Embed Size (px)
Citation preview

БИОЛОГИЯ И МЕДИЦИНА BIOLOGY AND MEDICINE
Андреенко, Емилия, Мима Николова. Особености в психометричната характеристика на пловдив-ски студенти
Andreenko, Emilia, Mima Nikolova. Peculiarities in psyhometric characteristics of Plovdiv students ____ 947 Андреенко, Емилия, Мима Николова. Сравнително проучване на основни кефалометрични пока-
затели при студенти от град Пловдив Andreenko, Emilia, Mima Nikolova. Comparative study of basic cephalometric indexes of university
students from Plovdiv _____________________________________________________________ 955 Давидова, Румяна, Стилянка Йочкова, Нана Нарлиева. Активни подходи в обучението по анато-
мия: Новата роля на преподавателите Davidova, Rumyana, Stilyanka Jochkova, Nana Narlieva. Active approaches in learning Anatomy: New
role of the faculty _________________________________________________________________ 965 Йорданова, Албена, Емилия Стоименова, Христина Петкова, Румен Тодоров, Йордан
Думанов, Здравко Лалчев. Влияние на сърфактантния протеин Sp-C върху повърхностното поведение на липидни монослоеве
Jordanova, Albena, Emilia Stoimenova, Hristina Petkova, Roumen Todorov, Jordan Doumanov, Zdravko Lalchev. Influence of surfactant protein SP-C on the surface behaviour of lipid monolayers 973
Йочкова, Стилянка, Дорка Вучидолова, Румяна Давидова. Разпределение на Brain-Derived
Neurotrophin Factor във вътрешните слоеве на развиваща се постнатална ретина на плъх Jochkova, Stilyanka, Dorka Vouchidolova, Rumyana Davidova. Expression of Brain-Derived
Neurotrophin Factor in the inner layers of developing postnatal rat retina _____________________ 984 Коев, Койчо, Иван Бакалски, Диана Калъчева. Морфо-анатомични особености на сладката сте-
вия (Stevia Repaudiana Bertoni) в условията на България Koev, Kojcho, Ivan Bakalski, Diana Kalacheva. Morfo-anatomical characteristics of Sweet Stevia
(Stevia Repaudiana Bertoni) in Bulgarian conditions ______________________________________ 994 Митова, Зорка. Антропометрична характеристика при деца и подрастващи според възрастово-
половата им принадлежност Mitova, Zorka. Anthropometrical characterization in children and adolescents according to their age –
gender belonging _________________________________________________________________ 1006 Младенова, Силвия, Мима Николова. Разпространение на свръхтеглото и затлъстяването сред
смолянските деца и подрастващи Mladenova, Silvia, Mima Nikolova. Distribution of the overweight and obesity among Smolyan children
and adolescents _________________________________________________________________ 1025

Пешева, М., О. Кръстанова, М. Димитров, П. Венков. Нов краткосрочен тест за определяне на
канцерогенни замърсители в почвени проби у нас Pesheva, M., O. Krastanova, M. Dimotrov, P. Venkov. New Ty1 test to Study Environmental Soil
Pollution with Carcinogens _________________________________________________________ 1034 Сренц, Агоп, Христо Христов, Диана Кирин. Състояние и проблеми за съхранение на защитена
територия “Голямо Смолянско езеро” Srentz, Agop, Khristo Khristov, Diana Kirin. Condition and problems for conservation on the preserve
territory “Big Smolyan’s Lake” _______________________________________________________ 1046 Стайкова, Жени, Иван Чавдаров, Таня Търновска, Галя Цолова. Здравни аспекти на затлъстя-
ването сред населението в гр. Кърджали Staykova, Jenny, Ivan Chavdarov, Tanya Ternovska, Galya Tsolova. Health aspects of the farrening
into a population in the town of Kardzhali ______________________________________________ 1054 Станоев, Иван. Санаториално лечение на болни от анкилозиращ спондилартрит в курорта Велинград Stanoev, Ivan. Sanatorium treatment of patients with ankylosing spondyloarthritis in Velingrad ________ 1060 Стефанова, Надежда, Веселина Москова, Таня Топузова-Христова, Ралица Тодорова, Ивайло
Иванов, Машенка Димитрова. Дипептидил ептидаза ІV – структура и функции. Флуоресцент-ни хисто- и цитохимични методи за локализация на ензима
Stefanova, Nadejda, Veselina Moskova, Tanja Topouzova-Hristova, Ralica Todorova, Ivailo Ivanov, Mashenka Dimitrova. Dipeptidylpeptidase IV – structure and functions. Fluorescent histo- and cytochemical methods for the enzyme localization _______________________________________ 1064
Стоев, Рачо, Силвия Младенова. Секуларни промени във физическото развитие на смолянски
подрастващи Stoev, Racho, Silvia Mladenova. Secular growth changes in physical development of Smolyan
adolescents __________________________________________________________________ 1076 Томова, Елена, Виктория Сарафян. Хетерогенна тъканна експресия на кръвно-груповите антиге-
ни А и В при гризачи Tomova, Elena, Victoria Sarafian. Heterogenic tissue expression of blood group antigens A and B in
rodents ________________________________________________________________________ 1082 Ekici, Nuran, Feruzan Dane. Calcium oxalate crystals in vegetative and floral organs of Galanthus sp.
(Amaryllidaceae) _________________________________________________________________ 1090 Kirin, Diana. Biological diversity and ecological evaluation of the fresh water ecosystems from the Arda
river, Bulgaria ___________________________________________________________________ 1099 Stanoev, Ivan, Diana Stanoeva. Studying the effect of ultraphonophoresis with “Vurdon”-gel and Feloran-
zel in diseases with degenerative dystrophy of spinal cord _________________________________ 1111 Stanoev, Ivan. Studying the effectiveness of climatictherapy and aerosolotherapy with mineral water in
patients with chronic bronchitis in condition of Velingrad __________________________________ 1116

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 947
SCIENTIFIC PAPERS
ОСОБЕНОСТИ В ПСИХОМЕТРИЧНАТА ХАРАКТЕРИСТИКА НА ПЛОВДИВСКИ СТУДЕНТИ
Емилия Андреенко, Мима Николова
Пловдивски Университет ”Паисий Хилендарски”
PECULIARITIES IN PSYHOMETRIC CHARACTERISTICS OF PLOVDIV STUDENTS
E. Andreenko, M. Nikolova
Plovdiv University “Paisii Hilendarski”
ABSTRACT: The aim of this research is to characterize psychometric status of young men and wimin from Plovdiv and to characterize specifically intersexual differences. Object of the study are 136 clinically healthy individuals (88 girls and 48 boys), students from different majors of the University of Plovdiv, at the average age of 20-21 years. Six-teen psychometric traits (12 direct and 4 derivative) were measured regarding: velocity of sensorimotor reaction in a given rate; maximum movement frequency with the hand (tap-ing test), tremometry (static and dynamic), quickness and skillfulness of fingers and atten-tion intensity (two correcture tests). The data are statistically processed. The authenticity of the obtained results was checked by means of ANOVA – tukey test for independent ex-tract of significance level p ≤ 0,05. There are established significant more fast velocity of sensorimotor reaction and maximum movement frequency with the hand in young men. The representatives of both sexes show high attention intensity, as though the same de-gree of emotional stability.
Key words: psychometric traits, students, intersexual differences ВЪВЕДЕНИЕ Основните направления в съвременните психологически проучвания е
изследване промените в нивото на дееспособността на индивидите, в зави-симост от тяхната възрастово-полова принадлежност, социална, спортна или трудова активност. Често тези промени се използуват за характеризи-ране степента на нервно-психическото натоварване и настъпващите проце-си на умора. В резултат на изучаването на психо-физиологическите харак-теристики на различни групи от населението, се получават показателни, сензитивни и обобщени оценки, които имат своето значение за по-пълното изясняване на взаимоотношенията между човека и заобикалящата го среда [1, 3, 4]. Във връзка с това се прилагат комплекс от различни психодиагнос-тични методики [2].

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 948
(ЦЕЛ) на настоящето проучване е да се изследват специфичните особености на
висшата нервна дейност на млади мъже и жени, студенти от Пловдивския Университет, както се характеризира спецификата на междуполовите разли-чия в техния психометричен статус.
МАТЕРИАЛ И МЕТОДИ Обект на изследване са 136 клинично здрави индивиди (88 момичета и 48
момчета), студенти от различни специалности на Пловдивския университет, на средна възраст 21 години. Проучени са общо 16 психометрични признака, от които12 директно измерени показатели и 4 производни индекси, изчислени по съответни формули.
1. Скорост на зрително-двигателна реакция в зададен темп (брой верни сигнали). С помощта на рефлексометъра на “Piorkowski – I.C. – 6” бе измерена бързината на реакцията и зрително-моторната координация при честота на подаваните светлинни сигнали – 75 имп./мин (по-бавен темп) и 107 имп./мин (по-бърз темп) и време на експозиция – 0,5 минути. Изследването автоматично прекъсва след изтичане на времето. За скоростта на реакцията се съди по броя на точните попадения на апарата по време на изпълнението на задачата, които се регистрират от вградения брояч.
2. Максимална честота на движение с ръката Тепинг – тест (бр. удари за 10 сек). Измерването се извърши на полския електро-механичен тестиращ апарат за психо-физиологически и технически изследвания “Тремометър”. Нанасят се възможно най-бързо удари с металически молив върху горната пластинка на апарата за 10 сек. Ръката е опряна с лакът върху масата, а дви-женията се изпълняват само от китката. Броят на ударите се регистрират ав-томатично, чрез вграден в апарата брояч.
3. Емоционална устойчивост – тремометрия (брой грешки) Измерването се извърши на същия апарат – “Тремометър”. На горната
страна на апарата е монтирана металическа таблица с тестовете за изс-ледване на статичен и динамичен тремор.
а/ Статичен тремор – изследваният държи с изпъната ръка метали-чески молив в отвърстие на апарата (5 мм) в продължение на 10 секунди, стремейки се да не допира до металическите стени. Всяка докосване задейст-вува звънец и се съпровожда със светване на контролна лампа за предуп-реждение относно допусната грешка. Резултатът се отчита по броя на докос-ванията (грешките) до стените на отвърстието.
б/ динамичен тремор – тестовете за динамичен тремор представляват прорези с начупена конфигурация, в различни форми и размери, създаващи различна трудност. Изследваният трябва така да води металическия молив по отделните прорези, че да не се получава допиране по техните стени. Всяко докосване на “писеца” до уреда се отчита за грешка. Скоростта на придвиж-ване на молива в прорезите е произволна.
4. Сръчност на пръстите (в секунди) Психомоториката от тази сфера бе оценена с помощта на специално
блок-табло. На него има гнезда за поставяне на щифтчета с три вида големи-на. Задачата е да се поставят тези щифтчета възможно най-бързо в техните гнезда. Мярка за сръчността на пръстите е времето, отчетено със секундомер, за което ще бъдат нареди всички извадени щифтчета обратно по местата им.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 949
5. Концентрация и устойчивост на вниманието Изследвани са качествата на вниманието чрез проследяване възмож-
ността то да бъде съсредоточено върху даден тестов материал за по- про-дължително време. Използувани са два вида коректурни теста: Тест 1 – “Преплетени линии” (по проф. К. К. Платонов) – изследваното лице прос-ледява само с очи (без помощта на молив или пръст) една след друга 25 преплитащи се линии, като в края на всяка от тях вписва цифрата, с която ли-нията е номерирана в началото. Времето за изпълнение е 5 минути, което се засича със секундомер.
Тест 2 – “Намиране на геометрични фигури” – на едната страна на този тест (страна А) има обозначени с цифри 105 различни по форма, очерта-ния и защрихованост геометрични фигури. На другата страна на теста (страна В) са обозначени само 25 от всички, но вече кодирани с друга номерация. За-дачата е последователно да се запомни всяка от 25-те геометрични фигури от страна В, да се намери цифрата, с която са обозначена на страна А, след кое-то да се нанесе цифрата в съответна таблица. Времето за изпълнение е 7 минути, което се засича със секундомер.
След приключване на двата теста се отчитат следните показатели : - брой на вярно попълнените знаци, брой на грешните знаци, брой на
пропуснатите знаци.За да се определят продуктивността и точността на из-пълнението на задачите. бяха изчислени коефициенти за точност (А) и за продуктивност (Р) при работата с двата теста.
А / коеф. за точност / = N / М. N – количеството на зачертаните при изпълнението на задачата знаци. М – количеството знаци, които по изискване на инструкцията трябва да се
зачертаят. Коеф. А има максимална стойност = 1
P / коеф. за продуктивност / = A. C. A – коефициент за точност. C – количеството на всички прегледани знаци от теста. Коеф. Р има максимална стойност = 25
Достоверността на получените резултати е проверена чрез ANOVA – Tukey test за независима извадка при уровен на значимост p ≤ 0,05.
РЕЗУЛТАТИ И ОБСЪЖДАНЕ • Дейността на всеки човек изобилствува със ситуации, в които решава-
нето на определени двигателни задачи зависи изключително от способността му да реагира възможно най-бързо на определени сигнали (дразнители). Средните стойности на проучваните скоростни параметри на нервната систе-ма при студентите, както и тяхното съпоставяне при двата пола са представе-ни на Таблица 1 и Фигура 1.
Бързината на реагиране зависи много от функционалното състояние на централната нервна система и от особеностите на висшата нервна дейност. При съпоставяне на данните от нашето изследване установяваме по-добри резултати от психодвигателните задачи за бързина при младите мъже. Нали-це са достоверни междуполови различия и при двата зададени темпа за реа-гиране – 75 имп./мин. и 105 имп.мин (p ≤ 0,05). Способността на младите мъ-

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 950
же да извършват по-голямо количество движения за кратко време (Тепинг –тест) е също достоверно по-висока в сравнение с тази на младите жени (средно 75,97 ±13,11 бр. почуквания при младите мъже и 65,83 ± 8,85 бр. по-чуквания при младите жени – p ≤ 0,05). Следователно при студентите се ре-гистрира по-голяма функционална подвижност и скорост на протичане на нер-вните процеси. Резултатите от последната скоростна задача – Сръчност на пръстите, при изпълнението на която главни компоненти са освен бързина и координация на движенията на ръцете и съобразителност, показват липса на достоверни различия между студентите и студентките. Средното време за подреждане на щифчетата при момчетате е 39,17 ± 8,16 сек., а при момичета-та – 37,60 ±5,78 сек (p ≥ 0,05). Явно по този параметър младите жени израв-няват своята скорост на реагиране и умението си да контролират собствения двигателен темп с тези на младите мъже.
• Специфична и важна черта на човека е неговата емоционална устойчи-вост, тясно свързана със способността за невъзприемчивост към емоционал-ни фактори, оказващи отрицателно въздействие върху психическото му със-тояние. Получените данни за Тремометрия в нашето изследване са предста-вени на Таблица 2 и Фигура 2. Въпреки регистрираните колебания в общата психо-емоционална устойчивост при двата пола, различията в стойностите на статичната и динамична тремометрия не достигат статистическа величина (p ≥ 0,05). Явно е, че при младите мъже и жени е налице близка координация на микродвиженията, а и следователно близки процеси на активно вътрешно за-държане.
• За допълнително характеризиране състоянието на психичната сфера на студентите бе изследвано устойчивостта и концентрацията на внимание-то. Както е известно вниманието е важно условие за успешното протичане на всяка човешка дейност. Експерименталното му изследване включва проучва-не на неговите характеристики. При това е важно практическите научни изс-ледвания да се ръководят от йерархичната концепция за вниманието (Мечков, К., 1987), съгласно която неговите качества трябва да бъдат разбирани като последователно подредени във възходящ йерархичен ред звена, в които вся-ко следващо предполага предшествуващото и го включва: концентрация – ус-тойчивост – обхват –превключваемост – разпределяемост и т.н. Според този йерархичен ред основното свойство на вниманието, условие за всички оста-нали е концентрацията. Устойчивостта е концентрация с определена продъл-жителност, обхватът е концентрация и устойчивост върху повече от един обект и пр. За целите на нашето изследване използувахме два коректурни теста: Тест1 “преплетени линии” и Тест 2 “геометрични фигури”.
На Таблица 3 и Фигура 3 са представени средните стойности на показа-телите за внимание при студентите от двата пола. Резултатите показват лип-сата на достоверни различия между тях, както по отношение на броя вярно зачертани знаци, така и по отношение на броя сгрешени и пропуснати знаци и при двата теста. Единствено прави изключение по-ниската при момчетата стойност на сгрешените отговори при работа с втория тест в сравнение с тази на момичетата, която достига статистическа величина (p ≤ 0,05). Тези резул-тати са показателни и отразяват наличието на общи и специфични особенос-ти в качествата на вниманието на младите мъже и жени. Близката и висока интензивност на протичане на психическата дейност и мисловните процеси и при двата пола са най- вероятно възрастово обусловена и могат да се свър-

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 951
жат с характера на тяхното образователно и интелектуално ниво. Използва-нето на различни средства и дейности в процеса на обучението им спомага за трениране на различните страни на вниманието- интензивност, устойчивост, обем, разпределеност и др.
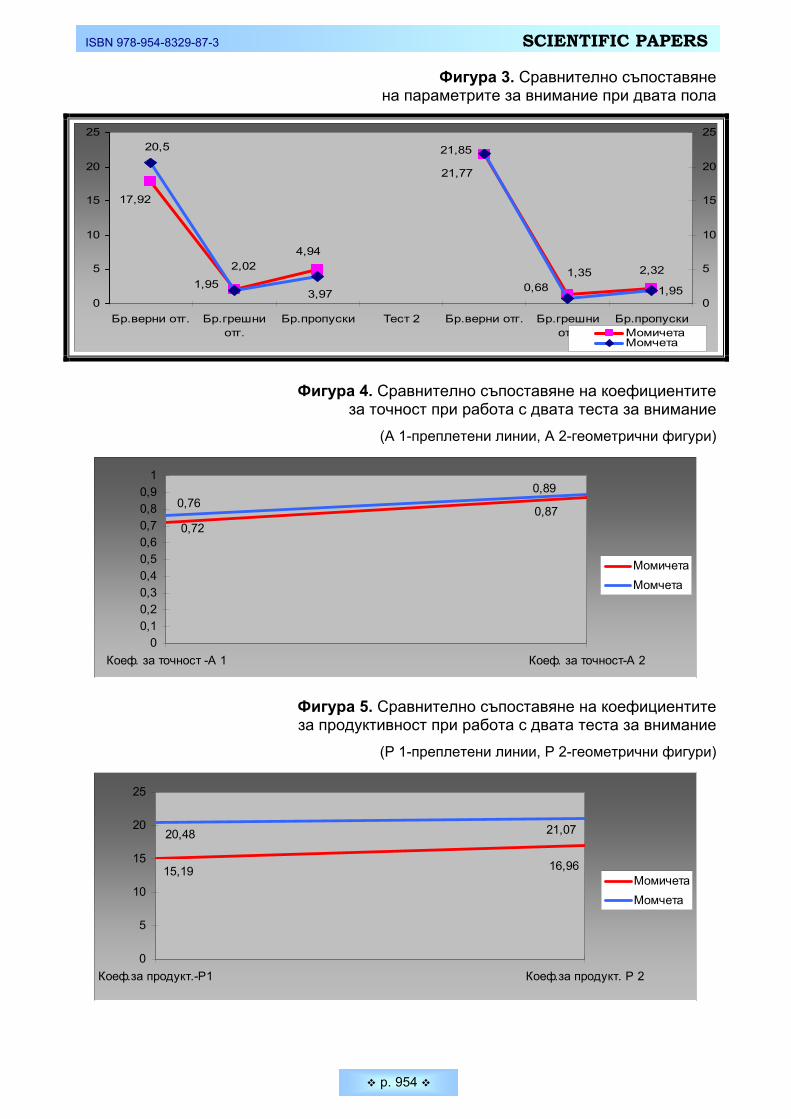
Резултатите от двата най-обобщаващи показатели за внимание – коефи-циентите за точност и продуктивност при работа с тестовете (Фигура 4 и Фигура 5) отново не показват наличие на достоверни различия между студентите и студентките. Решаването на мисловните задачи, обаче от втория геометричен тест протича с по-висока степен на изразеност на вниманието и при по-активна аналитико-синтетична дейност, за което съдим по получените по-добри коефи-циенти за точност и продуктивност и при двата пола при втория тест.
ЗАКЛЮЧЕНИЕ 1. Направена е характеристика на основния психометричен статус на
млади мъже и жени, студенти от Пловдивския университет, на средна възраст 21 години. Установени са достоверно по-голяма бързина на реакция и под-вижност на висшата нервна дейност (измерена с броя на максималните дви-жения ) при младите мъже.
2. За разлика от скоростните параметри на нервната дейност, при които се откроява тяхната зависимост от пола, при съобразителността и сръчността липсват ярки различия.
3. Установени са близки процеси на активно вътрешно задържане и еднаква степен на стабилност в емоционалната сфера при изследваните млади мъже и жени.
4. Представителите и на двата пола показват висок уровен на концент-рация и устойчивост на вниманието.
ЛИТЕРАТУРА [1] Pavlova, T. A. Principles and methods of occupational psycho-physiological
selection of deep water divers. Fiziol.Zh., 1991, 37, 1020107. Mick, J., J. Brick. Bit-slice Microprocessor Design. New York: MCGraw – Hill Book Company, 1980.
[2] Николов, Б., Г. Енева. Ръководство за работа с апаратура за психологи-чески изследвания. Благоевград, ВПИ, 1991, 163с.
[3] Пирьов, Г. Д. Психологически аспекти на екологията. Психология, 1991, 1, 3-5.
[4] Русинова, В., Л. Василева. Професионален стрес: Теоретичен и експери-ментален модел на Купър. Психология, 1995, 4, 39-48. гл. ас. д-р Eмилия Андреенко катедра Анатомия и Физиология на човека, ПУ “П. Хилендарски” тел: +359 32 261 533 е-mail: [email protected] проф. дбн Мима Николова катедра Анатомия и Физиология на човека, ПУ “П. Хилендарски” тел: +359 32 261 510 е-mail: [email protected]
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 952
ПРИЛОЖЕНИЯ
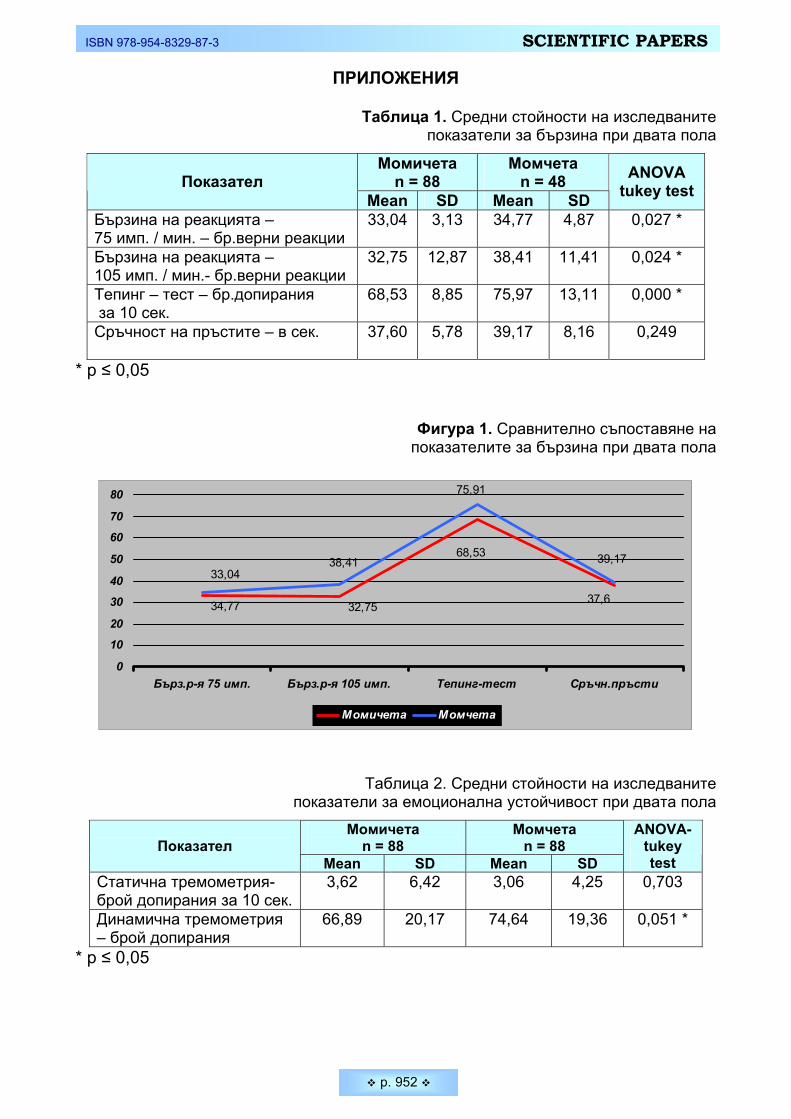
Таблица 1. Средни стойности на изследваните показатели за бързина при двата пола
Момичета n = 88
Момчета n = 48 Показател
Mean SD Mean SD
ANOVA tukey test
Бързина на реакцията – 75 имп. / мин. – бр.верни реакции
33,04 3,13 34,77 4,87 0,027 *
Бързина на реакцията – 105 имп. / мин.- бр.верни реакции
32,75 12,87 38,41 11,41 0,024 *
Тепинг – тест – бр.допирания за 10 сек.
68,53 8,85 75,97 13,11 0,000 *
Сръчност на пръстите – в сек.
37,60 5,78 39,17 8,16 0,249
* p ≤ 0,05
Фигура 1. Сравнително съпоставяне на показателите за бързина при двата пола
37,6
68,53
33,04
32,75
39,17
75,91
38,41
34,77
0
10
20
30
40
50
60
70
80
Бърз.р-я 75 имп. Бърз.р-я 105 имп. Тепинг-тест Сръчн.пръсти
Момичета Момчета
Таблица 2. Средни стойности на изследваните показатели за емоционална устойчивост при двата пола
Момичета n = 88
Момчета n = 88 Показател
Mean SD Mean SD
ANOVA- tukey test
Статична тремометрия- брой допирания за 10 сек.
3,62 6,42 3,06 4,25 0,703
Динамична тремометрия – брой допирания
66,89 20,17 74,64 19,36 0,051 *
* p ≤ 0,05
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 953
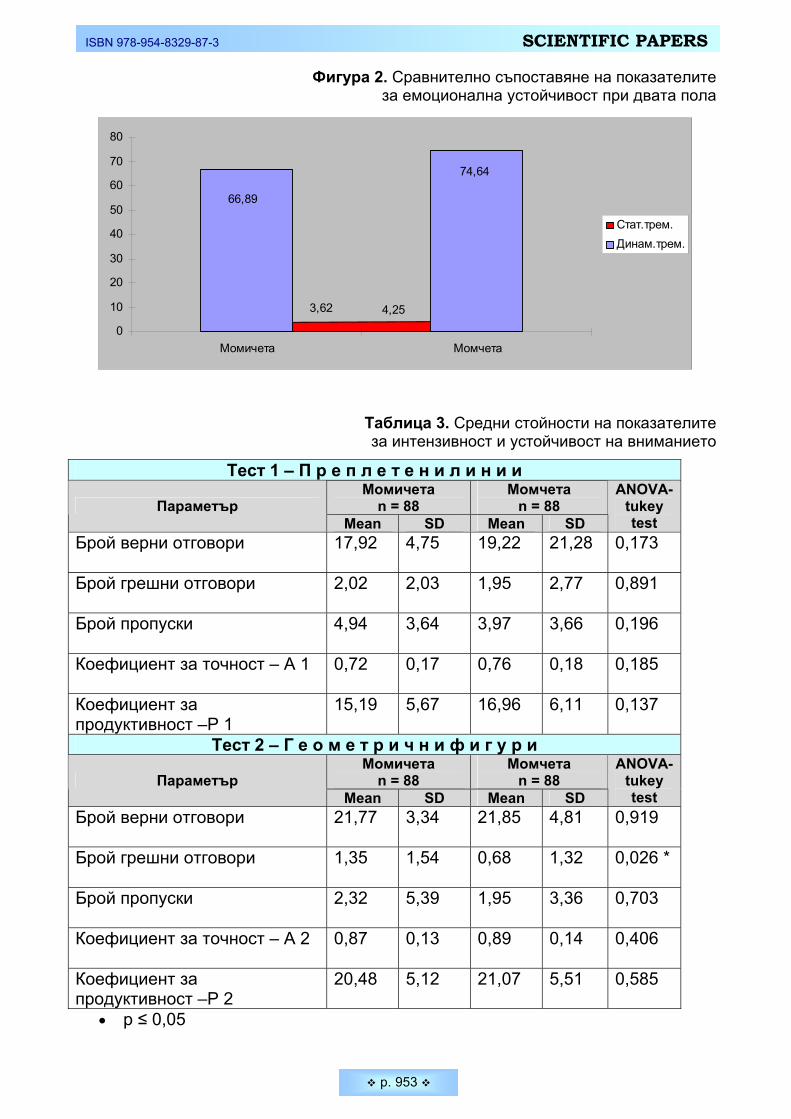
Фигура 2. Сравнително съпоставяне на показателите за емоционална устойчивост при двата пола
4,253,62
66,89
74,64
0
10
20
30
40
50
60
70
80
Момичета Момчета
Стат.трем.Динам.трем.
Таблица 3. Средни стойности на показателите за интензивност и устойчивост на вниманието
Тест 1 – П р е п л е т е н и л и н и и Момичета
n = 88 Момчета
n = 88 Параметър Mean SD Mean SD
ANOVA-tukey test
Брой верни отговори
17,92 4,75 19,22 21,28 0,173
Брой грешни отговори
2,02 2,03 1,95 2,77 0,891
Брой пропуски
4,94 3,64 3,97 3,66 0,196
Коефициент за точност – А 1
0,72 0,17 0,76 0,18 0,185
Коефициент за продуктивност –Р 1
15,19 5,67 16,96 6,11 0,137
Тест 2 – Г е о м е т р и ч н и ф и г у р и Момичета
n = 88 Момчета
n = 88 Параметър Mean SD Mean SD
ANOVA-tukey test
Брой верни отговори
21,77 3,34 21,85 4,81 0,919
Брой грешни отговори
1,35 1,54 0,68 1,32 0,026 *
Брой пропуски
2,32 5,39 1,95 3,36 0,703
Коефициент за точност – А 2
0,87 0,13 0,89 0,14 0,406
Коефициент за продуктивност –Р 2
20,48 5,12 21,07 5,51 0,585
• p ≤ 0,05
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 954
Фигура 3. Сравнително съпоставяне на параметрите за внимание при двата пола
2,024,94
17,92
21,77
1,35 2,321,95
3,97
20,5 21,85
0,68 1,950
5
10
15
20
25
Бр.верни отг. Бр.грешниотг.
Бр.пропуски Тест 2 Бр.верни отг. Бр.грешниотг.
Бр.пропуски0
5
10
15
20
25
МомичетаМомчета
Фигура 4. Сравнително съпоставяне на коефициентите за точност при работа с двата теста за внимание
(А 1-преплетени линии, А 2-геометрични фигури)
0,870,72
0,760,89
00,10,20,30,40,50,60,70,80,9
1
Коеф. за точност -А 1 Коеф. за точност-А 2
МомичетаМомчета
Фигура 5. Сравнително съпоставяне на коефициентите за продуктивност при работа с двата теста за внимание
(Р 1-преплетени линии, Р 2-геометрични фигури)
15,19 16,96
20,48 21,07
0
5
10
15
20
25
Коеф.за продукт.-Р1 Коеф.за продукт. Р 2
МомичетаМомчета

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 955
SCIENTIFIC PAPERS
СРАВНИТЕЛНО ПРОУЧВАНЕ НА ОСНОВНИ КЕФАЛОМЕТРИЧНИ ПОКАЗАТЕЛИ
ПРИ СТУДЕНТИ ОТ ГРАД ПЛОВДИВ
Емилия Андреенко, Мима Николова Пловдивски Университет ”Паисий Хилендарски”
COMPARATIVE STUDY OF BASIC CEPHALOMETRIC INDEXES
OF UNIVERSITY STUDENTS FROM PLOVDIV
E. Andreenko, M. Nikolova Plovdiv University “Paisii Hilendarski”
ABSTRACT: The aim of this research is to compare basically cephalometric features and indexes of young men and wimin from Plovdiv and to characterize specifically inter-sexual differences. Object of the study are 136 clinically healthy individuals (88 girls and 48 boys), students from different majors of the University of Plovdiv, at the average age of 20-21 years. The data of 8 cephalometric indicators and 3 basic indexes have been ana-lyzed, as well as their distribution by categories according to the rubrics generally ac-cepted in anthropology. Statistically significant intersexes differences were established ac-cording to the all indicators. The results show that the young men and wimin have mainly middle head (mesocephal), narrow nose (leptorrin) and long face (leptoprosop) so to wimen the leptorrin and the leptoprosop are stronger marked.
Key words: cephalometry, proportions, categories, students.ecological ВЪВЕДЕНИЕ Целенасочените проучвания на физическия облик на различни групи от
българското население са колкото традиционни, толкова и актуални, поради необходимостта от събирането на нови данни и използването на нови мето-дически подходи. Телесните и лицеви размери заемат важно място в интерп-ретацията на антропометричните данни на индивидите, те се променят в хода на онтогенезата и са специфични за различните възрастови периоди [1, 2, 3]. Основен принцип при снемането на метричните признаци на главата и лицето е измерването да се извършва без никакъв натиск върху меките тъкани. На-личието на такива меки, еластични и податливи форми в областта на главата на живия човек налагат строгото използване на определените мерни точки, тъй като често се касае да се установят минимални различия и нюанси. Абсо-лютните стойности на дадените метрични признаци, обаче, не са достатъчни да ни ориентират за взаимоотношенията им с други белези, поради което не могат да дадат ясни указания относно формата на съответната част от тяло-

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 956
то, към която те принадлежат. Това налага често в антропометричните проуч-вания на главата и лицето да се използват релативни величини и индекси, ко-ито дават по-цялостна и ясна представа за размерите, формата и пропорции-те на съответната телесна част.
(ЦЕЛ) на настоящето проучване е да се сравнят някои основни кефалометрич-
ни показатели и индекси на млади мъже и жени, студенти от град Пловдив, като се характеризират специфичните особености на междуполовите разли-чия, както и да се съпоставят данните на студентите от двата пола със стан-дартните стойности за съответната възрастова група от българското населе-ние [Янев, Б и кол., 1982].
МАТЕРИАЛ И МЕТОДИ Обект на изследване са 136 клинично здрави индивиди (88 момичета и 48
момчета), студенти от различни специалности на Пловдивския университет, на средна възраст 21 години. Използван е стандартен антропометричен инст-рументариум – малък допирателен пергел с обхват от 0-300 mm, малък плъз-гащ перегел с обхват от 0-250 mm, милиметрова лента. Анализирани са дан-ните на 8 кефалометрични показатели: обиколка глава, дължина глава (g-op), ширина глава (eu-eu), морфологична височина лице (n-gn), ширина лице (zy-zy), ширина долна челюст (go-go), височина нос (n- sn), ширина нос (al-al). Из-числени са и 3 основни кефалометрични индекса :
1. Индекс на главата [ ширина глава / дължина глава х 1 00 ]. 2. Индекс на лицето [ височина лице / ширина лице х 1 00 ]. 3. Индекс на носа [ ширина нос / височина нос х 100 ]. Анализирано е разпределението на студентите по категории, според об-
щоприетите в антропологията рубрикаци. Данните са обработени статисти-чески. Достоверността на получените резултати е проверена чрез ANOVA – tukey test за независима извадка при уровен на значимост p ≤ 0,05.
РЕЗУЛТАТИ И ОБСЪЖДАНЕ Средните стойности на проучваните кефалометрични признаци при сту-
дентите от двата пола са представени на Таблица 1 и Фигура 1. Средната обиколка на главата на младите мъже е 58,31±1,70 см, а на
младите жени – 55,72±1,72 см. При студентите обиколката на главата се дви-жи от 55,00 до 62,50 см, а при студентките – от 51,00 до 59,50 см. Средната обиколка на главата при мъжете е с 2,59 см или 4,44 % по-голяма от тази на жените. Различията между двата пола са статистически достоверни (p ≤ 0,05).
Данните от изследваните абсолютни главови размери – дължина глава и ширина глава, показват достоверно по-ниски стойности при младите жени. Средната дължина на главата при тях е 18,63 ± 0,75 см. Стойността на този признак се движи в границите от 16,40 см до 20,40 см. Средната дължина на главата при младите мъже е 19,29 ±1,57 см и е с 0,66 см или 3,42 % по-голяма от тази на жените. Стойността на този признака при студентите варира от 11,50 см до 21,50 см – т.е. вариабилитетът при мъжете е по-голям.
Средната ширина на главата при студентките е 14,80 ±0,70 см, а при студентите - 15,40 ± 0,98 см. Разликата между двата пола е 0,60 см или 3,89 % и е статистически значима в полза на мъжете (p ≤ 0,05).

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 957
Средната морфологична височина на лицето при младите жени е 11,47 ± 0,62 см, а при младите мъже – 12,25 ± 0,84 см. Разликата в средните стойности между двата пола е 0,78 см или 6,37 % и е достоверно по-висока при мъжете
Средната скулова ширина (бизигоматична) на лицето при младите жени е 12,20 ± 0,80 см, а при младите мъже - 13,01 ± 0,87 см. Разликата между най- малката и най-голямата измерена скулова ширина при студентите е 46 мм, а при студентките – 42 мм. Средната скулова ширина при мъжете е достоверно по-голяма с 0,81 см или 6,23 % от тази на жените (p ≤ 0,05).
По отношение и на другия ширинен признак на лицето – ширина долна челюст, тенденцията се запазва същата. По-голяма средна стойност имат младите мъже- средната ширина на долната челюст при тях е 11,20 ± 0,93 см и е в границите между 7,30 см и 13,00 см. При младите жени средната стойност на същия признак е 10,48 ± 0,66 см, като варира от 8,50 см до 12,50 см. Ширината между ъглите на долната челюст при студентките е с 0,72 см или 6,43 % по-малка в сравнение с тази на мъжете. Междуполовите различия са статистически значими (p ≤ 0,05).
Що се отнася до размерите на носа, резултатите показват достоверни различия между двата пола по отношение на ширината на носа (p ≤ 0,05) и незначими различия по отношение на неговата височина (p ≥ 0,05). Средната височина на носа при младите мъже е 5,19 ± 0,49 см при 4,10 см минимална и 6,00 см максимално измерена височина на носа. Съответните стойности при младите жени са 5,05 ± 0,47 см, минимум и максимум 3,10 см – 6,20 см. Средната ширина на носа при студентите варира от 3,00 см до 4,10 см при средна стойност 3,43 ± 0,28 см, а при студентките от 2,50 см до 4,10 см при средна стойност 3,16 ± 0,28 см. Ширината на носа при младите жени е досто-верно по-малка с 0,27 см или 7,87 % от тази на младите мъже (p ≤ 0,05).
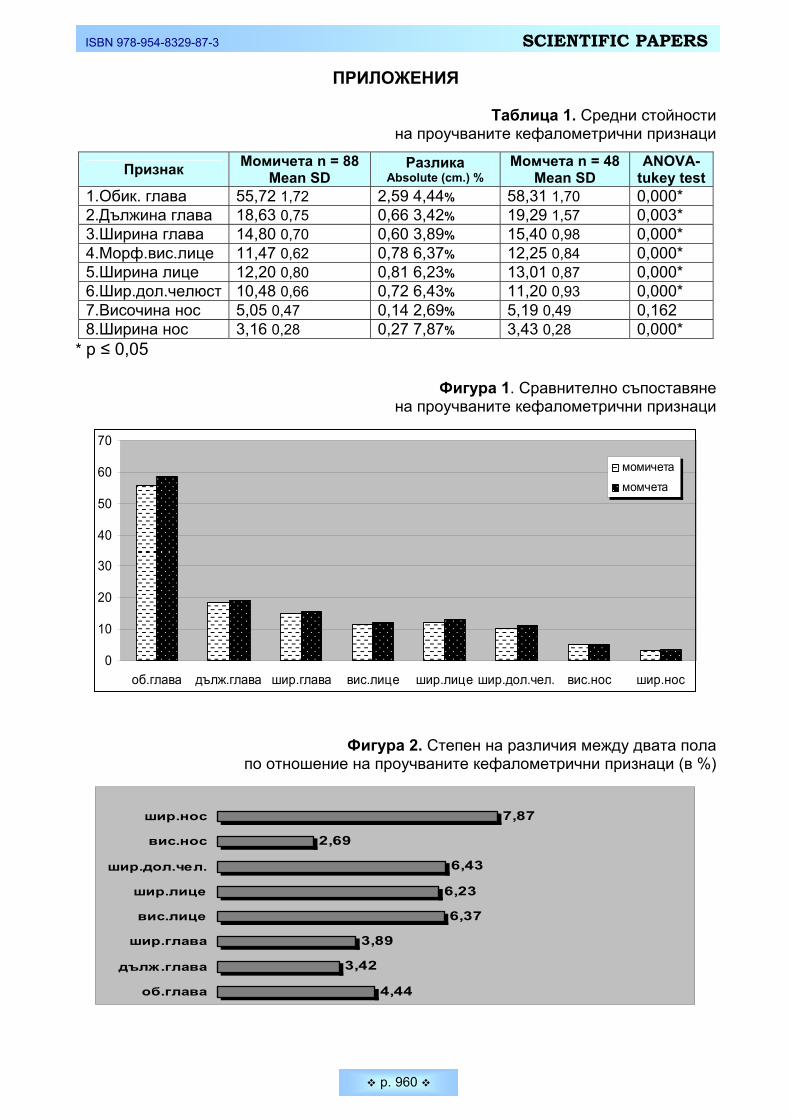
Получените резултати от кефалометричните измервания показват, че средните стойности на всички показатели са достоверно по-високи при мла-дите мъже в сравнение с младите жени, с изключение на височината на носа. Твърде интересно бе да установим по кои от признаците различията в абсо-лютните стойности между двата пола са по-отчетливо изразени. На Фигура 2 е представено степента на различие (в %) между студентите и студентките по отношение на главовите и лицеви показатели. Младите мъже и жени се раз-личават в по-голяма степен по отношение на своята ширина на носа и в приз-наците, характеризиращи лицето (височина лице, ширина лице, ширина долна челюст) и в по-малка степен в признаците, характеризиращи главата (дължи-на глава, ширина глава, обиколка глава).
Въз основа на получените абсолютните размери на главата и лицето бя-ха изчислени и три основни кефалометрични индекса, даващи представа как-то за взаимоотношението между отделните признаци, така и за формата и пропорциите на дадени части от главата (Таблица 2). Резултатите показват, че според средните стойности на индексите младите мъже и жени имат средна глава, тесен нос и дълго лице, като при жените дължината на лицето е по-изразена.
По-съществени различия между студентите от двата пола се установяват при съпоставяне на процентното им разпределение по категории според об-щоприетите рубрикации на тези индекси (Таблица 3).

