunyai pH yang sangat rendah pula.
Sementara jenis-jenis pohon Bakau (Rhizophora spp.)
tumbuh baik pada tanah yang salinitasnya (kadar garam)
tinggi. sedangkan Gelam (Melaleuca leucadendron) dapat
tumbuh pada tanah gambut rusak yang kerkadar pirit
tinggi dan sangat beracun bagi tumbuhan lain.
3. Tinggi Tempat
Setiap jenis pohon juga mempunyai persyaratan
tumbuh berdasarkan ketinggian tempat dari permukaan
laut, mulai dari pinggir pantai sampai pegunungan dan
mempunyai selang tolerasi terhadap ketinggian yang
berbeda-beda pula. Ketinggain tempat ini berkaitan
dengan suhu dan tekanan udara yang berbeda-beda dari
satu ketinggian ke ketinggian yang lain. Pemilihan
jenis pohon sebaiknya memenuhi kisaran ketinggian
tempat ini.

4. Kebutuhan Cahaya
Setiap jenis pohon ada yang memerlukan cahaya
penuh selama pertumbuhan, ada yang tahan terhadap
naungan selama pertumbuhan dan ada yang sewaktu muda
perlu naungan tetapi setelah dewasa perlu cahaya penuh,
atu sering disebut pohon intoleran, pohon toleran dan
pohon setengah toleran (semi toleran).
5. Kesarangan Tanah
Kesarangan tanah ini terutama adalah kondisi
tanah dalam hubungannya dengan tergenang tidaknya oleh
air atau berhubungan dengan baik buruknya drainase
tanah. Pada tanah-tanah yang jenuh dengan air seperti
tanah rawa, tanah gambut dan tanah payau jelas
berdrainase jelek. Setiap jenis mempunyai toleransi
sendiri-sendiri terhadap kondisi tanah tersebut.
Selain hal-hal diatas hal lain yang perlu
mendapatkan perhatian dalam pemelihan jenis pohon
adalah :
1. Biaya pembangunan tanaman dan manajemennya,
2. Kewaspadaan teerhadap serangan hama, penyakit dan
kebakaran.
3. Dampak terhadap positif dan negatif yang akan
ditimbulkan baik dampak sosial-ekonomi maupun dampak
terhadap faktor-faktor lingkungan
4. Cukup tersedia biji bermutu baik (unggul) dari
sumber yang kualitasnya terjamin
5. Penguasaan terhadap silvikultur jenis terpilih
6. Mampu berintegarasi dengan penggunaan lahan lain,
terutama dalam rangka peningkatan taraf hidup
masyarakat setempat misalnya dengan sistem tumpang
sari
7. Mudah diremajakan/regenerasi
Tabel Hubungan antara jenis-jenis pohon dengan keadaan ekologis (Soerianegara dan Indrawan, 1998)
No. Jenis Tipe Hujan
Kebutuhan Cahaya
Ketinggian (mdpl)
Keadaan Tanah Kedalaman Kesuburan
1 Acacia auriculiformis C,D Intoleran 0-800 Toleran terhadap tanah dangkal
Toleran terhadap tanah kurus
2. Acacia decurens A,B,C Intoleran 1000-2000 Tidak diketahui Tidak diketahui 3. Acacia catechu B,C,D Intoleran 0-800 Toleran terhadap
tanah dangkal Toleran terhadap tanah kurus
4. Agathis boornensis A,B Semitoleran 0-400 Toleran terhadap tanah dangkal
Membutuhkan tanah subur
5 Agathis labillardieri A,B Semitoleran 0-800 Membutuhkan tanah dalam
Membutuhkan tanah subur
6. Agathis lorantifolia A,B Semitoleran 400-1200 Membutuhkan tanah dalam
Membutuhkan tanah subur
7. Albizia falcataria A,B,C Intoleran 0-1200 Toleran terhadap tanah dangkal
Membutuhkan tanah subur
8. Albizia lebbeck C,D Intoleran 0-800 Membutuhkan tanah dalam
Toleran terhadap tanah kurus
9. Altingia excelsa A Toleran 600-1600 Membutuhkan tanah dalam
Membutuhkan tanah subur
10 Anthocephalus cadamba A,B,C,D Intoleran 0-1200 Toleran terhadap tanah dangkal
Tidak diketahui
11 Cassia siamea C,D Intoleran 0-800 Toleran terhadap tanah dangkal
Toleran terhadap tanah kurus
12 Castamea javanica A Toleran 300-1600 Membutuhkan tanah dalam
Membutuhkan tanah subur
13 Casuarina equisetifolia A,B,C,D Intoleran 0-400 Membutuhkan tanah dalam
Toleran terhadap tanah kurus
14 Casuarinas junghuhniana A,B,C,D Intoleran 400-1200 Membutuhkan tanah dalam
Toleran terhadap tanah kurus
15 Dalbergia latifollia B,C,D Intoleran 0-800 Toleran terhadap tanah dangkal
Toleran terhadap tanah kkurus
16 Dalbergia sisso B,C,D Intoleran 0-800 Membuthhkan tanah dalam
Toleran terhadap tanah kurus
17 Dryobalanops aromatica A Semitoleran 0-400 Tidak diketahui Tidak diketahui 18 Eucalyptus alba C,D Intoleran 0-800 Membutuhkan
tanah dalam Toleran terhadap tanah kurus
19 Eucalyptus alba Subspec. platyphylla
D Intoleran - Toleran terhadap tanah dangkal
Toleran terhadap tanah kurus
20 Eucalyptus deglupta A,B Intoleran 0-800 Membutuhkan tanah dalam
Toleran terhadap tanah kurus
21 Eucalyptus grandis C,D Intoleran 800-1200 Membtuhkan tanah dalam
Toleran terhadap tanah kurus
22 Eucalyptus salina C,D Intoleran 800-1200 Membutuhkan tanah dalam
Toleran terhadap tanah kurus
23 Eucalyptus umbellata C,D Intoleran 800-1200 Membutuhkan tanah dalam
Toleran terhadap tanah kurus
24 Gmelina arborea B,C,D Intoleran 0-800 Membutuhkan tanah dalam
Membutuhkan tanah subur
25 Lagerstomia speciosa A,B,C Semitoleran 0-400 Membutuhkan tanah dalam
Toleran terhadap tanah kurus
26 Maesopsis eminii B,C,D Intoleran 400-1200 Tidak diketahui Tidak diketahui 27 Melalueca leucadendron A,B,C,D Intoleran 0-400 Toleran terhadap
tanah dangkal Toleran terhadap tanah kurus
28 Pinus caribaea B,C,D Intoleran 0-800 Toleran terhadap tanah dangkal
Toleran terhadap tanah kurus
29 Pinus insularis B,C Intoleran 800-1200 Toleran terhadap tanah dangkal
Toleran terhadap tanah kurus
30 Pinus khasya B,C,D Intoleran 800-1200 Toleran pada tanah dangkal
Toleran terhadap tanah kurus
31 Pinus merkusii B,C,D Intoleran 200-1700 Toleran terhadap tanah dangkal
Toleran terhadap tanah kurus
32 Podocarpus imbricatus A,B,C Semitoleran 1200 Membutuhkan tanah dalam
Toleran terhadap tanah kurus
33 Pterospermum javanicum A,B,C Intoleran 0-400 Membutuhkan tanah dalam
Toleran terhadap tanah kurus
34 Santalum album C,D Intoleran 0-800 Toleran pada tanah dalam
Toleran terhadap tanah kurus
35 Schimaa noronhae A,B Toleran 800-1200 Membutuhkan tanah dalam
Membutuhkan tanah subur
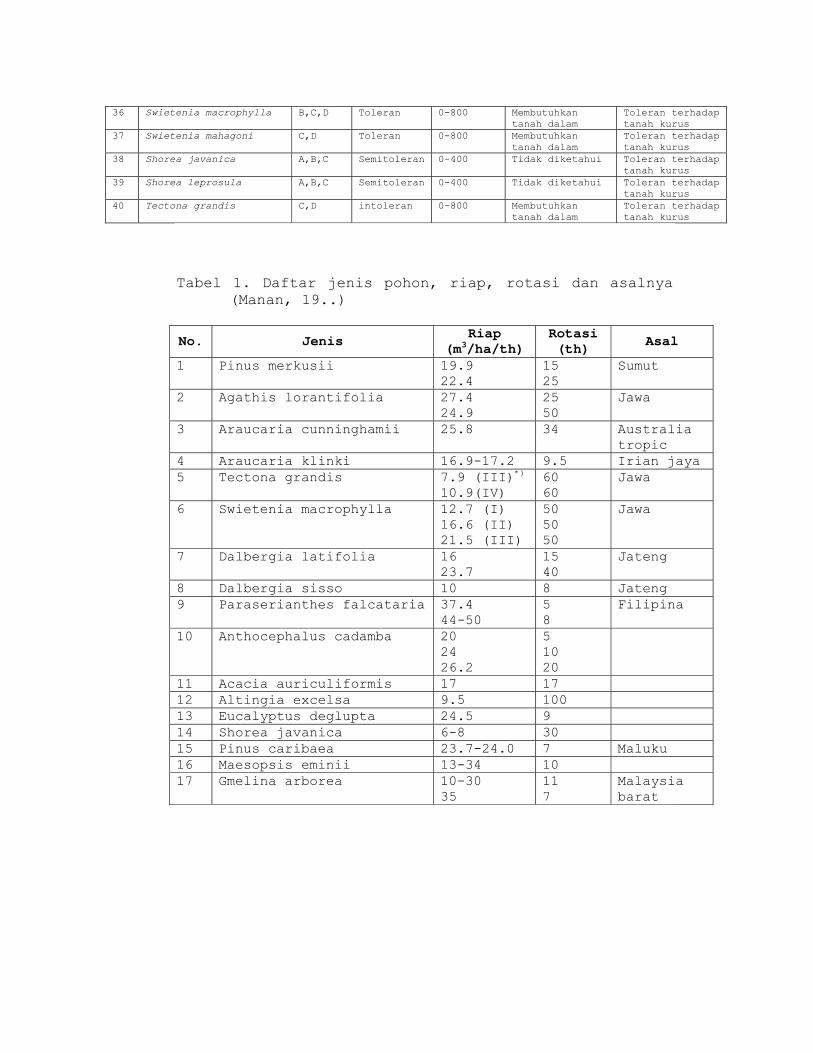
36 Swietenia macrophylla B,C,D Toleran 0-800 Membutuhkan tanah dalam
Toleran terhadap tanah kurus
37 Swietenia mahagoni C,D Toleran 0-800 Membutuhkan tanah dalam
Toleran terhadap tanah kurus
38 Shorea javanica A,B,C Semitoleran 0-400 Tidak diketahui Toleran terhadap tanah kurus
39 Shorea leprosula A,B,C Semitoleran 0-400 Tidak diketahui Toleran terhadap tanah kurus
40 Tectona grandis C,D intoleran 0-800 Membutuhkan tanah dalam
Toleran terhadap tanah kurus
Tabel 1. Daftar jenis pohon, riap, rotasi dan asalnya
(Manan, 19..)
No. Jenis Riap (m3/ha/th)
Rotasi (th) Asal
1 Pinus merkusii 19.9 22.4
15 25
Sumut
2 Agathis lorantifolia 27.4 24.9
25 50
Jawa
3 Araucaria cunninghamii 25.8 34 Australia tropic
4 Araucaria klinki 16.9-17.2 9.5 Irian jaya 5 Tectona grandis 7.9 (III)*)
10.9(IV) 60 60
Jawa
6 Swietenia macrophylla 12.7 (I) 16.6 (II) 21.5 (III)
50 50 50
Jawa
7 Dalbergia latifolia 16 23.7
15 40
Jateng
8 Dalbergia sisso 10 8 Jateng 9 Paraserianthes falcataria 37.4
44-50 5 8
Filipina
10 Anthocephalus cadamba 20 24 26.2
5 10 20
11 Acacia auriculiformis 17 17 12 Altingia excelsa 9.5 100 13 Eucalyptus deglupta 24.5 9 14 Shorea javanica 6-8 30 15 Pinus caribaea 23.7-24.0 7 Maluku 16 Maesopsis eminii 13-34 10 17 Gmelina arborea 10-30
35 11 7
Malaysia barat
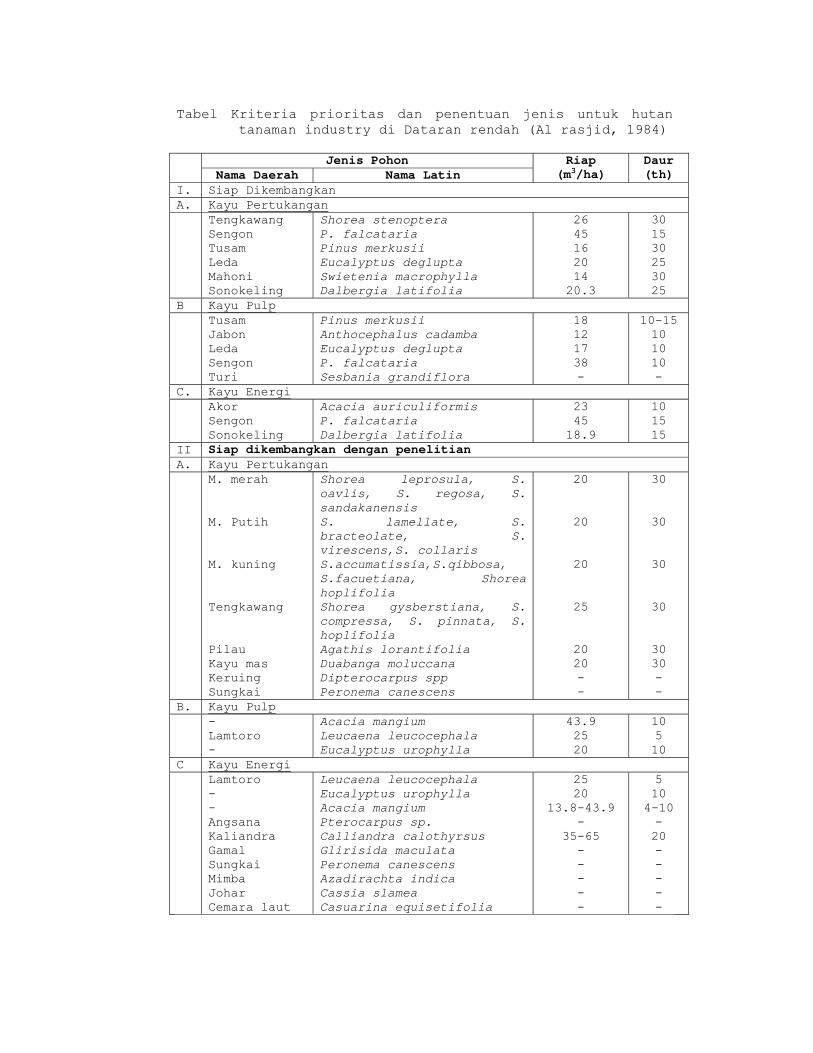
Tabel Kriteria prioritas dan penentuan jenis untuk hutan tanaman industry di Dataran rendah (Al rasjid, 1984)
Jenis Pohon Riap
(m3/ha) Daur (th) Nama Daerah Nama Latin
I. Siap Dikembangkan A. Kayu Pertukangan Tengkawang
Sengon Tusam Leda Mahoni Sonokeling
Shorea stenoptera P. falcataria Pinus merkusii Eucalyptus deglupta Swietenia macrophylla Dalbergia latifolia
26 45 16 20 14 20.3
30 15 30 25 30 25
B Kayu Pulp Tusam
Jabon Leda Sengon Turi
Pinus merkusii Anthocephalus cadamba Eucalyptus deglupta P. falcataria Sesbania grandiflora
18 12 17 38 -
10-15 10 10 10 -
C. Kayu Energi Akor
Sengon Sonokeling
Acacia auriculiformis P. falcataria Dalbergia latifolia
23 45 18.9
10 15 15
II Siap dikembangkan dengan penelitian A. Kayu Pertukangan M. merah
M. Putih M. kuning Tengkawang Pilau Kayu mas Keruing Sungkai
Shorea leprosula, S. oavlis, S. regosa, S. sandakanensis S. lamellate, S. bracteolate, S. virescens,S. collaris S.accumatissia,S.qibbosa, S.facuetiana, Shorea hoplifolia Shorea gysberstiana, S. compressa, S. pinnata, S. hoplifolia Agathis lorantifolia Duabanga moluccana Dipterocarpus spp Peronema canescens
20 20 20 25 20 20 - -
30 30 30 30 30 30 - -
B. Kayu Pulp -
Lamtoro -
Acacia mangium Leucaena leucocephala Eucalyptus urophylla
43.9 25 20
10 5 10
C Kayu Energi Lamtoro
- - Angsana Kaliandra Gamal Sungkai Mimba Johar Cemara laut
Leucaena leucocephala Eucalyptus urophylla Acacia mangium Pterocarpus sp. Calliandra calothyrsus Glirisida maculata Peronema canescens Azadirachta indica Cassia slamea Casuarina equisetifolia
25 20
13.8-43.9 -
35-65 - - - - -
5 10 4-10 - 20 - - - - -
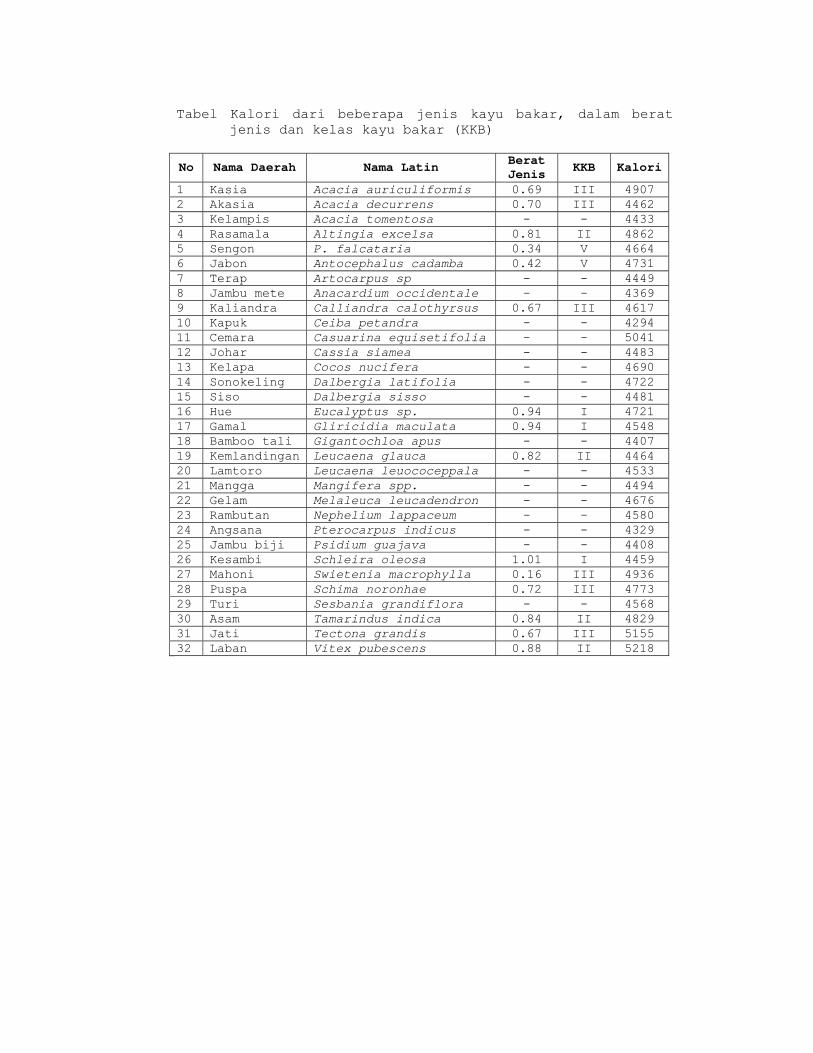
Tabel Kalori dari beberapa jenis kayu bakar, dalam berat jenis dan kelas kayu bakar (KKB)
No Nama Daerah Nama Latin Berat Jenis KKB Kalori
1 Kasia Acacia auriculiformis 0.69 III 4907 2 Akasia Acacia decurrens 0.70 III 4462 3 Kelampis Acacia tomentosa - - 4433 4 Rasamala Altingia excelsa 0.81 II 4862 5 Sengon P. falcataria 0.34 V 4664 6 Jabon Antocephalus cadamba 0.42 V 4731 7 Terap Artocarpus sp - - 4449 8 Jambu mete Anacardium occidentale - - 4369 9 Kaliandra Calliandra calothyrsus 0.67 III 4617 10 Kapuk Ceiba petandra - - 4294 11 Cemara Casuarina equisetifolia - - 5041 12 Johar Cassia siamea - - 4483 13 Kelapa Cocos nucifera - - 4690 14 Sonokeling Dalbergia latifolia - - 4722 15 Siso Dalbergia sisso - - 4481 16 Hue Eucalyptus sp. 0.94 I 4721 17 Gamal Gliricidia maculata 0.94 I 4548 18 Bamboo tali Gigantochloa apus - - 4407 19 Kemlandingan Leucaena glauca 0.82 II 4464 20 Lamtoro Leucaena leuococeppala - - 4533 21 Mangga Mangifera spp. - - 4494 22 Gelam Melaleuca leucadendron - - 4676 23 Rambutan Nephelium lappaceum - - 4580 24 Angsana Pterocarpus indicus - - 4329 25 Jambu biji Psidium guajava - - 4408 26 Kesambi Schleira oleosa 1.01 I 4459 27 Mahoni Swietenia macrophylla 0.16 III 4936 28 Puspa Schima noronhae 0.72 III 4773 29 Turi Sesbania grandiflora - - 4568 30 Asam Tamarindus indica 0.84 II 4829 31 Jati Tectona grandis 0.67 III 5155 32 Laban Vitex pubescens 0.88 II 5218

