Embed Size (px)
Citation preview

Behavioural Brain Research, 36 (1990) 41-51 41 Elsevier
BBR 00997
Behavioural effects of neocortical and cingulate lesions in the Mongolian gerbil
Colin G. Ellard, Dwight J. Stewart*, Steven Donaghy** and C.H. Vanderwolf* Department of Psychology, Mount Allison University, Sackville, New Brunswick (Canada)
(Received 22 February 1989) (Revised version received 22 May 1989)
(Accepted 23 June 1989)
Key words: Frontal cortex; Cingulate cortex; Ventral marking; Social behaviour; Foot-stomping; Type I behaviour; Type II behaviour; Gerbil
The experiments examine the effects of cortical lesions on a variety of behaviours in the Mongolian gerbil (Meriones unguiculatus). Gerbils with either large anterior or posterior lesions were compared with normal gerbils by administering a battery of tests of rodent behaviours such as grooming, eating, social interaction, ventral marking and foot-stomping. In a second experiment, a more detailed investigation was carried out of the effects of focal cortical lesions on ventral marking. The results of these experiments suggest that anterior cortical lesions in gerbils give rise to a number of different deficits in behaviour. The results further suggest that cingulate cortex is part of the neural substrate for ventral marking behaviour. The implications of these results for contemporary theories of frontal lobe function are discussed.
INTRODUCTION
Research on the effects of lesions on natural classes of behaviour such as grooming, loco- motion, manipulation and social behaviour has been used to good effect to investigate the function of the cerebral cortex of rodents (see ref. 28 for review). Lesion studies 24 have sug- gested that large cortical lesions in the rat may affect the control of behaviours defined as Type 122 on the basis of electrophysiological criteria. Such behaviours, which include loco- motion, postural changes and object manipu- lation, are invariably accompanied by neocortical
low voltage fast activity and by hippocampal rhythmical slow activity. These electrical patterns appear to be produced by joint cholinergic and serotonergic inputs to the cerebral cortex 18,23. During Type II behaviours, including grooming, chewing, licking, scratching, and immobility, neocortical low voltage fast activity and hippo- campal rhythmical slow activity may be produced by the cholinergic inputs acting alone or may be absent altogether.
After complete decortication, rats are able to perform most movement patterns quite normally but often fail to make the movements in environ- mentally appropriate contexts and in the normal
* Present address: Department of Psychology, The University of Western Ontario, London, Ontario, Canada, N6A 5C2. ** Present address: Department of Psychology, University of Windsor, Windsor, Ont., Canada. Correspondence: C.G. Ellard, Department of Psychology, Mount Allison University, Sackville, New Brunswick, Canada E0A 3C0,
0166-4328/90/$03.50 © 1990 Elsevier Science Publishers B.V. (Biomedical Division)

42
sequence z4. More recent studies have shown that the extent to which certain of these deficits appear may be dependent upon environmental condi- tions z9.
The purpose of the present experiments was to determine whether the effects of large cortical lesions in the gerbil were similar to those that have been observed in the rat. We were also able to test two behaviours that are not present in the rat. Ventral marking 1'19 consists of a distinctive set of movements that gerbils use to deposit scent on environmental objects. Foot-stomping consists of a vigorous rhythmic tapping with the hindlimbs that is carried out when gerbils are alarmed or are engaging in sexual activity. Ventral marking and foot-stomping provide an interesting test of the relative dependence of the control of Type I and Type II behaviours on the cerebral cortex. Ventral marking occurs during the course of locomotion and is always associated with hippocampal rhyth- mical slow activity, thus defining it as a Type I behaviour. Foot-stomping, on the other hand, is a Type II behaviour because hippocampal rhyth- mical slow activity is usually absent when the behaviour occurs in an immobile gerbil 1z'25.
EXPERIMENTI
Method
Subjects Twenty-five male Mongolian gerbils (Meriones
unguiculatus) of approximately 90 days of age were used as subjects. The gerbils were maintained on a 12:12 light-dark schedule and were housed individually in Plexiglas cages. The gerbils had free access to Purina Lab Chow and water.
Surgery Subjects were anesthet ized with sodium pen-
tobarbital (60 mg/kg) and placed in a stereotaxic apparatus. For the anterior cortical lesions, the skull was exposed and a small hole was drilled just anterior to bregma and just lateral to the midline. The hole was enlarged with bone ron- geurs until a region of bone had been removed that extended 3 mm anterior to bregma, 2 mm
posterior to bregma, laterally to the temporal bone ridge and medially to within 1 mm of the midline. The dura was then incised and the exposed cortex was removed by aspiration. An attempt was made to remove all of areas SI and MI as illustrated by Shook 17. For the posterior lesions, bone was re- moved from a point just anterior to the lambdoidal suture to 3 mm anterior to lambda and from the midline to the temporal bone ridge. Again, the dura was incised and the exposed cor- tex removed by aspiration.
Following all cortical lesions, the damaged area was covered with a small piece of Gelfoam to control bleeding and the area was flushed with isotonic saline. The wound was then suture- closed, and the gerbils were allowed two weeks for recovery before testing.
