Embed Size (px)
Citation preview

Bb
QJ
a
ARRA
KAFMS
1
gp2Cmtetfibbmoec
0d
Applied Soil Ecology 47 (2011) 67–75
Contents lists available at ScienceDirect
Applied Soil Ecology
journa l homepage: www.e lsev ier .com/ locate /apsoi l
iocontrol of Fusarium wilt disease for Cucumis melo melon usingio-organic fertilizer
ingyun Zhao, Caixia Dong, Xingming Yang, Xinlan Mei, Wei Ran, Qirong Shen, Yangchun Xu ∗
iangsu Provincial Key Lab of Organic Solid Waste Utilization, Nanjing Agricultural University, Nanjing 210095, China
r t i c l e i n f o
rticle history:eceived 23 April 2010eceived in revised form 28 July 2010ccepted 29 September 2010
eywords:ntagonistic microbesusarium oxysporum f. sp. meloniselon
oil-borne disease
a b s t r a c t
Fusarium wilt disease in melon (Cucumis melo L.) is widespread, responsible for serious economic losses.Pot and field experiments were performed to investigate the effects of different bio-organic fertilizers(BIOs) made from organic fertilizer and different antagonistic microbes. BIOs decreased the incidence offusarium wilt disease and increased melon yield. The disease incidence of treatments with double appli-cation (BIOs applied both in the nursery and in the pot soil) was 20%, much lower than control (80%).Moreover, double application of a mix termed BIOII resulted in a 17.4-fold increase in plant fresh weightcompared to control. Application of BIOs strongly reduced the number of pathogen colony-formingunits (cfu) in stems and roots of melon. Pathogen populations were 2.27 × 103 and 6.67 × 103 cfu g−1 FW(fresh weight) on BIOII-treated stems and roots, respectively, and 8.17 × 104 and 3.67 × 104 cfu g−1 FWon control stems and roots, respectively; i.e., cfus were reduced by 97% and 82%, respectively. Microbialcommunity structure was ameliorated by all BIOs. The number of bacteria and actinomycota in rhizo-
sphere soil increased markedly under all BIO applications compared to control. In contrast, pathogenand fungal density was dramatically higher in the rhizosphere of control plants. The activities of defenseenzymes in the leaves of melons receiving double application of BIOII were lower than those of con-trol plants. The highest melon yield was achieved by plants that received a double application of BIOIIin field experiments (7225 kg ha−1 vs. 4447 kg ha−1 for control plants). In conclusion, the most effectivetreatment was double application of BIOII, which minimized the incidence of wilt disease, maximizedaltere
biomass production, and. Introduction
Melon (Cucumis melo L.) is one of the most important fruit cropsrown in China. It is susceptible to fusarium wilt disease whenlanted in the same field without rotation (Soriano-Martın et al.,006). An intensive farming practice aggravated wilt disease inhina. Melon fusarium wilt caused by Fusarium oxysporum f. sp.elonis Snyder and Hansen is a destructive vascular disease leading
o serious economic losses (Martyn and Amador, 1987; Champacot al., 1993). The pathogen can exist in soil for several years throughhe production of chlamydospores (Garret, 1970), and is thus dif-cult to control. Traditionally crop rotations have been proven toe an effective strategy to control many soil-borne diseases, butecause most of these pathogens (like Fusarium oxysporum f. sp.
elonis) can survive for a long period of time, the effectivenessf crop rotation is limited once a disease outbreak occurs (Kingt al., 2008). Grafting is a promising practice in soil-borne diseasesontrol in vegetable farming, particularly for watermelon in Japan,
∗ Corresponding author. Tel.: +86 25 8439 6824; fax: +86 25 8439 6824.E-mail address: [email protected] (Y. Xu).
929-1393/$ – see front matter © 2010 Elsevier B.V. All rights reserved.oi:10.1016/j.apsoil.2010.09.010
d microbial community structure.© 2010 Elsevier B.V. All rights reserved.
Korea, and Taiwan. However added cost still limits its feasibility toChinese farmers. Soil disinfection using various chemicals (Cebollaet al., 2000) or pesticides (Fravel et al., 2005) is also a traditionalpractice but must phase out because of environmental and humanhealth concerns (Brimner and Boland, 2003; Boutler et al., 2000).Soil solarization (Tamietti and Valentino, 2006) is an effective strat-egy too but is not readily applicable for intensive vegetable farmingsystems where time to solarize soil is very limited. Furthermore,soil solarization is often limited by local climate constrains.
Biological control was an alternative strategy to control fusar-ium wilt disease (De Cal, 1995; Larkin and Fravel, 1998). Manyantagonistic strains have been proved to be effective biocontrolagents in controlled laboratory or greenhouse conditions. Rhizoc-tonia (Muslim et al., 2003), Bacillus spp. (Gong et al., 2006; Hervaset al., 1998; Phae et al., 1990), Penicillium spp. (De Cal, 1995; DeCal et al., 1997, 2000; Sabuquillo et al., 2006; Larena et al., 2003),Aspergillus spp. (Suárez-Estrella et al., 2007; Kandhari et al., 2000),
Trichoderma spp. (Kaur and Mukhopadhyay, 1992; Cotxarrera et al.,2002; Tawfic and Allam, 2004; Rojo et al., 2007) are a few among thelong lists. Despite these efforts, effectiveness of biocontrol agentsis often site-specific. There has been no any biocontrol agent thatis universally effective everywhere, because of genetic diversity of
6 oil Ec
rttaoaW(plafci
mi(sds1tb
ftbt
2
2a
fC(siSi2idoftpifbroPpptTaoils
8 Q. Zhao et al. / Applied S
esponsible pathogens (Mishra et al., 2010). In addition, antagonis-ic strains must be combined with a suitable substrate to improveheir biocontrol efficacy because they can survive for a long timefter applied to soil (El-Hassan and Gowen, 2006). Compost notnly plays an important role in providing a suitable substrate, butlso serves as a growth-promoting medium (Raviv et al., 1998).ood shavings help reduce Fusarium wilt incidence in carnations
Cebolla and Pera, 1983). The microorganisms in compost mayroduce antibiotics (Whipps, 1997), compete for nutrients and eco-
ogical niches (Hoitink et al., 1997; Hoitink and Boehm, 1999),nd/or induce resistance in plants (Weller et al., 2002). There-ore developing a new bio-organic fertilizer with improved fieldonsistency is continuous process and mandatory for several andndividual ecological niches.
Malondialdehyde (MDA) content is considered as an index ofembranous lipid peroxidation (Morsya et al., 2007), and generally
ncreased under stressed conditions such as pathogenic infectionHeber et al., 1996). Besides MDA content, stress-related enzymesuch as peroxidases (POD), polyphenoloxides (PPO) and superoxideismutase (SOD) were induced and increased when the plants wereubjected to pathogenic attack (Zhao et al., 2005; Rossum et al.,997). Therefore, CAT, POD, PPO and SOD activities are usually usedo evaluate physiological and biochemical responses of plants toiotic and abiotic stresses (Gechev et al., 2003).
The objectives of this work were: (1) to develop a bio-organicertilizer for controlling fusarium wilt of melon (Cucumis melo L.)hat has been widespread in China, (2) to test the efficacies of theio-organic fertilizer in greenhouse as well as in field by two con-inuous cropping seasons.