Daftar nama jenis pohon untuk ditanam bagi berbagai tujuan (manan dalam Departemen Kehutanan, 1992) A. Untuk Rehabilitasi lahan kritis 1. Hutan Lindung a. Schima noronhae, b. Altingia excelsa, c. Schima bancana, d. Peronema canescens, e. Acacia decurrens, f. Cassia siamea, g. Pterocarpus indicus, h. Duabanga maluccana, i. Schima wallchii, j. Melaleuca leucadendron, k. Eucalyptus deglupta, l. Quercus sp. , m. Shorea atenoptera, n. Aleurites inaluccana, o. Arenga pinnata, p. Anacardium occidentale, q. Parkia speciosa, r. Bambussa sp., s. Gnetum gnemon, t. Artocarpus Integra, u. Gosampinus heptaph, v. Durio zibethinus.
2. Hutan Konservasi a. Ficus benjamina , b. Tamarindus indloa, c. Antidesma bunius, d. Adenanthera pavonin e. Mangifera sp., f. Canarium sp., g. Eugenia sp. , h. Lagers trosnia sp., i. Durio sp. , j. Artocarpus integra 3. Hutan Produksia. Tectona grandis, b. Eucalyptus deglupta, c. E. urophylla, d. Acacia mangium, e. A. decurrens, f. Swietenia macrophylla, g. Pterocarpus indicus, h. Dalbergia latifolia, i. Pinus merkusii j. Paraserianthes falcataria
B. Untuk Areal Bekas Tebangan HPH
a. Dryobalanopa aromatica, b. Shorea acuminatisima , c. S. leprosula, d. S. parvifolia, e. S. pinanga, f. Gonystilus bancanus, g. Dipterocarpus lowii, h. Shorea zeylanica, i. Diospyros celebica, j. Agathis borneensis, k. A. hamii.
C. Untuk Hutan Tanaman Industri 1. Kayu Pertukangan a. Tectona grandis, b. Swietenia macrophylla, c. Parashorea sp., d. Pinus merkusli, e. Acacia mangiiun, f. Eucalyptus deglupta, g. Shorea sp., h. Dipterocarpus sp. , i. Dryobalanops aromatica, j. Gonystilus bancanus, k. Pometia pinnata, l. Peronema canescens, m. Santalum album, n. Diospyros celebica.
2. Pulp dan Rayona. Pinus merkusii b. Agathis lorantifolia, c. Eucalyptus urophylla, d. Acacia mangium e. Eucalyptus deglupta, f. Paraserianthes falcataria, g. Anthocephalus chinensis, h. Araucaria ciminghamii

3. Kayu Energi/Kayu Bakara. Duabanga moluccana, b. Leucaena leucocephala, c. Acacia mangium, d. Eucalyptus urophylla, e. Gmelina arborea, f. Acacia auriculiformis, g. Sesbania grandifolia, h. Calliandra calothyrsus, i. Casuarina equisetifolia, j. C. junghuniana, k. Glirisidia sepium, l. Rhizophora sp.
4. kayu Mewah/Indah dan Rot a. Calamus sp., b. Santalum album, c. Diospyros celebica, d. Maniikara kauki, e. Gonystilus bancanus, f. Dalbergia latilolia, g. Pericopsis moniana, h. Eusideroxylon zwageri, i. Dracontomelon mangifera, j. Pometia pinnata
Daftar Jenis asing yang mudah beradaptasi dan asalnya a. Pinus caribeae (Honduras) b. Finns oocarpa (Mexico, Guatemala) c. Pinus kesiya/insularis (Filipina, Myanmar) d. Dalbergia latifolia (India) e. Swletenia macrophylla (Amerika Tengah) f. Gliricidia spp. (Amerika Tropik) g. Calliandra spp (Guatemala) h. Gmelina arborea (India, Burma, Thailand) i. Paulownia spp. (Taiwan, Jepang) j. Leucaena leucocephalus (Amerika Tengah) k. Maesopsis eminii (Afrika Tropik) l. Khaya spp. (Afrika Tropik) m. Eucalyptus spp. (Australia) n. Acacia duurrens (Australia) o. Ochroma lagopus (Amerika tropik) Daftar jenis asli Indonesia yang mudah dibudidayakan a. Pinus merkusii (Aceh, Tapanuli) b. Tectona erandis (Jatim, NT, Muna) c. Agathis iorantifolia (Maluku) d. Agathia boorneenis (Sampit) e. Paraseriantes falcataria (Maluku) f. Aleurithes moluccana (Jawa, Wetar, Timor) g. Anthocephalus cadamba (Kepulauan Indonesia) h. Altingia excelsa (Bukit Barisan, Priangan) i. Schima noronhae (Jabar) j. Peronema can&scens (Sumsel, Kalsel) k. Dyospyros celebica (Sulawesi) l. Manilkara kauki (Ball) m. Eucalyptus deglugta (Sulawesi) n. Casuarina equsetifolia (daerah pantai Indonesia) o. Casuarina mqntana (Jatim, Nusa Tenggara, Sulawesi) p. Macadamia hildebrandii (Sulawesi) q. Podocarpuf imbricatus (Jawa) r. Fragraea fragrans (Sumatera) s. Eucalyptus alba (Timor) t. Shorea leprosula (Sumatera, Kalimantan) u. Shorea javanica (Sumsel, Tapanuli, Subah) v. Santalum album (Timor)

XI. BIOLOGI REPRODUKSI TUMBUHAN DI HUTAN TROPIS
PENDAHULUAN
Penyerbukan pohon-pohon hutan, liana dan tanaman merambat,
epifita, semak belukar dan herba pada hutan tropis melibatkan semua
antar hubungan yang biasa antara tanaman dengan hewan antopilus yang
tampak pada daerah-daerah temperate (beriklim sedang), dengan
penyerbukan tumbuhan oleh kelelawar. Juga, sebaliknya dari banyak
formasi hutan temperate, penyerbukan angin jarang {tidak biasa}
sebagai mekanisme seleksi (Whitehead, 1969) dan penyerbukan diri
sendiri tampaknya kurang umum daripada penyerbukan silang (Bawa,
1974, 1979). Demikian juga, penyebaran benih/biji oleh angin lebih
jarang daripada penyebaran biji oleh burung dan mamalia yang setelah
memakan daging buahnya lalu membuang bijinya lewat muntahan atau
kotoran, pada jarak tertentu dari tanaman induk (Van der Pijl, 1969;
Janzen, 1975).
Aspek lain dari biologi reproduksi hutan ini (termasuk
pembentukan semaian) terdapat sejumlah peningkatan yang sama dalam
interaksi biotik yang tampak pada penyerbukan dan penyebaran benih.
Oleh karena itu pertimbangan reproduksi dari sebuah ekosistem
haruslah menjadi tujuan akhir kita (Baker, 1979}. Namun, hal ini baru
dicobakan pada sejumlah kecil penelitian hutan tropis Amerika, Asia
dan Afrika. Oleh karena itu, bab ini akan berisi sebuah pertimbangan
mengenai penyerbukan dan penyebaran benih, di bawah/dalam berbagai
pokok bahasan yang beragam, dimana individu spesies atau kelompok
spesies lebih menjadi bahari pertimbangan daripada ekosistem;
sintesis dan uraian tentang sifat-sifat yang muncul ketika kita
berpindah dari tingkat individu menuju suatu penanganan ekosistem
yang holistik (menyeluruh) untuk sebagian besar belum mungkin
diperoieh. Laporan ini tidak akan merujuk pada semua kepustakaan
tentang individu spesies yang ada, bagaimanapun, karena topik ini
terpencar pada berbagai jurnal/majalah yang sebagiannya sulit
didapat. Meskipun demikian, di bawah masing-masing rubrik akan

diupayakan bahasan yang menyeluruh, ilustrasi dengan contoh juga
diberikan, sering dari hutan Amerika Tengah yang sangat penulis
kenal.
REPRODUKSI BENIH DAN PERKEMBANGBIAKAN VEGETATIF
Pohon-pohon dan palma
Kadang-kadang proporsi benih yang dapat terus hidup yang
dihasilkan oleh sebuah pohon hutan tropis ternyata amat rendah
[misalnya: 0,1% untuk Endospermum (Dalbergia malaccensis) (Fabaceae,
Faboideae) dan Vemnonia arborea (Asteraceae) (Ng dalam Whitniore,
1978). Namun demikian, semaian dari pohon-pohon sering terlihat di
lantai-lantai hutan basah pada sebagian besar waktu dalam setahun,
dan pada hutan-hutan kering sekurang-kurangnya selama musim hujan.
Distribusi semaian tersebut dan anak pohon yang tumbuh darinya telah
menjadi subyek/bahan diskusi yang sunggun-sungguh dan beberapa
penemuan (kepustakaan diringkas oleh Hubbel, 1979). Hal yang kurang
jelas adalah kapasitas beberapa pohon untuk bertunas dari
tangkai/batang bawah tanah atau sistem akar (Richards, 1952,
Schnell, 1970;Janzen, 1975; Hartshorn, 1978). Kedua mekanisme
reproduksi mungkin penting dalam menjembatani perbedaan pertunasan
pohon dari tangkai/bawah tanah atau sistem akar.
Kadang-kadang pohon yang runtuh masih berhubungan sebagian
dengan sistem akamya, masih tertanam pada tanah dan "pohon-pohon"
baru (rarnets) akan bertumbuh dari tunas yang tertidur/terhenti
sepanjang batang pohon yang melintang. Sebuah contoh dibenkan oleh
Pentaclethra macroloba (Fabaceae, Mimosoideae) yang tumbuh pada
tanah basah di Finca La Selva, Costa Rica (H.G.Baker,
pers.obs.:Hartshorn, 1972,1978).
Strangler figs terkenal, sejenis gulma seperti beberapa pohon
lainnya yang mulai pembentukkannya sebagai epifit, berkembang biak
dengan biji yang dibawa oleh burung, kelelawar, atau mamalia "non-
volant" ke cabang-cabang pohon korbannya.
Tanaman palma, dimana biasanya tidak mempunyai alat reproduksi

vegetatif maka tiap generasi harus berkembang biak dengan benih,
Usia pertama kali berbunga untuk pohon-pohon hutan tropis belum
banyak diketahui. Satu penelitian (Ng, 1966) terhadap Dipterocarpa
yang dicangkok memperlihatkan bahwa banyak pohon tersebut mulai
berbunga dan berbiji sebelum berumur 30 tahun.
Semak Belukar, Pohon Pakis dan Bambu
Bagi kebanyakan semak belukar di hutan tropis, produksi
biji/benih jelas penting daiam perkembangbiakan. Meski banyak
cabang/tangkai akan berkembang, Rhizoma dan pembentukan tunas baru
dari akar samping lebih jarang dibandingkan pada hutan temperate.
Pohon pakis, dapat berkembang biak dengan cara vegetatif (Shnell,
1970).
Mengenai bambu, keadaannya benar-benar berbeda. Rumput yang
terlalu besar tersebut adatah "monocarpic" (semelparous), dengan
suatu periode waktu pertumbuhan vegetatif sebelum berbunga, yang
dapat berlangsung sampai 120 tahun pada Phyllostachys bambusoides
(McClure, 1966b; Janzen, 1976a,b). Selama waktu tersebut, rhizoma
yang luas dan pertumbuhan tunas udara dapat menghasilkan suatu semak
belukar yang sulit diternbus. Biasanya tanaman ini mati setelah
berbunga dan berbuah dan akan digantikan oleh semaian. Janzen
(1976b) menganggap lambatnya pembentukan bunga ini kurang lebih
sinkron (selaras/serernpak) pada sebuah spesies, diikuti oleh
produksi "caryopsis" yang sangat banyak, sehingga sebuah penyesuaian
yang memungkinkan untuk pembentukan dan penyebaran buah-buahan
berbiji tunggal sebelum pemangsa biji dapat menemukan mereka,
berpesta dengannya dan mengembangkan populasi besar.
Di hutan-hutan Afrika Barat, spesies dikotiledon dapat
membentuk semak belukar yang besar melalui
perkembangbiakan vegetatif dari "air-layering" (lapisan
udara) seperti pada Anthonota (Macrolobium) macrophylla
{Fabaceae, Faboidea) dan Scaphopetalum amoenum
(Sterculiaceae) {Longman dan Jenik, 1974). Contoh-contoh

juga dapat diberikan untuk tiap-tiap benua.
Tanaman merambat dan Liana
Seperti pohon ara pencekik, tanaman merambat dan liana
dapat berkembang biak melalui biji yang berkecambah pada
sebuah cabang atau batang sebuah pohon yang akhirya akan
jadi penyokongnya. Dari asal udara ini, tunas atau akar
bisa turun ke tanah. Tanaman rambat dan liana lain dapat
berkecambah di darat dan mernbuat jalan tumbuh ke atas
(Richards, 1952; Schnell, 1970; Longman dan Jenik, 1974;
Whitmore, 1975). Oleh karena tingkat keberhasilan yang
rendah dalam pembenlukan pohon merambat dan liana ini,
maka diperlukan pembentukan biji yang berlebih-lebihan.
Meskipun demikian, pohon rambat dan liana yang telah jadi
dapat ditumbuhkan kembali melalui pembentukan tunas dari
tunas yang dorman. (Janzen, 1975). Ini salah satu alasan
keberhasilan penggunaan tanaman merambat di pinggiran
hutart neotropis -Fabaceae- dalam pembentukan padang
rumput buatan di wilayah tropika Amerika dan Australia
(Baker, 1978b).
Epifit dan Parasif Tanqkai
Epifit, karena habitatnya alami, membatasi reproduksi
biji atau spora. Hal ini tidak hanya berlaku pada epifit
herbaceous yang terkenal (anggrek, bromeliads,
pteridophytes, dan lainnya) dan hemiparasit {mistletoes,
dll} tetapi juga bagi pohon-pohonn semak belukar epifit
[(misainya spesies dari Clusia (Hepericaceae) dan Blakea
(Melastomataceae)]. Reproduksi melalui epifit secara
karakteristik disempurnakan oleh produksi biji yang amat
kecil dalam jumlah yang amat besar {misainya Piperaceae,
Orchidaceae, Bromeliaceae, dil) yang dapat disebarkan oleh
angin atau, lebih sering lagi oleh burung dan mamalia yang
menelan biji dan kemudian memuntahkan atau membuang
kotorannya tepat pada cabang pohon yang menjadi inangnya.

Herba_dan Parasitakar
Peluang bagi pembentukan bunga dan penyebaran biji
oleh tanaman herba dari lantai hutan yang matang secara
khas terjadi bilamana sebuah pohon tumbang meningkatkan
iluminasi di tingkat dasar/tanah. Penerangan oleh manusia
juga memberikan fungsi persiapan yang sama. Sebaliknya, di
pedalaman hutan, perkembangbiakan dan penyebaran vegefatif
melalui pertumbuhan rhizomata merupakan hal yang umum.
[(misalnya Heiiconia (Musaceae), Caiathea (Marantaceae).
Zingiber (Zingiberaceae, dll)]
Pada intensitas pencahayaan lantai hutan yang rendah,
saprofit dan parasit mernpunyai sebuah keuntungan.
Terutama untuk paleotropis, anggota-anggota
Balanophoraceae merupakan parasit obligat pada akar-akar
tanaman kayu. Perhatian/penekanan pada reproduksi
vegetatif di sini, oleh karena pada kasus parasit akar
Thonianningia sanguinea dari Afrika, yang termasuk
dioecious, kejadian bersama benangsari dan putik hampir
tidak dikenal (Bullock, 1948), sehingga perkembangbiakan
itu harus secara eksklusif dengan cara vegetatif.
REPRODUKSI BIJI/BENIH SISTEM PERKAWINAN
Riset mengenai sistem perkawinan pada tanaman hutan
tropis dipusatkan terutama pada pohon-pohon dan belukar
hutan {East, 1940, Baker, 1959, 1976; Ashton, 1969, 1977;
Styles, 1972; Bawa, 1974; Tomlinson, 1974; Bawa dan Opler,
1975; Styles dan Khosla, 1976; Soepadmo dan Eow, 1977) dan
sebagian tanaman perkebunan (Ferwerder dan Wit, 1969;
Purseglove, 1968; Baker, 1976; Simmonds, 1976). Hanya
sedikit data tentang tanaman herba hutan, kecuali
Heliconia (J.Kress, pers.comm, 1979) dan Marantaceae
(Kennedy, 1978). Liana dan epifit hampir tidak pemah
ditetiti.
Isu/masalah utama dalam pembahasan sistem perkawinan
tanaman tropis adalah peran masing-masing dari inbreeding
(perkawinan sejenis) dan outbreeding (perkawinan antar

jenis) pada hutan dan dampaknya pembentukan populasi yang
berbeda dan khusus. (Baker, 1959; Fedorov, 1966; Ashton,
1969,1977; Bawa, 1974,1976). Sebagai akibatnya, tujuan
dari kebanyakan penelitian adalah untuk menentukan apakah
kebanyakan tanaman hutan tropis melakukan
fertilisasi/penyerbukan sendiri atau
fertilisasi/penyerbukan silang. Perlu dicatat, hal ini
berbeda dari penentuan apakah tanaman tersebut "self-
incompatible". Sebuah spesies yang self-incompatible dapat
saja menyediakan kebanyakan bijinya melalui penyerbukan
silang. Di pihak lain, harus ditekankan bahwa perkawinan
silang bisa sama dengan perkawinan yang secara genetik
sejenis jika dilakukan antar tanaman yang memiliki asal
yang sama. Jadi jelas sejumlah faktor yang berkaitan
dengan perilaku penyerbukan dan seleksinya mungkin penting
dalam evolusi sistem perkawinan pada pohon-pohon hutan
tropis (Bawa dan Opler, 1975; Frankie et al., 1976;
Janzen, 1977a,b; Bawa, 1980; Bawa dan Beach, 1981; Beach,
1981).
Geitonogamy dan Autogamy
Tingkat penyerbukan sendiri atau silang pada tanaman
Zoophilus lergantung pada sebagian besarnya dari aktivitas
pencarian makanan para penyerbuk (polinator) dimana
perilaku tersebut dipengamhi oleh variasi menurut ruang
dan waktu dari bunga (nektar/tepung). Namun demikian, pada
saat yang sama, sebuah pohon besar di hutan dapat
mengandung ratusan bunga yang terbuka siap menunggu
kedatangan penyerbuk potensial. Hampir sernua tamu
(penyerbuk) yang datang pada sebuah pohon tertentu akan
menemui banyak bunga sebelum meninggalkan pohon tersebut.
Geitonogamy, perpindahan tepung sari dari satu bunga
kepada bunga lainnya pada tanaman yang sama (sebagai
akibat penyerbukan sel telur bunga yang belakangan)
merupakan akibat pergerakan penyerbuk. Autogamy,
perpindahan tepung sari dari "anther" ke "stigma" pada

bunga yang sama, diikuti oleh fertilisasi dapat terjadi
dengan atau tanpa bantuan penyerbuk pada pohon tersebut.
Secara genetik, akibat dari geitonomi atau autogami sama,
kecuali untuk kasus yang jarang dimana bagian mahkota yang
berbeda dari sebuah pohon dapat berbeda secara genetik
melalui mutasi somatik (Bawa, 1979).
Monoecism, produksi terpisah benangsari dan putik
bunga pada pohon yang sama (seperti pada banyak Meliaceae,
Euphorbiaceae dan Arecaceae), memajukan/meningkatkan
"outcrossing" (persilangan luar) namun masih membuka
kemungkinan dari geitonomi. Tetapi, geitonomi dan autogami
hanya bisa muncui juka fungsi dari tepung sendiri pada
stigma (putik) dicegah oleh suatu mekanisrne self
incompatibilitas. Meskipun banyak pohon-pohon hutan tropis
mempunyai selfincompatibility (ketidakcocokan dalam diri
sendiri), ini bisa tidak mencukupi, oleh karena itu
penyerbukan diri dan silang mungkin terjadi pada pohon
yang sama, dan pada waktu yang sama (Bawa, 1974; Gan dkk.,
1977).
Bagi beberapa spesies pohon, terdapat laporan-laporan
yang bertentangan tentang terjadinya sistem perkawinan
yang menyatakan bahwa hal ini dapat bervariasi antara
individu atau populasi [cf. Opler dkk., (1076a, untuk
Cordiaalliodora (Boraginaceae); dan Valmajor dkk, (1965);
Baker (1970), dan Soepadmo dan Eow (1977), dan Durio
zibethinus (Bombacaceae)]
Luas fertilisasi sendiri dan silang mungkin juga
tergantung dari pengaruh iklim dan ketersediaan penyerbuk,
dan juga pada ketersediaan pohon-pohon tertentu daiam
wilayah terbang penyerbuk. Sebagai akibatnya, bagi
kebanyakan pohon-pohon hutan suiit diukur besarnya/luasnya
"outcrassing" (persilangan luar).
Outcrossinq 1: Mekanisme fisik dan waktu penyebab
outcrossing
Pada bunga besar dari beberapa pohon Bombacaceae

(misalnya Ceiba acuminata), putik terletak begitu jauh
dari benang sari pada sekuntum bunga yang mekar sehingga
tepung sari tidak dapat disimpan/diletakkan padanya tanpa
pertolongan binatang yang dimaksud. Pada kasus C.
acuminata, binatang yang dimaksud adalah kelelawar dan
burung kolibri (Baker dkk., 1971). Pemisahan secara fisik
benangsari dari putik (secara teknik/sebuah contoh
herkogamy) meningkatkan kemungkinan penyerbukan silang
(atau geitonogarni) dan dapat sesuai bagi banyak pohon-
pohon tropis lain. Sebuh contoh bagus herkogami pada
tumbuhan rambat dapat diperoleh dari Gliriosa superba
(Liliaceae).
Pelepasan oleh benangsari dan penerimaan oleh putik
dapat dibagi menurut waktu (dichogamy): pelepasan tepung
sari mendahului kesediaan putik (protandry) atau
sebaliknya (protogyny). Protandri khususnya terkenal pada
Fabaceae. Jadi, pada Hymenaea courbaril
(Caesalpinioideae), di hutan neotropis, ketika "anthesis"
pada awal malam benang sari "dehisce" sementara "style"
masih terguiung) dengan putik terlindung baik pada pusat
"coil" (gelung). Kemudian pada waktu malam, style uncoiled
dan tersajilah sebuah putik yang siap (G.W, Frankie, pers.
Obs.).
Pada rumpun Gardeniaceae dan Rubiaceae, dengan
mekanisme "protandrous" yang berbeda, style
memanjang/terulur saat "anthesis", mendorong seperti
sebuah pengisap (piston) antara benang sari yang diketahui
membelah dan serbuk sari disajikan untuk serangga
pendatang pada putik yang tidak reseptif (siap menerima).
Belakangan, ketika serbuk sari telah terlepas, putik
menjadi reseptif (siap) terhadap serbuk sari yang dibawa
dari tempat lain (Baker, 1958).
Dimana protandri melibatkan secara relatif bunga-bunga
besar di situ biasanya tidak ada sinkronisasi antara
bunga-bunga pada pohon yang sama sehingga meskipun
autogamy bisa dicegah, geitogamy tetap dimungkinkan.

