Embed Size (px)
Citation preview

SCIENCE CHINA Earth Sciences
© Science China Press and Springer-Verlag Berlin Heidelberg 2010 earth.scichina.com www.springerlink.com
*Corresponding author (email: [email protected])
• RESEARCH PAPER • December 2010 Vol.53 No.12: 1800–1810
doi: 10.1007/s11430-010-4071-3
Biodiversification of Late Ordovician Hirnantia Fauna on the Upper Yangtze Platform, South China
ZHAN RenBin1, LIU JianBo1,2*, Ian G. PERCIVAL3, JIN JiSuo4 & LI GuiPeng1
1 State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing 210008, China;
2 Department of Geology, Peking University, Beijing 100871, China; 3 Geological Survey of New South Wales, NSW Department of Primary Industries, 947-953 Londonderry Road, Londonderry,
NSW 2753, Australia; 4 Department of Earth Sciences, University of Western Ontario, London ON N6A 5B7, Canada
Received March 16, 2010; accepted July 6, 2010
The Hirnantia Fauna is a globally-represented, cool water brachiopod fauna that originated and flourished in shallow marine environments at intermediate to low latitudes after the first episode of the end-Ordovician mass extinction. It was well-developed, widely distributed, long in stratigraphical range, and complex in palaeoecological differentiation on the Upper Yangtze Platform of the South China paleoplate, where it has been extensively studied. Qualitative and quantitative analyses show that the FAD of the Hirnantia Fauna in South China becomes stratigraphically higher from nearshore to offshore locali-ties on the Yangtze Platform, while the range of the fauna becomes shorter whereas the taxonomic diversity increases. Over its stratigraphical range the diversity of the Hirnantia Fauna at nearshore localities consistently decreases, but trends in the oppo-site direction at offshore, deeper water localities. The fauna was represented by different communities, subcommunities or as-sociations with changing environmental factors (such as water depth and substrate) at different localities or horizons.
Hirnantia Fauna, biodiversification, Ordovician, Hirnantian, Upper Yangtze Platform, South China
Citation: Zhan R B, Liu J B, Percival I G, et al. Biodiversification of Late Ordovician Hirnantia Fauna on the Upper Yangtze Platform, South China. Sci China Earth Sci, 2010, 53: 1800–1810, doi: 10.1007/s11430-010-4071-3
The Hirnantia Fauna, a shallow marine brachiopod fauna developed at many localities in the world during the Hirnantian (Late Ordovician), is represented by the epony-mous genus Hirnantia, together with some characteristic constituents such as Kinnella, Draborthis, Leptaenopoma and Paromalomena, and some cosmopolitan components (e.g., Dalmanella, Cliftonia, Eostropheodonta, Fardenia, and Hindella) [1]. Its uniqueness in geological history and its wide distribution, both paleogeographically and pa-leoecologically, have made it a focus of study by evolution-ary paleobiologists and biostratigraphers. With its contem-
porary trilobite fauna characterized by Dalmanitina, the association is often combined together as the Hirnan-tia-Dalmanitina Fauna [2].
Such a fauna was originally recognized as the Hirnant Fauna from the Upper Ordovician near the small town of Hirnant in northern Wales [3], consisting of some typical components, such as Hirnantia sagittifera, Dalmanella testudinaria, Eostropheodonta hirnantensis, and Plecto-thyrella fissicostata. Subsequently, similar fossil assem-blages with these characteristic brachiopods, occupying a restricted geologic range and a wide geographic distribution, were reported at so many localities in Europe that Temple [4] formally called this association the “Hirnantia Fauna”.

ZHAN RenBin, et al. Sci China Earth Sci December (2010) Vol.53 No.12 1801
The same unique brachiopod fauna of end-Ordovician age had been investigated by some Chinese experts since the early 1960s, based on abundant material from South China (Rong Jiayu, personal communication in recent years). Un-fortunately, some events intervened, and delayed publica-tion of the relevant research until the late 1970s (e.g., Rong [5]), nearly 15 years behind the international pace.
After decades of intense investigation, the Hirnantia Fauna is now well-known for its global geological and stratigraphical distributions and its significance in the study of end-Ordovician macroevolution (e.g., [6, 7] and refer-ences therein). There are more than 100 localities in nearly 30 countries and regions where the Hirnantia Fauna has been documented [5, 7]. The fauna was biogeographically further differentiated into the typical Hirnantia Fauna (the Kosov Province) and the atypical Hirnantia Fauna (the Bani Province) [6]. A contemporary fauna without Hirnantia that developed in equatorial regions was called the Midcontinent Fauna (the Edgewood Province) [6].
The Hirnantia Fauna is limited mostly to the Hirnantian (uppermost Ordovician) globally, and is slightly older or younger only at a few localities [8, 9]. The fauna is com-monly confined to the Normalograptus extraordinarius-N. ojsuensis Biozone, or to lower part of the succeeding Nor-malograptus persculptus Biozone, or spans the boundary between these two biozones. On the Upper Yangtze Plat-form of South China, the Hirnantia Fauna spans these two biozones. Its occurrence at Honghuayuan of Tongzi from the Kuanyinchiao Formation (now spelled Guanyinqiao) was thought to have the longest stratigraphical range from the upper N. extraordinarius-N. ojsuensis Biozone to the lower Akidograptus ascensus Biozone (the lowermost Silu-rian) [7, 10], but further investigation confirms that there is a hiatus between the topmost Ordovician and the lowermost Silurian at this section, i.e., the Hirnantia Fuana here is ended in the middle N. persculptus Biozone (Figure 1) (Rong Jiayu, 2009, personal communication). So, the Hirnantia Fauna from the lower Skelgill Formation at
Figure 1 High-resolution correlation between series of Ordovician-Silurian boundary sections yielding Hirnantia Fauna, mainly in South China and one each in Sibumasu and Tibet. Shaded part of columns spans the interval between the first appearance horizon (FAH) and last appearance horizon (LAH) of the Hirnantia Fauna at each site. Lower YP, Lower Yangtze Platform; ZXS, Zhexi Slope developed during the Late Ordovician [11]; KY Fm, Kuanyinchiao Formation. 1. Ludiping, Songtao; 2. Donggongsi, Zunyi; 3. Yanzikou, Bijie; 4. Shichang, Renhuai; 5. Yangliugou, Renhuai; 6. Ganxi, Yanhe; 7. Honghua-yuan, Tongzi; 8. Gusong, Xingwen; 9. Shuanghe, Changning; 10. Wangjiawan, Yichang; 11. Guantangyuan, Wuning; 12. Beigong, Jingxian; 13. Yankou, Lin’an; 14. Wanyaoshu, Luxi; 15. Zhiwazuogou, Xainza.