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 958
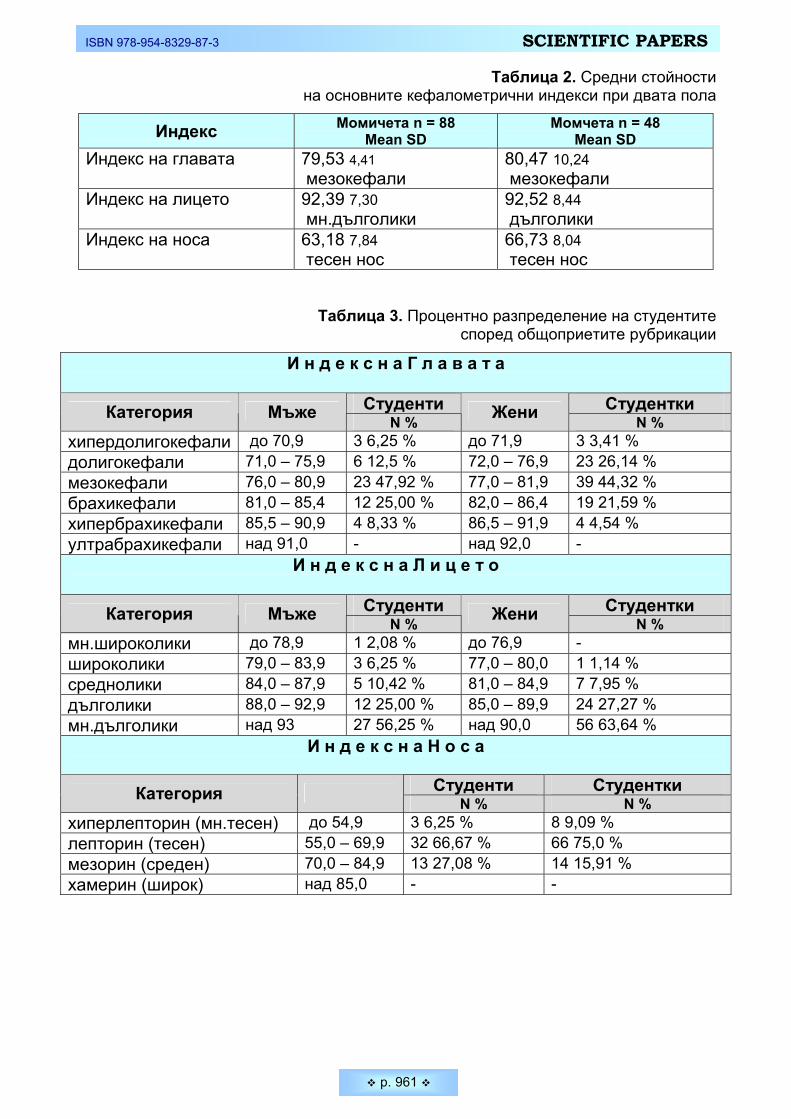
Индексът на главата представлява изразеното в проценти съотношение между ширината и дължината на главата. Той ни дава представа за формата на главата, гледана отгоре. Това е един от първите въведени в антропологич-ната наука индекси, който и до днес остава един от най-често изчисляваните и най-много използвани величини при антропологично изучаване на народите и расите (Фигура 3). При младите мъже най-голям процент са мезокефалите (47,92%), следвани от брахикефалите (25,00 %). Почти половината от младите жени също се отнасят към категорията - средноглави (44,32 %), но тенденция-та при тях е в обратна посока – към долигокефалията (26,14 %).
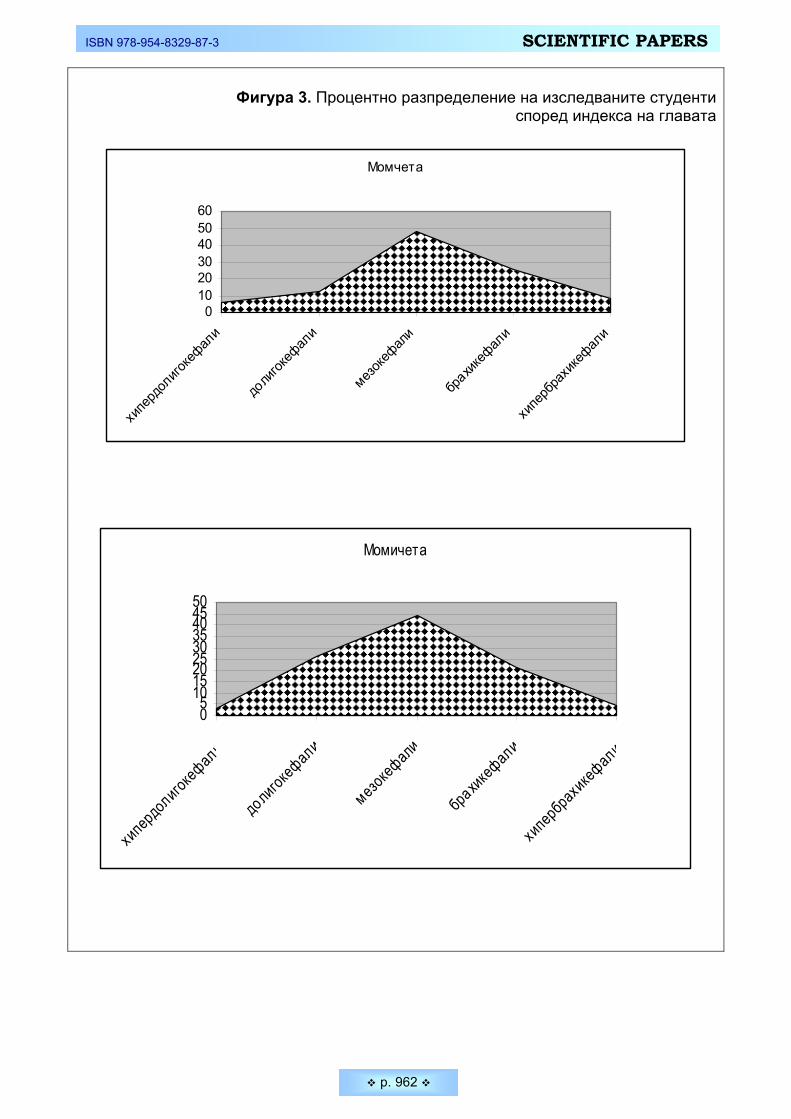
Индексът на лицето представлява морфологичната височина на лицето, изразена в проценти спрямо ширината на лицето (скуловата ширина). Инте-ресни са резултатите, които ни дава разпределението на изследваните млади мъже и жени в петте възприети от антропологичната наука категории (Фигура 4). Разпределението на студентите и от двата пола е явно асиметрично и силно изместено в посока към лептопрозопия (дълголики). Най-голям брой измерени млади мъже спадат към категорията много дълголики - 56,25 %. Този процент е още по-висок при младите жени - 63,64 %. Следователно при бол-шинството индивиди и от двата пола относително намалява скуловата шири-на при по-голяма абсолютна морфологична височина на лицето, в следствие от което по-силно се подчертава дълголикостта – т.е налице е тенденция към грацилизация и лептопрозопия. Тази тенденция е по-ясно подчертана при младите жени.
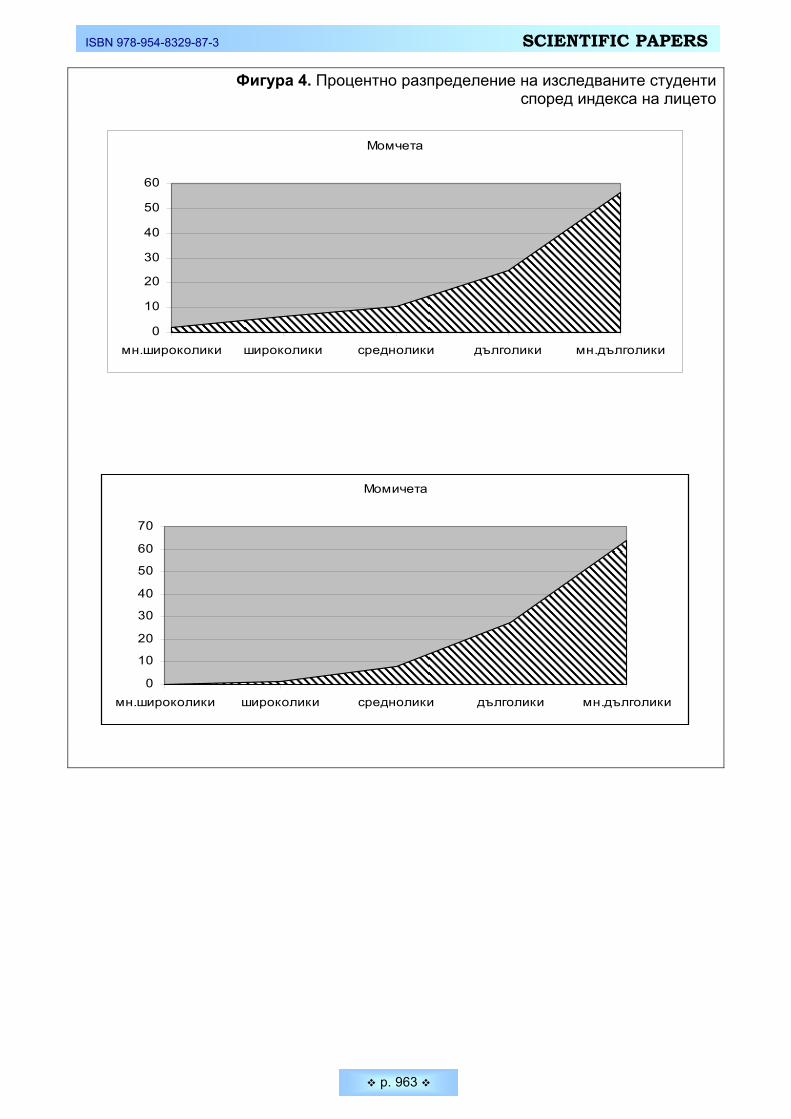
Третият проучван индекс в нашето изследване е индексът на носа, кой-то представлява изразеното в проценти отношение на ширината на носа спрямо височината му (Фигура 5). Поради важното значение, което се отдава в антропологията на размерите на носа, данните в литературата са твърде богати. Както е известно формата, строежът и пропорциите на носа представ-ляват извънредно важен антропологичен белег. Редица особености на носа отличават трите големи човешки раси една от друга (европеидната, монголо-идната и негро-австралоидната). Високите и тесни носове (лепториния) са ха-рактерни за европеидните антропологични типове. Мезоринията се среща главно всред монголоидните народи, а за негро-австралоидната раса е ти-пично силно изразеното широконосие (хамериния). Голямото мнозинство от измерените млади мъже и жени спадат към категорията лепториния, като при жените тенденцията към тесноносие е по-силно изразена - 75,0 % от жените и 66,67 % от мъжете. В същото време средноносите жени (мезорини) са 15,91 %, докато младите мъже от същата категория са почти два пъти повече – 27,08%. Това е най-вероятно във връзка с установената при тях по-голяма ширина на носа. Нито един от измерените от нас студенти не попадна в кате-горията на хамерините. Получените резултати за индекса на носа са напълно очаквани и закономерни, показващи характерните за европейските народи стойности.
Данните на Пловдивските студенти бяха съпоставени със стандартните стойности за съответната възрастова група от българското население по про-учването на Б.Янев (1982). В Таблица 4 сме отразили само тези признаци, за които намерихме данни в достъпната литература. Разликите в стойностите на ширина глава и ширина лице не достигат статистическа величина (p ≥ 0,05). Стойностите за обиколка глава и ширина глава на изследваните млади мъже и жени са статистически значимо по-високи от стандартните стойности (p ≤

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 959
0,05). Получените резултати ни покзват известна тенденциа към по-големи общи размери на главата при студентите предимно в сагитална равнина. За по-голямо прецизиране на резултатите, обаче е необходимо използването на данните и на другите кефалометрични показатели, с които не разполагаме в настоящата извадка.
ЗАКЛЮЧЕНИЕ 1. Направена е характеристика на основния кефалометричен статус на
млади мъже и жени, студенти от Пловдивския университет, на средна възраст 21 години. Средните стойности на всички показатели са досто-верно по-високи при младите мъже в сравнение с младите жени, с изклю-чение на височината на носа.
2. По-голяма информация за междуполова диференциация показват приз-наците, характеризиращи височината и ширината на лицето, отколкото признаците, характеризиращи големината на главата.
3. Младите мъже и жени имат предимно средна глава (брахикефалия), те-сен нос (лепториния) и дълго лице (лептопрозопия), като при жените леп-торинията и лептопрозопията са по-силно изразени.
4. От абсолютните главови размери само обиколката на главата и ширината на главата на изследваните млади мъже и жени са статистически значи-мо по-високи от стандартните стойности за българското население.
ЛИТЕРАТУРА [1] Barli, O., Elmali D., Gedik, T. Anthropometry of male and female children in
Greches in Turkey.Collegium Anthropologicum, 2005, 29 (1). [2] Кондова, Н., Зл. Филчева. Възрастови промени в кефалометричната и
кефалоскопичната характеристика на софийски деца в периода 7-11г. Journal of Anthtopology, BAS, 1999, 2, 117-124.
[3] Младенова, С., Антропологична характеристика на растежа и развитието на деца и подрастващи от Смолянски регион в съвременните условия на жи-вот. Дисертационен труд, 2003, 151 с. гл. ас. д-р Eмилия Андреенко катедра Анатомия и Физиология на човека, ПУ “П. Хилендарски” тел: +359 32 261 533 е-mail: [email protected] проф. дбн Мима Николова катедра Анатомия и Физиология на човека, ПУ “П. Хилендарски” тел: +359 32 261 510 е-mail: [email protected]
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 960
ПРИЛОЖЕНИЯ
Таблица 1. Средни стойности на проучваните кефалометрични признаци
Признак Момичета n = 88 Mean SD
Разлика Absolute (cm.) %
Момчета n = 48 Mean SD
ANOVA- tukey test
1.Обик. глава 55,72 1,72 2,59 4,44% 58,31 1,70 0,000* 2.Дължина глава 18,63 0,75 0,66 3,42% 19,29 1,57 0,003* 3.Ширина глава 14,80 0,70 0,60 3,89% 15,40 0,98 0,000* 4.Морф.вис.лице 11,47 0,62 0,78 6,37% 12,25 0,84 0,000* 5.Ширина лице 12,20 0,80 0,81 6,23% 13,01 0,87 0,000* 6.Шир.дол.челюст 10,48 0,66 0,72 6,43% 11,20 0,93 0,000* 7.Височина нос 5,05 0,47 0,14 2,69% 5,19 0,49 0,162 8.Ширина нос 3,16 0,28 0,27 7,87% 3,43 0,28 0,000*
* p ≤ 0,05
Фигура 1. Сравнително съпоставяне на проучваните кефалометрични признаци
0
10
20
30
40
50
60
70
об.глава дълж.глава шир.глава вис.лице шир.лице шир.дол.чел. вис.нос шир.нос
момичета момчета
Фигура 2. Степен на различия между двата пола по отношение на проучваните кефалометрични признаци (в %)
4,44
3,42
3,89
6,37
6,23
6,43
2,69
7,87
об.глава
дълж .глава
шир.глава
вис.лице
шир.лице
шир.дол.чел.
вис.нос
шир.нос
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 961
Таблица 2. Средни стойности на основните кефалометрични индекси при двата пола
Индекс Момичета n = 88 Mean SD
Момчета n = 48 Mean SD
Индекс на главата 79,53 4,41 мезокефали
80,47 10,24 мезокефали
Индекс на лицето 92,39 7,30 мн.дълголики
92,52 8,44 дълголики
Индекс на носа 63,18 7,84 тесен нос
66,73 8,04 тесен нос
Таблица 3. Процентно разпределение на студентите според общоприетите рубрикации
И н д е к с н а Г л а в а т а
Студенти Студентки Категория Мъже N % Жени N % хипердолигокефали до 70,9 3 6,25 % до 71,9 3 3,41 % долигокефали 71,0 – 75,9 6 12,5 % 72,0 – 76,9 23 26,14 % мезокефали 76,0 – 80,9 23 47,92 % 77,0 – 81,9 39 44,32 % брахикефали 81,0 – 85,4 12 25,00 % 82,0 – 86,4 19 21,59 % хипербрахикефали 85,5 – 90,9 4 8,33 % 86,5 – 91,9 4 4,54 % ултрабрахикефали над 91,0 - над 92,0 -
И н д е к с н а Л и ц е т о
Студенти Студентки Категория Мъже N % Жени N % мн.широколики до 78,9 1 2,08 % до 76,9 - широколики 79,0 – 83,9 3 6,25 % 77,0 – 80,0 1 1,14 % среднолики 84,0 – 87,9 5 10,42 % 81,0 – 84,9 7 7,95 % дълголики 88,0 – 92,9 12 25,00 % 85,0 – 89,9 24 27,27 % мн.дълголики над 93 27 56,25 % над 90,0 56 63,64 %
И н д е к с н а Н о с а
Студенти Студентки Категория N % N % хиперлепторин (мн.тесен) до 54,9 3 6,25 % 8 9,09 % лепторин (тесен) 55,0 – 69,9 32 66,67 % 66 75,0 % мезорин (среден) 70,0 – 84,9 13 27,08 % 14 15,91 % хамерин (широк) над 85,0 - -
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 962
Фигура 3. Процентно разпределение на изследваните студенти
според индекса на главата
Момчета
0102030405060
хипердолигокефали
долигокефали
мезокефали
брахикефали
хипербрахикефали
Момичета
0510
1520253035404550
хипердолигокеф
али
долигокефали
мезокефали
брахике
фали
хипербрахикеф
али
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 963
Фигура 4. Процентно разпределение на изследваните студенти според индекса на лицето
Момчета
0
10
20
30
40
50
60
мн.широколики широколики среднолики дълголики мн.дълголики
Момичета
0
10
20
30
40
50
60
70
мн.широколики широколики среднолики дълголики мн.дълголики
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 964
Фигура 5. Процентно разпределение на изследваните студенти според индекса на носа
Момчета
0
10
20
30
40
50
60
70
80
мн.тесен тесен среден широк
момичета
0
10
20
30
40
50
60
70
80
мн.тесен тесен среден широк
Таблица 4. Сравнително съпоставяне на някои кефалометрични признаци със стандартните стойности
за съответната възрастова група от българското население
М о м и ч е та М о м ч е та
Признак Наши из-следвания
Mean SD
Янев и кол.
Mean SD
ANOVA tukey test
Наши изследвания
Mean SD
Янев и кол.
Mean SD
ANOVA tukey test
обик.глава 55,72 1,72 54,70 2,00 p ≤ 0,05 58,31 1,70 56,90 2,00 p ≤ 0,05 дълж.глава 18,63 0,75 17,90 1,00 p ≤ 0,05 19,29 1,57 18,50 1,00 p ≤ 0,05 шир.глава 14,80 0,70 14,70 1,30 p ≥0,05 15,40 0,98 15,20 1,20 p ≥0,05

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 965
SCIENTIFIC PAPERS
АКТИВНИ ПОДХОДИ В ОБУЧЕНИЕТО ПО АНАТОМИЯ: НОВАТА РОЛЯ НА ПРЕПОДАВАТЕЛИТЕ
Румяна Давидова, Стилянка Йочкова, Нана Нарлиева,
Медицински университет – Плевен
ACTIVE APPROACHES IN LEARNING ANATOMY: NEW ROLE OF THE FACULTY
Rumyana Davidova, Stilyanka Jochkova, Nana Narlieva,
Medical University of Pleven
ABSTRACT Activity of bought subjects in the learning process – students and academic staff, is
the precondition of its success. The role of the teacher and teaching is been reexamined in the new high-technological world.
The aim of the present study is to reveal the role and the specific tasks of the anat-omy teacher in the conditions of active learning.
The responsibility of the faculty in the processes of motivation, regulation and as-sessment is very important. Some fears appear among the teaching staff – fear of un-known, and loss of superiority they have in classic teaching. Some students do not accept the new way of learning but still prefer the paternal learning model.
Teaching in active learning conditions includes not only presenting the information but also other activities as setting aim and tasks, student motivation and stimulation, plan-ning the learning process, assessing student activity and teaching results.
Key words: active learning, anatomy, faculty, new role ВЪВЕДЕНИЕ Същността на обучението се изразява в процесите на учене и препо-
даване. Тези два процеса са многомерни и се изучават в педагогически, психологически, социален и технологичен аспект в тяхното единство [8]. Познаването характеристиките на обучението дава възможност на препо-давателя успешно да ръководи процеса като се основава на научни подхо-ди. Активността на двата субекта на обучение – учещия и преподавателя, е необходима предпоставка за ефективността му. [7]. В основата на учебния процес е ученето, реализирано от субекта – обучавания, но “предизвикано, организирано, насочено и ръководено от преподаването, реализирано от другия субект” – преподавателя. Учене се извършва и самостоятелно (са-мообучение), но преподаването няма смисъл, “ако не предизвиква учене, ако е изолирано от ученето” [6].

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 966
В условията на съвременния високотехнологичен свят се преоценява ро-лята на преподаването и на преподавателя в процеса на обучението. В мина-лото преподаването е било главен, даже в някои моменти единствен източник на познанието, то днес ролята му се е променила коренно, като то вече е по-мощник на ученето, който го облекчава, улеснява, стимулира и контролира [2]. Надделява схващането, че “преподаването по същество означава, да се учи другия да учи” [5]
ЦЕЛТА на настоящето проучване e на основата на обширен литературен
обзор, да се очертаят ролята и специфичните задачи на преподавателя по анатомия в условията на прилагане на активни методи на обучение на сту-дентите.
МАТЕРИАЛ И МЕТОДИ Направен е обширен литературен обзор от над 104 източника (13 на бъл-
гарски език и 91 – на английски) под ключовите думи “активно учене”, “препо-даване”, “анатомия”, “роля на преподавателя”. Източниците на английски език са от различни база-данни: Medscape, Medline, Springer, Google и др. Матери-алите са публикувани в периода 1995 – 2006 г. Резултатите са синтезирани и обобщени.
РЕЗУЛТАТИ И ОБСЪЖДАНЕ Основните функции на преподавателя във висшето учебно заведение са
обучаваща, възпитателна, организираща и изследователска [10]. Специфика-та на учебната дейност изисква студентите да се научат да учат, да се спече-лят за съвместната дейност, да разберат нашето доброжелателно отношение, доверие и уважение към личността им, да се създадат мотиви и желание за изучаване на преподаваната учебна дисциплина [1].
Ефективността на учебния процес зависи от активността на всеки инди-вид при взаимодействието на обучавания и обучаващия – двата субекта на възпитанието. Преподавателят трябва добре да познава и да се съобразява с променените нагласи, очаквания и роли на студентите в така бързо променя-щия се съвременен свят във всеки от елементите на учебния процес – моти-вация, регулация и оценяване [4].
Ролята на преподавателя за мотивирането на студентите Положителната мотивация за учене на студентите определя до голяма
степен ефективността на учебната работа и зависи от редица фактори, не на последно място от уменията на преподавателя. Мотивирането се разбира ка-то умение за “елегантно” принуждаване на студентите да превръщат целите на обучение, предначертани от преподавателя във вътрешно осъзната от тях потребност от познание [3].
Новопостъпилите студенти могат да се разпределят в три групи според отношението им към необходимостта да се учи [3].
Първата група са студенти с нестабилна мотивация за учене – често попаднали във ВУЗ поради външни причини. Много от тях са с ниско самочув-ствие или отричат напълно необходимостта от образование, не виждат полза и смисъл от това, което учат и то за тях е само загуба на време. Често демон-

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 967
стративното им пренебрежение към образованието е защитна реакция на по-малко или повече осъзната неспособност да се справят с учебните задачи.
Втората група са “прагматично ориентирани”. За тях има смисъл да се учи медицина, защото дипломата осигурява бъдещето и дава възможност за по-добра самоиздръжка, по-добър жизнен стандарт и социален статус; дипломата отваря по-широко вратите за професионална реализация и изди-гате в обществото.
Третата група са “академичен тип” младежи. За тях образованието е ценност във всяко време и късметът е на страната на знаещите. Личност-ното им развитие и стремежът към професионализъм в избраната профе-сия са важна част от житейската им философия. За тази група студенти е присъщо желанието за самопроверка, самодоказване, постигане на по-голямо самочувствие, самоувереност, независимост, самореализация и са-моуважение.
Отделните групи студенти налагат преосмисляне на организацията на учебния процес, с оглед формиране и поддържане на положителна мотивация за учене.
Някои нерешени проблеми в организацията и провеждането на учебния процес могат да изиграят роля на демотиватори, тъй като отдалечават и за-мъгляват образа на професионалиста. Такива са недостатъчния брой практи-чески знания, незачитане мнението на студентите по отношение на интереси-те и предпочитанията им, претоварване на учебните програми, недобре ба-лансирано учебно разписание и ограничен достъп до технически средства за обучение извън занятията.
Преподавателят като основен мотивиращ фактор носи голяма отговор-ност за създаване и поддържане на положителна учебна мотивация. Основ-ните подходи, чрез които да се стимулира мотивацията на студентите мо-гат да се систематизират в няколко пункта. Ограничаването на авторита-ризма мотивира чрез създаване на психоклимат, основан на доверие, разби-рателство, удоволствие от ученето. Акцентирането върху отговорността на студентите за собственото им учене формира чувство за отговорност, за удовлетворение и признание, обвързване на знанията с личните потреб-ности. Осигуряването на свобода на студентите при планиране и изпълне-ние на задачите създава чувство за отговорност, за удовлетворение и приз-нание, организационни умения.
Възможността за допълнителни консултации и общуване извън заняти-ята повлиява вътрешната удовлетвореност, води до ефект на развитие и по-зитивно критично мислене. Степенуването на задачите по трудност и до-пълнителните задачи за по-бързо справящите се довежда до последовател-ност в усилията, ефект на успеха и развитие според собствения ритъм. По-ощряването на изявата на по-притеснителните студенти мотивира ак-тивността на цялата група, повишава самочувствието и също води до удов-летвореност от изявите. Подпомагането на студентите за успешна подго-товка за изпитите подсилва сигурността в собствените възможности и спо-собности и спомага за преодоляването на изпитния стрес. Поощряването на изразяване на собствено позиция на студентите по изучавания материал формира критично мислене, търсене на допълнителна информация, преодо-ляване на консуматорското отношение към ученето, развива личностно и гражданско самосъзнание.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 968
Има някои специфични изисквания към дейността на преподавате-лите, насочена към положителната мотивация за учене на студентите.При студентите от първа група, отричащи ползата от учене, трябва да се изясни дали поведението им се дължи на нежелание или на липса на по-тенциал за учене. Важно е установяването на причините за отношени-ето им към учебния процес като условие за намиране на верния подход за формиране на положителна мотивация. Необходима е индивидуална рабо-та с насърчаване на инициативността им и възлагане на специфични за-дачи за вдъхване на увереност в собствените сили. При студентите от втора група с добър потенциал за учене, но насочен предимно към извли-чане на полза от висшето образование подходите трябва да са по-различни. Те се справят с лекота със задачите, но без да влагат амбиции и усилия, учат избирателно само онова, което ще им бъде от полза и толкова, колкото им е необходимо за един успешно взет изпит. Често са формални лидери в групата, стремят се да бъдат забелязани и да компенсират с по-казна активност пропуските в знанията си. За удовлетворяване на необхо-димостта им от изява могат да им се възлагат самостоятелни презента-ции пред групата, да се стимулира приемането на ролята на говорители при груповата работа и ръководството на занятията без преподавател. Студентите от трета група са вътрешно мотивирани за задълбочено учене. Те не се ограничават само с предложената им от преподавателя ин-формация, а търсят допълнителни източници и се отличават със сериозна теоретична подготовка. За тях трябва да се създадат условия за проверка на силите и възможностите им на практика и да обвържат теоретичните си познания с практически умения. Самостоятелно създаденият учебен продукт мотивира допълнително студентите за самоусъвършенстване, по-вишава самочувствието и увереността им, че ще се справят в бъдещата си професионална реализация.
Ролята на преподавателя в регулацията на учебния процес Регулацията изпълнява функциите на информиране, формиране и кон-
трол. Разграничават се три типа регулация: интерактивна (интегрирана в
учебната ситуация), ретроактивна (програмиране на коригиращи действия след процеса на оценяване) и проективна (включва бъдещите формиращи дейности, които в основата си са насочени към обособяване и доразвиване способностите на студентите, отколкото към преодоляване на специфични проблеми) [3] . Отговорността при регулация пада предимно върху препода-вателя. “Луксът” да я прилага му струва доста скъпо, защото той трябва да се намесва непрекъснато в процеса на учене, за да извлича информация от него и да я осъществява пълноценно. Затова са създадени стратегии за подпома-гане регулацията на процеса на обучение. Те основно целят да научат сту-дентите как да се учат, така че да се чувстват по-независими и по-продуктивни в процеса на обучение.
Всеки индивид си има своя собствена система на учене, която постепен-но е изградил. Една основна дидактическа стратегия при регулацията на обу-чението е да се преподава на студентите така, че да развиват своята са-мостоятелност и да изграждат свой собствен, личен стил на действие.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 969
За да постигнем регулация, която основно да се осъществява от човека, който се учи, трябва да имаме предвид някои педагогически приоритети. На първо място изключително важно е обсъждане на целите, които трябва да се следват и доказване, че студентите ги разбират. Студентите трябва да са на-ясно какво ще учат, защо определени дейности ще им бъдат предлагани и защо тези дейности са структурирани по определен начин; да имат идея какъв е очакваният краен продукт, какви са резултатите, които се очаква да бъдат достигнати. Особено важен е контролът на предварителните и планира-щите действието стъпки от страна на учащия. Ако един студент е спосо-бен да контролира тези стъпки, това означава, че той е в състояние да си представи мисловно действията, които би трябвало да бъдат извършени, по-нятията и теориите, които да бъдат изучени, за да бъде изпълнена успешно предложената задача. Третият важен компонент е възприемане на критери-ите и средствата за оценяване. Студентът трябва да приеме и усвои стан-дартите и критериите, според които преподавателят решава дали студентът е разбрал понятието, дали е схванал конкретната процедура, дали отношение-то му е очакваното или не.
Някои френски автори [3] предлагат т.нар. дидактически контракт, т.е договаряне между студентите и преподавателя, и между самите студенти, като това се прави свободно, по начин и правила ясни и за двете страни. Той позволява развитие на автономност при учене, уточняване на целите на обучението и избиране на учебни стратегии за достигане на целите. Об-съждането на договора с преподавателя дава възможност за свободна изя-ва на мнение, за избор, за поемане на инициатива в учебния процес. Той е също така средство за внедряване на диференциация и позволява разгра-ничаване на съдържанието на материала от начините, по които студентите ще преминат през него.
При Методът на проектите, студентите изпълняват една цялостна дейност, т.е. по този начин те са принудени да имат идея за изпълнение, да планират задачите необходими за постигане на целта, да изпълнят или да материализират идеята, да я представят в семинарната зала, да оценят про-цеса и резултата. Външната изява на целта е материализирането на нещо реално и полезно. Срокът на продължителност зависи от броя на целите и степента на уменията, които трябва да бъдат придобити. Студентите работят в малки групи. Групата е отговорна за изпълнение на проекта. Всеки индивид поема определени функции в организационната структура и изпълнява спе-цифичните за нея задачи. Могат да се формулират пет етапа на проекта: оформяне на групите, откриване на идеята, подготовка на проекта, изпълне-ние и представяне и крайно оценяване.
Професионалното израстване на медицинските студенти започва още през първата гадина на обучение. Затова обучението по анатомия може да се използва за представяне на проблеми, отнасящи се до критичното мис-лене и професионализма. Тук широко се прилага практиката на “отражението” и критичното мислене, включващо и рефлексията [12].
Истинския рефлективен процес изисква обучението да дава възмож-ност за непрекъсната отвореност към въпросите “какво”, “как” и “защо” нещата се правят. Ученето на тежкия анатомичен материал се осъществява чрез въпроси, изследване, оценяване, анализиране, теоретизиране, търсене на обратна връзка и внедряване на идеи и виждания от членовете на екипа. В

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 970
контекста на анатомията “отвореността” може да се използва като средство в изучаването на клинична анатомия, която окуражава студентите към учене по време на дисекции, където наблюдението е основно средство за теоретично-то разбиране. “Отвореността” позволява на студентите да мислят отвъд структурните отговори и дава възможност да прилагат теоретичните си позна-ния за оценяване на клинични ситуации.
Описват се 3 стадия на рефлексия [12,13]. В първия стадий студентът може да усвоява основни знания в учебни условия. Това позволява на пре-подавателя да оценява приложения учебен метод. Втората фаза включва анализ на специфични учебни дейности и демонстрира тяхната ефектив-ност върху студентското учене. Третата фаза е критично отражение, което позволява на студента и преподавателя изпробване на по-обширни когни-тивни процеси като етични проблеми свързани с практиката и вземане на решения.
Рефлективно учене в програмата по анатомия се постига успешно с из-ползването на клиничен материал в изучаването на обща анатомия. Описват се 5 основни подхода за използване на активни форми в обучението по ана-томия.
1. Самонасочено учене 2. Оценъчни стратегии 3. Иновационни учебни дейности 4. Ориентация към етични проблеми 5. Запазване на дисекциите в програмата Използването на форми на активно обучение изисква добра организация,
планиране и отдаденост. Преподавателите имат две основни роли: да об-лекчават ученето чрез задаване на индиректни стимулиращи въпроси и да дават обратна информация чрез обобщаване, откриване на пропуските и поправяне на всяка грешна информация. Стимулиращата среда е особено важна при практикуването на активни форми на учене. Преподавателите мо-гат да подпомагат тази среда чрез разчупване на леда, споделяне на очаква-нията си и даване на възможност на студентите да споделят своите, устано-вяване на основни изисквания за отсъствията, закъсненията, бавната работа, разделението, поведението; осигуряване на достъп до учебни средства в библиотеката, ръководства, помощна литература и др.
За преподавателите работили по традиционния начин, промяната от ди-дактично преподаване към самонасочено учене е не само промяна на мис-ловните нагласи, но и привикване към преподаване в такива програми.
Кои са най-честите страхове при преминаване към активни форми на обучение? Много преподаватели се страхуват, че губят чувството на пре-възходство в процеса на обучение, което имат при класическата форма. Съ-ществува също страх от неизвестното и от новата роля при сменената форма на обучение. Студентите също не приемат лесно промените. Много от тях предпочитат да следват патерналистичния начин на учене, отколкото да са отговорни за собственото си обучение.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 971
ЗАКЛЮЧЕНИЕ Същността на преподаването не се свежда само до поднасяне на ин-
формация, а включва и други действия, насочени към ученето. Преподавате-лят трябва да постави целта и да мотивира учещите, да подпомогне планира-нето на дейността и подбора на средства за реализирането ù, да подпомага и стимулира учещите, да контролира изпълнението на учебните задачи, да оце-ни дейността на учещите и резултатите им [3, 9].
Отговорните задачи на преподавателя включват също организиране на процеса, стимулиране на своите възпитаници, мотивиране за работа, при не-обходимост коригиране и оценяване резултатите от дейността им [4].
Основни личностни качества, които трябва да притежават преподава-телите по анатомия в медицинските училища са професионализъм, отго-ворност, организираност и дисциплинираност, способности за работа в екип, етичност, комуникативност, интелигентност, тактичност и съвременно мислене [4].
Новата роля на преподавателите в тази най-стара медицинска дисципли-на може да се обобщи със следните няколко характеристики и способности:
• Полагане на усилия за правилното функциониране на учебния процес • Поддържане на процеса • Мотивиране на персонала за подпомагане на учебния процес • Осигуряване на активно участие на всички страни в учебния процес • Подкрепяне на новите инициативи • Оценка на плюсовете и минусите на новите решения • Откриване на проблемите и навременно намиране на разрешения за
тях • Осигуряване на успешно завършване на работата [11] ЛИТЕРАТУРА
[1] Батоева, Д. Педагогическото общуване в учебния процес като средство за неговата оптимизация. Годишник на Висш педагогически институт, Благо-евград, 1985, том ІІ, кн. 4, стр. 27-30
[2] Герджиков, К. Дидактическа същност на преподаването. София, 1999, изд. “Фактум”, 136 с.
[3] Гюрова, В., Г. Дерменджиева, Е. Георгиев, С. Върбанова. Провокацията учебен процес. Аскони издат – София, 1997; 213 (12-20)
[4] Милчева, Хр. Взаимоотношенията в Медицински колеж в етико-деонтологичен аспект. сп. “Здравни грижи”, 2006, кн. 1, 62-67
[5] Милков, Л. Дидактика. Шумен, 2002, изд. “Ейро Клима”, 500 с. [6] Михова, М. Преподаването и ученето. Велико Търново, 2002, изд. “Астар-
та”, 219 с. [7] Петров, П. Дидактика. София, 1998, изд. “Веда Словена ЖГ”, 455 с. [8] Попов, Т. Педагогика Теория на обучението Дидактика. София, 2005, изд.
“Типографика” ООД, 526 с. [9] Попов, Т., Хр. Милчева, Г. Петрова, Принципи и методика на обучението,
изд. “Кота Принт”, Ст.Загора, 2006

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 972
[10] Смирнов, С. Д. Педагогика и психология висшего образования. Москва, 2003., изд “Академия”
[12] Barbero, M.G., M.T.A. Rosa, M.C.M. Moratalla, How to develop educational programs for health professionals, 1998, World Health Organization, Copen-hagen, 181 p.
[13] Five G, Building reflective practice into student learning, School of Biomedical science , Curtin University, 2005,
www.ecu.edu.au/conferences/tft/2002/pub/doc [14] Lachman, Nirusha, Pawlina Wojciech, Integrating professionalism in early
medical education: the theory and application of reflective practice in the anat-omy curriculum, Clinical anatomy, 2006, 19(5):456-460 д-р Румяна Иванова Давидова, гл. асистент, д-р Стилянка Димитрова Йочкова, гл асистент, доц. д-р Нана Василева Нарлиева, кмн, доцент, катедра “Анатомия, хистология и цитология” МУ – Плевен ул. “Св Климент Охридски”, 5800, Плевен Тел.: 064 884238

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 973
SCIENTIFIC PAPERS
ВЛИЯНИЕ НА СЪРФАКТАНТНИЯ ПРОТЕИН SP-C ВЪРХУ ПОВЪРХНОСТНОТО ПОВЕДЕНИE
НА ЛИПИДНИ МОНОСЛОЕВЕ
Албена Йорданова, Емилия Стоименова*, Христина Петкова**, Румен Тодоров**, Йордан Думанов*, Здравко Лалчев*
Институт по Биофизика, БАН; **Институт по физикохимия, БАН; *Катедра по биохимия, Биологически Факултет, СУ “Св. Климент Охридски”
INFLUENCE OF SURFACTANT PROTEIN SP-C ON THE SURFACE BEHAVIOR OF LIPID MONOLAYERS
Albena Jordanova, Emilia Stoimenova*, Hristina Petkova**, Roumen Todorov**,
Jordan Doumanov*, Zdravko Lalchev*
Institute of Biophysics, Bulgarian Academy of Sciences, **Institute of Physical Chemistry, Bulgarian Academy of Sciences,
* Biological Faculty, Sofia University “St. Kliment Ohridski”
ABSTRACT: The alveolar surfactant (AS) is a lipoprotein film, lining the alveoli and lowering the surface tension at the air/liquid interface thus preserving the lung from col-lapse during expiration, i.e. from respiratory distress syndrome (RDS). RDS, induced by inactivation of AS, is lipid related disorder connected with phospholipid deficiency and/or change of lining film structure. Besides phospholipids, AS includes the specific surfactant proteins SP-A, SP-B, SP-C and SP-D. In the other hand the phosphatidylethanolamines (PE) are minor lipid class in AS that may form both lamellar and non-lamellar structures under physiological conditions and thus the PE polymorphic phase behavior may have a crucial role in the arrangement and effective functioning of AS.
In this study mixed monolayers at the air-water interface formed by hydrophobic surfactant protein SP-C and lipid in different phase state (lamellar Lα and hexagonal HII) from dipalmitoleoylphosphatidylethanolamine (DPoPE) were investigated. The SP-C was isolated from porcine lung and it was detected by SDS-PAAGE. Differences in the degree of interaction between spread monolayers formed from specific surfactant protein SP-C and adsorption monolayers of DPoPE have been established. It has been shown that the dynamic surface properties (gamma max and gamma min) of mixed monolayers of SP-C+DPoPE (in Lα and HII phases) are significantly different and follow entirely different ten-dency during continuous compression/decompression depending on the lipid phase state.
Key words: alveolar surfactant, phospholipid monolayer, surfactant proteins, monolayers

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 974
ВЪВЕДЕНИЕ Способността на алвеоларния сърфактант (АС) да понижава повърхност-
ното напрежение на въздушно-водната фазова граница, стабилизирайки ал-веолите и предпазвайки ги от колапс при издишване, е от основно жизнено значение. АС се състои главно от фосфолипиди (90%) и белтъци (10%), като 4 от протеините са специфични за АС – SP-A, SP-B, SP-C и SP-D. Редица авто-ри моделират функциите на АС на фазовата граница, като изследват свойст-вата на индивидуални фосфолипиди (ФЛ), главно на най-застъпените ФЛ на АС – дипалмитоилфосфатидилхолин (ДПФХ) и дипалмотоилфосфатидилгли-церол (ДПФГ) [1]. В други изследвания се използват смеси от ДПФХ и ДПФГ с пречистени или рекомбинантни специфични сърфактантни белтъци [2], но в тези случаи не се взема под внимание ролята на минорните фосфолипидни компоненти на АС, а в допълнение се използват значително по-високи от ус-тановените in vivo белтъчни концентрации. Етаноламините са минорна фрак-ция в състава на АС – те съставят около 3,5 % от общото количество на сър-фактантните фосфолипиди [3]. Способността им обаче да формират както ламеларни, така и неламеларни структури, ги правят привлекателен обект за целите на настоящото изследване.
Вече е общоприето, че алвеоларният сърфактант формира монослойни, бислойни и мултислойни структури на граничната алвеоларна повърхност [4, 5]. За да изпълнява ефективно функциите си на въздушно-водната граница в белия дроб АС трябва да се трансформира бързо от хипофазата в повърх-ностно-активен филм на повърхността. Главният фосфолипиден компонент на АС, ДПФХ, се адсорбира изключително бавно и той сам не може да изпълнява ролята на ефективен сърфактант. Присъствието на други ФЛ, главно ненаси-тени, подобрява адсорбцията на ДПФХ, но тя остава незадоволителна и е да-леч под установената за естествения сърфактант. Вече е доказана ключовата рола в това отношение на специфичните хидрофобни сърфактантни белтъци SP-B и SP-C, които улесняват и ускоряват разтичането на фосфолипидите на алвеоларната повърхност [6, 7].
Във всички изследвания и публикации, засягащи повърхностното поведе-ние и свойствата на най-хидрофобния природен белтък SP-C в липидно обк-ръжение, е доказана важната му роля. Интерес за нас представляваше да се изолира от свински бял дроб и за първи път да се изследват взаимодействия-та между сърфактантния протеин SP-C и адсорбционни ФЛ монослоеве от различни течно-кристални фазови състояния на DPoPE – ламеларна и обрат-на хексагонална фази. Съществуват литературни данни, че при взаимодейст-вие с ФЛ в гелно състояние молекулата на SP-C се оказва по-къса от дебели-ната на липидния бислой и може да предизвика фазово разделяне на пептиди и липиди [8]. В бислоеве, формирани от гелни фази на ДПФХ/ДПФГ, SP-C претърпява агрегация, но в течно-кристални бислоеве белтъка е в мономерно състояние [9]. Определено може да се каже, че SP-C предпочита да взаимо-действа преимуществено с ФЛ в течно-кристално фазово състояние, и то с ФЛ с палмитоилови мастно-кисели вериги, но при взаимодействие със същите ФЛ в гелна фаза, той се изключва от бислоевете [10, 11]. Този ефект на изключ-ване може задоволително да се обясни със способността на агрегираните SP-C пептиди да разрушават липидното пакетиране в гелната фаза, докато теч-но-кристалните мембранни бислоеве са идеално пригодени са малките хидрофобни молекули на SP-C [12]. Установено е, че в ДПФХ-монослой SP-C

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 975
е силно наклонен спрямо интерфазата и по някакакъв начин предопределя виско конформационната подреденост на липидните въглеводорородни опашки. Това предполага, че в монослой SP-C може би изпълнява ролята на хидрофобен “лост”, който закотвя и задържа около себе си максимален брой фосфолипидни молекули [13].
ЦЕЛ Целта на представената работа е да се изследва влиянието на специ-
фичния сърфактантен протеин SP-C върху повърхностното поведение на адсорбционни липидни монослоеве, образувани от различни течно-кристални фази на DPoPE (ламеларна Lα и обратна хексагоналнана HII) на въздушно-водната фазова граница.
МАТЕРИАЛИ И МЕТОДИ ПОЛУЧАВАНЕ НА ЛАМЕЛАРНИ И НЕЛАМЕЛАРНИ ФАЗИ НА DРoPE: За
целта на настоящето изследване бе използван ненаситеният фосфолипид 1,2-Dipalmitoleoyl-sn-glycero-3-phosphoethanolamine (DPoPE) – продукт на Avanti Polar Lipids (Alabaster, USA), който сравнително лесно може да форми-ра различни течно-кристални фази при съответни температурни и електро-литни условия. Липидни дисперсии в 0.14 М NaCl (Merck) са приготвени при стайна температура чрез механично разбъркване. Както показват резултатите от диференциалната сканираща калориметрия (DSC) и рентгено-структурният анализ, ламеларната течно-кристална фаза на DPoPE се наблюдава в доста широк температурен интервал – от –35°С до +42°С. Водни дисперсии на об-ратната хексагонална HII фаза са приготвени при нагряване от стайна темпе-ратура до над 39°С (където се осъществява кооперативният фазов преход от ламеларна към хексагонална фаза). При охлаждане HII показва хистерзис и се запазва до 29°С, така че при проведените експериментите при 37°С диспер-сията е в обратна хексагонална фаза.
МЕТОД НА LANGMUIR ЗА ФОРМИРАНЕ И ИЗСЛЕДВАНЕ НА МОНОС-ЛОЕВЕ: Монослойният модел на Langmuir в комбинация с метода на Wilhelmy за измерване на повърхностното напрежение на течности, е широко използван в мембранологията. Везната на Langmuir-Wilhelmy се състои от ва-на, в която се формира монослоят от изследваното вещество върху течна подложка и чиято повърхност може да се променя с помощта на подвижна ба-риера (от 100% до 20% от първоначалната площ). По този метод могат да се образуват както нанесени, така и асдорбционни монослоеве. Нанесените мо-нослоеве от SP-C и DPoPE се получават чрез накапване по подходящ начин на точно определени количества върху чистата течна подложка, в резултат на което всички молекули се стремят да се разполагат термодинамично най-изгодно на въздушно-водната повърхност, като се ориентират в неразтворим мономолекулен слой. Адсорбционните монослоеве са получени чрез адсорб-ция на молекулите на DPoPE от обема на разтвора към граничната повърх-ност, където е термодинамично най-изгодно за тях да бъдат разположени. За получаване на адсорбционни монослоеве подходящи количества от липидна-та дисперсия (в различните течно-кристални фази) се инжектират през специ-ален отвор на дъното на ваната и се следи кинетиката на адсорбцията и разг-раждането на структурите на въздушно-водната повърхност, както и ефекта

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 976
от взаимодействието им на повърхността с предварително нанесения монос-лой от SP-C.
ИЗОЛИРАНЕ НА SP-C: Белодробен повърхностно активен материал се получава чрез 3-кратна промивка на свински бели дробове с физиологичен разтвор на стайна температура. Полученият лаваж се центрофугира некол-кократно по протокол, предложен от Ng и сътрудници [14]. Крайният етап на пречистване се осъществява чрез градиентно центрофугиране в захарозен градиент (0.68М и 0.33М захароза, 78 000 g, 60 min, 4°C) и получената фрак-ция се подлага на диизопропилов етер/n-бутанолова екстракция за отделяне на хидрофобните белъци SP-B и SP-C във водната фаза [15]. Следва изпаря-ване под азот на тази водна фаза и полученият сух материал се екстрахира с диетилов етер/етанол. Получената утайка след центрофугиране на 18 000g, за 30 min и температура 4°C, отново се екстрахира със система от органични разтворители: хлороформ/метанол/HCl (1:1:0.5М). Долният органичен (хлоро-формен) слой съдържа SP-C. След отделянето му този разтвор се съхранява на –20°C, и преди провеждане на експериментите се темперира и задължи-телно се изчаква (при формиране на нанесени монослоеве) поне 20 минути, за да се изпари напълно органичният разтворител.
РЕЗУЛТАТИ И ОБСЪЖДАНЕ Формирането на ламеларната течно-кристална фаза на DРoPE става
чрез механично диспергиране на липида в подбрания разтвор при стайна температура. Както показаха измерванията от DSC (Фигура 1) и данните от рентгено-структурния анализ (Фигура 2), водната дисперсия от DРoPE има фазов преход Lα → HII при температура 42 °С. При охлаждане се наблюдава хистерезис от около 10-15 °С (Фигура 1 и 2). DРoPE дисперсиите в 0.5 М NaCl имат фазов преход Lα → HII при температура 39°С (Фигура 1), т.е. под тази температура DРoPE съществува в ламеларна течно-кристална фаза Lα, а над 39 °С липида формира обратна хексагонална фаза HII. В резултат на гореспо-менатия хистерезис след първото нагряване на DРoPE-дисперсията при тем-ператури над фазовия преход, т.е. над 39°С, и след последващо охлаждане до 37°С, където са проведени голяма част от експериментите, липида форми-ра спонтанно обратна хексагонална фаза. При нагряване обаче на първона-чалната дисперсия от стайна температура до 37 °С, DPoPE е в ламерано теч-но-кристално фазово състояние.
За целите на нашето изследване е изолиран SP-С от свински белодробен лаваж с различни системи органични разтворители, при което протеина се ко-екстрахира с фосфолипидите (Виж Материали и методи). Ниското съдържа-ние на специфичните хидрофобни белтъци (под 1 % от хлороформ-разтво-римата фракция на АС [16, 17] прави изключително трудно получаването им в чист вид в достатъчни количества. Някои методи за изолирането на SP-C включват разделянето му от съпътстващите го липиди и SP-В чрез молекул-но-ситова хроматография, с използване на Sephadex LH-20 и/или LH-60 [2], което обаче може да доведе до ковалентни модификации на молекулата му. Поради тази причина ние предпочетохме прилагането на диизопропилов етер/n-бутанолова екстракция за отделяне на двата хидрофобни белъка SP-B и SP-C във водната фаза [15]. Нискомолекулните хидрофобни белтъци SP-B и