Dimana protandri terlihat pada spesies yang banyak bunga-
bunga kecil berkumpul ke dalam inflorescences yang padat,
dan secara fungsional merangsang bunga besar di situ bisa
terdapat sinkroni dengan inflorescence. Jadi, pada spesies
Parkia (Fabaceae, Mimosoideae) paleotropik dan neotropik
terdapat sinkroni. Pada P. clappertoniana, inflorescence
cembung seukuran bola tenis terjadi pada benangsari bunga-
bunga di suatu malam dan pada putik di malam berikutnya
(Baker dan Harris, 1957). Meski begitu, tidak terdapat
sinkroni antara infloresensi dan sekali iagi meski
autogamy bisa dicegah, geitogamy dapat terjadi bilamana
keleiawar penyerbuk melakukannya.
Infloresensi Parkia memperlihatkan pembagian kerja
yang menarik antara bunga-bunga, sebagian berfungsi hanya
sebagai produser nectar (penghasil madu) sementara
sebagian Iagi sebagai penghasil buah (Baker dan Harris,
1957; Baker, 1978a).
Protogini, yang kurang sering dibandingkan protandri
boleh jadi merupakan sebuah mekanisme yang iebih efektif
untuk rnemajukan outcrossing, oleh karena itu serbuk
'asing" yang mencapai putik sebelum benangsari bunga itu
sendiri "dehisce" akan lebih disukai/menang dalam
perlombaan pada batang benang sari menuju ovula,
mengalahkan semua serbuk yang belakangan dilepaskan bahkan
jika pada waktu itu putik masih hams disiapkan. Dua
penelitian yang baik sebagai contoh protogini diberikan
oleh Persea americana (Lauraceae) dan Annona cherimolia
(Annonaceae) dan , tentunya, spesies Ficus (Moraceae) yang
sangat khusus (McGregor, 1976).
Outcrossing 2: Sistem incompatibiljtas
Self-inkompatibilitas (inkompatibilitas sendiri)
homomorfik ditemukan pada banyak pohon-pohon tropis yang
memperlihatkan mekanisme penghindaran/penceganan autogarni
dan geitonogami ini, tetapi terdapat juga pohon-pohon
belukar, herba dan liana yang heteromorfik pada hutan-

hutan tropis (East, 1940; Baker, 1958; Bawa, 1974; Arroyo,
1976; Opler dkk., 1976a; Haber dan Frankie, 1982).
Pada sebuah hutan yang rontokdan kering di Costa Rica,
Bawa (1974) menemukan 27 dari 34 spesies pohon berbunga
hermaphrodit merupakan "self-incompatible" sebagai sebuah
hasil percobaan bagging, selfing dan crossing. Arroyo
(1976) dan Zapata dan Arroyo (1978) menemukan suatu
proporsi yang serupa pada sebuah hutan tanah rendah di
Venezuela. Penelitlan-penelitian yang dilakukan pada hutan
hujan tropis tanah rencah di Malaya (Ashton, 1977} dan
hutan basah di Costa Rica (Bawa, unpub.) juga menunjukkan
adanya oto-inkompatibilitas pada sebuah proporsi yang
tinggi dari pohon-pohon contoh lebih jauh spesies
individual yang diperlihatkan oteh East (1940), Purseglove
(1968), Hedegart (1976), Mori dan Kallunki (1976),
Simmonds (1976) dan Gan dkk. (1977),
Sedikit sekali penentuan nubungan kompatibilitas pada
semak belukar hutan, kecuali untuk heterostylous (tumbuhan
berakar serabut) dan Fubiaceae lainnya di Afrika Barat
{Bakaer, 1958) dan di Costa Rica (Bawa dan Beadh, 1981)
dimana inkompabilitas sendiri banyak terjadi, dan untuk
spesies Cordia (Boraginaceae) yang heterostylous (self-
incompatible) dan hoostylous (self-compatible) di hutan
neotropis (Opler dkk., 1976a). Pada sebuah hutan tropis
rendah tanah basah di Costa Rica, L. McDade (pers.comm.,
1979) menemukan oto-inkompatibilitas pada sekurang-
kurangnya 4 buah spesies Aphelandra (Acanthaceae), tetapi
J.Kress (pers.comm.) telah membuktikan bahwa Heliconia
(Musaceae) mempunyai spesies-spesies baik yang self-
compatible maupun self-incomptible. Pada hulan yang sama,
K. Grove (pers.comm., 1977) menemukan beberapa herba yang
self-compatible.
Di antara liana dan tanaman merambat, Pass/flora
mucronata ditemukan self-incompatible dan self-compatible
ini banyak terjadi pada Bignoniaceae yang merambat
(Gentry, 1974a).

Pada banyak spesies, barier inkompatibilitas tidak
lengkap atau mudah menjadi rusak (contohnya, lihat Baker,
1958; Lee, 1967; Purseglove, 1968, 1975; Bawa, 1974;
Hedegart, 1976). Pada Cord/a alliodora (opier dkk., 1976a)
dan Luehea seemannii (Haber dan Frankie, 1982), terdapat
bukti eksperimental bahwa sebagian pohon self compatibel
dan sebagian lagi self-inkompatibel.
Pada hutan-hutan tropis, tampak bahwa musim berbunga
dan usia bunga secara individu berpengaruh besar pada
integritas sistem self-inkompatibilitas. Sebagai contoh,
pada Byrsonima crassifolia (Malpighiaceae) satu dari 5
pohon yang digunakan dalam percobaan awalnya ditemukan
sebagai self-incompatibel tetapi ketika masa berbunga
hampir selesai, bunga-bunga yang mengalami penyerbukan
sendiri menghasilkan buah {Bawa, unpubt.) Pada Piscidia
carthagenensis {Fabaceae, Faboideae), bunga-bunga berusia
satu hari hasil penyerbukan sendiri secara buatan, tidak
menghasilkan buah, tetapi bunga yang berusia dua hari
membentuk buah. Pohon-pohon dari spesies ini biasanya
mengalami penyerbukan siiang (Frankie dan Haber, unpubl.).
Pada Luehea seemanii (Tiliaceae), bunga-bunga yang
mengalami penyerbukan sendiri menghasilkan buah lebih
banyak pada 12-16 jam setelah "anthesis" dibandingkan pada
2-4 jam dari "anthesis" (Haber dan Frankie, 1982).
Meskipun demikian, frekuensi kerusakan sistem
inkompatibilitas dan faktor lingkungan penyebab kerusakan
tersebut rnemerlukan penelitian lebih lanjut (Baker, 1955,
1965a, 1974; Stebbins, 1957; Jain, 1976). "Selfing" yang
melestarikan genotipe; "crossing" menyebabkan
rekombinasinya, sehingga sistem perkawinan sebuah pohon
hutan tropis akan tercermin pada struktur populasi dan
variasinya. Penelitan tentang variasi ini, yanng dibuat
melalui elektroforesis protein yang terkandung dalam sel,
baru saja mulai dilakukan pada pohon-pohon tropis (Can
dkk., 1977).
Sistem self-inkompatibilitas yang heteromorfik untuk

sebagian besarnya terbatas pada tanaman herba dan pohon-
pohon kecil (misalnya: Rubiaceae, Erythroxylaceae,
Oxalidaceae, Boraginaceae) dan bisa tidak ditemukan pada
pohon-pohon tinggi. "Distyly" lebih umum daripada
"tristyly" pada tumbuhan hutan, "tristyly" terbukti hanya
pada bentuk turunan dari Averrhoa carambota dan A. bilimbi
(Oxalidaceae) (Baker, unpubl.). Pada peneiitian-
penelitian, dua ekosistem hutan tanah rendah di Costa
Rica, Bawa (unpubl.) menemukan 2 s/d 3% spesies pohon dari
setiap ekosistem adalah "distylous".
Pada beberapa tanaman kayu tropis "heterostyty"
tampaknya berkembang menjadi "dioecism" (Baker, 1958,
1959; Carlquist, 1966; Bawa dan Opler, 1975; Opler dkk.,
1976a; Bawa, 1980). Jadi, pada Mussaenda spp. (Rubiaceae),
menghambat (mencekik) pembuluh corolla dengan rambut
artinya adalah bahwa pada tanaman ber-"style" panjang,
serbuk sari jarang diambil dari benangsari yang bawah dan
pada tanaman ber-"style” pendek serbuk jarang ditaruh pada
putik. Jadi, tanaman ber"style" panjang berfungsi sebagai
individu "pistillate" (benangsari) dan menghasilkan biji,
sementara (anaman ber"style” pendek berfungsi sebagai
putik (Baker, 1958, 1959). Gambaran yang sama terlihat
pada Cord/a (Opler dkk., 1976a). Sebagai akibat,
pengamatan pembentukan buah dan penelitian morfologi bunga
adalah penting untuk memastikan apakah mereka berfungsi
sebagai hermaprodii (dan heterostyle) atau berfungsi
uniseksual.
Pada spesies dengan self-inkompatibilitas yang
homomorfik, sistem biasanya tampak sebagai tipe yang
dikontrol secara gametophyt, dan sebaliknya pada sistem
in-heteromorfik yang dikontrol secara sporafit. Jadi pada
Theobroma (Sterculiaceae), sistemnya mempunyai ciri yang
tidak biasa yaitu bahwa pembuluh tepung sari dari semua
penyerbukan tumbuh sama cepat dengan kantung embrio dan
perbedaan antara penyerbukan compatibel dan inkompatibel
hanya menjadi nyata ketika garnet jantan yang

terakhir/belakangan gagal melakukan pembuahan sel telur
(Cope, 1962a). Cope (1962b) memperlihatkan bahwa dari
sebuah populasi yang terisolasi (dan Theobroma cacao
memiliki distribusi "spotty" di hutan Amazon) menghasilkan
pohon-pohon self-compatibel, yang berbuah banyak, akhirnya
harus menggantikannya dengan self inkompatibilitas. Namun
demikian, akan terjadi diskriminasi selektif yang kuat
melawan tipe-tipe self compatibel (Purseglove, 1968) dan
di dekat pusat asa! spesies yang diperkirakan berada pada
lereng ttmur Andes, semua klon yang sejauh ini telah
ditefiti terbukti merupakan self incompatible. Di lain
pihak, semakin jauh pengumpulan dibuat dari pusat asal,
semakin besar proporsi pohon-pohon self compatibel.
Purseglove (1969) mempertimbangkan bahwa self
incompatibility akan menolong menyebarkan spesies ke dalam
daerah baru.
Keberadaan self incompatibility pada tumbuhan tidak
menghilangkan secara keseluruhan kemungkinan proses
inbreeding secara genetis, dimana ini tidak tergantung
pada pembuahan sendiri, ukuran populasi kecil yang efektif
disebabkan oleh terbatasnya sejumlah individu dimana polen
mungkin diterima, pernbentukan bunga yang tidak sinkron
dan penyebaran benih terbatas dapat memperbesar tingkat
inbreeding. Aliran gen dalam waktu dan ruang sulit untuk
dimonitor pada populasi ponon tropis, dan studi-studi yang
sesuai masih dalam taraf penyelesaian. Akan tetapi, studi
terbaru mengenai pohon-pohon di hutan hujan Malaya
menunjukkan bahwa varian genetik dan seedling menurun
dengan cepat satu langkah pada suatu pohon induk Shorea
leprosula (Dipterocarpaceae), dan Xerospermum intermedium
(Sapindaceae) (Gan, et al, 1977). Ini sesuai dengan
pendapat Ashton (1969) bahwa pohon-pohon di tropika basah
kebanyakan menukar gen-gen dengan tetangga-tetangga dekat
mereka dalam suatu rumpun, meskipun kadang-kadang dapat
juga terjadi antar anggota-anggota dari rumpun-rumpun yang
berbeda. Untuk pembuktian, setidaknya di hutan kering di

Costa Rica, pohon-pohon dapat berumpun, lihat Hubbel
(1979), ditunjukkan bahwa rumpLin-rumpun dari pohon hutart
dipterocarpaceae Malaysia (yang terbentuk karena
terbatasnya penyebaran benih} menghasilkan lebih banyak
buah dibandingkan pohon-pohon yang terisolasi sebagai
individu-individu.
Outcrossing 3: Monoesisme dan dioecisme
Banyak tumbuhan tropis mendorong outcrossing dengan
monoesisme (contohnya beberapa Meliaceae; Styles, 1972;
Styles dan Khosla, 1976) atau merubah monoesisme harus
dengan dioesisme (contoh, Cariaceae; Baker, 1976). Pada
bunga-bunga uniseksual dari beberapa taksa monoecious
telah berkembang baik, tetapi organ-organ seks lawannya
lidak berfungsi, membuatnya sulit untuk menduga sifat alam
seksualitas dari pemeriksaan kasual morfologi bunga.
Dengan demikian, berdasarkan studi berikut, banyak spesies
dari hutan-hutan tropis yang bersifat hermaprodit pada
bunga-bunga tua, telah ditemukan dapat menghasilkan bunga-
bunga yang berfungsi secara uniseksual (Styles, 1972; Bawa
dan Opler, 1975; Styles dan Khosia, 1976; Bawa, tidak
dipublikasikan). Kesamaan antara bunga-bunga staminate
dengan pistiltat (bertentangan dengan yang ditemukan pada
proses penyerbukan oleh angin pada pohon-pohon hutan
temperate) kemungkinan dibutuhkan kedua macam bunga untuk
rnenyesuaikan diri dalam mencari bentuk yang sama oleh
hewan polinator-polinator (serangga, burung, kelelawar)
dari pohon-pohon tropis ini.
Uniseksualitas dari bunga-bunga tersebut akan lebih
jelas pada spesies berkayu dibandingkan pada taksa
herbaceous (termasuk epifit). Sebuah survei flora di Pulau
Barro, Colorado, Croat (1978) melaporkan proporsi dari
spesies diocious dalam flora (semua bentuk hidup bersarna)
adalah 9%, (115 dari 1265 spesies). Penulis yang sama
(Croat, 1979) telah membuat analisis lebih jauh dan
menunjukkan bahwa diantara pohon yang berukuran sedang

hingga besar, 21% adalah diocious (berumah dua) dan 15%
adaiah monoecious (berumah satu). Angkanya rnengecil
menjadi masing-masing 7% dan 12%, untuk pohon-pohon kecil
(dan semak), 8% dari tumbuhan scandent adalah dioecious
dan 12% adalah monoecious.
Dalam studi mengenai hutan kering di propinsi
Guanacaste, Costa Rica, Bawa dan Opler (1975) menemukan
22% dari spesies pohon adalah diocious. Mereka mengulangi
dan dana diperoleh 40% untuk hutan hujan di Nigeria dan
26% (termasuk beberapa spesies hermaprodit dikogamus)
untuk hutan hujan campuran dipterocarpa dataran rendah di
Serawak Tengah (masing-masing adalah data dari Jones, 1955
dan Ashton, 1969). Jelasnya, diosisme lebih lazim diantara
pohon-pohon daerah-daerah temperate, dan kemungkinan lebih
lazim pada pohon-pohon besar dibandingkan pada bentuk
hidup lainnya. Alasan-alasan ini telah dikemukakan oleh
Bawa dan Opler (1975), Bawa (1981) dan Beach (1981).
Pohon-pohon dioecious tropis menunjukkan variabilitas
yang tidak teratur daiam ekspresi seks, dimana manifestasi
dari satu seks (biasanya staminate) menghasilkan beberapa
bunga yang hermaprodit atau lebih jarang dari seks
lawannya. Selanjutnya dalam Carica papaya, ada
kecenderungan mempakan pohon-pohon staminate, pada akhir
periode pernbentukan bunga, untuk menghasilkan bunga-bunga
hermaprodit dimana buah-buah yang membawa benih terbentuk.
Benih-benih dari buah-buah ini meningkatkan progeni
staminate mapun pistillate, yang artinya datam
mempertahankan keberadaan kedua seks dalam areal lokal
telah diteliti oleh Baker (1976).
Pada beberapa spesies dari pohon-pohon hutan hujan
tropis, individu-individu hermaprodit, sama halnya dengan
pistillate (bunga berkelarnin betina) dan staminate (bunga
berkelamin jantan), dapat terjadi pada dasar reguler
(contohnya, Coccoloba padiform'ts (Polygonaceae), atau
individu-individu pohon dapat beragam dalam keadaan
seksual dari waktu ke waktu. Beberapa dari taksa pohon

didaftar oleh Yampolsky dan Yampolsky (1922) sebagai
polyamodioecious. Di areal Pulau Barro, Colorado (Panama),
Croat (1978) menemukan ada 54 spesies (4% dari flora)
dalam suatu kondisi. Belum jelas arti adaptif dari variasi
semacam itu dalam ekspresi seks.
Pohon-pohon tropis dioecious menunjukkan beberapa
hubungan seks dimorphisme dalam ciri reproduktif seperti
jumlah bunga per inflorescecncec, ukuran bunga, warna dan
bentuk dari petal (kebanyakan khususnya pada Ccarica,
dimana bunga staminatenya adalah gamopetalus sedangkan
bunga pistillatenya adalah polypetalous), dalam sejumiah
nektar yang dihasilkan, dan dalam tingkat herbivory pada
bagian-bagian floral (Bawa dan Opler, 1975; 1978; Baker,
1976). Bawa dan Opler (1975) telah mencatat perbedaan-
perbedaan dalam jumlah nektar yang dikeluarkan oleh bunga-
bunga staminate dan pistillate, tetapi Carica spp,
menunjukkan opposite lengkap, disini bunga-bunga
pistillate tidak menghasitkan nektar sedangkan suplainya
baik pada bunga-bunga staminate (Baker, 1976). Dalam kasus
Carica, dikatakan bahwa ini membuat pengunjung bunga
berbahaya, seperti hummingbird (Trochilidae) dan lebah
Trigonia, jauh dari bunga-bunga pistillate dimana ovari
mudah rusak mengisi bunga. Penyerbukan yang bemasil
dilakukan oleh ngengat yang mengunjungi bunga pistillate
karena kesalahan dalam pencahayaan yang kurang.
Baker (1978a) telah mertunjukkan bahwa mungkin ada
perbedaan-perbedaan nyata antara pohon-pohon pistillate
dan staminate dari spesies yang sama pada rasio sukrosa
terhadap heksosa dalam nektar bunga mereka (contohnya pada
Triplaris americana, Polygonacceae). Arti dan tingkat
perbedaan-perbedaan semacam itu akan terbukti dengan studi
lebih lanjut.
Kebanyakan pohon dioeccious tropis yang telah diteliti
menunjukkan rasio seks yang bias, umumnya berlaku seks
staminate (Opler dan Bawa, 1978). Pada beberapa kasus,
hanya tumbuhan bunga berkefamin betina yang diketahui

(Tomlinson, 1974) dan disini apomiksis harus dicurigai.
Apomiksis
Reproduksi apomiksis, biasanya melibatkan agamospermi
(pembentukan embrio tanpa fusi/peleburan seksual) yang
telah diketahui untuk individu tropis spesies berkayu dan
herbaceous sejak abad ke-19. Reproduksi apomiksis pertama
kali ditemukan oleh Smith (1841), yang mengamati
penyusunan biji pada tumbuhan berbunga betina (bunga
berkelamin betina) yang diisolasi, yaitu spesies yang
sekarang dikenai sebagi Alchornea ilicifolia
(Euphorbiaceae). Daftar spesies dengan satu mekanisme
apomistik atau lebih telah dipublikasikan oleh Nygren
(1954, 1967) dan sekurangnya terdapat 30 genera tropis
(terutama rumput-rumputan) tercantum dalam daftar ini.
Mekanisme yang lazim/umum adalah embriony liar
(adventitious), yang berlawanan dengan apomiksis di
bagian-bagian dunia dimana musim pertumbuhan singkat, yang
bergantung pada diplospori atau apospori pada setiap kasus
yang diikuti oleh diplotid partenogenesis (Baker, tidak
dipubl.). llustrasi mengenai mekanisme embriony liar
(adventitious) pada pohon hutan tropis ditunjukan oleh
Pachira oleaginea (sekarang dikenai sebagai Bombacopsis
glabra (Bombacaceae) oleh Baker (1960)}.
Penjelasan-penjelasan eko-evolusioner untuk perbedaan
yang berhubungan dengan iklim ini akan dipertirnbangkan di
bagian lain, tetapi yang perlu mendapat perhatian adalah
beberapa macam apomiksis dapat "menghentikan"
heterozygositas (juga heterosis) yang telah dihasilkan
dari persilangan di luar pada reproduksi seksual
(amphimistik) tumbuhan induk(moyang). Akibatnya,
demonstrasi oleh Ashton (1977) dan Kaur dkk. (1978)
tentang tingkat kemungkinan apomiksis tinggi pepohonan di
hutan klimaks Malaysia, dimana reproduksi amphimistik
mungkin sulit karena pemisahan fisik dari individu-
individu conspesifik, menyatakan bahwa frekuensi

terjadinya sistem perkembangbiakan ini harus diselidiki
secara teliti di hutan tropis lain.
S. Appanah (dalam Kavanagh, 1979) menemukan bahwa
pepohonan Dipterocarpaceae di Malaysia diserbuki oleh
trips (Thysanoptera). Jumlah serangga ini tidak banyak,
dan akan meningkat pesat pada saat sekumpulan spesies
dipterokarpa berbunga. Namun demikian, spesies pohon
pertama dalam unjtan pembentukan bunga, contohnya Shorea
macmptera, dihadapkan pada kelangkaan trip sehingga
kenyataan tersebut menunjukkan pohon-pohon dari spesies
ini adalah apomistik.
Ekologi penyerbukarn Anthecology)
Awal pembentukan dan perkembangan bunga (antesis)
Permulaan pembentukan bunga melibatkan dua fase
berbeda: "induksi" pada tumbuhan dengan kesiapan untuk
menghasilkan kuncup-kuncup bunga (seringkali dikendalikan
oleh fotoperiode, sepanjang suhunya cocok), dan
"diferensiasi" ujung vegetatif sebelum menjadi kuncup
bunga (Hilman, 1962; Salisbury, 1963}. Antesis
(perkembangan bunga} mungkin langsung terjadi atau
tertunda (dormansi kuncup}.
Pendorong fotoperiode, jika ada, pada tumbuhan tropis
dapat berasal dari perubahan yang sangat kecil pada
proporsi cahaya terhadap gelap pada lamanya hari
(McClelland, 1924; Sunning, 1948; Njoku, 1958; dll)
Tetapi, apapun faktor pemicunya, mungkin "induksi",
"diferensiasi" atau "antesis" kebanyakan tumbuhan tropis
berbunga secara diskontinu, dengan kecenderungan berbunga
pada musim tertentu (Holturn, 1952; Daubenmire, 1972;
Frankie dkk., 1974 Opler dkk. 1976b, 1980; Stiles, 1978;
dan referensi lain dalam Frankie dkk., 1974}. Tumbuhan
minoritas pada habitat-habitat lebih lembab berbunga
secara kontinu, setidaknya pada dasar popuiasi, dan inilah
yang paling sering dijumpai pada komunitas-komunitas seral
pionir pada zona-zona hutan yang lebih basah.