. Materials and methods
.1. Isolation of Fusarium oxysporum f. sp. melonis andntagonistic microbes
The pathogen F. oxysporum f. sp. melonis (FOM) was isolatedrom tissue of a diseased melon plant (collected from Hexianounty, Anhui Province, China) using Fusarium-selective mediumKomada, 1975). The infected plant was washed three times withterile distilled water (SDW), cut into small pieces, homogenizedn a sterile muller and filtered. The filtered liquid was diluted withDW into appropriate dilution rates, spread onto a plate contain-ng Fusarium-selective medium (Komada, 1975) and incubated at8 ◦C for 4 days. A colony showing typical FOM morphologies was
solated and confirmed as the responsible pathogen by proceduresescribed in Koch’s postulation (by pot experiments). For isolationf antagonistic microorganisms, a rhizosphere soil was sampledrom a healthy melon plant, 10-fold series diluted, spread ontohe plates containing nutrient agar (NA, for bacteria), or onto thelates containing potato dextrose agar (PDA, for fungi), and were
ncubated at 28 ◦C for 4–6 days. Purified colonies were screenedor antagonists against FOM on plates using the method describedy Suárez-Estrella et al. (2001). For screening antagonist bacte-ia, a 8-mm plug from the leading edge of a 7-day-old culturef FOM cultured on PDA was placed onto the center of a newDA plate. The isolated bacteria were inoculated adjacent to thelug, and further incubated at 28 ◦C. For screening antagonist fungi,lugs taken from the perspective fungi were inoculated adjacento the centered FOM plug and were further incubated at 28 ◦C.wo bacteria and two fungi showing the strongest antagonisms
gainst FOM (Fig. 1) were selected as antagonistic agents to fortifyrganic fertilizer. The 16S rRNA gene sequences of one bacterialsolate, designated as y10, shows 99% identity to Paenibacil-us polymyxa strain ISSDS-851 as BLASTed against all nucleotideequences in NCBI database. The other isolate, designated as yIV1,ology 47 (2011) 67–75
shows 100% identity to Bacillus subtilis strain L4. Sequences ofboth strains were deposited into GenBank under accession noGQ849013 and GQ475486, respectively. Two fungi isolates, des-ignated as P1 and A1, respectively, were tentatively identified asPenicillium sp. and Aspergillus sp., respectively, according to theirmorphologies.
2.2. Preparation of bio-organic fertilizer
Spore suspensions of two selected antagonistic fungi P1 and A1were used as inocula to fortify organic fertilizer. To produce sporesuspensions, a 6 mm-diam disc obtained from the periphery of thecolony, in areas of active growth were separately inoculated into a1000-ml flask containing 200 ml potato dextrose broth (PDB) andincubated in a shaker at 180 rpm at 28 ◦C for 48 h. The concentrationof the spore in the suspension was determined using a hemacy-tometer, and adjusted to final concentration of 105 spores per ml bydilution with sterile water. Cell suspension of two selected antag-onistic bacteria were produced separately by inoculating into a1000-ml flask containing 200 ml beef extract peptone broth andshaken at 180 rpm at 34 ◦C for 24 h. The final concentration of bac-terial cells was 109 cfu ml−1. Suspensions produced as such wereused to fortify organic fertilizer.
The spore suspensions of two selected antagonistic fungi P1 andA1 were separately sprayed onto a pile of organic fertilizer (com-mercially purchased from Jiangsu Tianniang Ltd., China) at the rateof 100 ml suspension kg−1 organic fertilizer. Cell suspensions of y10and yIV1 were separately sprayed onto another pile of organic fer-tilizer at the same rate. The two piles of organic fertilizer fortifiedas such were combined into one and mixed thoroughly. This prod-uct was coded as BIOI. Another fortified organic fertilizer, codedas BIOII, was formulated by mixing BIOI with a SQR21-fortifiedproduct at 1:1 ratio (w/w), later of which was kindly provided byLing (for details, ref to Ling et al., in press). Both BIOI and BIOIIwere used for later pot and field studies. In field experiments,the SQR21-fortified product was also included and was designatedhereafter in this paper as BIOIII. The original organic fertilizer(non-fortified) commercially purchased from Jiangsu TianniangLtd., China, designated as OF, was used in later pot experiments aswell.
2.3. Preparation of pathogen inocula
FOM was grown in PDA plates at 28 ◦C for 1 week till sporeswere densely produced. The spore suspension was obtained byadding 15 ml SDW into each plate, scraped, collected into a tube,and filtered through eight layers of sterile gauze to remove solids.Spore concentration was determined using a hemacytometer, andadjusted to 5 × 106 cfu ml−1 by dilution with SDW. This spore sus-pension was stored at 4 ◦C until use.
2.4. Pot experiment design
Two soils were used in this study. One soil collected from a fieldthat had never been planted with melon (considered as healthy soil)was used for nursery growth. Another soil, having been previouslygrown three seasons of melon successively and consequently beingseriously infested with FOM was used for pot experiments. FOMspore concentration in this soil was 5.3 × 104 cfu g−1 fresh soil andwas thus considered as infested soil. The soils were passed througha 6-mm sieve before put into pots.
Melon seeds (Baisha Mi) were surface-sterilized with H2O2 (5%)for 30 min, washed with SDW three times, placed in Petri dishescontaining wet filter paper for germination at 30 ◦C for 30 h. Thegerminated seeds were moved into plates each containing 300 ghealthy soils. By 4-leaf age (∼25 days growth), the seedlings were

Q. Zhao et al. / Applied Soil Ecology 47 (2011) 67–75 69
F rum f.( illus p
twg
eOtBe(BAicssubewt
2
ppan((
ig. 1. The antagonistic activities were tested by co-culture FOM (Fusarium oxyspoB) FOM co-culture with P1 (Penicillium sp.). (C) FOM co-culture with y10 (Paenibac
ransplanted to pots. Each pot containing 12 kg of infested soilsas transplanted with one seedling. The plants were grown in a
reenhouse.Treatments applied to both nursery and pot soils. In the nurs-
ry phase, soils were treated with either none (CKn), or 2% (w/w)F (OFn), or 2% (w/w) BIOI (BIOIn), or 2% (w/w) BIOII (BIOIIn). In
he pot phase, soils were treated with none (CKp), or 0.5% (w/w)IOII (BIOIIp). Combination of two phased treatments resulted inight treatments, expressed as nursery treatment + pot treatment:1) CKn + CKp; (2) CKn + BIOIIp; (3) OFn + CKp; (4) OFn + BIOIIp; (5)IOIn + CKp; (6) BIOIn + BIOIIp; (7) BIOIIn + CKp; (8) BIOIIn + BIOIIp.ll these amendments were thoroughly mixed with soils before put
nto nursery plates or pots. Urea, superphosphate and potassiumhloride was also applied to each pot at the rate of 0.15 g N kg−1
oil, 0.05 g P kg−1 soil and 0.10 g K kg−1 soil, respectively. To ensureufficiently high density of FOM population, all pot soils were inoc-lated with FOM spores at 105 cfu g−1 soil before put into pots. Twolocks were randomly laid out to replicate the eight treatments andach treatment with ten replicates (pots) was randomly arrangedithin each block. This pot experiment was carried out from July
o September in 2008.
.5. Field experimental design
Field experiments were carried out in Xinyi city, Jiangsurovince, China. Plots, each 7 m × 3 m in size, planted with 45
lants, were randomly distributed. Field treatments, expresseds nursery treatment + field treatment, included: (1) CKn + CKf:ursery and field soils treated with chemical fertilizer (control);2) BIOIn + CKf: BIOI-treated nursery soil, untreated field soil;3) BIOIIIn + CKf: BIOIII-treated nursery soil, untreated field soil;sp. melonis) and antagonistic strains. (A) FOM co-culture with A1 (Aspergiluls sp.).olymyxa). (D) FOM co-culture with IVI (Bacillus subtilis).
