Embed Size (px)
Citation preview

Beta Receptor Occupancy
ASSESSMENT IN THE INTACT ANIMAL
CHARLES J. HoMcY, H. WILLIAM STRAUSS, and SUSAN KOPIWODA, Cardiac Unit,Department of Medicine and the Division of Nuclear Medicine, Department ofRadiology, Massachusetts General Hospital, and the Harvard Medical School,Boston, Massachusetts 02114
A B S T R A C T Organ uptake of 1251-hydroxybenzyl-pindolol, a potent beta adrenergic antagonist, wasdetermined after intravenous administration. Pre-treatment with the beta agonist, epinephrine, inhibitedan almost identical fraction of 1251-hydroxybenzyl-pindolol binding as did the antagonist, propranolol.Specific beta receptor binding accounted for 50% oftotal uptake in the lung and demonstrated the followingcharacteristics. The dose-response curve for propranololinhibition of 1251-hydroxybenzylpindolol bindingduplicated that reported for its physiologic action.Simultaneous serum propranolol levels as determinedby a sensitive radioimmunoassay allowed an apparentdissociation rate constant -7 nM to be obtained thatcorrelated closely with the results reported frommembrane binding studies. Alpha blockade had noeffect and inhibition of 1251-hydroxybenzylpindololbinding by propranolol demonstrated stereospecificity.After chemical sympathectomy with reserpine or 6-OHdopamine, there was a 100% increase in receptorspecific binding. Finally, a scintillation camera wasemployed to visually and quantitatively detect 1251_hydroxybenzylpindolol displacement from the lungduring intravenous propranolol administration in theliving animal. Reversal of binding was rapid and an invivo inhibition curve was generated. Such a methodprovides the potential for longitudinally assessing betareceptor occupancy and apparent affinity directly in man.
INTRODUCTION
Significant progress has been made in the identifica-tion and characterization of cell membrane receptorsin the past decade. Compounds have been synthesizedthat can be labeled to high specific activity and willstill bind to receptors with high affinity (1). Theseagents have been used primarily for the in vitro
Receivedfor publication 11 June 1979 and in revisedform10 December 1979.
identification and localization of receptors in specifictissues. In the study of the beta adrenergic receptor,antagonists have been preferentially used in that theymay demonstrate less nonspecific membrane bindingand do not undergo chemical oxidation in the samemanner as catecholamines (2-4). The synthesis,several years ago, of 3-indoloxy-l-(2-p-hydroxybenzyl-propyl-2-amino) isopropanol, called hydroxybenzyl-pindolol (5), provided an agent that could readily belabeled with iodine and still maintain its high affinitybinding to beta receptors with a reported dissociationrate constant (KD)' of -0.1 nM (6, 7). In vitro character-ization of cell membrane receptors, however, does notprovide data concerning receptor occupancy by endog-enous agonists in the intact animal and consequentlyabout mechanisms that may be important in the regula-tion of synaptic activity.
This study was undertaken to define the in vivodistribution of 1251-hydroxybenzylpindolol and to deter-mine whether this could be used to measure betareceptor binding in a particular organ. Specific bindingwas identified by comparing the distribution datafrom control animals with that from animals who hadpreviously received the beta blocking agent, pro-pranolol. To further validate that such changes in 125I_hydroxybenzylpindolol distribution are related to betareceptor binding, several lines of evidence weregenerated. Agonist pretreatment with epinephrinewas shown to inhibit 1251-hydroxybenzylpindolol bindingin a manner identical to that for the antagonist, pro-pranolol. Propranolol inhibition of 1251-hydroxybenzyl-pindolol binding followed a dose-response patternthat duplicated its physiological effects. An apparentKD as determined from simultaneous serum propranolollevels was similar to that obtained from direct membranebinding studies. Alpha blockade had no effect on1251-hydroxybenzylpindolol binding. Propranolol-
I Abbreviation used in this paper: KD, dissociation rateconstant.
J. Clin. Invest. (© The American Society for Clinical Investigation, Inc. * 0021-9738/80/05/1111/08 $1.00 1111Volume 65 May 1980 1111-1118

inhibitable binding possessed stereospecificity. Further-more, this approach detected changes in beta receptoroccupancy produced by pharmacologic interventionssuch as chemical sympathectomy. Finally, specific betareceptor binding was externally quantitated in a livinganimal.
METHODS
Hydroxybenzylpindolol was supplied as a reagent gradechemical from Sandoz Ltd. (Basel, Switzerland). Thecompound was iodinated using a modification of thechloramine-T method (6). The labeling efficiency averaged40%. The iodinated product was purified to a theoreticalspecific activity of -2,000 Ci/mmol using a paper chromato-graphic technique previously described (6). The retardationfactor (Rf) (0.50) of the iodinated product by thin-layerchromatography on silica gel plates in diethylamine:toluene(5:6) was similar to the starting material.