1802 ZHAN RenBin, et al. Sci China Earth Sci December (2010) Vol.53 No.12
Yewdale Beck of northern England is now the youngest known representative of this fauna, where some typical constituents of the Hirnantia Fauna (e.g., Kinnella, Miror-this, Hirnantia, Paromalomena, Hindella, and Plecto-thyrella, Figure 2) occur interbedded with graptolites of the Parakidograptus acuminatus Biozone [8].
Detailed investigation of the Hirnantia Fauna on a global scale has already made a significant contribution to discus-
sion about the position of the Ordovician-Silurian boundary, as well as biotic macroevolution across this boundary, and most importantly has enabled recognition of the essence of the end-Ordovician mass extinction. In the past decade, geochemical analyses (mainly carbon isotopes) have been conducted for this particular interval at several key sections around the world, e.g., Dob’s Linn, Scotland [12], Anticosti, eastern Canada [13], Nevada, southwestern United States
Figure 2 Representative brachiopods of the typical Hirnantia Fauna. All figured specimens were collected from the Kuanyinchiao Formation (Hirnantian, Upper Ordovician) at Wangjiawan, northern Yichang, western Hubei Province, at which the GSSP site of the Hirnantian is located. 1, 2. Kinnella kielanae, ventral and dorsal internal moulds, NIGP136755, 136756; 3, 4. Paromalomena polonica, ventral and dorsal internal moulds, NIGP136757, 136758; 5. Dal-manella testudinaria, dorsal internal mould, NIGP136765; 6. Cliftonia oxoplecioides, ventral internal mould, NIGP136767; 7. Triplesia yichangensis, ven-tral internal mould, NIGP136764; 8. Aegiromena ultima, conjoined ventral and dorsal internal moulds of a single individual, NIGP136768; 9. Hindella crassa incipiens, ventral internal mould, NIGP136771; 10, 11. Eostropheodonta parvicostellata, dorsal and ventral internal moulds, NIGP136769, 136770; 12. Leptaena trifidum, ventral internal mould, NIGP136772; 13, 14. Hirnantia sagittifera, ventral and dorsal internal moulds, NIGP136774, 136775. Scale bars = 1 mm unless otherwise noted.

ZHAN RenBin, et al. Sci China Earth Sci December (2010) Vol.53 No.12 1803
[14], Valga and Kaugatuma, Estonia [15–17], and Wang-jiawan, Yichang, central China [18]. Such research has made international correlation of this particular interval more reliable and precise, and also has substantially assisted our detailed understanding of correlation between biodiver-sity and environmental changes during the Hirnantian.
Recently, a unique shelly fauna of very small shells (mostly less than 8 mm in width) dominated by brachiopods has been reported from yellowish green mudstone of the upper Yankou Formation at Yuhang of Hangzhou, Zhejiang Province (paleogeographically on the upper Zhexi Slope (western Zhejiang)) [19]. Some key elements of the fauna (e.g., the plectambonitoid Aegiromena planissima and the trilobite Dalmanitina (Songxites) cf. wuningensis), together with graptolites from underlying strata (i.e., lower part of the Yankou Formation) and the overlying Anjie Formation, confirm that this fauna lies biostratigraphically in the upper Normalograptus persculptus Biozone. Thus, it is the youngest known Ordovician shelly fauna anywhere in the world. It is provisionally called the Leangella-Dalmanitina Assemblage, and is thought to have lived in a deep water benthic regime corresponding to BA5 (or slightly deeper) on muddy substrates [20]. In macroevolutionary terms, it is significant in bridging the gap between the Hirnantia Fauna and the earliest Silurian shelly fauna.
South China, particularly the Upper Yangtze Platform, is the only place globally where the Hirnantia Fauna has a wide distribution within a single terrane, being represented at many localities, with a long stratigraphical range, high taxonomic diversity, and pronounced ecological differentia-
tions. Having been intensely collected and studied for seve- ral decades by Chinese paleontologists as well as some of their foreign colleagues, the Hirnantia Fauna is now one of the best known Ordovician shelly faunas in South China, and indeed in the world [21]. The major objective of this paper is to summarize the geographical and stratigraphical distribution of the Hirnantia Fauna on the Upper Yangtze Platform of South China, and to analyze its palaeoeco- logical differentiation from nearshore to offshore localities.
1 Geographical and geological distribution of the Hirnantia Fauna on the Upper Yangtze Plat-form
In the 1950s, Chinese biostratigraphers began to pay atten-tion to a thin but distinctive unit of shelly rocks near the O/S boundary, naming it the Kuanyinchiao Bed with type local-ity at Guanyinqiao of Qijiang County, Chongqing District [2]. It was also informally called the Dalmanitina Bed [22, 23] due to the abundance of this famous trilobite. Subse-quently, similar rocks and fauna were found on the entire Yangtze Platform and at some localities of western Yunnan (northern Sibumasu paleoplate) and Tibet. Among nearly one hundred localities yielding the Hirnantia Fauna of South China, most are in Guizhou, Sichuan, and Hubei provinces of the Upper Yangtze Platform (Figure 3). There was a long debate among many Chinese paleontologists and stratigraphers about the validity of the Kuanyinchiao Bed as a formal lithological unit because of its thickness. Its strati-
Figure 3 Location map of the Kuanyinchiao Formation and its equivalents, containing the Hirnantia Fauna in South China. 1. Tiemenkan section near Shuanghe town, Changning County, southern Sichuan; 2. Helong, Shuanghe, Changning, southern Sichuan; 3. Gusong, Xingwen, southern Sichuan; 4. Yan-zikou, Bijie, northwestern Guizhou; 5. Yangliugou and Shichang, Zhangjiaping, Renhuai, northern Guizhou; 6. Honghuayuan, Tongzi, northern Guizhou; 7. Jiancaohe, Zunyi, northern Guizhou; 8. Zhujiapo, Zunyi, northern Guizhou; 9. Wulipo, Meitan, northeastern Guizhou; 10. Dongkala, Fenggang, northeastern Guizhou; 11. Donghuaxi, Si’nan, northeastern Guizhou; 12. Yingwuxi, Si’nan, northeastern Guizhou; 13. Shichang’ao, Ganxi, Yanhe, northeastern Guizhou; 14. Shatuo, Yanhe County, northeastern Guizhou; 15. Datianba, Xiushan, Chongqing; 16. Ludiping, Songtao, northeastern Guizhou; 17. Heshui, Yinjiang, northeastern Guizhou; 18. Shanxi, Yinjiang, northeastern Guizhou, 19. Leijiatun, Shiqian, northeastern Guizhou; 20. Wenjiadian, Si’nan, northeastern Guizhou; 21. Zhiwazuogu, Xainza, south-central Xizang; 22. Wanyaoshu, Luxi, western Yunnan; 23. Wangjiawan, Yichang, western Hubei; 24. Guantan-gyuan, Wuning, northern Jiangxi; 25. Beigong, Jingxian, southern Anhui; 26. Yankou, Lin’an, northwestern Zhejiang. Black square represents higher diver-sity Hirnantia Fauna; black triangle is lower diversity Hirnantia Fauna; hollow triangle indicates the Kuanyinchiao Formation and its equivalents without Hirnantia Fauna; circle with a cross shows location of a hiatus across the Ordovician and Silurian rocks without Kuanyinchiao Formation or its equivalents and Hirnantia Fauna. Thick dashed line in (b) indicates postulated paleoshoreline, and fine dashed lines circling localities 11 and 12, and locality 14 indi-cates postulated uplifted areas.