Procedure Ventral marking. The apparatus consisted of
a square transparent colony cage (36x 30 × 18 cm), on the floor of which were fixed 6 reotan- gular foam rubber pegs (4 x 3 x 1 cm). Under dim illumination, the gerbil was placed in thecage for a period of 5 min. In such an apparatus, the ventral marking episodes of normal gerbils con- sist of an approach phase, which normally in- cludes sniffing the peg, and a marking phase, in which the gerbil walks over the peg with its legs spread slightly and lowers its abdomen so that its ventral sebaceous gland drags over the top of the peg. This entire sequence ofm0vements was clas- sified as a single episode of ventral marking, and the number of times that this sequence occurred in the time period was recorded. Each gerbil received 3 trials, one every 24 h. Between trials, the apparatus was thoroughly cleaned with 100~o ethanol in order to remove the sebum from the apparatus.
Social interaction. A normal female was placed in a dimly lit colony cage Idled vAth sawdust. After a period of 10 ~ , either a normal mate o r a lesioned male was placed in the cage with ~the female. The observation p ~ o d was divided into 6 consecutive 5-min periods. After 30 min, the male was removed and, after a rest period for the female of 15 min, a new male was placed in the cage with the female. If the first male had been

43
normal, the second male was in the lesion group, and vice versa. Each gerbil was presented to the same female on two occasions, with the order of presentation reversed on the second occasion. In each observation period, the number of times that one member of the pair approached and made contact with the other member of the pair was recorded.
Foot-stomping. Gerbils were placed in a Plexi- glas chamber that contained a shock grid on the floor. Gerbils received up to five 2-s shocks separated by 1 min recovery and observation periods. Following the shocks, the incidence and duration of foot-stomping was recorded. Once foot-stomping had occurred, no further trials were carried out.
Seed eating. In the first phase of the seed-eating tests, gerbils were presented with sunflower seeds in the shells, one at a time in their home cages. The time required to shell and fully consume a seed was recorded and a count was made of the number of times that the seed was dropped. This procedure was carried out 10 times for each gerbil.
In the second phase of the seed-eating tests, each gerbil was deprived of food for 24 h and then placed inside a Plexiglas enclosure that rested upon a glass platform. The underside of the gerbil was videotaped by means of a 45 degree mirror in order to observe the manipulatory forepaw move- ments that occur during seed shelling and eating. Seeds were scattered over the floor of the enclo- sure, and the gerbil was videotaped as it picked up, shelled, and ate the seeds. Manipulatory movements were classified as either flips, in which the seed was rotated 180 degrees around its short axis (from end to end), or spins, in which the seed was rotated around its long axis. The number of flips and spins per seed were recorded. An attempt was made to gather data from the con- sumption of 3 seeds per gerbil both prior to and subsequent to surgery. This was not always possible because of the reticence of some gerbils to feed in the apparatus and because of the extreme difficulty that they had in picking up seeds after surgery.
Grooming. Gerbils were immersed briefly in water at room temperature (19-21 ° C) and were
placed in their home cages under warm lights. Gerbils were videotaped from two camera lo- cations for a period of 20 min. The latency to the first grooming movements was subsequently de- termined from the videotapes, and the incidence of the following movements was recorded: face- washing, body shaking, licking of fore- or hind- limbs, licking of flank or anogenital region, and rolling. The duration and number of elements in a grooming sequence was also recorded, with a sequence being defined as a series of movements uninterrupted by locomotion.
Histology At the conclusion of testing, all gerbils were
deeply anesthetized with sodium pentobarbital and perfused through the heart with isotonic sa- line and 4~o buffered formalin. Their brains were removed and fixed for at least 24 h in formalin. They were frozen and sectioned at 40 #m on a sliding microtome. Every fifth section was mounted on slides coated with chrom-alum and stained with gallocyanin.
Results
Anatomical results Fig. 1 shows dorsal view reconstructions of
representative anterior and posterior cortical lesions. Comparison of our lesions with Shook's ~7 cytoarchitectonic map of gerbil cortex suggests that the anterior lesions destroyed large areas of primary somatosensory and motor cor- tex, parts of prefrontal cortex, cingulate cortex, parietal cortex, and, in at least one case, the rostral part of the cortical visual areas.
Posterior lesions destroyed most of primary visual cortex (with fairly consistent sparing of the caudal pole of occipital cortex) and, in a few cases, small regions of temporal cortex as well.
Examination of retrograde degeneration in the thalamus generally substantiated these estimates of the regions of cortex that received damage. Gerbils with anterior cortical lesions consistently showed cell loss and gliosis in the nucleus ventralis medialis, but in two of these cases, the damage was unilateral. In gerbils with anterior lesions, cell loss was evident in the posterior part

44
A
B
Fig. 1. Representative cases of lesions of sensorimotor cor- tex and visual cortex in Expt. I. A: sensorimotor cortical lesions. This lesion probably includes a large portion of pri- mary motor and somatosensory cortex. Some damage is also likely to have been inflicted on parietal and prefrontal corti- cal areas. B: visual cortical lesions. This lesion includes most of primary visual cortex (excluding the posterior pole) and large areas of secondary visual areas (areas 18a and 18b).
of the nucleus ventralis lateralis. In two of these cases, this cell loss was quite diffuse, and in one case, it was unilateral. In one case, there was unilateral gliosis in the dorsal part of the lateral geniculate nucleus.
Retrograde thalamic degeneration in the gerbils with posterior lesions appeared to be limited to visual areas. Thus, in all cases, heavy gtiosis was evident in the lateral genicuiate nucleus, and in most cases, diffuse cell loss had occurred in the lateral posterior nucleus.