(4) BIOIIn + CKf: BIOII-treated nursery soil, untreated field soil;(5) CKn + BIOIf: untreated nursery soil, BIOI-treated field soil; (6)BIOIn + BIOIf: BIOI-treated nursery and field soils; (7) CKn + BIOIIIf:untreated nursery soil, BIOIII-treated field soil; (8) BIOIIIn + BIOIIIf:BIOIII-treated nursery and field soils; (9) CKn + BIOIIf: untreatednursery soil, BIOII-treated field and (10) BIOIIn + BIOIIf: BIOII-treated nursery and field soils. The application rate of bio-organicfertilizer was 2% (w/w) in nursery soils and was 1000 kg ha−1 in fieldplots. All plots received an application of 180 kg N ha−1 (as urea),40 kg P ha−1 (as superphosphate) and 120 kg K ha−1 (as muriate ofpotash). Two blocks were randomly laid out and each treatmentwith three plots was randomly arranged within each block.
2.6. Sampling and analytical methods
Thirty-four days after transplanting, pot soils and plants weredestructively sampled. Three healthy plants in each treatment weresampled to measure plant fresh weight. Plant fresh weight (includ-ing roots) was determined by washing out soils on roots with tapwater and the water in excess was absorbed with filter papers.The rest plants in each treatment were also sampled to determineother parameters. Shoots and roots were separated for enumera-tion of FOM populations in the tissues. Plant leaves were frozenin liquid nitrogen and kept at −20 ◦C for later measurement of theactivities of polyphenol oxidase (PPO), peroxidase (POD), superox-ide dismutase (SOD), and malondialdehyde (MDA) concentration.
Roots were taken from the pots and carefully shaken by hand toremove soil; these soil samples represented rhizosphere soils. Soilthat still adhered to roots, and the roots themselves, were consid-ered rhizoplane samples. The big pot soil that free of roots werealso collected and termed bulk soils.
7 oil Ec
dwmw1tt0(rmSstfisbi
owa2tt5M
d1aibbT2a
incidences (percent of wilted plants over total plants) estimated
FpteB
0 Q. Zhao et al. / Applied S
The microorganisms were enumerated by the standard 10-foldilution method. Soil suspensions at appropriate dilution ratesere inoculated onto plates containing appropriate media. Agaredia for growing bacteria, actinomycota, fungi, and Fusariumere beef extract peptone, Gause NO.1 (soluble starch 20 g, KNO3g, NaCl 0.5 g, K2HPO4 0.5 g, MgSO4 0.5 g, FeSO4 0.01 g, agar 20 g,
ap water 1000 ml, pH 7.2), Martin medium (glucose 10 g, pep-one 5 g, KH2PO4 1 g, MgSO4 0.5 g, rose bengal 30 �g, streptomycin.03 g, distilled water 1000 ml), and Fusarium-selective mediumKomada, 1975), respectively. The FOM populations in shoots andoots were measured as described by Muslim et al. (2003) withodifications. Briefly, the shoots and roots were washed with
DW three times and homogenized using a juice extractor in 1:9hoot/root:water ratio (w/v). The homogenized liquid was filteredhrough two layers of sterile gauze. Residues were discarded. Theltered liquid was diluted 10–1000 times. Aliquots of 0.1 ml werepread onto Fusarium-selective medium (Komada, 1975), incu-ated at 28 ◦C for 4–6 days. All measurements were based on 3
ndependent samples (3 replicates).MDA concentration was determined according to the meth-
ds described by Hartman et al. (2004). Fresh leaves in 0.5 gere homogenized in a solution containing 0.6% thiobarbituric
cid (TBA) and 5% trichloroacetic acid (TCA), heated at 100 ◦C for5 min and centrifuged at 8000 × g for 10 min after cooled to roomemperature. MDA concentrations were measured with a spec-rophotometer based on the differences in absorbance at 600,32, and 450 nm. The values were calculated from the formula:DA = [6.45 (A532 − A600) − 0.56 A450]/leaf fresh weight.SOD enzyme activity in leaves was measured by the procedures
escribed by Garcia-Limones et al. (2002). Fresh melon leaves ing were homogenized in 5 ml 50 mM phosphate buffer (pH 7.8)nd centrifuged at 10,000 × g for 10 min at 4 ◦C. The supernatantn 0.05 ml was added into a mixture containing 25 mM phosphateuffer (pH 7.8), 13 mM l-methionine (Met), 75 �M nitrotetrazolium
lue tetrazolium (NBT), 10 �M EDTA-Na2, and 2.0 �M riboflavin.he mixture was placed into a light incubator, incubated at 25 ◦C for0 min under 54 �mol m−2 s−1 light intensity, and measured withspectrophotometer at 560 nm. One unit of SOD activity equals theig. 2. Effect of different treatments on (A) plant Fusarium wilt disease incidence and (B)ot soil unamended, organic fertilizer, BIOI and BIOII, but the soil in the big pots was unamo nursery pot soil unamended, organic fertilizer, BIOI and BIOII while the soil in all big pach time with 10 plants per treatment. Data were subjected to Duncan’s ANOVA test. Baars with different letters indicate significant differences (p = 0.05).
ology 47 (2011) 67–75
amount of enzyme that inhibits the rate of NBT reduction by 50%under these conditions.
POD activity was determined following the methods of Nakanoand Asada (1981). The supernatant in 0.1 ml, same as above, wasadded into a mixture containing 50 mM phosphate buffer (pH7.8), 50 mM guaiacol, and 10 mM H2O2. The reaction product wasmeasured spectrophotometrically at 470 nm. One unit of enzymeactivity was designed as the change in absorbance of 0.01 for 1 gfresh weight per minute.
PPO activity was measured following Tao et al. (2007) withslight modifications. Fresh leaves in 1.0 g was homogenized in 10 ml100 mM citric acid phosphate buffer (pH 6.0) containing 0.20 gpolyethylene glycol and centrifuged at 10,000 × g for 10 min at 4 ◦C.The supernatant in 0.6 ml was added into a mixture containing 3 mlcitric acid phosphate buffer and 0.02 mol l−1 catechol. The reactionproduct was measured spectrophotometrically at 420 nm for 5 min.One unit of PPO activity equals the change in absorbance at 420 nmfor 1 g fresh weight per minute.
2.7. Data analysis
Data obtained were analyzed statistically using SPSS 16.0program. Data were subjected to Duncan’s analysis of variance(ANOVA) and means were separated by Duncan’s multiple rangetests at p ≤ 0.05.
3. Results
3.1. Effect of different BIO applications on fusarium wilt diseaseincidence and plant biomass production
The symptoms of melon wilt appeared 14 days after trans-planting into the pots (containing FOM-infested soil). The disease
at 34 days after transplanting, were 80% and 70% in the CKn + CKpand OFn + CKp treatments and 20% and 30% in the BIOIn + CKp andBIOIIn + CKp treatments, respectively (Fig. 2A). In the treatmentsthat seedlings received BIOI or BIOII amendments (BIOIn + BIOIIp,
plant fresh weight. CKn + CKp, OFn + CKp, BIOIn + CKp, BIOIIn + CKp refer to nurseryended, respectively; CKn + BIOIIp, OFn + BIOIIp, BIOIn + BIOIIp, BIOIIn + BIOIIp referots was amended with BIOII, respectively. The experiment was carried out twice,rs are standard errors according to Duncan’s least significant difference at p = 0.05.