Pecahnya masa dormansi kuncup bunga yang mengarah pada
antesis pada tumbuhan tropis telah menjadi bahan
perdebatan. Kelihatannya, lebih dari satu jenis stimulus
yang terlibat dalam pemecahan masa dormasi kunbup bunga.
Api (Hopkins, 1963), perubahan fotoperiode (juga Hopkins,
1963), penurunan suhu (Kerling, 1941; Holttum, 1952; Went,
1957}, dan pergerakan tekanan air (Aivim, 1960,1964;
Holdsworth, 1961; Daubenmire, 1972; Opler dkk., 1976a,
1980; Opler, 1981), semuanya dilaporkan memecahkan masa
dormansi. Kelihatannya ada juga ritme endogenous yang
terlibat, sehingga beberapa tumbuhan hutan tropis berbunga
lebih dari satu kali setahun (Holttum, 1852; Frankie dkk.,
1974; Opler dkk., 1976a, 198C; Opler, 1981), sedangkan
yang lainnya turnbuh untuk beberapa tahun atau dalam waktu
yang lama tanpa pembentukan bunga (termasuk bambu telah
disinggung sebelumnya). Bambu, dan beberapa pohon, adalah
monokarpik (Foster, 1977). Di Ghana, pohon-pohon dari
ekotipe savanna Ceiba pentandra (Bombacaceae) berbunga
setahun sekali; pohon-pohon besar dari ekotipe hutan tetap
vegetatif untuk beberapa tahun antara tahapan pembentukan
bunga (Baker, 1965b). Gentry (1974b) telah mensurvei tipe-
tipe pembentukan bunga pada Bignoniaceae neotropis.
Sinkroni berikut yang dipicu oleh stimulus lingkungan
yang kuat seperti badai hujan tropis pada akhir musim
kering sangat penting dalam mempertahankan aliran gen
intraspesifik dalam spesies pohon tertentu yang agak umum.
Juga memungkinkan pembentukan bunga oleh spesies yang
berbeda dengan perpindahan temporal/sementara yang
sebaliknya akan menyaingi penyerbuk tertentu yang sama
(Frankie, 1975; Opler dkk. 1976a; Frankie dan Haber, in
pre.). Stiles (1978) memperlihatkan bahwa dalam areal
hutan basah (La Selva, costa Fiba), urutan pembentukan
bunga oleh spesies hummingbird penyerbuk akan berubah dari
tahun ke tahun akibat perbedaan iklim namun kontinuitas
suplai sumberdaya dipertahankan.
Antesis biasanya terjadi pada waktu tertentu siang

atau malam untuk setiap spesies tumbuhan berbunga dan
mungkin memakan waktu 24 jam penuh. Sesuai dengan
kehangatan malam di daerah tropis, proporsi tumbuhan hutan
yang berbunga pada malam hari lebih tinggi dibandingkan
pada daerah temperate, sehingga kelelawar dan ngengat
mempunyai peran penting sebagai penyerbuk. Kehangatan juga
dtrefleksikan dengan membuka/mekarnya beberapa bunga
diurnal khusus seperti Thevetia ovata (Apocynaceae) pada
kegelapan sebelum senja. Tumbuhan malam seperti Inga vera
(Fabaceae, Minosoideae) membuka/memekarkan bunganya pada
tengah sore hari dan tetap mekar sepanjang malam, tumbuhan
tersebut memiliki sejumlah pengunjung mulai dari lebah
hingga kelelawar (Salas, 1974). Bunga-bunga dari beberapa
tumbuhan dioecious menunjukan ciri khusus, karena bunga
jantan (staminate) mekar beberapa jam sebetum bunga betina
(bunga berkelamin betina)(Bawa dan Opler, 1975).
Panjang waktu (lamanya) yang diperlukan bunga untuk
tetap dapat menghasilkan serbuk/tepung sari (polen) atau
lebih penting lagi untuk menerimanya, sangat bervariasi
antar spesies. Anggrek epifit terkenal karena bunganya
tetap segar dalam waktu yang lama; kebalikannya yang
sangat ekstrim ialah spesies Passiflora yang bunganya
hanya dapat bertahan beberapa jam dan penyerbukannya hanya
beberapa menit setelah bunga mekar (Janzen, 1968). Namun
demikian, secara umum benar bahwa bunga tunggal pada pohon
tropis biasanya bertahan kira-kira 1 hari. Pengecualian
untuk aturan ini diperlihatkan oleh beberapa spesies
protandous dan protoginus yang bunganya memiliki androecia
dan gynaecia yang berfungsi pada hari (malam) yang
berbeda. Beberapa spesies memiliki bunga yang terus
menyokong (kontribusi) daya tarik irtflorescense (warna)
walaupun telah "dipakai'Vdiserbuki. Misalnya Lantana
camara (Verbenaceae) dimana bunga-bunga yang sedang
diserbuki berwarna kuning, dan bunga yang telah diserbuki
dan masih bertahan beberapa hari berwarna oranye
kemerahan. Contoh yang lebih jauh (further) adalah

Byrsonima crassifolia (Malpighiaceae) dengan perubahan
warna yang tidak terlalu mencolok antara bunga yang belum
dan sudah diserbuki.
Individu tumbuhan spesies monoecious biasanya
memperlihatkan pemisahan temporal dalam antesis bunga
jantan dan betina. Demikianlah Schmid (1970) menemukan
ciri tersebut pada palma Asterogyne martiana di hutan
basah Costa Rica. Pada Capania guatemalensis
(Sapindaceae), Bawa (1977) menunjukkan bahwa setiap
individu tumbuhan mempunyai dua periode anthesis yang
terpisah, namun dalam waktu singkat, dimana bunga jantan
mekar dan di antara periode tersebut bunga betina mekar
dalam waktu yang lebih tama.
Fenologi Dan Keteraturan Masa Berbunga
Sampai saat ini perilaku periodik tumbuhan di
lingkungan tropik kurang mendapat perhatian (Holttum,
1952; Rees, 1964; McClure, 1966a; Gibbs dan Leston, 1970;
Nevling, 1971; Burger, 1974; Frankie dkk., 1974; Opler
dkk., 1980). Tanggapan mengenai kejadian-kejadian fenologi
yang terkait dengan biologi reproduktif seringkali hanya
sebagai Catalan singkat dalam sebuah tulisan panjang untuk
topik lainnya (contohnya Beard, 1946; Ducke dan Black,
1953). Informasi lain mengenai kejadian-kejadian fenologi
khusus dapat diternukan pada perlakukan-perlakuan
floristik (contohya Alien, 1956; Little dan Wadsworth,
1964; dan yang sangat terkenal pada Croat, 1978). Sumber
informasi fenologi lainnya terdapat pada tulisan yang
mengetengahkan kepentingan ekonomi suatu spesies (yaitu
Broekmans, 1957; Rees, 1964; Lamb, 1966; Purglove, 1968,
1975). Pada beberapa studi, data mengenai periodisitas
dari sejumlah kecil spesies telah dikumpulkan sebagai
usaha untuk merefleksikan kecenderungan fenologi umum pada
tipe-tipe vegetasi tertentu (sebagai contoh J.R.Baker dan
I. Baker, 1936; Hopkins, 1963; Daubenmlre, 1972; berbagai
referensi pada Richards, 1952).

Baru-baru ini beberapa usaha dilakukan untuk dapat
membedakan pola-pola komuniktas secara umum dalam
pembentukan daun, bunga dan buah untuk banyak spesies yang
secara khusus membentuk tipe-tipe hutan tertentu. Beberapa
studi telah dilakukan di Afrika oleh Boaler (1966), pada
lahari hutan deciduous miombo di Tanzania, dan oteh Burger
(1974) pada 4 tipe hutan di Ethiopia. Di Asia, Ng (1977)
mempelajari fenologi pada hutan dipterocarpaceae di
Malaya, dengan studi-studi lain di Malaya oleh Medway
(1972) dan di Sri Langka oleh Koelmeyer (1959). Di daerah
neotropik, studi fenologi diiakukan di hutan lembab semi-
evergreen Panama oleh Croat (1969,1978} dan oleh Foster
(1974). Di Costa Rica, studi fenologi pepohonan telah
diiakukan oleh Janzen (1967) di hutan kering, di hutan
lembab oleh Fournier dan Salas (1966), dan di hutan basah
dan kering oleh Frankie dkk. (1974) dan Opler dkk. (1980)
melakukan studi fenologi semak dan treelet. Nevling (1971)
mempelajari fenologi hutan elfin di Puerto Rico, sedangkan
Jackson (1978) mengamati hutan hujan pegunungan yang lebih
rendah (secara teknik hanya pada subtropis) di Brazil.
Dengan beberapa pengecualian dari penelitian di Sri Langka
oleh Koelmeyer (1959) dan di Malaya oleh Medway (1972),
seperti studi-studi di Costa Rica yang diiakukan Baker,
pola-pola ini diternukan hanya dalam jangka pendek
(biasanya sekitar 2 tahun).
Analisis-analisis fenologi pohon hutan Costa Rica
(Frankie dkk., 1974), fenologi semak dan treelet (Opler
dkk.. 1980) dapat diringkaskan. Pada hutan basah (Finca La
Selva), terdapat dua puncak rnasa berbunga yang tampak
pada spesies kanopi dan tiga puncak di tingkat bawah. Masa
berbunga terjadi di dua musim basah dan pada musim-musim
yang tidak terlalu kering, dengan sedikit kesesuaian
(synchrony) di antara dua lapisan. Semak dari hutan basah
menunjukkan masa berbunga yang lebih sering dan bunganya
secara umum dapat digolongkan sebagai "aseasonal" {tidak
bermusim). Pada hutan kering (Guanacaste), dua puncak masa

berbunga terlihat pada pepohonan; satu periode yang luas
selama musim kering yang panjang dan puncak kedua pada
awal musim hujan. Semak hutan kering, sebaliknya
menunjukkan hanya sekali puncak masa berbunga utama yaitu
pada awal musim hujan, sebelum dinaungi oleh munculnya
daun-daun muda pohon desiduous. Selama musim kering yang
panjang semak cenderung berada pada kondisi dorman,
kemungkinan karena kekeringan lebih berat bagi semak
dibandingkan bagi pohon yang berakar lebih dalam. Hutan
ripari di Guanacaste cenderung menunjukkan perilaku
fenologi semak, dan pepohonan intermediate (menengah)
sesuai dengan ketersediaan suplai air yang cukup.
Di hutan basah, setiap bulannya dalam.setahun
ditemukan sejumlah besar pohon yang berbuah masak,
meskipun puncak masa berbuah pada musim kering kedua
(Agustus-Oktober). Pada hutan kering, puncak produksi buah
masak terjadi pada akhir musim kering yang panjang (dengan
hasil benih akan tersedia pada lantai hutan pada awal
musim hujan). Semak di hutan kering memperlihatkan dengan
jelas pola pembentukan buah bimodal, puncaknya pada
pertengahan musim kering dan musim basah. Hutan ripari di
Guanacaste, yang kelembabannya berada di antara hutan
kering dan basah, umumnya juga menunjukkan pola fenologi
intermediate "menengah".
Di Panama, pola fenologi di Pulau Barro Colorado juga
intermediate, saat ini secara klimatik antara hutan basah
dan kering Costa Rica, dan pengamatan fenologi oleh Croat
(1978) pada bentuk hidup semua Eumbuhan menunjukkan bahwa
herba lantai hutan mencapai puncak masa berbunga pada awal
musim hujan, dan epifit sebagian besar berbunga pada
pertengahan hingga akhir musim kering. Tumbuhan perambat
memiliki pola pembentukan bunga yang lebih menyebar. Liana
rnencapai puncak masa berbunga pada awal musim kering:
puncak masa berbunga untuk pohon yang lebih besar berada
pada musim kering. Puncak masa berbunga pada semak dicapai
pada awal musim hujan. Musim berbuah dari berbagai bentuk

kehidupan ini berjalan mengikuti musim berbunga (herba,
pada pertengahan hingga akhir musim hujan; epifit terutama
pada akhir musim kering; liana juga akhir musim kering;
pohon-pohon menunjukkan dua puncak, pada awal musim basah
dan pertengahannya; semak pada musim basah).
Di hutan Cagar Alam Pasoh Malaysia, pohon-pohon
Dipterocarpaceae menunjukkan suatu urutan masa berbunga
yang tumpang tindih dengan durasi dua sampai tiga minggu
untuk setiap spesies, meskipun pematangan dan jatuhnya
buah terjadi bersamaan (H.T. Chan dalam Kavanagh, 1979).
Rangkaian/urutan masa berbunga berhubungan dengan
pemanfaatan penyerbuk yang sama (trip) oleh berbagai pohon
dipterokarpa.
Berbagai usaha telah dilakukan untuk meneliti aspek-
aspek fenologi dari interaksi-interaksi yang terjadi
antara tumbuhan dan binatang pada tingkat komunitas, pada
paleotropis oleh Putz (1979) dan yang lain, dan pada
neotropis oleh beberapa peneliti seperti Snow (1965),
Janzen (1967), Smythe (1970), Gentry (1974b, 1976),
Heithaus (1974, 1979), Stiles (1975, 1978), dan Frankie
(1975, 1976). Studi-studi terinci mengenai ekologi
hummingbird dalam hubungannya dengan fenlogi tumbuhan
telah dibuat oleh Snow dan Snow (1972). Feinsinger (1976,
1978), Stiles (1978) dan Feinsinger dan Colwell (1978).
Rangkaian tulisan Frankie dan Haber (in prep.)
mengkarakteristikkan interaksi-interaksi antara pengunjung
anthofilus dengan pohon-pohon di hutan kering Costa Rica
(juga semak, liana dan tumbuhan perambat terpilih).
Tulisan ini ingin menunjukkan bahwa kelimpahan musiman
lebah-lebah besar dan ngengat rajawali (hawkmoth)
berkaitan erat dengan pola temporal dan spasial
ketersediaan sumber pembentukan bunga.
Penemuan-penemuan Stiles (1978) didasarkan pada studi
pembentukan bunga oleh 59 spesies dari tumbuhan yang
didatangi hummingbird selama kira-kira 4 tahun di hutan
basah Finca La Selva, Costa Rica. Puncak ketersediaan

bunga terjadi pada musim kering dan awal musim basah,
sedikitnya bunga terjadi pada bulan-bulan terbasah pada
setiap tahun. Meskipim demikian, spesies tumbuhan pangan
yang berbeda secara bersamaan akan memenuhi kebutuhan
burung-burung sepanjang tahun. Yang menarik, variasi curah
hujan dari tahun ke tahun menyebabkan beberapa perubahan
dalam urutan masa berbunga suatu spesies, namun
kontinuitas tetap dipertahankan.
Di masa yang akan datang, kita mengharapkan studi-
studi fenologi dapat berkembang datam jangka waktu yang
lebih panjang, karena dapat memberikan pengetahuan yang
lebih mendalam tentang penyebab bervariasinya pola-pola
dari tahun ke tahun, dan pengaturannya yang mendasar. Hal
ini juga memungkinkan pengertian yang lebih baik mengenai
pola pembentukan bunga dan buah pada tumbuhan yang tidak
berbunga setiap tahun. Pemicu yang membawa kepada antesis
akan dapat teriihat lebih jelas. Studi-studi perbandingan
mengenai pola fenologi pada habitat yang terganggu dan
tidak terganggu penting agar kita dapat mengatur hutan
tropis dengan lebih tepat, demikian pula untuk memahami
interaksi-insteraksi antara berbagai organisme yang
mendiarninya. Interaksi-interaksi kompetitif antara
tumbuhan dalam hubungannya dengan penggunaan polen dan
vektor benihnya (dan penghindaran interaksi kompetitif
dengan pembuatan jarak sementara) akan dipelajari lebih
intensif pada batasan yang diusulkan oleh penelitian
Stiles (1975, 1978). Feinsinger (1978), Feinsinger dan
Colwell (1978) untuk penyerbuk, dan oleh Foster (1974),
Howe dan Primack (1975), Howe (1977) dan Howe dan
Eslabrook (1977) untuk penyebar buah.
Vektor-vektor polen/serbuk sari
(1) Penyerbukan oleh angin. Sebagian besar studi mengenai
penyerbukan di hutan tropis tertuju pada tumbuhan
zoophilous dan binatang yang berhubungan dengannya,
karena sebagian besar tumbuhan hutan tropis diserbuki

binatang. Penyerbukan oleh angin agak jarang terjadi
di hutan yang lebih basah dimana penyebaran individual
conspesifik secara luas, ditambah lagi dengan
penampakan fisik pohon-pohon yang tidak berhubungan
satu sama lain, menjadikan anemofili sebagai suatu
sistem yang tidak efisien (Whitehead, 1969; Janzen,
1975). Pada tingkat/level permukaan tanah, kurangnya
pergerakan udara mungkin merupakan faktor yang
menghalangi penyerbukan oleh angin, lagipula rumput-
rumputan yang ditemukan pada tingkat ini, yang tumbuh
berkala, sebagian besar diserbuki oleh serangga
(Sodestrom dan Calderaon, 1971; Karr, 1976).
Anemofili telah dijelaskan pada beberapa pohon di
hutan deciduous yang dibatasi oleh savana dimana
faktor-faktor yang tidak menguntungkan berada pada
tingkat minimum (Daubenmire, 1972; Bawa dan Opler,
1975). Beberapa spesies pohon Moraceae di hutan semi-
desiduous tropis di Afrika dan Asia juga termasuk
anemofili (D. Leston, kom.prib., 1978; Corner, 1952),
demikian pula beberapa Rhizophoraceae di hutan-hutan
mangrove (Tomlinson dkk., 1978). Penyerbukan angin di
hutan hijau abadi (basah) dataran rendah neotropis
baru-baru ini telah dilaporkan oleh Bawa dan Crisp
(1980) untuk Trophis involucrata (Moracaceae) dan ada
indikasi beberapa spesies lainnya berada pada tingkat
bawah dari hutan basah Amerika Tengah (Bawa, tidak
dipubl.). Beberapa palma termasuk Trinax spp.,
tampaknya diserbuki oleh angin (Uhl dan Moore, 1977),
tetapi apa yang umumnya dipercayai, yaitu bahwa
Arecaceae secara keseluruhan anemophilus, ternyata
tidak benar (Schmid, 1970; Uhl dan Moore, 1977).
Tekanan-tekanan selektif yang berkaitan dengan
evolusi anemofili pada beberapa spesies di daerah
lembab di hutan evergreen tidak jelas. Kompetisi untuk
penyerbuk-penyerbuk dan pembatas energetik pada
produksi bunga dan alat-alat pemikat penyerbuk, dapat

diseleksi untuk penyerbukan oleh angin. Pada lapisan
tingkat bawah tidak seluruhnya kurang angin dan dalam
kenyataannya angin yang tertalu keras mengganggu
efisiensi penyerbukan. Hal ini juga memungkinkan
anemofili berkembang pada famili seperti Moraceae dan
Arecaceae di bawah kondisi lingkungan yang berbeda,
namun berlangsung setelah migrasi ke dalam hutan.
(2) Penyerbukan oleh serangga, Kisaran serangga antofilus
di hutan tropis adalah kumbang, lebah (minimal 2
kategori fungsional yang dapat dibedakan), tawon,
ngengat (dengan sphingids dan "ngengat pengatur'"
(settling moth), kupu-kupu dan berbagai jenis talat.
Namun, hampir seluruh informasi yang berhubungan
dengan penyerbukan oleh kelompok serangga ini
diperoleh dari pengamatan-pengamatan yang dilakukan
agak tergesa-gesa. Survei-survei terhadap sistem
penyerbukan pada tingkat ekosistem kebanyakan
dilakukan sangat khusus di neotropik, meskipun akhir-
akhir ini survei di Malaysia telah dapat
mengimbanginya. Studi-studi intensif tentang taksa
tertentu tidak terlalu banyak dan penelitian jarang
dilakukan.
Sindrom-sindrom karakter yang mengadaptasi bunga-
bunga terhadap berbagai serangga penyerbuk di hutan
tropik dapat dibandingkan dengan yang berada di daerah
sedang (temperate) (cf. Van der Pijt, 1960-61; Baker
dan Hurd, 1968; Faegri dan van der Poijt, 1971,1978).
Namun, beberapa korelasi tropis yang murni lebih
terkenal. Selanjutnya Bawa dan Opler (1975) menyatakan
bahwa bunga-bunga putih kecil dengan suplai nektar
terbatas merupakan ciri umum spesies pohon dioceous
yang sebagian besar diserbuki oleh lebah berlidah
pendek. Pada Cordia (Boraginaceae), Opler dkk. (1976a)
menemukan spesies bunga paling kecil C. inermis yang
memiliki pengunjung bunga sebanyak 300 jenis.