Direct surgical damage to subcortical areas was rare in both groups. In one gerbil with an anterior lesion there was unilateral damage to the dorsal fornix, and in another the dorsal part of the caudate nucleus was damaged unilaterally. In all gerbils with posterior lesions, the hippocampus and superior colliculus were intact.
Behavioural results Ventral marking. On initial placement in the
apparatus, normal gerbils and gerbils with visual
cortical lesions moved rapidly around the arena, rearing at the sides and sniffing the pegs. Marking usually began after about 1 min. An episode of marking often began with sniffing at a peg. This was followed by the gerbils' dragging their ventral scent gland over the peg.
The gerbils with anterior cortical lesions display significantly fewer marks per trial (mean --2.1) than did those with posterior lesions (mean -- 8.4). In fact, ventral marking was com- pletely abolished in 9 of the gerbils with anterior cortical lesions, whereas all 8 of the gerbils with posterior cortical lesions marked on at least one of the three trials (Mann-Whitney U--29 , P < 0.01).
Social interaction. When introduced into a cage with a normal female, normal males usually came into contact with the female within 1 min. The initial contact usually consisted of an extended period (20-30 s) of face to face contact in which one gerbil pressed its snout into the lateral area of the lip margin of the other gerbil. Episodes of genital sniffing and licking were seen less fre- quently. This type of contact often terminated in vigorous chases around the perimeter of the cage and, on occasion, attempts by the male to mount the female. These attempts were, without ex- ception, terminated by the female either by evasion or by an upright defensive stance and a period of 'boxing'.
One gerbil would often attempt to groom the other gerbil by chewing and licking the fur, usually on the flanks or near the back of the head just posterior to the ears. When this occurred, the groomed gerbil was immobile until the contact was terminated by the groomer.
There were obvious qualitative differences in behaviour between normal gerbils and gerbils with anterior cortical lesions. Lesioned males often spent the entire observation period rearing at the sides of the cage or digging in the sawdust. When contacts between the male and the female occurred, they were likely to have been initiated by the female. On those few occasions when male- initiated chases occurred, they terminated very quickly as the male appeared to be unable to follow the complicated evasive maneuvres of the females. Further differences were noted in the
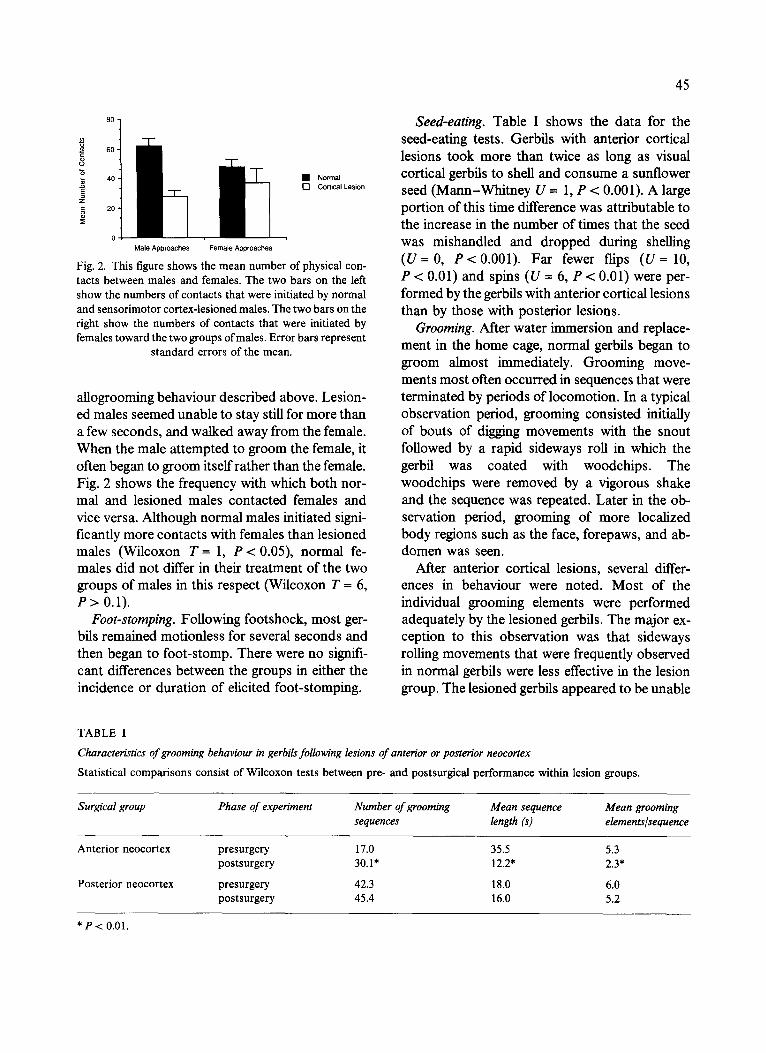
8 0
o~
6o 8 '5
40 • Normal E [ ] Cortical Lesion
~ 2o '5
0
Male Approaches Female Approaches
Fig. 2. This figure shows the mean number of physical con- tacts between males and females. The two bars on the left show the numbers of contacts that were initiated by normal and sensorimotor cortex-lesioned males. The two bars on the right show the numbers of contacts that were initiated by females toward the two groups of males. Error bars represent
s tandard errors of the mean.
allogrooming behaviour described above. Lesion- ed males seemed unable to stay still for more than a few seconds, and walked away from the female. When the male attempted to groom the female, it often began to groom itself rather than the female. Fig. 2 shows the frequency with which both nor- mal and lesioned males contacted females and vice versa. Although normal males initiated signi- ficantly more contacts with females than lesioned males (Wilcoxon T-- 1, P < 0.05), normal fe- males did not differ in their treatment of the two groups of males in this respect (Wilcoxon T = 6, P > 0.1).