Biodistribution studies were performed in type CD-1 miceweighing 30-40 g, obtained from the Charles River BreedingLaboratories, Inc., Wilmington, Mass. In all studies, theamount of '25I-hydroxybenzylpindolol injected was -1 pmol(activity of 1-2 uCi). The propranolol-treated animalsreceived the indicated dose of propranolol intravenouslyeither 10 or 15 min before the administration of 1251-hydroxy-benzylpindolol.An initial study was performed in control and propranolol-
pretreated animals to define the distribution of 1251-hydroxy-benzylpindolol over time. Tracer distribution was determinedfollowing sacrifice of the animals with an overdose of ether;samples of blood, kidney, heart, liver, spleen, stomach, smallbowel, thyroid, pancreas, bone, fat, skeletal muscle, and brainwere obtained, weighed, and counted in a gamma counterwith a window of 20-60 keV. In addition to the tissuesamples, an aliquot of injected dose was counted at the sametime and results expressed as percent injected dose perorgan. Data is expressed as mean ±its standard duration. Com-parison of groups was made using the Student t test.
In other experiments, agonist-inhibitable binding wasdetermined following pretreatment with epinephrine. 50-,ug doses of epinephrine were used because higher dosesresulted in a significant mortality rate among the treatedanimals. To insure that the serum level of the drug wasmaximal, experiments were performed in which the epi-nephrine was administered simultaneously with the labeledhydroxybenzylpindolol. The same degree of inhibition (50%)of hydroxybenzylpindolol binding in the lung occurred aswhen the epinephrine was administered 15 min beforehydroxybenzylpindolol injection. Reserpine-pretreated ani-mals received three 15-,ug doses intraperitoneally at 48,36, and 24 h before the study; 6-OH dopamine was administeredintraperitoneally in two doses of 5 mg per animal at 36 and24 h before the study (8-10).External counting. Imaging and external measurement of
'251-hydroxybenzylpindolol in an anesthetized rabbit was ac-complished with an Ohio-Nuclear 420/550 gamma camera (Ohio-Nuclear, Inc., Subsid. Technicare Corp., Solon, Ohio) using alow-energy parallel hole collimator. High voltage was maxi-mized and a 50% window was used that included the gammaand x-ray emission of 1251. Data was analyzed on a nuclearmedicine data system VP450, Ohio-Nuclear, Inc. Regions ofinterest in the lungs were outlined with a light pen program.Curves representing pulmonary '25I-hydroxybenzylpindololactivity vs. time after injection were then generated. Non-specific binding was determined by measuring lung accumula-tion of 1251-hydroxybenzylpindolol in control rabbits of
identical weight that were pretreated with intravenouspropranolol (1.5 mg/kg) 15 min before the injection of thelabel. The curves in Fig. 6 were computed after subtractingthis value for each time point.Radioimmunoassay. Antibodies that recognize propranolol
with high affinity were raised in the following manner. Thebromohydrin of alprenolol was synthesized according to themethod of Hoebeke et al. (11). This was confirmed bynuclear magnetic resonance which showed loss of the olefinproton signal and by elemental analysis of the product(Galbraith Laboratories, Knoxville, Tenn.). The bromohydrinwas then reacted with bovine serum albumin that had pre-viously been modified by reaction with N-acetylhomocysteine-thiolactone. This resulted in a product with 2 mol ofalprenololbound per mol of albumin (as later determined by radio-immunoassay ofthe hydrolyzed product). New Zealand Whiterabbits were immunized with 0.5 mg of this immunogenevery 3 wk. Approximately 3-4 mo later, antiseruim washarvested that bound both [3H]alprenolol and [3H]propranololwith an apparent KD of 5 nM similar to that previously re-ported (11, 12). The final assay employed 100 ,ul sera, 50 ,ul4 nM [3H]propranolol (final concentration, 1 nM) and 50 Ilof a 1:10 or 1:50 dilution of antiserum.2 Separation of boundfrom free ligand was performed by a double antibody pre-cipitation technique. In the following manner, it was alsoshown that these antibodies primarily recognized the pro-pranolamine side chain. Thus 4-OH propranolol (an effec-tive beta blocker), alprenolol, and propranolol were recognizedwith similar affinities despite their different ring structures(apparent KD 5 nM) whereas naphthoxyacetic acid, whichpossesses an identical ring structure to propranolol, butlacks the requisite side chain, was not recognized by thisantibody class. Catecholamines were recognized as a resultof the similarity of their ethanolamine side chain but witha 500-fold lower affinity. A variety of other drugs (aldomnet,digoxin, and tricyclic antidepressants) did not react at con-centrations at least 10,000-fold greater or not at all.Catecholamine determination. Serum epinephrine and
norepinephrine were measured by the radioisotopic enzymaticmethod of DaPrada and Zurcher (13), using an ethanolamine-chloroform-methanol solvent system. In this system, bothcompounds are 0 [3H]methylated and then separated by thin-layer chromatography providing resolution from each otherand from circulating precursors. Some hemolysis of samplesoccurred because of the inability to obtain adequate vol-umes from the same mice used in the above protocols. How-ever, these measurements were aimed at simply showing amajor reduction in catecholamine concentrations followingthese protocols. Norepinephrine levels in peripheral bloodwere in the range of 10 ng/ml with epinephrine concentra-tions averaging 1-3 ng/ml which, to some extent, representsthe stress response that these animals undergo (14).