1804 ZHAN RenBin, et al. Sci China Earth Sci December (2010) Vol.53 No.12
graphic status was also controversial such that it was re-ferred to variously as the Kuanyinchiao Member within the Wufeng Formation or the Kuanyinchiao Formation. More-over, the lithostratigraphic and chronostratigraphic rela-tionships with its underlying Wufeng and overlying Lung-machi (now spelled as Longmaxi) formations are also un-resolved at some localities. Although some of these prob-lems remain to be settled, recent opinion is for the unit to be designated the Kuanyinchiao Formation [24].
Among several tens of localities on the Upper Yangtze Platform, the Hirnantia Fauna at Ludiping of Songtao County, northeastern Guizhou Province (Section 16 in Fig-ure 3) has the oldest FAD (first appearance datum) near the bottom of the Normalograptus extraordinarius-N. ojsuensis Biozone, i.e., the base of Hirnantian [7, 10] (Figure 1). The Kuanyinchiao Formation at this site is 1.52 m thick and is composed of calcareous mudstone and argillaceous lime-stone. The brachiopod fauna is typical of Hirnantia Fauna, including ten species and genera, i.e., Dalmanella testudi-naria, Hirnantia sagittifera, Kinnella kielanae, Leptaena trifidum, Paromalomena polonica, Eostropheodonta parvi-costellata, Fardenia cf. F. modica, Cliftonia sp., Plecto-thyrella crassicosta, and Hindella crassa incipiens. Its LAD (last appearance datum) at this locality is close to the top of the N. extraordinarius-N. ojsuensis Biozone [7].
The Kuanyinchiao Formation at Jiancaohe, Donggongsi, Zunyi, northern Guizhou (Section 7 in Figure 3) is com-paratively thin and is composed mainly of calcareous mud-stone [25, 26]. The Hirnantia Fauna at this site is of low diversity but is very abundant with large shell sizes. Repre-sentative constituents are Dalmanella testudinaria, Hirnan-tia sagittifera, Kinnella kielanae, Eostropheodonta ultrix, and Plectothyrella crassicosta. The underlying and overly-ing strata are the Wufeng and Lungmachi formations re-spectively, both containing graptolites which confirm that the age of the Kuanyinchiao Formation and the Hirnantia Fauna at this site ranges from the basal to the upper part of the Normalograptus extraordinarius-N. ojsuensis Biozone [5, 27] (Figure 1).
The Kuanyinchiao Formation at Zhonggou of Yanzikou, Bijie, western Guizhou (Section 4 in Figure 3) is 1.35 m thick and contains mainly bioclastic micritic limestone [28]. Brachiopod fossils from the upper 40 cm of the formation are mostly silicified, including Dalmanella testudinaria, Eostropheodonta sp., Fardenia modica, Dorytreta longicr- ura, Hindella crassa incipiens, and Plectothyrella crassico- sta, as well as many rugose corals, trilobites, and gastropods [28]. Although Hirnantia itself has not yet been found at this site, this association is also representative of the typical Hirnantia Fauna. Graptolites from the underlying Wufeng and overlying Lungmachi formations constrain the age of the Hirnantia Fauna here precisely from the lower part to the top of the Normalograptus extraordinarius-N. ojsuensis Biozone [28] (Figure 1).
The Kuanyinchiao Formation and the Hirnantia Fauna
are represented at several localities in Renhuai County, northern Guizhou. Two sections at Yangliugou and Shichang (Locality 5 in Figure 3) have been carefully measured and collected. The thickness of the Kuanyinchiao Formation is between 2.4 and 2.6 m, and the formation is composed mainly of a lower calcareous mudstone with limestone nodules, and an upper bioclastic limestone. Hirnantia Fauna with an intermediate level of diversity was found in the lower part of the formation, with predominant brachiopods including Pseudopholidops partibilis, Dal-manella testudinaria, Hirnantia sagittifera, Kinnella kie-lanae, Aegiromena? sp., Leptaena trifidum, Paromalomena polonica, Eostropheodonta parvicostellata, Fardenia cf. F. modica, Dorytreta longicrura, and Hindella crassa in-cipiens. As confined by graptolites from the underlying Wufeng and overlying Lungmachi formations, this fauna covers the time span represented by the sequence from near the base of the Normalograptus extraordinarius-N. ojsuen-sis Biozone to the lower part of the N. persculptus Biozone [7] (Figure 1).
The Kuanyinchiao Formation at Shichang’ao of Ganxi, Yanhe County, northeastern Guizhou (Section 13 in Figure 3) is close to 1 m thick, and consists mainly of bioclastic limestone with little argillaceous components. Very few shelly fossils were found, mainly from the lower and upper parts of the formation, where brachiopods are the dominant group, including Dalmanella testudinaria, Hirnantia cf. H. sagittifera, Leptaena trifidum, Paromalomena polonica, Eostropheodonta parvicostellata, Cliftonia sp., Plecto-thyrella crassicosta, and Hindella crassa incipiens. Grapto-lites from the underlying Wufeng and overlying Lungmachi formations confirm that the Hirnantia Fauna at this site is confined to the lower to middle parts of the Normalograptus extraordinarius-N. ojsuensis Biozone [7].