Foot-stomping. Following footshock, most ger- bils remained motionless for several seconds and then began to foot-stomp. There were no signifi- cant differences between the groups in either the incidence or duration of elicited foot-stomping.
45
Seed-eating. Table I shows the data for the seed-eating tests. Gerbils with anterior cortical lesions took more than twice as long as visual cortical gerbils to shell and consume a sunflower seed (Mann-Whitney U = 1, P < 0.001). A large portion of this time difference was attributable to the increase in the number of times that the seed was mishandled and dropped during shelling (U- -0 , P<0.001) . Far fewer flips ( U = 10, P < 0.01) and spins (U = 6, P < 0.01) were per- formed by the gerbils with anterior cortical lesions than by those with posterior lesions.
Grooming. After water immersion and replace- ment in the home cage, normal gerbils began to groom almost immediately. Grooming move- ments most often occurred in sequences that were terminated by periods of locomotion. In a typical observation period, grooming consisted initially of bouts of digging movements with the snout followed by a rapid sideways roll in which the gerbil was coated with woodchips. The woodchips were removed by a vigorous shake and the sequence was repeated. Later in the ob- servation period, grooming of more localized body regions such as the face, forepaws, and ab- domen was seen.
After anterior cortical lesions, several differ- ences in behaviour were noted. Most of the individual grooming elements were performed adequately by the lesioned gerbils. The major ex- ception to this observation was that sideways rolling movements that were frequently observed in normal gerbils were less effective in the lesion group. The lesioned gerbils appeared to be unable
TABLE I
Characteristics of grooming behaviour in gerbils following lesions of anterior or posterior neocortex
Statistical comparisons consist of Wilcoxon tests between pre- and postsurgical performance within lesion groups.
Surgical group Phase of experiment Number of grooming Mean sequence Mean grooming sequences length (s) elements~sequence
Anterior neocortex presurgery 17.0 35.5 5.3 postsurgery 30.1" 12.2* 2.3*
Posterior neocortex presurgery 42.3 18.0 6.0 postsurgery 45.4 16.0 5.2
* P < 0.01.

46
TABLE I1
Characteristics of seed-eating behaviour in gerbils after anterior or posterior lesions of the neocortex
Surgical group Mean time to eat a seed (s) Mean number of drops Mean number of flips Mean number of spins
Anterior neocortex 59.6** 3.3** 0.3* 0.3* Posterior neocortex 25.8 0.5 0.8 0.8
* P < 0.01, Mann-Whitney; ** P < 0.001, Mann-Whitney.
to execute the rapid sideways rolls with anything more than a slight exposure of the flank to the woodchips. Because this movement was one of the major means by which gerbils removed exces- sive water from their coats, the lesioned gerbils were noticeably wetter than normal gerbils at the end of the observation period.
A more dramatic effect of the lesions, however, involved the apparent inability of gerbils with an- terior cortical lesions to organize coherent se- quences of grooming movements. Grooming in these gerbils was characterized by very short series of movements that were frequently inter- rupted by walking. Gerbils with visual cortical lesions, on the other hand, showed no differences in grooming behaviour after the lesions. These differences are summarized in Table II.
Discussion
Ventral marking Although the frequency of ventral marking epi-
sodes was dramatically reduced in the experimen- tal group, on those few occasions on which they occurred, the details of the movements appeared to be unaffected by the lesions. Although the function of marking behaviour in general is usually said to be related to territorial defense, scent deposits of most mammals are a source of attraction rather than repulsion 9. Daly 7 has argued that the function of ventral marking in the gerbil is to reduce intersexual aggression and hence to facilitate mating. One possibility, there- fore, is that the observed deficits were related to impairments in social behaviours that have been reported after frontal lesions 3'~4'15. The ventral marking behaviour consists of a stereotyped se-
quence of discrete movements consisting of approach, sniffing, and marking. It is also possi- ble, therefore, that the deficits were caused by the inability of frontal animals to chain together certain types of movements into appropriate se- quences 24,2s.
Social interaction The finding that gerbils with lesions of anterior
cortex were less likely to make contact with fe- males is in agreement with the literature on the effects of such lesions on sexual behaviour in rats. Most of the rat studies have focussed on the role of cortex in copulation. For example, a detailed investigation of the movements of male-female pairs of rats both immediately before and during copulation 27 revealed that decorticate rats show very few deficits in specific movement sequences required for copulation. Whishaw and K o l b 27
argue that the delayed initiation of mating in de- corticates may be partially due to their inability to perform a stereotyped maneuvre involving clasping of the female's back between the fore- paws. This clasping manenvre seems to somehow precipitate the rest of the copulatory sequence.