RESULTS
The blood disappearance of 1251-hydroxybenzylpindololwas rapid, and at 5 min after administration in thecontrol animals, <6% of the injected dose was presentin the entire blood volume of the mouse. Data on thedistribution of 1251-hydroxybenzylpindolol are sum-marized in Fig. 1. The liver was the organ with the
2 Homcy, C., S. Rockson, R. Cody, and E. Haber. 1980. Asensitive radioimmunoassay potentially applicable to all,8-blocking drugs. American Heart Association Meeting.Accepted for presentation.
1112 C. J. Homey, H. W. Strauss, and S. Kopiwoda

Ioo
5
3co-A
cF._
do
AR
40-
10-
propronolol i.v.
* corrected for nonspecific binding
i 4 i A 10 1'2 14 16 18
thu. (minutes)
FIGURE 1 Mice were pretreated either with normal saline, D,L-propranolol (15 mg/kg), or
epinephrine (1.5 mg/kg) in an identical volume (0.1 ml); 15 min later, '25I-hydroxybenzylpindolol(1 ,Ci) was injected intravenously and 15 min thereafter, the animnal was killed. The organs were
removed and their activity determined in a Packard gamma counter (Packard Instrument Co.,Inc.). At least three experiments, each with six mice in the control and six in the propranololgroup, were performed. Activity (percent dose per organ) has been normalized with thecontrol being 100%. Standard deviation bars are shown. Each bar had included within itthe percent dose per organ under control conditions (lung, P < 0.001; spleen, P < 0.05; pancreas,
P < 0.01; no significant changes were detected in other organs shown; n = 18). O, total; 0,propranolol; *, epinephrine.
highest concentration of radiopharmaceutical, followedby skeletal muscle, kidney, bone, and bowel. Over thecourse of time, the concentration in most organs exceptthyroid, fatty tissue, and small bowel decreased. Ingeneral, maximal uptake in all organs occurred within 5min and gradually decreased over the next 25 min. Theeffects of propranolol were greatest in the lung, spleen,pancreas, thyroid, and blood. Approximately 50% of'251-hydroxybenzylpindolol binding in the lung wasconsistently blocked by propranolol whereas only10-20% was consistently inhibited in these otherorgans. Therefore, further studies detailed below con-centrated on i251-hydroxybenzylpindolol binding inthe lung only.
It was expected that maximal agonist (epinephrine)inhibition of 125I-hydroxybenzylpindolol binding
would duplicate the effects of the antagonist, pro-pranolol. Therefore, the same protocol was employedwith the exception that the beta agonist, epinephrine,was used to define specific binding. As can be seenin Fig. 1, the same fraction and distribution of receptorspecific binding resulted as had been obtained utilizingpropranolol.
Inhibition of 1251-hydroxybenzylpindolol binding bypropranolol pretreatment was a saturable process andoccurred at doses of propranolol at which the physio-logical effects of the drug are observed (15). A com-petitive inhibition curve relating serum propranololconcentrations to hydroxybenzylpindolol binding isshown in Fig. 2. It can be seen that 50% inhibitionof hydroxybenzylpindolol binding occurs at a D,L-propranolol concentration of 7 nM. It should be noted
Beta Receptor Occupancy 1113

-0
._
.2
.Cc
am
1 M lOnM 0.QiUM 1'AM 1OUM
Propronolol
- 3.0
- 2.7
Ca
1.- 2.4 0
- 2.1 00
- 1.8
c
0N
(n00
-00-
5.0
4.0
L 1.5
FIGURE 2 Doses of 0.001 mg/kg, 0.01 mg/kg, 0.05 mg/kg, 0.1mg/kg, and 1 mg/kg D,L-propranolol were uised. The meanpropranolol concentration from the three experiments wasused for each dosage. The standard error at 0.001 mg/kg, 0.01mg/kg, 0.05 mg/kg, and 0.1 mg/kg was <10% and at 1.0 mg/kgwas <15%. Blood samples were collected before sacrificeand serum propranolol levels were determined by radio-immunoassay. Standard deviation bars for '251-hydroxy-benzylpindolol binding are shown. Five animals were used ateach dose in every experiment. The results from threeindividual experiments are included.