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 977
SP-C са детектирани чрез денатурираща SDS-PAAGE и визуализирани със сребърно оцветяване. Резултатите от електрофоретичния анализ са предста-вени на Фигура 3. В Панел А, старт 2, е идентифицирано присъствието и на двата хидрофобни белтъка SP-B и SP-C преди крайното им разделяне. На Фигура 3, Панел Б, стартове 1,3 и 4 показват присъствие само на SP-C в крайните фракции. Тези фракции се явяват доста удобен модел за изследва-не чрез сравнителен анализ на ролята на хидрофобните сърфактантни бел-тъци за поведението на различно-фазови ФЛ компоненти на АС в монослой.
На Фигура 4 са показани резултатите от взаимодействието между пред-варително формиран нанесен монослой от SP-C (с концентрация, отговаряща на 0,4 молни % спрямо липидната концентрация, съгласно литературни данни [18, 19, 20]). Монослоят от SP-C, формиран върху чист физиологичен разтвор при температура 37 °С е с площ 706 mm2, и показва понижаване на повърх-ностното напрежение до 53.4 mN/m, спрямо чистата субфаза (Таблица 1). До достигане на тази равновесна стойност се изчаква 20 минути, за да се изпари хлороформа, в който е разтворен и съхраняван белтъка. След отчитане на равновесната стойност γeq на дъното на ваната се инжектира съответната течно-кристална липидна фаза (с крайна обемна концентрация в субфазата 5 µg/ml) и се проследява промяната на повърхностното напрежение с времето до достигане на крайна равновесна стойност на формирания смесен монос-лой. Трябва да отбележим, че получените равновесни стойности на γ за адсорбционни монослоеве, формирани само от Lα и HII при същите експери-ментални условия са съответно 34.3mN/m и 44.0mN/m (Таблица 1).
Данните от експеримента показват, че след инжектирането на DPoPE в различните фази, вследствие взаимодействието им на повърхността със SP-C, се достига до нови, значително по-ниски стойности на повърхностното нап-режение, като трябва да подчертаем, че при смесените SP-C+Lα монослоеве крайната отчетена равновесна стойност на повърхностното напрежение е 28.1mN/m, а за монослоевете, формирани от SP-C+HII тази стойност е 29.6mN/m. Т.е. ефектът на взаимодействие на DPoPE със специфичния сър-фактантен белтък SP-C е значителен и за двете фази, като се отчита малка разлика между тях.
На следващите две фигури са показани резултатите от три последова-телни цикъла на компресия/декомпресия на адсорбционни липидни моносло-еве (формирани съответно от Lα и от HII при крайна обемна концентрация 5µg/ml), както и на смесени монослоеве (липида е инжектиран в субфазата под нанесен монослой на SP-C). Проследена е промяната на γmax (Фигура 5) и γmin (Фигура 6) от номера на цикъла. Данните са представени и в Таблица 1 и от тях можем да констатираме, че докато за ламеларния монослой промяната на γmax от първия до третия цикъл на компресията е съответно от 34.3 mN/m до 36.8 mN/m (леко покачване), то при формиране на смесен с SP-C монослой тези стойности са съответно 28.1mN/m и 23.1mN/m (силно понижаване) (Фигу-ра 5). Освен че се понижава значително γmax за смесения Lα+SP-C, се забе-лязва и тенденция към достигане на все по-ниски стойности на γmax във всеки следващ цикъл за този смесен монослой.
При смесените SP-C с обратната хексагонална фаза монослоеве се за-белязва друга тенденция. При измерване на γmax на монослоеве, формирани от HII се регистрира значително повишаване на γmax от първи до трети цикъл

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 978
съответно от 44.0 mN/m до 50.9 mN/m (Фигура 5). При компресията на смесе-ните (HII+SP-C) монослоеве стойностите на γmax се покачват съответно от 29.6 mN/m в първия цикъл до 36.2mN/m в третия цикъл, за разлика от смесените ламеларни+SP-C монослоеве, при които тенденцията е стойността на γmax да намалява във всеки следващ цикъл.
Промяната на γmin от номера на цикъла за изследваните монослоеве е показана на Фигура 6. Най-голямо понижение на тази стойност се наблюдава при смесените с SP-C ламеларни монослоеве. Докато чистите адсорбционни ламеларни монослоеве показват незначителна промяна при динамични усло-вия на γmin от първия до третия цикъл (от 27.1 mN/m до 26.5 mN/m), то при смесване с SP-C тези стойности намаляват чувствително: от 25.6 mN/m в пър-вия цикъл до 17.5 mN/m в третия цикъл на компресия.
Обратно на чистата Lα фаза, при HII фазата γmin намалява при циклиране. Също така, обратно на поведението на смесените Lα+SP-C монослоеве, къде-то γmin силно намалява, при смесените HII+SP-C монослоеве γmin е почти пос-тоянно с увеличаване на броя на циклите.
ЗАКЛЮЧЕНИЕ Сравнителният анализ на резултатите от повърхностното взаимодейст-
вие между SP-C и DPoPE в монослой показват, че ефектът на много силно понижение на γ се достига много по-бързо при взаимодействие с ламеларната фаза, докато при обратната хексагонална фаза се наблюдава постепенно достигане (до 4-тата минута) до равновесната стойност на смесения монослой (Фигура 4). По отношение на динамичните характеристики на смесените мо-нослоеве, в зависимост от липидното фазово състояние, тенденциите на про-мяна на стойностите на γmax и γmin се различават в значителна степен, като ре-зултат от различната степен на взаимодействие на ламеларната и хексаго-налната фази с SP-C (Фигури 5 и 6).
Представените резултати са първи опит за сравнителен анализ на ре-гистрираните равновесни стойности на повърхностното напрежение при адсорбция на Lα и HII липидни фази към нанесен монослой на SP-C, както и на динамичните характеристики между смесени ламеларни+SP-C от една страна и обратно-хексагонални+SP-C от друга, монослоеве.
Данните от проведените изследвания потвърждават и уникалната спо-собност на специфичния сърфактантен белтък SP-C силно да улеснява и ус-корява адсорбцията на фосфолипиди на гранични повърхности.
Благодарности Настоящата експериментална работа е осъществена с финансовата под-
крепа на договор 29/2006 фонд Научни изследвания при СУ “Св. Кл. Охридс-ки”.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 979
ЛИТЕРАТУРА [1] Notter, R., Holcomb, S., Mavis, R., Chem. Phys. Lipids, 27:305-311, 1980 [2] Taneva S.G., Keough K.M.W., Biophys. J., 66:1149–1157, 1994 [3] Yu S.H., Smith N., Hardihg P.G.R., Possmayer F., Lipids, 18:522-527, 1983 [4] Schurch S., Green F.H.Y., Bachofen H., BBA, 1408, 180-202, 1998 [5] Scarpelli E., Mautone A.J., De Fouw D.O., Clutario B.C., Anat. Rec., 246, 245-
270, 1996 [6] Shou-Hwa Yu, Possmayer F., BBA, 1046: 233-241, 1990 [7] Ross M., Krol S., Janshoff A., Galla H.J., Eur. Biophys. J., 31(1):52-61, 2002 [8] Johansson, J., Curstedt, T., Eur. J., Biochem. 244:675-693, 1997 [9] Horowitz, A.D., Baatz, J.E. and Whittset, J., Biochemistry, 32:9513-9523, 1993 [10] Perez-Gil J., Casals C., Marsh D., Biochemistry, 34:3964-3971, 1995 [11] Horowitz A.D., Chem. Phys. Lipids, 76:27-39, 1995 [12] Johansson J., Szyperski, T., Wuthrich K., FEBS Lett., 362:261-265, 1995 [13] Gericke A., Flach C.R., Mendelsohn R., Biophys. J., 73:492-499, 1997 [14] Ng V., Herndon V., Mendelson C, Snyder J., BBA 754, 218-226, 1983 [15] Cham B., Knowles B., J. Lipid. Res., 17, 176-181, 1976 [16] Curstedt T., Jornvall H., Robertson B., Bergman T., Berggren P., Eur. J.
Biochem, 168:255-262, 1987 [17] Hall S.B., Venkitaraman A.R.,Whitsett J.A., Holm B.A., Notter R.H., Am. Rev.
Respir. Dis., 145:24-30, 1992 [18] Amrein M., Nahmen A von, Sieber M., Eur. Biophys. J., 26:349-357, 1997 [19] Nahmen A. von, Post A., Galla H.J., Sieber M, Eur. Biophys. J., 26:359-369, 1997 [20] Krol S., Janshoff A., Ross M., Galla H.J., Phys. Chem. Chem Phys. 2:4586-
4593, 2000 Н.с. I ст. Албена Георгиева Йорданова Институт по Биофизика, БАН 1113 e-mail: [email protected], 02 8167345 Ас. Емилия Кирчева Стоименова Катедра по биохимия, Биологически Факултет СУ “Св. Климент Охридски” 1164 Спец. Христина Тонкова Петкова Институт по физикохимия, БАН 1113 Н.с. I ст. Румен Кънчев Тодоров Институт по физикохимия, БАН 1113 Ас. д-р Йордан Атанасов Думанов Катедра по биохимия, Биологически Факултет СУ “Св. Климент Охридски” 1164 Проф. дбн Здравко Иванов Лалчев Катедра по биохимия, Биологически Факултет СУ “Св. Климент Охридски” 1164
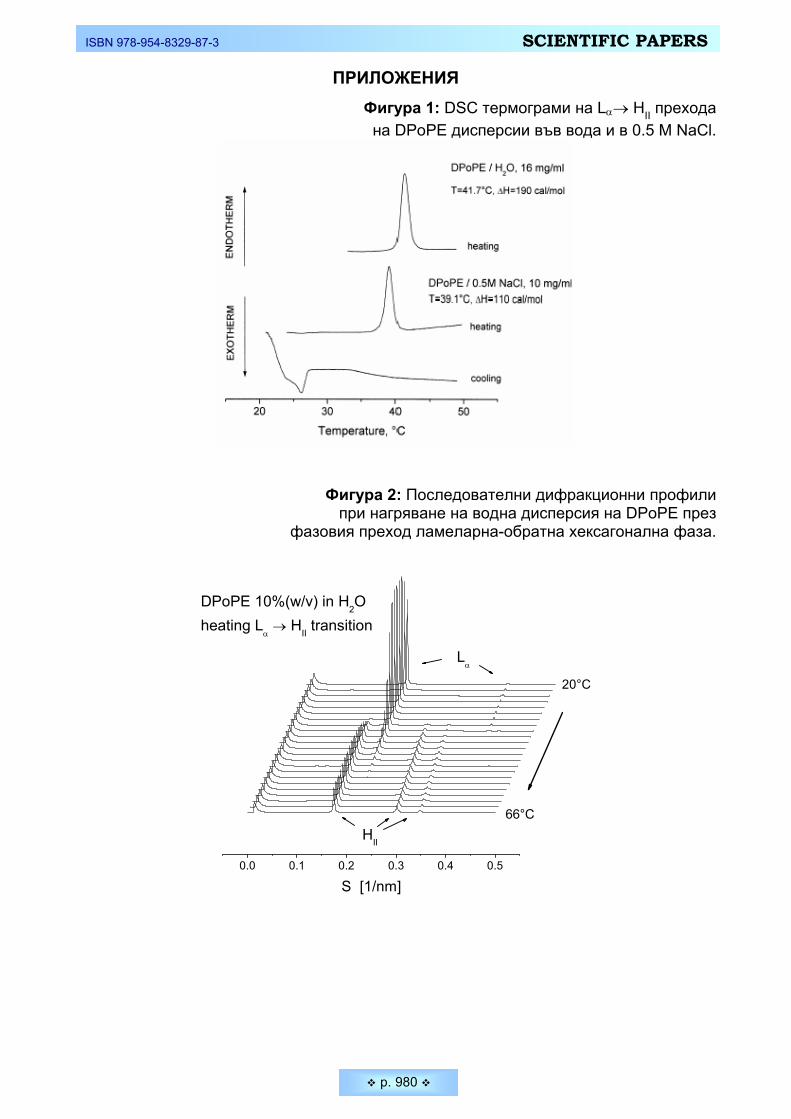
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 980
ПРИЛОЖЕНИЯ Фигура 1: DSC термограми на Lα→ HII прехода на DPoPE дисперсии във вода и в 0.5 М NaCl.
Фигура 2: Последователни дифракционни профили при нагряване на водна дисперсия на DPoPE през
фазовия преход ламеларна-обратна хексагонална фаза.
0.0 0.1 0.2 0.3 0.4 0.5
DPoPE 10%(w/v) in H2Oheating L
α → HII transition
66°C
20°C
Lα
HII
S [1/nm]
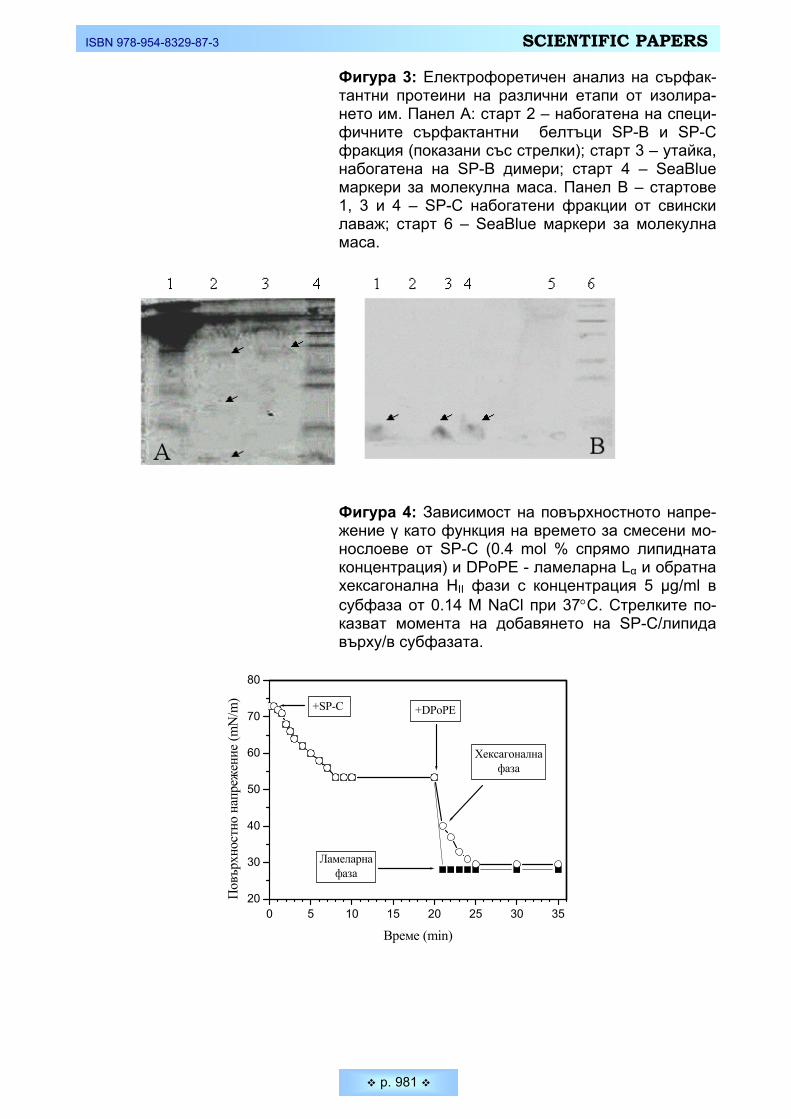
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 981
Фигура 3: Електрофоретичен анализ на сърфак-тантни протеини на различни етапи от изолира-нето им. Панел A: старт 2 – набогатена на специ-фичните сърфактантни белтъци SP-B и SP-C фракция (показани със стрелки); старт 3 – утайка, набогатена на SP-B димери; старт 4 – SeaBlue маркери за молекулна маса. Панел B – стартове 1, 3 и 4 – SP-C набогатени фракции от свински лаваж; старт 6 – SeaBlue маркери за молекулна маса.
Фигура 4: Зависимост на повърхностното напре-жение γ като функция на времето за смесени мо-нослоеве от SP-C (0.4 mol % спрямо липидната концентрация) и DPoPE - ламеларна Lα и обратна хексагонална HII фази с концентрация 5 µg/ml в субфаза от 0.14 M NaCl при 37°C. Стрелките по-казват момента на добавянето на SP-C/липида върху/в субфазата.
0 5 10 15 20 25 30 3520
30
40
50
60
70
80
Ламеларнафаза
Хексагоналнафаза
+DPoPE+SP-C
Повърхностно напрежение
(mN
/m)
Време (min)
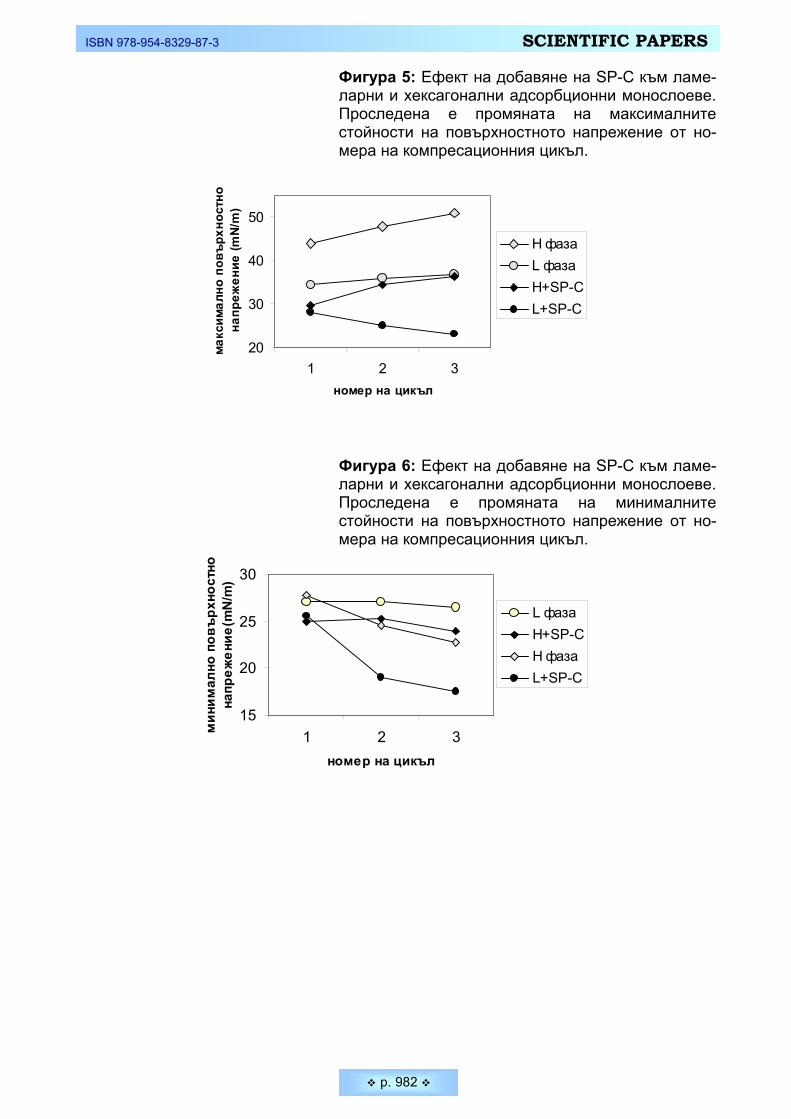
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 982
Фигура 5: Ефект на добавяне на SP-C към ламе-ларни и хексагонални адсорбционни монослоеве. Проследена е промяната на максималните стойности на повърхностното напрежение от но-мера на компресационния цикъл.
20
30
40
50
1 2 3номер на цикъл
максима
лно по
върх
ностно
на
преж
ение
(mN
/m)
H фазаL фазаH+SP-CL+SP-C
Фигура 6: Ефект на добавяне на SP-C към ламе-ларни и хексагонални адсорбционни монослоеве. Проследена е промяната на минималните стойности на повърхностното напрежение от но-мера на компресационния цикъл.
15
20
25
30
1 2 3номер на цикъл
мин
имал
но пов
ърх
ностно
напр
ежен
ие(m
N/m
)
L фазаH+SP-CH фазаL+SP-C
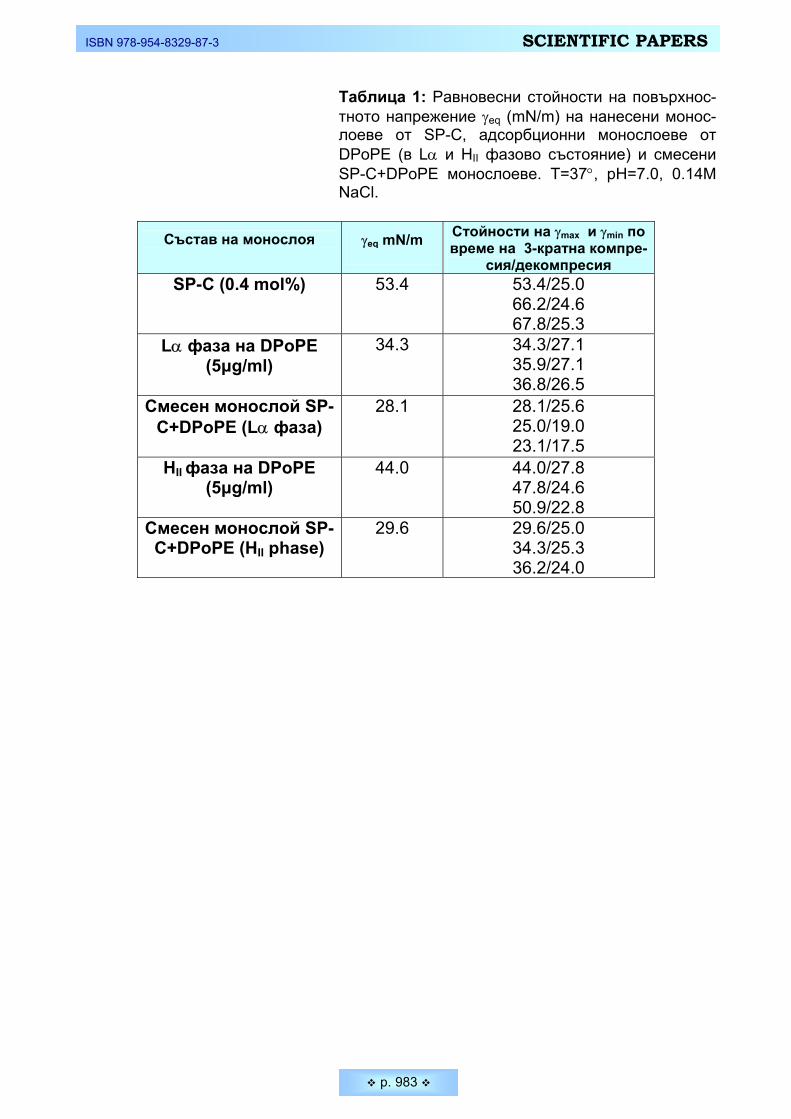
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 983
Таблица 1: Равновесни стойности на повърхнос-тното напрежение γeq (mN/m) на нанесени монос-лоеве от SP-C, адсорбционни монослоеве от DPoPE (в Lα и HII фазово състояние) и смесени SP-C+DPoPE монослоеве. Т=37°, pH=7.0, 0.14М NaCl.
Състав на монослоя
γeq mN/m
Стойности на γmax и γmin по време на 3-кратна компре-
сия/декомпресия SP-C (0.4 mol%)
53.4
53.4/25.0 66.2/24.6 67.8/25.3
Lα фаза на DPoPE (5µg/ml)
34.3
34.3/27.1 35.9/27.1 36.8/26.5
Смесен монослой SP-C+DPoPE (Lα фаза)
28.1
28.1/25.6 25.0/19.0 23.1/17.5
HII фаза на DPoPE (5µg/ml)
44.0
44.0/27.8 47.8/24.6 50.9/22.8
Смесен монослой SP-C+DPoPE (HII phase)
29.6
29.6/25.0 34.3/25.3 36.2/24.0

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 984
SCIENTIFIC PAPERS
РАЗПРЕДЕЛЕНИЕ НА BRAIN-DERIVED NEUROTROPHIN FACTOR ВЪВ ВЪТРЕШНИТЕ СЛОЕВЕ НА РАЗВИВАЩА СЕ
ПОСТНАТАЛНА РЕТИНА НА БЯЛ ПЛЪХ
Ст. Йочкова, Д. Вучидолова, Р. Давидова Катедра по Анатомия, Хистология и Ембриология,
Медицински Университет, Плевен, България
EXPRESSION OF BRAIN-DERIVED NEUROTROPHIC FACTOR IN THE INNER LAYERS OF DEVELOPING
POSTNATAL RAT RETINA
St. Jochkova, D. Vouchidolova, R. Davidova Department of Anatomy, Histology and Embryology,
Medical University, Pleven, Bulgaria ABSTRACT Brain-Derived Neurotrophic Factor (BDNF) promotes differentiation, maturation, syn-
aptic plasticity and survival of neurons in central and peripheral central nervous system. The expression of Brain-Derived Neurotrophic Factor (BDNF) was studied in inner
layers of developing postnatal rat retina. We investigated BDNF in retinas of rats on postnatal day 1, 3, 5, 7, 9, 12, 18, 30.
Changes in BDNF expression and localization in developing retina were assessed by im-munohistochemistry.
The immunohistochemical analyses revealed that BDNF expression occurred in reti-nal ganglion cells on P1 and increase between P12 and P30. Amacrine cells be-gin immunopozitive on P1, bipolar cells on P5. Horizontal cells are well-stained on P9. BDNF immunoreaktivity in processes of Müller cells was observed in inner nuclear layer (INL) on P3. In plexiform layers the BDNF express early on P1 in inner and on P5 in outer plexiform layer.
The distribution of expression of BDNF in postnatal retina suggest that it plays role in differentiation of retinal cells of inner layers and their synaptogenesis.
Key words: BDNF, neurotrophins, rat retina, postnatal retina development, retinal inner layers Невротрофините, включващи BDNF, NGF, NT3 и NT4 и техните рецепто-
ри TrkB, TrkA и TrkC подпомагат диференциацията, матурацията и оцелява-нето на невроните в периферната и централната нервни системи .
Ние изследваме ретиналната BDNF експресия и локализация чрез иму-ноцитохимия, за да проследим промените в експресията и локализацията на BDNF в диференциращата се ретина на бял плъх в първите тридесет постна-тални дни.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 985
МАТЕРИАЛ И МЕТОДИ Плъхове от порода Wistar бяха убивани под етерна наркоза на 1, 3, 5, 7,
9, 12, 18, 30 постнатални дни (Р1, Р3, Р5, Р7, Р9, Р12, Р18 и Р30). Енуклеира-ните очи бяха фиксирани в Carnoy разтвор за 1-2 часа и включени в течен па-рафин. Депарафинираните срезите с дебелина 5 µm бяха депарафинизирани и инкубирани за 20 часа с първично anti-rabbit BDNF с разреждане 1:50 (Santa Cruz Biotechnology, California). За визуализиране на антитялото беше използ-вана ABC Staining System (Santa Cruz Biotechnology, California ) и оцветяване-то беше извършено с пероксидазна 3-3′ диаминобензидинова (DAB) реакция. Готовите препарати бяха наблюдавани с микроскоп, тип Leica и заснети с ди-гитален фотоапарат от микроскопа.
РЕЗУЛТАТИ И ОБСЪЖДАНЕ На Р1 BDNF имунопозитивност се наблюдаваше в ганглийните клетки
(GC), слоя на нервните влакна (NF) и вътрешния влакнест слой (IPL). Слабо се позитивираха изместени амакринни клетки (AC), разположени в близост до IPL (фиг. 1). На трети постнатален ден се забелязва слаба имунопозитивна реакция на израстъците на мюлеровите клетки (MC) във вътрешния подслой на невробластните (вентрикуларни) клетки, диференциращи се във фоторе-цепторни и клетъчните популации на INL (фиг. 2). BDNF експресия във външ-ния влакнест слой (OPL) започва с началото на формирането на този слой на Р5 и се засилва в следващите две постнатални седмици. Клетъчните попула-ции, разположени във вътрешния нуклеарен слой (INL) са слабо имунопози-тивни на Р5. В средата на INL добре позитивирани се наблюдават биполарни клетки (ВС) на Р7 (фиг. 3). На Р9 добре се очертават по големите тела на хоризонталните клетки (HC), разположени в най- външната част на INL и из-растъците на мюлеровите клетки във слоя на пръчиците и колбичките (ONL) (фиг. 4). Експресията на BDNF в клетъчните популации от вътрешните слоеве на ретината на белия плъх и двата влакнести слоя достига максимум между Р12 и Р18 (фиг. 5 и фиг. 6) и се запазва до първия поснатален месец. Стената на кръвоносните съдове в NF и OPL проявява имунопозитивност в първите поснатални дни и също достига максимум на Р18.
Невротрофичния фактор BDNF упражнява различни ефекти в ембрио-налния период, след раждането и в зрелия период, чрез свързването си с ви-сокоафинитетния тирозин киназен рецептор B (TrkB).
В нашите проучвания беше проследена експресията на BDNF във вът-решните слоеве на ретината на плъх в постнаталния период. Диференциаци-ята на ганглийните клетки започва преди раждането на Е12 и продължава в първите две постембрионални седмици. От клетките на вентрикуларния слой се развиват амакринните, биполарните и хоризонталните неврони. Морфоге-незата на хоризонталните клетки от първи тип започва на Р2-3, а на тези от втори тип на Р4-7. Биполарните се диференцират между Р4-Р8. Между Р3-8 амакринните клетки, разположени в ганглийния слой мигрират и заемат крайното си положение в най- вътрешната част на вътрешния нуклеарен слой [4]. Имунопозитивната реакция за BDNF изразена в етапите на диференциа-цията и заемането на дифенитивните позиции на определени ретинални кле-тъчни популации във вътрешните слоеве на ретината през първите две седмици след раждането, подсказват участието и важността на BDNF в тези процеси. В ембрионалния период BDNF и TrkB играят роля за оцеляване на

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 986
ганглийните клетки ин витро и ин виво [3, 6, 8, 11, 14]. Във възрастни животни BDNF има отношение към контролиране на дегенерацията на ганглийните клетки [7, 9, 10]. BDNF повишава броя на ретиналните допаминергични синап-си в плъховата ретина Плътността на ретиналната допаминергична инерва-ция, развитието на амакринните клетки от втори тип и някои субпополации на ганглийните клетки се контролира от екзогенно приложен BDNF в гризачи (5). Нашите резултати подчертават ролята на BDNF предимно в процеса на ди-ференциация на определени ретиналните типове в постнаталния период.
Синаптичната пластичност, е процес на функционална модификация на синапсите в развиващата се и зрялата централна нервна система, често придружена от анатомични промени в тези места. Невротрофините модулират функционалния статус на невроните от повлияващата се синаптогенеза, ак-тивно зависимата синаптична пластичност и синтеза на невротрансмитерите и невромодулаторите. Процесът на синаптичната пластичност, който присъства през ембрионалния период и продължава през живота на организма е актив-но-зависим [1, 2, 12, 13]. BDNF и TrkB играят роля в постнаталната ретино- тектална пластичност в гълъби след излюпването им, докато в пилешка рети-на те се наблюдават в по раннните периоди. Ние наблюдаваме ранна имуно-позитивност на слоя на нервните влакна и вътрешния влакнест слой на Р1 както и на външния влакнест слой на Р5, корелираща с времето на развитие-то и формирането им. Тези резултати подчертават ролята на BDNF в процеса на синаптогенезата и пластичността на синапсите.
Мюлеровите клетки на бозайниците съдържат ендогенен BDNF и са път за предаването на невротрофина до ганглийните клетки. Наличието на BDNF в Мюлеровите клетки подсказва тяхната поддържаща роля, като източник на BDNF в ретиналните неврони [3]. Експресията на BDNF в Мюлеровите клетки и техните израстъци започва и се засилва в дните на диференциацията им и моментите на формиране и заемане на дефинитивните места на фоторецеп-торните и ганглийните клетки.
В заключение може да кажем, че BDNF участва предимно в процесите на диференциацията и синаптогенезата на клетките от вътрешните слоеве на ретината на плъх в постнаталния период.
ЛИТЕРАТУРА
[1] Barbara Lom and Susana Coham-Cory, Brain-derived Neuthophic Factor Differentially Regulates Retinal Ganglion Cells Dendritic and Axonal Arborization in Vivo, Jornal of Neuroscience, 1999, November 15, 19(22), 9928-9938
[2] Barbara Lom, J. Cogan, A. L. Sanchez, Thuy Vu and Susana Coham-Cory, Local and Targed Brain-derived Neuthophic Factor exert opposing effect on the Dendritic Arborization of Retinal Ganglion Cells in Vivo, , Jornal of Neuroscience, 2002, September 1, 22(17), 7639-7649
[3] Baerbel Rohrer, Matthew M. LaValt, Kevin R. Jounes and Louis F. Reichardt, Neutrophin Reseptor TrkB Activation Is Not Required for the Postnatal Survaval of Retinal Ganglion Cells in Vivo, Experimental Neurology, 2001, 172, 81-91

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 987
[4] Вучидолова Д., С. Йочкова, К. Койчев, Постнаталното развитие на кле-тъчните типове във вътрешните слоеве на ретината на плъх, Българска медицина, 1996, том IV, брой 5-6, 63-66
[5] Cellerino A, Pinzon-Duarte G, Carroll P, Kohler K, Brain-Derived Neurotrofic Factor Modulates the Development of the Dopaminergic Network in the Rodent Retina The J. of Neuroscience 1998 May 1, 18, 3351-3362
[6] Frade J M et all, Control of early death in chick retina, Development, 1997, 124, 3313-3320
[7] Hua Gaoq Xiaoxi Qiao, Louis B. Cantor, Darirell WuDunn, Up-Regulation of Brain-derived Neuthophic Factor Expression by Brimordine in Rat Retinal Ganglion Cells, Arch ophalmology, vol 120, June 2002, 797803
[8] Michael Neal, Joanna Cunningham, Isobel Lever, Sophie Pezet, and Marszia Malcangio, Mechanism by which Brain-derived Neuthophic Factor Increase Release from the Rabbit Retina, Investigative Ophtalmology and Visual Science, 2003, 44, 791-798
[9] Monica Garsia, Valerie Foster, David Hick and Elena Vecino, In Vivo Exspression of Neurotrophins and Neurotrophin Reseptors Is Conversed in Adult Porcine Retina In Vitro, Investigative Ophtalmology and Visual Science, 2003, 44, 4532-4541
[10] Pollock Graeme S., Regine R., Kristina A. B. et all, TrkB Receptor Signaling Regulates Developing Death Dynamics, Not Final Number, of Retinal Ganglion Cells, The J. of Neuroscience2003 Noember 5, 10137-10145
[11] Roher B. et all (2001) Neurotrophin Receptor TrkB Activation Is Not Required for the Postnatal Survival of Retinal Ganglion Cells in vivo Experimental Neurology 2001, 172, 81-91
[12] Rohrer B., J. I. Korenbrot, Matew M. LaVail, Louis F. Reichardt, Baoji Xu, Role of Neutrophin Receptor TrkB in the Maturation of Rod Photoreceptors and Establishment of Synaptic Transmission to the Inner Retina, The J. of Neuroscience, 1999, October15, 19(20), 8919-8930
[13] Susana Coham-Cory and Barbara Lom, Neurotrophic regulation of retinal ganglion cells synaptic connectivity& from axons and dendrites to synapses, Int. J. Dev. Biol., 2004, 48, 947-956
[14] Theiss C, Onur Gunturkun, Distribution of BDNF, NT-3, trkB and trkC in the developing retino-testal system of the pigeon /Columba livia/ Anat Embryol,2001 204: 27-37. Д-р Стилянка Димитрова Йочкова-Василева, гл. асистент Д-р Дорка Михайлова Вучидолова, кмн Д-р Румяна Иванова Давидова, гл. Асистент катедра Анатомия, хистология и цитология МУ – Плевен, ул. “Св. Климент Охридски” № 1, 5800, Плевен Тел.: 064884145

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 988
ПРИЛОЖЕНИЯ
Фиг. 1. BDNFимунореактивност в постнаталната ретина на плъх на Р1

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 989
Фиг. 2. BDNFимунореактивност в постнаталната ретина на плъх на Р3

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 990
фиг. 3. BDNFимунореактивност в постнаталната ретина на плъх на Р7

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 991
фиг. 4. BDNFимунореактивност в постнаталната ретина на плъх на Р9

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 992
фиг. 5. BDNFимунореактивност в постнаталната ретина на плъх на Р12
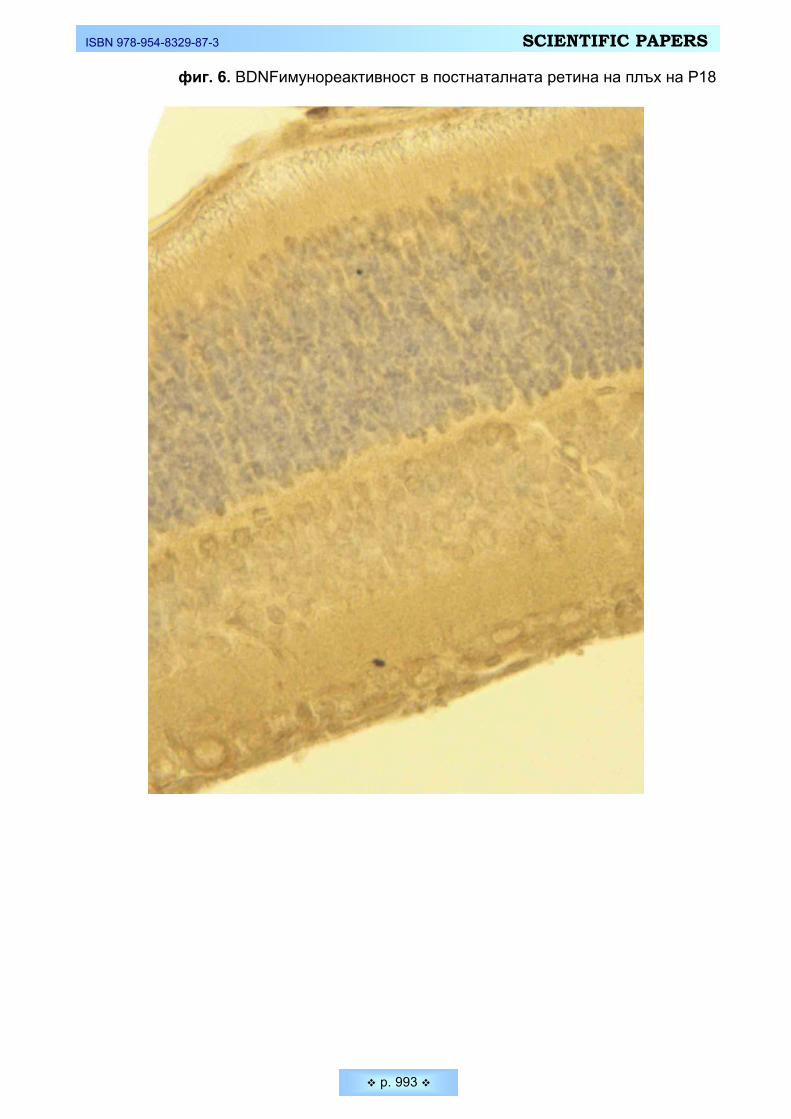
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 993
фиг. 6. BDNFимунореактивност в постнаталната ретина на плъх на Р18

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 994
SCIENTIFIC PAPERS
МОРФО-АНАТОМИЧНИ ОСОБЕНОСТИ НА СЛАДКАТА СТЕВИЯ (STEVIA REBAUDIANA BERTONI) В УСЛОВИЯТА НА БЪЛГАРИЯ
Койчо Коев, Иван Бакалски, Диана Калъчева
ПУ “Паисий Хилендарски”, гр. Пловдив, България
MORFO-ANATOMICAL CHARACTERISTICS OF SWEET STEVIA (STEVIA REBAUDIANA BERTONI)
IN BULGARIAN CONDITIONS
Kojcho Koev, Ivan Bakalski, Diana Kalacheva University of Plovdiv, Plovdiv, Bulgaria
ABSTRACT The investigation includes morphological and anatomical analysis of the species Ste-
via rebaudiana Bertoni, which is very perspective for the pharmacological and foodflavor industry.The experiment had been carried out with wild-nature /field/ growing plants origi-nating from Paraguay and the cultivated form from the botanical garden in Praga. The macro- and microscopic investigation was accomplished upon fully developed vegetative organs – roots, leafs and stems. The results show, that the structural characteristics of the sweet Stevia, growing in the Bulgarian environmental conditions, are similar to the pa-rameters of the wild nature growing plants. The observed specific features in leafs’ struc-ture determine the species as hydromorphic and suitable for cultivation predominantly in greenhouse conditions or nearby large water-filled areas, where the humidity would be enough to satisfy its needs.
Key words: sweet stevia, morphological analysis, anatomical analysis, leaf structure ВЪВЕДЕНИЕ Сладката стевия (Stevia rebaudiana Bertoni) се отнася към сем. Сложноц-
ветни (Asteraceae). Родът Stevia е много богат на видове – над 150. По-широко представени обаче са: Stevia eupatoria, Stevia salicifolia, Stevia ovata, Stevia serrata, Stevia plumerea и естествено обектът на настоящото изследва-не – Stevia rebaudiana.
Популяризирането на вида Сладка стевия е свързано с името на д-р Мо-исей Сантяго Бертони, преподавател в аграрния колеж в гр. Асунсион в Па-рагвай. През далечната 1887г. той се запознава с широко използваното от местните индианци–Гуарани, растение “кааихее”, като подсладител и арома-тизатор на техните традиционни напитки. След щателно проучване на вида д-р Бертони го отнася към род Stevia и през 1899 г. го публикува като нов за на-уката вид, който назовава – rebaudiana, на името на своя колега и приятел, парагвайския химик Ребауди.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 995
Разпространението на род Stevia е предимно в Южна и Централна Америка, но Stevia rebaudiana е ограничена единствено в Парагвай и Боли-вия. Най-големите находища са по долината на р. Мондей в Североизточен Парагвай, където обитава кисели, блатисти почви с ниско плодородие. В естествените си местообитания видът се развива в условия на субтропи-чен, влажен климат при среднодневни температури–24 °C и температурен диапазон 21–43 °C. Количеството на валежите достига до 1375 mm/m2, кое-то определя вида като изискващ висока почвена и въздушна влажност и оп-ределено леки, наносни почви.
Широкото разпространение на Stevia rebaudiana Bertoni в Северна Аме-рика, Азия и Европа е свързано с откритието на двата извънредно сладки гли-козида–ребаудиозид и стевиозид, превишаващи сладкия вкус на захарта от 200 до 400 пъти.Почти веднага след това, първо в Япония, а по-късно в Кана-да, Мексико и Китай, започват опити за култивация на вида.
Българския контакт със Stevia rebaudiana Bertoni води началото си от 80-те години на ХХ век, когато с генетичен материал от Япония в тогавашния Инс-титут по захарно цвекло в гр. Шумен се създава звено за изследване на вида. За съжаление разработките не се довеждат до благополучен край и Сладката стевия не бива въведена в култура. По същото време украински учени, изпол-звайки растителният материал от България създават огромна база за научно–приложна дейност към Института сахарной свеклы в гр. Киев. Задълбочените изследвания обаче са насочени предимно към химичния анализ на сладките гликозиди и размножителните методи без да се спира вниманието върху бо-таническите и екологическите параметри на Stevia. Тези изследвания естест-вено не създават добра основа за разработка на подходяща за условията на България технология за култивация и преработка на коментирания вид.
ЦЕЛ Проследяване на морфологичните и структурни белези на вегетативните
органи на Сладката стевия в условията на България. Така поставените цели бяха разпределени в следните задачи: 1. Морфологичен анализ на вегетативните органи на растението; 2. Анатомичен анализ на завършилите развитието си стъбло, лист и ко-
рен. МАТЕРИАЛИ И МЕТОДИ За целите на експеримента са използвани два типа генетичен материал: 1. Семена от Stevia с произход от ботаническата градина в гр. Прага, Р.
Чехия; 2. Растения от гр. Хамбург, Германия – отглеждани в полеви условия; Опитната постановка включва двегодишен период, обхващащ вегетаци-
онните сезони на 2005 и 2006 г. Целите на настоящата разработка определят следните варианти, а
именно: 1.Отглеждане на Stevia rebaudiana Bertoni с вместимост 0,5 l и1l, в оран-
жерийни условия; (Вариант 1), (Снимка № 1 и 2); 2.Отглеждане на растения от Stevia rebaudiana Bertoni в полеви условия
при директна сеитба; (Вариант 2), (Снимка № 4).