oil Ecology 47 (2011) 67–75 71
B2nOBoOofa
3
BhTrn
BBtdtdaa
3
Bl6la
B
Table 1Effects of application of BIOs on microorganism density (cfu g−1 soil) in rhizoplanesoil.
Treatments Bacteria (×109) Actinomycota (×107) Fungi (×105)
CKn + CKp 1.5 ± 0.35d 3.7 ± 0.58c 18.0 ± 2.68bCKn + BIOIIp 6.7 ± 1.15c 3.6 ± 1.04c 40.0 ± 10.15aOFn + CKp 2.4 ± 0.40d 8.4 ± 1.25b 17.0 ± 2.56bcOFn + BIOIIp 16.0 ± 2.64ab 4.1 ± 0.26c 10.3 ± 0.87cdBIOIn + CKp 6.9 ± 2.27c 14.7 ± 2.52a 5.0 ± 1.00dBIOIn + BIOIIp 17.6 ± 1.70a 11.6 ± 3.76ab 5.4 ± 1.19dBIOIIn + CKp 8.1 ± 2.95c 11.4 ± 1.51ab 5.3 ± 0.64dBIOIIn + BIOIIp 14.0 ± 1.95b 11.0 ± 3.72ab 6.3 ± 1.06d
Note: CKn + CKp, OFn + CKp, BIOIn + CKp, BIOIIn + CKp refer to treatments with nurs-ery pot soil unamended, organic fertilizer, BIOI and BIOII but all big pot soilunamended, respectively. CKn + BIOIIp, OFn + BIOIIp, BIOIn + BIOIIp, BIOIIn + BIOIIp
Frrt
Q. Zhao et al. / Applied S
IOIIn + BIOIIp and BIOIn + CKp), disease incidences were about0%, obviously lower than those treatments that seedlings didot receive BIOI or BIOII amendments (CKn + CKp, CKn + BIOIIp,Fn + CKp and OFn + BIOIIp). Plant fresh weights in treatments ofIOIn + CKp, BIOIIn + CKp, BIOIn + BIOIIp, and BIOIIn + BIOIIp werebviously higher than those in treatments of CKn + CKp andFn + CKp (10.4–32.2 g) (Fig. 2B). This indicates that applying bio-rganic fertilizers to nursery pots can significantly increase plantresh weight; moreover, double application of BIOII in the nurserynd field phases had a stronger effect than a single application.
.2. FOM in shoots and roots
FOM populations in shoots in treatments of BIOIn + CKp,IOIn + BIOIIp, BIOIIn + CKp, BIOIIn + BIOIIp, and CKn + CKp wereighly decreased compared with control (CKn + CKp) (Fig. 3A).hese four treatments decreased FOM by 84%, 97%, 98%, and 97%,espectively. This demonstrates that applying BIOI or BIOII can sig-ificantly reduce FOM in shoots.
FOM populations in roots showed a similar trend (Fig. 3A and). FOM populations in treatments of BIOIn + CKp, BIOIIn + CKp,IOIn + BIOIIp, and BIOIIn + BIOIIp were lower compared to con-rol (CKn + CKp), these four treatments decreased FOM populationensity by 67%, 60%, 71%, and 82%, respectively. These data showhat nursery application or double application of BIOs significantlyecreased the number of FOM in shoots and roots, whereas BIOIIpplied only to pot soil had no significant effect on FOM populations compared to control.
.3. FOM isolated from the rhizoplane, rhizosphere, and bulk soil
The FOM populations in rhizoplane soil in treatments ofIOIn + CKp, BIOIIn + CKp, and BIOIIn + BIOIIp were significantly
ower compared with control (Fig. 4A), resulting in 81%, 77.6% and
4.8% decrease, respectively. A significant decrease of FOM popu-ation was observed in the treatments of BIOIn + CKp, BIOIIn + CKp,nd BIOIIn + BIOIIp, but not in treatment BIOIn + BIOIIp (Fig. 4A).
The FOM population in rhizosphere soil in treatments ofIOIn + CKp and BIOIIn + CKp did not differ significantly from the
ig. 3. Pathogen populations isolated from (A) plant fresh shoots, and (B) plant fresh roots. Tefer to nursery pot soil unamended, organic fertilizer, BIOI and BIOII, but the big pot soil waefer to nursery pot soil unamended, organic fertilizer, BIOI and BIOII while all the big potest. Bars show standard errors of the means. Columns with different letters are statistica
refer to treatments with nursery pot soil unamended, organic fertilizer, BIOI andBIOII whereas all big pot soil were amended with BIOII. Data were expressed asmean ± standard error. The data in a column with a different letter differ significantlyat Duncan’s significance level 0.05.
control. However, the populations in treatments of BIOIn + BIOIIpand BIOIIn + BIOIIp differed significantly from that of CKn + BIOIIp,respectively, resulted in 63% and 94% decrease (Fig. 4B). FOM pop-ulations in bulk soils were insignificantly different among all thesetreatments (data not shown).
3.4. Total microbial counts after application of BIOs
Total bacterial counts in the rhizoplane soils of treatmentsBIOIIn + CKp and BIOIn + CKp were 4.4 and 3.6 times increases com-pared to control. Conversely, total fungal counts in the rhizoplanesoils of treatments BIOIIn + CKp and BIOIn + CKp decreased by 70%and 72% as compared to that of control, respectively. In addition,double application of BIOs also increased total bacterial counts ascompared to nursery-only application (Table 1).
The changes in microbial counts in the rhizosphere soil showed
similar trends (Tables 1 and 2), although the values were gener-ally lower than those in the rhizoplane soils. For example, the totalcounts of bacteria and actinomycota in treatment BIOIIn + BIOIIpwere 170% and 72% lower in the rhizosphere soil than those inrhizoplane soil (Table 2).he cfu refer to colony-forming units. CKn + CKp, OFn + CKp, BIOIn + CKp, BIOIIn + CKps unamended, respectively. CKn + BIOIIp, OFn + BIOIIp, BIOIn + BIOIIp, BIOIIn + BIOIIp
soil was amended with BIOII, respectively. Data were subjected to Duncan’s ANOVAlly different at p = 0.05.