At Honghuayuan of Tongzi, northern Guizhou (Section 6 in Figure 3), the Kuanyinchiao Formation is well developed and comparatively thicker (5.76 m) than most of other lo-calities on the Upper Yangtze Platform. Graptolites are also abundant in the calcareous mudstone of its lower part, and shelly fossils occur throughout the formation with brachio-pods as the dominant group. Genera representatives at vari-ous horizons through this section include Pseudopholidops partibilis, Orbiculoidea sp., Dalmanella testudinaria, Mi-rorthis sp., Hirnantia sagittifera, Leptaena trifidum, Paro-malomena polonica, Eostropheodonta parvicostellata, Fardenia sp., Cliftonia sp., Plectothyrella cf. crassicosta, Hyattidina? sp., Hindella crassa incipiens, and Eospirifer sp. Graptolites from the underlying Wufeng Formation, the overlying Lungmachi Formation and within the Kuany-inchiao Formation itself confirm that the Hirnantia Fauna at this locality ranges from the lower to middle parts of the Normalograptus extraordinarius-N. ojsuensis Biozone to the middle Normalograptus persculptus Biozone, and a hia-tus exists during the O/S transition at this site for at least two graptilitic biozones (Rong Jiayu, 2009, personal com-

ZHAN RenBin, et al. Sci China Earth Sci December (2010) Vol.53 No.12 1805
munication). The level of brachiopod diversity at the Honghuayuan
section is intermediate between shallow water settings to-wards the shoreline, and high diversity in deeper water en-vironments further offshore with up to twice the generic diversity. Although the Kuanyinchiao Formation most likely was accumulated here at shallower water depths than prevailed during deposition of the underlying Wufeng For-mation, clearly the difference in water depth was not as marked as in other sections on the Yangtze Platform, as only in the Honghuayuan section do graptolites occur within the Kuanyinchiao Formation (particularly in its lower part). The fauna includes Mirorthis, which is a component of the deeper water Mirorthis Association recognized in the more offshore setting of the Shuanghe section of Changning [29], and also is present in the Wangjiawan section of Yichang (offshore locality). We therefore interpret the Kuanyinchiao Formation in the Honghuayuan section as having initially formed in a BA4-5 setting, that subsequently got shallower slightly (perhaps to lower BA3 depths) before deepening again with a brief recurrence of graptolitic facies in its up-per half; this was succeeded by continued shallowing (BA3-4) into the basal Silurian while the Upper Yangtze Platform elsewhere was subject to a widespread deep water inundation. To explain this unique history, we suggest that the Honghuayuan section was initially deposited in a local-ized small subsiding basin, bounded by growth faults, which would explain the presence of graptolite-rich mudstones forming its lower part and the comparatively thick sediment accumulation. Then, as the second major transgression took place across the Yangtze Platform in latest Ordovician to earliest Silurian time, this localized area must have main-tained a slight elevation relative to the surrounding sea floor (possibly produced by reverse fault movement) in order to have continued to provide a suitable depth for habitation by the Hirnantia fauna. This depth would not have been as shallow as that when the Hirnantia fauna first populated the Upper Yangtze Platform, because by mid-Hirnantian time the fauna had apparently evolved to inhabit lower BA3 or even BA4 environments further offshore, as shown in this study. Although regional geological maps of the Tongzi area do in fact reveal surface traces of faults that lie parallel to the trend of the Ordovician shoreline, without seismic sections across the region it is impossible to confirm whether growth faults existed in Late Ordovician as we postulate. Therefore, the model suggested here awaits veri-fication, and other explanations to account for the extended deposition of the Kuanyinchiao Formation in the Honghua-yuan section may be possible.
At the opposite end of the spectrum, the Kuanyinchiao Formation at Gusong of Xingwen County, southern Sichuan Province (Section 3 in Figure 3), is only 38 cm thick. Here the Kuanyinchiao Formation is also well exposed, consist-ing of argillaceous limestone yielding very abundant shelly fossils of comparatively high diversity. Brachiopods include
Philhedra sp., Deliella sp., Pseudopholidops sp., Dal-manella testudinaria, Trucizetina sp., Dysprosorthis sp., Draborthis caelebs, Hirnantia sagittifera, Kinnella kielanae, Triplesia sp., Cliftonia cf. C. oxoplecioides, Leptaena trifi-dum, Eostropheodonta parvicostellata, Fardenia sp., Dory-treta cf. D. longicrura, Plectothyrella crassicosta, Sphenot-reta sp., and Hindella crassa incipiens. Biostratigraphically, it ranges between the upper Normalograptus extraordi-narius-N. ojsuensis Biozone and the basal Normalograptus persculptus Biozone, as determined by graptolites from the underlying Wufeng and overlying Lungmachi formations [7].
The Kuanyinchiao Formation near Shuanghe town, Changning County, southern Sichuan Province (Section 1 in Figure 3) has been reported many times by different re-searchers [5–7, 10, 30]. We have investigated this section on several occasions during the past decade, re-measuring the 65-cm-thick Kuanyinchiao Formation and making five detailed collections. The dominant lithology of the Kuany-inchiao Formation here is dark grey or black calcareous mudstone and argillaceous limestone yielding very abun-dant shelly fossils. Brachiopods, according to our prelimi-nary systematic study, are the dominant group and include Aegiromena sp., Cliftonia cf. oxoplecioides, Dalmanella testudinaria, Deliella sp., Dolerorthis sp., Drabovia sp., Eostropheodonta parvicostellata, Epitomyonia sp., Hirnan-tia sagittifera, Kinnella kielanae, Mirorthis mira, Onniella sp., Paracraniops sp., Paromalomena polonica, Plecto-thyrella sp., Rostricellula sp., Strophomena sp., Thebesia sp., and Triplesia sp. The underlying Wufeng and overlying Lungmachi formations are very rich in graptolite fossils, so its age is well constrained from the upper Normalograptus extraordinarius-N. ojsuensis Biozone to the basal N. persculptus Biozone (refs. [5, 7], Fan Junxuan, 2009, per-sonal communication) (Figure 1).
In the Yichang district of western Hubei Province (northeastern Upper Yangtze Platform, comparatively far away from the shoreline: Section 23 in Figure 3), the Kuanyinchiao Formation is between 10 and 30 cm thick and is composed mainly of argillaceous limestone or calcareous mudstone. Weathered exposures appear as yellowish green mudstone, yielding abundant brachiopod fossils of high taxonomic diversity, together with some trilobites, bryo-zoans, and stems of crinoids. The brachiopods, which have been intensely collected and studied during the past several decades, include Pseudopholidops partibilis, Philhedra sp., Deliella sp., Dalmanella testudinaria, Onniella? yichangen-sis, Mirorthis mira, Dysprosorthis sinensis, Draborthis caelebs, Hirnantia sagittifera, Kinnella kielanae, Triplesia yichangensis, Cliftonia obovata, Aegiromena sp., Leptaena trifidum, Paromalomena polonica, Eostropheodonta parvi-costellata, Fardenia sp., Dorytreta cf. D. longicrura, Sphe-notreta sp., Plectothyrella crassicosta, and Hindella crassa incipiens (refs. [1, 10, 18] and references therein). Abundant, well-preserved graptolites from the underlying Wufeng and

1806 ZHAN RenBin, et al. Sci China Earth Sci December (2010) Vol.53 No.12
overlying Lungmachi formations help clarify the age of the Hirnantia Fauna in the Yichang district as defined by the sequence from the top of the Normalograptus extraordi-narius-N. ojsuensis Biozone to the base of the N. persculp-tus Biozone, a very short time interval [1, 5, 10, 18] (Figure 1).