that the concentration of the active isomer is in therange of3 to 4 nmol. These serum levels were achievedby increasing the dose from 0.001 to 0.1 mg/kg. There-after, a 10-fold increase in the dose to 1.0 mg/kg resultedin a serum level of 5 ,LM with no fuirther inhibitionof binding. Residual binding at this serum propranololconcentration was considered to represent nonspecificbinding and was subtracted from total binding at eachpoint to determine specifically bound counts.Alpha blockade did not affect 1251-hydroxybenzyl-
pindolol binding. Fig. 3 demonstrates that high dosepretreatment with an alpha blocker, phentolamine(3 mg/kg), using the same protocol as that for pro-pranolol, did not significantly alter lung '251-hydroxy-benzylpindolol binding. A simultaneous experimentinvolving propranolol pretreatment is shown forcomparison.
Specific 1251-hydroxybenzylpindolol binding demon-strates stereospecificity as seen in Fig. 4. Doses of 0.1mg/kg of L-propranolol inhibit -45-50% of 1251_hydroxybenzylpendolol binding, whereas the D-isomerblocks only 10% ofthe binding at a dose fivefold greater.
Additional studies were performed in chemicallysympathectomized animals to determine whetherchanges in 1251-hydroxybenzylpindolol binding in vivomight reflect alterations in beta receptor occupancyor number. Fig. 5 depicts the result obtained fol-lowing such a chemical sympathectomy. In theseexperiments, groups of mice were treated either withreserpine or 6-OH dopamine. Then the typical protocoloutlined previously was followed whereby 1251-hydroxy-
1.0
Lung
FIGURE 3 A similar protocol was also employed in theseexperiments with a phentolamine dose of 3.0 mg/kg and apropranolol dose of 1.5 mg/kg. The mean and its standarddeviation is shown (P < 0.001 for propranolol-treated grotups(Pr) vs. control groups (C); P < 0.1 for phentolamine-treatedgrouips (Ph) vs. control grouips; n = 10).
benzylpindolol binding in saline-treated controls wascompared with that in propranolol-treated animals.After the reserpine treatment, the concentrations of1251-hydroxybenzylpindolol in the lungs rose markedlyas shown in Fig. 5. Furthermore, propranolol-inhibitablebinding compromised all of this increase, consistentwith the proposal that beta receptor binding wasbeing delineated. Additional confirmation of this findingis that identical results were obtained after 6-OH
41
31a.
CL
control L-prop D-prop L-prop D-prop0.1 mg/kg 0.5 mg/kg
FIGURE 4 The protocol described previously was used withthe doses of the individual isomers shown. The mean andits standard error is shown. Differences between L-propranolol-treated and control groups were significant at the P < 0.001level (n = 10). HBP, hydroxybenzylpindolol.
1114 C. J. Homcy, H. W. Strauss, and S. Kopiwoda
C I Ph I Pr

SPECIFIC BINDING
Control Reserpine 6 O Dopomine
+4Con- Reser- 6 OHtrol pine Dopamine
FIGURE 5 Mice were pretreated with either reserpine or 6-OH dopamine as described inMethods. The same protocol as described previously was then used. The first graph depicts datafrom control, reserpine, and 6-OH dopamine-treated animals with and without propranololpretreatment (1.5 mg/kg). The second graph shows specific binding only for each of these groups.Differences between the propranolol-treated and control animals in each group were significant atthe P < 0.001 level (n = 10).
dopamine infusion which produced a chemical sympa-
thectomy via a different mechanism. Both drugsresulted in a significant depletion of norepinephrinelevels measured from peripheral blood samples(reserpine 17% of control and 6-OH dopamine 19%ofcontrol). Reserpine also decreased serum epinephrinelevels to 24% of control, whereas 6-OH dopamineactually resulted in an increase of 50%. However,epinephrine only accounted for 10-20% of total serumcatecholamines.
Detection ofbeta receptor binding was also attemptedin a living animal and an in vivo competitive inhibi-tion curve was generated. A 3-kg rabbit was anesthetizedand injected with 150 ,uCi of 1251-hydroxybenzyl-pindolol via an ear vein. At 2 min after injection, thelungs were identified from the whole body imageutilizing a scintillation camera and total counts were
collected for 1 min from a defined area ofinterest withinthe lung fields. 2 min after the injection, a 200-,ug bolusof propranolol was injected and again counts were
collected for 1 min from the previously defined area ofinterest. This procedure was repeated every 2 min over
a 20-min period. The counts remaining in the definedarea of interest at each time point were first cor-
rected for nonspecific binding as indicated in Methodsand then expressed as a percentage of the initial ac-
tivity recorded at 2 min after 1251-hydroxybenzylpindololinjection and plotted vs. time. The count collectionrecorded from the initial 2-min image was consistentlyin the range of 2,500-3,000 cpm. A control curve froman experiment in which no propranolol was injectedis shown for comparison. The greatest effect was seen
after the first dose as might be predicted in that a rela-tively high serum propranolol concentration resulted.