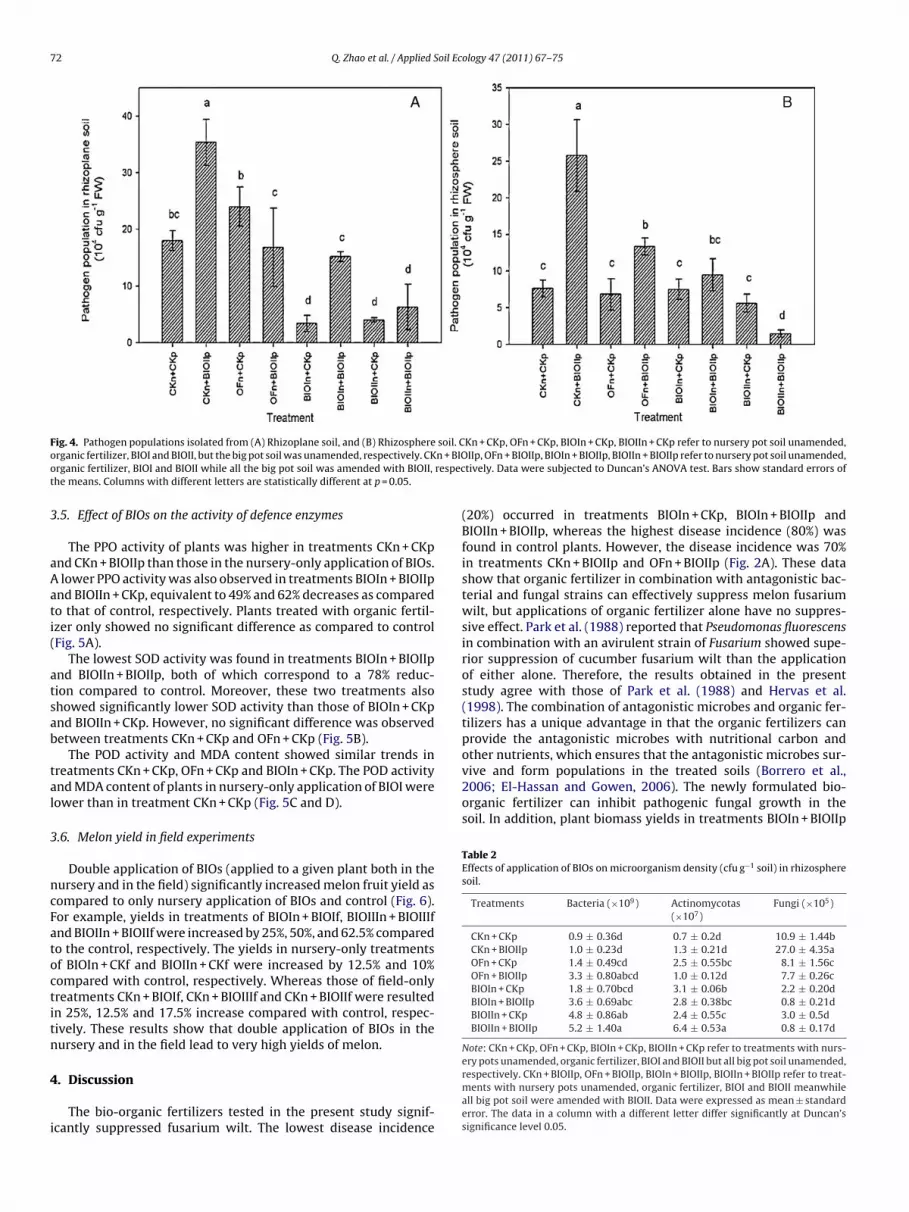
72 Q. Zhao et al. / Applied Soil Ecology 47 (2011) 67–75
F soil. Co n + BIOo respect
3
aAati(
atsab
tal
3
ncFatoctitn
4
i
other nutrients, which ensures that the antagonistic microbes sur-vive and form populations in the treated soils (Borrero et al.,2006; El-Hassan and Gowen, 2006). The newly formulated bio-organic fertilizer can inhibit pathogenic fungal growth in thesoil. In addition, plant biomass yields in treatments BIOIn + BIOIIp
Table 2Effects of application of BIOs on microorganism density (cfu g−1 soil) in rhizospheresoil.
Treatments Bacteria (×109) Actinomycotas(×107)
Fungi (×105)
CKn + CKp 0.9 ± 0.36d 0.7 ± 0.2d 10.9 ± 1.44bCKn + BIOIIp 1.0 ± 0.23d 1.3 ± 0.21d 27.0 ± 4.35aOFn + CKp 1.4 ± 0.49cd 2.5 ± 0.55bc 8.1 ± 1.56cOFn + BIOIIp 3.3 ± 0.80abcd 1.0 ± 0.12d 7.7 ± 0.26cBIOIn + CKp 1.8 ± 0.70bcd 3.1 ± 0.06b 2.2 ± 0.20dBIOIn + BIOIIp 3.6 ± 0.69abc 2.8 ± 0.38bc 0.8 ± 0.21dBIOIIn + CKp 4.8 ± 0.86ab 2.4 ± 0.55c 3.0 ± 0.5dBIOIIn + BIOIIp 5.2 ± 1.40a 6.4 ± 0.53a 0.8 ± 0.17d
Note: CKn + CKp, OFn + CKp, BIOIn + CKp, BIOIIn + CKp refer to treatments with nurs-
ig. 4. Pathogen populations isolated from (A) Rhizoplane soil, and (B) Rhizosphererganic fertilizer, BIOI and BIOII, but the big pot soil was unamended, respectively. CKrganic fertilizer, BIOI and BIOII while all the big pot soil was amended with BIOII,he means. Columns with different letters are statistically different at p = 0.05.
.5. Effect of BIOs on the activity of defence enzymes
The PPO activity of plants was higher in treatments CKn + CKpnd CKn + BIOIIp than those in the nursery-only application of BIOs.lower PPO activity was also observed in treatments BIOIn + BIOIIp
nd BIOIIn + CKp, equivalent to 49% and 62% decreases as comparedo that of control, respectively. Plants treated with organic fertil-zer only showed no significant difference as compared to controlFig. 5A).
The lowest SOD activity was found in treatments BIOIn + BIOIIpnd BIOIIn + BIOIIp, both of which correspond to a 78% reduc-ion compared to control. Moreover, these two treatments alsohowed significantly lower SOD activity than those of BIOIn + CKpnd BIOIIn + CKp. However, no significant difference was observedetween treatments CKn + CKp and OFn + CKp (Fig. 5B).
The POD activity and MDA content showed similar trends inreatments CKn + CKp, OFn + CKp and BIOIn + CKp. The POD activitynd MDA content of plants in nursery-only application of BIOI wereower than in treatment CKn + CKp (Fig. 5C and D).
.6. Melon yield in field experiments
Double application of BIOs (applied to a given plant both in theursery and in the field) significantly increased melon fruit yield asompared to only nursery application of BIOs and control (Fig. 6).or example, yields in treatments of BIOIn + BIOIf, BIOIIIn + BIOIIIfnd BIOIIn + BIOIIf were increased by 25%, 50%, and 62.5% comparedo the control, respectively. The yields in nursery-only treatmentsf BIOIn + CKf and BIOIIn + CKf were increased by 12.5% and 10%ompared with control, respectively. Whereas those of field-onlyreatments CKn + BIOIf, CKn + BIOIIIf and CKn + BIOIIf were resultedn 25%, 12.5% and 17.5% increase compared with control, respec-ively. These results show that double application of BIOs in theursery and in the field lead to very high yields of melon.
. Discussion
The bio-organic fertilizers tested in the present study signif-cantly suppressed fusarium wilt. The lowest disease incidence
Kn + CKp, OFn + CKp, BIOIn + CKp, BIOIIn + CKp refer to nursery pot soil unamended,IIp, OFn + BIOIIp, BIOIn + BIOIIp, BIOIIn + BIOIIp refer to nursery pot soil unamended,tively. Data were subjected to Duncan’s ANOVA test. Bars show standard errors of
(20%) occurred in treatments BIOIn + CKp, BIOIn + BIOIIp andBIOIIn + BIOIIp, whereas the highest disease incidence (80%) wasfound in control plants. However, the disease incidence was 70%in treatments CKn + BIOIIp and OFn + BIOIIp (Fig. 2A). These datashow that organic fertilizer in combination with antagonistic bac-terial and fungal strains can effectively suppress melon fusariumwilt, but applications of organic fertilizer alone have no suppres-sive effect. Park et al. (1988) reported that Pseudomonas fluorescensin combination with an avirulent strain of Fusarium showed supe-rior suppression of cucumber fusarium wilt than the applicationof either alone. Therefore, the results obtained in the presentstudy agree with those of Park et al. (1988) and Hervas et al.(1998). The combination of antagonistic microbes and organic fer-tilizers has a unique advantage in that the organic fertilizers canprovide the antagonistic microbes with nutritional carbon and
ery pots unamended, organic fertilizer, BIOI and BIOII but all big pot soil unamended,respectively. CKn + BIOIIp, OFn + BIOIIp, BIOIn + BIOIIp, BIOIIn + BIOIIp refer to treat-ments with nursery pots unamended, organic fertilizer, BIOI and BIOII meanwhileall big pot soil were amended with BIOII. Data were expressed as mean ± standarderror. The data in a column with a different letter differ significantly at Duncan’ssignificance level 0.05.