In other regions of South China outside the Upper Yang-tze Platform are some sporadic reports of the Hirnantia Fauna, which is usually low in diversity and abundance. Generally, only 1–3 brachiopod genera are found at each locality and up to now no Hirnantia itself is known from any other areas outside the Upper Yangtze Platform on the South China paleoplate. The rocks yielding these brachio-pods are generally stratigraphically from the top of the Normalograptus extraordinarius-N. ojsuensis Biozone to the base of the N. persculptus Biozone [7]. For example, only a few tiny brachiopods occur in the top part of the Xinkailing Formation at Beigong of Jingxian County, Anhui Province (Section 25 in Figure 3) and the top Wufeng For-mation at Guantangyuan of Wuning County, Jiangxi Prov-ince [31, 32] (paleogeographically both on the Lower Yangtze Platform, Section 24 in Figure 3), comprising Ae-giromena ultima, Paromalomena polonica, and Fardenia sp., all of which are typical components of the Hirnantia Fauna [7]. A few dwarfed individuals of Paromalomena cf. polonica occur together with some trilobites and graptolites in the lower Yankou Formation at Yankou, Lin’an, north-western Zhejiang Province (Section 26 in Figure 3) (paleo-geographically on the upper Zhexi Slope [11]). The accom-panying graptolites confirm that the age of these fossils is defined by the sequence from the middle-upper Normalo-graptus extraordinarius-N. ojsuensis Biozone to the basal N. persculptus Biozone [7]. Unfortunately, neither Hirnantia itself nor other brachiopods were found.
From the above analysis, the general trend of the geo-graphical distribution and stratigraphical range of the Hirnantia Fauna in South China seems to be clear: the closer the locality is to the paleo-shoreline, the older is the FAD of the Hirnantia Fauna (Figure 1). From Guizhou and Sichuan in southwest China, northeastward to Hubei, Ji-angxi and Anhui provinces, the base of the Kuanyinchiao Formation and its equivalents and the FAD of the Hirnantia Fauna progressively becomes higher, while the thickness of the stratum concurrently decreases and the stratigraphical range narrows. The upper boundary of the Kuanyinchiao Formation and the Hirnantia Fauna is commonly close to the base or in the lower part of the Normalograptus persculptus Biozone, except for the locality Honghuayuan of Tongzi, northern Guizhou Province. In southwest China, on the inner shelf of the Upper Yangtze Platform, a similar temporal and spatial trend also existed along the pa-leo-shoreline (Figure 3). For example, at those near shore localities such as Ludiping of Songtao County, Donggongsi of Zunyi, and Yanzikou of Bijie, the base of the Kuany-inchiao Formation and the FAD of the Hirnantia Fauna is
coincident with or close to the base of the Normalograptus extraordinarius-N. ojsuensis Biozone. Slightly further away from the shoreline, at localities of Yanhe and Renhuai counties, the base of the Kuanyinchiao Formation and the FAD of the Hirnantia Fauna is slightly higher, but still in the lower part of the N. extraordinarius-N. ojsuensis Bio-zone. More remote from the shoreline at Changning of southern Sichuan and Wangjiawan of northern Yichang, western Hubei, the base of the Kuanyinchiao Formation and the FAD of the Hirnantia Fauna moved to the middle and upper parts of the N. extraordinarius-N. ojsuensis Biozone respectively.
2 Diversity change of the Hirnantia Fauna on the Upper Yangtze Platform
Although the Kuanyinchiao Formation of South China is of centimeter to meter-scale thickness and relatively simple in lithology, it was subdivided into several layers and intensely collected since the early 1980s [1]. It was found that the Hirnantia Fauna at each locality on the Upper Yangtze Platform changes in diversity in different layers, and there are also some general trends in its geographical distribution.
Figure 4 shows the number of brachiopod genera in each collection at representative localities of the Hirnantia Fauna on the Upper Yangtze Platform. At nearshore localities, the diversity of the Hirnantia Fauna is generally low, and fur-ther decreases upward stratigraphically. For localities inter-preted as offshore, the diversity of the fauna persistently increases upward, with brachiopod diversity near the top of the Kuanyinchiao Formation almost ten times higher than that of the basal part at a few localities (e.g., Huanghua-chang section of Yichang) [1]. The diversity of the Hirnan-tia Fauna at Honghuayuan of Tongzi is comparatively stable at an intermediate level, containing 9–11 brachiopod genera throughout the 5.76-m-thick strata. Such opposite trends of the brachiopod diversities at nearshore and offshore locali-ties might be related to their different paleoecological and regional geological settings when the Cathaysian and the Yunnan-Guizhou oldlands were gradually expanding north-westward and northward, leading to a drop in diversity. Those relatively nearshore areas were originally quite suit-able for the Hirnantia Fauna, but subsequently became too shallow to support any normal marine benthic faunas like the Hirnantia Fauna. On the contrary, although those off-shore and comparatively deeper localities were originally not very suitable for the Hirnantia Fauna while it was de-veloping in nearshore environments, with ongoing regional tectonic movement they eventually became the most favor-able places for the Hirnantia Fauna to flourish. Thus, there is a definite macroevolutionary trend for the Hirnantia Fauna on the Upper Yangtze Platform to gradually move from nearshore to offshore localities.