DISCUSSION
The distribution of the beta antagonist 1251-hydroxy-benzylpindolol in the mouse in these experiments issignificantly different from that of the 'IC-labeledcatecholamine, norepinephrine, as studied by Ansariet al. (16). Ansari found the highest concentration of['1C]norepinephrine in the heart, followed by lung,liver, and kidney. Because the binding of such agentscould be based on either specific binding to the betareceptor or nonspecific binding to other sites on the cellmembrane or within the cell, receptor binding was
defined in our studies by comparing control binding
Beta Receptor Occupancy 1115
z4
LZ00
w(I)00

with that following pretreatment with the beta receptorantagonist propranolol. The most significant changes inthe distribution of 1251-hydroxybenzylpindolol followingsuch pretreatment occurred in the lung with lesserchanges in thyroid, spleen, and blood. These dataindicate that the total organ distribution reflectsprimarily nonspecific binding.We have not yet determined whether specific binding
in the lung is primarily in vascular endothelium orparenchymal epithelium. That it does not representbinding to blood elements is based on the simple ob-servation that a much greater degree of specific bindingwas present in the lung than either in blood or in adensely vascular organ such as spleen. In addition,Bylund et al. (17) have demonstrated in a perfusedmouse lung model that the great majority of specificallybound '25I-hydroxybenzylpindolol is associated withtissue elements other than blood. In contrast, therelatively low uptake in the heart and other organs aswell as the lack of effect of propranolol pretreatmentmay be the result of a variety of factors. To someextent this probably reflects the relatively small frac-tion of the total cardiac output that these organs receiveas compared with the lung. The actual dose of 1251_hydroxybenzylpindolol administered is extremelysmall (-1 pmol) and thus the relatively high uptakein the lung as compared with the heart and otherorgans may reflect a bolus effect after intravenousadministration. Furthermore, Bylund et al. (17) reportedthat nonspecific binding in heart tissue was significantlyhigher (approximately fivefold) than in lung, evenwhen determined during in vitro binding experi-ments using crude membrane preparations. In par-ticuLlar, the high nonspecific uptake of 125I-hydroxy-benzylpindolol in the liver almost certainly reflects therole of this organ in metabolizing drigs of this nature.High first-pass extraction and metabolism of propranololand other beta blockers by the liver is a well-knownphenomenon.
Specific and nonspecific binding was also evaluatedby Bylund et al. (17) in mice by comparing the tissueconcentration of 125I-hydroxybenzylpindolol 5 min afterintravenous injection in control mice with that in micetreated with 1 mg/kg of propranolol. In those studies,specific binding was assessed by removing the tissuesand performing in vitro analyses of '251-hydroxy-benzylpindolol displacement by propranolol on crudeimembrane preparations. Using this approach, specificbinding in the brain averaged 66%, whereas in theheart this was 58% and in the lungs 92%. Their resultsalso indicate that as the amount of 1251-hydroxybenzyl-pindolol injected was increased, specific bindingdecreased. Furthermore, such a method suffers in thatsome loss of receptor-bound label will occur as thee(luilibrium is disturbed during the minutes requiredfor the preparation of even a crude membrane fraction.
In an attempt to keep nonspecific binding to a minimum,the amount of 125I-hydroxybenzylpindolol administeredto each animal in our studies was held constant at -1pmol (1-2 ,uCi). We have also observed that by deter-mining specific binding at earlier time points (5-10min) after injection, one can consistently inhibit 50%of total binding with propranolol pretreatment. Ourinitial experiments, as demonstrated in Fig. 1, con-sistently indicated that 40-50% of total binding wasspecific when determined at 15 min after '251-hydroxy-benzylpindolol administration. Later experimentsusing an earlier time point demonstrated a higherdegree of specific binding in the 50% range (Figs. 2and 4). The basis for this finding becomes more obviousafter examination of the in vivo binding curves inFig. 6. Most of the decrease in total binding over thefirst 10-20 min appears to represent depletion of thereceptor-b)ound ligand pool.The marked increase in specific binding in the lung
after chemical sympathectomy could have been theresult of an actual increase in the number of receptorsalthough physiologic data suggest that this may occurover a longer time period (10). In vitro stuidies havedocumented this phenomenon of upregulation followingeither chemical sympathectomy with 6-OH dopaminewhich produced a decrease in tissue norepinephrineconcentrations similar to ours (18), or after chronicreceptor blockade (19). Alternatively, in simply depletingendogenous agonists, a greater fraction of the receptorpopulation may have been available for '251-hydroxy-benzylpindolol binding. Recently, a functional parallelfor these observations in the lung was reported byJengo et al. (20) who showed that a greater degree ofsupersensitivity to exogenous catecholamines occturredin the pulmonary vascular bed than in the systemicbed after chemical sympathectomy. This led them topostulate that the lung has a greater concentration ofsympathetic nervous system innervation or of catechol-amine receptors than does the systemiiic vasculature.