Janzen (1975) mengatakan bahwa proporsi jenis dan
biomasa lebah tropik yang besar adalah makhluk sosial,
termasuk beberapa genera seperti Apis, Trigona dan
Melipona. Namun demikian, mereka yang kurang berperan
secara proporsional sebagai penyerbuk, sering menjadi
pemakan bangkai (scavenger) polen dan nektar yang
ditinggalkan setelah proses penyerbukan bunga
dilakukan, atau mengambilnya dari bunga-bunga dimana
penyerbukan yang efisien membutuhkan pengumpul nektar.
Lebah-lebah sosial ini juga mahir membersihkan polen
di badannya ke dalam pellet, sehingga yang tersisa
dibawa dari bunga ke bunga dalam posisi
penyerbuk/polinator (Janzen, 1975).
Lebah-lebah soliter (yang hidup menyendiri) yang
besar sangat penting sebagai penyerbuk pohon berkayu
di daerah tropis. Studi-studi lebah Cyclocopid di Asia
Tenggara dilaporkan in extenso oleh Van der Pijl
(1954). Lebah jantan euglosine pengumpul wewangian
telah diselidiki oleh C.H. Dodson, R.I. Dressier dkk.
(ringkasan pada Dodson, 1975). Vogel (1968) juga
menjetaskan tentang hal tersebut. Traplining (yaitu
perkunjungan pada sekumpulan tumbuhan) oleh lebah
euglosine betina, pertama kali digambarkan oleh Janzen
(1971), yang juga menyatakan bahwa lebah-lebah
tersebut tampaknya sanggup untuk terbang sejauh 23 km
setiap hari, sehingga penyebaran dari tumbuhan bunga
yang dikunjungi seharusnya bukan merupakan rintangan
yang tak mungkin diatasi pada pembuahan silang.
Sindrom karakter trapliner dan tumbuhan yang
didatanginya digambarkan oleh Janzen (1971,1974b).
Penelitian Frankie dan Colville (1979)
menunjukkan bahwa lebah-lebah yang besar dari jenis
yang berbeda mencari makan pada ketinggian di atas
tanah yang berbeda di hutan kering Costa Rica. Dengan
pengertian ini, pembagian sumber nektar dan polen
dapat dicapai. Peran interaksi teritorial dan

interaksi agresif antar lebah di hutan tropik
digambarkan oleh Dodson dan Frymire (1961), Frankie
dan Baker (1974) dan Frankie (1976). Pengaruh reaksi
insting lebah dalam mendukung penyerbukan silang ini
dengan memindahkan individu akan dijelaskan pada
bagian Selanjutnya. Yang juga berhubungan dengan
pendukung penyerbukan silang adalah pernyataan Gentry
(1978) bahwa spesies serangga penyerbuk bunga banyak
(masal) dapat mencurahkan energi untuk menghasilkan
surplus penyerbuk, dan selanjutnya akan menarik
burung-burung pemakan serangga yang rnembantu
penyerbukan siiang dengan menakuti serangga tersebut
sehingga terbang/pindah ke tumbuhan lain.
"Pembentukan bunga masal" dimana individu
tumbuhan memproduksi sejumlah besar bunga dalam jangka
waktu yang singkat, paling sesuai untuk pohon-pohon
bertajuk dan liana fmisalnya Tabebuia dan Pterocarpus
di Amerika Tengah) yang menunjukkan kumpulan warna
pada binatang yang dapat terbang di atas tajuk pohon.
Dalam hal ini burung-burung dan Iebah masuk termasuk
di dalamnya. Sebaliknya, pohon dan semak di bawah
kanopi, tumbuhan perambat, beberapa liana dan epifit
dapat lebih baik dilayani oleh sindrom "traplining".
Studi komprehensif mengenai penyerbukan serangga
dari beberapa anggota famili tertentu belum sebanyak
penelilian yang dilakukan di daerah temperate.
Tercatat penyelidikan dari Bignoniaceae oleh Gentry
(1974a), dan pada Lecythidaceae oleh Prance (1976) dan
lainnya. Pada kedua penelitian ini, Sebah paling
berpengamh untuk sebagian besar aktivitas
penyerbuk/polinator.
Hymenoptera lain, terutama jenis tawon tropika
temyata lebih banyak berperan dalam penyerbukan
dibandingkan dengan yang disadah sebelumnya (lihat
Faegri dan Van der Piji, 1978 hal. 107-109). Harus
diingat bahwa mutualisme yang ekstrim antara jenis

Ficus dan penyerbuk-penyerbuknya melibatkan tawon dari
Agaonidae (ringkasan pada Ramirez, 1970; Galil dan
Eisikowitch, 1971). Semut yang melimpah di dalarn dan
pada pohon-pohon hutan dapat juga menjadi penyerbuk
siiang yang jarang/langka.
Lepidoptera banyak terdapat di hutan tropik,
meskipun tidak seluruhnya sebagai pencari nektar.
Kupu-kupu besar dan berwama-warni, dari genus Morpho
memakan buah busuk (Young, 1972). Di Asia Tenggara,
Banziger (1971) menggambarkan ngengat penghisap darah.
Perilaku kupu-kupu betina dari genus Heliconius yang
agak aneh, diperlihatkan oleh Gilbert (1972, 1975).
Kupu-kupu ini mengambil nektar dari bunga-bunga
kemudian mengumpulkan butiran polen dari bunga jantan
tumbuhan perambat Anguria dan Gurania (Cucurbitaceae)
lalu memuntahkan nektar ke atasnya, sehingga asam
amino tersebar keluar dalam nektar yang kemudian
dihisap oleh kupu-kupu. Dari hasil penelitian terkini,
De Vries (1979) memiliki bukti-bukti tidak langsung
bahwa kupu-kupu pada genera Paridesa dan Battus dapat
melakukan hal yang sarna seperti di Costa Rica. Kupu-
kupu jenis lain di hutan tropis memiliki kebiasaan
yang lebih konvensional dalarn mengumpulkan nektar,
dan hal ini serupa dengan "ngengat pengendap
settling". Nektar-nektar dari bunga hasil kunjungan
Lepidoptera-lepidoptera ini kaya asam amino, (lihat
bawah) (Baker, 1978a).
Ngengat sphigid (rajawali) biasa terdapat di
hutan-hutan tropis setelah matahari terbenam. Beberapa
bukti menunjukkan pada ketinggian yang lebih rendah,
puncak aktivitas mencari makan mungkin dicapai dalam
beberapa jam di waktu pagi (W.A. Haber, G.W. Frankie
dan P.A Opler, pengamatan pribadi) tapi di hutan yang
berawan (cloud forest) di Costa Rica aktivitasnya
terbesar pada senja dan fajar. Kemungkinan sphingid
mengikuti lajur "traplining", tetapi pernyataan ini

memerlukan bukti (lihat Linhart dan Medenhall, 1977).
Sindrom ciri bunga hawkmoth (ngengat rajawali)
dapat dibandingkan dengan yang dijeiaskan secara luas
di bagian lain untuk tumbuhan-tumbuhan temperate
(misalnya Van der Fiji. 1960-1961; Baker dan Hurd,
1968; Faegri dan Van der Fiji, 1966, 1971, 1978),
sebagai tambahan beberapa bunga berwarna kusam dan
berbau tidak sedap dapat pula menank perhatian ngengat
(lihat bawah). Bunga-bunga hawkmoth tropis, termasuk
beberapa yang memiliki tangkai mahkota (corola) paling
panjang, yang sesuai dengan panjang belalai ngengat.
Tercatat bahwa Xanthopan morgani f. praedicta di hutan
Madagaskar, yang menyerbuki anggrek Angraecum
sesquipedale, memiliki panjang belaiai 25 - 30 cm.
Kumbang dari berbagai jenis merupakan penyerbuk
yang secara proporsionai lebih penting di daerah
tropis dibandingkan di daerah temperate (cf. Ban der
Fiji, 1960-1961), 1969; Gottsberger, 1974; dll.) Di
daerah tropis mungkin sangat spesifik dengan kumbang
yang tertarik dengan bau khas tumbuhan (misalnya
Amorphophallus titanium dan kumbang sphigid besar dari
genus Diamesus yang mendatangi inflorescenses dan
seeing terperangkap; Faegri van der Fiji, 1978 hal
101).
Namun demikian, Thien (1980) yang telah
menyelidiki penyerbukan biologi pohon-pohon
angiosperrnae "primitif, terutama di hutan-hutan Asia
Tenggara dan Pulau-pulau lain di Lautan Pasifik,
menyimpulkan bahwa beberapa Diptera dikategorikan
sebagai penyerbuk pionir. Kemungkinan, lalat-lalat
tebih penting sebagai penyerbuk di hutan tropis
daripada yang selama ini disadari; lalat bukanlah
organisme yang secara estetika menarik perhatian dan
ialat tidak selalu mengunjungi bunga secara khusus,
sehingga diabaikan oleh kebanyakan pengamat. Sebagai
contoh adalah pengenalan yang lambat terhadap peran

lalat-lalat Syrphid dalam penyerbukan palma hutan
basah Asterogyne martiana di Costa Rica (Schmid,
1970).
(3) Penyerbukan oleh Burung. Sebuah literatur berguna
tentang ikhtisar penyerbukan oleh burung disajikan
oleh Faegri dan Van der Fiji (1978, hal. 123 kedua).
Pohon-pohon dan semak-semak di hutan neotropis mungkin
saja diserbuki oleh hummingbird (Trochilidae) atau
oleh berbagai burung passerine (perching)
(Passeriformes), termasuk vireos (Vireonidae),
warblers (Sylviidae), tanager (Thraupidae), finches
(Coerebidae) (Toledo, 1977). Honeycreepers yang
disebutkan terakhir adalah yang paling mungkin
bersaing dengan hummingbirds dalam memperoleh
nektardari bunga-bunga hutan (Colwetl dkk., 1974). Di
Afrika dan Asia, sun bird akrobatidk dan tidak
melayang (Nektahniidae) sangat penting sebagai
penyerbuk (Wolf, 1975; Faegri dan Van der Fiji, 1978;
dli), di Asia mereka dapat dijumpai bersama shite-eyes
(Zosteropidae). Di wilayah Indo-Malaya, pemakan madu
(Neliphagidae) di Hawai Drepanididae memakan
nektaryang berasal dari pohon hutan (Metrosideros,
Sophora, dll). Yang agak berbeda yaitu pada cara
mereka mengambil nektar (dan juga dalam pemanfaatan
serbuk sari sebagai sumber makanan yang kaya akan
protein) adalah brush-longued lorikeets
(Trichoglossidae) Australia.
Pada hutan basah di Costa Rica, studi yang
dilakukan oleh Slud (1960), Linhart (1973) dan Stiles
(1975) menunjukkan bahwa dalam Trochilidae, burung-
burung jantan dari banyak spesies mempertahankan
daerah teritorial tempat makanannya dalam pembukaan
hutan, dan lain-lain, sedangkan "pertapa"yang tidak
memiiiki teritorial berlaku lebih seperti seorang
penangkap (trapliner) di hutan.

Toledo (1977) menjelaskan untuk hutan hujan Vera
Cruz di Meksiko, bahwa hummingbirds cenderung
mendatangi pohon-pohon pada strata yang lebih rendah,
semak-semak dan herba (Heliconia adalah herba
tertinggi yang dapat dipertimbangkan}. Sebaliknya
pohon-pohon yang lebih tinggi dan liana yang menempati
tempat yang lebih tinggi cenderung lebih banyak
didatangi oleh burung Perching. Burung tersebut
menunjukkan ketertarikan yang sangat besar pada nektar
sebagai bahan makanan selama masa berbuah rendah dan
ketersediaan serangga minim. Penuiis yang sama
(Toledo, 1975) melaporkan bahwa variabilitas suplai
nektar sepanjang tahun bagi hummingbirds berhubungan
dengan perilakunya dalam mencari makan dan
berkembangbiak.
Pada areal cloud-forest di Costa Pica, Feinsinger
{1976, 1978) mempelajari pola hummingbirds mencari
makanan {kebanyakan dalam komunitas terganggu) dan
dihubungkan dengan pola penyediaan nektar oleh
tumbuhan. Feinsinger dan Colwell (1978) menghubungkan
perbedaan-perbedaan perilaku dalam mencari makanan di
dalam dan atar spesies pada ketinggian yang berbeda
seiring dengan perubahan suhu, kerapatan udara dan
faktor-faktor lainnya.
Van der Pijl {1937} dan Faegri dan Van der Pijl
(1978) menjelaskan perbedaan-perbedaan dalam
organisasi bunga antara bunga paleotropik burung
perching dan inflorescens di satu sisi, sedangkan pada
sisi lain bunga neotrropik hummingbirds. Terdapat
ketetapan tempat berdiri pada yang pertama, berlawanan
dengan daerah terbuka di sekitar bunga sehingga dapat
ditempati oleh hummingbirds yang sedang terbang.
Perbedaan ini juga dapat dilihat dengan membandingkan
antara spesies yang diserbuki oleh hummingbirds dan
passerine yaitu Etythrina di neotropis {Cruden dan
Toledo, 1977).

Beberapa spesies pohon Ornithopilous mernpunyai
bunga berbentuk rnangkok yang sangat besar (seperti
Spathodea sampanulata, Bignoniaceae, dari hutan
deciduous di Afrika Barat, dengan diameter iebih
kurang 10 cm dan sama-sama dalam). Jenis-jenis inni
memiiiki begitu banyak nektar (dan air hujan) sehingga
dapat digunakan sebagai tempal minum oleh burung-
burung yang lebih besar.
Hadiah yang dapat disediakan untuk burung-burung
yang datang oleh bunga hutan tropis akan dibahas pada
bagian berikutnya. Bunga-bunga tersebut dapat
diproduksi sepanjang tahun, karena memberi makan
burung-burung residen (cf. Stiles, 1978, untuk
hummingbirds), atau untuk setengah tahun, ketika
mereka dimanfaalkan oleh burung-burung migran, yang
mungkin bergerak atas dasar musiman secara lintang
atau ketinggian (Janzen, 1975).
(4). Kelelawar dan Mamalia Lainnya sebagai polinator.
Pernyataan pertama yang muncul mengenai kelelawar
sebagai poiinator di areal hutan tropis telah dibuat
oleh W. Burck, di Taman Botani (Kebun Raya) Buitenzorg
(sekarang Bogor) pada tahun 1892. Mulai dari itu
hingga tahun 1954, relatif sedikit perhatian ditujukan
kepada polinator crepuscular dan nocturnal ini
(kecuali oleh Van der Pijl, 1936, di Asia Tenggara).
Selanjutnya dengan adanya photografi flash elektronik,
dan dewasa ini telah ada alat-alat penguat gambar
penampakan malam bagitu pula alat radio tracking telah
memungkinkan studi yang lebih sering dan mendetait dan
telah memberikan kepada kita apresiasi yang nyata dari
luasnya sistem penyerbukan yang hampir terbatas pada
daerah tropis. Jaeger (1954), Baker dan Harris (1957,
1959), Harris dan Baker {1958, 1959}, Carvalho (1960),
Vogel (1958, 1968-1969), Baker (1970, 1973), Ayensu
(1974), Heithaus ef a/. (1974, 1975}, Sazima dan

Sazima (1977, 1978}, Lack (1978), dan Gould (1977,
1978) adalah diantara mereka yang telah memberikan
contoh-contoh dan bahasan mengenai penyerbukan
kelelawar di hutan tropis. Sebuah resensi umum dan
diskusi disajikan oleh Start dan Marshall (1976), yang
juga meliputi banyak data utarna dari Malaysia.
Bunga yang didatangi kelelawar pada paleotropik
terbatas pada sub order Megachiroptera (sementara
anggota dari sub order Microchiroptera adalah
insectivonous) {Baker dan Harris, 1957; Vigel, 1968-
1969; Baker, 1973}. Namun demikian, Megachiroptera
tidak menyebar ke neotropic dan Microchiroptera (yang
menyebar} menemukan ternpat kosong di hutan tropis
Amerika dan menyebar kedalamnya (Baker, 1973). Sussman
dan Raven (1978) berpendapat bahwa bunga yang
didatangi oleh mamalia non-volant yang didahului oleh
penyerbukan kelelawar, dan cukup besar digantikan oleh
penyerbukan kelelawar ketika mamalia yang terbang
lebih efektif menjadi tersedia. Jika hal seperti ini
adalah kasusnya, sisa keterlibatan mamalia yang lebih
tua ini masih terlihat di Madagaskar, dimana lemur
(kukang) boleh jadi merupakan pollen-vektor, dan di
Australia, dimana marsupial benar-benar terlibat
(Morcombo, 1968, d!l). Faegri dan Van der Pijl (1978,
hal. 122) memberikan contoh lebih lanjut mengenai
penyerbukan oleh mamalia non-volant
Tumbuhan yang diserbuki kelelawar di hutan tropis
biasanya pohon-pohon, liana atau kadang-kadang epifit
dengan bunga-bunga inflorescenses atau individu-
individu bunga yang menggantung pada kekuatan
peduncles atau pedicels sehingga terdapat ruang
terbang bagi kelelawar bebas dari ranting atau cabang.
Contohnya Parkia dappertoniana (Fabaceae, Mimosoideae)
(Baker dan Harris, 1957, Baker, 1978a) dan Mucuna
andreana {fabaceae, Faboideae) (Baker, 1970).
Kemungkinan lain, bunga-bunga tersebut mungkin berada

dalam satu tandan dekat ujung cabang, sehingga
kelelawar dapat hinggap pada inflorescences dan
rnerayap disana, meminum nektar dan memakan pollen,
seperti pada Ceiba petandra (Bombacaceae) (Haris dan
Baker, 1959; Baker, 1963). Susunan ini terlihat dalam
bentuk ganjil pada inflorescences-rachises yang kaku
pada Oroxylum indicum (Bignoniaceae) di Malaysia
(Gould, 1978). Beberapa bunga kelelawar menghasilkan
cauliflorously (seperti Crescentia spp.,
Bignoniaceae}. Banyak pohon-pohon dari bunga yang
diserbuki oleh kelelawar berada dalam kondisi tanpa
daun (yang memungkinkan kelelawar terbang lebih bebas)
dan hal ini mungkin merupakan alasan rnengapa
penyerbukan kelelawar kelihatan lebih sering terjadi
pada hutan kering daripada hutan basah.
Terdapat dua sindrom bunga kelelawar yang cukup
berbeda (Faegri dan Van der Pijl, 1978). Bersama
dengan anthesis nocturnal, warna abu-abu kernerah-
merahan atau keputih-putihan, dan bau yang tajam, agak
tidak enak, yang mungkin terdapat pada bunga-bunga
bermahkota besar, atau pada bunga-bunga tunggal yang
kokoh {contohnya Kigeha, Bignoniaceae) atau bunga tipe
semak (contohnya Adansonia digitata, Bombacaceae) atau
inflorescences (contohnya Parkia clappertonia). Volume
yang sangat besar dari nektar agak mencair pada maiam
hari dan polien juga mencair dalam jumlah yang cukup
besar, merupakan hal yang sangat penting karena
keduanya membentuk suplai terbesar asam amino
pembangun protein untuk kelelawar tersebut (banyak).
Dalam mengumpulkan hadiah-hadiah ini, kelelawar
dapat terbang dalam jarak yang jauh. Start dan
Marshall (1976) menemukan bukti bahwa megachiroptera
Eonyderis speiea mungkin mencari makan pada mangrove
Sonneratia alba (Sonneratiaceae) pada lebih dari 38 km
dari tempat bertenggernya. Migroglossus minimus
mencari makan hanya pada mangrove ini dan terbatas

untuk bertengger sampai 3 km dari sumber makanan.
Gould (1977) memperkirakan bahwa, di Malaysia juga,
Pteropus vampirus, dari suborder kelelawar yang sama,
dapat terbang dalam jarak yang jauh anlara tempat
bertenggernya dengan pohon-pohon Durio zibethinus,
dimana mereka akan makan. Pencarian makan oleh
Microchiroptera neotropic mungkin juga pada jarak 16km
(Janzen, 1975).
Di neotropic, kelelawar microchiroptera kecil
mencari makan satu demi satu, seperti misalnya
Glossophaga soridna dari Markea sp. (Solanaccea)
(dilaporkan di dalam kesalahan sebagai species dari
genus tetangganya Trianaceae dalam Baker (1973), atau
sekawanan, seperti pada Artheus jamaicensis pada
Bauhinia paulitia (Fabaceae, Faboideae; Heithaus, et
al., 1974). Sazima dan sazima (1977) mencatat bahwa di
Brazil bagian Tenggara, Phyllostomus discolor mungkin
saja mencari makanan secara soliter atau mengelompok,
tergantung pada jumlah nektar yang tersedia.
Terdapat bukti-bukti secara tidak langsung bahwa
pencarian makan oleh beberapa kelelawar neotropic
mengikuti pola traplining (Baker, 1973; Heithaus et
al., 1975,, Sazima dan sazima, 1978). Di palcotropic
pemberian makanan "kesempatan" dari kelelawar
megachiroptera pada pohon-pohon "pembentukan bunga
besar" boleh saja merupakan suatu keadaan yang sangat
biasa (seperti pada Ceiba pentandra, Parkia
clappertoniana) (Baker, 1973) tetapi Gould (1978)
telah menunjukkan traplining oleh megachiroptera, sama
halnya di Malaysia (lihat juga, Start dan marshall,
1976), Gould (1978) juga mengklaim bukti mengenai
pertahanan teritorial dari Eonycteris.
Howell (1978) menunjukkan bahwa pohon-pohon yang
menghasilkan bunga kelelawar di Guanacaste (Costa
Rica) menghasilkan aliran nektar pada waktu yang
berbeda-beda sepanjang malam, sehingga kelelawar yang

datang dapat merupakan spesialis untuk waktu tertentu
dan oleh karena itu kemungkinan polinator-polinator
siiang yang lebih efisien, belum umum dalam
mernperoleh makanan dari berbagai sumber. Dia juga
menemukan bukti bahwa kelelawar dapat mengangkut
pollen dari jenis-jenis yang berbeda ini dari tanaman
pada bagian-bagian tubuh yang berbeda sampai menarnbah
efisiensi kelelawar sebagai pollen-vektor. Gould
(1978) menunjukkan bahwa Oroxylum indicum dan Musa
acuminata dapat juga "bekerja sama" dalam memberi
makan kelelawar megachiroptera Eonycteris spelaca di
Malaya.
Kebutuhan untuk bunga atau buah-buah ringan
dimaksudkan agar kelelawar tetap bertahan hidup
sepanjang tahun, di paleotropic diisi oleh prosesi
pembentukan bunga dan buah oleh species yang tidak
bertalian dan hal ini dimungkinkan oleh iklim (Alien,
1939; Van der Fiji, 1969; Baker, 1973; Start dan
Marshall, 1976; Gould, 1978). Di Afrika Barat, Eidolon
helvum mencapai tahap akhir dengan migrasi kelompok
dari daerah berbunga yang satu ke daerah berbunga yang
lain (Alien, 1939). Prilaku sepanjang tahun dari
microchiroptera yang mendatangi bunga dan buah
neotropis telah dipertimbangkan oleh Heithaus et al.
(1975). Dalam hal in, Corollia perspiciilata mungkin
dapat mempertahankan dirinya pada buah-buahan selama
rnusim basah sedangkan Glossophaga soricina dapat
mempertahankan kecenderungan nektarivorous.
Daya Tarik Baqi Polinator
(1) Warna. Macam-macam warna dan pola warna dijumpai
di hutan tropis meskipun suasana dari warna tersebut
mungkin hanya tampak pada waktu-waktu tertentu dalarn
setahun, seperti ketika tajuk Cochlospermum atau
tabebuta berbunga di hutan kering Amerika Tengah, atau
ketika Warszewiczia (Rubiaceae) berbunga di hutan yang

lebih basah. Warna merah adalah warna yang paling umum
diantara species yang diserbuki oleh burung
(hummingbirds dan passerine). Bunga untuk kupu-kupu
berkisar dari putih hingga kuning dan pink (dan bahkan
merah) tetapi bukan biru. Bunga untuk ngengat biasanya
berwarna pucat, atau putih. Bunga untuk lebah
barangkali mempunyai hampir semua warnd, kecuali
rnerah murni {seperti yang terlihat oleh mata manusia)
yang biasanya disertai oleh pancaran ultraviolet(tidak
dapat dilihat oleh kita). Bunga-bunga untuk tawon
sering berwarna merah keunguan.
Meskipun hampir dari semua wama tersebut
berasosiasi dengan jenis polinator dengan cara yang
sama seperti pada daerah temperate, sedikitnya
hubungan yang baru telah terungkap (sama halnya dengan
pada bunga-bunga yang diserbuki kelelawar). Kemudian,
di hutan basah dan kering Costa Rika, beberapa species
yang beradaptasi dengan ngengat memiliki bunga
berwarna pink, merah atau warna lavender daripada
warna putih atau cream yang biasanya rnerupakan bagian
dari sindrom penyerbukan ngengat {Haber, Frankie,
Opier dan Bawa, tidak dipubl.). Contoh-contoh dari
hutan kering meliputi Calliandra spp. dan
Pithecellobium saman {baik Fabaceae maupun
Mimosoideae), Hum crepitans (Euphorbiaceae), Schoepfia
schreberi (Olacaceae) dan Sloanea temifoiia
(Elaeocarpaceae). Di hutan basah, Guarea spp.
(Meiiaceae), Pithecellobium gigantiiolia dan P.
catenatum adalah anggotanya. Di Afrika Barat, Harris
dan Baker (1958) telah menyaksikan dan memotret
sphingid ngengat yang mendatangi bunga-bunga berwarna
ungu tua dari Kigetia africana (Bignoniaceae).
Sebaliknya, beberapa species pohon yang diserbuki
tawon memperlihatkan bunga-bunga berwarna cream
daripada keungu-unguan yang biasa tarnpak, corolla
berstekstur daging, misalnya Casearia sylvestris dan