The geographical distribution of the Hirnantia Fauna on
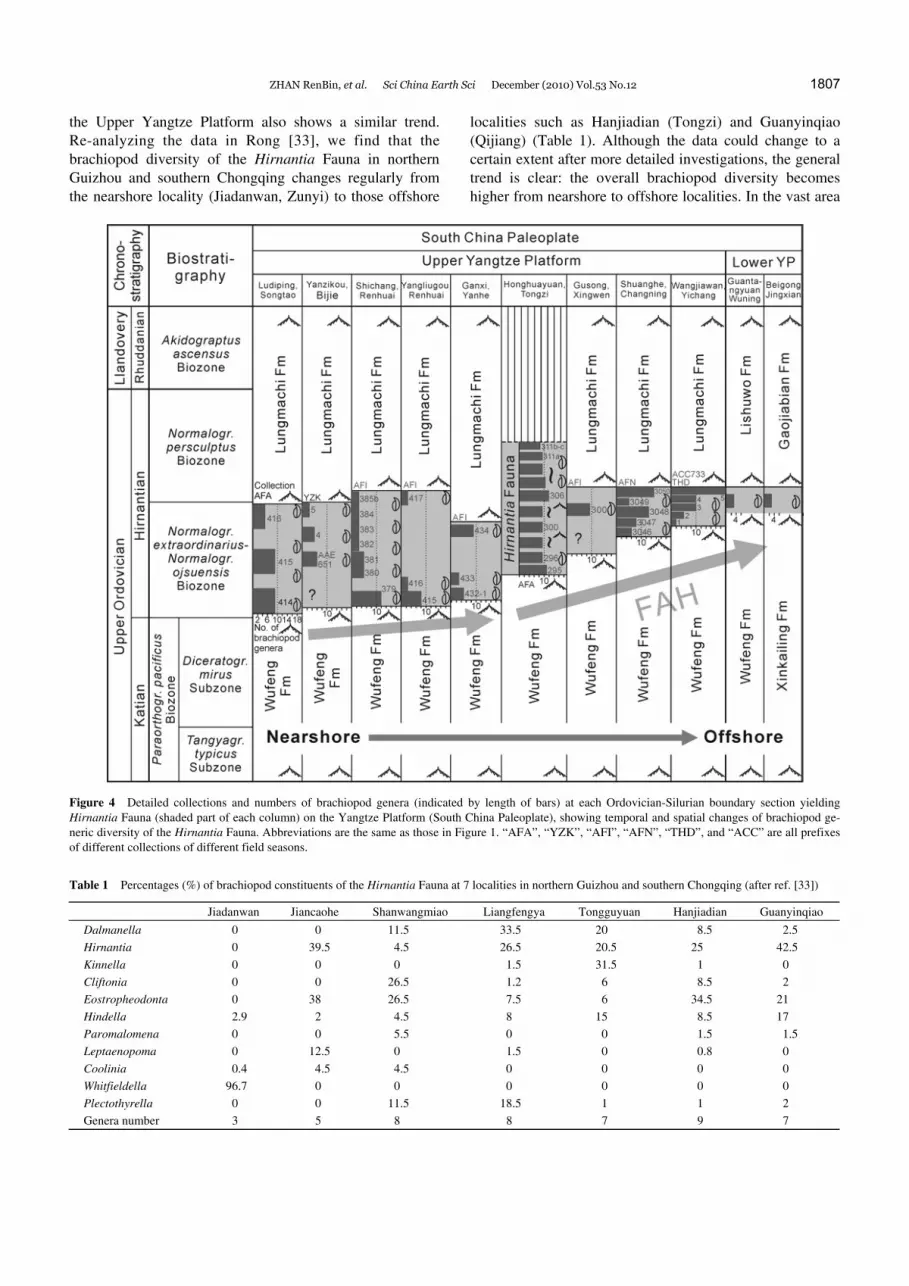
ZHAN RenBin, et al. Sci China Earth Sci December (2010) Vol.53 No.12 1807
the Upper Yangtze Platform also shows a similar trend. Re-analyzing the data in Rong [33], we find that the brachiopod diversity of the Hirnantia Fauna in northern Guizhou and southern Chongqing changes regularly from the nearshore locality (Jiadanwan, Zunyi) to those offshore
localities such as Hanjiadian (Tongzi) and Guanyinqiao (Qijiang) (Table 1). Although the data could change to a certain extent after more detailed investigations, the general trend is clear: the overall brachiopod diversity becomes higher from nearshore to offshore localities. In the vast area
Figure 4 Detailed collections and numbers of brachiopod genera (indicated by length of bars) at each Ordovician-Silurian boundary section yielding Hirnantia Fauna (shaded part of each column) on the Yangtze Platform (South China Paleoplate), showing temporal and spatial changes of brachiopod ge-neric diversity of the Hirnantia Fauna. Abbreviations are the same as those in Figure 1. “AFA”, “YZK”, “AFI”, “AFN”, “THD”, and “ACC” are all prefixes of different collections of different field seasons.
Table 1 Percentages (%) of brachiopod constituents of the Hirnantia Fauna at 7 localities in northern Guizhou and southern Chongqing (after ref. [33])
Jiadanwan Jiancaohe Shanwangmiao Liangfengya Tongguyuan Hanjiadian Guanyinqiao
Dalmanella 0 0 11.5 33.5 20 8.5 2.5
Hirnantia 0 39.5 4.5 26.5 20.5 25 42.5
Kinnella 0 0 0 1.5 31.5 1 0
Cliftonia 0 0 26.5 1.2 6 8.5 2
Eostropheodonta 0 38 26.5 7.5 6 34.5 21
Hindella 2.9 2 4.5 8 15 8.5 17
Paromalomena 0 0 5.5 0 0 1.5 1.5
Leptaenopoma 0 12.5 0 1.5 0 0.8 0
Coolinia 0.4 4.5 4.5 0 0 0 0
Whitfieldella 96.7 0 0 0 0 0 0
Plectothyrella 0 0 11.5 18.5 1 1 2
Genera number 3 5 8 8 7 9 7

1808 ZHAN RenBin, et al. Sci China Earth Sci December (2010) Vol.53 No.12
of the normal marine benthic regime (from Honghuayuan of northern Guizhou to Guanyinqiao of southern Chongqing), diversity variations are of small scale that is irrelevant to the changing of substrates or the distance away from the shore-line.
3 Paleoecological differentiation of the Hirnan-tia Fauna on the Upper Yangtze Platform
The wide distribution and rich abundance and diversity of the Hirnantia Fauna on the Upper Yangtze Platform of South China paleoplate enable us to further investigate its paleoecological differentiation. Based on data published by Rong [33] (see its text-fig. 7), a multivariate dataset is es-tablished taking the seven localities as different cases and the ten representative brachiopod genera as variables. Prin-cipal Component Analysis (PCA) and Cluster Analysis (CA) have been conducted using the computer software PAST [34] (Figure 5). The results confirm that the Hirnantia Fauna could be differentiated into the Whitfieldella and Hirnantia communities [33], and furthermore CA also shows that these two communities are distinctly different from each other with very low similarity coefficient. The eponymous genus of the Whitfieldella Community consti-tutes about 97% of the collection, accompanied by very rare individuals of Coolinia and Hindella, which indicates that this community lived in a shallow water benthic regime corresponding to upper BA2 [33]. The Hirnantia Commu-nity could be further differentiated into the Hirnan-tia-Eostropheodonta Subcommunity and the Dalmanella- Hirnantia Subcommunity. The Hirnantia-Eostropheodonta Subcommunity has a low brachiopod diversity, and is dominated by the two eponymous genera (accounting for more than 80% of the collection). In contrast, the Dal-manella-Hirnantia Subcommunity has no components that are more than one-third of the total, although it also has some “dominant” constituents. It is thought to have occu-pied a lower BA2 to BA3 environment with argillaceous or lime mud substrates [33].