Zeleznik and Roth (21) have recently detected in vivobinding of 1251-insulin to a high affinity receptor inrabbits. In results analogous to ours, administration oftinlabeled insulin before injection of tracer decreasedthe distribution space of the labeled insulin, con-sistent with inhibition of binding resulting from priorsaturation of available receptors with the unlabeledhormone. This process was dose dependent and satui-rable paralleling our observations with propranolol-mediated inhibition of '251-hydroxybenzylpindololbinding. Furthermore, by monitoring serum tracerlevels they were able to detect apparent reversal of1251-insulin binding to its receptor during infusion ofunlabeled insulin. Our findings utilizing externalimaging techniques to quantitate 1251-hydroxybenzyl-pindolol binding to pulmonary beta receptors are inline with their observations. The decrease in hydroxy-
1116 C. J. Homcy, H. W. Strauss, and S. Kopiwoda
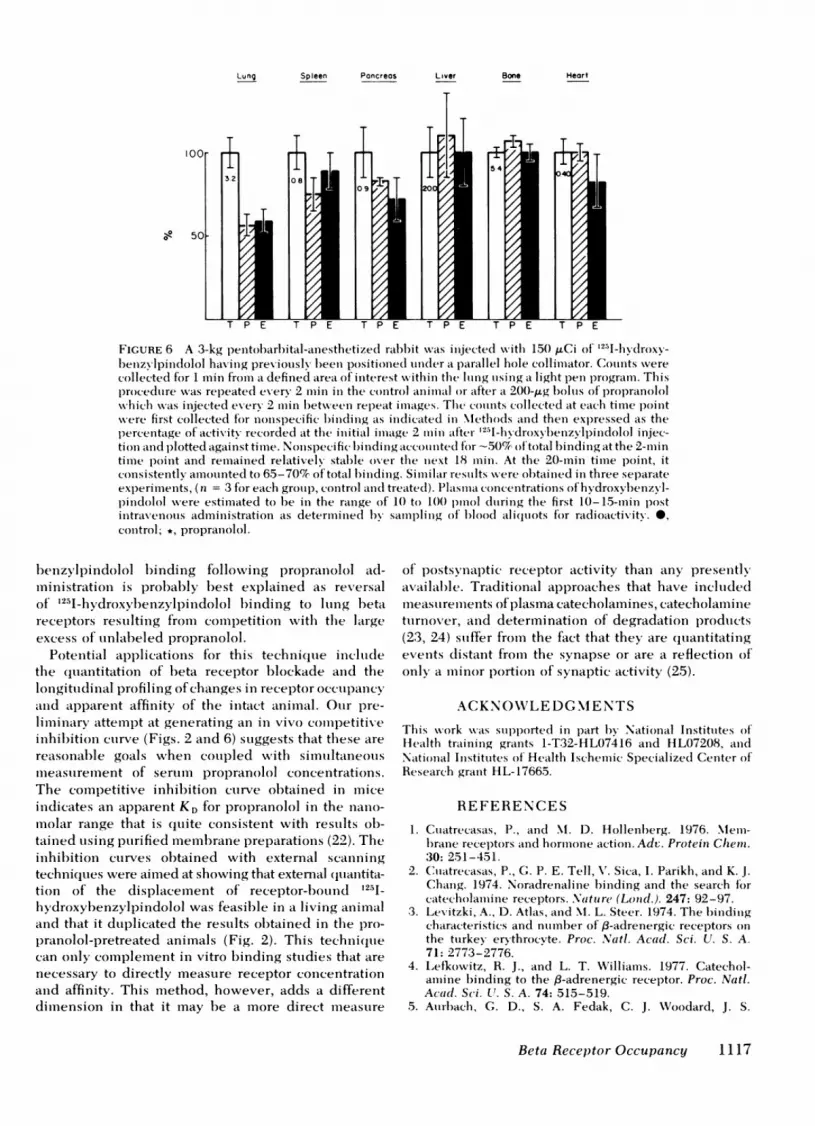
Lung Spleen Pancreas Liver
FIGURE 6 A 3-kg )enitol)arl)ital-ainestlietize(l ral)b)it was iiijected wvith 150 ,uCi of '251-hydroxy-Ibenzyvpincdolol having previously l)een positioinedl iunler a parallel hole collimiaitor. Couints wverecollected for 1 min fromil a defined area of interest within the Iling uising a lighit pen program. Thisproceduire w-cas repeate(d every 2 min in the control animall or after a 200-,ug bolus of propranololwhich was in-jected every 2 immin hetween rep)eat imiawges. The counts collected at eatchl tiimie pointwere first collected for nonispecific lbinding ais indlicated in MIethods and then expressed as thepercentage of activity recorded aIt the initial imiaige 2 iniln after 1251_11ydroxvbenzvlpindolol injec-tion aind plotted against time. Nonspecific lbinding accounted f'or -50% of'totail binding at the 2-mintime point and remainied relatively stahle over the next 18 min. At the 20-mim time point, itconsistently' amouintecl to 65-70% of total hinding. Siiliar res;ilts were ohtained in three separateexperiments, (n = 3 for eatch grouil), control and treatedl). Platsimaii concentrations of hydroxvl)enzN l-pindolol were estimated to be in the range of 10 to 100 pmol duiring the first 10-15-mmin postintravenous adminiistration as determi nedl 1y samilpling of' 1)lood ali (liots f'or radioactivity. *,conitrol; *, propraniolol.
benzylpindolol binding following propranolol ad-ministrationi is probably best explained as reversalof' i2I-hydroxybhenzylpindolol binding to luing betareceptors restiltinig from comI)etition wvith the latrgeexcess of unlabeled propranolol.