Xytosma sp. (Flacourtiaceae}, dan Karwinskia calderoni
dan Ziziphus guatemalensis (Rhamnaceae} (Haber,
Frankie, Opier dan Bawa, tidak dipubl. Banyak species
lainnya dengan bunga berwarna putih atau cream
diserbuki oleh kombinasi lebah-lebah kecil dan tawon
(seperti beberapa species Cord/a. Opier et al.,
1976a).
Beberapa penelitian lanjutan mengenai warna bunga
dan polanya yang memperhitungkan pantulan atau
absorpsi ultraviolet yaitu berubahnya pola warna
seiring dengan penuaan bunga, dan hubungan variabel-
variabel ini terhadap pola mencari makanan pada
pengunjung bunga masih tidak umum. Kevan (1978}
membahas proses pewarnaan bunga dengan mengacu pada
anthecology. Barrows (1977) mempelajari perubahan
warna (dari lavender gelap sampai putih) yang terjadi
pada bunga Pachyptera hymenaea {Bignoniaceae) ketika
polen dan nektar tidak lagi tersedia, tetapi bunga-
bunga tersebut masih didatangi oleh lebah selagi
nodanya masih bersisa, sehingga arti biologi dari
perubahan tersebut masih belum jelas. Untuk beberapa
masalah, eksperirnen rnungkin dapat memberikan
jawabnya. Sebuah indikator dari apa yang mungkin
dibutuhkan didemonstrasikan lewat eksperimen Jones dan
Buchrnann {1974) pada Caesalpinia eriostachys dan
Parkinsonia aculeata {Fabaceae, Caesalpiniodeae) pada
tumbuhgn di alam. Mereka menunjukkan bahwa hanya ada
satu petal dalam satu bunga yang mengabsorpsi ultra
violet dan dengan manipulasi posisi petal (dengan
pembedahan dan penempelan kembali), mereka menunjukkan
bahwa pendaratan yang biasa dilakukan oleh lebah
polinator dapat salah arah.
Jones dan Rich (1972) telah memberikan perhatian
kepada kenampakan yang luar biasa dari adaptasi
Columnea florida (Gesneriaceae) pada penyerbukan
hummingbird di Costa Rica, dimana burung-burung

tersebut tertarik oleh bintik-bintik merah pada
belakang daun dimana bunga tersembunyi. Disini
terdapat kesamaan pada pewarnaan merah terang dari
daun dan bagian penguat lainnya yang terdapat di
sekitar bunga pada banyak euphorbia tropis.
Penelitian masa depan tentang pewarnaan bunga
dalam konteks penarik penyerbukan mungkin dibuat pada
basis family, seperti yang dimulai oleh Gentry (1974a)
untuk Bignoniaceae dan oleh Prance dan kelompoknya
(Prance, 1976, dll) untuk Lecythidaceae.
(2) Bau Bunga. Di hutan tropis terdapat asosiasi yang
biasa dari bau-bauan yang enak dengan bunga-bunga yang
diserbuki lebah dan kupu-kupu, dan ketidakadaan bau
yang biasa pada species yang diserbuki burung. Bau
aminoid berkaitan dengan bunga-bunga yang diserbuki
kumbang, dan bunga-bunga yang diserbuki hewan terbang
lainnya (termasuk bunga yang besar sekali dari
Rafflesia) memiliki kecenderungan yang familiar
tertiadap bau yang tidak enak bagi hidung manusia.
Seperti Van der Fiji (1936) pertama menunjukkan, bau
dari bunga yang diserbuki kelelawar (sebagaimana buah
yang didistribusikan kelelawar) biasanya agak tidak
enak, dan bahkan telah dibandingkan dengan bau
kelelawar itu sendiri. Species yang diserbuki oleh
ngengat biasanya mempunyai bau yang enak (begitu pula
relatif temperate mereka) tetapi hal ini tidak dapat
dipercayai karena bau yang tidak enak dari bunga Durio
zibethinus (Bombacaceae) menarik perhatian ngengat
sebagaimana pada kelelawar (Baker, 1970}. Bunga-bunga
yang diserbuki kelelawar, Kigelia africana, di Afrika
Barat, mempunyai bau busuk (dan berwama ungu kemerah-
merahan) tetapi mereka juga menarik perhatian ngengat
sebagaimana kelelawar (Harris dan Baker, 1958).
Kerja dari C.H. Dodson, R.L. Dressier dkk.
(diringkas dalam Dodson, 1975) dan Vogel (1968)

mengenai pengumpulan substansi bau-bauan dari petal
berbagai anggrek tropis oleh lebah auglosine jantan
telah menunjukkan kombinasi karakteristik "minyak-
minyak essensiaf yang secara berbeda menarik perhatian
lebah, sehingga terdapat mating assortive dari
pollinia dan stigma anggrek-anggrek ini.
Overland (1960) telah mempelajari ritme
endogenous yang ada pada pembukaan dan produksi bau
pada bunga dari blooming nocturnal Cesfrum noctumum
(Solanaceae). Ritme endogenous semacam itu, dan
kesempurnaan mereka ke dalam siklus 24 jam oieh
faktor-faktor lingkungan dapat dilihat juga dengan
keuntungan pada species diurnal. Salas (1974),
melaporkan bahwa nektar Inga vera var, spuria
(Fabaceae, Mimosoideae) tidak berbau saat pertama kati
dihasilkan (diakhir senja) dan menarik perhatian
beragam pengunjung bunga. Namun demikian, bunga-bunga
tersebut rnenjadi berbau tidak enak setelah beberapa
jam dan kemudian bunga-bunga tersebut didatangi
kelelawar.
Hadiah Bagi Pengunjung Bunga
(1) Nektar. (a) Bahan Kimia dari nektar. Aspek-aspek
bahan kimia penyerbukan biologi tumbuhan berkayu di
daerah tropis telah disebutkan oleh Baker (1978a),
dengan menekankan pada bahan kimia nektar. Nektar
diketahui lebih dari sekedar air gula: nektar
mengandung beberapa atau semua bahan-bahan kimia :
gula, protein asam amino, lipid, asam organik
antioksidan, beragam substansi nutrisi organik lainnya
dalam jumlah yang sangat kecil, sebagaimana bahan
kimia lainnya yang mungkin mengandung efek pencegah
pada beberapa pemakan nektar yang potensial {lihat
bagian mencegah dibawah} (Baker dan Baker, 1975;
Baker, 1978a).
Energi produksi atau nilai pembangun jaringan

nektar bergantung pada volume nektar dan konsentrasi
bahan kimia di dalamnya dalam pertanyaan (lihat di
bawah). Ada beberapa bukti dari sarnpel yang di arnbil
dari hutan kering Costa Rica, dimana konsentrasi gula
pada nektar meningkat dari dasar hingga puncak pohon
(dengan perkiraan gradien terbalik untuk asam amino)
(Baker, 1978a).
Rasio sukrosa terhadap heksosa dalam nektar
menunjukkan hubungan dengan perilaku polinator,
menjadi tinggi untuk bunga hummingbirds dan bunga
ngengat, tetapi rendah untuk bunga passerine yang
diserbuki burung dan yang diserbuki kelelawar (Baker,
1978a; Baker dan Baker, 1981). Bunga untuk kupu-kupu,
dominasi sukrosa kurang jelas terlihat, dan untuk
bunga lebah cukup beragam. Kadangkala ada phylogenetic
yang kuat; nektar dari Asteraceae biasanya didominasi
heksosa, nektar Ranunculaceae kaya sukrosa, tanpa
melihat tipe polinator (Baker dan baker, 1981).
Konsentrasi asam amino dalam nektar cenderung
lebih besar jika nektar adaiah satu-satunya (atau yang
terbesar) sumber material pembangunan protein untuk
para pengunjung bunga biasa dibandingkan jika
pengunjung mempunyai alternatif yang melimpah (Baker,
1978a). Di Costa Rica, ngengat, kupu-kupu dan tawon
(termasuk pula beberapa kelornpok tawon) bergantung
pada nektar untuk pemeliharaan diri mereka, dan nektar
dari bunga yang mereka kunjungi mempunyai konsentrasi
asam amino yang relatif tinggi. Bunga yang dikunjungi
kelelawar di neotropis membuat penggunaan getah buah
dan pollen sebagai sumber dari bahan-bahan penyusun
protein dan mengkonsumsi beberapa serangga; nektar-
nektar yang mereka konsumsi kurang dalam asam amino.
Burung kolibri betina khususnya pada waktu reproduksi
gemar memakan serangga. Bunga-bunga kemungkinan tidak
dapat menyediakan mereka dengan suplai alternatif yang
nyata dari bahan-bahan penyusun protein dan merekapun

tidak.
Nektar dari tumbunan tropik berkayu dapat terdiri
dari 2-24 asam amino yang dapat dideteksi, sesuai
dengan spesies masing-masing (Baker, 1978a). Terdapat
beberapa asam amino "non protein" yang lebih sering
ada diantara nektar bunga dari pohon-pohon tropik dan
liana (55%) daripada tanaman di temperate (36%). Jika
asam-asam amino ini mempunyai pengaruh toksik yang
dipercaya ada saat mereka terdapat pada benih-benih
(Rehr, dkk., 1973a,b), mereka mungkin berperan sebagai
penangkal terhadap pengunjung bunnga-bunga yang tidak
diharapkan.
Pada tahun 1969, Vogel menitikberatkan bahwa
sejumlah tanaman di Amerika Selatan, yang meliputi
anggota dari famili Malpighiaceae hutan tropik,
menghasilkan minyak yang dikumpulkan oleh lebah
anthophorine tertentu. Ketenjar yang mengeluarkan
minyak ini diserbuki "elaiophors" dan ia menyatakan
bahwa produksi minyak in sebagai alternate untuk
produksi nektar. Lebah-lebah termasuk dan genus
Centris, mengangkut minyak ke sarang mereka dimana
Vogel (1968,1971,1974) percaya bahwa minyak ini
dicampur dengan polen dan digunakan pada pemberian
makan larva.
Namun demikian, pada tahun 1973, Baker dan Baker
(1973, 1975) melaporkan bahwa nektar yang terdiri dari
lipid-cairan yang terdiri dari lipid dalam suspensi
dan juga terdiri dari gula biasa dan asam amino,
seperti bahan-bahan larut air lainnya dan dikatakan
bahwa minyak Vogel "alternatif bagi nektar adalah
benar-benar nektar yang secara luar biasa kaya akan
lipid. Di hutan Costa Rica dataran rendah, nektar yang
terdiri dari lipid ditemukan paling sering pada pohon-
pohon. Di antara pohon-pohon dan liana, lipid nektar
ditemukan terutama sering ditemukan pada
Caesalpinioideae dan Bignoniaceae (Baker, 1978a).

Bahan-bahan lain pada nektar bunga, yang secara
potensial penting bagi nutrisi pengunjung bunga di
dalam hutan, termasuk antioksidan (sebagian besar asam
askorbik, vitamin C), dan ini terutama sering ada pada
nektar yang mengandung lipid di mana mereka rnampu
mencegah berkembangnya ransiditas (rasa anyir) (Baker
dan Baker, 1975}. Sebagian besar bahan kimia yang
disebut, dapat juga mempengaruhi "rasa" dari nektar.
(b) Volume Nektar dan Konsentrasi Gula. Konsentrasi
gula nektar dari bunga-bunga yang beradaptasi pada
klas-klas pengunjung bunga yang berbeda mempunyai
selang yang luas (Fahn, 1949; Meeuse, 1961; Percival,
1965,1974; Baker, 1975, 1977,1978a; Cruden, 1979),
ketika volume nektar pada bunga-bunga dengan selang
adaptasi polinasi yang berbeda melewati beberapa
tingkat jarak (Cruden, dkk., 1981; Opler, 1981). Dua
variabel ini terkait erat karena sejumlah gula yang
diproduksi di dalam nektar sebagai hasil konsentrasi
beberapa volume (yang terakhir biasanya diukur dengan
refractometer dalam "sucrose equivalent"; Baker,
1975,1977,1978a; Cruden, dkk., 1981; Bolten, dkk.,
1979).
Konsentrasi gula dari nektar hutan tropik berada
pada selang 5-80% (sucrose equivalent - sebagai berat
per berat total). Pada serangkaian determinasi pada
hutan kering /pada musirn kering (Baker, 1978a),
menemukan bahwa nektar bunga burung kolibri, ngenngat
sphingid dan kelelawar mempunyai konsentrasi yang
rendah (masing-masing x ~ 21%, 24% dan 17%) daripada
nektar bunga kupu-kupu (x = 29%) dimana, pada akhirnya
kurang pekat dibandingkan bunga-bunga settling moth
dan lebah (masing-masing x = 41 % dan 46 %).
Konsentrasi gula yang rendah pada nektar bunga kolibri
menunjukkan nilai yang tidak semestinya dengan laju
metabolik yang tinggi dari burung-burung kecil ini,

tetapi setidaknya tiga penjelasan yang mungkin telah
dikemukakan. Baker (1975) merumuskan bahwa viskositas
yang rendah penting untuk memungkinkan pengambilan
nektar dengan cepat oleh burung kolibri, sphingid dan
kelelawar yang hanya menghabiskan periode yang singkat
pada bunga (dan, di dalam kasus burung kolibri dan
ngengnat sphinngid nectar seharusnya mengalir pada
saluran yang sempit saat dipindahkan dari bunga).
Sebagai alternatif, disarankan oleh W.A. Calder
(komunikasi pribadi, 1980) bahwa suatu nektar cair
diambil oleh burung dalam volume yang cukup besar
untuk memuaskan kebutuhan energi mereka, dapat
bermanfaat karena ia mengurangi tekanan air saat
burung berada di bawahnya. Penjelasan ini tidak dapat
diaplikasikan pada hutan tropika basah. Dinyatakan
oleh Bolten dan Feinsinger (1978} penetesan dari
nektar cair akan memperkecil kemungkinan lebah dari
'perampokan' bunga yang dibutuhkan burung kolibri
untuk keberhasilan polinasi. Bagaimanapun juga,
konsentrasi gula dalam nektar (dart Trinidad) yang
mereka asumsikan terlalu rendah untuk lebah. Sekalipun
demikian, adalah benar bahwa lebah, tawon, kupu-kupu
dan ngengat lebih suka dan dapat berhadapan dengan
nektar yang pekat, dan bahkan mernuntahkan cairan
untuk mencairkan nektar yang sangat pekat sebelum
mencernanya.
Volume nektar dari bunga hutan tropik biasanya
meningkat seiring dengan biomassa bunga (Opler, 1981),
walaupun terdapat pengecualian-pengecualian terkenat,
contohnya Cochlosperum vitifolium (Cochlorspermaceae)
dan Bixa orellana (Bixaceae) pada hutan kering neo-
tropik mempunyai bunga-bunga dengan diameter sebesar 7
cm, tetapi tidak mernproduksi nektar. Beberapa lebah
kecil, kupu-kupu atau ngengat bunga, seperti pada
beberapa Cordia spp., mernproduksi lebih sedikit
mikroliter nektar pada suatu saat, beberapa perbedaan

yang besar lainnya, yaitu bunga-bunga besar pada pohon
balsa neo-tropik (Ochroma pyramidalis, Bombacaceae)
dapat memproduksi 15 ml datam satu malam dalam kondisi
terbuka. Di Asia bunga staminate dari Musa paradisiaca
(Musaceae) memproduksi beberapa mililiter selama satu
malam (Fahn, 1949), saat inflorescences tunggal dari
Parkia clappertariiana Afrika Barat dapat memproduksi
sebanyak 15 ml (dimana semua mengalir ke tempat
biasanya) dalam satu malam (Baker dan Harris, 1957).
Ochroma, Musa dan Parkia semuanya dipolinasi oleh
kelelawar Malaya: Oroxytum indicum 1,8 ml; Durio
zibethinus 0,36 ml; Musa acuminata 0,63 ml.
Volume nektar yang disediakan oleh bunga
berhubunngan erat denngan ukuran pengunjung bunga
(Opler, 1981) dan ini diperoleh bukan dengan secara
kebetulan, terdapat korelasi yang terbalik antara
konsentrasi gula nektar dan volume nektar yang
tersedia, bagaimanapun juga, gula yang dihasilkan
bahkan pada permukaan dilusi biasanya lebih besar pada
bunga yang menyediakan nektar umurnnya untuk burung-
burung yang relatif besar, kelelawar dan ngengat.
Di Artik, Hocking (1953,1968) menghitung produksi
gula nektar per hektar dari tundra Artik dan menduga
berapa yang dapat mendukung penerbangan serangga dan
diperoleh hasil yang agak menakjubkan, sebagai contoh
bahwa satu catkin dari Salix arctophila (Salicaceae)
dapat tersebar untuk 950 "km nyamuk" setiap hari. Hal
ini akan sangat membantu untuk data kuantitatif hutan
tropik.
(2) Polen. Polen yang dihasilkan oleh kunjungan bunga
digunakan sebagai nutrisi oieh kumbang, lalat, lebah
(untuk mereka sendiri dan untuk anak-anaknya) dan oleh
kelelawar, tetapi kelihatannya sangat jarang, bagi
sebagian besar burung pengunjung bunga (lorikeet dari
Australia adalah pengecualian dalam pencernaan polen

yang disengaja: Churchill dan Cristensen, 1970).
Gilbert (1972, 1975} rnenunjukkan bahwa kupu-kupu
betina dari genus Heliconius menggunakan pollen
melalui pengumpulan sampel bunga-bunga staminate dari
tumbuhan perambat diocious pada genera Anguria dan
Gurania {Cucurbitaceae}. Kemudian dilakukan pemuntahan
nektar di atasnya sehingga asam amino meresap ke dalam
nektar dan kemudian diminum oleh kupu-kupu.
Bagaimanapun juga fenomena ini masih terbatas pada
hutan neo-tropik dimana Helioconius berada (lihat juga
Dunlap-Pianka, dkk, 1971; De Vries, 1979).
Banyak bunga dari tumbuhan hutan digunakan oleh
lebah sebagai sumber nektar dan polen sedangkan polen
hanya sebagai hadiah untuk pengunjung. Di hutan
noetropik, dicontohkan seperti Cochlospermum
vitifolium (Cochbrspermaceae) yang terdapat diantara
pohon-pohon, 8/xa orellana (Bixaceae), Sotanum spp.
(Solanaceae) dan banyak Melastomataceae diantara
semak. Cassia spp. yang tidak bernektar (Fabaceae,
Caesalpiniodeae) diternukan pada bentuk kehidupan dari
herba hingga pohon yang kesemuanya menghasilkan polen
yang berlimpah.
Bunga yang tidak bernektar dari anggota
Annonaceae sangat atraktif untuk kurnbang. Sebaliknya
"keuntungan" dari bunga tidak bernektar seperti
Cassia, Swartzia dan Melastomataceae sangat erat dalam
beradaptasi dengan lebah penngumpul polen yang
dilayani oleh "anther-anther makan", berbeda dengan
"anther-anther polinasi" yang menyimpan polen pada
badan lebah ketika sedang makan. Pada Solatium, polen
lolos dari anther melalui pori-pori ujung dan beberapa
polen "menderu" disebabkan karena lebah menggetarkan
badannya saat kontak dengan adroecium dan mengguncang-
guncang polen yang berbentuk seperti debu tersebut.
Masih sangat sedikit pengetahuan tentang
komposisi bahan kimia polen tanaman tropis, tetapi

dinyatakan oleh Howell {1974) bahwa polen yang diambil
sebagai makanan oleh kelelawar mungkin sangat kaya
akan protein, dan mungkin berhubungan dengan
konsentrasi asam amino yang rendah pada nektar bunga
keleiawar {Baker, 1977,1978a). Beberapa studi yang
menguntungkan telah dilakukan pada kisaran urnum kimia
polen dari tumbuhan hutan tropis dan sebagai tahap
awal telah dibuat baru-baru ini (H.G. Baker dan J.
Baker, tidak dipublikasikan)
(3) Hadiah padatan lainnya. Padatan "tubuh makanan",
menyediakan makanan untuk pengunjung bunga dengan
bagian-bagian mulut pengunyah (mungkin untuk
memindahkan perhatian pengunjung dari androecium dan
gynoecium), telah digarnbarkan untuk sejumlah spesies
tropik. Di Asia Tenggara, Freycinetia arborea
(Pandanaceae) memberi makan burung-burung dengan
dedaunan berdagingnya. Di Hawaii, burung-burung ini
dan tikus yang makan pada inflorescence (sekumpulan
bunga pada (anaman), dapat menjadi polinator juga
sebagai agen perusak (Faegri dan Van der Pijl, 1978).
Pada inflorescence "trapping" dan beberapa Araceae
tropik bahkari pada Amorphophatlus variabilis non
trapping, kumbang polinalor dan serangga lainnya
mengkonsumsi bahan padat yang dihasilkan didasar
tampuk bunga (spathe). Banyak kasus telah diketahui
dari peningkatan anak kumbang pada jaringnan berdaging
dari bunga-bunga yang hidupnya panjang, atau lebih
sering disebut inflorescence (Faegri dan Van der Pijl,
1978}, tetapi terdapat suatu keseimbangan yang tidak
stabil disini yaitu antara keuntungan polinasi dengan
kerusakan jaringan-jaringan.
Ini juga terdapat pada kategori hadiah padatan
yaitu yang pertumbuhannya tidak normal didalam synonim
Ficus, tanpa interaksi siklus hidup biasanya juga ada
dan ini akan rumit apabila antara tanaman dan tawon

agaonid, tidak disempurnakan.
(4) Tipuan dari pengunjung bunga. Pada sebagian besar
tumbuhan, pemberitahuan keberadaan hadiah yang tampak
kepada pengunjung bunga melalui warna, bau, dan bentuk
dari bunga didukung oleh penyediaan hadiah itu
(biasanya nekltar atau pollen). Namun demikian tipuan
telah diketahui dengan baik pada biologi polinasi,
khususnya dalam hubungannya dengan serangga pada
kekuatan diskriminator yang kurang berkembang.
Tipuan parsial benar-benar ada diantara pohon-
pohon tropik pada kasus-kasus tersebut dirnana bunga
staminate dari spesies monoecious atau dioecious
menyediakan hadiah (nektar, polen atau keduanya)
sedang bunga pistilat tidak tersedia. Bunga pistilat
dengan "tidak sengaja" dikunjungni oleh polinator yang
mengantisipasi hadiah yang sama dan mereka peroleh
dari bunga staminate (Baker, 1976). Contoh dari
polinasi "sengaja" disediakan oleh spesies Car/ca
(Caricaceae), diaman bunga staminate menawarkan polen
dan nektar untuk berbagai pengunjung (burung kolibri,
lebah, lalat, kupu-kupu dan ngengat}, tapi bunga
pistilat tidak menyediakan hadiah dan dikunjungi
secara singkat oleh ngengat pada saat hampir setengah
gelap di awal malam. Kasus polinasi "sengaja" lainnya
kemungkinan akan ditemukan dilain tempatdi hutan
tropik.
Kasus-kasus yang jelas ada tipuan total, dimana
tidak disediakan hadiah sama sekali, terlihat bahwa
yang paling baik sangat jarang diantara pohon-pohon
besar, dan hal ini telah dinyatakan (Baker, 1978a)
bahwa untuk pohon besar energi yang digunakan untuk
pembentukan bunga begitu besar sedangkan simapanannya
kecil sehingga tidak bisa mencukupi hadiah untuk
polinator. Ini mungkin akan seimbang jika polinator
tidak tertipu. Secara nyata, kami membutuhkan lebih

banyak informasi tentang biaya energi pada penyediaan
hadiah.
Penangkal-penangkal terhadap Pengunjung Bunga yang
tidak Dikehendaki
(1) Penangkal fisik. Ketika bunga beradaptasi pada
polinator-polinator tertentu, diharapkan akan terdapat
seleksi penampakan yang kemudian menurunkan kemampuan
ketersediaan nektar dan polen pada polinator, atau
terhadap yang tidak efisien sebagai vektor polen. Pada
kasus yang sama, ini dicapai oleh perubahan dalam
morfologi bunga. Selanjutnya bunga pengunjung kupu-
kupu dan ngengat settling hingga burung kolibri
mungkin terhalangi oleh bidang datar yang kurang untuk
tempat berdiri pada tubular bunga dalam keadaan
horisontal. Sebaliknya tabung corolla yang panjang dan
sempit mungkin cocok untuk Lepidoptera tapi tidak bagi
burung kolibri yang paruhnya membutuhkan tabung yang
lebih luas. Semua bunga yang bertabung panjang
melindungi nektarnya dari jilatan lebah berlidah
pendek, lalat, kumbang dan lain-lainnya, dan pengaruh
yang sama dihasilkan dengan mensekresi nektar ke dalam
taji (spur). Serangga-serangga merayap sering menjauh
dari tabung-tabung ini melalui pengaturan rambut-
rambut (yaitu Musaenda spp., Rubiaceae, pada
paleotropik; Baker, 1958).
Pengunjung yang tidak resmi, yang disebut pencuri
nektar, menyerbu dasardari corolla atau spur atau
mengunyah melalui pucuk yang belum terbuka untuk
memperoleh hadiah-hadiah yang kaya nektar atau polen.
Lebah, kumbang dan burung kolibri {juga burung-burung
lainnya) termasuk dalam kategori ini; mereka tidak
berperan dalam polinasi. Kehilangnan hadiah pengunjung
resmi kadang-kadang terjadi melalui kalik penjepit
atau bract tebal {sebagai contoh beberapa
Cucurbitaceae untuk kaliks, dan Acanthaceae untuk
bract).