The Kuanyinchiao Formation near Tiemenkan of Shuanghe town, Changning County (section 1 in Figure 3), is lithologically composed of dark grey and greyish black calcareous mudstone and argillaceous limestone. Brachio-pod fossils are very abundant and diverse, and were qualita-tively named the “Paracraniops”-Paromalomena Commu-nity [35] or the Hirnantia-Draborthis Community [7], in-terpreted as living in a lower BA3 environment. In the field seasons of 2006 and 2007, we re-measured this section, subdivided the Kuanyinchiao Formation into five layers, and made intense collections at each layer. Altogether more than 3000 fossil specimens of different groups were ob-tained, amongst which more than 90% are brachiopod fos-sils (3114 specimens). Nineteen brachiopod genera are recognized in this preliminary study, representing a typical
occurrence of the Hirnantia Fauna. Using numerical methods (e.g., PAST), two brachiopod-dominated associations are provisionally recognized, i.e., the Dalmanella-Kinnella As-sociation and the Mirorthis Association. The Dal-manella-Kinnella Association is thought to be first flour-ished in a comparatively deeper environment (correspond-ing to lower BA3), and subsequently shifting into a normal marine benthic regime corresponding to middle BA3 [29]. The Mirorthis Association, developed in the upper Kuany-inchiao Formation at Shuanghe, represents a short deepen-ing during the accumulation of the Kuanyinchiao Formation at this particular site, and might have lived in an environ-
Figure 5 Principal component analysis (a) and cluster analysis (b) of seven representatives of Hirnantia Fauna in northern Guizhou and southern Chongqing. a, Hirnantia Community including the Hirnantia-Eostro- pheodonta Subcommunity and the Dalmanella-Hirnantia Subcommunity; b, Whitfieldella Community. 1. Dalmanella; 2. Hirnantia; 3. Kinnella; 4. Cliftonia; 5. Eostropheodonta; 6. Hindella; 7. Paromalomena; 8. Lep-taenopoma; 9. Coolinia; 10. Whitfieldella; 11. Plectothyrella.

ZHAN RenBin, et al. Sci China Earth Sci December (2010) Vol.53 No.12 1809
ment possibly as deep as upper BA4 with argillaceous sub-strates.
In summary, on the Upper Yangtze Platform at both nearshore and offshore localities, the brachiopod diversity of the Hirnantia Fauna displays relatively simple trends, and always changes in a single direction. For example, at those nearshore localities close to the oldlands, the diversity of the Hirnantia Fauna consistently decreases, whereas it is progressively increases at offshore localities, such as those in the Yichang district [1].
4 Concluding remarks
The Hirnantia Fauna developed in normal marine benthic regimes at many localities in the world of intermediate to low latitudes in latest Ordovician times. It is a typical cool water shelly fauna with its entire macroevolutionary history limited between the two episodes of the end-Ordovician mass extinction [36]. Located in low latitudes with argilla-ceous or lime mud benthic environments, the Upper Yang-tze Platform of the South China paleoplate was eminently suitable for the Hirnantia Fauna to become the best devel-oped, most widely distributed, longest in stratigraphical range, and most complicated in paleoecological differentia-tion, and it is in this region that the Hirnantia Fauna has been most thoroughly studied [21].
The case study conducted on the Upper Yangtze Plat-form shows that taxonomic composition and ecological dif-ferentiation of the Hirnantia Fauna changes from one local-ity to another, and at almost each locality the fauna has its own unique macroevolutionary story. The general trend is that the closer to the shoreline, the lower the FAD, and the more restricted the brachiopod diversity; the further away from the shoreline, the shorter the stratigraphical range, and the higher the brachiopod diversity. At nearshore localities, the diversity of the fauna decreases stratigraphically upward. In contrast, at offshore localities, the brachiopod diversity increases upward although the strata are much thinner and the stratigraphical range is considerably shorter. Generally, brachiopod diversity at offshore localities is much higher than that of nearshore localities. The Hirnantia Fauna of the Kuanyinchiao Formation at Honghuayuan of Tongzi, north-ern Guizhou is an exception to these trends; in this unique case, the brachiopod diversity of the Hirnantia Fauna main-tains an intermediate level (with 9–11 genera).
Qualitative and numerical analyses on the paleoecology of the Hirnantia Fauna indicate that a series of diagnostic brachiopod-dominated communities and subcommunities developed on the Upper Yangtze Platform from nearshore to offshore settings. At some offshore localities, the fauna was manifested by different subcommunities or associations corresponding to minor fluctuation of environments (e.g., at Shuanghe of Changning), but they are all closely related in brachiopod constituents.
The analysis conducted in this paper shows that the FAD of the Hirnantia Fauna becomes progressively higher from the Upper to Lower Yangtze Platform, from nearshore to offshore localities, whereas its paleoecological environment increases in water depth. These trends provide support to the concept that the deep water environment could be an important place for benthic communities to have survived the end-Ordovician mass extinction [19].
Fan Junxuan and Wu Rongchang are thanked for their considerable assis-tance in the field. Ian Percival publishes with permission of the Director, Geological Survey of New South Wales, NSW Department of Primary Industries. Two reviewers and Dr. Miao Desui helped us a lot in improving the manuscript both scientifically and linguistically. This study was sup-ported by Chinese Academy of Sciences (Grant No. KZCX2-YW-Q05-01), National Natural Science Foundation of China (Grant No. 40825006), and Ministry of Science and Technology of China (Grant Nos. 2006CB806402, 2006FY120300-5).