Potenitial applications for this techniiquie includlethe quiantitation of' beta receptor blockade and thelongitudinal profiling of changes in receptor occupancyanld apparenit affiinity of the intact animnal. Ouir pre-liminiary attemllpt at generating an in vivo comiipetitixeinhil)ition culrve (Figs. 2 and 6) suggests that these arereasonal)le goals wheni couipled wvith simuiltanieotismiieasturemnent of serum propranolol concentrations.The competitive inhibition cutrve obtained in mieeindicates an apparent KD for propranolol in the nianio-molar range that is quite conisistent with restults ob-tainied using ptirified mem-brane preparations (22). Theinhil)ition curves obtained with external scaniniiigtechnii(quies were aimed at showing that external (Ijuantita-tion of the displacement of receptor-bound 1251_hydroxybenzylpindolol was feasible in a living animaland that it duplicated the results obtained in the pro-I)ranolol-pretreated aniInals (Fig. 2). This techniq(uiecan only complemiient in vitro binding studies that arenecessary to directly measure receptor coneentrationand affinity. This method, however, adds a differentdimiension in that it may be a more direct measure
of postsynaptic receptor activity than any presentlyavailable. Traditional approaches that have incltudecdmeasuiremiients ofplasma catecholamines, catecholami neturnover, anid determination of degradation produlcts(23, 24) suiffer from the fact that they are (Ijuantitatinigevenits distant from the synapse or are a reflection ofonly a minor portioni of synaptic activity' (25).
ACKNOWL E DGMENTS
This work wNas supported in part hy Nationial Instituites ofHealth traininig granits 1-T32-HL07416 anid HL07208, andN'ationial Instituites of Health Isehemic Specializedl Center ofResearch granit HL-17665.
REFERENCES
1. Ctiatrecasas, P., and MI. D. Hollenberg. 1976. Mem-hrane receptors aind horimione action. Adv. Protein Chlevm.30: 251-451.
2. (Ctiatrecasas, P., G. P. E. Tell, V. Sica, I. Parikh, and K. J.Chang. 1974. Noradrenialinle binding and the search forcateeholamiine receptors. Natu re (Lotid.). 247: 92-97.
3. Levitzki, A., D. Atlas, and MI. L. Steer. 1974. The bindingcharacteristics and number of,8-adrenergic receptors onthe turkey erythrocyte. Proc. Natl. Acad. Sci. U. S. A.71: 2773-2776.
4. Lefkowitz, R. J., and L. T. Williams. 1977. Catechol-amine binding to the 8-adrenergic receptor. Proc. Natl.Acad. Sci. U. S. A. 74: 515-519.
5. Auirbach, G. D., S. A. Fedak, C. J. \Voodard, J. S.
Beta Receptor Occupancy 1117
Bon Heart

Hauser, and F. Troxler. 1974. j-adrenergic receptor:stereospecific interaction of iodinated 18-blocking agentwith high affinity site. Science (Wash. D. C.). 186:1223-1224.