Although previous experiments have focussed on the behavioural components of copulation (mounts and intromissions) and upon the chase behaviour that is closely associated with these acts, we chose to examine the behaviour of newly introduced male-female pairs. There is evidence that gerbils undergo an extensive period of courtship before mating 16, and our own observa- tions with a breeding colony have suggested that the failure rate of courtships is quite high. It is likely, therefore, that successful mating depends upon correct intersexual behaviours and possibly

some form of male display. Such behaviours gain adaptive significance by veridically signalling male competence 2, so they are likely to be difficult and complex. This experiment identified two components of the early interaction of opposite sex pairs in which males with anterior cortical lesions behave abnormally. The first of these was the lowered rate of bodily contact initiated by males. Our observations indicate that one possi- ble cause of this lowered rate of contact was the difficulty that males appeared to experience in locating the female when she was moving rapidly. Experiments concerned with spatial behaviour have shown that rats with such lesions are deficient in the performance of place learning, but not cue learning, in a Morris maze 26. Because the location of a visible female in a colony cage is a cue learning task, one might expect the gerbils with lesions to perform quite well. Vanderwolf et al.24 noted a certain sluggishness in response to peripherally presented visual stimuli in decorti- care rats, and Whishaw and K o l b 27 reported that males seemed to be 'less efficient' in their tracking of females during sexual encounters. Both of these reports are similar to our own observations and suggest that the high temporal demands of the location task in our experiment were responsible for the observed deficit.
An alternative and more conservative explan- ation for the inability of males to make contact with females is that the problem is one of the control of locomotion rather than of localization. The lesioned males may be unable to walk reliably toward a target female or to inhibit locomotion when they arrive at the female's location. This explanation fits well with the abnormal loco- motion observed during grooming.
The second abnormality consisted of the male's apparent inability to maintain immobility while allogrooming was taking place. Although not quantified in our experiment, this deficit was strikingly obvious and was observed repeatedly. It is likely that such behaviour could delay or rule out the onset of mating by signalling to the female the incompetence of the male. This deficit is re- miniscent of the well-known deficits in response inhibition after frontal lesions ~°.
47
Seed-eating It is well known that lesions of motor cortex
lead to deficits in manipulatory movements of the digits 5 and of the forelimbs 11,28 in rats, even with quite small lesions restricted to the medial aspect of motor cortex. It is not surprising, therefore, that our large anterior lesions produced such deficits in the gerbil. The forelimb and digit usage that is required to successfully shell a sunflower seed is quite intricate. Gerbils with lesions of motor cor- tex were rarely able to execute such movements, and consequently, they dropped each seed a num- ber of times before it was successfully shelled. Although we did not record the time to shell and the time to consume a seed separately (this is quite a difficult distinction to make because seeds are often partially consumed before the shell is fully removed), it is possible that some of the increased time required by the gerbils to fully shell and con- sume the seed was attributable to deficits in move- ments of the tongue 6. It is worth emphasizing that, as in rats, the movements that were impaired by the anterior lesions were specifically those that could be classified as manipulatory movements. Other types of forelimb movements, such as those occurring during locomotion or face-washing, were functional, and appeared to be performed normally.
Grooming The observation that grooming sequences were
short and impoverished in gerbils with frontal lesions is consonant with previous findings in rats 24"28. Like these findings, there was a higher incidence of bouts of locomotion resulting in shorter grooming sequences. One difficulty of interpretation in the grooming results concerns the relatively large differences between the preoperative scores of the two lesion groups on some of the measures used. Even before surgery, animals in the posterior group produced shorter and more frequent grooming sequences than ani- mals in the anterior group. On the other hand, the number of grooming elements per sequence and the total time spent grooming were roughly com- parable in these groups. The inevitable conclusion from this seems to be that, before surgery, there

48
were differences between the groups in the time spent on each movement in a grooming sequence. The reason for these differences is not at all clear. Regardless of these differences, it remains true that the lesions affected differentially the structure of grooming sequences in the two groups. Pos- terior neocortical lesions had no effect on the preoperative pattern of results, but anterior neocortical lesions produced shorter sequences with fewer elements. Although wetting the fur is a reliable method of eliciting long bouts of grooming, there is evidence to suggest that this procedure may be at least partially responsible for the observed changes in grooming. Whishaw et al . 29 found that the grooming behaviour of dry decorticate rats did not differ from that of con- trois with respect to sequence lengths. They suggest that the wetting procedure may increase the likelihood of locomotion in response to the novel stimulus of the wet fur, or that the lesions may affect the orthokinetic response that is characteristic of wet animals in general as an adaptive thermoregulatory response 4.
EXPERIMENT II
In the In'st experiment, we discovered that large anterior cortical lesions abolished or reduced the frequency of ventral marking in gerbils. We, there- fore, decided to conduct a second, more detailed investigation of this particular deficit. To this end, we made small lesions of 3 discrete areas of cor- tex: primary motor cortex, primary somato- sensory cortex, and cingulate cortex.
Method
Subjects The subjects were 24 male Mongolian gerbils
between 3 and 6 months of age. These gerbils were housed individually on a 12 : 12 light : dark schedule with continuous access to Purina Lab Chow and water.
Apparatus A clear Plexiglas arena (36 x 30 × 18 era) was
used for testing. Plexiglas inserts were construct- ed to fit the bottom of the arena and 6 roughened
Plexiglas pegs (3 x 2 x 0.6 cm) were glued to the inserts in two rows of three, at equal spacings from each other and the walls of the arena.