Q. Zhao et al. / Applied Soil Ecology 47 (2011) 67–75 73
Fig. 5. Activities of defense-related enzyme and concentrations of malondialdehyde measured in leaves. (A) Polyphenoloxidase (PPO), (B) Superoxide dismutase (SOD), (C)Peroxidase (POD), (D) Malondialdehyde (MDA). CKn + CKp, OFn + CKp, BIOIn + CKp, BIOIIn + CKp refer to nursery pot soil unamended, organic fertilizer, BIOI and BIOII, butt IIp, Bw to Dul
aBwnyBwct
camniMuldB(
he big pot soil was unamended, respectively. CKn + BIOIIp, OFn + BIOIIp, BIOIn + BIOhile all the big pot soil was amended with BIOII, respectively. Data were subjected
etters are statistically different at p = 0.05.
nd BIOIIn + BIOIIp were higher than those of BIOIn + CKp andIOIIn + CKp, demonstrating that BIOs not only reduced fusariumilt, but also promoted plant growth. BIOs applied to both theursery and the field phases significantly increased melon fruitield compared to single applications (except for CKn + BIOIf andIOIn + BIOIf) (Fig. 6). The highest yield was found in treatmentsith double applications of BIOII. This is possibly because the BIOs
ontain biologically active substances such as amino acid and pep-ides, which stimulate the growth of plant roots (Zhang et al., 2008).
The pathogen densities in plant shoots and roots were signifi-antly lower in treatments BIOIn + CKp, BIOIIn + CKp, BIOIn + BIOIIp,nd BIOIIn + BIOIIp compared to the control and CKn + BIOIIp treat-ents (Fig. 3A and B). This implies that application of BIOs in the
ursery phase could significantly inhibit the invasion of pathogensnto plant roots and shoots. These results support the report by
uslim et al. (2003), who indicated that tomato seedlings inoc-
lated with hypovirulent binucleate Rhizoctonia had significantlyower pathogen densities for all diseases studied. The pathogenensities in the rhizoplane soil significantly decreased in allIOs treatments except BIOIn + BIOIIp, compared to control plantsFig. 4A). Furthermore, melon wilt incidence was closely related
IOIIn + BIOIIp refer to nursery pot soil unamended, organic fertilizer, BIOI and BIOIIncan’s ANOVA test. Bars show standard errors of the means. Columns with different
to FOM densities in plant shoots (R2 = 0.9535), roots (R2 = 0.9542),and the rhizoplane soil (R2 = 0.7191). For example, when thedisease incidence was 70% in treatment OFn + BIOIIp (Fig. 2A),the corresponding FOM densities were 3.77 × 104, 1.7 × 104, and1.68 × 105 cfu g−1 in plant shoots, roots, and the rhizoplane soils,respectively (Figs. 3A, B and 4A). In contrast, when the diseaseincidence was 20% in treatment BIOIIn + BIOIIp (Fig. 2A), the FOMdensities were 2.27 × 103, 6.67 × 103, and 6.3 × 104 cfu g−1 in plantshoots, roots, and rhizoplane soil, respectively (Figs. 3A, B and 4A).These results suggest that plants are more easily attacked by wiltdisease when the pathogen densities are more than 104 cfu g−1 inshoots and roots or more than 105 cfu g−1 in the rhizoplane soil,whereas disease is much less severe when pathogen densities areless than 103 cfu g−1 in shoots and roots or lower than 104 cfu g−1
in the rhizoplane soils. Pathogen densities were insignificantly dif-ferent among any treated rhizosphere soil except BIOIIn + BIOIIp,
compared to that of control (Fig. 4B). This indicates that, althoughthe pathogen could survive in soil treated with antagonists, it wasunable to attack the melon roots. De Cal (1995) and Bark (1981)found that biological control via competition has little or no effecton the viability of pathogens in soil; however, in the present study,
74 Q. Zhao et al. / Applied Soil Ec
Fig. 6. Effect of different treatments on melon fruit yield. The first term refers totreatment in the nursery (CKn, BIOIn, BIOIIIn or BIOIIn) and the second term referstDsd
clob(
r(t(pzgbahIciCziscrmn
ptrsiwtrdpdp
o the treatment in the field (CKf, BIOIf, BIOIIIf or BIOIIf). Data were subjected touncan’s ANOVA test. Bars indicate standard errors according to Duncan’s least
ignificant difference at p = 0.05. Columns with a different letter were significantlyifferent.
ompetition among the antagonistic strains Aspergillus spp, Penicil-ium spp, and B. subtilis may have played protective roles against F.xysporum f. sp. melonis. In addition, antagonistic strains of Paeni-acillus polymyxa also show effective inhibition against pathogensLing et al., in press).
BIOs application altered microbial community structure asevealed by plate count method (Tables 1 and 2). Borrero et al.2004) reported that microbial activity in soil plays an impor-ant role in suppressing tomato fusarium wilt. Sturz and Christie2003) confirmed that an optimum microbial community canromote soil defence capability. Dommergues (1978) classified rhi-osphere microorganisms into beneficial, harmful, or ineffectiveroups based on their actions on plant growth. Many strains ofacteria and actinomycota can cause plant disease but more dam-ges are caused by fungi (Brussaard et al., 2007). Some bacteriaave the potential for pathogen suppression (Boulter et al., 2002).
n the present study, the populations of bacteria and actinomy-ota in both the rhizosphere and the rhizoplane soils dramaticallyncreased after nursery application of BIOs as compared to control.onversely, fungal populations in both the rhizoplane and the rhi-osphere soil were markedly lower after treatments with BIOs thann control soil (Tables 1 and 2). Bacterial densities in the rhizoplaneoil of treatments BIOIn + BIOIIp and BIOIIn + BIOIIp were dramati-ally higher than those of BIOIn + CKp and BIOIIn + CKp. The presentesults demonstrate that BIOs treatments can alter microbial com-unity structure. Boulter et al. (2002) also reported that higher
umbers of microorganisms in compost can improve plant health.The activities of plant defence enzymes generally increase when
lants are subjected to injury, pathogens, or extreme tempera-ures (Chen et al., 2000; Garcia-Limones et al., 2002). MDA contentepresents the degree of damage to plant cell membranes undertress (Morsya et al., 2007). Enzymes such as SOD and POD arenvolved in eliminating reactive oxygen species (Asada, 1992),
hereas PPO may be induced to produce lignin and other phenolicso strengthen cell walls (Avdiushko et al., 1993). Wu et al. (2008)
eported that the activities of defence-related enzymes in leaves areirectly related to the resistance of the plant to biotic stress. In theresent study, MDA content and the activities of SOD, PPO, and PODecreased significantly after BIOs treatments compared to controllants and treatment OFn + CKp (Fig. 5A–D). These results agreeology 47 (2011) 67–75
with Chen et al. (2000), who showed that when plants are attackedby pathogens, defence enzyme activities increase over time as thedisease develops. Ren et al. (2008) also showed that watermelon,when intercropped with rice, had significantly lower MDA contentand defence-related enzyme activity compared to watermelon inmonoculture. The MDA content of leaves treated with bio-organicfertilizer was much lower than that of control plants (Wu et al.,2008). These results demonstrate that the BIOs used in the presentstudy significantly inhibited the invasion of pathogens into plantroots, so that the disease-free plants had low activities of relateddisease-response enzymes. This further confirms that the BIOs usedin this study can effectively control fusarium wilt disease, not byincreasing resistance of the plants to the pathogens but by protect-ing the roots from pathogen invasion.