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 996
Етапите на изследването са продиктувани от една страна от поставените задачи, а от друга зависят от фенологичната характеристика на разглеждания вид. Комбинацията от тези два фактора определят и следната схема на ця-лостния експеримент:
1. Морфологични характеристики на надземната част на Stevia rebaudiana;
2. Анатомичен анализ на листата, стъблото и корена на опитното расте-ние;
3. Проследяване на прираста на надземната част в различни опитни ус-ловия;
Морфологичните и анатомичните анализи са извършени по класичес-ките методи на Metcalf and Cholck, 1952. Микроскопските изследвания са направени на микроскоп PZO при увеличение-×156,25 и ×625. Адаптирането на окоренените резници в почвена среда е осъществено в хранителна смеска включваща: 30 % четиригодишен оборски тор, 10 % перлит, 10 % промит пясък, 10 % угнили дървесни стърготини (от иглолистна дървесина) и 40 % градинска почва. Посочената смеска константно поддържа pH=6,0–6,5. Този ниско кисел характер доближава едафичните условия на експери-мента до естествените за вида условия [7], [8], [14].Типът на почвата при “полски вариант“ е алувиално–ливаден с pH=6,9–7,3.
РЕЗУЛТАТИ И ОБСЪЖДАНЕ Морфологичен анализ Стъбло Височината на стъблото в условията на България се явява функция на
прираста за един вегетативен сезон. Установената максимална височина при двугодишни растения достига абсолютна стойност от 170 сm. (Таблица 1).
Средната дължина на стъблото през първата и втората година на експе-римента не надвишава 133,1 сm. Междувъзлията варират в границите 2,5–6,7 сm, като по–значителни са на нискостоящите между 2-ра, 3-та, 4-та и 5-та двойка листа (Таблица 1). Връхните части на стъблените разклонения са видимо по–облистени, ефект получаващ се от по-късите междувъзлия. (Снимка № 1). Стъблото е цилиндрично по форма. Размерите му в диаметрална посо- ка ва-рират от 0,3 до 0,72 сm. Не се установява правопропорционална зависимост между височината и дебелината на коментирания орган. Разклоняването на стъблото е типично симподиално. Разклоненията се формират от пазвени пъпки, което определя и псевдодихотомичния му тип, имайки предвид срещу-положното листоразположение на вида. Необходимо е да бъде отбелязан от-четения ефект на задължителен листопад в основата на всяка новоформира-ща се клонка, която се проявява след 15-20 дни от развитието на разклонени-ята. Най-често разклоненията са от 2-ри порядък. Осите от тази категория не са равнозначни по размери. Нискостоящите достигат и изпреварват височина-та на осите от 1-ви порядък, докато тези във връхната част преустановяват развитието си на фаза 2-ра-3-та двойка листа.
Лист Листата са с типично дорзи–вентрална характеристика. Петурата се ха-
рактеризира с ланцетна форма (Фиг. 2). В основата на листната петура посте-пенно преминава в много къса листна дръжка (Табл. 2) 0,2–0,3 сm, създаващ

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 997
впечатление за приседналост на листата. Прилистници не се развиват. Лист-ния ръб е ясно напилен, като размерите намаляват в посока от основата към върха на листа (Фиг. 1).
Жилкуването е типично перест тип / Фиг. 2 /, с ясно изпъкнали към долна-та листна повърхност централна жилка и страничните ù разклонения. Разме-рите на листата, със средна дължина–5,8 сm и ширина 1,6 сm са константни над 4-тия стъблен възел / Табл. 2 /. Листните параметри противно на очаквани-ята не се променят под влияние на екологичните особености. Засенчването оказва видим ефект по-скоро в удължаване на междувъзлията, отколкото в промяна на параметрите на листната повърхност.
Корен Stevia rebaudiana Bertoni развива силна алоризна коренова система при
растения получени от семена. При разклоняването на главния корен не се ус-тановява етажност (Фиг. 3). Страничните корени са равномерно разпределени по цялото протежение на главния корен. При допира на обезлистените разк-лонения с почвата се формират единични адвентивни корени в зоната на въз-лите и в мястото на прекия контакт между растителния орган и субстрата, кое-то постепенно променя кореновата система до смесен тип.
Опитите за вегетативно размножаване чрез резници показват поява на адвентивни корени по междувъзлията. Броят им е между 5 и 11, и причинява развитие на хоморизна коренова система. По време на двугодишния период на изследване разклонения по добавъчните корени не са установени. Форми-ране на генеративни пъпки, съцветия и плодове при всички варианти на екс-перимента не са наблюдавани, което може би се дължи на ювенилния харак-тер на отглежданите растения.
Морфологичния анализ на отглежданите в условията на Южна България растения от Stevia rebaudiana Bertoni показва запазване на характерните за вида основни морфологични белези–форма на стъблото и листата тип на ко-реновата система, листоразположение, разполагане на стъблото [22], [17], [19], [1].Постоянни морфологични белези са и тези свързани с репродуктивни-те органи–цветове и плодове [16], [15] които в нашия експеримент не са уста-новени.
Морфометричните показатели на вегетативните органи обаче (височина на стъблото, дължина и ширина на листата) отбелязват съществени отлики от установените в предишни изследвания [11], [12], [6], [7]. Така при култивира-ните растения височината от 0,8–1,0 m (в естествени условия) е значително надвишена с повече от 30–40 %. Подобна тенденция се отбелязва и за фоли-арните параметри изпреварващи тези на развиващите се в естествена среда растения с повече от 25 %.
Анатомичен анализ Стъбло Вътрешния строеж на стъблото с първичен произход е типично с несноп-
често устройство на проводящите тъкани.Външния цилиндър е сведен до 7-10 реда клетки, от които най-външния слой е епидермален,сравнително дребни клетки покрити с тънък кутикулен слой с характерна скулптуризация (Фиг. 4).
Под епидермиса се разполага двуреден рехав коленхим. Навътре от ме-ханичната тъкан се наблюдава ендодермис от 3-4 реда клетки. Перицикълът с

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 998
пунктационно разполагане има вид на масивни снопчета от по няколко десет-ки склеренхимни фибри. Дървесната проводяща тъкан съдържа почти равни количества трахеи и механични влакна. Процента на дървесния паренхим не надвишава 10 от общия обем на ксилема.
Централната стъблена част е заета от основен паренхим, чийто клетки се отличават със сферична форма, тънкостенни са и оформят помежду си зна-чителни междуклетъчни пространства (Фиг. 4). Преходът към вторично уст-ройство се отбелязва в количествени изменения на тъканите.
В състава на кората групичките от склеренхимни влакна в резултат на диаметралния растеж се раздалечават. Не се установяват механични тъкани с вторичен произход.
Вътрешния цилиндър е представен от плътен вторичен ксилем, отнасящ се към типа пръснатопореста дървесина (Снимка № 5 и № 6). Количеството на проводящите елементи в тази тъкан е в границите на 15-20 % от общия обем на ксилема. Останалата част се пада на дебелостенен дървесен парен-хим и механични влакна. В центъра на вторично устроеното стъбло се запаз-ва основната паренхимна тъкан, остатък от стъблото с първичен произход и естествено отличаваща се с вече коментираните цитологични характеристики.
Лист Листата на Stevia rebaudiana Bertoni са с ясно изразен бифациален ха-
рактер. Горния епидермис е сравнително едроклетъчен с тънка кутикула по външната клетъчна стена. По форма клетките са изодиаметрични с нагънати стени, плътно допрени помежду си (Фиг. 5). Макар и в по-малко количество в сравнение с долната листна повърхност по горния епидермис се наблюдават устица (Табл. 3), (Фиг. 6).
Покривната тъкан по долната листна повърхност формира жлебове, по чийто изпъкналости са разположени устицата върху листната повърхност (Фиг. 7).
Мезофилът е двуслоен (Снимка № 8). Горен, от палисаден хлоренхим, който е съставен от един ред силно удължени клетки и заема до 40 % от вът-решната листна структура. Гъбчест отличаващ се с незначителни междукле-тъчни пространства (Фиг. 7), които са около два пъти по–големи от клетките на аериращата тъкан единствено в зоните на въздухоносните стаички на ус-тицата по долната листна повърхност (Табл. 5).
Долният листен епидермис е съставен от основни клетки подобни по форма на тези при горния, но малко по–големи и повече на брой (Табл. 4).
Необходимо е да се отбележи наличието на оребряване на долния епи-дермис. По оформящите се изпъкналости се разполагат устицата, в най-дисталната част на структурата (Фиг. 7). По долния и горния листен епидер-мис се разполагат многоклетъчни, едноредни, покривни трихоми, които са с еднаква гъстота.
Централната жилка е силно изпъкнала към долната повърхност (Фиг. 8) При младите листа проводящите тъкани са представени от затворено колате-рално снопче, съотношението на двете проводящи тъкани в него е 8/2 в полза на ксилема, което най-вероятно е свързано с огромния разход на вода от лис-тата на Сладката стевия. Над проводящите тъкани, а също и към долната по-върхност се оформят масивни склеренхимни дъги (по–силно изразени под

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 999
проводящото снопче) (Снимка № 7). Останалата част от жилката е заета от основен паренхим, съставен от изодиаметрични клетки с задебелени стени.
Устицата и по двете листни повърхности са от аномоцитен тип. Отлича-ват се със сравнително големи размери както на затварящите клетки, така и на остиолума.
Корен Вътрешния строеж на корена е типичен за двусемеделните представите-
ли.Сложното радиално снопче заема не повече от 15 % от общия обем на осевия орган (Снимка № 9). Снопчето е от трикомпонентен тип (три ксилемни и три флоемни участъка), като централната му част е заета от дебелостенен паренхим (Снимка № 10).
Граничните тъкани (между външния и вътрешния цилиндър), ендодермис и перицикъл не са ясно диферинцирани. Кората е съставена изключително от тънкостенен резервен паренхим. Клетките на тази тъкан са едроразмерни, изодиаметрични по форма и със значителни междуклетъчни пространства. Външната обвивка на корена е представена от 2-3-редов екзодермис, прите-жаващ цитологични характеристики на модифицирана покривна тъкан.
Проследяването на анатомичните характеристики на Сладката стевия, което за първи път се извършва в настоящия експеримент определят Stevia rebaudiana, като мезоморфен към отклонения на хигроморфен вид.
Основание за такова заключение намираме в някои скали за епидерма-лен анализ [5], [2], [4].
Особеностите в строежа на мезофила, дебелината на кутикулата, поло-жението на устицата, се допълват и от такива значими показатели, като коли-чеството на устицата на единица повърхност и размери на основните епидер-мални клетки [3], [13], [4], [21].Съвкупността от коментираните структурни по-казатели дава основание изследвания вид да бъде отнесен към групата на мезофилите с определено отклонение към хидрофитната екологична катего-рия.Доказаната ниска толерантност на Сладката стевия към температури под 15 °С [7], [10], [8], [16], [14] в ранните съобщения за вида, се доказва и като пряка зависимост от условията за култивация. Така растенията отглеждани в оранжерия при t >30 °С се отличават с по-голям прираст на височина, по-висока степен на разклоняване на стъблото и по-значителна облистеност на клоните от втори, трети и по-висш порядък. Тези особености определят и по-вишената продуктивност (количество суха листна маса), при моделирани ус-ловия на отглеждане. Получените от нас резултати са близки до тези при пол-ските условия на Индонезия и Танзания [19], [18] а също и до оранжерийната култивация в Япония [20] и Канада [9].
ЗАКЛЮЧЕНИЕ 1. Морфологична характеристика на растенията от Stevia rebaudiana
Bertoni отгледани в условията на България ги доближава в по-голяма степен до диворастящия аналог отколкото до културните форми създадени в Азия и Америка;
2. Структурните особености на листата определят вида като мезоморфен с отклонения към хигроморфен;

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1000
ЛИТЕРАТУРА [1] Крумов И., М. Върбанов, 1984, Растението стевия (Stevia rebaudiana
Bertoni) нов вид в културната флора на България. Известия,ДКНТП, Со-фия, 21, 162-171;
[2] Куртева М., 1982, Биологическии особености древесных растении в усло-вия промышленого загразнения атмосферы зоне Украины, (автореферат) Киев, стр. 24;
[3] Николаевский В. С., 1963, О показателях газаустойчивости растений, Труд. Ин-та биологии УФАН, вып, 31;
[4] Нинова Д., 1970, Върху анатомическите показатели за димоустойчевост на някои дървесни видове, Горско стоп. Наука, 4, 9-16;
[5] Сергейчик С., 1980, Устойчивосты древесных растений ксероводороду, двуокиси серы и биологическая очистка атмосферного воздуха в услови-ях Белорусии (автореферат), Днепропетровск, стр. 20
[6] Чудновский Б., О. Запольский, 1990, Способы выращивании рассады сте-вии из материала, полученного in vitro, в Ввдение в културу стевииисточ-ника низкокалорийного заменителя сахара, Киев ВНСИ, стр.9-13;
[7] Bertoni M., 1905, Anales Cient Paraguayos, Vol. Ser. I, 5 (1); [8] Bertoni M., 1918, Stevia rebaudiana Bertoni, Steviana and rebaudiana, new
sweetening substance. Anales Cient Paraguayos, Enero Vol. II (2) 1-4; [9] Boxus P., 1987, In vitro vegetative propagation of plants, Nestle Res., News,
Vevey, p. 73-79; [10] Dieterich M., 1909, Composition of Eupatorium rebaudianum. Chemist and
druggiest, p. 1; [11] Katayama O., O. Sumida, T. Hayashi and Mutsuhashi, 1976, The practical ap-
plication of Stevia and research and development data (English translation) I.S.U. Company, Japan. 747pp
[12] Lewis W., 1992, Early uses of Stevia rebaudiana (Asteracea) leaves as a sweetener in Paraguay Econ. Bot. 46: 336-337;
[13] Mansfield T., O. Majernic, 1970, Can stomata play a part in protecting plants agains air pollutans?, Environ. Pollut., 80: 959;
[14] Melville, 1941, Stevia rebaudiana as a substitute for sugar and saccharum memo to the director 1p. England Royal Botanic Gardens (English) H;
[15] Oddone B., 1997, How to grow stevia. Technical manual. Guarani botanicals, Pawtucket, CT;
[16] Robinson B. L., 1930, Contributions from the Gray Harvard University, Cam-bridge;
[17] Rocha, 1966, Effects of photoperiod and growth regulators on growth and flowering of Stevia rebaudiana Bertoni Jap. J. Crop. Sci. 46: 243-248;
[18] Saxena S., and Ming, 1988, Preliminary harvesting characteristics of Stevia. Phys. Prop. Agric. Mat. Prod. 3: 299-303;
[19] Shock C., 1982, Experimental cultivation of Rebaudi’s Stevia in California. Univ. California, Davis Agron. Progr.Rep.122;
[20] Shyu Y., Liu, Lu, Wu, Su, 1994, Effects of harvesting dates on the characteristics yield and sweet components of Stevia lines. J. Agric. Res. China 43: 29-39;
[21] Swiecki T., A. Endress, O. Taylor, 1989, Hystologycal effects of aqueous acids and gaseous hydrogen chloride on bean leaves, Amer.J. Bot., 69: 141-149;
[22] ValioI., 1966, Effects of photoperiod and growth and flowering of Stevia re-baudiana Bertoni Jap. J crop. Sci. 46: 243-248.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1001
ПРИЛОЖЕНИЯ Снимка 1 Снимка 2 Контейнерно отглеждане в Полеви експеримент оранжерия Снимка 3 Снимка 4. Вторично устройство на стъбло Напречен пререз от централна жилка на лист Снимка 5 Напречен пререз от корен
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1002
Fig. 1 Онтогенетично развитие на лист (1-ва – 20-та двойка листа) Fig. 2 Fig. 3 Форма и жилкуване на лист Тип на кореновата система Fig. 4 Напречен пререз на първично устроено стъбло
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1003
Fig. 5 Fig. 6 Лист – горен епидермис Лист – долен епидермис Fig. 7. Напречен пререз на листна пластинка Fig. 8 Напречен пререз на централна жилка
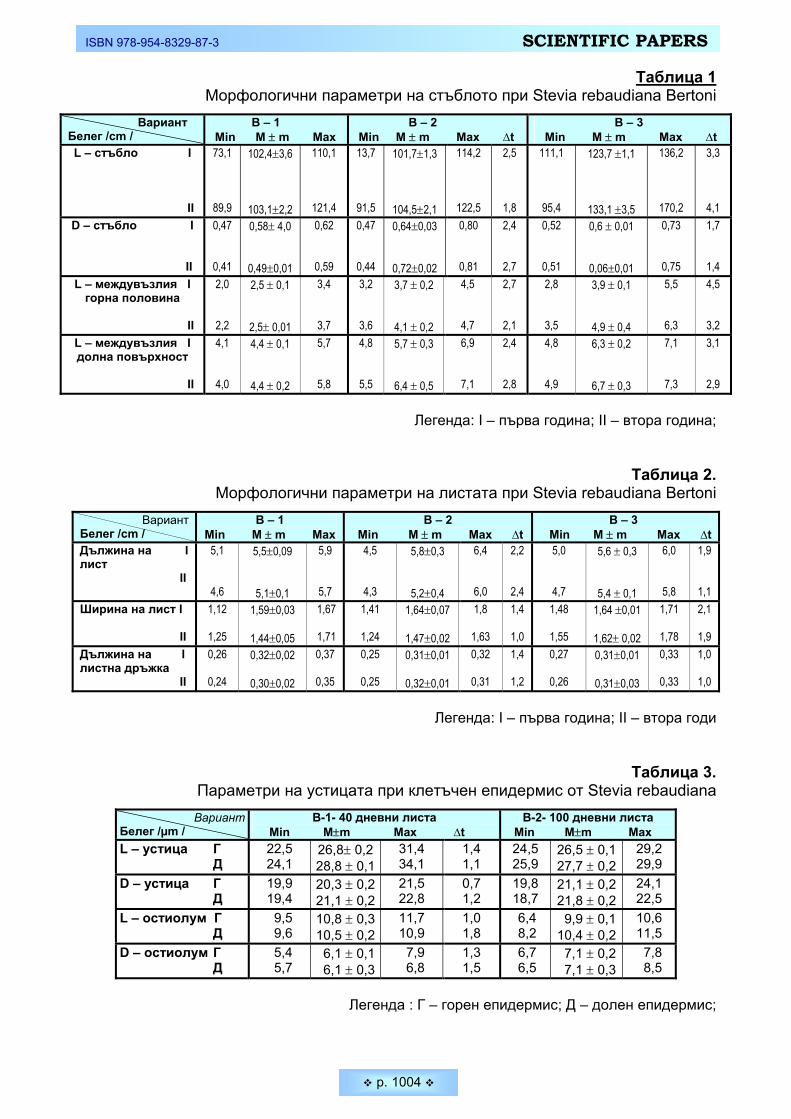
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1004
Таблица 1 Морфологични параметри на стъблото при Stevia rebaudiana Bertoni
Вариант Белег /cm /
B – 1 Min M ± m Max
B – 2 Min M ± m Max ∆t
B – 3 Min M ± m Max ∆t
L – стъбло І
ІІ
73,1
89,9
102,4±3,6
103,1±2,2
110,1
121,4
13,7
91,5
101,7±1,3
104,5±2,1
114,2
122,5
2,5
1,8
111,1
95,4
123,7 ±1,1
133,1 ±3,5
136,2
170,2
3,3
4,1 D – стъбло І
ІІ
0,47
0,41
0,58± 4,0
0,49±0,01
0,62
0,59
0,47
0,44
0,64±0,03
0,72±0,02
0,80
0,81
2,4
2,7
0,52
0,51
0,6 ± 0,01
0,06±0,01
0,73
0,75
1,7
1,4 L – междувъзлия І горна половина
ІІ
2,0
2,2
2,5 ± 0,1
2,5± 0,01
3,4
3,7
3,2
3,6
3,7 ± 0,2
4,1 ± 0,2
4,5
4,7
2,7
2,1
2,8
3,5
3,9 ± 0,1
4,9 ± 0,4
5,5
6,3
4,5
3,2 L – междувъзлия І долна повърхност
ІІ
4,1
4,0
4,4 ± 0,1
4,4 ± 0,2
5,7
5,8
4,8
5,5
5,7 ± 0,3
6,4 ± 0,5
6,9
7,1
2,4
2,8
4,8
4,9
6,3 ± 0,2
6,7 ± 0,3
7,1
7,3
3,1
2,9
Легенда: І – първа година; ІІ – втора година;
Таблица 2. Морфологични параметри на листата при Stevia rebaudiana Bertoni
Вариант Белег /cm /
B – 1 Min M ± m Max
B – 2 Min M ± m Max ∆t
B – 3 Min M ± m Max ∆t
Дължина на І лист ІІ
5,1
4,6
5,5±0,09
5,1±0,1
5,9
5,7
4,5
4,3
5,8±0,3
5,2±0,4
6,4
6,0
2,2
2,4
5,0
4,7
5,6 ± 0,3
5,4 ± 0,1
6,0
5,8
1,9
1,1 Ширина на лист І ІІ
1,12
1,25
1,59±0,03
1,44±0,05
1,67
1,71
1,41
1,24
1,64±0,07
1,47±0,02
1,8
1,63
1,4
1,0
1,48
1,55
1,64 ±0,01
1,62± 0,02
1,71
1,78
2,1
1,9 Дължина на І листна дръжка ІІ
0,26
0,24
0,32±0,02
0,30±0,02
0,37
0,35
0,25
0,25
0,31±0,01
0,32±0,01
0,32
0,31
1,4
1,2
0,27
0,26
0,31±0,01
0,31±0,03
0,33
0,33
1,0
1,0
Легенда: І – първа година; ІІ – втора годи
Таблица 3. Параметри на устицата при клетъчен епидермис от Stevia rebaudiana Вариант
Белег /µm / B-1- 40 дневни листа
Min M±m Max ∆t B-2- 100 дневни листа
Min M±m Max L – устица Г Д
22,5 24,1
26,8± 0,228,8 ± 0,1
31,4 34,1
1,4 1,1
24,5 25,9
26,5 ± 0,1 27,7 ± 0,2
29,2 29,9
D – устица Г Д
19,9 19,4
20,3 ± 0,221,1 ± 0,2
21,5 22,8
0,7 1,2
19,8 18,7
21,1 ± 0,2 21,8 ± 0,2
24,1 22,5
L – остиолум Г Д
9,5 9,6
10,8 ± 0,310,5 ± 0,2
11,7 10,9
1,0 1,8
6,4 8,2
9,9 ± 0,1 10,4 ± 0,2
10,6 11,5
D – остиолум Г Д
5,4 5,7
6,1 ± 0,1 6,1 ± 0,3
7,9 6,8
1,3 1,5
6,7 6,5
7,1 ± 0,2 7,1 ± 0,3
7,8 8,5
Легенда : Г – горен епидермис; Д – долен епидермис;
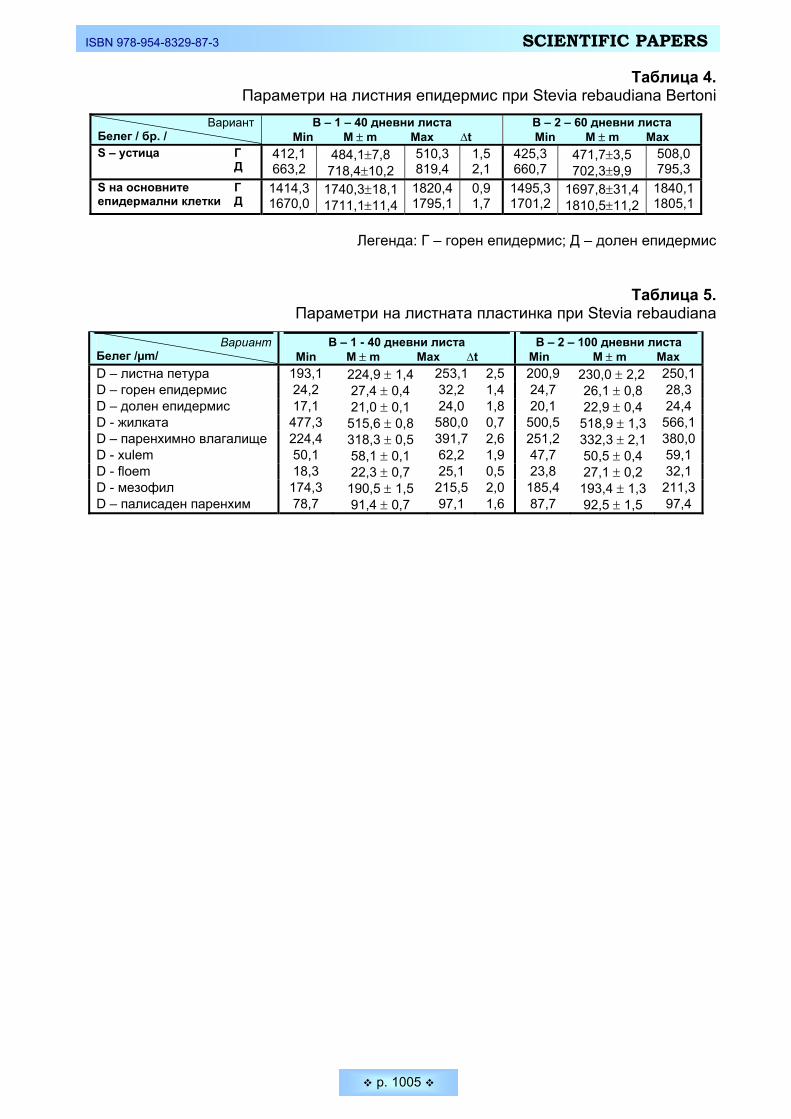
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1005
Таблица 4. Параметри на листния епидермис при Stevia rebaudiana Bertoni
Вариант Белег / бр. /
В – 1 – 40 дневни листа Min M ± m Max ∆t
B – 2 – 60 дневни листа Min M ± m Max
S – устица Г Д
412,1 663,2
484,1±7,8 718,4±10,2
510,3 819,4
1,5 2,1
425,3 660,7
471,7±3,5 702,3±9,9
508,0 795,3
S на основните Г епидермални клетки Д
1414,3 1670,0
1740,3±18,11711,1±11,4
1820,41795,1
0,9 1,7
1495,31701,2
1697,8±31,4 1810,5±11,2
1840,11805,1
Легенда: Г – горен епидермис; Д – долен епидермис
Таблица 5. Параметри на листната пластинка при Stevia rebaudiana
Вариант Белег /µm/
B – 1 - 40 дневни листа Min M ± m Max ∆t
B – 2 – 100 дневни листа Min M ± m Max
D – листна петура 193,1 224,9 ± 1,4 253,1 2,5 200,9 230,0 ± 2,2 250,1D – горен епидермис 24,2 27,4 ± 0,4 32,2 1,4 24,7 26,1 ± 0,8 28,3 D – долен епидермис 17,1 21,0 ± 0,1 24,0 1,8 20,1 22,9 ± 0,4 24,4 D - жилката 477,3 515,6 ± 0,8 580,0 0,7 500,5 518,9 ± 1,3 566,1D – паренхимно влагалище 224,4 318,3 ± 0,5 391,7 2,6 251,2 332,3 ± 2,1 380,0D - xulem 50,1 58,1 ± 0,1 62,2 1,9 47,7 50,5 ± 0,4 59,1 D - floem 18,3 22,3 ± 0,7 25,1 0,5 23,8 27,1 ± 0,2 32,1 D - мезофил 174,3 190,5 ± 1,5 215,5 2,0 185,4 193,4 ± 1,3 211,3D – палисаден паренхим 78,7 91,4 ± 0,7 97,1 1,6 87,7 92,5 ± 1,5 97,4

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 1006
SCIENTIFIC PAPERS
АНТРОПОМЕТРИЧНА ХАРАКТЕРИСТИКА ПРИ ДЕЦА И ПОДРАСТВАЩИ
СПОРЕД ВЪЗРАСТОВО-ПОЛОВАТА ИМ ПРИНАДЛЕЖНОСТ
Зорка Митова Институт по експериментална морфология и антропология с музей,
БАН, София, България
ANTHROPOMETRICAL CHARACTERIZATION IN CHILDREN AND ADOLESCENTS ACCORDING
TO THEIR AGE – GENDER BELONGING
Zorka Mitova Institute of Experimental Morphology and Anthropology with Muzeum,
BAS, Sofia, Bulgaria
ABSTRACT: In a cross-sectional study of growth, 1142 schoolchildren (569 boys, 573 girls) from
the city of Sofia (Bulgaria) were measured. Five linear features (stature, biacromial breadth, bicristal breadth, chest breadth and chest depth), three circumferences (chest, waist and hip) and two indices (acromiocristal and waist to hip ratio) characterizing body building were studied in the age span of 9 to 15 years. A significant average annual in-crease of stature were observed up to 15 years of age in boys and 13 years of age in girls, showing that girls had a shorter growing period. Between 9 and 11 years of age, boys were overgrown by girls in stature (statistically significant at the age of 11) due to the ear-lier onset of puberty in girls.The growth gains for girls when compared with those for boys show different patterns across variables. The girls’ growth advantage remained in two-three years period for stature and other features. The boys’ statistically significant pre-dominance in size had an onset at the age 11 for waist circumference and at the age 12 for biacromial breadth, bicristal breadth, chest breadth, chest depth and chest circumfer-ence. During the entire investigated period – 9–15 years, girls have larger hip circumfer-ence, than boys (statistically significant at the age of 10, 11 and 13). The sing of sexual dimorphism and growth dynamics in stature and some of the investigated features during growing up years are similar to those observed in other populations living in Balkan Penin-sula.
Key words: adolescents, growth patterns, gender, anthropometry, body building
ВЪВЕДЕНИЕ Един от най-интересните онтогенетични периоди при човека е този, през
който в детския организъм започват бурни процеси на растеж, развитие и по-лово съзряване [26]. В резултат на това за няколко години при подрастващите се формират фенотипните белези, характерни за мъжкия и женския тип те-

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1007
лостроеж. Установяването на особеностите във физическото развитие на де-цата и подрастващите е важно условие за изясняване на връзката между нас-ледствената информация и адаптационните възможности за промени в те-лостроежа при конкретните условия на живот [12, 13, 16, 17, 18, 26]. Поради това въпросите, свързани с възрастовата изменчивост имат изключително го-ляма медико-биологична и социална значимост. В тази връзка натрупването на богата база от антропологични данни за всяко ново поколение дава въз-можност за коректно сравняване на резултатите и установяване на промените в техния антропометричен статус. Актуалността на изследванията, свързани с процесите на растеж и развитие, определя постоянния интерес и на нашите специалисти към ученици от различни региони на страната [3, 5, 6, 7, 8, 11, 14, 15, 23]. Особено ценни в това отношение са антропометричните изследвания, осъществени на границата между две хилядолетия и в преходен период на социално-икономическо развитие.
Това определи и целта на настоящата статия, а именно – да се предста-вят особеностите във физическото развитие на деца и подрастващи от Со-фия, живеещи в началото на 21-ви век, като се оцени кога и как се осъществя-ва полово-свързаната диференциация в техния телостроеж между 9-15 го-дишна възраст.
МАТЕРИАЛ И МЕТОДИ При трансверзално антропометрично проучване проведено в столицата
през периода март 2001 – юни 2002 г. са изследвани 569 момчета и 573 мо-мичета, на възраст между 9 и 15 години (разпределени средно по 82-ма души, в по седем възрастово-полови групи). Всяка едногодишна възрастова група включва подрастващи на навършена възраст от n години до n години, 11 ме-сеца и 29 дни (при трансформиране в десетична възраст – напр. от 9,00 до 9,99 години).
От подробната програма на изследването, в настоящия анализ са вклю-чени десет от най-често използваните основни анропометрични показатели за мъжкия и женския тип телостроеж. Тези показатели характеризират метрично височината, широчинното и обемно развитие на горната и долната части на тялото, а именно: ръст, биакромиален диаметър (БАД), бикристален диа-метър (БКД), акромиокристален индекс (АИ), представляващ процентното отношение на бикристалния към биакромиалния диаметър, напречен (НДГК) и сагитален диаметър на гръдния кош (СДГК), гръдна обиколка при пауза (ОГКП), обиколките на талията (ОТ) и ханша (ОХ), както и отношението талия/ханш (Т/ХО). За всеки от тях са дадени: средната стойност ( x ), стан-дартното отклонение (SD), стандартната грешка на средната (SEM), мини-малната (Min), максимална (Max) стойност и коефициента на вариация (CV).
Обективната, сравнителна оценка на междувъзрастовите различия се ос-новава на статистическата достоверност на метричните различия при всеки признак на подрастващите от последователните възрастови групи. Тя е опре-делена чрез едномерен дисперсионен анализ ANOVA, при който се осъщест-вяват всички възможни подвойкови сравнения на разликите в извадковите средни, като се запазва предварително фиксираната вероятност за грешка от тип Іα непроменена при определено ниво на значимост. В случая е приложена най-гъвкавата Post Hoc процедура – Мultiple comparison Scheffe test, чието из-ползване се препоръчва при подвойкови сравнения на извадки с различен

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1008
обем [2]. Тъй като методът на Scheffe е консервативен, проверката на теста и анализа на резултатите са извършени при ниво на значимост p=0.10 [2]. Пре-димството на Post Hoc Scheffe метода се изразява и в това, че той обединява в еднородни групи тези извадки, които са хомогенни по дисперсии и вариа-ции на съответните признаци и чрез средните им стойности онагледява това групиране в табличен вид. В текста при анализа на резултатите сме използ-вали понятието “еднородна размерна група” (ЕРГ), което в конкретния слу-чай отразява биологичния смисъл на това групиране.
Достоверността на междуполовите различия при отделните признаци е установена чрез t-теста на Student (при р<0.05, p<0.01 и p<0.001).
За всеки измерен признак допълнително са изчислени абсолютния (АГП, в cm) и относителния годишен прираст (ОГП, в %). Въпреки, че при транс-верзални проучвания тези показатели до известна степен са условни, те да-ват определена информация за абсолютната и относителната растежна ско-рост на съответните признаци. Изчислени са по следните формули:
АГП = ( x n+1 – x n); ОГП = 2 * ( x n+1 – x n) * 100 / ( x n + x n+1), където x n е средната стойност на признака при подрастващите от определена възрас-това група, а x n+1 – на съответната предходна възрастова група.
Относителната сила на междуполовите различия е определена чрез ин-декса за междугрупови сравнения (ИМР), по формулата:
ИМР = 2 * ( x ♂ – x ♀) * 100 / ( x ♂ – x ♀), където с x ♂ е означена средната стойност на признака при момчетата и с x ♀ – при момичетата от същата въз-растова група. Относителните стойности на ИМР са представени в индексни единици (±ИЕ) като положителният знак пред ИЕ показва метричен приоритет за момчетата, а отрицателният – метричен приоритет за момичетата.
РЕЗУЛТАТИ И ОБСЪЖДАНЕ Биостатистическите данни за анализираните антропометрични признаци
са представени по възрастово-полови групи в Таблица 1. Статистически дос-товерните междувъзрастови и междуполови различия според теста на Sсheffe и t – теста на Student са отбелязани съответно с “∗” и “#”.
Ръст Ръстът е основен показател за физическото развитие на индивида, който
през периода на усилен растеж обобщено определя спецификата на настъп-ващите с възрастта промени в антропометричната характеристика на подрас-тващите (Фиг. 1).
Годишното нарастване на ръста при момчетата е достоверно през целия период между 9-та и 15-та година, а при момичетата – от 9-та до 13-та година. Максимална растежна скорост на височината на тялото при момичетата се отчита между 10 и 11 годишна възраст или две-три години по-рано, отколкото при момчетата – между 12 и 13 годишна и между 13 и 14 годишна възраст. При 14 и 15 годишните момичета нарастването на височина е минимално.
При групирането на момчетата според височината на тялото, Scheffe теста разграничава всяка извадка от седемте възрастови групи в самостоя-телна еднородна размерна група по този признак, отразявайки постоянно го-лямата растежна скорост на техния ръст. (Табл. 2.1). В самостоятелни едно-родни ръстови групи са разграничени и четирите извадки на момичетата от 9 до 12 годишна възраст (Табл. 2.2). Поради хомогенността на дисперсиите и

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1009
близките средни стойности на ръста, Post hoc Scheffe теста обединява данни-те на 13, 14 и 15 годишните момичета в петата ЕРГ по този признак.
Резултатите от междуполовите сравнения (илюстрирани на Фиг. 11) по-казват, че от 9 до 11 годишна възраст момичетата са по-високи от момчетата, статистически достоверно само на 11 годишна възраст (ИМР= -1,7 ИЕ). По-ускореното нарастване на височината на тялото при момичетата през тези възрасти отразява по-ранното начало на тяхното пубертетното развитие. След 11-та година момчетата нарастват по-ускорено и на 14 и 15 годишна въз-раст те вече са статистически достоверно по-високи от момичетата (съответ-но с 5,0 ИЕ и 7,3 ИЕ).
По-ранното настъпване на пубертетният растежен скок (ПРС) на ръста при момичетата – между 10 и 11 годишна възраст се изразява в достоверното надрастване по височина на момчетата, с максимален абсолютен годишен прираст от 7,6 cm. При момчетата началото на пубертетният растежен скок на ръста се отчита между 11 и 12 годишна възраст, с последващото надрастване на момичетата по височина. При 13 и 14 годишните момчета са установени максимални стойности на абсолютния годишен прираст от 7,35 cm.
ПРС на височината на тялото при момичетата продължава до 13 годиш-на, а при момчетата до 14 годишна възраст, след което нарастването при момчетата плавно намалява, а при момичетата почти спира.
При подрастващите коефициентите на вариация на ръста по време на ПРС са най-високи. Към края на пубертетното им растежно ускорение между-половите различия вече са с характеристики за израсналите мъже и жени. Също така вариациите във височината на тялото при момчетата и момичета-та са в по-тесни граници, дори и от тези при извадките на 9 и 10 годишна въз-раст.
Широчинни размери на тялото Широчинните размери на раменния пояс, на гръдния кош и на таза, опре-
делят размерността и развитието на горната и долната части на тялото и са основни характеристики при оценка спецификата на мъжкия и женски тип те-лостроеж (Фиг. 2-5).
При подрастващите момчета достоверното годишно нарастване на ши-рочинните размери на тялото започва между 10 и 11 годишна възраст, проя-вено при размерите на БАД, БКД и НДГК. Между 10-та и 11-та година са отчете-ни и максимални АГП и ОГП при тазовата широчина и напречната широчина на гръдния кош. До края на изследвания период (11-15 годишна възраст) при момчетата ежегодно достоверно нараства само биакромиалната раменна ши-рочина, като най-голяма е растежната скорост между 13-та и 14-та година. Това е междувъзрастовият етап, през който таза и гръдния кош отново нарастват достоверно в трансверзално направление. Последният нараства вече и в са-гитално направление (Табл. 6.1).
При момичетата широчинните размери на раменете, гръдния кош и таза нарастват достоверно само между 10 и 11 и между 12 и 13 годишна възраст. Максималната растежна скорост при БАД и БКД е отчетена между 10-та и 11-та година, докато при НДГК – между 12-та и 13-та година, когато при тях е регист-рирано и единственото достоверно годишно нарастване на предно-задния ди-аметър на гръдния кош.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1010
При подрастващите и от двата пола ПРС на широчинните размери на тя-лото започва между 10 и 11 годишна възраст, докато краят му при момичетата настъпва на 13-та, а при момчетата - на 14-та година. Единствено растежната скорост на БАД (подобно на ръста) при момчетата остава сравнително голяма до 15 годишната им възраст. При момчетата от последната възрастова група стойностите на CV на широчинните размери на тялото намаляват до тези, ус-тановени при извадката на 9 годишните.
Тези резултати са обективизирани и от теста на Scheffe с групирането на подрастващите в еднородни размерни групи по съответните признаци (Табл. 3-6). При всички широчинни признаци на тялото извадките на 9 и 10 годишни-те момчета и момичета са обединени в първата ЕРГ. След началото на ПРС, извадките на 11, 12 и 13 годишните момчета формират две (при БКД и НДГК) или три (при СДГК) препокриващи се (при извадката на 12 годишните момче-та) еднородни размерни групи. Това препокриване илюстрира от една страна по-плавното и недостоверно нарастване на съответните размери при момче-тата между 11 – 12 и между 12 – 13 годишна възраст, а от друга – достовер-ните различия между момчетата от несъседните възрастови групи (между 11 и 13 годишните момчета). С други думи, достоверни са различията по съот-ветните признаци само между момчетата от извадките, намиращи се извън зоната на препокриване на две еднородни размерни групи.
Във връзка с по-тесните граници на ПРС при момичетата, почти при всички широчинни размери на тялото (с изключение на СДГК) теста на Scheffe обединява извадките на 11 и 12 годишните в една еднородна размерна група, която не се препокрива с останалите две (с първата ЕРГ, включваща 9 и 10 годишните и третата ЕРГ, включваща 13, 14 и 15 годишните момичета). Тъй като годишното нарастване на СДГК при момичетата е достоверно само към края на ПРС (на 13-та година), съответно първите две ЕРГ се препокриват (при размерите на 10 годишните момичета), а в третата еднородна размерна група (вече без препокриване) са обединени данните за предно-задния диаметър на гръдния кош отново на 13, 14 и 15 годишните момичета.
Междуполовите различия и при четирите общотелесни диаметъра са с подобен характер (илюстрирано на Фиг. 11), като между 9-та и 11-та година те са минимални или незначителни. За първи път статистически достоверни междуполови различия се установяват на 12 годишна възраст, когато момче-тата вече са с по-големи широчинни размери на тялото от момичетата. Силно проявени междуполови различия, с подчертан приоритет за момчетата, се ус-тановяват при 14 и 15 годишните подрастващи. На 15 годишна възраст меж-дуполовите различия са най-интензивни при СДГК (ИМР=10,0 ИЕ), следвани от тези при НДГК и БАД (с по 9,4 ИЕ), а при широчината на таза те са с интен-зивност от по-ниска степен (ИМР=6,3 ИЕ).
Акромиокристален индекс (АИ) Чрез този индекс се обективизира съразмерността между широчинните
размери в долната и горната части на туловището, което улеснява сравня-ването на измененията във формата на тялото през различните възрасти (Фиг. 9).
При момчетата от началото до края на проучвания период акромиокрис-талният индекс намалява недостоверно. За разлика от тях, при момичетата между 9-та и 14-та година относителният дял на бикристалния, спрямо биакро-

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1011
миалния диаметър постепенно се увеличава (недостоверно). При 15 годишни-те момичета, обаче акромиокристалният индекс намалява до стойностите, от-четени на 9-та и 10-та година.
В конкретния случай по-информативни данни за изменението в съотно-шението между широчините на долната и горната части на туловището се по-лучават от междуполовите сравнения (Фиг. 11). Момичетата от всички възрас-тови групи (с изключение на 11 годишните) са с по-високи стойности на АИ, в сравнение с тези при момчетата. Според t-теста на Student статистически достоверни по този показател са междуполовите различия от 12-та до 15-та го-дишна възраст, когато относителния дял на бикристалния, спрямо биакроми-алния диаметър при момчетата плавно намалява, а при момичетата остава сравнително постоянен. Тези резултати показват, че още на 12 годишна въз-раст при подрастващите вече са формирани специфичните за младите израс-нали мъже и жени особености в типа телостроеж, а именно – при момчетата пропорциално по-широк раменен, а при момичетата съответно пропорциално по-широк тазов пояс.
Обиколни размери на тялото Анализът на данните за телесните обиколки допълва антропометричната
характеристика за формата на тялото при подрастващите, като дава инфор-мация и за неговата обемност (Табл. 7-9 и Фиг. 6-8).
При момчетата обиколните размери на гръдния кош, на талията и на ханша се увеличават статистически достоверно с еднаква, максимална рас-тежна скорост между 10-та и 11-та им година. До края на изследвания период следващо достоверно, годишно нарастване при тях се установява между 13 и 14 годишна възраст само на гръдната обиколка.
При групирането на момчетата, според достигнатите обиколни размери на тялото, тестът на Scheffe разграничава четири (при ОГКП и ОТ) и пет (при ОХ) еднородни размерни групи. При 9 и 10 годишните момчета данните и за трите обиколки на тялото са обединени в първата ЕРГ, без препокриване.
След началото на ПРС (между 10 и 11 годишна възраст) Scheffe тестът включва данните за обиколните размери на 11, 12 и 13 годишните момчета при формирането на две (при ОГКП и ОТ) или на три (при ОХ) препокриващи се (при извадките на 12-13 годишните), еднородни размерни групи. Данните за трите обхватни признака при 14 и 15 годишните момчета отново се обединя-ват от теста в последната ЕРГ, като само при ОГКП не се наблюдава препок-риване.
При момичетата от анализираните обиколни размери на тялото, статис-тически достоверно годишно нарастване се отчита само в областта на ханша (между 10-та и 11-та и между 12-та и 13-та им годишна възраст). В случая най-голяма е растежната скорост при последната възрастова група.
Специално внимание заслужава фактът, че в началото на изследвания период (между 9 и 10 годишна възраст) и трите обиколни признака на тялото при подрастващите момичета нарастват с по-голяма интензивност, в сравне-ние с тази при момчетата. При момичетата, между 9-та и 10-та им година, е от-четена максимална растежна скорост на гръдната обиколка и обиколката на талията. След 13 годишна възраст нарастването на тези телесни характерис-тики при тях спира, дори се наблюдава намаляване на гръдната обиколка. Може да се предположи, че тази външна проява на пубертетното развитие

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1012
при момичетата през посочения период, е отражение на преразпределянето на подкожната им мастна тъкан в тази област.
По-плавното и недостоверно годишно нарастване на обиколните размери в горната част на тялото (гръдната обиколка и обиколката на талията) при подрастващите момичета е онагледено и от теста на Scheffe (Табл. 7.2 и 8.2) с групирането на седемте извадки в три взаимно препокриващи се ЕРГ. От препокриването на трите ЕРГ следва, че достоверни различия в размерите на ОГКП и на ОТ има само между момичетата от извадките на първата ЕРГ (включваща 9 и 10 годишните) и тези от включените в третата ЕРГ, които са извън зоната и на препокриване с втората ЕРГ (на 13, 14 и 15 годишните). Данните за ОХ при момичетата са групирани от теста на Scheffe също в три ЕРГ, но вече без препокриване между тях (както при БАД, НДГК и БКД), отра-зявайки по-голямата интензивност в нарастването на обиколните им размери в долната част на тялото (Табл. 9.2).
Резултатите от междуполовите сравнения на обиколните размери при подрастващите показват, че от 11 годишна (при обиколката на талията) и от 12 годишна (при обиколката на гърдите) до 15 годишната си възраст, момче-тата са с достоверно по-големи размери от тези при момичетата.
Най-големи междуполови различия се установяват при 15 годишните подрастващи, когато ИМР показва приоритет за момчетата с 13,1 ИЕ (при обиколката на гърдите) и с 10,6 ИЕ (при обиколката на талията) (Фиг. 12).
През целият изследван възрастов период при подрастващите момичета обиколката на ханша е единственият от проучените признаци с постоянно по-големи размери, отколкото при момчетата (статистически достоверно на 10, 11 и 13 годишна възраст, ИМР съответно 4,5 ИЕ; 3,5 ИЕ и 4,5 ИЕ). Този факт може да се обясни с определящата роля на наследствеността при формира-нето на женския тип телостроеж в тазовата област, още преди началото на пубертета.
Талия/ханш отношение (Т/ХО) От началото до края на изследвания възрастов период при подрастващи-
те Т/ХО намалява. Това е отражение на по-интензивното нарастване на оби-колката на ханша в сравнение с тази на талията, по-силно изразено при мо-мичетата (Фиг. 10). Достоверни междувъзрастови различия при Т/ХО се уста-новяват между 10 и 11 годишна възраст (само при момичетата) и между 12 и 13 годишна възраст (и при двата пола).
Тъй като при подрастващите момчета по-интензивни са растежните про-цеси в горната, а при момичетата – в долната половина на тялото, постояннен приоритет в междуполовите различия и при Т/ХО (от 5.1 ИЕ на 9 годишна до 10,1 ИЕ на 15 годишна възраст) имат момчетата.
Според морфологичното развитие на изследваните соматометрични
признаци, пубертетното развитие при софийските момичета започва средно на 10,5 годишна възраст. Последната съответства на публикувани в литера-турата данни за възрастта, през която започва появата и на първите вторични полови белези при момичетата [5, 6, 9, 11, 22].
Възрастовата динамика на ръста и основните широчинни и обиколни признаци на тялото при софийските подрастващи по време на тяхното пубер-