(2) Penangkal kimia. Bahan kimia nektar hanya pada
awalnya bekerja, tapi mungkin diasumsikan bahwa
alkaloid, fenolik, glikosida dan asam amino non
protein akan tidak menyenangkan atau bersifat toksik
terhadap beberapa pengunjung bunga. Semuanya telah
ditemukan dalam nektar bunga dari tumbuhan hutan
tropis (Baker dan Baker, 1975; Baker, 1977, 1978a).
Suatu perbandingan dari proporsi nektar bunga yang
terdiri dari asam amino non protein, alkaloid dan
bahan fenolik, masing-masing diantara sampel dari
spesies hutan tropik di Costa Rica dan sampel dari
California dan dari tundra Alpine di Gunung Rocky
Colorado menunjukkan bahwa nektar tropik mempunyai
proporsi yang lebih besar dari masing-masing (Baker,
1977,1978a).
Baru-baru ini terdapat banyak pertentangan
mengenai bagaimana spesies hutan tropik menjaga nektar
bunga mereka dari pengambilan semut non polinasi
(Janzen, 1877c; Baker dan Baker, 1978; Feinsinger dan
Swarm, 1978; Shubart dan Anderson, 1978). Kelihatan
bahwa sebagian besar dari beberapa spesies bunga
tertutup dapat "menyembunyikan" nektar dari semut pada
saat frekuensinya lebih sedikit karena mungkin
terdapat pada bahan kimia penolak semut di dalam
nektar atau di dalam jaringan yang berdekatan
(sehingga nektar lebih mudah terkontaminasi oleh
bahan-bahan ini, W. Haber, komunikasi pribadi, 1979;
lihat juga Guerrant dan Fiedler, 1981). Van der Pijl
(1955) yang memberitahukan dan menunjukkan adanya
penolakan semut oleh petal, yang disebut secara khusus
sebagai bahan-bahan berbau. Bagaimanapun juga, pada
beberapa kasus, semut memasuki bunga dan memindahkan
nektar.
Pencegahan dari perusakan jaringan bunga oleh
herbivores metalui keberadaan cluster dari kristal
kalsium oksalat atau pemupukan tanin pada bagian bunga

yang tepat dinyatakan oleh kerja morfologi dan anatomi
dari Uhl dan Moore (1977) untuk beberapa spesies
palma.
(3) Penangkal Biotik. Konsep dari "penjaga semut" yang
makan pada nektar bunga ekstra dan melindungi bunga
dari kehilangan nektar bunga mereka terhadap
pengunjung bunga tidak resmi (yang paling nyata mereka
yang membuat lubang metalui corolla untuk memindahkan
nektar)-terlihat seperti yang dikemukakan oleh Van de
Fiji (1955). Dukungan terhadap kemampuannya dalam
menjaga fekunditas (kesuburan) tumbuhan disediakan
melalui percobaan oleh Keeler (1977) pada Ipomoea
carnea (Convolvulaceae). Oleh Bentley (1977) pada Bixa
oretlana (Bixaceae) dan oleh Schemske (1978) pada
Cosfus woodsonii (Zingiberaceae). Bentley (1977b)
telah meninjau subjek yang lebih luas dari
perlindungan tumbuhan dari berbagai macam herbivora
oleh semut.
Interaksi intra dan interspesifik diantara lebah-
lebah yang mendatangi pohon-pohon pada hutan tropis
rnemperoleh perhatian akhir-akhir ini (Frankie dkk.,
1976; Frankie, 1976). Interaksi ini beragam dari yang
agresif satu lawan satu sampai yang sederhana dijumpai
diantara individu hingga aktivitas massa yang
melibatkan kelompok-kelompok individu (Frankie dan
Baker, 1974),
Penghadapan satu lawan satu, yang sering mengarah
pada satu ke individu yang dipindahkan dari bunga dan
kadang-kadang didorong keluar dari tumbuhan, umumnya
telah diamati diantara lebah-lebah soliter dari
Anthophorinae (khususnya Centris spp.) dan Bombinae
(beberapa anggota dari Euglossinae) dari daerah tropis
Amerika Tengah (Frankie, dipubl.). Spesies tertentu
dari lebah tanpa sengat (secara kelompok) pada hutan
yang sama juga diketahui menunjukkan perilaku intra
dan inter agresif (Johnson dan Hubell, 1974). Pada

kasus ini, penyerangan berhubungan dengan makanan.
Johnson dan Hubell menyatakan bahwa perbedaan
interspesifik pada agresi diantara keberadaan lebah-
lebah tanpa sengat menentukan perbedaan perilaku
pengambilan makanan, sekurang-kurangnya dalam jangka
pendek.
Beberapa serangga anthopilous diketahui mempunyai
beberapa daerah pertahanan yang berdekatan dengan
sumber bunga di hutan tropik, sementara yang disusun
oleh semacam burung Kolibri jantan telh dipelajari
oleh Stiles dan Wolf (1970), Linhart (1973) dan Stiles
(1973). Hal ini dapat mengurangi aliran polen antara
wilayah (memotongnya di kecualikan untuk aktivitas
polinator silang jantan dan pengganggu lainnya ke
dalam wilayah yang ditinggalkan untuk beberapa saat
sebelum diusir dan keberuntungan di wilayah lainnya).
Interaksi-interaksi ini telah diamati pada kumbang
(Frankie, 1987; Rauscher dan Fowler, 1979) dan pada
semut (G.W. Frankie, pengamatan pribadi). Banyak
pengamatan wilayah yang telah dilakukan pada lebah-
lebah soliter (sebagian besar Anthaphoridae) di
Amerika Tengah (Frankie dan Baker, 1974; Frankie,
1976, dan tidak dipubl.; Frankie dkk., 1976) dan di
Amerika Selatan (Dodson dan Frymire, 1961; Dodson,
1962,1975).
Pada suatu studi di wilayah hutan kering Costa
Rica, Frankie (in. prep.) mengamati bahwa sebagian
besar spesies pohon beradaptasi dengan polinasi lebah-
lebah besar (sekitar 35 spesies) mernpLinyai satu atau
lebih wilayah lebah {sebagian besar dart spesies
Centris). Hal ini terlihat berbeda dari wilayah burung
Kolibri pada saat mereka berhubungan terutama dengan
perilaku pasangannya, tetapi mereka tidak hanya menuju
kembali pada polinasi dalam wilayah tersebut, tetapi
kadang pula terhadap polen yang dibawa oleh
perpindahan lebah ke pohon lainnya.

Pengambilan makanan dari kelompok oleh beberapa
lebah individu dari Anthophorinae telah diamati pada
hutan kering Costa Rica (Frankie dan
Baker,1974;Frankie1976). Pengambilan makanan semacam
itu mungkin menyebabkan kekacauan yang besar diantara
lebah penyimpan makanan yang bukan kelompok lain ke
suatu titik dimana mereka meninggalkan kluster-kluster
bunga yang diberikan dan memindahkannya ke pohon lain.
Gentry (1978) menaruh perhatian terhadap bentuk
lain dari interaksi agresif diantara pengunjung bunga
pada suatu species dengan proses pembentukan bunga
yang terjadi secara besar-besaran pada daerah tropis.
Dia mengamati bahwa dengan serangga anthophilous,
burung-burung pemakan serangga juga tertarik pada
sumber-sumber flora yang besar ini. Interaksi yang
terjadi antara predator dengan mangsanya dihasilkan
pada penyebaran terakhir, yang mungkin membawa polen
ke pohon yang lain.
Lebah-lebah tropis tertentu (kebanyakan
Anthophoridae) telah diamati mendatangi beberapa bunga
pada suatu tumbuhan tetapi mengabaikan atau hanya
menyentuh sebentar yang lainnya. Terutama jika ada
tidaknya nektar tidak lagi terlihat mengundang lebah,
dimungkinkan bahwa bau yang ditinggalkan oleh
pengunjung-pengunjung sebelumnya adalah yang menjadi
penyebab. Zona temperate baru bekerja dengan lebah
tukang kayu (carpenter bee) (Xylocopa spp.) pada
Passiflora (dari genus yang cukup baik mewakili hutan
tropik) telah mempelihatkan bahwa lebah betina
rnenandai bunga dengan sekresi dari kelenjar Dufour
(frankie dan Vinson,1977;Vinson et al.,1978). Bunga-
bung yang ditandai dapat dikenali hingga 14 menit.
Penghindaran yang sama dari bunga yang terlihat
ditandai dengan bau-bauan yang telah diamati Xylocopa
gualanensis betina pada Passiflora pulchella di hutan
kering dataran rendah Costa Rica, dan dengan Epicharis

sp. jantan Passiflora adenopoda pada ketinggian sedang
(1500 m) di cloud forest (Frankie, tidak dipubl.).
Di paleotropik, Burkill (1907 ) di India, dan Van
der Pijl (1954, dan kom. Prib.,1975) di Indonesia,
telah mengamati bahwa lebah jantan xylocopid
menghindari bunga-bunga yang baru saja mereka kunjungi
atau oleh individu-individu conspesifik lainnya.
Penghindaran terhadap bunga yang baru didatangi
tidak hanya berakibat pada proses mencari makanan yang
lebih efisien oleh serangga tetapi juga dapat
memberikan sumbangan pada peningkatan polinasi silang.
Hasil ini mungkin dapat dicapai dalam jangka waktu
yang agak lebih lama, dengan mengubah warna bunga
(atau dari "pembimbing nektar" pada bunga) yang
merupakan petunjuk bahwa pengunjung bunga selanjutnya
oleh binatang-binatang anthophilous tidak akan
memperoleh apa-apa (dan bahkan kemungkinan merusak
tumbuhan). Perubahan warna bunga mungkin teriihat,
sebagai contoh, pada bunga Gloriosa superba
(Liliaceae) atau pada beberapa species Hibiscus
(Malvaceae).
PEMBENTUKAN BENIH DAN PENGUGURAN BUAH SENDIRI
Pembentukan bunga, bahkan produksi bunga
hermaprodit, tidak selalu diikuti dengan pematangan
buah dan benih di hutan tropis. Karena itu, pada
Hymenaera caurbarit (Febaceae, Caesalpinioideae},
walaupun pembentukan bunga sering terjadi setiap
tahun, pembentukan buah pada pohon tertentu berlimpah
hanya pada satu tahun dalam jangka waktu lima tahun
(Janzen,1978a). Pada tahun-lahun berselang, tumbuhan
berfungsi terutama sebagai donor atas garnet jantan,
Pohon-pohon "Sub-adult" (sub dewasa) juga berlaku
sebagai penghasil polen, dan produksi benih juga
berkurang pada hutan relatif terhadap pohon-pohon yang
sedang tumbuh di tempat terbuka (Janzen,1978a).

Gejala yang umum adalah penguguran buah yang
berkembang beberapa saat sebetum kematangannya. Hal
ini nyata sekali dimana jumlah yang sangat besar dari
bunga-bunga kecil mulai diproduksi, seperti pada sub
famili Mimosoideae dari Febaceae. Contohnya, Parkia
clappertoniana di Afrika Barat, mempunyai sebanyak
2.000 bunga fertil yang potensial dalam inflorencence
tunggal, tetapi jarang untuk 4 atau 5 buah untuk
terbentuk dari setiap inflorencence (Baker dan
Harris,1957).
Pada kasus Cassia grandis (Fabaceae,
Caesalpinioideae), di Amerika Tengah, kurang dari 1%
bunga memproduksi buah (meskipun pada tahun "benih"),
dan sedikit atau tidak sama sekali dalam tahun-tahun
yang berselang. Benih yang akan dipanen membutuhkan
waktu setahun atau lebih untuk rnatang (Janzen,
1978a).
Lamanya proses buah dan benih-benih menjadi
rnatang di pohon hutan tropis bervariasi dari beberapa
hari hingga setahun, seperti pada Pithecellobium saman
dan Enterolobium cyclocarpum (Fabaceae, Mimosoideae)
(Janzen, 1978a).
Srnythe (1970) telah melakukan suatu studi
tentang buah dan benih yang jatuh selama periode 17
bulan di hutan Borro Colorado (Panama). Buah-buah
dengan benih kecil matang pada sekuen sepanjang tahun,
sehingga burung-burung yang makan disitu tetap
terpenuhi dan persaingan untuk mendapatinya minimum.
Pada saat buah-buah menghasilakan benih dalam jumlah
besar, kemungkinan kerusakan benih oleh binatang
Frugivorous (dan oleh predator benih) meningkat dan
untuk mereka, pembentukan buah yang sinkron didalarn
dan antar species rnemiliki keuntungan "memenuhi
pasarnya", terutama dimana binatang-binatang penimbun
menyebar (scatter hoarding animal), seperti agoutis
(Dasyprocta punctata). Persembunyian benih yang

tertupakan mungkin dapat berkecambah.
Penampakan yang mencolok dari buah-buahan yang
diproduksi oleh pohon-pohonan hutan tropis, biasanya
pematangnya cepat hingga beberapa titik tertentu,
diikuti oleh beberapa periode perkembangan yang
tertunda. Kasus yang menarik diniana kecepatan
diferensial dari pematangan terjadi pada dua spesies
dari genus pohon yang sama dijumpai pada Spondias
(Anacardiaceae) (Croat, 1974J. S. radlkoferi, buahnya
berwarna hijau dan tidak manis, berbeda dengan
beberapa spesies yang ancestral (leluhurnya) yaitu S.
mombin, dimana buahnya berwarna orange dan manis.
Ketika species tersebut dijumpai di Borro Colorado, S.
mombin mematangkan buahnya diawal musim hujan,
sedangkan S. radikoferi membutuhkan waktu lebih lama
untuk mencapai kematangan dan matang pada waktunya (di
akhir musim hujan) saat terjadi kekurangan makanan
pada mamalia yang mencari makan, walaupun memiliki
warna yang tidak menarik dan rasa yang tidak manis,
tetap saja secara aktif dimakan oleh beberapa monyet
dan agoutis.
Rangkaian pematangan buah dalam skala besar
ditunjukkan oleh Snow (1965) dengan 18 spesies Miconia
(Melastomataceae) dari lembah Trinidad.
Interprestasinya adalah bahwa telah terjadi seleksi
alam yang menurunkan kompetisi bagi agen-agen
menyebar, dalam haf ini burung yang memakan buah dan
mengangkut benih secara endozootical. Bagaimanapun,
hal ini juga merupakan hasil yang tidak langsung dari
periode pembentukan bunga pada waktu yang sama.
PENYEBARAN BENIH
Metode Penyebaran Benih
Penyebaran benih merupakan bagian yang penting
dalam proses reproduksi dan menjadi subjek studi
selama manusia masih menaruh perhatian pada pertanian,

holtikultura dan kegiatan-kegiatan kehutanan. Untuk
banyak pohon tropis, banyak ada seleksi karakter yang
mengurangi penyebaran, contohnya, seleksi
(dimungkinkan oleh orang-orang Afrika Barat) pohon-
pohon Ceiba pentandra dengan buah polongnya yang tidak
pecah saat matang, yang merupakan alasan kapok dapat
dipanen dengan mudah (Baker, 1965b).
Saat para naturalis mulai tertarik pada tumbuhan
liar, hingga mereka mengukur penyebaran benih dengan
kemungkinan akan rnengurnpulkan benih jauh dari
tumbuhan induk. Pendapat lain menyatakan bahwa
meskipun penyebaran jarak jauh penting dalam
meningkatkan range spesies, sebagian besar tujuan
penyebaran adalah cukup untuk membawa faenih ke titik
perkecambahan cukup jauh dari tumbuhan induk agar
terhindar dari persaingan dengannya (dan untuk lepas
dari predator benih atau seedling yang mungkin
terpusat pada tumbuhan induknya) adalah tepat, dimana
akan cenderung untuk menjatuhkan benih pada
rnikrohabitat yang sama, atau rnirip dengan tempat
induknya. Tinjauan atas pra-penyebaran benih diberikan
oleh Janzen (1978a).
Akibatnya, kita mengharapkan dapat melihat hasil
seleksi dari mekanisme yang membawa sebagian besar
benih pada jarak yang cukup dari tumbuhan induknya,
dengan kadangkala benih dibawa dalam jarak yang lebih
jauh. Penyebaran tidak selalu seragain pada semua arah
dan areat yang terisi oleh hujan benih seringkali
deisebut sebagai "bayangan benih", balasan yang patut
disayangkan karena kebanyakan "bayangan", contohnya
bayangan cahaya atau bayangan hujan, masing-masing
merujuk pada ketidak-adaan cahaya dan hujan.
Sebagai hasil dari kekuatan untuk bertahan hidup
pada seedling yang sangat dekat dengan tumbuhan
induknya, pohon-pohon conspesifik dihutan tropis
mungkin diharapkan mendekati suatu penyebaran seragam,

dan bukti yang mendukung hal ini diberikan oleh Janzen
(1970) dan oleh Conneil (1971). Dari studi terbaru
penyebaran pohon hutan kering, banyak spesies memiliki
lebih kurang distribusi berumpun {Hubbell, 1970, lihat
juga Ashton, 1969).
Metode aktual penyebaran bibit di hutan tropis
melibatkan angin, air atau penyebaran oleh burung atau
mamalia pemakan buah atau pemakan biji. Buah atau biji
mungkin dibawa oleh vektor binatang langsung dari
pohon, atau buah mungkin jatuh ke permukaan dan
terbelah/terbuka disana, yang memungkinkan benih-benih
tersebut disebarkan secara sekunder oleh hewan yang
berkeliaran. Teristimewa sekali adalah penyebaran yang
dilaporkan terjadi pada spesies hutan Amazon oleh ikan
yang menelan benih dari pohon riparian dan tumbuhan
perambat yang jatuh ke dalam air (Gottsberger, 1978).
Beberapa spesies dari genus Ficus (Moraceae) di Afrika
dapat mengalami predasi hebat oleh hama ligaeid
(Hemiptera, Lygaeidae) disini distribusi terbatas pada
jalan air dimana buah-buahan mungkin jatuh di air yang
mengalir dan didistribusikan ketempatnya berkecambah
jauh dari predator (Slater, 1972). Spesies riparian
lainnya dari famili lain, memperlihatkan distribusi
riverine dalampenyebaran benihnya yang berat tapi
mengambang di air (seperti liana Entada scandens.
Fabaceae, Mimosoideae)(Ridle, 1930).
Banyak literatur mengenai penyebaran benih
termasuk penyebaran pohon-pohon huian tropis,
diringkas pada buku II N. Ridley, Penyebaran Tumbuhan
di Seluruh Dunia, dipubiikasikan pada tahun 1930.
Penekanan tropis dalam buku ini berasal dari
pengalaman penulisnya sebagai Direktur Royal Botanic
Gardens, Singapura. Pada tahun 1972, L.van der Pijl
membuat tinjauan mengenai subjek tersebut, kembali
dengan penekanan tropis didasarkan pada pengalamannya
di Indonesia. Peneliti penyebaran benih tropis lain