Surgery General surgical procedures were the same as
those of Expt. I. There were 4 surgical groups as follows: sham (n = 6), somatosensory cortex (n = 6), primary motor cortex (n = 6) and cingu- late cortical lesions approximately 5 mm in length and centred on bregrna (n = 6). Following surge- ry, gerbils were given 2 weeks to recover during which time they were weighed daily and fed wet mash from a shallow Petri dish.
Procedure During each trial, a gerbil was placed in the
apparatus for a period of 5 min under dim illumi- nation (a single shielded 60-W lightbulb) in a quiet room. During habituation trials, no data were recorded, but the experimenter sat nearby in order to habituate the gerbil to his presence. During test trials, the 5-min period was divided into 30-s in- tervals, and the experimenter recorded each oc- casion of ventral marking. At the end of a trial, the gerbil was removed and returned to its home cage. The plastic insert was removed from the test chamber, washed with strong detergent and water, and allowed to dry overnight. A sufficient number of inserts were used so that no insert was used twice in one day. The allocation of inserts to individual gerbils was randomized daily. Between trials, the test chamber was thoroughly washed with a solution of 70% ethyl alcohol and allowed to dry.
Each gerbil received 3 habituation trials and 3 test trials over the 6 days immediately preceding surgery. Following surgery and recovery, the same 6-trial regimen was repeated. At the con- clusion of testing, gerbils were sacrificed by over- dose of sodium pentobarbital and the same histological procedure as described in Expt. I was carried out.
Results
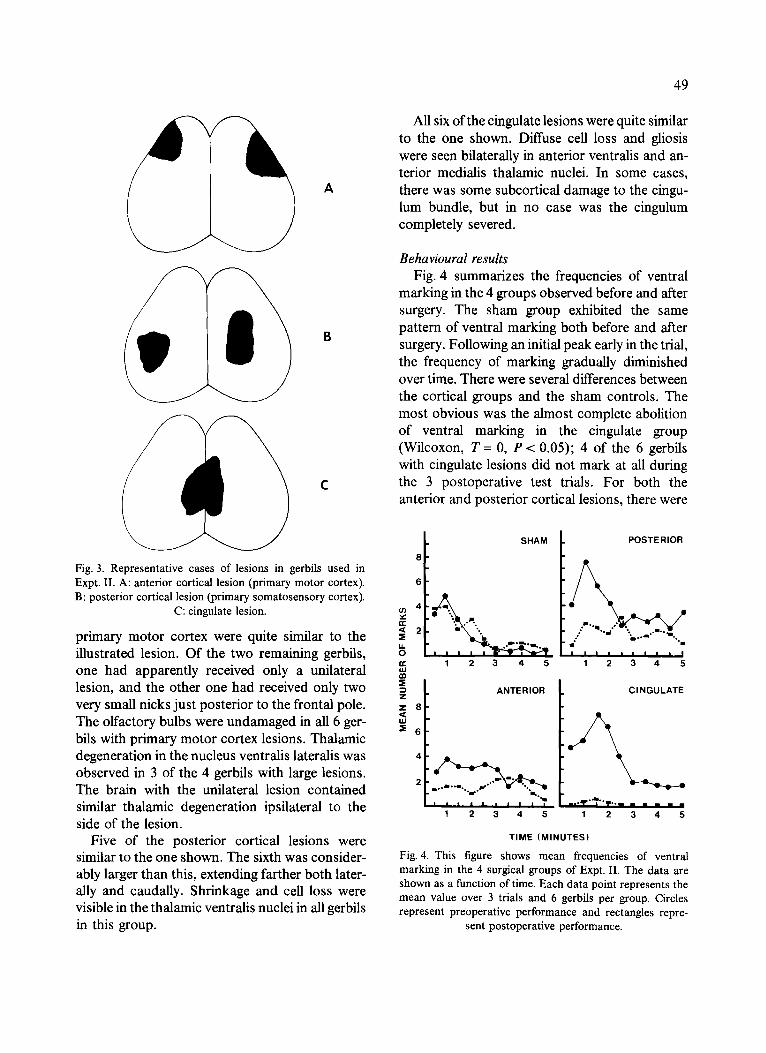
Anatomical results Fig. 3 illustrates a representative lesion from
each of the 3 lesion groups. Four of the lesions of
A
B
C
Fig. 3. Representative cases of lesions in gerbils used in Expt. II. A: anterior cortical lesion (primary motor cortex). B: posterior cortical lesion (primary somatosensory cortex).
C: cingulate lesion.
primary motor cortex were quite similar to the illustrated lesion. Of the two remaining gerbils, one had apparently received only a unilateral lesion, and the other one had received only two very small nicks just posterior to the frontal pole. The olfactory bulbs were undamaged in all 6 ger- bils with primary motor cortex lesions. Thalamic degeneration in the nucleus ventralis lateralis was observed in 3 of the 4 gerbils with large lesions. The brain with the unilateral lesion contained similar thalamic degeneration ipsilateral to the side of the lesion.
Five of the posterior cortical lesions were similar to the one shown. The sixth was consider- ably larger than this, extending farther both later- ally and caudally. Shrinkage and cell loss were visible in the thalamic ventralis nuclei in all gerbils in this group.
49
All six of the cingulate lesions were quite similar to the one shown. Diffuse cell loss and gliosis were seen bilaterally in anterior ventralis and an- terior medialis thalamic nuclei. In some cases, there was some subcortical damage to the cingu- lum bundle, but in no case was the cingulum completely severed.