In conclusion, the BIOs used in the present study effectively sup-press fusarium wilt disease in melon. The most effective methodis double application (i.e., in the nursery and in the field phases).It reduced pathogen populations in stems, roots and rhizoplanesoil. Moreover, it altered microbial community structure. BIO treat-ments lead to low disease incidence. Consequently, the activities ofdefense-related enzymes in plant leaves were lower in treatmentswith BIOs.
Acknowledgements
This work was supported by the project “Basic Research onFertiliser Saving and Efficiency Improvement for Sustainable Uti-lization of Farmland” (2007CB109304), and the 863 Program(2006AA10Z416) of the China Science and Technology Ministry. Wewould like to acknowledge Professor Hans Lambers for revising thismanuscript.
References
Asada, K., 1992. Ascorbate peroxidase – a hydrogen peroxide scavenging enzyme inplants. Physiol. Plantarum 85, 235–241.
Avdiushko, S.A., Ye, X.S., Kuc, J., 1993. Detection of several enzymatic activities inleaf prints of cucumber plant. Physiol. Mol. Plant Pathol. 42, 441–454.
Bark, R., 1981. Ecology of the fungus Fusarium: competition. In: Nelson, P.E., Tousson,T.A., Cook, R.J. (Eds.), Fusarium Diseases, Biology, and Taxonomy. PennsylvaniaState University Press, University Park, pp. 245–249.
Borrero, C., Trillas, M.I., Ordovas, J., Tello, J.C., Aviles, M., 2004. Predictive factors forthe suppression of fusarium wilt of tomato in plant growth media. Phytopathol-ogy 94, 1094–1101.
Borrero, C., Ordovas, J., Trillas, M.I., Aviles, M., 2006. Tomato fusarium wilt suppres-siveness. The relationship between the organic plant growth media and theirmicrobial communities as characterized by Biology (R). Soil Biol. Biochem. 38,1631–1637.
Boutler, J.I., Boland, G.J., Trevors, J.T., 2000. Compost: a study of the developmentprocess and end-product potential for suppression of turfgrass disease. WorldJ. Microb. Biot. 16, 115–134.
Boulter, J.I., Trevors, J.T., Boland, G.J., 2002. Microbial studies of compost: bacterialidentification, and their potential for turfgrass pathogen suppression. World J.Microb. Biot. 18, 661–671.
Brimner, T., Boland, G., 2003. A review of the non-target effects of fungi used tobiologically control plant diseases. Agr. Ecosyst. Environ. 100, 3–16.
Brussaard, L., Ruiter, P.C., Brown, G.G., 2007. Soil biodiversity for agricultural sus-tainability. Agr. Ecosyst. Environ., Available online at www.sciencedirect.com.
Cebolla, B., Busto, J., Ferrer, A., Miguel, A., Maroto, J.V., 2000. Methyl bromide alter-natives on horticultural crops. Acta Hort. 532, 237–242.
Cebolla, V., Pera, J., 1983. Suppressive effects of certain soils and substrates againstFusarium wilt of carnation. Acta Hort. 150, 113–119.
Champaco, E.R., Martyn, R.D., Miller, M.E., 1993. Comparison of Fusarium solani andFusarium oxysporum as causal agents of fruit rot and root rot of muskmelon.Hort. Sci. 28, 1174–1177.
Chen, C., Richard, R., Nicole, B., Paulitz, T.C., 2000. Defense enzymes inducedin cucumber roots by treatment with plant growth-promoting rhizobacteria(PGPR) and Pythium aphanidermatum. Physiol. Mol. Plant Pathol. 56, 13–23.
Cotxarrera, L., Trillas-Gay, M.I., Steinberg, C., Alabouvette, C., 2002. Use of sewage
sludge compost and Trichoderma asperellum isolates to suppress fusarium wiltof tomato. Soil Biol. Biochem. 34, 467–476.De Cal, A., 1995. Biological control of Fusarium oxysporum f. sp. lycopersici. PlantPathol. 44, 909–917.
De Cal, A., Pascual, S., Melgarejo, P., 1997. Involvement of resistance induction byPenicillium oxalicum in the biocontrol of tomato wilt. Plant Pathol. 46, 72–79.

oil Ec
D
D
E
F
G
G
G
G
H
H
H
H
H
K
K
K
K
L
L
L
M
M
M
10526-008r-r9168-7.
Q. Zhao et al. / Applied S
e Cal, A., García-Lepe, R., Melgarejo, P., 2000. Induced resistance by Penicilliumoxalicum against of Fusarium oxysporum f. sp. lycopersici: histological studies ofinfected and induced tomato stems. Phytopathology 90, 260–268.
ommergues, Y.R., 1978. The plant–microorganism system. In: Dommergues, Y.R.,Krupa, S.V. (Eds.), Interactions between Nonpathogenic Soil Microorganisms andPlants. Elsevier, Amsterdam, Netherlands, pp. 1–37.
l-Hassan, S.A., Gowen, S.R., 2006. Formulation and delivery of the bacterial antag-onist Bacillus subtilis for management of lentil vascular wilt caused by Fusariumoxysporum f. sp. lentis. Phytopathology 154, 148–155.
ravel, D.R., Deahl, K.L., Stommel, J.R., 2005. Compatibility of the biocontrol fun-gus Fusarium oxysporum strain CS-20 with selected fungicides. Biol. Control 34,165–169.
arcia-Limones, C., Hervas, A., Navas-Cortes, J.A., Jimenez-Diaz, R.M., Tena, M., 2002.Induction of an antioxidant enzyme system and other oxidative stress markersassociated with compatible and incompatible interactions between chickpea(Cicer arietinum L.) and Fusarium oxysporum f. sp. ciceris. Physiol. Mol. PlantPathol. 61, 325–337.
arret, S.D., 1970. Pathogenic Root-Infecting Fungi. Cambridge University Press,London, UK.
echev, T., Willekens, H., Montagu, M.V., Inze, D., Camp, W.V., 2003. Differentresponses of tobacco antioxidant enzymes to light and chilling stress. J. PlantPhysiol. 160, 509–515.
ong, M., Wang, J.D., Zhang, J., Yang, H., Lu, X.F., Pei, Y., Cheng, J.Q., 2006. Study ofthe antifungal ability of Bacillus subtilis strain PY-1 in vitro and identification ofits antifungal substance (iturin A). Acta Biochim. Biophys. Sin. 38, 233–240.
artman, G.L., Huang, Y.H., Li, S., 2004. Phytotoxicity of Fusarium solani culture fil-trates from soybeans and other hosts assayed by stem cuttings. Aust. J. PlantPathol. 33, 9–24.
eber, U., Miyake, C., Mano, J., Ohno, C., Aada, K., 1996. Monodehydroascorbate rad-ical detected by electron paramagnetic resonance spectrometry is a sensitiveprobe of oxidative stress in intact leaves. Plant Cell Physiol. 37, 1066–1072.
ervas, A., Linda, B., Datnoff, L.E., Jimenez-Diaz, R.M., 1998. Effect of commercialand indigenous microorganisms on Fusarium wilt development in chickpea. Biol.Control 13, 166–176.