adalah E.J.H. Corner, juga berhubungan untuk beberapa
tahun dengan taman botanik di Singapura, yang
pertimbangan-pertimbangan teoritisnya dikristalkan
dalam "Teori Durian" dari evolusi tumbuhan berbunga
(Corner, 1949,dll.).
Akan tetapi baru-baru ini kerja eksperimen
kuantitatif dan pekerjaan yang memperjelas subjek
tersebut sebagian besar dilakukan di neotropis.
Penyebaran biji di hulan tropis oleh burung telah
banyak dipelajari oleh ahli ilmu burung, terutama
mereka yang tertarik dalam burung-burung frugivorous
(contohnya Me Diarmid dkk.,1977; Howe, 1977; Howe dan
Estabrook, 1977; Howe dan Van de Kerckhove, 1979;
Cant, 1979). Studi-studi juga telah dilakukan
rnengenai penyebaran benih oleh kelelawar di
paleotropik (Van der Pijl, 1957) dan neotropis
(Vasquez-Yanes dkk., 1975; Heithaus dkk., 1975; Janzen
dkk., 1976; Fleming dkk., 1977; Janzen, 1978b).
Penyebaran benih oleh manusia non volant kurang
mendapat perhatian (lihat Van der Pijl, 1972).
Dalam semua studi, hanya terdapat sedikit jumlah
dari pengarnatan yang pertama tetapi tidak sama dengan
penyerbukan bunga, yang mungkin sering dipelajari di
semua tahap dalam beberapa jam, buah-buahan hutan
hujan dapat tetap melekat dengan tumbuhan induk untuk
beberapa bulan dan kemudian menghilang tiba-tiba. H.F.
Chan (dalam Kavanagh, 1979) melaporkan bahwa
Dipterocarpaceae Malaysia memperlihatkan pembentukan
bunga yang mengejulkan tetapi bersamaan dengan
jatuhnya buah. Ng dan Loh (1974) mengukur panjang
waktu antara pembentukan bunga dan buah dari 93
spesies pohon di hutan Malaysia. Panjangnya berkisar
dari tiga minggu (Pterocymbium javanicum,
Sterculaceae) sampai sebetas bulan (Disspyros
mainganyi, Ebenaceae). Jadi, akan sulit mengamati
agen-agen penyebar terhadap pekerjaan, sebagai

akibatnya, pengganti untuk pengamatan yang pertama
telah dicari
Tabel 12.1 Klasifikasi sistem penyebaran diaspor dalam
skema oleh Dansereau dan Lems (1957)
Auxochore Cyclochore Pterrochore1
Pogonochore Desmochore1 Sarcochore1 Sporochore2
Sclerochore2
Barochore Ballochore
penanaman diaspor oleh induk tanpa disartikulasi diaspor sangat besar, berisi udara (termasuk tumbleweed diaspor dengan anggota badan seperti sayap atau saccate diaspor dengan anggota badan seperti rambut panjang atau plumose diaspor perekat (duri, burr, oleh kelenjar) diaspor dengan lapisan getah atau lapisan luar berdaging diaspor secara morfologi tidak khusus, sangat ringan diaspor secara morfologi tidak khusus, berat sedang diaspor secara morfologi tidak khusus, sangat berat diaspor dikeluarkan atau dibuangi dari induk
Keterangan : 1) diaspor biasanya buah-buahan; 2) diaspor biasanya benih
Pada 1957, Dansereau dan Lems, meninjau literatur
penyebaran benih dan rnemperkenalkan sistem
klasifikasi mereka berdasarkan morfologi diaspor (unit
penyebaran benih, buah, bagian tumbuhan atau bahkan
keseluruhan tumbuhan) (label 12.1). Meskipun, sebagai
contoh dimilikinya buah berdaging oleh tanaman
(Sarcochore) tidak membuktikan bahwa buah ini
disebarkan oleh burung atau mamalia pemakan buah,
kehadiran banyak tumbuhan yang buahnya berdaging dari
berbagai spesies pada komunitas membuktikan pentingnya
penyebaran endozootik pada komunitas itu, khususnya
jika mengalami kekurangan buah-buahan atau benth
bersayap (pterochore) atau benih-benih ditanam atau
disediakan dengan rambut-rambut panjang
(progonochore). Benih-benih atau buah-buahan yang
mempunyai kait atau alat-alat melekat lai pada bulu
atau penampakan (desmochore) akan juga membuktikan
pentingnya penyebaran binatang (dalam kasus epizootik)
pada komunitas dimana mereka ada.
Metode yang sesuai dalam menampilkan proposi
spesies dengan setiap jenis sistem penyebaran diaspora
analog sebagai "spektrum biologis" sebagaimana
Raunkiaer (1934) menampilkan analis-analisnya mengenai
bentuk-bentuk kehidupan. Spektra penyebaran semacam

ini dibuat untuk tegakan hutan atau untuk lapisan-
lapisan (sinusia) dalam hutan.
Penyebaran diaspore di hutan primer
Dalam literatur lain, pernyataan-pernyataan
kualitatid menyatakan (Richards, 1952, hal. 93-94)
bahwa penyebaran diaspore oleh angin dibatasi pada
beberapa pohon-pohon emergen dan pohon-pohon konopi
atas. Schnell (1970, vol. I, hal. 83-84) menunjukkan
bahwa penyebaran angin dan penyebaran binatang
keduanya terlihat pada pohon-pohon besar di hutan
tropis, sedangkan pohon-pohon yang lebih sedikit
memiliki buah-buah besar/berat (barochore) atau
berdaging (sarcochore), dan jarang yang bersifat
anemochorus, penyebaran angin tidak terjadi pada
lantai hutan. Namun, pengecualian-pengecualian
sementara terjadi pada setiap tingkat, pohon-pohon
besar dari genus Mimusops (Sapotaceae) dan beberapa
anggota lebih besar dari Lecythidaceae memiliki
barochore.
Di Costa Rica, di hutan basah La Selva, 71% dari
pohon-pohon kanopi memiliki sarcochore, sedangkan 18%
memiliki barochore, dan hanya pterochore (10%)
diantara kategori-kategori diaspore lain menunjukkan
lebih dari 1% {Baker, Frankie dan Opier, tidak
dipublikasikan). Pada tingkat semak, persentase
sarcochore naik hingga 93% dan ballochore menjadi
terbentuk modestly (4%). Pada tingkat herba, hanya 73%
dari spesies memiliki sarcochore, penekanannya pada
penyebaran epizootik {daripada endozootik), dengan 18%
desmochore dan 9% ballochore (Baker, Frankie dan
Opier, tidak dipublikasikan}.
Hartshorn (1978), menegaskan angka-angka ini
untuk La Selva, melaporkan bahwa sekitar 49% dari
spesies pohon memiiiki diaspor penyebaran angin,
sedangkan 13% disebarkan oleh keleiawar, 3% oleh

burung dan kelelawar, dan 9% oleh angin. Sisanya 26%
tidak dikategorikan, tetapi agaknya, memiliki buah-
buah berat yang jatuh ke permukaan dan terbelah,
sehingga benihnya secara sekunder disebarkan oieh
binatang.
Dalam hutan tropical kering kanopi lebih mudah
patah oleh angin atau juga pada saat pengguguran daun.
Konsekuensinya, tidak adanya kemantapan dari proporsi
penyebara diaspore oleh angin lebih tinggi. Baker,
Frankie dan Opier (tidak dipubl.) menemukan bahwa di
Guanacaste, Costa Rica disposisi dari pohon dan Palm
adalah sebagai berikut; sarcochores hanya 49 %,
barochore 8 %, pterochore 25 %, pogonochore 5 %,
sclerochore 9%, dan ballochore 9 %. Pada tingkatan
tersebut, persentase sarcochore meningkat menjadi 61 %
dan diaspor yang diperoleh dari penyebaran angin akan
berkurang. Pada herba persenrase sarcochore menurun
lagi menjadi 40 % dan rerata sclerochore 20 % dengan
pterochore, pogonochore dan sporochore (masing-masing
10 %) menunjukkan pentingnya penyebaran oleh angin
lebih efektif membuka areal hutan. Desmochore (juga 10
%} lebih balk dibandingkan dengan strata lainnya yang
lebih tinggi.
Penyebaran diaspore dan hutan sekunder
Richards (1952, hal. 382) menyatakan bahwa "Dalam
suatu hal pengamatan ditemukan bahwa mayoritas
karateristik pohon hutan sekunder muda memiliki benih
atau buah-buah yang sangat beradaptasi dengan
pengangkutan oleh angin atau binatang". Dalam hal ini,
ia memberikan perhatian pada ketidakhadiran dari apa
yang sekarang kita sebut barochore. Contohnya, di
neotropis, Vismia guianensis (Hypericaceae),
disebarkan oleh burung dan kelelawar, sedang burung-
burung menyebarkan benih Dydymopanax morototoni
(Araliaceae), Guazuma ulmifolia (Sterculaceae),

Miconia spp. (Melastomaceae) dan Byrsonima spp.
(Malpighiaceae). Di hutan sekunder Afrika Barat,
Musanga cecropioides (Moraceae) disebarkan oleh burung
dan kelelawar (seperti counterpart neotropisnya
Cecropia), sedangkan Pyenanthus angolensis
(Myristicaceae) dan Macaranga barteri (Euphorbiaceae)
disebarkan oleh burung. Di Asia, Melastoma
malabathricum (Melastomaceae) dan spesies Macaranga,
Mallotus (Euphorbiaceae), Trema (Ulmaceae) dan
Rhodamnia dan Rhodomylitus (Myrlaceae) disebar oleh
burung. Di antara spesies hutan sekunder yang
disebarkan oleh angin adalah Ochroma (Bombacaceae) di
neotropis, Ceiba baik di neotropis niaupun Afrika dan
Alstonia (Apocynaceae) dan Anthocephalus (Rubiaceae)
di Asia.
Di Nigeria, Keay (1957) menemukan spesies pohon
dan liana yang disebar oleh angin umumnya hanya pada
sinusia lebih atas dari hutan sekunder tua, kebanyakan
spesies ini menjadi spesies yang intoleran terhadap
naungan peninggalan (relict) dari tahap serai awal.
Di dua areal hutan Costa Rica, tumbuhan yang
pertama mengalami reproduksi dalam pernbebasan buatan
(meskipun tidak mesti yang paling pertama memiliki
sclerochore dengan beratbenih yang relatif ringan
(Opler dkk., 1977,1980). Hal ini secara cepat berhasil
dalam turnbuhan dengan tipe penyebaran yang berbeda
khususnya sarcochore dan dalam periode tiga tahun,
proporsi sclerochore jatuh hingga mencapai tingkat
awal hutan dewasa, berdekatan meskipun spesies yang
ditampilkan berbeda.
Symington (1933) menjadi pernerhati pertama pada
cadangan (reservoir) benih-benih tumbuhan hutan
sekunder yang hidup pada tanah hutan yang tidak
terganggu di Malaya. Selanjulnya, Budowski (1965,
1970), Guevara dan Gomez Pompa (1972) dan Cheke dkk,
(1979) telah membuat penemuan yang sama masing-masing

di Amerika Tengah, Meksiko dan hutan Thailand.
Budowski (1965, 1970) mencatat bahwa benih-benih dari
pionir cenderung membutuhkan rangsangan cahaya untuk
perkecambahan, sama halnya dengan pencahayaan saat
seedling tumbuh menjadi tumbuhan dewasa. Pada
akhirnya, baik yang berada di puncak pohon pada tahap
serai akhir atau datang untuk menguasai posisi sebagai
pohon-pohon kanopi atau emergen pada hutan dewasa. Di
Afrika, pohon-pohon Musanga cecropioides mulai mati
ketika berumur antara 15 dan 20 tahun dan tidak rnampu
beregenerasi dalam naungannya sendiri (Ross, dalam
Richard, 1952).
Dalam cerah terang, bertolak belakang dengan
pembebasan di hutan basah La Selva, Costa Rica,
Hartshorn (1978) menemukan bahwa mayoritas spesies
yang berkernbang dalam celah dan memiliki benih-benih
yang disebarkan oleh burung atau kelelawar. Herwitz
(1979) telah menemukan, di Taman Nasional Corcovado,
Costa Rica, regenerasi pada celah terang disebabkan
oleh jatuhnya pohon tunggal yang kebanyakan berasal
dari seedling atau sapling yang sudah menjadi
pertumbuhan bawah dan mempertahankan jatuhnya pohon.
Bahan kimia dari cadangan benih
Benih-benih mengandung cadangan makanan (dalam
endossperma atau dalam kotiledon) yang akan mensuplai
energi yang dibutuhkan dan material-material pembangun
untuk embrio saat perkecambahan terjadi dan benih
berubah menjadi seedling. Dalam situasi dan kondisi
intensitas yang kurang terang, sebagaimana yang
seringkali terjadi di hutan tropis, seedling rnungkin
tidak dapat menghasilkan jumlah fotosintat yang cukup
sampai dengan sejumlah yang besar daun teiah
dikeluarkan. Sehingga akan ada pembagian yang sama
dari bahan-bahan makanan daiam benih yang pada
akhirnya berarti akan ada seleksi benih-benih yang

lebih besar dan lebih berat.
Studi-studi di daerah temperate (Salisbury, 1942,
di Inggris, Baker, 1972, di California) telah
menunjukkan bahwa berat individu-individu benih lebih
besar untuk spesies-spesies yang berkecambah; (a),
pada naungan berat, (b). dalam keadaan kompetisi antar
tanaman yang hebat, dan (c). dalam keadaan dimana
kekeringan mungkin berlaku segera setelah
perkecambahan. Namun demikian, pasti ada kompromi
antara persediaan cadangan makanan yang melimpah pada
benih dan jumlah benih yang kurang diharuskan karena
batasanpada ketersediaan substansi-substansi makanan
tergabung didaiarnnya. Tambahan "penjualan (trade-
oil)" dibutuhkan antara ukuran benih dan kemampuan
tersebarnya.
Janzen (1977d) telah menyatakan bahwa variasi
dalam berat antara benih-benih pada tumbuhan yang sama
(dimana rasio dari kecil hingga besar mungkin 100 %,
seperti pada kasus Mucuna andreana, tumbuhan perambat
dari Fabaceae, Faboideae) mungkin menguntungkan dimana
menyebar benih "bayangan" dalam suatu kisaran jarak.
Janzen (1970, dkk) telah lebih dulu menyatakan bahwa
jumlah dan berat benih yang dihasilkan oleh tumbuhan
hutan tropis dapat juga dikendalikan oleh kegiatan-
kegiatan predator benih. Dua alternatif "strategi"
yang telah disusun rnungkin mengatasi predasi. Benih-
benih kecil mungkin dihasilksn pada jumlah yang besar
dimana populasi predator benih tidak cukup untuk
menghancurkan mereka semua. "Strategi" ini akan
efektif jika benih tidak dihasilkan pada interval-
interval reguler (banyak pohon-pohon hutan), atau
dihasilkan hanya pada interval-interval panjang
(contohnya bambu). Kemungkinan lain, tumbuhan dapat
mengeluarkan lebih banyak energi dalam pertahanan
kimia dari benih (oleh asam amino non protein, seperti
pada Mucuna andreana, oleh alkaloid,, cynogenic

glycoside, tanin, dll.}. Pada "strategi" terakhir
mungkin terdapat batasan pada jumlah substansi-
substansi defensif yang dapat dihasilkan dan, untuk
alasan ini, jumlah benih yang dihasilkan mungkin lebih
kecit dibandingkan dengan strategi kejenuhan predator.
Namun demikian, karena adanya sedikit benih, dan
dilindungi secara kimia, maka jumlahnya akan lebih
besar dan mengandung lebih banyak cadangan makanan,
sehingga akan berbentuk seddling awal yang baik dalarn
kehidupan. Meskipun selalu ada kemungkinan bahwa satu
atau lebih predator benih mungkin menyusun alat
detoxity protektan kimia dan kernudian akan dapat
meningkatkan kerusakan benih.
Sifat aktual dari penyadiaan energi dan cadangan
pembangunan jaringan dalam benih tumbuhan hutan tropis
telah rnenjadi subjek dari beberapa analisis. Untuk
hutan basah dan kering di Costa Rica, ada hubungan
antara berat benih dengan cadangan simpanan utama;
untuk benih-benih besar, mungkin pada zat tepungnya,
untuk benih-benih kecil biasanya minyak (yang terakhir
menyediakan surnber energi yang febih padat) (Baker
dkk., in prep.}. Benih-benih kecil dari Piper spp.
(Piperaceae) mengalami penyimpanan zat tepung. Protein
selalu tersedia dan akan sangat jelas terlihat pada
benih-benih dari tumbuhan Fabaceous (Leguminus).
Proporsi spesies dengan benih yang menyimpan
minyak pada sampel-sampel dari tumbuhan hutan Costa
Roca adalah 76 % pads pohon dan palma. 85 % pada semak
dan 91 % pada herba. Kira-kira 35 % dari semua spesies
ini menyimpan zat tepung (dalam banyak kasus sebagai
tambahan dari minyak). Hampir semua benih mengandung
cukup protein untuk memberi hasil positif dalam
pengujian kasartiengan reagen Millon.
Ada juga hubungan antara minyak sebagai cadangan
benih dan phanerocotyly (sebaliknya dikenal sebagai
perkecambahan epigcal), kemungkinan dihasilkan dari

kenyataan bahwa benih yang berkecamfaah pada
phsncrocotylar biasanya kecil dibandingkan dengan
benih dengan germinasi (hypogeal).
PENGADAAN SEEDLING
Pada biologi reproduklif, disadari bahwa
pengadaan seddling perlu jika genotip baru akan
dikenalkan ke dalam suatu struktur hutan (untuk
kenyataan bahwa persistensi atau penyebaran melalui
propagasi vegetatif tidak akan dilakukan).
Ng (1978) telah membahas "strategi-stratsgi
pengadaan" pada pohon-pohon hutan hujan Malaysia.
Benih-benih besar dengan perkecambahan yang cepat,
seringkali epigeal cocok untuk pengadaan seedling
besar dengan taproot yang tebal pada situasi bernaung,
ini paling sering ditemukan. Karakteristik dari
sebagian besar pohon, bertolak belakang dengan susunan
ksrakter dari sebagian kecil "nomad" yang cenderung
ditemukan pada pembebasan dan pada margin hutan. Ini
secara ekslusif epigeal. Terbentuknya diameter akar
yang membuat mereka membentuk seedling pada permukaan
yang lebih keras dibandingkan pemanfaatan oleh spesies
hutan yang memiliki benih yang besar.
Untuk hutan basah La Selva, Costa Rica, Hartshorn
(1978) menduga bahwa rata-rata waktu untuk pergantian
pohon kira-kira 118 tahun. Untuk menyediakan
pergantian ini harus ada celah gelap pada interval-
interval yang cukup karena Hartshorn menemukan bahwa
75 dari 104 spesies pohon kanopi yang diteliti
tergantung pada celah terang untuk keberhasilan
regenerasinya. Hartshorn (dalam Whitmore, 1978)
rnelaporkan bahwa proporsi spesies yang tergantung
pada celah untuk regenerasi menurun seperti
perpindahan dari emergen dan kanopi lebih atas melalui
pohon-pohon tingkat bawah.
Di daerah hutan kering Guanacaste, dinegara yang

sama, Hatheway dan Baker (1970) mengamati bahwa pohon-
pohon besar Pithecellobium saman dan Enterolobium
cydocarpum (Fabaceae, Mimosoideae) terlihat tidak
menjadi seedling sampai sapling dalam naungan hutan,
tetapi ini berlaku dalam situasi pencahayaan penuh
(yang juga dilindungi dari penggembalaan).
Whitmore (1975, 1978) melaporkan bahws di hutan-
hutan Asia Tenggara, celah kecil di hutan dewasa diisi
oleh sapling yang ada atau pohon-pohon "tertekan",
sedang dalam celah besar ada flora suksesi dari benih
yang dibaws ke dalam atau sudah ada dalam tanah.
Dormansi yang ada dari benih-benih pohon terlihat
jarang di pohon-pohon hutan basah di Afrika Barat.
Benih-benih yang ada tidak berkecambah daiam tanah di
lantai hutan yang hanya membutuhkan stimulus cahaya
untuk berkecambah (Longman, 1978). Ini adalah
pengalaman dari Frankie, Baker dan Opler (tidak
dipublikasikan} di hutan basah di Costa Rica. Dua
paper oleh Ng (1973, 1975} membcrilan data mcngenai
laju germinasi benih-benih segar dari hampi 200
spesies dsri pohon-pohon yang lumbuh di hutan-hutan
Malaysia. Germinasi "cepat" (dalam 12 minggu dari
sceciiing), "intermediet" dan "tertunda" secara kasar
dalam proporsi 10:3:1 yang selama ini dipercaya bahwa
perkecambahan merupakan hal yang biasa ditemukan di
hutan-hutan Malaysia, tetapi memberikan dugaan yang
tersebar luas bahwa germinasi tertunda jarang atau
tidak mungkin (No, 1&73, hoi. 54).
Percoban-percobaan dengan benih-benih dari pohon-
pohon yang tumbuh di hutan kering Costa Rica
menunjukkan bahwa spesies dari pohon yeng mematangkan
benih-benih mereka di musim basah telah menyusun
mekanisme dormansi yang mencegah germinasi pada
sekurangnya beberapa benih di musim basah yang sama
(Frankie dkk., 1974). Dalam kasus Casearia aculata,
Eugema salamensis var. hiracifolia dan Spondias

mombin, waktu yang dibuluhkEn untuk 50 % dari benih
berkecambah adalah kira-kira 150 hari. Dormansi
semacam itu cenderung untuk menjamin bahwa seedling
akan muncul pada waktu-waktu itu yang akan
memungkinkan perkembangan akar cukup sebelum permulaan
dari musim kering pendek (Juli hingga Agustus) dan
musim kering yang menggantikannya (akhir November
hingga Mei).
RINGKASAN
Perlakuan dari biologi reproduktif tumbuhan di
hutan tropis ini bukan pendalaman dari informasi yang
dipublikasikan, meskipun diharapkan bahwa suatu cara
ke dalarn belukar literatur teiah tersedia. Akan jelas
bahwa lebih banyak kerja tetap akan selesai sebelum
hubungan-hubungan individu dari tumbuhan dengan
penyebar dari polen atau benihnya dapat digabungkan
pada apa yang benar-benar disebut studi ekosistem.
Namun demikian, kita dapat melihat bahwa, meskipun
prinsip-perinsip biologi dasar yang diterapkan pada
hutan temperate dan tropis serupa, ada perbedaan-
perbedaan nyata dalam detail dan penekanan bahwa hasil
dari pernbedaan musiman yang kurang dari iklim di
hutan-hutan tropis, dan dari kerapatan tumbuhan dan
binatang yang lebih besar (dan kelimpahan lebih besar
dari beberapa yang terakhir dihasilkan dalam
penyempitan niche dan peningkatan interaksi-inleraksi
biotik). Perbaikan-perbaikan dalam pengertian kita
dari biologi reproduktif tumbuhan hutan tropis akan
terjadi melalui aplikasi teknik-teknik kuantitatif
modern dari penyelidikan dan manipulasi mcncukupi dari
konstituen ekosistem (sebagai contoh percobaan dalam
tambahan pada pengamatan). Pengetahuan yang didapat
mungkin penting dalam mengembangkan rencana-rencana
manajerial untuk jumlah hutan tropis yang relatif
kecil yang sepertinya mungkin untuk mempertahankan

pengaruh yang terus meningkat dari ekspansi populasi
manusia.





