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1013
тетно развитие също е с подобни характеристики на тези, публикувани в ли-тературата [3, 4, 8, 18, 26, 27].
Поради липса на актуални референтни нормативи за физическото разви-тие на децата и подрастващите в България, резултатите от настоящото изс-ледване са сравнени с последните литературни данни за 9-15 годишни со-фийски деца и подрастващи, измервани през периода 1984-1987 година [21]. При съпоставката на абсолютните размери на ръста и анализираните широ-чинни и обиколни признаци на тялото при момчетата и момичетата от тези две генерации се установяват от много близки до еднакви стойности на: ръс-та, биакромиалния диаметър, напречните и предно-задните размери на гръдния кош. За разлика от изброените признаци, при сравнението на данните за бикристалния диаметър и за обиколката на ханша при учениците, обхвана-ти от двете изследвания се установяват противоположни тенденции. Подрас-тващите, измервани през 2001-2002 г. в сравнение с връстниците им, измер-вани през 1984-1987 г. имат: от една страна сравнително по-големи размери на бикристалния диаметър (при момчетата - от 1,5 cm на 12 годишна възраст до 1,0 cm на 15 годишна възраст, а при момичетата - от 2,6 сm на 12 годишна възраст до 0,4 cm на 15 годишна възраст); а от друга - по-малки размери на обиколката на ханша (при момчетата - от ~2,0 до 3,0 cm, а при момичетата - от ~ 0,7 до 5,0 cm).
Подобни резултати са установени и при сравнението на физическото развитие на две поколения подрастващи от гр. Смолян, трансверзално проу-чени през 1986 г. и в периода 1999-2001 г. [10].
Резултатите от настоящото изследване потвърждават установените, как-то у нас, така и в международен мащаб тенденции на затихване на акцелера-ционните процеси и грацилизация на подрастващите, живеещи в края на вто-рото хилядолетие [1, 13,19, 20, 24, 25].
ЗАКЛЮЧЕНИЕ Описаните резултати дават представа за темповете на растеж, както и за
характера и степента на междуполовите различия във физическото развитие на 9–15 годишните деца и подрастващи от София, изследвани през периода 2001 – 2002 г. Тези резултати показват следното:
Момичетата между 9-та и 11-та година са по-високи от момчетата (досто-верно на 11 годишна възраст). След 11-та година момчетата нарастват на ви-сочина с по-бързи темпове и на 14-та и 15-та година те вече достоверно над-растват момичетата.
Широчините на раменете, гръдния кош и таза между 9-та и 11-та година са относително еднакви при подрастващите и от двата пола.
При момичетата като цяло темповете на нарастване са по-ускорени през първата половина на проучвания период (до 12 годишна възраст), а при мом-четата през втората.
След 12 годишна възраст при момчетата по-ускорено нарастват широчи-ната на раменете и гръдния кош, което води до пропорционално по-силно развитие на горната част на тялото, докато при момичетата продължава по-интензивното нарастване на тазовия пояс. При изследваните подрастващи се очертава подобна тенденция и при обиколните им размери в горната и долна-та части на тялото.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1014
Пубертетният растежен скок на височинно–широчинните и обиколните размери на тялото при момичетата се проявява между 10 и 13 годишна въз-раст, а при момчетата между 10-та и 14-та им година.
Пубертетното растежно ускорение при софийските подрастващи момиче-та продължава средно около 3 години. След достигане на 13 годишна възраст растежната скорост при всички общотелесни размери (с изключение на оби-колката на ханша) рязко намалява до минимални стойности.
При подрастващите момчета пубертетното растежено ускорение про-дължава средно около 4 години и има максимални стойности между 13-та и 14-
та им година. При 15 годишните момчета статистически достоверно остава го-дишното нарастване само на височината на тялото и на биакромиалния диа-метър.
При софийските подрастващи, родени в края на 20-ти век и живеещи в на-
чалото на 21-ви век, още на 12 годишна възраст са формирани специфичните особености в типовете телостроеж, характерни за младите израснали мъже и жени. Тези особености в техния телостроеж се открояват все повече с въз-растта и на 14 и 15 годишна възраст момчетата имат достоверно: по-висок ръст, по-широк раменен пояс, по-обемен гръден кош и по-голяма обиколка на талията; а момичетата - достоверно по-широк тазов пояс и по-голяма седа-лищна обиколка.
Потвърждават се установените у нас и в международен мащаб тенден-ции на затихване на акцелерационните процеси и грацилизация на подраст-ващите, живеещи в края на второто хилядолетие.
ЛИТЕРАТУРА
[1] Година, Е. Ауксология человека – наука ХХI века: проблемы и перспек-тивы. В: Антропология на пороге III тысячелетия /Ред. Алексеева, Т. И., Балановская, Е. В., Година, Е. З., Дубова, Н. А./, Москва, Российское от-деление ЕАА, 2003, т. 2, 529-566..
[2] Калинов, К. Статистически методи в поведенческите и социалните науки. София, Нов български университет, 2001, 232-275.
[3] Младенова, С. Антропологична характеристика на растежа и развитието на деца и подрастващи от Смолянски регион в съвременните условия на живот. Автореферат на канд. дис., Пловдив, 2003, 1-58.
[4] Николаидис, Т. Динамика на физическото развитие и физическата деес-пособност на учащи се от Балканския регион (Гърция, България, Турция) (Възраст: 7- 18 години). Докт. дис., С., 2000, 1-202.
[5] Рашкова-Андреева, П. Физическо развитие на учениците от Великотър-новски окръг. Канд. дис., С, 1978, 1-176.
[6] Сеизов, Хр. Пубертетното развитие по първични и вторични полови приз-наци у софийските ученички и ученици през 1953 година. Известия на ИФВУХ, С., 1957, 2, 183-207.
[7] Слънчев П., Б. Янев, Ф. Генов, П. Щерев, П. Боев, Д. Сепетлиев и Б. За-хариев. Физическо развитие, физическа дееспособност и нервно-психическа реактивност на населението на България. С., 1992.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1015
[8] Станимирова, Н. Растеж и пубертетно развитие – норми и физиологични отклонения. Автореферат на докт. дис., С, 1997, 1-59.
[9] Станчев, Здр., Н. Станимирова. Пубертет у момчета. І. Поява на вторич-ните полови белези и физическо развитие. Педиатрия, 1980, 3, 238-245.
[10] Стоев, Р., С. Младенова. Секуларни промени във физическото развитие на смолянските деца и подрастващи. Юбилейна национална научна кон-ференция с междунарадно участие “20 години СУБ – клон Смолян”, Смо-лян, 20-21 октомври 2006. Доклади, Смолян, СУБ /под печат/.
[11] Султов, К. Лонгитудинално проучване на растежа и половото съзряване на децата и юношите от Благоевград. Канд. дис., С., 1980, 1-207.
[12] Шестернина, Ж., С. Михайлова. Влияние экологических и социальных факторов на формирование физического и полового развития девочек Горного Алтая. Рос. акад. мед. наук, М., 2004, 1-91.
[13] Ямполская, Ю. Физическое развитие школников – жителей крупного мега-полиса в последние десятилетия: состояние, тенденции, прогноз, методи-ка скрининг-оценки. Автореферат на докт. дис., М., 2000, 1-76.
[14] Янев, Б., П. Щерев, П. Боев, Р. Семерджиева и Д. Сепетлиев. Физическо развитие и дееспособност на населението в България от раждане до двадесет и шест години. Издателство на БАН, С., 1965.
[15] Янев, Б., П. Щерев, П. Боев, Ф. Генов, Д. Сепетлиев, И. Попов и Б. Заха-риев. Физическо развитие, физическа дееспособност и нервно-психическа реактивност на населението. Медицина и физкултура, С., 1982.
[16] Healy, M. Statistic of growth standards. In: Human growth a comprehensive treatise. Methodology ecological, genetic and nutritional effects on growth. /Eds. Folkner F. & J. M Tanner/. New York and London: Plenum press, 1987, 3, 47-58.
[17] Malina, R. Regional Body Composition: Age, sex, and ethnic variation. In: Human body composition /Eds. Roche, F. Alex, Heymsfield, B. Steven, Lohman, G. Timothy/. Human Kinetics, 1996, 217 – 255.
[18] Satake, T., R. Malina, Sh. Tanaka and F. Kikuta. Individual variation in the sequence of peak velocity in seven body dimension. Am. Journal of Human Biology, 1994, 6, 356-367.
[19] Štefančič, M., P. Leben-Seljak. Dynamic of physical growth and development during puberty. Auxology '94, Humanbiol. Budapest, 1994, 25, 333-340.
[20] Štefančič, M., U. Arko, V. Brodar, F. Dovečar, M. Juričič, M. Macarol-Hiti, P. Leben-Seljak. An assessment of physical growth and development in children and youths in Ljubljana. Ljubljana, 1996, 23-53.
[21] Stoev, R. Physical development of adolescents from the city of Sofia and town of Smolyan in 1980s. Acta morf. et anthrop., S., 2001, 6, 164-171.
[22] Stoev, R., T. Kazakova, L. Zhelkova and L. Tsacheva. Sexual maturation and sociofamilial conditions in school boys and girls in Sofia. Anthrop. Közl, 1989/90, 131-135.
[23] Stoev, R., Y. Yordanov. Secular trend in Bulgaria. In: Secular growth in Europe /Eds. Bodzár, B. E. and C. Susanne/. Budapest, Eötvös univ. press, 1998, 65-73.
[24] Tomazo-Ravnik, T., D. Zerbo. Secular growth changes of Ljubljana schoolchildren in the period from 1958 to 1994 (Longitudinal series). Anthrop. Közl., 1996/97, 38, 61-68.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1016
[25] Tomazo-Ravnik, T. Secular trend in growth of schoolchildren in Ygoslavia. Coll. Anthrop., 12, 1, 1988, 135-140.
[26] Van Loan, M. Total body composition: Birth to old age. In: Human body composition /Eds. A. Roche, S. Heymsfield, J. Lohman/. Human Kinetics, 1996, 205–215.
[27] Živičnjak, M., N. Narančić, L. Szirovicza, D. Franke, J. Hrenović and V. Bišof. Gender-specific growth patterns for stature, sitting height and limbs length in Croatian children and youth (3 to 18 years of age). Coll. Anthrop. 27, 2003, 1, 321-334.
н. с. II ст. Зорка Митова секция “Антропология”, ИЕМАМ - БАН тел: +359 02 979 23 17 е-mail: [email protected]
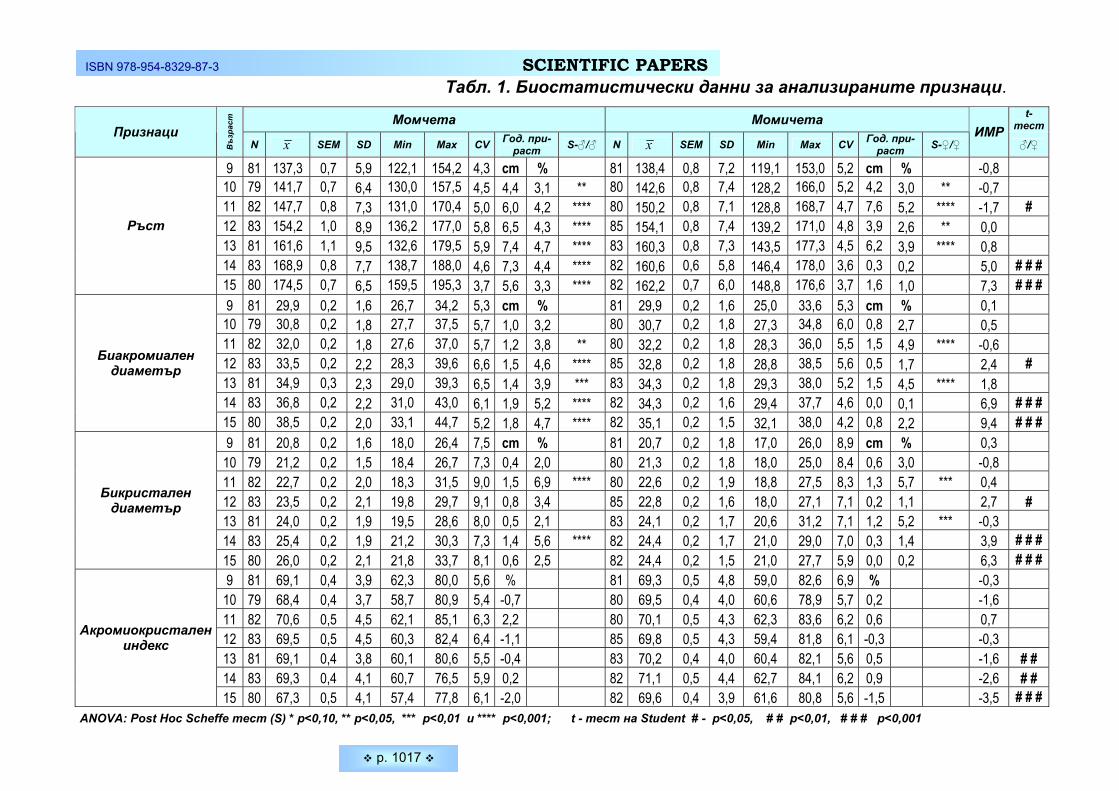
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1017
Табл. 1. Биостатистически данни за анализираните признаци.
Момчета Момичета t-тест Признаци
Възраст
N x SEM SD Min Max CV Год. при-раст S-♂/♂ N x SEM SD Min Max CV Год. при-
раст S-♀/♀ ИМР
♂/♀
9 81 137,3 0,7 5,9 122,1 154,2 4,3 cm % 81 138,4 0,8 7,2 119,1 153,0 5,2 cm % -0,8 10 79 141,7 0,7 6,4 130,0 157,5 4,5 4,4 3,1 ** 80 142,6 0,8 7,4 128,2 166,0 5,2 4,2 3,0 ** -0,7 11 82 147,7 0,8 7,3 131,0 170,4 5,0 6,0 4,2 **** 80 150,2 0,8 7,1 128,8 168,7 4,7 7,6 5,2 **** -1,7 # 12 83 154,2 1,0 8,9 136,2 177,0 5,8 6,5 4,3 **** 85 154,1 0,8 7,4 139,2 171,0 4,8 3,9 2,6 ** 0,0 13 81 161,6 1,1 9,5 132,6 179,5 5,9 7,4 4,7 **** 83 160,3 0,8 7,3 143,5 177,3 4,5 6,2 3,9 **** 0,8 14 83 168,9 0,8 7,7 138,7 188,0 4,6 7,3 4,4 **** 82 160,6 0,6 5,8 146,4 178,0 3,6 0,3 0,2 5,0 # # #
Ръст
15 80 174,5 0,7 6,5 159,5 195,3 3,7 5,6 3,3 **** 82 162,2 0,7 6,0 148,8 176,6 3,7 1,6 1,0 7,3 # # # 9 81 29,9 0,2 1,6 26,7 34,2 5,3 cm % 81 29,9 0,2 1,6 25,0 33,6 5,3 cm % 0,1
10 79 30,8 0,2 1,8 27,7 37,5 5,7 1,0 3,2 80 30,7 0,2 1,8 27,3 34,8 6,0 0,8 2,7 0,5 11 82 32,0 0,2 1,8 27,6 37,0 5,7 1,2 3,8 ** 80 32,2 0,2 1,8 28,3 36,0 5,5 1,5 4,9 **** -0,6 12 83 33,5 0,2 2,2 28,3 39,6 6,6 1,5 4,6 **** 85 32,8 0,2 1,8 28,8 38,5 5,6 0,5 1,7 2,4 # 13 81 34,9 0,3 2,3 29,0 39,3 6,5 1,4 3,9 *** 83 34,3 0,2 1,8 29,3 38,0 5,2 1,5 4,5 **** 1,8 14 83 36,8 0,2 2,2 31,0 43,0 6,1 1,9 5,2 **** 82 34,3 0,2 1,6 29,4 37,7 4,6 0,0 0,1 6,9 # # #
Биакромиален диаметър
15 80 38,5 0,2 2,0 33,1 44,7 5,2 1,8 4,7 **** 82 35,1 0,2 1,5 32,1 38,0 4,2 0,8 2,2 9,4 # # # 9 81 20,8 0,2 1,6 18,0 26,4 7,5 cm % 81 20,7 0,2 1,8 17,0 26,0 8,9 cm % 0,3
10 79 21,2 0,2 1,5 18,4 26,7 7,3 0,4 2,0 80 21,3 0,2 1,8 18,0 25,0 8,4 0,6 3,0 -0,8 11 82 22,7 0,2 2,0 18,3 31,5 9,0 1,5 6,9 **** 80 22,6 0,2 1,9 18,8 27,5 8,3 1,3 5,7 *** 0,4 12 83 23,5 0,2 2,1 19,8 29,7 9,1 0,8 3,4 85 22,8 0,2 1,6 18,0 27,1 7,1 0,2 1,1 2,7 # 13 81 24,0 0,2 1,9 19,5 28,6 8,0 0,5 2,1 83 24,1 0,2 1,7 20,6 31,2 7,1 1,2 5,2 *** -0,3 14 83 25,4 0,2 1,9 21,2 30,3 7,3 1,4 5,6 **** 82 24,4 0,2 1,7 21,0 29,0 7,0 0,3 1,4 3,9 # # #
Бикристален диаметър
15 80 26,0 0,2 2,1 21,8 33,7 8,1 0,6 2,5 82 24,4 0,2 1,5 21,0 27,7 5,9 0,0 0,2 6,3 # # # 9 81 69,1 0,4 3,9 62,3 80,0 5,6 % 81 69,3 0,5 4,8 59,0 82,6 6,9 % -0,3
10 79 68,4 0,4 3,7 58,7 80,9 5,4 -0,7 80 69,5 0,4 4,0 60,6 78,9 5,7 0,2 -1,6 11 82 70,6 0,5 4,5 62,1 85,1 6,3 2,2 80 70,1 0,5 4,3 62,3 83,6 6,2 0,6 0,7 12 83 69,5 0,5 4,5 60,3 82,4 6,4 -1,1 85 69,8 0,5 4,3 59,4 81,8 6,1 -0,3 -0,3 13 81 69,1 0,4 3,8 60,1 80,6 5,5 -0,4 83 70,2 0,4 4,0 60,4 82,1 5,6 0,5 -1,6 # # 14 83 69,3 0,4 4,1 60,7 76,5 5,9 0,2 82 71,1 0,5 4,4 62,7 84,1 6,2 0,9 -2,6 # #
Акромиокристален индекс
15 80 67,3 0,5 4,1 57,4 77,8 6,1 -2,0 82 69,6 0,4 3,9 61,6 80,8 5,6 -1,5 -3,5 # # # ANOVA: Post Hoc Scheffe тест (S) * p<0,10, ** p<0,05, *** p<0,01 и **** p<0,001; t - тест на Student # - p<0,05, # # p<0,01, # # # p<0,001
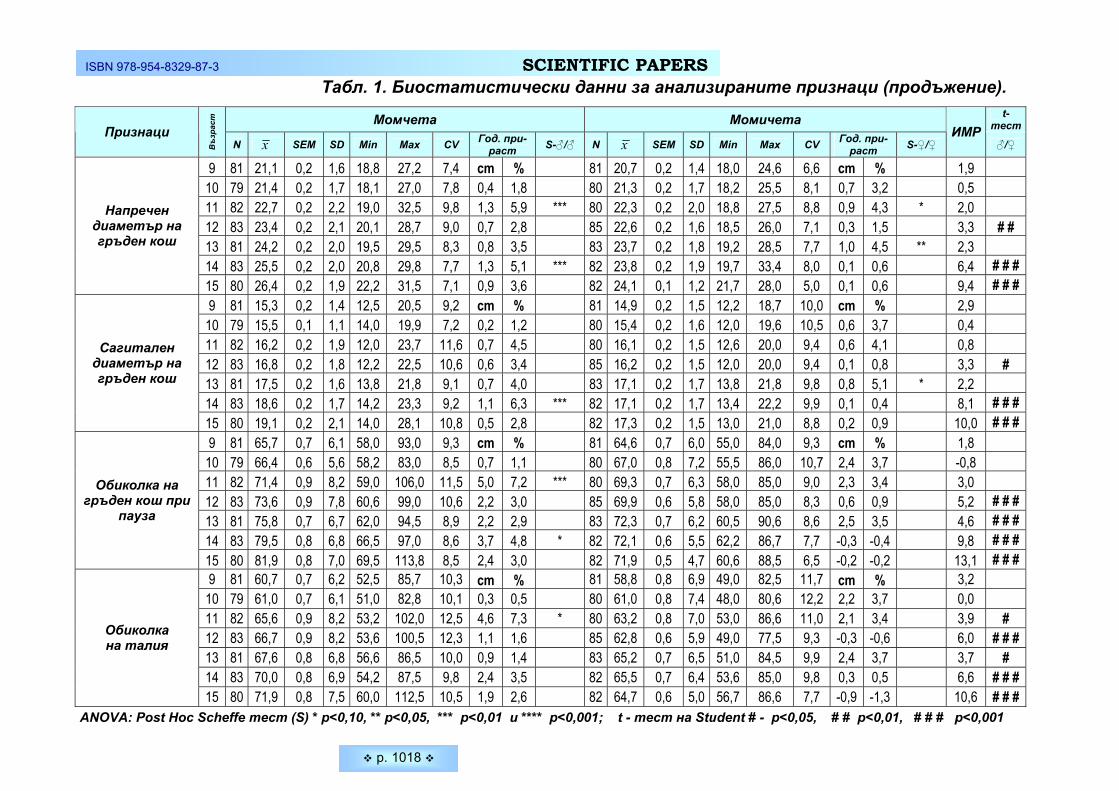
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1018
Табл. 1. Биостатистически данни за анализираните признаци (продъжение).
Момчета Момичета t-тест Признаци
Възраст
N x SEM SD Min Max CV Год. при-раст S-♂/♂ N x SEM SD Min Max CV Год. при-
раст S-♀/♀ ИМР
♂/♀
9 81 21,1 0,2 1,6 18,8 27,2 7,4 cm % 81 20,7 0,2 1,4 18,0 24,6 6,6 cm % 1,9 10 79 21,4 0,2 1,7 18,1 27,0 7,8 0,4 1,8 80 21,3 0,2 1,7 18,2 25,5 8,1 0,7 3,2 0,5 11 82 22,7 0,2 2,2 19,0 32,5 9,8 1,3 5,9 *** 80 22,3 0,2 2,0 18,8 27,5 8,8 0,9 4,3 * 2,0 12 83 23,4 0,2 2,1 20,1 28,7 9,0 0,7 2,8 85 22,6 0,2 1,6 18,5 26,0 7,1 0,3 1,5 3,3 # # 13 81 24,2 0,2 2,0 19,5 29,5 8,3 0,8 3,5 83 23,7 0,2 1,8 19,2 28,5 7,7 1,0 4,5 ** 2,3 14 83 25,5 0,2 2,0 20,8 29,8 7,7 1,3 5,1 *** 82 23,8 0,2 1,9 19,7 33,4 8,0 0,1 0,6 6,4 # # #
Напречен диаметър на гръдeн кош
15 80 26,4 0,2 1,9 22,2 31,5 7,1 0,9 3,6 82 24,1 0,1 1,2 21,7 28,0 5,0 0,1 0,6 9,4 # # # 9 81 15,3 0,2 1,4 12,5 20,5 9,2 cm % 81 14,9 0,2 1,5 12,2 18,7 10,0 cm % 2,9
10 79 15,5 0,1 1,1 14,0 19,9 7,2 0,2 1,2 80 15,4 0,2 1,6 12,0 19,6 10,5 0,6 3,7 0,4 11 82 16,2 0,2 1,9 12,0 23,7 11,6 0,7 4,5 80 16,1 0,2 1,5 12,6 20,0 9,4 0,6 4,1 0,8 12 83 16,8 0,2 1,8 12,2 22,5 10,6 0,6 3,4 85 16,2 0,2 1,5 12,0 20,0 9,4 0,1 0,8 3,3 # 13 81 17,5 0,2 1,6 13,8 21,8 9,1 0,7 4,0 83 17,1 0,2 1,7 13,8 21,8 9,8 0,8 5,1 * 2,2 14 83 18,6 0,2 1,7 14,2 23,3 9,2 1,1 6,3 *** 82 17,1 0,2 1,7 13,4 22,2 9,9 0,1 0,4 8,1 # # #
Сагитален диаметър на гръдeн кош
15 80 19,1 0,2 2,1 14,0 28,1 10,8 0,5 2,8 82 17,3 0,2 1,5 13,0 21,0 8,8 0,2 0,9 10,0 # # # 9 81 65,7 0,7 6,1 58,0 93,0 9,3 cm % 81 64,6 0,7 6,0 55,0 84,0 9,3 cm % 1,8
10 79 66,4 0,6 5,6 58,2 83,0 8,5 0,7 1,1 80 67,0 0,8 7,2 55,5 86,0 10,7 2,4 3,7 -0,8 11 82 71,4 0,9 8,2 59,0 106,0 11,5 5,0 7,2 *** 80 69,3 0,7 6,3 58,0 85,0 9,0 2,3 3,4 3,0 12 83 73,6 0,9 7,8 60,6 99,0 10,6 2,2 3,0 85 69,9 0,6 5,8 58,0 85,0 8,3 0,6 0,9 5,2 # # # 13 81 75,8 0,7 6,7 62,0 94,5 8,9 2,2 2,9 83 72,3 0,7 6,2 60,5 90,6 8,6 2,5 3,5 4,6 # # # 14 83 79,5 0,8 6,8 66,5 97,0 8,6 3,7 4,8 * 82 72,1 0,6 5,5 62,2 86,7 7,7 -0,3 -0,4 9,8 # # #
Обиколка на гръден кош при
пауза
15 80 81,9 0,8 7,0 69,5 113,8 8,5 2,4 3,0 82 71,9 0,5 4,7 60,6 88,5 6,5 -0,2 -0,2 13,1 # # # 9 81 60,7 0,7 6,2 52,5 85,7 10,3 cm % 81 58,8 0,8 6,9 49,0 82,5 11,7 cm % 3,2
10 79 61,0 0,7 6,1 51,0 82,8 10,1 0,3 0,5 80 61,0 0,8 7,4 48,0 80,6 12,2 2,2 3,7 0,0 11 82 65,6 0,9 8,2 53,2 102,0 12,5 4,6 7,3 * 80 63,2 0,8 7,0 53,0 86,6 11,0 2,1 3,4 3,9 # 12 83 66,7 0,9 8,2 53,6 100,5 12,3 1,1 1,6 85 62,8 0,6 5,9 49,0 77,5 9,3 -0,3 -0,6 6,0 # # # 13 81 67,6 0,8 6,8 56,6 86,5 10,0 0,9 1,4 83 65,2 0,7 6,5 51,0 84,5 9,9 2,4 3,7 3,7 # 14 83 70,0 0,8 6,9 54,2 87,5 9,8 2,4 3,5 82 65,5 0,7 6,4 53,6 85,0 9,8 0,3 0,5 6,6 # # #
Обиколка на талия
15 80 71,9 0,8 7,5 60,0 112,5 10,5 1,9 2,6 82 64,7 0,6 5,0 56,7 86,6 7,7 -0,9 -1,3 10,6 # # # ANOVA: Post Hoc Scheffe тест (S) * p<0,10, ** p<0,05, *** p<0,01 и **** p<0,001; t - тест на Student # - p<0,05, # # p<0,01, # # # p<0,001
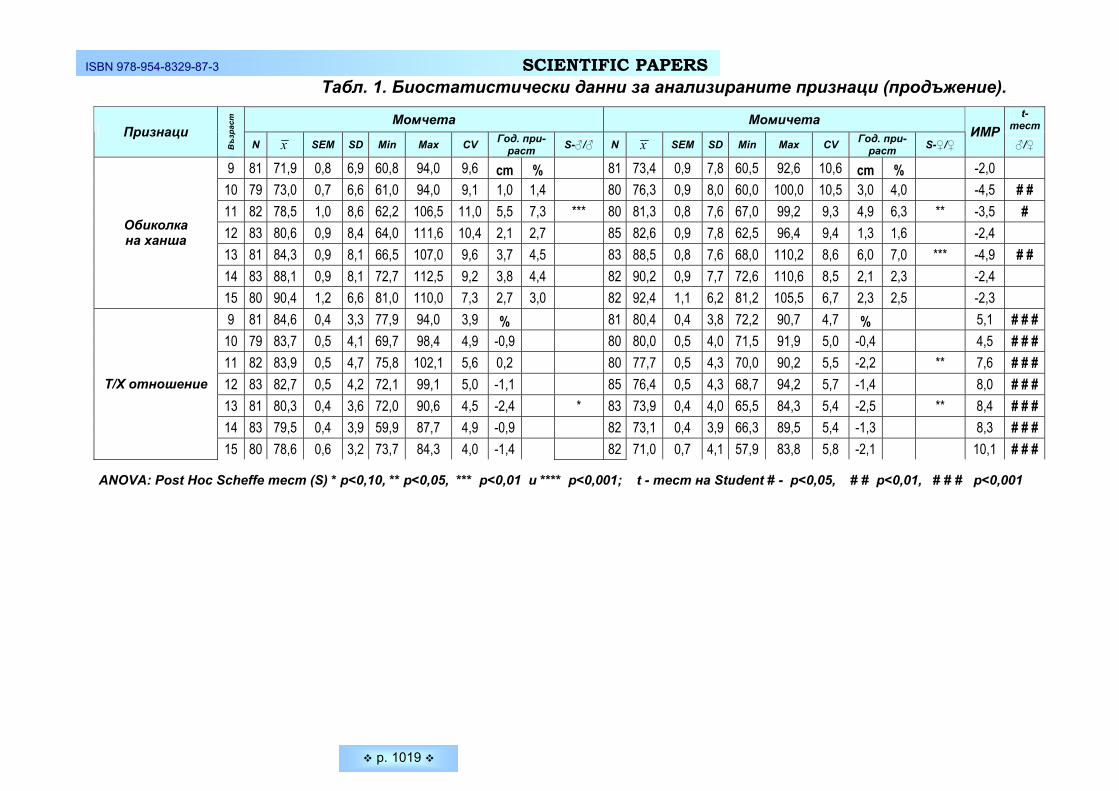
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1019
Табл. 1. Биостатистически данни за анализираните признаци (продъжение).
Момчета Момичета t-тест Признаци
Възраст
N x SEM SD Min Max CV Год. при-раст S-♂/♂ N x SEM SD Min Max CV Год. при-
раст S-♀/♀ ИМР
♂/♀
9 81 71,9 0,8 6,9 60,8 94,0 9,6 cm % 81 73,4 0,9 7,8 60,5 92,6 10,6 cm % -2,0 10 79 73,0 0,7 6,6 61,0 94,0 9,1 1,0 1,4 80 76,3 0,9 8,0 60,0 100,0 10,5 3,0 4,0 -4,5 # # 11 82 78,5 1,0 8,6 62,2 106,5 11,0 5,5 7,3 *** 80 81,3 0,8 7,6 67,0 99,2 9,3 4,9 6,3 ** -3,5 # 12 83 80,6 0,9 8,4 64,0 111,6 10,4 2,1 2,7 85 82,6 0,9 7,8 62,5 96,4 9,4 1,3 1,6 -2,4 13 81 84,3 0,9 8,1 66,5 107,0 9,6 3,7 4,5 83 88,5 0,8 7,6 68,0 110,2 8,6 6,0 7,0 *** -4,9 # # 14 83 88,1 0,9 8,1 72,7 112,5 9,2 3,8 4,4 82 90,2 0,9 7,7 72,6 110,6 8,5 2,1 2,3 -2,4
Обиколка на ханша
15 80 90,4 1,2 6,6 81,0 110,0 7,3 2,7 3,0 82 92,4 1,1 6,2 81,2 105,5 6,7 2,3 2,5 -2,3 9 81 84,6 0,4 3,3 77,9 94,0 3,9 % 81 80,4 0,4 3,8 72,2 90,7 4,7 % 5,1 # # #
10 79 83,7 0,5 4,1 69,7 98,4 4,9 -0,9 80 80,0 0,5 4,0 71,5 91,9 5,0 -0,4 4,5 # # # 11 82 83,9 0,5 4,7 75,8 102,1 5,6 0,2 80 77,7 0,5 4,3 70,0 90,2 5,5 -2,2 ** 7,6 # # # 12 83 82,7 0,5 4,2 72,1 99,1 5,0 -1,1 85 76,4 0,5 4,3 68,7 94,2 5,7 -1,4 8,0 # # # 13 81 80,3 0,4 3,6 72,0 90,6 4,5 -2,4 * 83 73,9 0,4 4,0 65,5 84,3 5,4 -2,5 ** 8,4 # # # 14 83 79,5 0,4 3,9 59,9 87,7 4,9 -0,9 82 73,1 0,4 3,9 66,3 89,5 5,4 -1,3 8,3 # # #
Т/Х отношение
15 80 78,6 0,6 3,2 73,7 84,3 4,0 -1,4 82 71,0 0,7 4,1 57,9 83,8 5,8 -2,1 10,1 # # # ANOVA: Post Hoc Scheffe тест (S) * p<0,10, ** p<0,05, *** p<0,01 и **** p<0,001; t - тест на Student # - p<0,05, # # p<0,01, # # # p<0,001
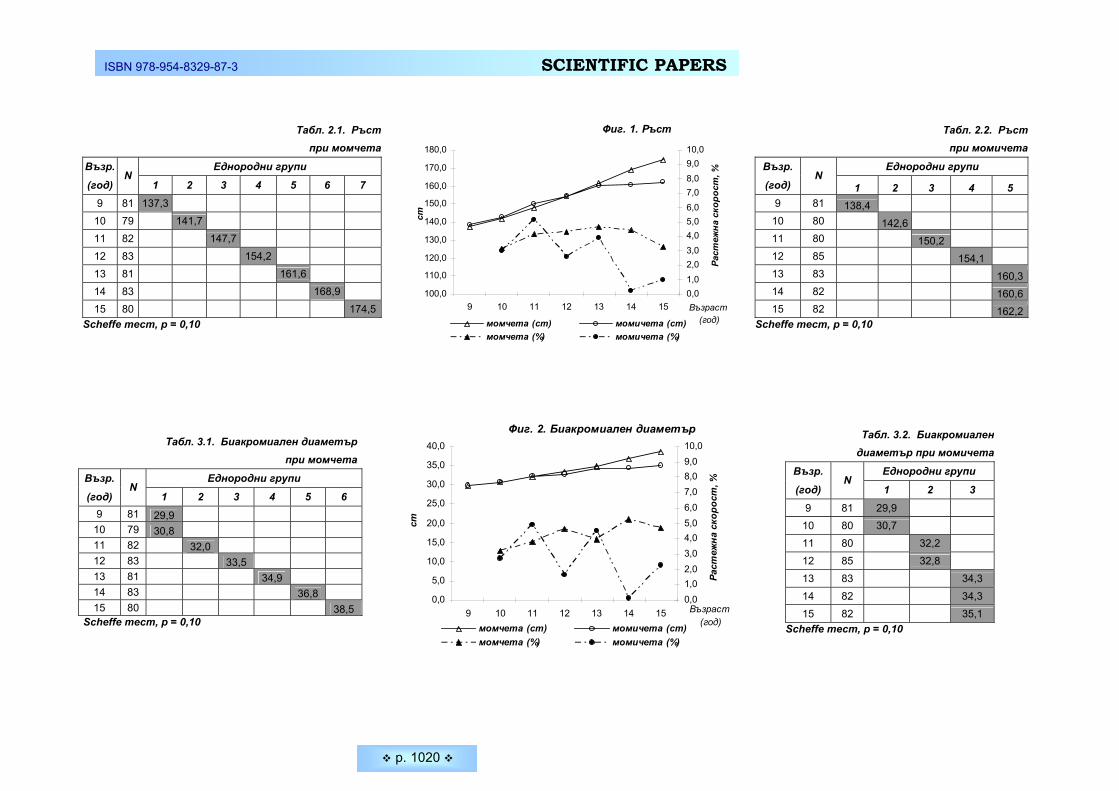
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1020
Табл. 2.1. Ръстпри момчета
Възр. Еднородни групи
(год) N
1 2 3 4 5 6 7 9 81 137,3
10 79 141,7 11 82 147,7 12 83 154,2 13 81 161,6 14 83 168,9 15 80 174,5
Scheffe тест, p = 0,10
Фиг. 1. Ръст
100,0
110,0
120,0
130,0
140,0
150,0
160,0
170,0
180,0
9 10 11 12 13 14 15 Възраст (год)
cm
0,01,02,03,04,05,06,07,08,09,010,0
момчета (cm) момичета (cm)момчета (%) момичета (%)
Раст
ежна
ско
рост
, %
Табл. 2.2. Ръстпри момичета
Възр. Еднородни групи
(год) N
1 2 3 4 5 9 81 138,4
10 80 142,6 11 80 150,2 12 85 154,1 13 83 160,3 14 82 160,6 15 82 162,2
Scheffe тест, p = 0,10
Табл. 3.1. Биакромиален диаметърпри момчета
Възр. Еднородни групи
(год) N
1 2 3 4 5 6 9 81 29,9
10 79 30,8 11 82 32,0 12 83 33,5 13 81 34,9 14 83 36,8 15 80 38,5
Scheffe тест, p = 0,10
Фиг. 2. Биакромиален диаметър
0,0
5,0
10,0
15,0
20,0
25,0
30,0
35,0
40,0
9 10 11 12 13 14 15 Възраст (год)
cm
0,01,02,03,04,05,06,07,08,09,010,0
момчета (cm) момичета (cm)момчета (%) момичета (%)
Раст
ежна
ско
рост
, %
Табл. 3.2. Биакромиалендиаметър при момичета
Възр. Еднородни групи
(год) N
1 2 3 9 81 29,9
10 80 30,7 11 80 32,2 12 85 32,8 13 83 34,3 14 82 34,3 15 82 35,1
Scheffe тест, p = 0,10
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1021
Табл. 4.1. Бикристален диаметърпри момчета
Възр. Еднородни групи
(год) N
1 2 3 4
9 81 20,8
10 79 21,2 11 82 22,7 12 83 23,5 23,5 13 81 24,0 14 83 25,4 15 80 26,0
Scheffe тест, p = 0,10
Фиг. 3. Бикристален диаметър
0,0
5,0
10,0
15,0
20,0
25,0
30,0
9 10 11 12 13 14 15 Възраст (год)
cm
0,01,02,03,04,05,06,07,08,09,010,0
момчета (cm) момичета (cm)момчета (%) момичета (%)
Раст
ежна
ско
рост
, %
Табл. 4.2. Бикристален диаметърпри момичета
Възр. Еднородни г рупи
(год) N
1 2 3
9 81 20,7
10 80 21,3 11 80 22,6 12 85 22,8 13 83 24,1 14 82 24,4 15 82 24,4
Scheffe тест, p = 0,10
Табл. 5.1. Напречен диаметърна гръден кош при момчета
Възр. Еднородни групи
(год) N
1 2 3 4 9 81 21,1
10 79 21,4 11 82 22,7 12 83 23,4 23,4 13 81 24,2 14 83 25,5 15 80 26,4
Scheffe тест, p = 0,10
Фиг. 4. Напречен диаметър на гръдния кош
0,0
5,0
10,0
15,0
20,0
25,0
30,0
9 10 11 12 13 14 15 Възраст (год)
cm
0,01,02,03,04,05,06,07,08,09,010,0
момчета (cm) момичета (cm)
момчета (%) момичета (%)
Раст
ежна
ско
рост
, %
Табл. 5.2. Напречен диаметър на гръден кош при момичета
Възр. Еднородни групи
(год) N
1 2 3 9 81 20,7
10 80 21,3 11 80 22,3 12 85 22,6 13 83 23,7 14 82 23,8 15 82 24,1
Scheffe тест, p = 0,10
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1022
Табл. 6.1. Сагитален диаметър на гръден кошпри момчета
Възр. Еднородни групи
(год) N
1 2 3 4 5 9 81 15,3
10 78 15,5 15,5 11 82 16,2 16,2 12 83 16,8 16,8 13 81 17,5 14 83 18,6
15 80 19,1
Scheffe тест, p = 0,10
Фиг. 5. Сагитален диаметър на гръдния кош
0,02,04,06,08,0
10,012,014,016,018,020,0
9 10 11 12 13 14 15 Възраст (год)
cm
0,01,02,03,04,05,06,07,08,09,010,0
момчета (cm) момичета (cm)момчета (%) момичета (%)
Раст
ежна
ско
рост
, %
Табл. 6.2. Сагитален диаметър на гръден кош при момичета
Възр. Еднородни групи
(год) N
1 2 3 9 81 14,9
10 80 15,4 15,4 11 80 16,1 12 85 16,2 13 83 17,1 14 82 17,1
15 82 17,3
Scheffe тест, p = 0,10
Табл. 7.1. Гръдна обиколка при паузапри момчета
Възр. Еднородни групи
(год) N
1 2 3 4 9 81 65,7
10 79 66,4 11 82 71,4 12 83 73,6 73,6 13 81 75,8 14 83 79,5 15 80 81,9
Scheffe тест, p = 0,10
Фиг. 6. Гръдна обиколка-пауза
-5,0
5,0
15,0
25,0
35,0
45,0
55,0
65,0
75,0
85,0
9 10 11 12 13 14 15 Възраст (год)
cm
-1,00,01,02,03,04,05,06,07,08,09,010,0
момчета (cm) момичета (cm)момчета (%) момичета (%)
Раст
ежна
скорост
, %
Табл. 7.2. Гръдна обиколка при пауза при момичета
Възр. Еднородни групи
(год) N
1 2 3 9 81 64,6
10 80 67,0 67,0 11 80 69,3 69,3 12 85 69,9 69,9 15 82 71,9 14 82 72,1 13 83 72,3
Scheffe тест, p = 0,10
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1023
Табл. 8.1. Обиколка на талияпри момчета
Възр. Еднородни групи
(год) N
1 2 3 4 9 81 60,7
10 79 61,0 11 82 65,6 12 83 66,7 66,7 13 81 67,6 67,6 14 83 70,0 70,0 15 80 71,9
Scheffe тест, p = 0,10
Фиг. 7. Обиколка на талия
-5,0
5,0
15,0
25,0
35,0
45,0
55,0
65,0
75,0
9 10 11 12 13 14 15 Възраст (год)
cm
-2,0-1,00,01,02,03,04,05,06,07,08,09,010,0
момчета (cm) момичета (cm)момчета (%) момичета (%)
Раст
ежна
ско
рост
, %
Табл. 8.2. Обиколка на талияпри момичета
Възр. Еднородни групи
(год) N
1 2 3 9 81 58,8
10 80 61,0 61,0 12 85 62,8 62,8 11 80 63,2 63,2 15 82 64,7 13 83 65,2 14 82 65,5
Scheffe тест, p = 0,10
Табл. 9.1. Обиколка на ханшпри момчета
Възр. Еднородни групи
(год) N
1 2 3 4 5 9 79 71,9
10 79 73,0 11 75 78,5 12 83 80,6 80,6 13 81 84,3 84,3 14 78 87,7 87,7
15 29 90,4
Scheffe тест, p = 0,10
Фиг. 8. Обиколка на ханш
0,010,020,030,040,050,060,070,080,090,0
100,0
9 10 11 12 13 14 15 Възраст (год)
cm
0,01,02,03,04,05,06,07,08,09,010,0
момчета (cm) момичета (cm)момчета (%) момичета (%)
Раст
ежна
скорост
, %
Табл. 9.2. Обиколка на ханш при момичета
Възр. Еднородни групи
(год) N
1 2 3
9 77 73,4 10 79 76,3 11 80 81,3 12 83 82,6 13 81 88,1 14 77 90,2 15 31 92,4
Scheffe тест, p = 0,10
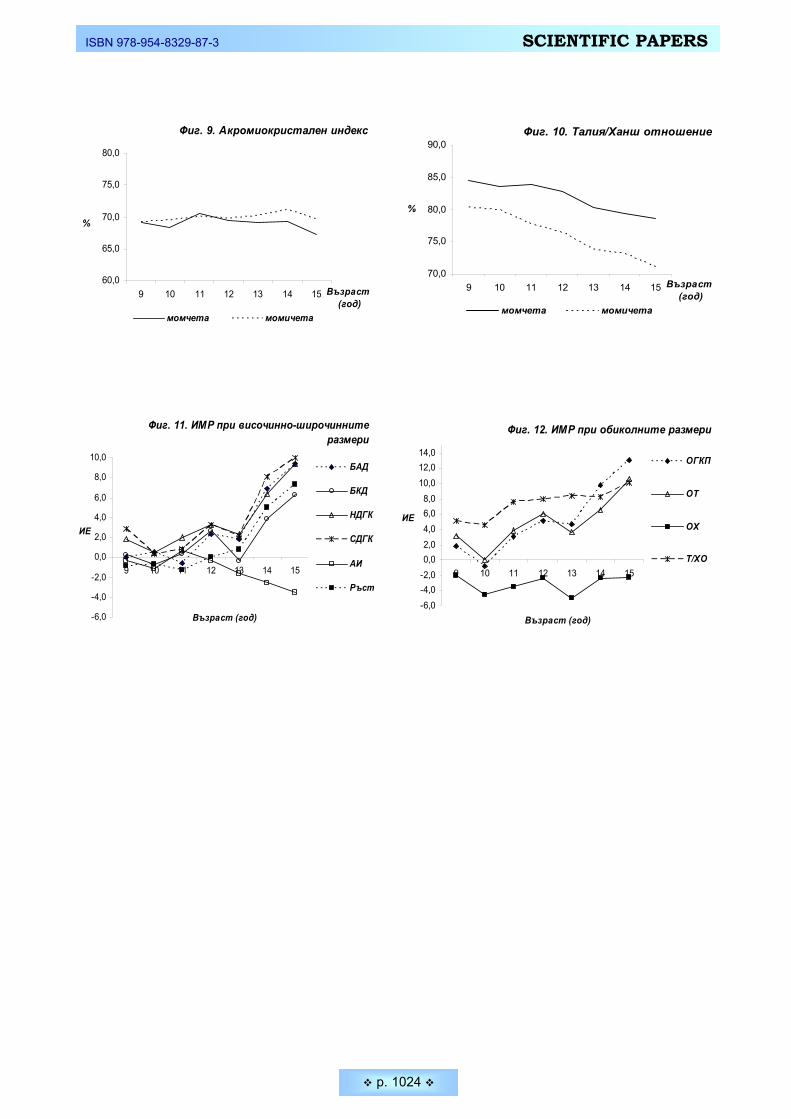
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1024
Фиг. 9. Акромиокристален индекс
60,0
65,0
70,0
75,0
80,0
9 10 11 12 13 14 15 Възраст (год)
момчета момичета
%
Фиг. 10. Талия/Ханш отношение
70,0
75,0
80,0
85,0
90,0
9 10 11 12 13 14 15 Възраст (год)
момчета момичета
%
Фиг. 11. ИМР при височинно-широчинните размери
-6,0
-4,0
-2,0
0,0
2,0
4,0
6,0
8,0
10,0
9 10 11 12 13 14 15
Възраст (год)
ИЕ
БАД
БКД
НДГК
СДГК
АИ
Ръст
Фиг. 12. ИМР при обиколните размери
-6,0-4,0-2,00,02,04,06,08,0
10,012,014,0
9 10 11 12 13 14 15
Възраст (год)
ОГКП
ОТ
ОХ
Т/ХО
ИЕ