Behavioural results Fig. 4 summarizes the frequencies of ventral
marking in the 4 groups observed before and after surgery. The sham group exhibited the same pattern of ventral marking both before and after surgery. Following an initial peak early in the trial, the frequency of marking gradually diminished over time. There were several differences between the cortical groups and the sham controls. The most obvious was the almost complete abolition of ventral marking in the cingulate group (Wilcoxon, T = 0, P < 0.05); 4 of the 6 gerbils with cingulate lesions did not mark at all during the 3 postoperative test trials. For both the anterior and posterior cortical lesions, there were
8
6
u) 4
n,-
,~ 2 I =E it 0
tu ~3 =E
z z 8
u.i
6 ¸
SHAM
. ~ .''-...,. . . . . .,.
1 2 3 4 5
ANTERIOR
| • i | | | | ! i m
1 2 3 4 5
POSTERIOR
m ~ m . o q , .
n n | | i a n | | | 1 2 3 4 5
CINGULATE
1 2 3 4 5
T IME (M INUTES)
Fig. 4. This figure shows mean frequencies of ventral marking in the 4 surgical groups of Expt. II. The data are shown as a function of time. Each data point represents the mean value over 3 trials and 6 gerbils per group. Circles represent preoperative performance and rectangles repre-
sent postoperative performance.

50
no significant differences between overall pre- and postsurgical marking frequencies (T = 4, P > 0.1 ; T = 3, P > 0.1). In both of these groups, however, there was some tendency for the frequency of ventral marking to be relatively low during the early part of the trials. Statistical tests showed that the pre- and postsurgical functions relating marking frequency and time were uncorrelated in the anterior and posterior lesion groups (r's = 0.14, 0.02, respectively, P > 0.1) and highly correlated in the sham and cingulate groups (r's = 0.84, 0.83, respectively, P < 0.01).
Discussion
Normal gerbils marked most frequently in the early part of the test trials. This finding is consonant with the results of other investigations (ref. 20 for review). Since an intratrial gradient of activity is usually observed in open-field studies 24, the observed gradient in the frequency of the marking response may be related to overall activity levels. In the absence of independent measurements of activity, this possibility is diffi- cult to evaluate.
The data suggest that cortical lesions might affect the ventral marking response in two differ- ent ways. Cingulate lesions resulted in an almost complete loss of ventral marking behaviour. However, on those infrequent occasions when cingulate gerbils marked, the event was most likely to occur in the early part of the trial, just as with normal gerbils. Lesions of primary motor and primary somatosensory cortex, on the other hand, affected particularly the frequency of marking in the early part of the test trials. This deficit may be secondary to a primary deficit in the efficiency with which the gerbils explored a some- what novel environment. Indeed, experiments with rats 28 have shown that large cortical lesions may alter patterns of open field behaviour. These effects, like those seen in grooming and in social situations, could best be described as the presence of excessive and 'aimless' locomotion.
GENERAL DISCUSSION
In general, the findings of these experiments with gerbils are consonant with other experiments
dealing with the effects of cortical lesions in rodents 1°. The effects of the large cortical ablations on manual dexterity are consonant with the well-documented fact that primary motor cor- tex plays a special role in the fine control of distal effectors 8a3. The effects of our lesions on other behaviours are also largely in agreement with the findings of other investigators, as reviewed in the discussions of specific experiments. So far as we know, the report of deficits in ventral marking after cortical lesions is a novel finding.
The major problem facing the investigator of cortical function is to extract some general state- ment from the data that describes what cortex does. One approach to this problem is to use electrophysiological criteria to categorize be- haviour that is or is not dependent on cerebral cortex. As described in the Introduction, Type I behaviours are accompanied by neocortical low- voltage fast activity and hippocampal rhythmical slow activity, whereas Type II behaviours may or may not be accompanied by such patterns of elec- trical activity. One attractive feature of this scheme is that it allows genuine a priori pre- dictions as to which behaviours will be affected by lesions and which ones will not. In the present experiments, the predictions that ventral marking would be affected and that foot-stomping would not be affected by frontal cortical lesions were made on the basis of electrophysiological evi- dence and were, in fact, borne out by the experi- ments. A second attraction is that the relatively colourless terminology (Type I and II) sidesteps the need to employ the language of psychology to describe the pattern of deficits. Without doubt, there will be a time when we will have to come to terms with the meaning of the Type I/Type II dis- tinction. The use of a neutral terminology that has its roots in brain architecture, however, makes it less likely that we will automatically assume that the structure-function relationships of the brain conform to the logical structures of scientific psy- chology 21.
ACKNOWLEDGEMENTS
This research received financial support from Natural Sciences and Engineering Research

Council of Canada research Grants A6313 (M.A. Goodale) and A0118 (C.H.V.).
REFERENCES
1 Baran D. and Glickman, S.E., 'Territorial marking' in the Mongolian gerbil: a study of sensory control and function, J. Comp. Physiol. Psychol., 17 (1970) 123-245.
2 Barash, D.P., Sociobiology and Behavior, Elsevier, New York, 1982.
3 Butter, C.M. and Snyder, D.R., Alterations in aversive and aggressive behaviors following orbital frontal lesions in rhesus monkey, Acta Neurobiol. Exp., 32 (1972) 115-156.