6. Maguire, M. E., R. A. Wiklund, H. J. Anderson, and A. G.Gilman. 1976. Binding of 1251 iodohydroxybenzylpindololto putative ,8-adrenergic receptors of rat glioma cells andother cell clones. J. Biol. Chem. 251: 1221-1231.
7. Brown, E. A., S. A. Fedak, C. J. Woodard, G. D. Aurbach,and D. Rodbard. 1976. 1-adrenergic receptor interactions.Direct comparison of receptor interaction and biologicalactivity. J. Biol. Chem. 254: 1239-1246.
8. Thoenen, H. 1972. In Catecholamines. H. Blaschko andE. Mescholl, editors. Handb. Exp. Pharmakol. Springer-Verlag, Berlin. 33: 813.
9. Lungborg, P. 1969. Effect of reserpine on the subcellulardistribution of 3H-a-methylnoradenaline in the mouseheart. Br. J. Pharmacol. 36: 386-392.
10. Kostrzewa, R. M., and D. M. Jacobwitz. 1974. Pharmaco-logical actions of 6-hydroxydopamine. Pharmacol. Rev.26: 199-288.
11. Hoebeke, J., G. Vauquelin, and A. D. Strosberg. 1978.The production and characterization of antibodies againstthe ,B-adrenergic antagonists. Biochem. Pharmacol.27: 1527-1532.
12. Rockson, S. G., C. J. Homcy, and E. Haber. 1979. Anti-alprenolol antibodies: a probe for the study off-receptor-ligand interactions. Clin. Res. 27: 441A. (Abstr.)
13. DaPrada, M., and G. Zurcher. 1976. Simultaneousradioenzymatic determination of plasma and tissueadrenaline, noradrenaline, and dopamine levels withinthe femtomole range. Life Sci. 19: 1161-1174.
14. Roizen, M. F., J. Moss, D. P. Henry, V. Weise, and I. J.Kopin. 1978. Effect of general anesthetics on handlingand decapitation-induced increases in sympathoadrenaldischarge.J. Pharmacol. Exp. Ther. 204: 11-18.
15. Adam, K. R., C. G. Pullman, and P. C. Scholfield. 1973.Isoprenaline-and-exercise-induced tachycardia in theassessment of,8-adrenoceptor blocking drugs; a comparison
between tolamolol, practolol and propranolol. Br. J.Pharmacol. 49: 560-563.
16. Ansari, A. N. 1974. Myocardial imaging with 11C-norepinephrine. In Cardiovascular Nuclear Medicine.H. W. Strauss, B. Pitt, and A. E. James Jr., editors. TheC. V. Mosby Company, St. Louis, p. 234.
17. Bylund, D. B., M. E. Charness, and S. H. Synder. 1977.Beta adrenergic receptor labeling in intact animals with1251-hydroxybenzylpindolol. J. Pharmacol. Exp. Ther.201: 644-653.
18. Spom, J. R., T. K. Harden, B. B. Wolfe, and P. M. Molinoff.1976. /8-adrenergic receptor involvement in 6-hydroxy-dopamine-induced sensitivity in rat cerebral cortex.Science (Wash. D. C.). 194: 624-626.
19. Glaubiger, G., and R. J. Lefkowitz. 1977. Elevatedbeta-adrenergic receptor number after chronic propranololtreatment. Biochem. Biophys. Res. Commun. 78: 720-725.
20. Jengo, J. A., C. E. Juratsch, and M. M. Laks. 1978.Increased pulmonary to systemic vascular reactivity afterchemical sympathectomy (CS): denervation hyperreactivity.Circulation. 58(Suppl. II): 82. (Abstr.)
21. Zeleznik, A. J., and J. Roth. 1978. Demonstration of theinsulin receptor in vivo in rabbits and its possible role asa reservoir for the plasma hormone. J. Clin. Invest. 61:1363- 1374.
22. U'Pritchard, D. C., D. B. Bylund, and S. H. Synder. 1978.( + )-[3H]epinephrine and ( - )-[3H]dihydroalprenololbinding to f3 and 182-noradrenergic receptors in brain,heart, and lung membranes. J. Biol. Chem. 253: 5090-5102.
23. Axelrod, J. 1971. Noradrenaline: fate and control of itsbiosynthesis. Science (Wash. D. C.) 173: 598-606.
24. Landsberg, L. 1976. In The Year in Endocrinology,1975-1976. S. H. Ingbar, editor. Plenum Press, NewYork. 177.
25. Specter, S., J. Tarver, and B. Berkowitz. 1972. Effectsof drugs and physiologic factors in the disposition ofcatecholamines in blood vessels. Pharmacol. Rev. 24:191-202.
1118 C. J. Homcy, H. W. Strauss, and S. Kopiwoda




