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 1025
SCIENTIFIC PAPERS
РАЗПРОСТРАНЕНИЕ НА СВРЪХТЕГЛОТО И ЗАТЛЪСТЯВАНЕТО СРЕД СМОЛЯНСКИТЕ ДЕЦА
И ПОДРАСТВАЩИ
С. Младенова*, М. Николова** *Филиал на ПУ ”Паисий Хилендарски”- гр. Смолян
**ПУ ”Паисий Хилендарски”, Биологически факултет
DISTRIBUTION OF THE OVERWEIGHT AND OBESITY AMONG SMOLYAN CHILDREN AND ADOLESCENTS
S.Mladenova*, M. Nikolova**
Branch Smolyan at Plovdiv University “Paisii Hilendarski”; Plovdiv University “Paisii Hilendarski”, Faculty if Biology,
Depatment of Human anatomy and physiology
ABSTRACT: Overweight and obesity reached epidemic proportions globally and be-
come the main public health problem. The prevalence of child obesity is increasing rapidly worldwide. It is associated with several risk factors for later heart diseases, orther chronic diseases, mortality and other. Because of their public health importance the trends of child overweight and obesity should be closely monitored. The most commonly used method for classifying an individual as overweight or obese of adult and children is based on body mass index (BMI, kg/m2). For definition of child overweight and obesity often used differ-ent age- and sex- growth curves or intrenational age- and sex specific cut-off point for BMI from 2-18 years, passed through the widely used cut off points of 25 and 30 kg/m2 for adult overweight and obesity, published by 2000 year of Cole et al.
The aim of this investigation is to determine the distribution of the overweight and obesity among children and adolescents from Smolyan region. The sample include 1589 chlidren ( 809 girls and 780 boys), aged 3-17, measured in period 1998-2001. By Martin-Saller's method two basic size features are measured -height and weight. BMI are calcu-late using the formula The overweight and obese on each age and sex group are define using international cut -off points of BMI, create of Cole et al (2000).
The results are shows that the prevalence of the overweight is different by age and sex, and it is often prevalence in the girls, than in the boys. Especially high percent girls with overweight are shows among 8- (25.49%) and 15 years old (11.74%), while the high-est percent overweight boys observed among sixteen years (18.86%). The highest percent obese boys are observe in group of 14 years old (11.32%), while more obese girls are ob-serve among 6 and 8 years old (5.30%).
The prevalence of obesity is often observed in group of the boys, compared with group of the girls, especially after 12 th years.
The our results also corroborated with our and literature data of the greater sensitivity of child organism on the different factors, more pronounced of the puberty period and among n boys group.
Key words: children, overweight, obesity, BMI

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1026
ВЪВЕДЕНИЕ: Затлъстяването е болестта на съвременната цивилизация. Тя има епи-
демичен характер, като по данни на СЗО, в световен мащаб над 1 милиард възрастни са със свръхтегло, а най-малко 300 милиона от тях са със затлъс-тяване. Затлъстяването е комплексно състояние със сериозни социални и психологични измерения, обхващащо всички възрастови и социално-икономически групи, бързо разпространяващо се в развитите и развиващите се страни. То е резултат от нарушеното равновесие между енергоприхода и енергоразхода на организма, вследствие прекомерната употреба на високо-калорична храна, съдържаща много животински мазнини и захари, и намале-ната физическа активност. Затлъстяването е на второ място след тютюнопу-шенето като причина за преждевременна смърт и рисков фактор за развитие на сърдечно-съдови заболявания, неинсулинозависим захарен диабет (тип 2), злокачествени и респираторни заболявания, психосоциални проблеми и др [8; 9; 5 and other].
В световен мащаб най-разпространените методи за мониторинг на физи-ческото развитие и оценка на хранителния статус на индивида се базират на използването на различни антропометрични показатели, между които основно индекса за телесна маса (BMI, kg/m2). Този индекс, който практически не зави-си от ръста се използва масово за диагностика на недохранването и затлъс-тяването, както при деца, така и при израснали [14; 7; 13; 4; 2], а така също и за вътрегрупови и междугрупови съпоставки [3, 12]. В литературата обаче се срещат и много критики и препоръки [15] по отношение използването на ин-декса на Quetelet, наречен през 1972 г. от Keys и кол. BMI [6] за диагностика на свръхтеглото и затлъстяването.
Проблемът със затлъстяването в детска възраст има важна здравна, об-ществена и социална значимост, тъй като е свързан с редица сериозни забо-лявания в детска възраст, а така също и с профилактика на редица сърдечно-съдови, метаболитни и други смущения в по-късна възраст.
На него са посветени много разработки [16; 8; 12; 1;11 и др.]. По-голямата част от авторите дефинират свръхтеглото и затлъстяването сред децата с помощта на различни популационни възрастово- и полово-специфични пер-сентили за BMI, а някои и чрез международните “cut-off” точки за BMI, разра-ботени от Cole и кол. през 2000г., и препоръчвани от СЗО за дефиниране на свръхтеглото и затлъстяването при деца от 2 до 18 години.
Значимостта на проблема за затлъстяването сред децата и подраства-щите, както и необходимостта от съвременна база данни за сравнение с други популации, определиха ЦЕЛТА на настоящата разработка, а именно да проу-чим разпространението на свръхтеглото и затлъстяването сред смолянските деца и подрастващи.
ИЗСЛЕДВАН КОНТИНГЕНТ: 1589 деца и подрастващи от Смолянски регион, от които 809 момичета
и 780 момчета на възраст от 3 до 17 години
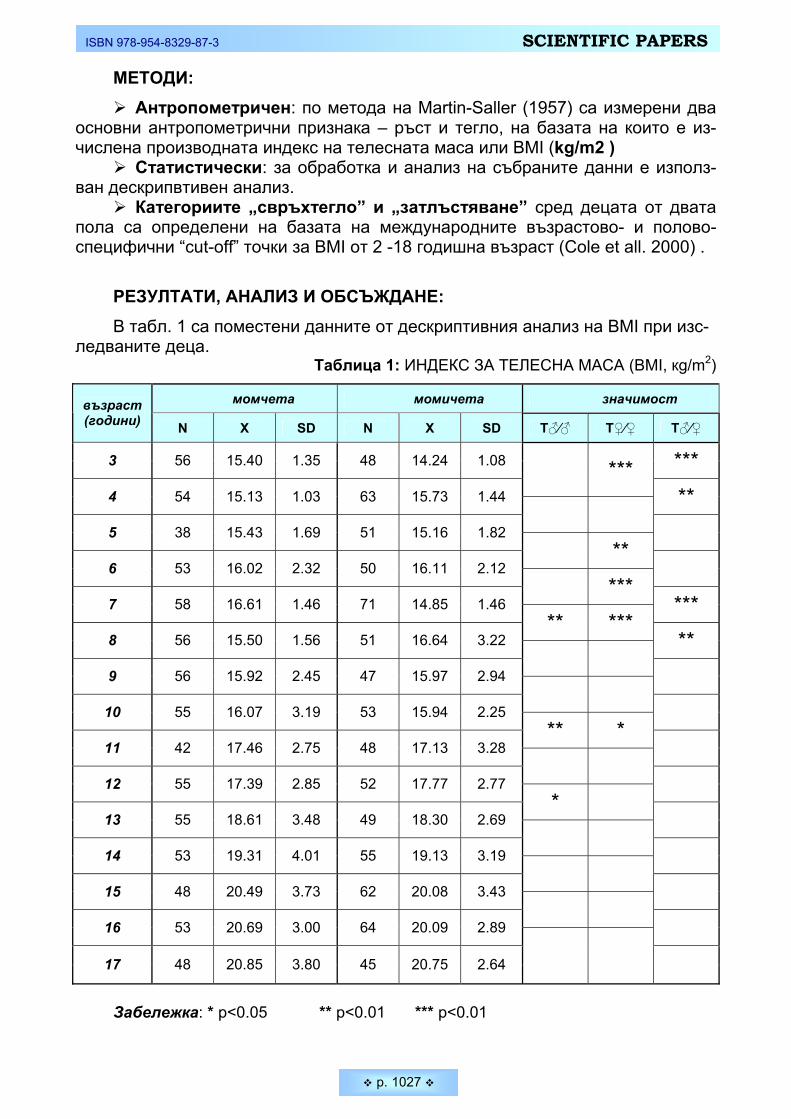
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1027
МЕТОДИ: Антропометричен: по метода на Martin-Saller (1957) са измерени два
основни антропометрични признака – ръст и тегло, на базата на които е из-числена производната индекс на телесната маса или BMI (kg/m2 )
Статистически: за обработка и анализ на събраните данни е използ-ван дескрипвтивен анализ.
Категориите „свръхтегло” и „затлъстяване” сред децата от двата пола са определени на базата на международните възрастово- и полово-специфични “cut-off” точки за BMI от 2 -18 годишна възраст (Cole et all. 2000) .
РЕЗУЛТАТИ, АНАЛИЗ И ОБСЪЖДАНЕ: В табл. 1 са поместени данните от дескриптивния анализ на BMI при изс-
ледваните деца. Таблица 1: ИНДЕКС ЗА ТЕЛЕСНА МАСА (BMI, кg/m2)
момчета момичета значимост възраст (години) N X SD N X SD T♂⁄♂ T♀⁄♀ T♂⁄♀
3 56 15.40 1.35 48 14.24 1.08 *** *** 4 54 15.13 1.03 63 15.73 1.44 **
5 38 15.43 1.69 51 15.16 1.82
** 6 53 16.02 2.32 50 16.11 2.12
*** 7 58 16.61 1.46 71 14.85 1.46 ***
** *** 8 56 15.50 1.56 51 16.64 3.22 **
9 56 15.92 2.45 47 15.97 2.94
10 55 16.07 3.19 53 15.94 2.25
** * 11 42 17.46 2.75 48 17.13 3.28
12 55 17.39 2.85 52 17.77 2.77
* 13 55 18.61 3.48 49 18.30 2.69
14 53 19.31 4.01 55 19.13 3.19
15 48 20.49 3.73 62 20.08 3.43
16 53 20.69 3.00 64 20.09 2.89
17 48 20.85 3.80 45 20.75 2.64
Забележка: * p<0.05 ** p<0.01 *** p<0.01
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1028
Анализът на резултатите, поместени в табл. 1 показва, че стойностите на BMI и при двата пола нарастват през целия растежен период, макар и с из-вестни флуктуации.
При момчетата сигнификантно по-голямо натрупване на kg телесна маса на единица ръст се наблюдава между 10-11-ата година-1.39 kg/m2, и 12-13-ата година -1.22 kg/m2 (p<0.05). Значимо намаляване на стойностите на изследва-ния индекс (1.11 kg/m2) при тях се наблюдава между 7-8-ата година, т.е. в пери-ода, когато те увеличават значимо телесната си височина (p<0.01).
При момичетата значимо натрупване на kg телесна маса на единица телесна височина се наблюдават в началото и средата на изследвания растежен период, а именно: между 3-4-ата година, когато увеличаването на телесната маса е с 1.49 kg/m2 (p<0.001), между 5- 6-ата година е 0.95 kg/m2, 7-8-ата – 1.79 kg/m2 и 10-11-ата година съответно 1.19 kg/m2. Но наред с пери-одите, в които се наблюдава прогресивно покачване на стойностите на ин-декса и при момичетата се наблюдават периоди, в които стойностите нама-ляват – между 4-5-ата година (p>0.05), 6-7-ата година (p<0.001), и 8-9-ата годи-на (p>0.05).
Анализът на междуполовите различия в стойностите на BMI (фиг.1) пока-за, че през по-голямата част от наблюдавания възрастов период представи-телите на мъжкия пол имат незначимо по–високи абсолютни стойности на по-казателя, с изключение на 4, 8, 9 и 12-годишните момичета.
5
7
9
11
13
15
17
19
21
BM
I (kg
/m^2
)
3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
възраст ( години)
фиг.1: Индекс за телесна маса (kg/m^2) при децата от Смолянски регион
момчета
момичета
Сигнификантно по-голямо количество телесна маса на единица телесна височина обаче се наблюдава само при 3, 7 и 8-годишните момчета и при 4 годишните момичета, в сравнение с тези на връстниците им от противопо-ложния пол.
Половите различия в стойностите на индекса са свързани с периодите на “окръгляне” и “източване” при децата от двата пола, както и с разликата във времето на настъпване на пубертетносвързаните трансформации в телесната им конституция. Нарастването на стойностите на индекса до края на наблю-давания период и при двата пола показва, че оформянето на терминалният им хранителен статус вероятно още не е завършило.
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1029
Фигури 2 и 3 отразяват особеностите в хранителния статус на децата от двата пола.
Относителният дял свръхтежки момчета (фиг. 2) варира в границите от 0 % при 14 годишните до 18.86 % при 16-годишните. Висок е относителен дял на свръхтежките момчета и сред 6, 11 и 15-годишните. Варира и относителния дял на затлъстелите момчета (фиг. 2). Във възрастовите групи на 3, 4 и 8 го-дишните момчета липсват деца със затлъстяване. Техният относителен дял се покачва от 1.81 % при 12 и 13-годишните до 11.32 % при 14-годишните момчета, където се наблюдава най-високият процент затлъстели.
фиг.2: Хранителен статус на момчетата
0%10%20%30%40%50%60%70%80%90%
100%
3 4 5 6 7 8 9 10 11 12 13 14 15 16 17възраст (години)
разпределени
е по
категории
(относителен
дял
)
свръхтегло затлъстяване без свръхтегло и затлъст.
При представителките на противоположния пол процентът свръхтежки
момичета варира от 0 % до 25.49 % (фиг. 3). Най-нисък е делът им във въз-растовата група на 7-годишните (2.81 %), а най-висок е при 8-годишните 25.49 %, следван от 9,15, 3, 6 и 17-годишните.
Относителният дял затлъстели момичета (фиг.3) сред изследваните въз-растови групи е нисък – на практика такива има само сред 6, 8, 9, 11, 12 и 14-годишните, като най-висок е техният дял при 6 и 8 годишните.
Анализът на междуполовите различия по отношение разпространението на свръхтеглото (фиг. 4) показа, че то е по-често срещано сред момичетата, и то през почти целия изследван възрастов период. Изключение правят 3, 7, 10, 11 и 16-годишните момчета.
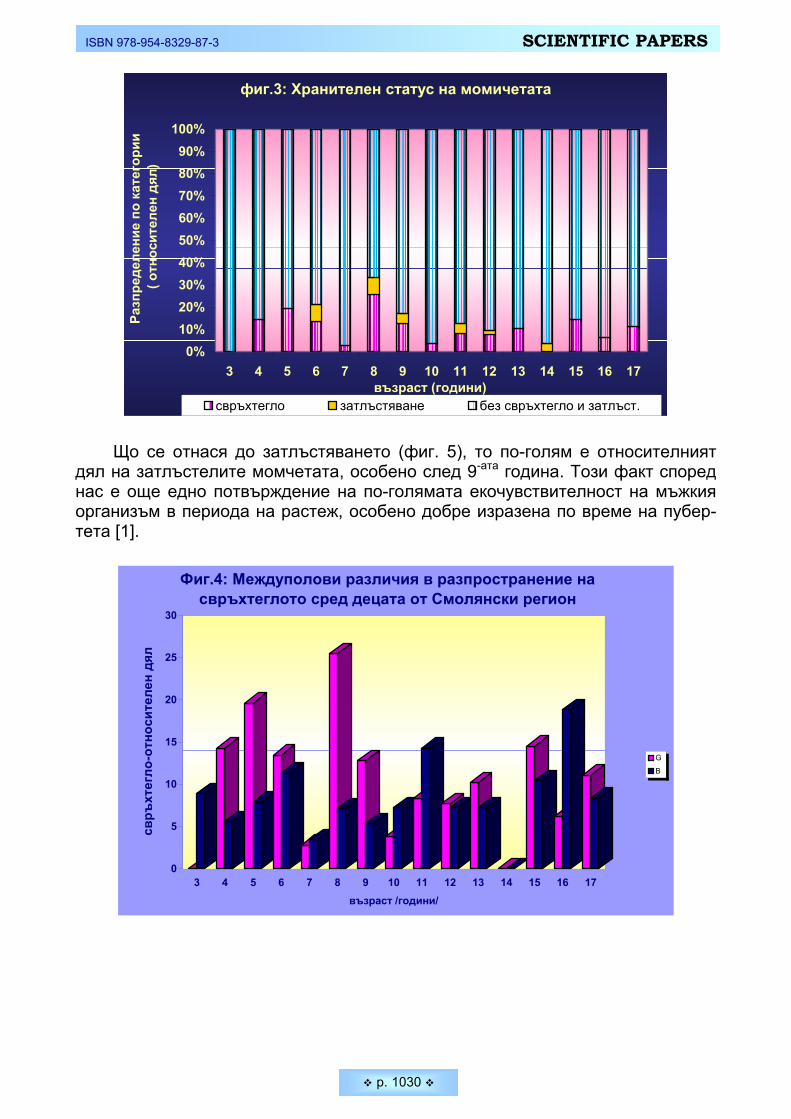
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1030
фиг.3: Хранителен статус на момичетата
0%10%20%30%40%50%60%70%80%90%
100%
3 4 5 6 7 8 9 10 11 12 13 14 15 16 17възраст (години)
Разпре
деле
ние по
категор
ии ( отно
сителе
н дя
л)
свръхтегло затлъстяване без свръхтегло и затлъст.
Що се отнася до затлъстяването (фиг. 5), то по-голям е относителният
дял на затлъстелите момчетата, особено след 9-ата година. Този факт според нас е още едно потвърждение на по-голямата екочувствителност на мъжкия организъм в периода на растеж, особено добре изразена по време на пубер-тета [1].
0
5
10
15
20
25
30
свръ
хтегло
-относ
ител
ен дял
3 4 5 6 7 8 9 10 11 12 13 14 15 16 17възраст /години/
Фиг.4: Междуполови различия в разпространение на свръхтеглото сред децата от Смолянски регион
G
B
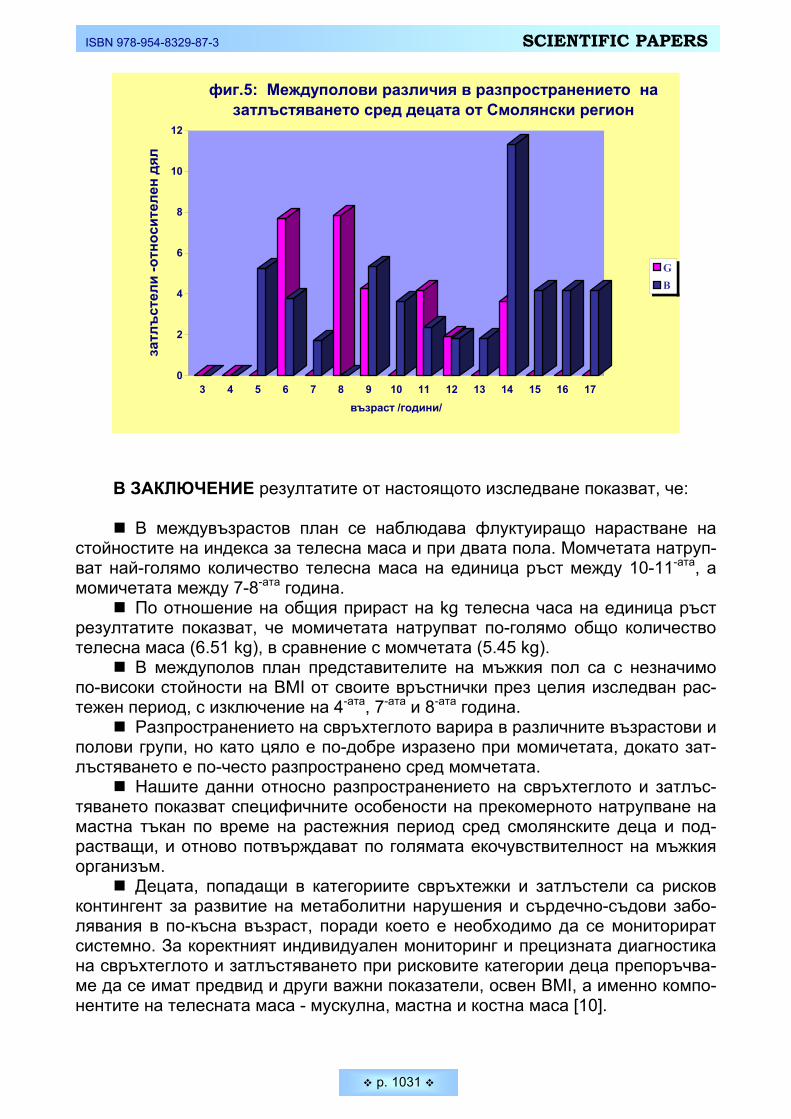
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1031
0
2
4
6
8
10
12
затлъстел
и -относител
ен дял
3 4 5 6 7 8 9 10 11 12 13 14 15 16 17възраст /години/
фиг.5: Междуполови различия в разпространението на затлъстяването сред децата от Смолянски регион
GB
В ЗАКЛЮЧЕНИЕ резултатите от настоящото изследване показват, че:
В междувъзрастов план се наблюдава флуктуиращо нарастване на стойностите на индекса за телесна маса и при двата пола. Момчетата натруп-ват най-голямо количество телесна маса на единица ръст между 10-11-ата, а момичетата между 7-8-ата година.
По отношение на общия прираст на kg телесна часа на единица ръст резултатите показват, че момичетата натрупват по-голямо общо количество телесна маса (6.51 kg), в сравнение с момчетата (5.45 kg).
В междуполов план представителите на мъжкия пол са с незначимо по-високи стойности на BMI от своите връстнички през целия изследван рас-тежен период, с изключение на 4-ата, 7-ата и 8-ата година.
Разпространението на свръхтеглото варира в различните възрастови и полови групи, но като цяло е по-добре изразено при момичетата, докато зат-лъстяването е по-често разпространено сред момчетата.
Нашите данни относно разпространението на свръхтеглото и затлъс-тяването показват специфичните особености на прекомерното натрупване на мастна тъкан по време на растежния период сред смолянските деца и под-растващи, и отново потвърждават по голямата екочувствителност на мъжкия организъм.
Децата, попадащи в категориите свръхтежки и затлъстели са рисков контингент за развитие на метаболитни нарушения и сърдечно-съдови забо-лявания в по-късна възраст, поради което е необходимо да се мониторират системно. За коректният индивидуален мониторинг и прецизната диагностика на свръхтеглото и затлъстяването при рисковите категории деца препоръчва-ме да се имат предвид и други важни показатели, освен BMI, а именно компо-нентите на телесната маса - мускулна, мастна и костна маса [10].

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1032
ЛИТЕРАТУРА: [1] Младенова, С., 2003. Антропологична характеристика на растежа и раз-
витието на децата и подрастващите от Смолянски регион в съвременните условия на живот., Дисертационен труд., Пловдив.
[2] Armstrong, J., 2003. The prevalence of obesity and undernutrition in Scotish Children: growth monitoring within the Child Health Sureillance Programme, Scotish Medical Journal, 48,2, 32-37
[3] Cole T. Y., Roede M .Y., 1999. Centiles of body mass index for Dutch children aged 0-20 years in 1980 a baseline to assess recent trends in obesity. Ann. Hum. Biol., 126,4, 303-308.
[4] Cole, T. J., M. C. Bellizzi, K. M. Flegal, W. H. Dietz. Establishing a standard definition for child overweight and obesity worldwide : International suwey., BMJ, 2000, 5, 320,1-6.
[5] Goran, M.I., Gower, B.A., 1999. Relation between visceral fat and disease risk in children and adolescents. Am.Journ.of Clin. Nutrit. 70, 1495-1565.
[6] Keys. A., Fidanza, F., Karvonen, M.J., Mirmura, M., Taylor, H.L., 1972. Indices of relative weight and obesity. J.Chron. Dis., 25:429-434.
[7] Martin R., Saller K. 1957. Lehrbuch der Anthropologie, Stuttgart: Custav Fisher Verlag, 1,661.
[8] Mc Carthy, HD., Ellis, S., Cole, T. 2003. Central overweight and obesiry in Brit-ish youth aged 11-16 years: cross sectional surveys of waist circumference. BMJ, 326 ( 7390):624.
[9] Mc Carthy, HD., Jarrett, KV., Grawley, HF. 2001. Development of waist cir-cumference percentiles in british children aged 5.0-16.9y., Eur. J. Clin. Nutr., 55, 902-907.
[10] Mladenova, S. Nikolova, M. 2005. Components of body mass and their rela-tions during the growth period of the boys. Proceedings of the Balkan scientific conference of biology in Plovdiv, Bulgaria., 138-149.
[11] Mladenova, S., Nikolova, M., Boydzhiev, D. 2005. Body mass index, some cir-cumference indices and their ratios for monitoring of physical development and nutritional status of children and adolescents., Acta morphologica et anthro-pologica, 10, 226- 229.
[12] Nacheva, A., lazarova, E., Yordanova, L., 2003. Body nutritional statius 7-17 years old schoolchildren from Sofia, Yourn. of Antrop. , Sofia, 4, 21-25
[13] Obesity: preventing and managing the global epidemic. Report of a WHO con-sultation on obesity.June, 305, 1997.WHO, Geneva. Aviable at: http:// dosei.who.int/., Accessed April, 18, 2002.
[14] Rolland- Cachera M. F., 1998. Body mass Index References. The Cambridge Encyclopedia of Human Crowth and Development., Eds. Ulijaszek S. Y., Yohnston F. E., Preece in M. A. Cambridge: Cambridge Univ. Press., 68-69.
[15] Ross, W.D., Eiben, O.G., 2002. BMI: A critique of its use in numan biology and the health professions., Humanobiol. Budapest, 27, 47-56.
[16] Tomazo-Ravnik, T., Zerbo, D., 1996/97. Secular growth changes of Ljubljana schoolchildren in the period from 1958 to 1994 (longitudinal series)., Anthrop. Kozl., 38, 61-68.
[17] World Health Organisation, 1995. Physical status: the use and interpretation of anthropometry, Report of a WHO Expert Committee, Geneva, 854.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1033
[18] Zsoffay, B.K. et all., 1998. Body height, body weight and BMI of the school-children in three urban areas of Hungary.,Anthrop. Kozl.39, 71-80.
гл.ас. д-р Силвия Нагулова Младенова ПУ “Паисий Хилендарски” – Филиал Смолян ул. “Дичо Петров” 32 4700 Смолян Телефон: 0301 6 23 39, вътр. 32 e-mail: [email protected]
проф. дбн. Мима Иванова Николова ПУ “Паисий Хилендарски” ул. “Цар Иван Асен" 24 4000 Пловдив Телефон: 032 261510 e-mail: [email protected]

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 1034
SCIENTIFIC PAPERS
НОВ КРАТКОСРОЧЕН ТЕСТ ЗА ОПРЕДЕЛЯНЕ НA КАНЦЕРОГЕННИ ЗАМЪРСИТЕЛИ
В ПОЧВЕНИ ПРОБИ У НАС
М. Пешева, О. Кръстанова, М. Димитров СУ “Св. Климент Охридски”, Биологически Факултет, Катедра Генетика
П. Венков Институт по криобиология и хранителни технологии
секция “Молекулярна екология”
NEW TY1 TEST TO STUDY ENVIRONMENTAL SOIL POLLUTION WITH CARCINOGENS
М. Pesheva. O. Krastanova, M. Dimitrov
Sofia University “St. Kliment Ohridski”, Faculty of Biology, Department of Genetics
P. Venkov Institute of Cryobiology and Food Technology Department of Molecular Ecology a
ABSTRACT: The Ty1 short-term test is based on induction the transposition of the oncovirus-like Ty1 retrotransposon in a gene-engineered Saccharomyces cerevisiae test strain. Previous studies have evidenced the positive response of the test to a number of laboratory carcinogens, including such that are without apparent genotypic activity and therefore undetectable by the other short – term tests and the negative response of Ty1 test to mutagens without carcinogenic potential. In this communication we report the re-sults from the application of Ty1 test to study environmental samples simultaneously ana-lyzed chemically for the presence of mutagens, carcinogens, heavy metals, polycyclic aromatic hydrocarbons, pesticides and polychlorinated biphenols. The Ty1 test was posi-tive in concentration – dependent and kinetics experiments with all samples that contained carcinogens above the ecological standards and gave negative results with samples free of carcinogens, although some of them contained noncarcinogenic mutagens in high amounts. These results evidenced that the property of Ty1 test for selective detection of carcinogens, previously found in experiments with laboratory mutagens and carcinogens, can be extended to environmental studies. It is concluded that the usage of Ty1 test to-gether with another short – term test for environmental studies is advantageous and will give a more complete picture of pollution in the studied regions. Key words: Saccharomyces cerevisiae, Ty1 test, mutagens, carcinogens, environmental pollution

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1035
ВЪВЕДЕНИЕ Нашата страна е на едно от първите места в света по честота на ракови
заболявания. Това до голяма степен се дължи на неконтролируемото и пов-семестно замърсяване на околната среда. Известно е, че развитието на ин-дустрията е свързано с отделяне на големи количества замърсители в приро-дата. Използването на пестициди и други химикали в земеделието увеличава опасността от активирането им в растителните и животински ензимни систе-ми.
Доказано е, че голям процент от замърсителите на околната среда имат мутагенно и канцерогенно действие. За отчитане на този ефект е необходима подходяща тест система. До сега тези изследвания са провеждани с използ-ване на бактериални тест системи [1]. Въпреки, че тези тестове са бързи и ев-тини, те не дават реална представа за дисбаланса човек/природа, тъй като: 1) бактеpиалните и животински клетки имат различна плоидност, което опре-деля и разнообразните отговори спрямо мутагенното и канцерогенно дейст-вие; 2) тестовете с използване на бактериални клетки, откриват главно гене-тични изменения като: реверсии, точкови мутации и други, които са най-тривиалните дефекти в молекулата на ДНК.
Тестовете, базирани на използване на еукариотни клетки, имат както пре-димства, така и недостатъци, но диплоидното състояние на еукариотите поз-волява откриването на по-копмплексни генетични повреди. Веществата, пре-дизвикващи канцерогенеза у животните и човека, които нямат изявена гено-токсична активност, обикновено трудно могат да бъдат доказани с теста на Еймс. За това са разработени тестове на базата на дрождени клетки [2]. Въп-реки, че дрождите са използвани за създаване на различни тестове за мони-торинг на околната среда, само някои от тези тестове [3] имат реално практи-ческо приложение. Главната причина за това е по-слабия отговор на дрожди-те, като индикаторен организъм, който се дължи на ограниченият пермеаби-литет на клетъчната им стена [4]. Дрождите Saccharomyces cerevisiae имат подвижни генетични елементи, наречени Ту, като най- добре проученият е Ту1. Подобно на известните онковируси, цялата ДНК-последоветелност на Ту1 се транскрибира в РНК-молекули. Ту1-РНК е междинна стъпка в транспо-зицията, като новото копие на Ту1-ДНК, получени чрез обратна транскриптаза може да се интегрира на различни места в генома. Включването на Ту1 тран-спозона създава геномна нестабилност, поражда различни мутации и е придружено от големи геномни реорганизации, подобни на тези, при канцеро-генезата. Така Ту1 е ретротранспозон с клетъчен цикъл, много подобен на то-зи на онковирусите [5].
Установено е, че от досега съществуващите тестове няма такъв, който да може да се прилага за контрол над замърсяването на околната среда, при из-следване на голям брой проби, за кратък период от време. Повечето тестове не различават мутагенно от канцерогенно действие, а такава разлика същест-вува, тъй като някои мутагени нямат канцерогенни свойства. За анализ на му-тагенното/канцерогенно действие на природни замърсители е конструиран от нас нов тест [6]. Новото тук е именно използването на генно-инженерен онко-вирус-подобен елемент Ту1 ретротранспозон. Конструираният от нас тест е много чувствителен и разграничава например канцерогенния бензо(а)пирен или шествалентния хром от неканцерогенните бензо(е)пирен или тривалент-ния хром [6]. Установено е [7], че транспозицията на Ту1 ретротранспозона е

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1036
индуцирана от различни вещества, имащи канцерогенно действие и зависи от функцията на гена rad9 и преминаването на G1-фазата от клетъчния цикъл. Дрожденият ген Rad9 е компонент на ДНК-репарационната система и е част от сигнално-трансдукционния път при дрождите, който стопира клетъчния ци-къл, при установяване на повреда в молекулата на ДНК. Генът rad9 се явява хомолог на главния човешки туморосупресорен ген р53 [8]. Тези данни свиде-телстват за съществуването на сходство при Ту1-ретротранспозицията и не-опластичната трансформация на клетките и доказват необходимостта от из-ползването на Ту1 теста за откриване и доказване на канцерогени.
ЦЕЛ Целта на настоящата работа е да покажем, че разработения от нас Ту1
тест може успешно да бъде прилаган за откриване на замърсители в околната среда, имащи канцерогенно действие и за разграничаване на канцерогени от мутагени. Прилагането на Ту1 теста, заедно с теста на Еймс дава една по-цялостна картина за статуса на замърсяване на различни изследвани от нас региони в нашата страна.
МАТЕРИАЛИ И МЕТОДИ 1.Събиране и анализ на почвените проби Почвените проби бяха събирани в периода 2004-2005 година, в обявени
от Националния център по екология различни региони в България, имащи ниски и високи нива на замърсяване. Пробите бяха взети от две села, нами-ращи се близо до София. За гара Яна е характерно силно замърсяване, за-щото се намира в близост до голям металургичен завод и Локорско, което има ниски нива на замърсяване.
Пробите бяха екстрахирани съгласно методика, публикувана от Rossi et al., [9]. Изсушените екстракти бяха разтваряни в метанол или диметилсулфок-сид. Всяка почвена проба е представителна за участък от 10m2. Непосредст-вено преди използване за анализа, пробите бяха концентрирани десеткратно през евапоратор и анализирани незабавно, като бяха направени няколко пов-торения в продължение на 15 дни. Пробите бяха съхранявани при температу-ра –20oС, като по време на съхранението не е наблюдавана промяна в тях.
Беше направен кратък химичен анализ на изследваните почвени проби, като резутатите са представени в таблица 1. В състава им са открити олово (Pb), мед (Cu), арсен (As), цинк (Zn), кадмий (Cd), нафтален, аценофтен, аце-нафтилен, флуорантен, пирен, бензо(а)антрацен, кризен, бензо(а)пирен, бен-зо(е)пирен, индено(1,2,3-Сd)пирен, дибензо(а)антрацен и петролни продукти. Открити бяха и други вещества с изключителна ниска концентрация и за това не са включени в таблицата. Почти във всички проби в минимални количества присъстваха пестициди (α-HCH, β-HCH, γ-HCH, ∆-HCH, ε-HCH ) хептахлор, алдрин, изодрин, еудрин, цис-хептахлор, епоксиди и други, както и полихло-рирани бифеноли, такива като: РСВ28, РСВ52, РСВ11 и т.н.
2. Използвани щамове и реактиви Всички използвани за анализ на пробите реактиви и химикали са закупе-
ни от Sigma. Като разтворители за приготвяне на сток-разтворите, непосредс-

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1037
твено преди всеки експеримент, бяха използвани вода или натриев винил-сулфонат, терт-бутилхидроквинолин, примидон, метанол.
Хранителните среди за култивиране на дрождения щам бяха приготвени съгласно методика на Sherman et al., [10], с използване на компоненти от Difco Chem. Co.
Използван беше щам Saccharomyces cerevisiae DG1141ts1 (MATα ura3-167 HIS3∆200: Tym HIS3AI), [11]. Алелът ts1 е мутантен алел на гена sec53, който увеличава клетъчния пермеабилитет към различни субстанции, влючи-телно към мутагени и канцерогени [6]. Това има значение за чувствителността на метода. Освен, че притежава температурочувствителна мутация, щамът DG1141ts1 има и Ту1 елемент, маркиран с индикаторен ген HIS3AI. Генът HIS3 съдържа един изкуствен интрон (AI-Artificial Introne), включен в антисенс ориентация по отношение на HIS3. Всяка транспозиция на маркерния Ту1 во-ди до поява на прототрофност по хистидин и изисква транскрипция на моди-фицирания Ту1, сплайсинг на AI, обратна транскрипция на Ту1-РНК, съдър-жащ HIS3 гена и включване на получената кДНК на ново място в генома. Това позволява появата на една HIS+ позитивна колония върху синтетичната среда без хистидин. По този начин, броят на HIS+ колониите е количествено измере-ние за честотата на транспозицията на маркерния Ту1 транспозон.
3. Тест за транспозиция на Ту1 ретротранспозона Клетките бяха култивирани в течна YPD-среда при 30оС за 24 часа. След
това бяха разреждани до 2х107 клетки/ml и в свежа YPD-среда и инкубирани повторно при същите условия до получаване на течна култура, с плътност около 4-6х107 клетки/ml. Получената култура беше разделена на части и клет-ките бяха третирани с различни концентрации на агента (канцероген, проба от околната среда и др.). Обемът на пробата, която изпитваме беше 200µl и се добавя към 3.8ml клетъчна суспензия. Следва култивиране за 30 min при 30оС, двукратно промиване на клетките с YPD-среда и ресуспендиране в све-жа YPD-среда и култивиране за 24 часа при 20оС, с цел завършване на Ту1-транспозицията. След съответно разреждане клетките бяха посявани на твърда YPD-среда за определяне на токсичното действие, на съответната концентрация от анализираното вещество (или почвена проба), като се опре-деля броя на преживелите клетки в контролата и опита. Клетките бяха пося-вани и на селективна среда без хистидин (SC-HIS) за определяне броя на ко-лониите, прототрофни по хистидин. Средната скорост и “пъти нарастване” на честотата на транспозицията се определя чрез уравнението на Дрейк [12]. За позитивен отговор на Ту1 транспозиционния тест се приема два или повече пъти нарастване на честотата на транспозицията, в сравнение с контролата.
РЕЗУЛТАТИ И ОБСЪЖДАНЕ Резултатите, получени при изследване на почвените проби са предста-
вени в таблица 2. Във всички проби, събрани от чисти региони липсват канце-рогенни съединения и дават отрицателни резултати на Ту1 транспозиционния тест (пъти нарастване на честотата на транспозицията по-ниско от 2.0), както и висока степен на преживяемост на дрождените клетки.
Пробите, събрани от райони със силно замърсяване, показват различни нива на токсичност, с тенденция към увеличаване на токсичността за пробите, съдържащи петролни продукти (S-8, S10-1). Всички проби показват позитивен

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1038
резултат на Ту1 теста, като честотата на транспозиция на Ту1 транспозона нараства от 4 до 8 пъти.
Пробите от замърсените райони (таблица 3), показват че даже относи-телно ниски концентрации имат силно токсичен ефект и дават позитивен ре-зултат в Ту1 теста. Резултатите от анализ на почвени проби, взети от сравни-телно чисти региони са различни. Слабото увеличение на средните стойности на Ту1 транспозицията се дължи на увеличението на концентрацията на ди-метилсуфоксида. В предишни опити [13] е показано, че при концентрация на диметилсулфоксид над 5% се увеличава и изменя индукцията на Ту1 транс-позицията. Същото бе потвърдено и в настоящото изследване (контролите в табл. 3). По тази причина концентрацията на диметилсулфоксида не трябва да превишава 5% от общия обем на пробата.
Зависимостта на увеличението на честотата на Ту1 транспозицията от времето за експониране с проби от замърсените райони е показана в табл. 4. Продължителното третиране корелира с увеличаване на честотата на Ту1 транспозицията. Химичният анализ на почвените проби (таблица 1) показва отсъствие на канцерогени в почвените проби от чистите региони и наличие на канцерогенни субстанции и тежки метали от замърсените райони. Специално внимание беше отделено на пробите S 1 и S 3, които съдържат нарастващи концентрации от флуорантен и показват негативен отговор на Ту1 теста, при увеличаване на концентрацията и времето за експониране.
Известно е, че флуорантена е потенциален мутаген в бактериални клетки и в клетки на бозайници в in vitro тест-системи, обаче получените резултати от изследвания в клетки на гризачи in vivo не показват канцерогенен ефект [15, 16].
Негативният резултат от почвените проби S1 и S3 на Ту1 теста, потвърж-дава нашите предишни резултати [14], получени за голям брой лабораторни неканцерогенни мутагени. Това още веднъж потвърждава възможностите на Ту1 транспозиционния тест за прилагане при осъществяване на контрол над замърсители на околната среда.
Химичният анализ на почвените проби от замърсените райони показва наличие на канцерогенни субстанции, което напълно корелира с положител-ните резултати, получени от нас на Ту1 теста. Изключение правят пробите S1 и S3 (от чисти райони), които дават отрицателен резултат на Ту1 транспози-ционния тест и не съдържат в състава си канцерогенни мутагени.
Чувствителността на Ту1 теста за откриване на канцерогенни замърси-тели изглежда е много висока. Всички проби, съдържащи канцерогени в кон-центрации, по-високи от екологично допустимите норми, показват и положи-телен резултат на Ту1 транспозиционния тест, характеризиращ се с от 4 до 8 пъти увеличение на честотата на транспозицията. Освен това концентрация-та на всеки канцерогенен замърсител на околната среда, може лесно да бъ-де изчислен в µg/ml . Например пробата S4 съдържа 16µg/ml бензо(а)пирен и показва положителен резултат на Ту1 теста, а S2 съдържа 85µg/ml бен-зо(а)антрацен.
Пробите от околната среда съдържат смес от различни канцерогени и мутагени, които могат да умножат ефекта на въздействие, който обикновено дават отделните субстанции. Досега, ние не сме установили как Ту1 теста, може да даде отговор за разграничаване на ефекта на действие на всеки