4 Cabanac, M., Thermoregulatory behavior. In J. Bligh and R. Moore (Eds.), Essays in Temperature Regulation, Elsevier, New York, 1972.
5 Castro, A.J., The effects of cortical ablations on digit usage in the rat, Brain Res., 73 (1972) 137-185.
6 Castro, A.J., The effects of cortical ablations on tongue usage in the rat, Brain Res., 45 (1972) 251-253.
7 Daly, M., Some experimental tests of the functional signi- ficance of scent-marking by gerbils (Meriones unguicula- tus), J. Comp. Physiol. Psychol., 91 (1977) 1082-1094.
8 Gentile, A.M., Green, S., Nieburgs A., Schmelzer, W. and Stein D.G., Disruption and recovery of locomotor and manipulatory behavior following cortical lesions in rats, Behav. Biol., 22 (1978) 417-455.
9 Johnson, R.P., Scent-marking in mammals, Anim. Behav., 21 (1973) 521-535.
10 Kolb, B., Functions of the frontal cortex of the rat: a comparative review, Brain Res. Rev., 8 (1984) 65-98.
11 Kolb, B. and Holmes, C., Neonatal motor cortex lesions in the rat: absence of sparing of motor behaviors and impaired spatial learning concurrent with abnormal cerebral morphogenesis, Behav. Neurosci., 97 (1983) 697-709.
12 Kramis, R.C. and Routtenberg, A., Rewarding brain stimulation, hippocampal activity and foot-stomping in the gerbil, Physiol. Behav., 4 (1969) 7-11.
13 Lawrence, D.G. and Kuypers H.G.J.M., The functional organization of the motor system in the monkey. I. The effects of bilateral pyramidal lesions, Brain, 91 (1968) 1-18.
14 Lubar, J.F., Herrman, T.F., Moore, D.R. and Shouse, M.N., Effect of septal and frontal ablations on species- typical behavior in the rat, J. Comp. Physiol. Psychol., 83 (1973) 260-270.
15 Nonneman, A.J. and Kolb, B., Lesions of hippocampus or prefrontal cortex alters species-typical behavior in the cat, Behav. Biol., 12 (1974) 41-54.
51
16 Norris, M.L. and Adams, C.E., Aggressive behaviour and reproduction in the Mongolian gerbil (Meriones unguicula- tus), relative to age and sexual experience at pairing, J. Reprod. Fertil., 31 (1972) 447-450.
17 Shook, B.L., Functional Organization of the Gerbil's Visual System: A Move to Revise Traditional Working Hypotheses of Brain Organization, Ph.D. Thesis, Department of Psy- chology, Brandeis University, 1983.
18 Stewart, D.J., MacFabe D.A. and Vanderwolf, C.H., Cholinergic activation of the electrocorticogram: role of the substantia innominata and effects of atropine and quinuclidinyl benzilate, Brain Res., 322 (1984) 219-232.
19 Thiessen, D.D., The roots of territorial marking in the Mongolian gerbil: a problem of species-common topogra- phy, Behav. Res. Methods and Instr., 1 (1968) 70-76.
20 Thiessen, D. and Yahr, P., The Gerbil in Behaviorallnvesti- gations, Texa Press, Austin, 1977.
21 Vanderwolf, C.H., The influence of psychological con- cepts on brain-behavior research, In T.E. Robinson (Ed.), Behavioral Approaches to Brain Research, Oxford, New York, 1983.
22 Vanderwolf, C.H., Role of the cerebral cortex and ascending activating systems in the control of behavior. In E. Satinoff and P. Teitelbaum (Eds.), Handbook of Be- havioral Neurobiology, Plenum, New York, 1983.
23 Vanderwolf, C.H. and Baker, G.B., Evidence that sero- tonin mediates non-cholinergic neocortical low voltage fast activity, non-cholinergic hippocampal rhythmical slow activity and contributes to intelligent behavior, Brain Res., 374 (1986) 342-356.
24 Vanderwolf, C.H., Kolb, B. and Cooley, R.K., Behavior of the rat after removal of the neocortex and hippocampal formation,J. Comp. PhysioL Psychol., 92 (1978) 156-175.
25 Whishaw, I.Q., Hippocampal electroencephalographic activity in Mongolian gerbil during natural behaviors and wheel running and in rat during wheel running and con- ditioned immobility, Can. J. Psychol., 26 (1972) 219-232.
26 Whishaw, I.Q. and Kolb, B., Decortication abolishes place but not cue learning in rats, Behav. Brain Res., 11 (1984) 123-134.
27 Whishaw, I.Q. and Kolb, B., The mating movements of male decorticate rats: evidence for subcortically generat- ed movements by the male but regulation of approaches by the female, Behav. Brain Res., 17 (1985) 171-191.
28 Whishaw, I.Q., Kolb, B. and Sutherland, R.J., The analy- sis of behavior in the laboratory rat. In T.E. Robinson (Ed.), Behavioral Approaches to Brain Research, Oxford, New York, 1983.
29 Whishaw, I.Q., Nonneman, A.J. and Kolb, B., Environ- mental constraints on motor abilities used in grooming, swimming, and eating by decorticate rats, J. Comp. Physiol. Psychol., 95 (1981) 792-804.





