1 Rong J Y. Brachiopods of latest Ordovician in the Yichang district, western Hubei, central China. In: Nanjing Institute of Geology and Palaeontology, Academia Sinica, ed. Stratigraphy and Palaeontology of Systemic Boundaries in China, Ordovician-Silurian Boundary (1). Hefei: Anhui Science and Technology Press, 1984. 111–176
2 Lu Y H. Subdivision and Correlation of the Ordovician rocks of southern China (in Chinese with English abstract). Beijing: Geologi-cal Publishing House, 1959. 113
3 Elles G L. The age of the Hirnant Beds. Geol Mag, 1922, 59: 409–414
4 Temple J T. Upper Ordovician brachiopods from Poland and Britain. Acta Palaeontol Polon, 1965, 10: 379–450
5 Rong J Y. The Hirnantia fauna of China with comments on the Or-dovician-Silurian boundary (in Chinese). J Stratigr, 1979, 3: 1–8
6 Rong J Y, Harper D A T. A global synthesis of the latest Ordovician Hirnantian brachiopod faunas. Trans Royal Soc Edinburgh Earth Sci, 1988, 79: 384–402
7 Rong J Y, Chen X, Harper D A T. The latest Ordovician Hirnantia Fauna (Brachiopoda) in time and space. Lethaia, 2002, 35: 231–249
8 Harper D A T, Williams S H. A relict Ordovician brachiopod fauna from the Parakidograptus acuminatus Biozone (Lower Silurian) of the English Lake District. Lethaia, 2002, 35: 71–78
9 Rong J Y, Zhan R B. Late Ordovician brachiopod mass extinction of South China. In: Rong J Y, Fang Z J, eds. Biotic Mass Extinction and Recovery Afterwards—Evidence from South China (in Chinese with English abstract). Hefei: China University of Science and Techno- logy Press, 2004. 71–96
10 Chen X, Rong J Y, Mitchell C E, et al. Late Ordovician to earliest Silurian graptolite and brachiopod zonation from Yangtze Region, South China with a global correlation. Geol Mag, 2000, 137: 623–650
11 Rong J Y, Chen X. Faunal differentiation, biofacies and lithofacies pattern of late Ordovician (Ashgillian) in South China (in Chinese with English abstract). Acta Palaeontol Sin, 1987, 26: 507–535
12 Underwood C J, Crowley S F, Marshall J D, et al. High-resolution carbon isotope stratigraphy of the basal Silurian Stratotype (Dob’s Linn, Scotland) and its global correlation. J Geol Soc, 1997, 154: 709–718
13 Bergström S M, Saltzman M M, Schmitz B. First record of the Hirnantian (Upper Ordovician) δ13C excursion in the North American Midcontinent and its regional implications. Geol Mag, 2006, 143: 657–678
14 Finney S C, Berry W B N, Cooper J D, et al. Late Ordovician mass extinction: A new perspective from stratigraphic sections in central Nevada. Geology, 1999, 27: 215–218

1810 ZHAN RenBin, et al. Sci China Earth Sci December (2010) Vol.53 No.12
15 Brenchley P J, Carden G A, Hints L, et al. High resolution isotope stratigraphy of Late Ordovician sequences: Constraints on the timing of bioevents and environmental changes associated with mass extinc-tion and glaciations. Bull Geol Soc Amer, 2003, 115: 89–104
16 Kaljo D, Hints L, Männik P, et al. The succession of Hirnantian events based on data from Baltica: Brachiopods, chitinozoans, cono-donts, and carbon isotopes. Estonian J Earth Sci, 2008, 57: 197–218
17 Kaljo D, Martma T, Saadre T. Post-Hunnebergian Ordovician carbon isotope trend in Baltoscandia, its environmental implications and some similarities with that of Nevada. Palaeogeog Palaeoclimat Pa-laeoecol, 2007, 245: 138–155
18 Chen X, Rong J Y, Fan J X, et al. The Global boundary Stratotype Section and Point (GSSP) for the base of the Hirnantian Stage (the uppermost of the Ordovician System). Episodes, 2006, 29: 183–196
19 Rong J Y, Zhan R B, Huang B, et al. Discovery of an end Ordovician deep water brachiopod fauna at Yuhang, Hangzhou, Zhejiang, China (in Chinese). Chin Sci Bull, 2007, 52: 2632–2637
20 Rong J Y, Huang B, Zhan R B, et al. Latest Ordovician brachiopod and trilobite assemblage from Yuhang, northern Zhejiang, East China: A window on Hirnantian deep-water benthos. Histor Biol, 2008, 20: 137–148
21 Harper D A T, Rong J Y. Completeness of the Hirnantian brachiopod record: Spatial heterogeneity through the end Ordovician extinction event. Lethaia, 2008, 41: 195–197
22 Sheng S F. Late Ordovician trilobites of Sichuan and Guizhou prov-inces with comments on the lower and upper boundaries of the Upper Ordovician Series (in Chinese). Newslett Palaeontol Soc China, 1962, 12: 46–47
23 Sheng S F. The age of the Dalmanitina Beds in China. In: Lu Y H, ed. Subdivision and Correlation of the Ordovician Rocks of Southern China (in Chinese with English abstract). Beijing: Geological Pub-lishing House, 1974. 53–95
24 Zhan R B, Jin J. Ordovician-Early Silurian (Llandovery) Stratigraphy and Palaeontology of the Upper Yangtze Platform, South China. Bei-jing: Science Press, 2007. 169
25 Zhang W T, Xu H K, Chen X, et al. Ordovician of northern Guizhou. In: Nanjing Institute of Geology and Palaeontology, ed. Palaeozoic Rocks of Northern Guizhou (in Chinese). Nanjing: Nanjing Institute
of Geology and Palaeontology, 1964. 33–78 26 Guizhou Geologic Bureau. Regional Geology of Guizhou Province
(in Chinese with English summary). Geol Memoir, 1987, 1: 1–698 27 Rong J Y, Harper D A T, Zhan R B, et al. Kassinella-Christiania
Associations in the early Ashgill Foliomena brachiopod fauna of South China. Lethaia, 1994, 27: 19–28
28 Rong J Y, Li R Y. A silicified Hirnantia fauna (Latest Ordovician brachiopods) from Guizhou, southwest China. J Paleontol, 1999, 73: 831–849
29 Li G P, Zhan R B, Wu R C. Response of Hirnantia Fauna to the en-vironmental changes before the second phase of Late Ordovician mass extinction: Example from the Kuanyinchiao Formation at Shuanghe, southern Sichuan, southwest China (in Chinese with Eng-lish abstract). Geol J China Univ, 2009, 15: 304–317
30 Mu E Z, Zhu Z L, Chen J Y, et al. Ordovician around Shuanghe, Changning, Sichuan (in Chinese). J Stratigr, 1978, 2: 105–215
31 Fang Y T, Liang S J, Zhang D L, et al. Stratigraphy and Graptolite Fauna of Lishuwo Formation from Wuning, Jiangxi (in Chinese with English summary). Nanjing: Nanjing University Publishing House, 1990. 155
32 Cao Q Y, Feng H Z, Fang Y T, et al. Graptolite zonation in Wufeng Formation of Xiushui Gorge, Jiangxi Province (in Chinese with Eng-lish abstract). Geol J China Univ, 1998, 4: 318–323
33 Rong J Y. Ecostratigraphy and community analysis of the Late Ordo-vician and Silurian in southern China (in Chinese with English sum-mary). In: Palaeontological Society of China, ed. Selected Papers from the 13th and 14th Annual Conventions of Palaeontological So-ciety of China. Hefei: Anhui Science and Technology Publishing House, 1986. 1–22
34 Hammer Ø, Harper D A T. PAST—Palaeontological statistics, 2008, ver. 1.89. http://folk.uio.no/ohammer/past (Denmark)
35 Wang Y, Boucot A J, Rong J Y, et al. Community palaeoecology as a geologic tool—The Chinese Ashgillian-Eifelian (latest Ordovician through early Middle Devonian) as an example. Spe Pap Geol Soc Am, 1987, 211: 1–100
36 Rong J Y, Harper D A T. Brachiopod survival and recovery from lat-est Ordovician mass extinction in South China. Geol J, 1999 34: 321–348





