oitink, H.A.J., Boehm, M.J., 1999. Biocontrol within the context of soil microbialcommunities: a substrate-dependent phenomenon. Annu. Rev. Phytopathol. 37,427–446.
oitink, H.A.J., Stone, A.G., Han, D.Y., 1997. Suppression of plant disease by composts.Hort. Sci. 32, 184–187.
andhari, J., Majumder, S., Sen, B., 2000. Impact of Aspergillus niger AN27 on growthpromotion and sheath blight disease reduction in rice. Pest Manage. Sci. (Decem-ber), 21–22.
aur, N.P., Mukhopadhyay, A.N., 1992. Integrated control of chickpea wilt complexby Trichoderma and chemical methods in India. Trop Pest Manage. 38, 372–375.
ing, S.R., Davis, A.R., Liu, W., Levi, A., 2008. Grafting for disease resistance.HortScience 43, 1673–1676.
omada, H., 1975. Development of a selective medium for quantitative isolation ofFusarium oxysporum from natural soil. Rev. Plant Protect. Res. 8, 114–125.
arena, I., Sabuquillo, P., Melgarejo, P., De Cal, A., 2003. Biocontrol of Fusarium andVerticillium wilt of tomato by Penicillium oxalicum under greenhouse and fieldconditions. Phytopathology 151, 507–512.
arkin, R.P., Fravel, D.R., 1998. Efficacy of various fungal and bacterial biocontrolorganisms for control of Fusarium wilt of tomato. Plant Dis. 82, 1022–1028.
ing, N., Xue, C., Huang, Q.W., Yang, X.M., Xu, Y.C., Shen, Q.R., in press. Developmentof a mode of application of bioorganic fertilizer for improving the biocontrolefficacy to Fusarium wilt. Biocontrol.
artyn, R.D., Amador, J., 1987. Fusarium wilt (Fusarium oxysporum f. sp. melonis Race
0) of muskmelon in Texas. Plant Dis. 71, 469.ishra, K., Kumar, A., Pandey, K., 2010. RAPD based genetic diversity among differentisolates of Fusarium oxysporum f. sp. lycopersici and their comparative biocontrol.World J. Microb. Biot. 26, 1079–1085.
orsya, M.R., Jouvebm, L., Hausmanbm, J., Hoffmannbm, L., Stewartam, J.M., 2007.Alteration of oxidative and carbohydrate metabolism under abiotic stress in two
ology 47 (2011) 67–75 75
rice (Oryza sativa L.) genotypes contrasting in chilling tolerance. Plant Physiol.164, 157–167.
Muslim, A.H., Horinouchi, Hyakumachi, M., 2003. Biological control of fusarium wiltof tomato with hypovirulent binucleate Rhizoctonia in greenhouse conditions.Mycoscience 44, 77–84.
Nakano, Y., Asada, K., 1981. Hydrogen peroxide is scavenged by ascorbate specificperoxidase in spinach chloroplasts. Plant Cell Physiol. 22, 867–880.
Park, C.S., Paulitz, T.C., Baker, R., 1988. Biocontrol of fusarium wilt of cucumber result-ing from interactions between Pseudomonas putida and non-pathogenic isolatesof Fusarium oxysporum. Phytopathology 78, 190–194.
Phae, C.G., Saski, M., Shoda, M., Kubota, H., 1990. Characteristic of Bacillus subtilisisolated from composts suppressing phytopathogenic micro-organisms. Soil Sci.Plant Nutr. 36, 575–586.
Raviv, M., Reuveni, R., Zaidman, B.Z., 1998. Improved medium for organic transplant.Biol. Agric. Hortic. 16, 53–64.
Ren, L.X., Su, S.M., Yang, X.M., Xu, Y.C., Huang, Q.W., Shen, Q.R., 2008. Intercroppingwith aerobic rice suppressed fusarium wilt in watermelon. Soil Biol. Biochem.40, 834–844.
Rojo, F.G., Reynoso, M.M., Ferez, M., Chulze, S.N., Torres, A.M., 2007. Biological controlby Trichoderma species of Fusarium solani causing peanut crown root rot underfield conditions. Crop Protect. 26, 549–555.
Rossum, M.W.P.C., Alberda, M., Plas, L.H.W., 1997. Role of oxidative damage in tulipbulb scale micropropagation. Plant Sci. 130, 207–216.
Sabuquillo, P., De Cal, A., Melgarejo, P., 2006. Biocontrol of tomato wilt by Penicil-lium oxalicum formulations in different crop conditions. Biol. Control 37, 256–265.
Soriano-Martın, M.L., Porras-Piedra, A., Porras-Soriano, A., 2006. Use of microwavesin the prevention of Fusarium oxysporum f. sp. melonis infection during the com-mercial production of melon plantlets. Crop Protect. 25, 52–57.
Sturz, A.V., Christie, B.R., 2003. Beneficial microbial allelopathies in the root zone:the management of soil quality and plant disease with rhizobacteria. Soil Till.Res. 72, 107–123.
Suárez-Estrella, F., Vargas-Garcia, C., Lopez, C.M.J., Capel, J.M., 2007. Antagonisticactivity of bacteria and fungi from horticultural compost against Fusarium oxys-porum f. sp. melonis. Crop Protect. 26, 46–53.
Suárez-Estrella, F., Elorrieta, M.A., Vargas-García, M.C., López, M.J., Moreno, J., 2001.Selective isolation of antagonist micro-organisms of Fusarium oxysporum f. sp.melonis. Biological Control of Fungal and Bacterial Plant Pathogens, Interna-tional Organization for Biological Control (IOBC) West Palaeartic Regional Sector(WPRS). Bulletin 24, 109–112.
Tao, F., Zhang, M., Yu, H.Q., 2007. Effect of vacuum cooling on physiological changesin the antioxidant system of mushroom under different storage conditions. J.Food Eng. 79, 1302–1309.
Tamietti, G., Valentino, D., 2006. Soil solarization as an ecological method for thecontrol of fusarium wilt of melon in Italy. Crop Protect. 25, 389–397.
Tawfic, A.A., Allam, A.D.A., 2004. Improving cumin production under soil infesta-tion with Fusarium wilt pathogen: II field trial of different landraces and seedtreatments. Assiut Univ. Bull. Environ. Res. 7, 47–63.
Weller, D.M., Raaijmakers, J.M., Mc Spadden Gardener, B.B., Thomashow, L.S., 2002.Microbial populations responsible for specific soil suppressiveness to plantpathogens. Annu. Rev. Phytopathol. 40, 309–348.
Whipps, J.M., 1997. Developments in the biological control of soilborne plantpathogens. Adv. Bot. Res. 26, 1–134.
Wu, H.S., Yang, X.M., Fan, J.Q., Miao, W.G., Ling, N., Xu, Y.C., Huang, Q.W., Shen, Q.R.,2008. Suppression of fusarium wilt of watermelon by a bio-organic fertilizer con-taining combinations of antagonistic microorganisms. BioControl, doi:10.1007/s
Zhang, S.S., Raza, W., Yang, X.M., Hu, J., Huang, Q.W., Xu, Y.C., Liu, X.H., Ran, W.,Shen, Q.R., 2008. Control of fusarium wilt disease of cucumber plants with theapplication of a bio-organic fertilizer. Biol. Fert. Soils 44, 1073–1080.
Zhao, H.C., Zhao, H., Wang, B.C., Wang, J.B., 2005. Effect of local stress induction onresistance-related enzymes in cucumber seeding. Colloid Surf. B 43, 37–42.





