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1039
един отделен канцероген, съдържащ се в сместа от различни канцерогени, установени в пробите.
Възможни са няколко обяснения: 1) Тестерният щам Saccharomyces cerevisiaе, съдържа маркерен Ту-
елемент, вкаран в геномната ДНК и е с увеличен пермеабилитет на дрожде-ната клетъчна стена към мутагените/канцерогените, който се дължи на ts1-мутацията [17]; [18], [6].
2) Ту1 е ретротранспозон, с жизнен цикъл и структура, много сходна с та-зи на известните ретровируси и онковируси [5]. Именно това позволява да бъ-де получен избирателен отговор към канцерогенните вещества. Докато пове-чето от съществуващите до сега тест-системи разкриваха индуцирането на специфични генетични изменения, Ту1 теста е способен да доказва онези повреди в молекулата на ДНК, които са характерни за неопластичните транс-формации.
3) Получените от нас резултати са доказателство за това, че при въз-действие с нарастваща концентрация на Ту1-позитивните канцерогени се ин-дуцират ROS (реактивни кислородни видове). Така например бензо(а)пирена и бензо(а)антрацена индуцират in vitro окислителен стрес, съпроводен с повре-ди в молекулата на ДНК, във фрагмента, съсържащ човешкия р53 туморсупре-сорен ген. Образуването in vivo на 8-оксо-7,8-дихидро-2’-дезоксигуанозин, е ин-дикатор за окислителни поврeди в молекулата на ДНК в телешки тимус [19].
Арсенът и други тежки метали, даващи положителен резултат на Ту1 тес-та [14], индуцират редокс-чувствителните транскрипционни фактори и генери-рат реактивни кислородни и азотни видове в тъканни култури от клетки на хамстер [20], [21]. Тези и други литературни данни отчетливо потвърждават, че позитивният отговор на Ту1 теста винаги е съпроводен с увеличаване про-дукцията на ROS. Резултатите от предишни и настоящи наши изследвания показват, че избирателността на Ту1 теста към канцерогени се губи в тестер-ни клетки, в които липсват функциониращи митохондрии, обозначени като rho- и rho0 и са производни на щама DG1141ts1, или в клетки с разрушени SCO1 ядрен ген, участващ в митохондриалното окислително фосфорилиране. Тези резултати показват ключовата роля на повишените нива на ROS за определя-не на чувствителността на Ту1 теста към канцерогени.
Необходими са допълнителни молекулярно-биологични експерименти (които провеждаме в момента) за по-детайлното изясняване на механизма на чувствителност на Ту1 теста към канцерогени.
ЗАКЛЮЧЕНИЕ Откриването на различни канцерогени и мутагени-замърсители на окол-
ната среда е задължителна и важна стъпка при възстановяването на баланса между природната среда и човека. Ту1 транспозиционния тест е вероятно първият тест за избирателно откриване на канцерогенни замърсители, които са особено опасни за здравето на човека и бозайниците. Той е евтин, бърз и чувствителен метод за откриване на канцерогенни замърсители в околната среда. Прилагането му, заедно с теста на Еймс, може да се използва за мони-торинг на околната среда и дава ясна и пълна картина на замърсяванията в изследваните региони.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1040
Тази разработка е финансиpана от проект SfP 977977 – NATO, да-ден на проф.д-р П. Венков.
ЛИТЕРАТУРА
[1] Maron D. M., B. Ames, 1983, Revised Methods for the Salmonella mutagenicity test., Mut. Res., 113., 173-215
[2] Ramel C., H. Cederberg., J. Magnusson, E. Vogel., A. Natarajan, L. Mulender, J. Nivard, J. Parry, A. Leyson, M. Comendador, L. Sierra, J. Ferreira, S. Consuegra, 1996, Somatic Recombination, Gene Amplification and Cancer., Mut. Res., 421., 117-120
[3] Zimmermann F., R. von Hakke., J. Parry., D. Siebert., G. Zetterberg., R. Barale., N. Leprieno, 1984, Testing of chemicals for genetic activity with Saccharomyces cerevisiae: a Report of the US Environmental protection agency gene-tox program., Mut. Res., 133, 199-214.
[4] Morita T., Y. Iwamoto., T. Shimizu., T. Maswzawa., Y. Yanagihara, 1989, Mutagenicity test with a permeable mutant of yeast on carcinogens showing false negative Salmonella assay., Chem. Pharm. Bull., 37, 407-409.
[5] Garfinkel D., 1992, Retroelements in microorganisms, in: J.A. Levy (Ed). Retroviridae, Plenum Press, NY., p. 107-136
[6] Pesheva M., Krastanova O., Staleva L., Dentcheva V., Hadzhitodorov M., Venkov P., 2005, The Ty1 transposition assay: A new short-term test for detection of carcinogens, Jour. of Microbiol. Methods, 61, 18.
[7] Staleva L. S, Venkov P.V., Activation of Ty transposition by mutagens, Mut. Res., 2001, 474, 93-103.
[8] BertramJ. S., 2001, The molecular biology of cancer., Mol.Asp.Medicine., 21.,167-223.
[9] Rossi C., Poli P.,Buschini A.,Cassoni F., Magnani F., Luartini . Tolomei S., Girbelli C., god Ocupacional genotoxicity assessment by mutagenicity assays, Toxicol.Lett., 77., 289-298.
[10] Sherman F., Fink G., Hiks G., 1986, Laboratory course manual for methods in yeast genetics. (Eds) CSHLP.
[11] Curcio M. J., Garfinkel D.J., 1999, New lines of host defense: Inhibition of Ty1 retrotransposition by Fus 3p and NER / TFII H., Trends of Genet.., 15., 43-45
[12] Drake J.W., The molecular bases of mutation. Holden Day, Ocland, CA., 1998., p.382
[13] Pesheva M., Venkov P., Dimethylsulfoxide has a recombinogenic effect on Saccharomyces cerevisiae cells, 2005, Biotechnology and Biotechnological Equipment, 19, 38-42.
[14] Pesheva M., Krastanova O., Stamenova R., Venkov P., Response of Ty1 transposition test to carcinogens, Mut. Res. In press.
[15] Goldman R., Enwold L., Pellizzari E., Beach G., Bowman E., Krishnan S., Shields P., 2001, Smoking increases carcinogenic polycyclic aromatic hydrocarbons in human lung tissue., Cancer Res., 61., 6064-6371
[16] Verschueren E., 2001, Fluoranthene., Handbook at Environmental data on Organic Chemicals., NY., John-Willey and Sons., Inc. 4th Ed. Vol.1., 1158-1161.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1041
[17] Staleva L., Walcheva L., Venkov P., 1996, Enchanced permeability increases the sensitivity of a yeast test for mutagens, Mut. Res., 370, 81-89.
[18] Terzyska A., Walcheva L., Venkov P., 2000, A new sensitive method based on yeast cells for studying environmental pollution., Environ. Poll., 109., 43-52
[19] Seike K., Murate M., Hirakawa K., Deyashiky Y., 2004, Oxidative DNA damage induced by benzo(a)anthracene dihydrodiols in the presence of dihydrodioldehydrogenate., Chem. Res. Toxicol., 17, 1445-1451.
[20] Ueno S., Sugiwama M., Susa N., Furukawa Y., 1993, Effect of dimrthyltiourea on chromatne (VI) - induced DNA single- strand breaks in Chinese hamster V-79 cells., Mut. Res., 346, 249-253
[21] Valko M., Morris H., 2000, Cronin N., Metals toxicity and oxidative stress., Current Med. Chemistry., 12 (1010), 1161-1208. гл.ас. д-р Маргарита Георгиева Пешева гл.ас. д-р Олга Микова Кръстанова студент-магистър Мартин Димитров Димитров Софийски университет “Св. Климент Охридски”, Биологически Факултет, Катедра Генетика, Бул. “Драган Цанков” № 8, 1421 София, 8167261 е-mail: [email protected] проф. д-р Пенчо Василев Венков Институт по криобиология и хранителни технологии секция “Молекулярна екология” бул. “Черни връх” № 53А, 1407 София,
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1042
ПРИЛОЖЕНИЯ
Таблица 1. Химичен анализ на почвени проби
Замърсители в почвени проби (mg/kg)
Чисти региони Замърсени региони
Химични съединения
Екол
огич
ни
станда
рти
(mg/
kg)
S1 S3 S5 S2 S4 S6 S8 S10-1 S10-4
Pb 50 38 34 39 334 346 317 215 ND 293 Cu 150 29 28 26 73 85 70 169 ND 338 As 25 19 11 9 103 73 70 11 ND 356 Zn 200 78 78 68 342 305 184 215 ND 257 Cd 2 0.4 0.4 0.4 2.2 2.2 1.0 1.0 ND 1.7 Ni 25 ND ND ND ND ND ND ND ND 29 Naphthalene 0.05 0.05 0.05 0.05 0.12 0.05 0.12 0.48 0.18 ND Aceno-phthene
0.03 0.003 0.003 0.002 0.007 0.02 0.01 0.08 0.03 ND
Acenaphtha-lene
0.03 <0.001 <0.001 0.001 <0.001 <0.001 0.009 0.023 0.04 ND
Fluorene 0.03 <0.001 <0.001 0.006 <0.001 <0.001 0.017 0.080 0.060 ND Phenan-threne
0.045 0.042 0.045 0.035 0.145 0.019 0.132 0.840 0.080 ND
Anthracene 0.05 <0.001 0.003 0.010 0.021 0.010 0.015 0.120 0.060 ND Fluoranthene 0.02 0.262 0.136 0.027 0.214 0.053 0.128 0.780 0.080 ND Pyrene 0.02 0.018 0.021 0.012 0.183 0.047 0.081 0.690 0.090 ND Benzo(a) anthracene
0.02 0.006 0.005 0.002 0.085 0.020 0.043 0.370 0.130 ND
Chrysene 0.02 0.019 0.002 0.002 0.117 0.057 0.053 0.340 0.100 ND Benzo(e) pyrene
0.015 0.011 0.015 0.001 0.092 0.176 0.001 0.280 1.000 ND
Benzo(a) pyrene
0.02 0.001 0.001 0.001 0.016 0.160 0.001 0.190 0.060 ND
Indeno(1,2,3-cd)pyrene
0.02 0.001 0.001 0.001 0.716 0.001 0.001 0.002 0.080 ND
Dibenzo(a,h) anthracene
0.02 0.001 0.001 0.001 0.001 0.001 0.001 0.003 0.004 ND
Benzo(ghi) pyrene
0.02 0.001 0.001 0.001 0.692 0.001 0.001 0.007 0.950 ND
Petrol products
50 ND ND ND ND ND ND 115 7809 ND
ND – не са определяни
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1043
Таблица 2. Ty1 транспозиционен тест за анализ на почвени проби
Проби
Преживя-емост (%) a)
Честота на Транспозицията
x 10-7 b)
Пъти нарастване на честотата на транспозицията
Мутагени/ канцерогени установени при химичния
анализ с) Контрола (5% MeSO) 100 (621) 3.4 ± 0.8 1.0 Чисти региони
S1 95 (590) 5.2 ± 1.1 1.6 Fluoranthene S3 93 (578) 4.4 ± 1.3 1.4 Fluoranthene S5 98 (609) 5.0 ± 0.8 1.5 None Замърсени региони
S2 82 (509) 10.6 ± 1.4 3.8 As,B(a)P, B(a)A, pyrene, cry-sene, benzo (ghi)perylene, B(e)P, fluorenthene, Pb
S4 59 (366) 8.6 ± 1.0 4.2 As, B(a)P, pyrene, crysene, B(e)P, fluorenthene, Pb
S6 71 (441) 9.5 ± 0.8 3.9 As, B(a)A, pyrene, crysene, fluoranthene, Pb
S8 55 (342) 12.2 ± 1.7 6.5 B(a)P, B(a)A, crysene, petrol products, B(e)P fluoranthene, Pb, A
S10-1d) 35 (217) 10.1 ± 0.9 8.5 B(a)P, B(a)A, benzo (ghi)perylene, crysene, petrol products fluoranthene, B(e)P
S10-4 73 (453) 11.9 ± 0.9 4.8 As, Ni, Cu, Pb
a) среден брой колонии b) усреднени стойности от 8 експеримента c) вещества, открити при химичния анализ (табл 1), кои-то показват стойности над екологично допустимите норми и окачествени като канцерогени
d) в теста е анализиран в 100 µl
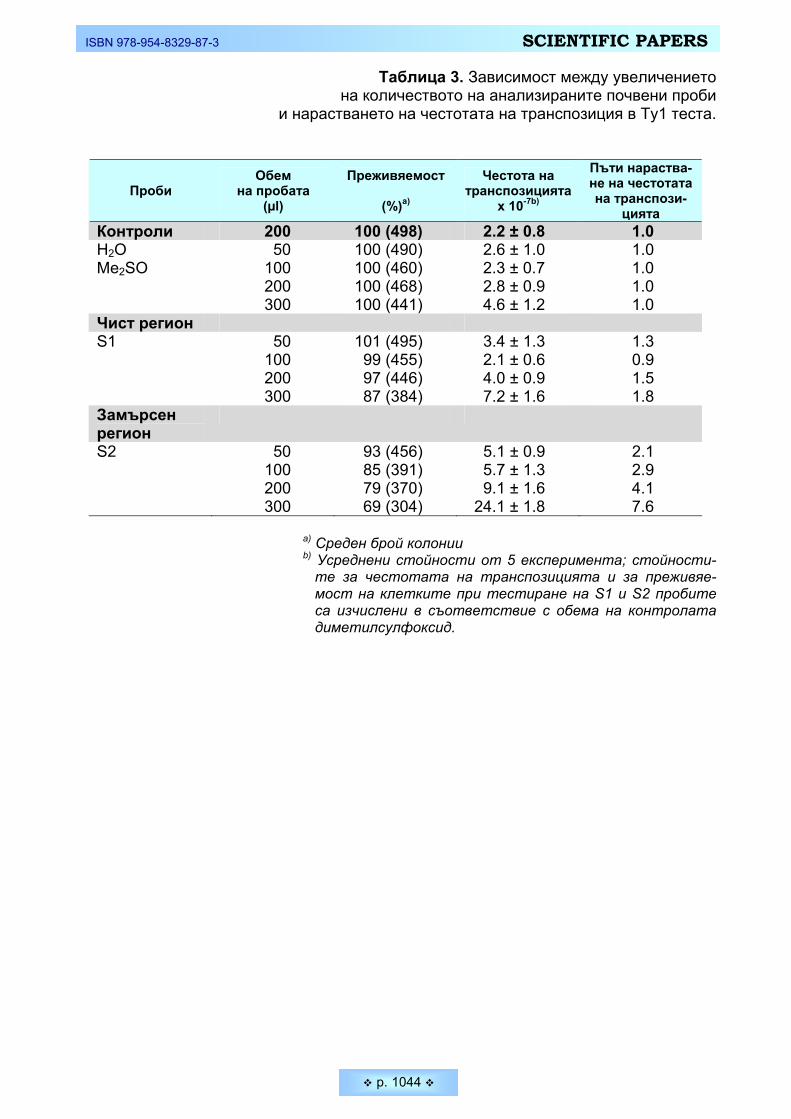
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1044
Таблица 3. Зависимост между увеличението на количеството на анализираните почвени проби
и нарастването на честотата на транспозиция в Ту1 теста.
Проби Обем
на пробата (µl)
Преживяемост
(%)a)
Честота на транспозицията
x 10-7b)
Пъти нараства-не на честотата на транспози-
цията Контроли 200 100 (498) 2.2 ± 0.8 1.0 H2O 50 100 (490) 2.6 ± 1.0 1.0 Me2SO 100 100 (460) 2.3 ± 0.7 1.0 200 100 (468) 2.8 ± 0.9 1.0 300 100 (441) 4.6 ± 1.2 1.0 Чист регион S1 50 101 (495) 3.4 ± 1.3 1.3 100 99 (455) 2.1 ± 0.6 0.9 200 97 (446) 4.0 ± 0.9 1.5 300 87 (384) 7.2 ± 1.6 1.8 Замърсен регион
S2 50 93 (456) 5.1 ± 0.9 2.1 100 85 (391) 5.7 ± 1.3 2.9 200 79 (370) 9.1 ± 1.6 4.1 300 69 (304) 24.1 ± 1.8 7.6
a) Среден брой колонии b) Усреднени стойности от 5 експеримента; стойности-те за честотата на транспозицията и за преживяе-мост на клетките при тестиране на S1 и S2 пробите са изчислени в съответствие с обема на контролата диметилсулфоксид.
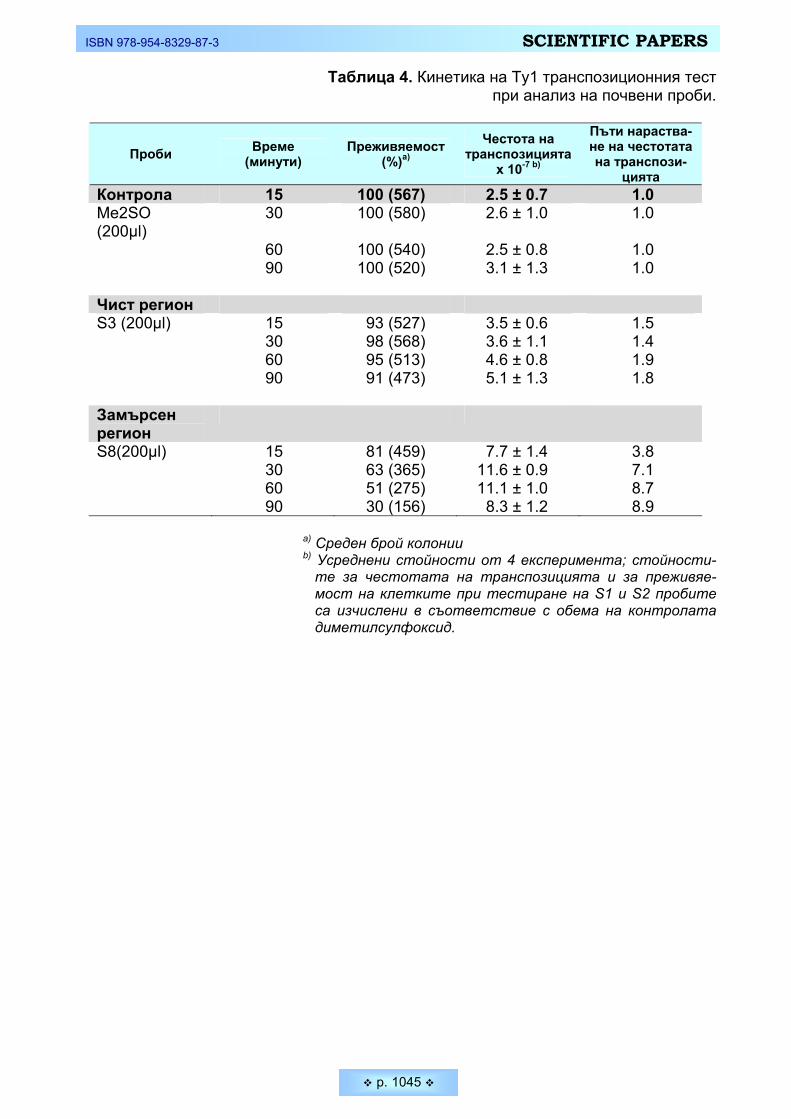
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1045
Tаблица 4. Кинетика на Ту1 транспозиционния тест при анализ на почвени проби.
Проби Време (минути)
Преживяемост (%)a)
Честота на транспозицията
x 10-7 b)
Пъти нараства-не на честотата на транспози-
цията Контрола 15 100 (567) 2.5 ± 0.7 1.0 Me2SO (200µl)
30 100 (580) 2.6 ± 1.0 1.0
60 100 (540) 2.5 ± 0.8 1.0 90 100 (520) 3.1 ± 1.3 1.0 Чист регион S3 (200µl) 15 93 (527) 3.5 ± 0.6 1.5 30 98 (568) 3.6 ± 1.1 1.4 60 95 (513) 4.6 ± 0.8 1.9 90 91 (473) 5.1 ± 1.3 1.8 Замърсен регион
S8(200µl) 15 81 (459) 7.7 ± 1.4 3.8 30 63 (365) 11.6 ± 0.9 7.1 60 51 (275) 11.1 ± 1.0 8.7 90 30 (156) 8.3 ± 1.2 8.9
a) Среден брой колонии b) Усреднени стойности от 4 експеримента; стойности-те за честотата на транспозицията и за преживяе-мост на клетките при тестиране на S1 и S2 пробите са изчислени в съответствие с обема на контролата диметилсулфоксид.

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 1046
SCIENTIFIC PAPERS
СЪСТОЯНИЕ И ПРОБЛЕМИ ЗА СЪХРАНЕНИЕ НА ЗАЩИТЕНА ТЕРИТОРИЯ
“ГОЛЯМО СМОЛЯНСКО ЕЗЕРО”
Агоп Сренц1, Христо Христов1, Диана Кирин2 1Пловдивски университет “Паисий Хилендарски”
2Аграрен Университет – Пловдив
CONDITION AND PROBLEMS FOR CONSERVATION ON THE PRESERVE TERRITORY
“BIG SMOLYAN'S LAKE”
Agop Srentz1, Khristo Khristov1, Diana Kirin2 1Plovdiv University 'Paissii Khilendarski'
2Agricultural University – Plovdiv
ABSTRACT: Samples from the plant tufts in the Big Smolyan’s lake were taken in
way allowing to trace the vertical distribution of Cs-137. The measurement were taken by precision gamma ray spectrometer. The specific and surface activity was calculated.
Key words: Smoyian’s lake, plant tufts, Cs-137, Potamogeton natans, Меnyanthes trifoliata ВЪВЕДЕНИЕ Едни от най-значимите влажни зони в България са Смолянските езера.
Разположени са в Западните Родопи, на 5 km северно от гр. Смолян. По про-изход те са свлачищни езера. Разположени са върху няколко свлачищни стъ-пала, на надморска височина от 1300-1500 m. Настоящото изследване се от-нася до състоянието и възможностите за опазване на "Голямо Смолянско езеро", наричано още в литературата Смолянско езеро – 2 или Тревистото езеро. Намира на малка надморска височина – 1500 m. Езерото е част от при-родната забележителност "Смолянски езера".
ЦЕЛ Разположено в район с реални перспективи за развитие на екотуризъм,
съхранението на този естествен водоем има безспорна и многостранна зна-чимост.
Това може да се постигне, ако наред с ограничителни мерки бъдат извършени и активни природозащитни мероприятия.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1047
МАТЕРИАЛ И МЕТОДИ Настоящото изследване е извършено в части от защитената територия
"Смолянски езера", представена от сладководни езерни екосистеми и приле-жаща вековна смърчова гора (Рicea abies (L.) Karsten), разположени на над-морска височина от 1300 до 1500 m.
Характеризирани са биологичното разнообразие на хидрофитната расти-телност, тяхната консервационна значимост, както и като цяло флористичните съобщества.
Екологичните характеристики при настоящите изследвания се основават на терминологията на Rosenberg [1] и Маgurran [2]. Лабораторните изследва-ния са извършени в лабораториите нa катедра Атомна физика, Пловдивски университет "Паисий Хилендарски" и катедра ”Агроекология”, Аграрен универ-ситет – Пловдив.
РЕЗУЛТАТИ И ОБСЪЖДАНЕ СЪСТОЯНИЕ НА ПРОУЧЕНОСТ НА ТЕРИТОРИЯТА Първа научна публикация за езерото е направено от проф. Вълка-
нов,1938г.[3]. Към настоящия момент броят на публикациите е над 30. Детайлни ботанически проучвания са направени от проф. Воденичаров и
проф. Киряков в периода 1965-1985 г., а през 1972 г. и 1974 г. са предприети подводни изследвания под ръководството на проф.Воденичаров, като съв-местно със Станцията на младите агробиолози, Смолян, организирани от Костадинка Сурова, е направено картиране на езерото.
Дискусия за необходимостта от действия, целящи опазването на този обект, е провеждана единствено в средствата за масова информация.
В периода 1996–1999 г. с проект “Орфей”, финансиран от фонд “Научни изследвания”, МОНТ, са предприети радиационни измервания в Смолянски регион, като една от целите е установяване динамиката на процеса на забла-тяване на езерото чрез изследване концентрацията на Cs-137. В периода 1999 – 2006 г. изследванията бяха продължени, с финансовата подкрепа на фонд “Научни изследвания” и участието на “Клуб за подводни изследвания” към Пловдивски университет.
Извършено бе: - картиране на езерото с GPS; - измерване на дълбочини; - сондиране на дъното и подготвяне на проби седиментни слоеве; - послойно определяне съдържанието на Cs-137; - експериментално частично отстраняване на растителност от крайб-
режна ивица и свободна повърхност; - периодично сондиране на растителните туфи. ПРОМЕНИ В СЪСТОЯНИЕТО НА “ГОЛЯМО СМОЛЯНСКО ЕЗЕРО” Обилната растителност и естествените процеси протичащи в езерото
имат като резултат неговото заблатяване. Отличителна особеност в този слу-чай е наличието на плаващи растителни туфи, поради което процеса на заб-латяване е иницаран не от бреговата ивица, а от централната му част.
През последните 10 години, процесите на заблатяване са обусловени още и от развитие на растителни видове в:

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1048
- свободната водна повърхност; - периферията на езерото – от брега до дълбочина 50–70 cm.
А) Биологично разнообразие и екологични особености на хидро-фитната растителност.
През периода на настоящото проучване, висшите водни растения в езер-ните екосистеми са представени основно от асоциациите на Potamogeton natans (фиг. 1), Lemna minor, Lemna trisulca, Utricularia vulgaris, Utricularia minor, Меnyanthes trifoliata (фиг. 2), Саrex rostrata, Саrex flava, Juncus bufonius, Equisetum limosum, Роtentila palustris, Sparganum minimum, Sphagnum sp. и др. [4, 5, 6, 7, 8] (Таблица 1). Те са представени с най-висока численост в лито-ралната зона на езерата и могат да служат като естествена бариера при пос-тъпване във водоемите на евентуални частични замърсявания, както и като инхибитори на сапрофитната микрофлора, участвайки по този начин в проце-сите на самоочистване на водите. Изключение правят асоциациите на Potamogeton natans, покриващи цялото водно огледало на Смолянско езеро – 2. Бързото развитие на висшата водна растителност е довело и до интензив-ни процеси на заблатяване в резултат на автохтонно биогенно замърсяване.
Сравнено с предходни изследвания трябва да се отбележи, че при нас-тоящото изследване са настъпили значителни изменения при Смолянско езе-ро-3, което е със значително увеличен дебит поради обилните снеговалежи и дъждове, което е изключително благоприятно на фона на наблюдаваните преди това изменения и естествени сукцесионни процеси, които го превръщат в ливада (обилните дъждове през лятото на 2005 г.).
От установените доминиращи таксони висши водни растения, Sphagnum sp. са биоиндикатори за олигосапробност; видовете Lemna minor, Utricularia vulgaris са биоиндикаторни за β—мезосапробност; Lemna trisulca — за 0-β—мезосапробност.
Видовете Кръглолистна росянка Drosera rotundifolia L. (Droseraceae), Богородична лъжичка, Горчивче Menyanthes trifoliata (Мenyanthaceae), Мочурно прозорче Роtentila palustris (Rosaceae), Малка ежова главичка Sparganum minimum (Sparganiaceae), Блатен плаун Lycopodiella inundata (Lycopodiaceae), Тинеста острица Саrex limosa (Cyperaceae), Малка мехурка Utricularia minor (Lentibulariaceae), са защитени видове (Закон за биологичното разнообразие, ДВ, бр. 77 от 2002 г.).
Установените видове Drosera rotundifolia L. (Droseraceae), Menyanthes trifoliata (Menyanthaceae), Potentila palustris (Rosaceae), Lemna minor (Lemnaceae) са лечебни растения (Закон за лечебните растения, ДВ, бр. 29 от 07.04.2000 г.).
Видовете Lycopodiella inundata (Lycopodiaceae), Саrex limosa (Cyperaceae), Utricularia minor (Lentibulariaceae), Utricularia vulgaris L., Drosera rotundifolia L. (Droseraceae), Potentila palustris (Rosaceae), Menyanthes trifoliata (Menyanthaceae), Sparganum minimum (Sparganiaceae), са включени в Черве-ната книга на България, том І .
Блатният плаун (Lycopodiella inundata; Lycopodiaceae) се среща в тор-фищни съобщества, рядко в асоциации на кисели треви, сфагнов торф, по периферията на млади мочурища, понякога в открита водна площ. Разпрост-ранен е в пояса на иглолистните гори на 1200–1600 m надморска височина. До сега у нас блатния плаун е установен само около Смолян включително и

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1049
Смолянско езеро – 2. На неговата територия се развиват малко екземпляри – предполага се, че се касае за начална фаза на залесяване, тъй като е устано-вено едва през последните няколко години. Отрицателно действащи фактори за развитието му са намаляването на торфищата след изсичане на горите и пресушаване на водните площи. Взети са мерки за опазване – находището на вида в езерата се намира в защитена територия.
Тинестата острица (Саrex limosa; Cyperaceae) се среща във високи пла-нински торфища и заторфени участъци в езерата на торфено-блатни почви, формирани върху силикатни скали от 1300 – 1800 m надморска височина. У нас тинестата острица се среща само в трофищата из високите части на Ви-тоша и територията на Смолянските езера. Отрицателно действащи фактори са бързото обрастване и затлачване на езерата, мелиоративните мероприя-тия и замърсяването.
Кръглолистната росянка (Drosera rotundifolia; Droseraceae) е едно от мал-кото растения – хищници у нас. Расте в торфени и мочурливи терени, пре-димно в субалпийския, иглолистния и буков пояс от 930–2000 m надморска височина. В района на Смолянско езеро – 2 росянката се среща масово и на места образува големи групи, които веднага правят впечатление с червеника-вия си оттенък. Отрицателно действащи фактори са отводняването на тор-фищата и общото засушаване на регионалния климат, вследствие на разреж-дане и намаляване на продуктивността и площта на горите.
Малката мехурка (Utricularia minor; Lentibulariaceae) е разпространена из блатата и локвите от предпланинския до високопланинския пояс, в Драгоман-ското блато, във водоеми на Западните Родопи, Пирин и Витоша. Отрицател-но действащи фактори – пресушаването на блатата и езерата, корекциите на речни корита, замърсяването на водоемите.
Обикновената мехунка (Utricularia vulgaris; Lentibulariaceae) е разпростра-нена в блата, езера и бавно течащи води от низините да иглолистния пояс. Среща се с поединични екземпляри, рядко. Отрицателно действащи фактори – пресушаването на езерата и блатата, корекциите на речните корита, замър-сяването на вътрешните водоеми с изкуствени торове, пестициди, промишле-ни и битови отпадъци.
Мочурното прозорче (Potentilla palustris; Rosaceae) обитава мочурливи, торфищни места върху преовлажнени планинско-ливадни, торфено-блатни почви от 800 до 2000 m надморска височина. Образува мозаични популации, представени от различни по брой на индивидите групи до единични екземпля-ри. Отрицателно деистващи фактори – отводняването на терените.
Богородичната лъжичка, горчивче (Menyanthes trifoliata; Gentianaceae). Обитава заблатени места и торфища 450–1600 m надморска височина.
Малката ежова главичка (Sparganium minimum; Sparganiaceae) е разпрос-транен в блата, езера в предпланинския и планинския растителен пояс 500–1600 m надморска височина. Расте поединично и на малки групи.
Б) Растителни туфи. Те се развиват върху повалени дървета и първоначално са плаващи. По-
ради обрастване, включително с дървесни видове, отделни участъци се срас-тват с дъното. Интензивно развиващата се растителна маса отмира, дебели-ната на туфите расте (сега е около 2 m).

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1050
При сравняване очертанията на туфите от 1974 г. с тази от 2004 г. се вижда, че хоризонталното нарастване е незначително.
През 1972 г. в периферията на плаващите туфи между езерното дъно и туфите имаше просвет (30-40 cm), който даваше възможност за хоризонтално проникване под туфите до 2-3 m. Сега това е възможно до 0,7 m.
Наблюденията на растителните туфи след 1998 г. показват: - те се развиват хоризонтално, в периферията, по свободната водна по-
върхност на разстояние 3–5 m; - достигат дебелина на слоя 30-50 cm; - поради отмиране на долния слой плаваемостта им става отрицателна
и те потъват. След 1-2 години загниват и се превръщат в тинеста маса; - През 1998 г. чрез сондиране и определяне съдържанието на Cs-137 в
седиментния слой бе направена преценка: този процес води до заблатяване на езерното дъно със скорост около 2 cm/година – доклад на конференцията “Човек, природа, здраве”, Пампорово, 2000 г.
Впоследствие вниманието бе насочено към определяне съдържанието на Cs-137 в самите растителни туфи. Измервания от 2001 г., 2004 г. и 2005 г. по-казват: общото количество на този радионуклид в туфите се запазва практически непроменено във времето.
Измерената от нас повърхностна активност на Cs-137 в растителните туфи е в границите 3 – 8,6 КBq/m2. Тя остава близка до стойността 3,2-13 КBq/m2, докладвана през 1986 г. за съседни райони [9].
Установеното присъствие на този изотоп в най-долните седиментни сле-ве следва да се отнесе към атмосферните отлагания от 1960 –1964 г. Анало-гично разпределение бе получено през 1998 г. и за точка с пясъчно дъно, в близост до южния бряг.
В) Растителни обраствания в свободната от туфи езерна повърх-ност.
След 1998 г., растителността плътно покрива езерото и създава впечат-ление за ливадна площ, въпреки наличието на прозрачна езерна вода под нея, на дълбочина 170 cm. Затова през последните 6 години усилията ни бяха насочени към ограничаване на нейното присъствие.
През 2001 г. на площ от 33 m2. с използуване на елементарна подводна екипировка бе отстранен плаващ ръждавец (Potamogeton natans) (фиг. 1) с биомаса около 460 kg мокро тегло (което съответствува на 0,28 g /cm3 сухо тегло).
Наблюденията след неговото отстраняване в отделни участъци показа-ха,че този растителен вид нараства в хоризонтална посока с около 1метър/година.
ЗАКЛЮЧЕНИЕ Като се основава на: - собствени наблюдения от 1972 г. досега; - моделни експерименти; - литературни данни [10]; - консултации с “Институт по екология” – БАН – София,
колективът достигна до следното становище.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1051
За да се съхрани тази природна забележителност с разностранна значи-мост и добие вид близък до този преди 50 години е необходимо да се извър-шат следните активни природозащитни мероприятия:
- Механически да се отстрани заблатяващата растителност по цялата южна брегова ивица на дълбочина 0–70 сm.
- Частично да се изкорени растителността в свободната от туфи езерна повърхност, разположена между брега и туфите.
- Периодично да се отстраняват плаващите прораствания на растител-ните туфи, с оглед да се предотврати тяхното потъване.
Считаме за подходящо мащабното осъществяване на тази дейност да бъде проведено от държавни и обществени структури в Смолян, тъй като гра-да разполага с такива кадри.
БЛАГОДАРНОСТИ Изследванията са финансирана от фонд “Научни изследвания” на
Пловдивски университет. ЛИТЕРАТУРА
[1] Rosenberg D., V. Resh. Introductoin to freshwater biomonitoring and bentic macroinvertebrates. In: M. D. Rosenberg and V. H. Resh (eds.). Freshwater biomonitoring and bantic macroinvertebrates. New York: Chapman and Hall, 1997, p. 1-9.
[2] Magurran A. Ecological diversity and its measurement. Princeton: Princeton University Press, 1988, 189.
[3] Вълканов, Ал. Смолянските езера, сп. Рибарски преглед, 1938, кн.7. [4] Воденичаров, Д., И. Киряков. Биометрични изследвания върху видове от
разред Yygnemales в НР България. ІІІ. Род Spirogyra Link. Научни трудове на ПУ ”Паисий Хилендарски”, 1981,19(4), 15-52.
[5] Воденичаров, Д., И.Киряков, Д. Темнискова-Топалова, С. Конаклиева. Хидробиологична характеристика на регионалните станции за биологичен мониторинг “Ропотамо”, “Рожен” и “Стенето”. І-ва Национална конферен-ция по проблемите на биологичния мониторинг, 1987,109-114.
[6] Киряков И. Водорасловата флора на Смолянските езера (Chlorococcales). Известия на института по Ботаника, 1971, 22:97-117.
[7] Киряков И. Принос към водорасловата флора на България. ІV. Научни трудове на ПУ”Паисий Хилендарски”, 1981,19(4): 109-120.
[8] Киряков И., Д. Воденичаров. Таксономичен състав на водораслите в Смо-лянските и Чаирските езера (Родопи пл.). І. Научни трудове на ПУ”Паисий Хилендарски”, 1984, 22(2):57-88.
[9] Маринов, В. Радиоекология на земеделието. С., БалБок, 1995. [10] “Състояние на околната среда 2001” – Смолянска община – РИОСВ,
МОСВ, С., 2001.
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1052
доцент, д-р Агоп Крикор Сренц, катедра "Атомна физика", Пловдивски университет " Паисий Хилендарски", ул. Цар Асен 24, тел. + 359 32 261 421 [email protected] доцент, д-р Христо Георгиев Христов, катедра "Атомна физика", Пловдивски университет " Паисий Хилендарски", ул. "Цар Асен" 24; тел. + 359 32 261 422 [email protected] доцент, д-р, Диана Атанасова Кирин катедра "Агроекология", Аграрен университет – Пловдив, ул. "Менделеев" 12
ПРИЛОЖЕНИЯ
Таблица 1 Видове висши растения във езерните екосистеми
на защитена територия “Смолянски езера” – консервационна значимост и ресурсно значение
Консервационна значимост Ресурсно значениеВидове хидрофити Закон за биологич-
ното разнообразие, Приложение 3
Червена книга на България,
т. І Лечебни растения
Drosera rotundifolia L. (Droseraceae)
+ + +
Menyanthes trifoliata L. (Menyanthaceae)
+ + +
Роtentila palustris L. (Rosaceae)
+ + +
Sparganum minimum L. (Sparganiaceae)
+ +
Lycopodiella inundata L. (Lycopodiaceae)
+ +
Саrex limosa L. (Cyperaceae)
+ +
Utricularia minor L. L. (Lentibulariaceae)
+ +
Utricularia vulgaris L. (Lentibulariaceae)
+
Lemna minor L. (Lemnaceae)
+
Lemna trisulca L. (Lemnaceae)
+

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1053
фиг. 1: Potamogeton natans, Плаващ ръждавец
фиг. 2: Menyanthes trifoliate, Водна детелина

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 1054
SCIENTIFIC PAPERS
ЗДРАВНИ АСПЕКТИ НА ЗАТЛЪСТЯВАНЕТО СРЕД НАСЕЛЕНИЕТО В ГРАД КЪРДЖАЛИ
Жени Стайкова – РИОКОЗ Кърджали Иван Чавдаров – РИОКОЗ Кърджали
Таня Търновска – МУ Пловдив Г. Цолова – НЦООЗ София
HEALTH ASPECTS OF THE FATTENING INTO A POPULATION IN THE TOWN OF KARDZHALI
Jenny Staykova – RIOKOZ Kardzhali Ivan Chavdarov – RIOKOZ Kardzhali
Tanya Tarnovska – MU Plovdiv Galya Tsolova – NCOOZ Sofia
Summary: The fattening is one of the most current up-to-date health problem, be-cause it is a prerequisite for increasing of the diseases and mortality-rate following of cardio-vascular and endocrine systems diseases, liver, pancreas, kidneys e.t.c. The aim of this research is to assess the dependency between hypertonics and over-normal body mass / fattening and lipidous status in a part of the lot attended the blood pressure control room in Kardzhali. It was assessed anthropometric and lipidous status of 102 people – regularly patients of the blood pressure control room. Results: It was found above aver-age levels of Body Mass Index (BMI) in all age groups, excluding women 20 to 44 years old. The laboratory research data shown difference by their nature dislipidomius better presented with women. Conclusion: 1. Over-normal body mass and fattening are among the basic risk factors of the hypertonics to both sexes. 2. Unhealthy model of nutrition (hy-perenergic accept) in women is closer related with hypertonic symptoms, than the men.
Затлъстяването е един от най-актуалните съвременни здравни проблеми
защото е предпоставка за увеличаване на заболеваемостта и смъртността от болести на сърдечно-съдовата и ендокринната системи, черния дроб, панкре-аса, бъбреците и пр. По данни на СЗО, над 1 милиард души в света са с над-нормено тегло, различна степен на затлъстяване и съпътстващите ги заболя-вания. Дори в страни, в които поради различни причини населението е с по-ниско тегло, се отбелязва тенденция към увеличена честота на затлъстяване (1). Отчита се, че в периода 1970-2000 г. децата с наднормено тегло, затлъс-тяване и метаболитен синдром в САЩ са утроили броя си (2). Днес приблизи-телно всеки седми от осем американци е с наднормено тегло. Последствията от затлъстяването са втората по значимост причина (след тютюнопушенето) за преждевременната смърт в САЩ (3).

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1055
В България, относителният дял на жените със затлъстяване е приблизи-телно 35 %, на мъжете – 25 %, на децата и юношите – 10-12 %. Високосте-пенното затлъстяване е два пъти по-често сред жените, в сравнение с мъжете (4).
Високата честота на заболяванията свързани със затлъстяването, както и тежките им здравни последствия от тях определят огромната социално-икономическа значимост на проблема. Затова са разработени, популяризират се и се изпълняват различни профилактични мероприятия. Населението на гр. Кърджали е сред участниците в интервенционната здравна програма СИНДИ, един от основните акценти, на която са здравните аспекти на затлъстяването.
Целта на това проучване е да се оцени зависимостта между хипертония-та и наднормената телесна маса/затлъстяването и липидния статус на част от контингента посещаващ кабинета за контрол на кръвното налягане в гр. Кърджали.
МАТЕРИАЛ И МЕТОДИ През м. февруари 2006 г. бяха измерени ръста, телесната маса, кръв-
ното налягане и беше определен индексът на телесната маса (ИТМ) на 102 души (66 жени и 36 мъже), системно посещаващи кабинета за контрол на кръвното налягане. На същите беше изследвани холестерол (HDL, LDL и VDL) и триглицериди. Данните са обработени със статистически пакет SPSS 11.
РЕЗУЛТАТИ И ОБСЪЖДАНЕ Изследваните показатели са представени в таблица № 1. Данните показват наднормени средни нива на ИТМ (тегло (kg) / ръст (m2)
>25) във всички възрастови групи с изключение на жените между 20 и 44 го-дини. Дори и в тази група обаче максималната стойност достига 31.64. От друга страна е очевидно, че сред изследваните липсват участници с високос-тепенно затлъстяване (ИТМ≥40).
Тревожен е фактът, че относителният дял на лицата с наднормена те-лесна маса в млада възраст е голям: 25-34 г. – 32,5 %; 35-44 г. – 55,1 %. Той е изключително висок във възрастовата група 55-64 г., съответно 76 % (жени) и 77 % (мъже). Като най-чести причини за наднорменото тегло се изтъкват хи-перергийният хранителен прием и ниската двигателна активност. Конкретно проучване за храненето на изследвания контингент не е правено, но тематич-ните проверки на РИОКОЗ върху храненето на различни организирани колек-тиви в града (непубликувани данни) показват, че в повечето случаи преобла-дават хиперергийни менюта за сметка предимно на мазнини и въглехидрати. Последните се внасят главно с традиционните за населението в района тес-тени и захарни изделия, животински и растителни мазнини. В същото време анамнестичните данни показват липса или минимална физическа активност при почти всички участници в изследването. Така двата основни фактора – хиперергийно хранене и ниска двигателна активност предпоставят високите средни нива на кръвно налягане, установени във всички възрастово-полови групи, с изключение на жените между 20 и 44 години, въпреки че и при тях максималните стойности са по-високи от референтните. Едновременно с това за хроничен дистрес, породен от тежкия им социално-икономически статус съобщават 80 % от изследваните. Не трябва да се пренебрегва сериозното и

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1056
многогодишно замърсяване на града с оловни аерозоли, което по литератур-ни данни би могло да бъде етиологично свързано с хипертонични прояви (5,6). Установиха се и единични случаи на бъбречни заболявания, които биха могли да бъдат основна причина за хипертонията.
Данните от лабораторните изследвания показаха различни по характер дислипидемии (табл. № 2).
Както се вижда, при жените отклоненията от референтните стойности са най-характерни за групата над 60 години; относителният им дял е 79%. Ана-логични са данните и за LDL-холестерола в тази група (56%); във възрастова-та група 20-44 г. данните са сравнително благоприятни – само 16% имат по-вишено ниво на общия холестерол и само 6,1% са с повишени нива на LDL-холестерол. При мъжете отклоненията от референтните стойности за общия холестерол и в трите възрастови групи е висок (от 50 до 67%), като във втора-та група (45-59 г.), той достига до 67%; при повече от половината (50,8%) от участниците в тази група отчитаме гранични или по-високи спрямо референт-ните нива на липопротеини с малко относително тегло (LDL).
Приложеният корелационен анализ установи права, положителна и дос-товерна зависимост между ИТМ и нивата на кръвно налягане (подчертано за систоличното) и при двата пола: систолично RR – жени r=0.62, p<0.00001, мъ-же r=0.56, p<0.00001; диастолично RR – жени r=0.51, p<0.00001, мъже r=0.31, p<0.07. Зависимостите между нивата на кръвното налягане и холестерола (HDL, LDL и VLDL) и между нивата на кръвното налягане и триглицеридите не са убедителни при мъжете:
RRsyst-Chol – r=-0.12, p=0.48; RRsyst-HDL – r=0.073, p=0.67; RRsyst-LDL – r=0.11, p=0.53; RRsyst-VLDL – r=0.067, p=0.71; RRsyst-Триглицериди – r=0.14, p=0.40.
При жените обаче, макар и с не много високи корелационни коефициенти те са с висока статистическа достоверност:
RRsyst-Chol – r=0.374, p<0.00001; RRsyst-HDL – r=-0.22, p=0.028; RRsyst-LDL – r=0.362, p<0.00001; RRsyst-VLDL – r=0.372, p<0.00001; RRsyst-Триглицериди – r=0.385, p<0.00001.
Това според нас показва, че докато при жените моделът на хранене, свързан вероятно с хиперергиен прием (мазнини, моно- и дизахариди) има водещо значение за хипертонията, то при мъжете тя вероятно е свързана с другите етиологични фактори.
ЗАКЛЮЧЕНИЕ: 1. Наднормената телесна маса и затлъстяването са сред основните рис-
кови фактори за хипертонията и при двата пола. 2. Нездравословният модел на хранене (хиперергиен прием) при жените
е по-тясно свързан с хипертоничните прояви, отколкото при мъжете.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1057
ЛИТЕРАТУРА: [1] Глобално затлъстяване. Медицина, 10.01.2005 г., http://www.martesbg.net/scilog.php [2] Harrell J. S., A. Jessup, N. Greene. Changing our future. Obesity and the
metabolic syndrome in children and adolescents. Journal of cardiovascular nursing, 21, 2006, 4, 322-330.
[3] American Heart Association. Heart disease and stroke statistics-2005 update. American Heart Association, TX, USA, 2004.
[4] Байкова Д. Затлъстяване. http://www.howtoeat.info/tema.asp?razdel=9&temaN=47. [5] Wedeen RP. Bone lead, hypertension, and lead nephropathy. Environ Health
Perspect., 1988, 78, 57-60. [6] Victery W. Evidence for effects of chronic lead exposure on blood pressure in
experimental animals: an overview. Environ Health Perspect, 1988, 78, 71–76.
д-р Жени Стайкова Николова гр. Кърджали, ул. ”ген. Вл. Стойчев” 2, РИОКОЗ тел.: 0361 / 60 297 e-mail: [email protected]
д-р Иван Вълков Чавдаров гр. Кърджали, ул. ”ген. Вл. Стойчев” 2, РИОКОЗ тел.: 0361 / 60 278 e-mail: [email protected]
д-р Таня Христова Търновска доц. д.м. гр. Пловдив МУ тел.: 0886 838 989 e-mail: [email protected]
д-р Галя Крумова Цолова д.м. гр. София НЦООЗ тел.: 02 / 58 12 201
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1058
Таблица № 1
Антропометрични показатели и кръвно налягане
Жени
Възраст Статистическипоказатели
20-44
Ръст Телесна маса ИТМ RR сист. RR ди-аст.
Х 36.19 1.66 62.62 22.72 116.15 76.35 SE 1.11 0.01 2.13 0.67 2.72 1.61 SD 5.66 0.07 10.86 3.42 13.88 8.19
Minimum 25.00 1.56 45.00 17.42 90.00 60.00 Maximum 44.00 1.80 85.00 31.64 150.00 90.00
N 26 26 26 26 26 26 45-59 Х 50.40 1.63 68.03 25.69 129.00 82.33
SE 0.76 0.01 1.84 0.57 3.30 1.64 SD 4.16 0.05 10.07 3.14 18.07 8.98
Minimum 44.00 1.55 53.00 20.96 90.00 60.00 Maximum 58.00 1.75 85.00 31.99 160.00 100.00
n 30 30 30 30 30 30 60+ Х 64.10 1.62 74.30 28.52 149.00 87.00
SE 0.89 0.01 2.63 1.06 2.56 1.33 SD 2.81 0.04 8.31 3.35 8.10 4.22
Minimum 60.00 1.56 64.00 23.53 135.00 80.00 Maximum 68.00 1.70 87.00 34.45 160.00 90.00
n 10 10 10 10 10 10
Мъже 20-44 Х 38.25 1.74 79.42 26.15 129.58 82.92
SE 1.92 0.02 1.88 0.65 3.11 1.99 SD 6.63 0.07 6.53 2.27 10.76 6.89
Minimum 30.00 1.65 70.00 21.97 115.00 75.00 Maximum 52.00 1.86 90.00 29.41 150.00 100.00
n 12 12 12 12 12 12 45-59 Х 51.91 1.73 85.50 28.44 135.45 82.95
SE 1.01 0.02 1.53 0.42 3.56 1.63 SD 4.75 0.08 7.20 1.96 16.68 7.66
Minimum 40.00 1.58 73.00 25.06 90.00 60.00 Maximum 59.00 1.87 95.00 32.05 170.00 95.00
n 22 22 22 22 22 22 60+ 1 63.00 1.70 86.00 29.76 150.00 100.00 2 74.00 1.76 87.00 28.09 140.00 90.00 Х 68.50 1.73 86.50 28.92 145.00 95.00
SE 5.50 0.03 0.50 0.84 5.00 5.00 SD 7.78 0.04 0.71 1.18 7.07 7.07
Minimum 63.00 1.70 86.00 28.09 140.00 90.00 Maximum 74.00 1.76 87.00 29.76 150.00 100.00
n 2 2 2 2 2 2
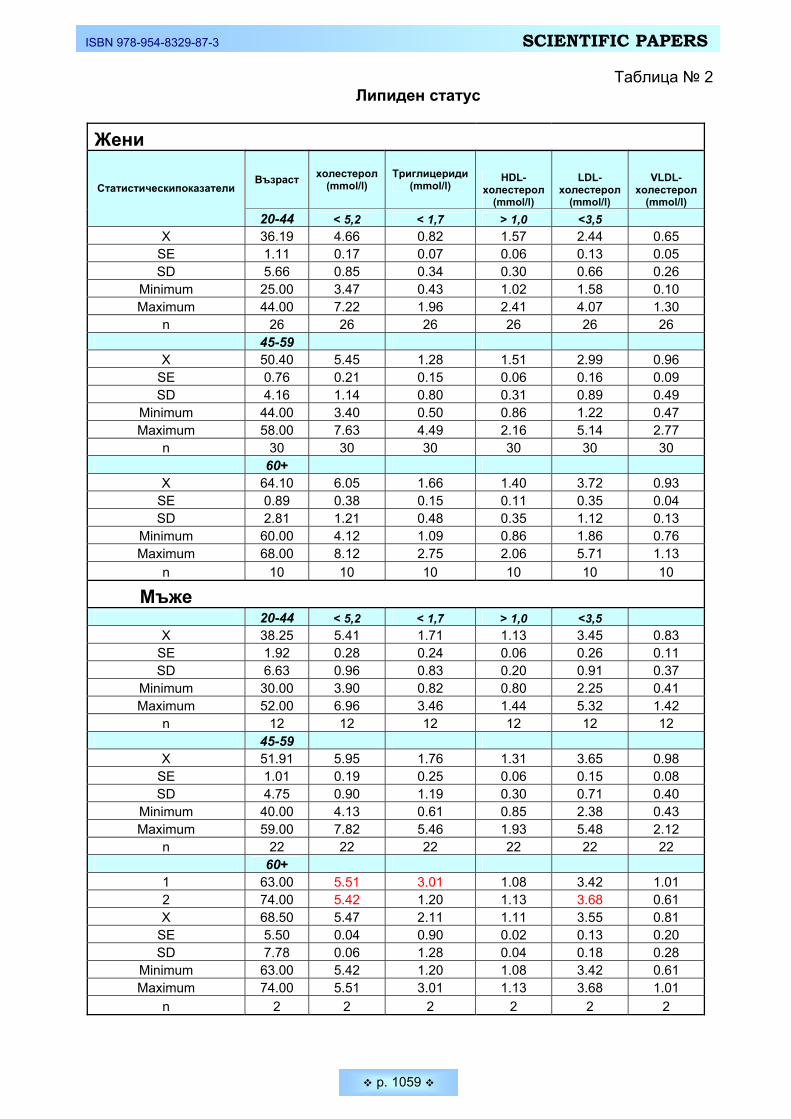
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1059
Таблица № 2 Липиден статус
Жени
Възраст холестерол (mmol/l)
Триглицериди (mmol/l)
HDL-холестерол
(mmol/l)
LDL-холестерол
(mmol/l)
VLDL-холестерол
(mmol/l) Статистическипоказатели
20-44 < 5,2 < 1,7 > 1,0 <3,5 Х 36.19 4.66 0.82 1.57 2.44 0.65
SE 1.11 0.17 0.07 0.06 0.13 0.05 SD 5.66 0.85 0.34 0.30 0.66 0.26
Minimum 25.00 3.47 0.43 1.02 1.58 0.10 Maximum 44.00 7.22 1.96 2.41 4.07 1.30
n 26 26 26 26 26 26 45-59 Х 50.40 5.45 1.28 1.51 2.99 0.96
SE 0.76 0.21 0.15 0.06 0.16 0.09 SD 4.16 1.14 0.80 0.31 0.89 0.49
Minimum 44.00 3.40 0.50 0.86 1.22 0.47 Maximum 58.00 7.63 4.49 2.16 5.14 2.77
n 30 30 30 30 30 30 60+ Х 64.10 6.05 1.66 1.40 3.72 0.93
SE 0.89 0.38 0.15 0.11 0.35 0.04 SD 2.81 1.21 0.48 0.35 1.12 0.13
Minimum 60.00 4.12 1.09 0.86 1.86 0.76 Maximum 68.00 8.12 2.75 2.06 5.71 1.13
n 10 10 10 10 10 10
Мъже 20-44 < 5,2 < 1,7 > 1,0 <3,5 Х 38.25 5.41 1.71 1.13 3.45 0.83
SE 1.92 0.28 0.24 0.06 0.26 0.11 SD 6.63 0.96 0.83 0.20 0.91 0.37
Minimum 30.00 3.90 0.82 0.80 2.25 0.41 Maximum 52.00 6.96 3.46 1.44 5.32 1.42
n 12 12 12 12 12 12 45-59 Х 51.91 5.95 1.76 1.31 3.65 0.98
SE 1.01 0.19 0.25 0.06 0.15 0.08 SD 4.75 0.90 1.19 0.30 0.71 0.40
Minimum 40.00 4.13 0.61 0.85 2.38 0.43 Maximum 59.00 7.82 5.46 1.93 5.48 2.12
n 22 22 22 22 22 22 60+ 1 63.00 5.51 3.01 1.08 3.42 1.01 2 74.00 5.42 1.20 1.13 3.68 0.61 Х 68.50 5.47 2.11 1.11 3.55 0.81
SE 5.50 0.04 0.90 0.02 0.13 0.20 SD 7.78 0.06 1.28 0.04 0.18 0.28
Minimum 63.00 5.42 1.20 1.08 3.42 0.61 Maximum 74.00 5.51 3.01 1.13 3.68 1.01
n 2 2 2 2 2 2

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 1060
SCIENTIFIC PAPERS
САНАТОРИАЛНО ЛЕЧЕНИЕ НА БОЛНИ ОТ АНКИЛОЗИРАЩ СПОНДИЛАРТРИТ
В КУРОРТА ВЕЛИНГРАД
Иван Ст. Станоев – MD.PHD Военна болница – Пловдив
SANATORIUМ TREATMENT OF PATIENTS WITH ANKYLOSING SPONDYLOARTHRITIS
IN VELINGRAD
Ivan Stanoev MD.PHD Militari Hospital – Plovdiv , Bulgaria
ABSTRACT The ankylosing spondiloarthritis complicates with ankylosis, deformation and diability Still the effective therapy is under investigation. Namely the sanatorium therapy regimen in the resort of Velingrad occupies a sub-
stantial role in the treatment of such patients. 24 patients suffering from ankylosing spondyloarthritis were subjected to the follow-
ing therapeutical scheme, under-water jet massage with mineral water from spring #10 from the resort Velingrad (hydrocarbonate-sulphate-alkaline-sodium with fluoride content), local paraffin therapy,paravertebral ultrasound therapy,suberythemal exposure to ultravio-let ray therapy,kinezistherapy.
All patients were with complete clinical and radiological examinations. The therapeutic scheme lasted 20 days independently of the weather conditions. The following indices were checked in due course of therapy: - clinical and anamnesis; - clinical and functional indices: centimeter, Tom-Mayer test, Forestie test, Ott;
Schober, Menel, flexibility. - haematological indices – SUE, leukocytes, red blood cells, Hb, urine profile. Results: 83.50 % of the patients were discharged with a good recovery and 16.50 % were with
no signs of improvement. 1. Patients with ankylosing spondyloarthritis achieve a good recovery as an effect
of the therapeutic regimen. 2. The physical therapeutic regimen is applied specifically to each case and ac-
cording to the stage of the disease. Key words: ankylosing spondyloarthritis, physiotherapy, balneology

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1061
ВЪВЕДЕНИЕ И ЦЕЛ. Анкилозиращият спондилартрит (АС) е автоимунна, системна, хронично-
възпалителна артропатия, която засяга главно сакроилиачни стави, интервер-тебрални, костовертебрални стави, междупрешлени дискове с прилежащи ли-гаменти и периферни стави на крайници и води до анкилози и деформации. [5, 7]
Проблемът за ефективно лечение на тази болест е актуален и изисква непрекъснато търсене на нови средства и методи за оптимално повишаване на заболяването. [3, 6]
Засяга 0,4 до 1 % от населението, предимно мъже и започва във въз-растта между 16 и 40 години. [5]
В системата на рехабилитациони мероприятия при тези болни важно място заема етапа на санаторно – курортно лечение [1, 2] във връзка с това проведохме и проучване за ефективността от балнеолечение и кинезотерапия в санаториални условия в курорта Велинград. [4]
Материали и методика На 24 болни от АС приложихме следният комплекс: 1. Подводно струеви масаж с минерална вода от извор № 10. С хидрокарбонатно – сулфатно – алкално- натриева с флуорно съдържа-
ние 37 °С, 5-15 мин., 2 атмосфери, 10 броя. 2. Парафинолечение 20 мин. 10 броя. 3. Ултразвук паравертебрално 0.2-0.4 w/cm2 5-10 мин. 10 броя и в зави-
симост от локализацията на процеса. 4. УВЛ (ултравиолетови лъчи). Суберитемни дози –общо облъчване. 5. Кинезитерапия Болните бяха в ранен стадии с минимална до умерена възпалителна ак-
тивност и среден стадии. Всички болни бяха с доказана клиника, съответно рентгенологична и лабораторна находка.
Проучванията са през всички сезони на годината за 20 дни. Резултатите от проведеното лечение бяха проследени по следните пока-
затели: а) Клинико-анамнестични данни: Скованост в седалищната и поясно-кръстната област. Болки в ребрено-
гръбначните стави с опасващ характер, което води до ограничаване амплиту-дата на дихателните движения.
Физическа умора, безапетитие, слабости в долни крайници, при някои болни субаребрална температура.
б) Клинично-функционални показатели: Сантиметрия, Том Майер, Форестие, Отт, Шобер, Менел, подвижност, ви-
тален капацитет, мануално тестуване. в) параклиника: СУЕ, левкоцити, еритроцити, хемоглобин, урина (албу-
мин седимент).
Резултати: По-голяма част от болните получиха значително подобрение от проведе-
ното лечение. Изписани болни с подобрение 83.50 % (20 болни), без промяна 16.50 % (4
болни).

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1062
Подобрението на болните е по-отчетливо в зависимост възрастта. Така например: от 20-год. възраст до 35-год. възраст подобрението е по-добре из-разено при проследяване състоянието на болните установихме, че подобре-нието се изразява в три направления:
а) значително намаляване на болките до отзвучаването им при някои болни (общо 83.50 %)
Общо укрепване на болните, подобрение на апетита, нормализиране на температурата, облекчаване на амплитудата на дихателните движения, по-добрение на общата подвижност на болните и общото самочувствие.
б) Подобряване параметрите на клиникофункционалните изследвания: като различните видове сантиметрия, Витален капацитет, мануално тества-не.(20 болни 85.00 %)
в) подобряване на параклиниките: стремеж за нормализиране стойности-те на СУЕ Р < 0,05. левкоцити р<0.05
Обсъждане : При тези болни добрите резултати се дължат на гледните фактори : - Физикалната терапия е съществен момент от целия комплекс за лече-
ние на АС. (Ултразвук, УВЛ) - Кинезотерапия като основен фактор в този комплекс: активни упражне-
ния за подобряване мускулатурата на гръбначния стълб, за подобряване на подвижността на периферните стави, Релаксиращи прийоми, аналитични уп-ражнения, дихателни упражнения.
Кинезитерапията е индивидуална и с масажни прийоми във връзка със стадия и активността на заболюването.
- Балнеолечението (подводноструеви масаж с минерална вода) повлиява болковия синдром, мускулния спазъм,подобрява трофиката. Балнолечението, не се прилагане при засягане на вътрешни органи и ексудативни явления от страна на ставите.
Заключение Получените резултати ни дават основания да направим следните изводи: 1. Болните от АС се подобряват в 83.50 % от приложеният лечебен комп-
лекс: - подводно струеви масаж с минерална вода от извор №10. - парафинолечение - ултразвук-паравертебрално - УВЛ-суберитемни дози общо облъчване - Кинезитерапия с масажни прийоми. 2. Комплекса се прилага индивидуално според възрастта, стадия на за-
боляването, липсата на засягане на вътрешни органи и общото състояние.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1063
ЛИТЕРАТУРА [1] Гачева, Й., Д. Костадинов. Основи на физикалната терапия. С., М.Ф.,
1971. [2] Дайски, А. Балнеология. С.,М.Ф., 1977. [3] Костадинов, Д., Д. Караколев. Главни балнеометични курорти. С., М.Ф.,
1984. [4] Научна конференция. Резюмета. 1-2 ноември 1990. [5] Рязкова, М., И. Кирова. Физикална терапия. София, 2002, с. 219-22 [6] Справочник за българските курорти. С., М.Ф., 1959. [7] Шейтанов, П. Болести на опорно-двигателния апарат. Във “Вътрешни
Болести”. Том 1. Медиц. и физк., С., 1988, с. 530-532 . Dr. Ivan St. Stanoev 4004, Plovdiv Blvd. “Nikola Vapzarov” № 122 Phone : 032 / 69 71 49

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 1064
SCIENTIFIC PAPERS
ДИПЕПТИДИЛПЕПТИДАЗА ІV – СТРУКТУРА И ФУНКЦИИ. ФЛУОРЕСЦЕНТНИ ХИСТО- И ЦИТОХИМИЧНИ МЕТОДИ
ЗА ЛОКАЛИЗАЦИЯ НА ЕНЗИМА
Надежда Стефанова**, Веселина Москова**, Таня Топузова-Христова**, Ралица Тодорова*, Ивайло Иванов***, Машенка Димитрова*
* Институт по експериментална морфология и антропология с музей, БАН
** Катедра “Цитология, хистология и ембриология”, Биологически факултет, СУ “Св. Климент Охридски”
*** Катедра “Хидробиология и ихтиология”, Биологически факултет, СУ “Св. Климент Охридски”
DIPEPTIDYLPEPTIDASE IV – STRUCTURE AND FUNCTINS. FLUORESCENT HISTO- AND CYTOCHEMICAL METHODS
FOR THE ENZYME LOCALIZATION
Nadejda Stefanova**, Veselina Moskova**, Tanja Topouzova – Hristova**, Ralica Todorova*, Ivailo Ivanov***, Mashenka Dimitrova*
* Institute of experimental morphology and anthropology with museum, BAS ** Department of Cytology, histology and embryology, Sofia University “St. Kl. Ohridski”
*** Department of Hydrobiology and Ichthyology, Sofia University “St. Kl. Ohridski” ABSTRACT: Dipeptidylpeptidase IV (DPP IV, EC 3.4.14.5) is a transmembrane pro-
tease, belonging to the prolyloligopeptidase family. Its enzyme activity is to remove selec-tively N-terminal dipeptides with proline or alanine in the second position. It also interacts with several proteins, as adenosine deaminase, fibronectine and collagen. It takes part in the activation of T-lymphocytes, HIV-infection and the processes of metastasis.
We synthesized new fluorogenic substrate, Gly-L-Pro-6-hidrazido-N-hexyl-1,8-naphthalimide for the in situ localization of the enzyme and used this substrate for detec-tion of DPP IV in different tissues and cell cultures.
Key words: DPP IV, enzyme markers, enzyme hystochemistry, fluorogenic substrates Дипептидил пептидаза ІV е описана за пръв път през 1966 г. в черен дроб
на плъх от V. K. Hopsu-Havu, който я нарича глицил-пролил-нафтиламидаза [1]. По-късно се използва наименованието дипептидил аминопептидаза ІV и е доказана в широк спектър от тъкани при гръбначните. Установено е, че ди-пептидил аминопептидаза ІV е идентична с маркера за Т-клетъчна диферен-циация CD26, а по-късно е доказано, че е идентична и с друга молекула, фун-кционираща в имунната система – аденозин деаминаза свързващия белтък

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1065
(ADAbp) [2]. Съвременното наименование, препоръчано от Комисията по Ен-зимната номенклатура, е дипептидил пептидаза ІV (DPP IV, ЕС 3.4.14.5) [3].
DPP ІV е член на пролил олигопептидазното семейство на сериновите протеази [4]. При човека DPP IV е гликопротеин с маса 110 kDa и се състои от 766 а.к. [5]. DPP ІV е повърхностен мембранен белтък тип ІІ, което означава, че основната част от молекулата ù, включваща С-краят, е насочена към из-вънклетъчното пространство [3]. N-краят формира къс цитоплазмен домен от 6 аминокиселини. Трансмембранният домен представлява хидрофобна спи-рала (остатъци 7-28). Извънклетъчната част е изградена от остатъци 29–766 и нея се различават един силно гликозилиран участък (остатъци 49-289) и един богат на цистеин участък (остатъци 290-510), който съдържа 9 от 12-те цистеи-на в състава на DPP ІV. Тези участъци образуват два домена: N-краен 8-листов β-витловиден домен (остатъци 61–495) и С-краен α/β-хидролазен до-мен – остатъци 39–55 и 497–766. Витловидният домен е свързан от една страна с трансмембранния чрез гъвкав участък, а от друга – с хидролазния, като каталитичната триада на хидролазния домен (Ser630, His740 и Asp708) остава на границата между тях [6, 4]. В нативно състояние мономерите на DPP IV асоциират и формират хомодимери. В това взаимодействие участват и двата извънклетъчни домена. Съществена роля за формирането на диме-рите играе една бримка в С-края на DPP-IV, която е много консервативна сред пролиловите дипептидази. Тя съдържа хистидинов остатък (His750), който има важно значение за поддържането на димерната стабилност [7]. (Фиг. 1)
Първичната секвенция на DPP ІV е консервативна в различните видове, като най-висока степен на идентичност има при С-крайния хидролазен домен [3]. Цистеиновите остатъци в β-витловидния домен също са много консерва-тивни като първите 6 от тях (в позиции 326, 337, 445, 448, 455, 473), както и Cys552 (друга много консервативна позиция) са ключови за формиране на третичната структура и за биологичните параметри на ензима [8].
Съществува и разтворима форма на DPP IV, която е открита в кръвната плазма и други телесни течности, както и свързана с извънклетъчния матрикс. При нея липсват цитоплазменият и трансмембранният домени [2, 3, 6, 9,]. Раз-творимата DPP ІV (при човека) съществува под формата на хомодимер с мо-лекулна маса от 210-290 kDa, но може да формира и високомолекулни комп-лекси с маса 900 kDa [6].
Кристалографската структура на свинската DPP ІV показва, че димериза-цията не е необходима, за да бъде завършена архитектурата на активния център. Образуването на димери и тетрамери, обаче, влияе върху взаимо-действието с други молекули, включително някои субстрати и аденозин деа-миназата (ADA) и е възможно да има значение за образуването на междукле-тъчни контакти [6].
DPPIV се синтезира от рибозоми, свързани с мембраната на зърнестата ендоплазмена мрежа. N-крайната сигнална секвенция, която направлява транслокацията на новосинтезираната полипептидна верига към лумена на ендоплазмената мрежа не се откъсва и функционира като мембранен закот-вящ домен. Процесирането на 100-kDa форма на DPP IV, богата на маноза, до 124-kDa силно гликозилирана зряла форма включва дълъг процес на O-гликозилиране, главно в богатия на Ser/Thr гъвкав стъбловиден участък, който следва непосредствено след мембраннния закотвящ домен [8]. Първичната

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1066
структура на човешката DPP IV съдържа 9 потенциални места за N-гликозилиране: Asp85, Asp92, Asp150, Asp219, Asp229, Asp281, Asp321, Asp520 и Asp685 [5]. Кристалографските изследвания доказват, че във всеки от тези девет аспаргинови остатъка става N-свързано гликозилиране [14]. Има две потенциални места за О-гликозилиране [3]. Чрез различни анализи е до-казано, че N-гликозилирането на DPP IV не оказва влияние върху каталитич-ната ù активност, формирането на хомодимерите и свързването с ADA, но участва в сортирането на белтъка в клетката [12].
Димеризацията на DPP IV става в ендоплазмената мрежа и е необходи-мо условие за транспорта към апарата на Голджи [8]. Установено е, че в ендо-телните и епителните клетки точната локализация на DPP IV е в апикалната мембрана. След зреенето в апарата на Голджи DPP IV се транспортира до апикалната мембрана или директно от транс-Голджи или чрез трансцитоза
през базолатералната мембрана. Механизмите на насочване не са добре изу-чени, но вероятно са клетъчно специфични. В N-крайния цитоплазмен и тран-смембранен домен има сигнал за апикално насочване [3]. В апикалното сор-тиране на човешката чревна дипептидил пептидаза IV участват N-свързаните и O-свързаните въглехидратни остатъци. И двата типа гликозилиране са клю-чови компоненти от сигнала за апикално сортиране [13]. Механизмът на сор-тиране включва свързването на DPP IV с детергент-неразтворими мембранни микродомени, съдържащи холестерол и сфинголипиди (липидни rafts) [13, 2]. Ефективното асоцииране с рафтовете зависи от присъствието на пълния ком-плект N- и O-свързани гликани, затова то протича след като белтъкът добие своята напълно гликозилирана форма [13].
Друга посттранслационна модификация на DPP IV е прибавянето на сиа-лова киселина. Изглежда, че сиализацията на DPP IV е клетъчно специфична: съдържанието на сиалова киселина в DPP IV в кръвоносните съдове и в разт-воримата форма е по-голямо, отколкото в DPP IV от бъбреци и тънко черво. При разтворимата форма сиализацията може би предпазва DPP IV от улавяне от чернодробните рецептори за асиалови гликопротеини и изваждането ù от циркулация [3]. Освен това е доказано, че сиализацията е важна за апикална-та секреция на разтворимата форма на DPP IV. При инхибиране на сиализа-цията в бъбречни и чревни епителни клетки е наблюдавано подтискане на апикалната секреция на DPP IV и усилване на секретирането ù през базола-тералната мембрана [14]. При здрави възрастни хора е установено засилено модифициране на DPP IV със сиалова киселина в мононуклеарни клетки от периферна кръв. Около 80-годишна възраст модифицирането в Т-клетките е от 3 до 5 пъти повече, отколкото при здрави 20-годишни индивиди. Тази раз-лика може би е свързана с нарушената имунокомпетентност. Хиперсиализа-цията на DPP IV в Т-лимфоцитите е още по-голяма при HIV-позитивни паци-енти със СПИН.
DPP ІV се експресира в много видове тъкани при бозайниците. Чрез изо-електрично фокусиране е установена голяма молекулна хетерогенност на DPP IV, но е открита само един тип иРНК. Това показва, че хетерогенността на молекулните форми се дължи на посттранслационните модификации на DPP IV [3].
Въпреки че естествените субстрати на DPP IV не са дефинирани оконча-телно, съществува почти безкраен списък от теоретично възможни субстрати. Сравняването на N-крайните секвенции на различни пептиди и белтъци по-

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1067
казва, че значителен брой цитокини, невропептиди и растежни фактори при-тежават консервативен X-Pro мотив в N-края си. Освен че има значение за вторичната структура и следователно в много случаи за биологичната актив-ност на тези молекули, N-крайната Х-Pro секвенция служи и за предпазване от неспецифично протеолитично разграждане, аналогично на С-крайното амиди-ране или ацетилиране [17].
Субстратната специфичност на DPP IV зависи и от свързването ù с други белтъци, които променят достъпността на активния ù център. Поради това различните изоформи на DPP IV могат да имат различна субстратна специ-фичност и е много трудно да се предскаже кои естествени субстрати са пред-почитани от тях [11].
Много от членовете на РАСАР/глюкагоновото семейство са субстрати за DPP IV. В това семейство за биологичната активност са необходими интактни N-краища (Tyr-Ala, His-Ala, His-Ser) и отстраняването им от DPP IV води до инактивиране [3]. При хора времето на полуживот на интактния глюкагон (1-29) в кръвта е 5-6 мин. Разграждането му в кръвната плазма е незначително. Още е спорно кои тъкани са отговорни за изваждането на глюкагона от цирку-лация, но изглежда, че бъбреците играят основна роля и вероятно DPP ІV е главният ензим, участващ в този процес [18].
Хемокините са семейство от малки цитокини със свойства на левкоцитни хемоатрактанти и активатори. Те играят фундаментална роля в развитието, хомеостазата и функционирането на имунната система. В N-краищата на го-лям брой хемокини са установени Pro или Ala във втора позиция, коeто ги прави подходящи субстрати за DPP IV. Ограниченото скъсяване на NH2-края на хемокините може да увеличи или да намали тяхната биологична активност [15]. Процесирането от DPP IV води до подтискане на провъзпалителните свойства на някои хемокини (еотаксин, MDC - макрофагиален хемокин и SDF-1 - стромално-клетъчен фактор-1) или до увеличаване на хемотаксичната ак-тивност (LD78β) [19].
Субстрат за DPP ІV е и аденозин деаминазата (ADA; EC 3.5.4.4), която е повсеместно разпространен разтворим глобуларен ензим, който се среща главно в цитозола и в серума. В много тъкани тя се открива и асоциирана с клетъчната мембрана, благодарение на взаимодейсто си с DPP IV. ADA ката-лизира необратимото дезаминиране на аденозина и 2`-дезоксиаденозина съ-ответно до инозин и 2`-дезоксиинозин. Недостигът ù при хората води до тежка комбинирана имунонедостатъчност (SCID), свързана с натрупване на токсич-ни съединения от пуриновия метаболизъм в клетките. Има предположение, че свързването на DPP ІV с ADA е механизъм за регулиране на клетъчната адхе-зия [10]. Експерименти с лимфоцити и епителни клетки показват, че прикреп-ването на голям брой Т-клетъчни линии зависи силно от DPP IV. Стимулира-нето на свързването на DPP IV с ADА чрез прибавяне на екзогенна ADA зна-чително редуцира адхезията [3].
DPP IV участва в процеса на Т-клетъчното активиране по няколко начина. Чрез ензимното си действие тя променя активността на някои хемокини, учас-тващи в активирането (например RANTES). Освен това DPP IV противодейст-ва на подтискането на пролиферацията от извънклетъчния аденозин чрез асоцииране на разтворимата аденозин деаминаза към повърхността на клет-ките. Освен всичко това DPP IV функционира като сигнален трансдуктор и ко-стимулаторна молекула при процеса на Т-клетъчна активация [20].

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1068
DPPIV участва в модулирането на Na+/H+ обмен в клетките на проксимал-ните бъбречни каналчета [44]. Установено е, че DPP ІV формира комплекс с една от изоформите на Na+/H+ ексчейнджъра и инхибирането на DPP ІV в клетките от проксималните бъбречни каналчета понижава активността и [21].
Установено е, че DPP IV може да свързва фибронектин [22]. Афинитетът на свързване е висок за фибронектина, асоцииран с повърхността на клетки-те, но не и за разтворения в плазмата. Адхезията на DPP IV с фибронектин играе основна роля в колонизирането на белия дроб от ракови клетки, прене-сени по кръвен път, например клетки от карцинома на млечната жлеза [23]. Поради адхезията на DPP ІV от повърхността на ендотелните клетки с фиб-ронектина по повърхността на раковите клетки, те се задържат в кръвоносни-те съдове на белия дроб [23,24].
В миши фибробласти и човешки Т-клетки е установено, че DPP IV функ-ционира и като рецептор за колагени тип І и ІІІ посредством богатия на цисте-ин β-витловиден домен.
DPPIV участва в развитието на някои неоплазии. Тя се свръхекспресира при някои агресивни T-клетъчни малигнени трансформации, хронична В-лимфоцитна левкемия и тироидна карцинома [25]. От друга страна експресия-та и чувствително намалява или даже напълно изчезва при тумори и клетъчни линии, произхождащи от епителните тъкани на белия дроб, черния дроб, де-белото черво и простатата, както и при меланомни клетки. Загуба на DPP IV-активност се наблюдава и при in vitro трансформацията на човешки мелано-цити и фибробласти. DPP IV-активността в серума е значително понижена при пациенти с рак на устната кухина, стомаха, панкреаса и дебелото черво [26].
Високата експресия на DPP IV е приета като маркер за някои по-агресивни Т-клетъчни лимфоми, както и за хронична В-лимфоцитна левкемия [27]. Това може да се обясни с факта, че DPP IV свързва ADА, отговорен за разграждането на аденозина и дезоксиинозина, което като цяло подтиска функциите на лимфоцитите. Раковите клетки при хронична лимфоцитна лев-кемия експресират високи нива на ADA и DPP IV, което може би им помага да преодолеят инхибиторния ефект на извънклетъчния аденозин. Ензимната ак-тивност на DPP IV не е задължително условие при това явление [28].
При някои видове Т-клетъчни левкемии обаче експресията и намалява (например остра Т-клетъчна лимфобластична левкемия, хронична Т-лимфоцитна левкемия и др.). DPP IV е предложена като специфичен маркер за диференциран тироиден карцином, тъй като здравата тироидна тъкан е DPP IV – негативна. Също така въпреки че обикновено DPP IV не е предста-вена нито върху нормалния, нито върху трансформирания епител на дебело-то черво, в някои от туморите му тя е извънредно експресирана [3].
Освен като маркер за някои малигнени трансформации (аденокарцином на щитовидната жлеза, агресивни Т-клетъчни лимфоми и др.) се предполага, че DPP IV може да служи и като маркер за специфичен Т-клетъчен отговор (ТН1 или ТН2) при автоимунни и възпалителни заболявания като ревматоиден артрит и мултитиплена склероза. При ревматоиден артрит DPP IV е свръхекс-пресирана в Т-лимфоцити от периферна кръв, докато нивото на разтворимата DPP IV е значително намалено и тази редукция е свързана с активността на болестта. При мултитиплена склероза също се наблюдават повишени нива на DPP IV-активност в серума, които корелират с тежестта на заболяването [27].

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1069
Намаляване на серумната DPP IV-активност се наблюдава при пациенти, страдащи от възпаления на червата. Това би могло да се използва при прос-ледяването на пациентите, за да се прецизира диагностиката им. Наскоро е доказано, че наднорменото тегло е свързано с увеличена активност на DPP IV [27]. Нивата на DPP IV са понижени при възрастни затлъстели пациенти с ди-абет. При пациенти с хроничен хепатит С серумните нива на DPP IV-активност са значително по-високи отколкото при здрави индивиди и намаляват при па-циенти, повлияващи се от терапията с интерферон [3].
Хисто- и цитохимичното локализиране на активността на DPP IV се про-вежда с помощта на хромогенни методи. Единственият известен досега флу-оресцентен метод се основава на субстрат Gly-L-Pro-амид на 4-метокси-2-нафтиламин и 5-нитро-салицилов алдехид като спомагателен реагент. Този метод не е възприет за рутинна употреба, тъй като има редица недостатъци – делокализация и грубо кристална структура на крайния флуорохром, невъз-можност активността на ензима да се визуализира на местата, където той е асоцииран с капилярен ендотел и др. [30].
В началото на 2006 г. ние синтезирахме нов флуорогенен субстрат за дипептидил пептидаза ІV: Gly-L-Pro-6-хидразидо-N-хексил-1,8-нафталимид (фиг. 2), на базата на който разработихме нов флуоресцентен метод за доказ-ване на ензима в хистологични срези [29]. N-хексил-6-хидразино-1,8-нафталимидът (HHNI) е флуорхром с максимум на поглъщане 430 нм и мак-симум на флуоресценция 540 нм. Чрез хидразидна връзка той е свързан с ди-пептида глицил-L-пролин. Съединението Gly-L-Pro-HHNI е подходящ субстрат за DPP IV, тъй като отговаря на стеричните изисквания на активния и център: L-Pro в позиция Р1, хидрофобен остатък в позиция Р2, транс-конфигурация на връзката Р2-Р1 и обемист незареден остатък в позиция Р1` [4].
Ензимът DPP IV хидролизира хидразидната връзка между дипептида и флуорохрома, при което се освобождава HHNI. Той е слабо разтворим във вода и областта му на флуоресценция съвпада с областта на излъчване на много флуорохроми в биологичните макромолекули. Когато HHNI взаимо-действа с ароматния алдехид пиперонал (3,4-метилендиоксибензалдехид), се получава неразтворим във вода флуоресцентен хидразон с максимум на пог-лъщане 540 нм и максимум на флуоресценция 620 нм, който не съвпада с об-ластта на флуоресценция на клетъчните флуорохроми. Този неразтворим продукт се отлага в местата на ензимна активност и може да бъде използван за хистохимична локализация на ензима. На фигура 2 е представен схематич-но принципът на метода.
Разработеният от нас метод е приложен успешно за доказване на актив-ността на DPP IV в хистологични срези от различни органи на плъх: бъбрек, черен дроб, тънко черво, слезка, сърце, бял дроб и др., както и на човешки клетъчни култури (фиг. 3, фиг. 4).
Нашите резултати при изследване на локализацията на DPP IV в човеш-ки белодробни клетъчни линии показаха, че при диплоидната култура Р флуо-ресцентният сигнал на продукта е сравнително еднакъв по интензитет в пове-чето клетки, а разпределението му в клетките е хомогенно (фиг. 4.Б). Точно такава картина би следвало да се очаква, ако DPP IV е равномерно разпреде-лен по мембраната на клетките.
При двете ракови линии обаче интензитета на флуоресценцията на про-дукта варира силно в различните клетки - при някои тя е доста по-силна от

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1070
средната, докато при други изобщо не може да бъде детектирана. Освен това разпределението на сигнала в клетките не е хомогенно, а зърнесто (фиг.4.Г и 4. Е).
Хетерогенността (генетична, морфологична и функционална) е отличи-телна черта на раковата тъкан, както in vivo, така и in vitro. Освен това при ра-ковите клетъчни линии броят на хромозомите варира в различните клетки и модалният брой хромозоми за линиите А549 и SK-MES-1 е съответно 12 и 17-20. Линията SK-MES-1 е класифицирана в групата с най-силно изразена хро-мозомна нестабилност. За линията А549 е установено, че се състои от най-малко два клетъчни субклона, които имат различни функционални характе-ристики (например различна чувствителност към оксиданти). За белодробните тумори има и конкретни изследвания върху възникването на мутации в хромо-зома 2q (където е локусът на гена за DPP IV). Те показват, че при т.нар. “недребно-клетъчни” белодробни карциноми (NSCLCs), към които принадле-жат линиите A549 и SK-MES-1, често става загуба на алели в хромозома 2q, като честотата на такива делеции нараства значително с развитието на тумо-рите и в метастазите. Дори генът за DPP IV да не е засегнат пряко от мутация, експресията му може да бъде повлияна косвено при нарушаването на други функции в клетката.
При А549 интензитетът на флуоресценция в съседните клетки варира силно, а при SK-MES-1 се различават групички от съседни клетки с еднакъв интензитет на флуоресценция. Както личи от фигура 4.Е, клетките от линията SK-MES-1 имат по-силно изразена склонност да образуват контакти със съсе-дите си и да формират плътни групи, отколкото клетките от линията А549, ко-ито вероятно имат и по-голяма подвижност. Предполагаме, че по-слабата подвижност и склонността към формиране на групички при линията SK-MES-1 е причина клетките с моноклонален произход да остават близо една до друга. Дори сред такива групички обаче понякога се наблюдават отделни клетки с променен флуоресцентен интензитет, вероятно поради възникване на спон-танни мутации, дължащи се на голямата хромозомна нестабилност на тази клетъчна линия.
Общо тези данни показват, че наблюдаваната разлика в нивото на DPP IV-активност в отделните клетки при тези две белодробни ракови линии не е изненадваща, а е пряко следствие от малигнеността.
Въз основа на получените от нас резултати смятаме, че предложения от нас нов флуорсцентен субстрат е подходящ за установяване на специфична-та активност на DPP IV, както и за изследване на разпределението на ензима в различни клетъчни типове.
Изследванията в настоящата работата са извършени с финансовата
подкрепа на Фонд “Научни изследвания” към МОН по проект 1527/05.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1071
ЛИТЕРАТУРА: [1] V.K. Hopsu-Havu, George G.Glenner, A New Dipeptide Naphpthylamidase
Hydrolysing Glycyl-Prolyl-β-Naphthylamide, Histochemie 7,197-201 (1966) [2] D.A. Pereira, L. Gomes, M.C. El-Cheikh and R. Borojevic. Dipeptidyl peptidase
IV (CD26) activity in the hematopoietic system: differences between the mem-brane-anchored and the released enzyme activity. Braz J Med Biol Res, Vol-ume 36(5) 567-578, (2003)
[3] Anne-Marie Lambeir, Christine Durinx, Simon Scharpé, Ingrid De Meester, Dipeptidyl-peptidase IV from Bench to Bedside: An Update on Structural Prop-erties, Functions, and Clinical Aspects of the Enzime DPP IV, Crit Rev Clin Lab Sci. 40(3):209-94, (2003)
[4] Kathleen Aertgeerts, Sheng Ye, Mike G. Tennant, Michelle L. Kraus, Joe Rogers, Bi-Ching Sang, Robert J. Skene, David R. Webb and G. Sridhar Prasad. Crystal structure of human dipeptidyl peptidase IV in complex with a decapeptide reveals details on substrate specificity and tetrahedral intermedi-ate formation. Protein Science, 13:412-421, (2004)
[5] Misumi Y,Hayashi Y,Arakawa F,Ikehara Y, Molecular cloning and sequence analysis of human dipeptidyl peptidase IV, a serine proteinase on the cell sur-face., Biochim Biophys Acta, 1131(3):333-6 (1992)
[6] Michael Engel , Torsten Hoffmann, Leona Wagner, Michael Wermann, Ulrich Heiser, Reiner Kiefersauer,, Robert Huber, Wolfram Bode, Hans-Ulrich De-muth , and Hans Brandstetter. The crystal structure of dipeptidyl peptidase IV (CD26) reveals its functional regulation and enzymatic mechanism. PNAS| vol. 100 | no. 9 | 5063-5068, (2003)
[7] Chia-Hui Chien, Li-Hao Huang, Chi-Yuan Chou, Yuan-Shou Chen, Yu-San Han, Gu-Gang Chang, Po-Huang Liang, and Xin Chen, One Site Mutation Dis-rupts Dimer Formation in Human DPP-IV Proteins, J. Biol. Chem., Vol. 279, Is-sue 50, 52338-52345, December 10, 2004
[8] Jörg Dobers, Sabine Grams, Werner Reutter and Hua Fan. Roles of cysteines in rat dipeptidyl peptidase IV/CD26 in processing and proteolytic activity. Eur. J. Biochem. 267, 5093-5100, (2000)
[9] Christine Durinx, Anne-Marie Lambeir, Eugène Bosmans, Jean-Bernard Fal-magne, Raf Berghmans, Achiel Haemers, Simon Scharpé and Ingrid De Meester. Molecular characterization of dipeptidyl peptidase activity in serum. Eur. J. Biochem. 267, 5608-5613, (2000)
[10] Wilhelm A. Weihofen, Jiango Liu, Werner Reutter, Wolfram Saenger, and Hua Fan. Crystal Structure of CD26/Dipeptidyl-peptidase IV in Complex with Adenosine Deaminase Reveals a Highly Amphiphilic Interface. J. Biol. Chem., Vol. 279, Issue 41, 43330-43335, (2004)
[11] Emil Boonacker, Cornelis J. F. Van Noorden, The multifunctional or moonlight-ing protein CD26/DPPIV. European Journal of Cell Biology 82, 53 ± 73 (2003)
[12] Kathleen Aertgeerts, Sheng Ye, Lihong Shi, Sridhar G. Prasad, Darbi Witmer, Ellen Chi, Bi-Ching Sang, Robert A. Wijnands, David R. Webb and Ronald V. Swanson, N-linked glycosylation of dipeptidyl peptidase IV (CD26): Effects on enzyme activity, homodimer formation, and adenosine deaminase binding. Protein Science, 13:145-154 (2004)
[13] Marwan Alfalah, Ralf Jacob, and Hassan Y. Naim. Intestinal Dipeptidyl Pepti-dase IV Is Efficiently Sorted to the Apical Membrane through the Concerted

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1072
Action of N- and O-Glycans as Well as Association with Lipid Microdomains. J. Biol. Chem., Vol. 277, Issue 12, 10683-10690, (2002)
[14] Slimane TA,Lenoir C,Sapin C,Maurice M,Trugnan G, Apical secretion and sia-lylation of soluble dipeptidyl peptidase IV are two related events. Exp. Cell. Res., 10;258(1):184-94 (2000)
[15] Paul Proost, Ingrid De Meester, Dominique Schols, Sofie Struyf, Anne-Marie Lambeir, Anja Wuyts, Ghislain Opdenakker, Erik De Clercq, Simon Scharpé, and Jo Van Damme. Amino-terminal Truncation of Chemokines by CD26/Dipeptidyl-peptidase IV. J Biol Chem, Vol. 273, Issue 13, 7222-7227, (1998)
[16] Hans-Ulrich Demutha, Christopher H.S. McIntosh, Raymond A. Pederson, Type 2 diabetes—Therapy with dipeptidyl peptidase IV inhibitors. Biochimica et Biophysica Acta 1751, 33 – 44 (2005)
[17] Greet Vanhoof, Filip Goossens, Ingrid De Meester, Dirk Hendriks, Simon Scharpe, Proline Motifs in Peptides snd Their Biological Processing.FASEB J.9:736-44 (1995)
[18] Simon Amadeus Hinke, J. Andrew Pospisilik, Hans-Ulrich Demuth, Susanne Mannhart, Kerstin Kühn-Wache, Torsten Hoffmann, Erica Nishimura, Raymond A. Pederson, and Christopher H.S. McIntosh. Dipeptidyl Peptidase IV (DPIV/CD26) Degradation of Glucagon. J Biol Chem, Vol. 275, Issue 6, 3827-3834, (2000)
[19] Paul Proost, Patricia Menten, Sofie Struyf, Evemie Schutyser, Ingrid De Meester, and Jo Van Damme, Cleavage by CD26/dipeptidyl peptidase IV con-verts the chemokine LD78b into a most efficient monocyte attractant and CCR1 agonist. Blood, VOL 96, NUM 5 (2000)
[20] Hideto Ikushima, Yasuhiko Munakata, Tomonori Ishii, Satoshi Iwata, Masazumi Terashima, Hirotoshi Tanaka, Stuart F. Schlossman, and Chikao Morimoto, Inter-nalization of CD26 by mannose 6-phosphate/insulin-like growth factor II receptor contributes to T cell activation. PNAS,vol.97,no.15,8439-8444 (2000)
[21] Adriana C. C. Girardi, Brenda C. Degray, Tamas Nagy, Daniel Biemesderfer, and Peter S. Aronson. Association of Na+-H+ Exchanger Isoform NHE3 and Dipeptidyl Peptidase IV in the Renal Proximal Tubule. J. Biol. Chem., Vol. 276, Issue 49, 46671-46677, December 7, (2001)
[22] Piazza GA, Callanan HM, Mowery J, Hixson DC. Evidence for a role of dipep-tidyl peptidase IV in fibronectin-mediated interactions of hepatocytes with ex-tracellular matrix. Biochem J. 262(1):327-34, (1989)
[23] Hung-Chi Cheng, Mossaad Abdel-Ghany, Randolph C. Elble, and Bendicht U. Pauli. Lung Endothelial Dipeptidyl Peptidase IV Promotes Adhesion and Me-tastasis of Rat Breast Cancer Cells via Tumor Cell Surface-associated Fi-bronectin. J Biol Chem, Vol. 273, Issue 37, 24207-24215, (1998)
[24] Hung-Chi Cheng, Mossaad Abdel-Ghany and Bendicht U. Pauli A Novel Con-sensus Motif in Fibronectin Mediates Dipeptidyl Peptidase IV Adhesion and Metastasis. J. Biol. Chem., Vol. 278, Issue 27, 24600-24607, (2003)
[25] Toshiko Yamochi, Tadanori Yamochi, Ugur Aytac, Tsutomu Sato, Kazuya Sato, Kei Ohnuma, Kathryn S. McKee, Chikao Morimoto, Nam H. Dang, Regulation of p38 Phosphorylation and Topoisomerase IIA Expression in the B-Cell Lymphoma Line Jiyoye by CD26/Dipeptidyl Peptidase IV Is Associated with Enhanced In vitro and In vivo Sensitivity to Doxorubicin. Cancer Res, 65: (5), (2005)
[26] Umadevi V. Wsley, Shakuntala Tiwari and Alan N. Houghton, Role for Dipepti-dyl Peptidase IV in Tumor Suppression of Human Non Small Cell Lung Carci-noma Cells. Int. J. Cancer: 109, 855–866 (2004)

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1073
[27] Linus Ho, Ugur Aytac, L. Clifton Stephens, Kei Ohnuma, Gordon B. Mills, Kath-ryn S. McKee, Christopher Neumann, Ruth LaPushin, Fernando Cabanillas, James L. Abbruzzese, Chikao Morimoto, and Nam H. Dang, In Vitro and in Vivo Antitumor Effect of the Anti-CD26 Monoclonal Antibody 1F7 on Human CD301 Anaplastic Large Cell T-Cell Lymphoma Karpas 299. Clinical Cancer Research, 7, 2031–2040 (2001)
[28] Petr Bušek, Radek Mal´ık, Aleksi Šedo, Dipeptidyl peptidase IV activity and/or structure homologues (DASH) and their substrates in cancer. The International Journal of Biochemistry & Cell Biology 36, 408–421 (2004)
[29] R. Todorova, I. Ivanov, M. Dimitrova, Fluorescent histochemical localization of dipeptidyl aminopeptidase IV using a newly developed substrate. Acta Morph. Anthr., 12, 2006 (in press)
[30] Reinhard Gossrau. Cytochemistry of membrane proteases. Histochemical Journal 17, 737-771 (1985)
Надежда Петрова Стефанова София 1164, бул. “Драган Цанков” 8 Биологически факултет, СУ “Св. Кл. Охридски”, лаб. 301 тел. 8167 302 е-mail: [email protected] Веселина Светославова Москова, ст. ас. София 1164, бул. “Драган Цанков” 8 Биологически факултет, СУ “Св. Кл. Охридски”, лаб. 227 тел. 8167 226 е-mail: [email protected] или [email protected] Таня Топузова-Христова, гл. ас. София 1164, бул. “Драган Цанков” 8 Биологически факултет, СУ “Св. Кл. Охридски”, лаб. 227 тел. 8167 226 е-mail: [email protected] Ралица Иванова Тодорова н.с. ІІІ ст. Институт по Експериментална Морфология и Антропология с Музей ул. “Акад. Г. Бончев” бл. 25, 1113 София тел. 9792307 е-mail: [email protected] Ивайло Петров Иванов, доц. д-р София 1164, бул. “Драган Цанков” 8 Биологически факултет, СУ “Св. Кл. Охридски” тел. 8167 214 Машенка Борисова Димитрова, ст.н.с. ІІ ст. д-р Институт по Експериментална Морфология и Антропология с Музей ул. “Акад. Г. Бончев” бл. 25, 1113 София тел. 9792307 е-mail: [email protected]
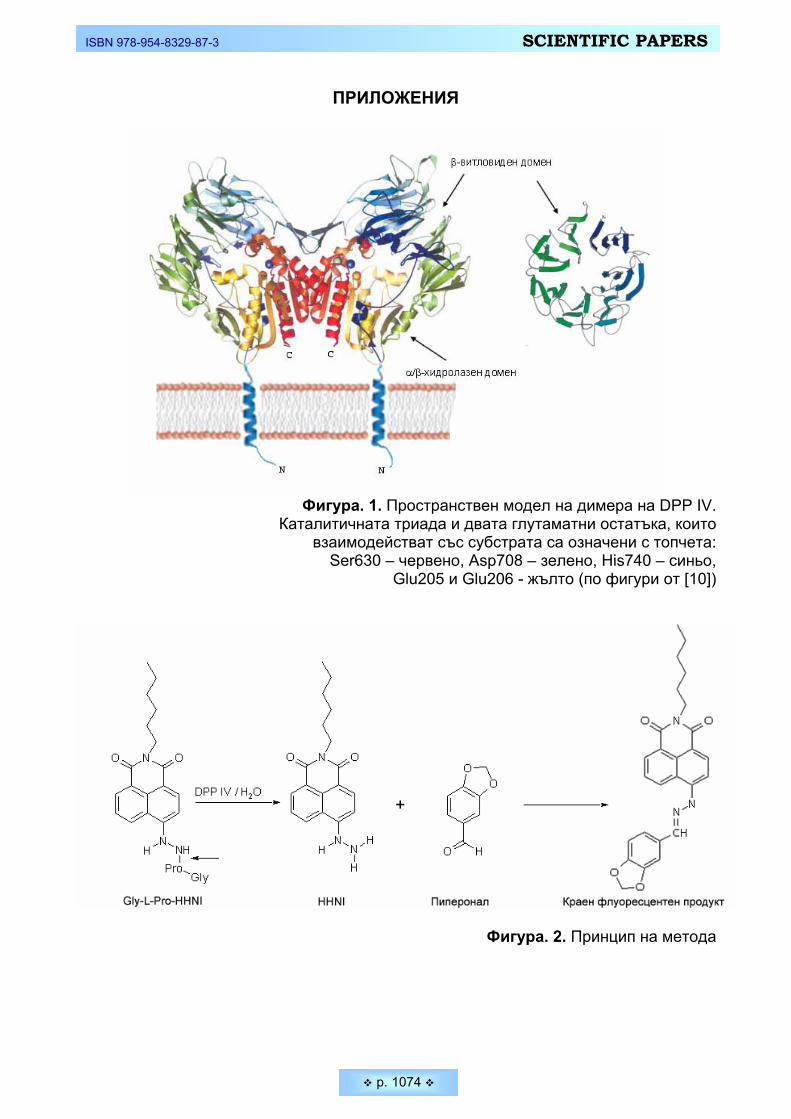
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1074
ПРИЛОЖЕНИЯ
Фигура. 1. Пространствен модел на димера на DPP IV.
Каталитичната триада и двата глутаматни остатъка, които взаимодействат със субстрата са означени с топчета:
Ser630 – червено, Asp708 – зелено, His740 – синьо, Glu205 и Glu206 - жълто (по фигури от [10])
Фигура. 2. Принцип на метода
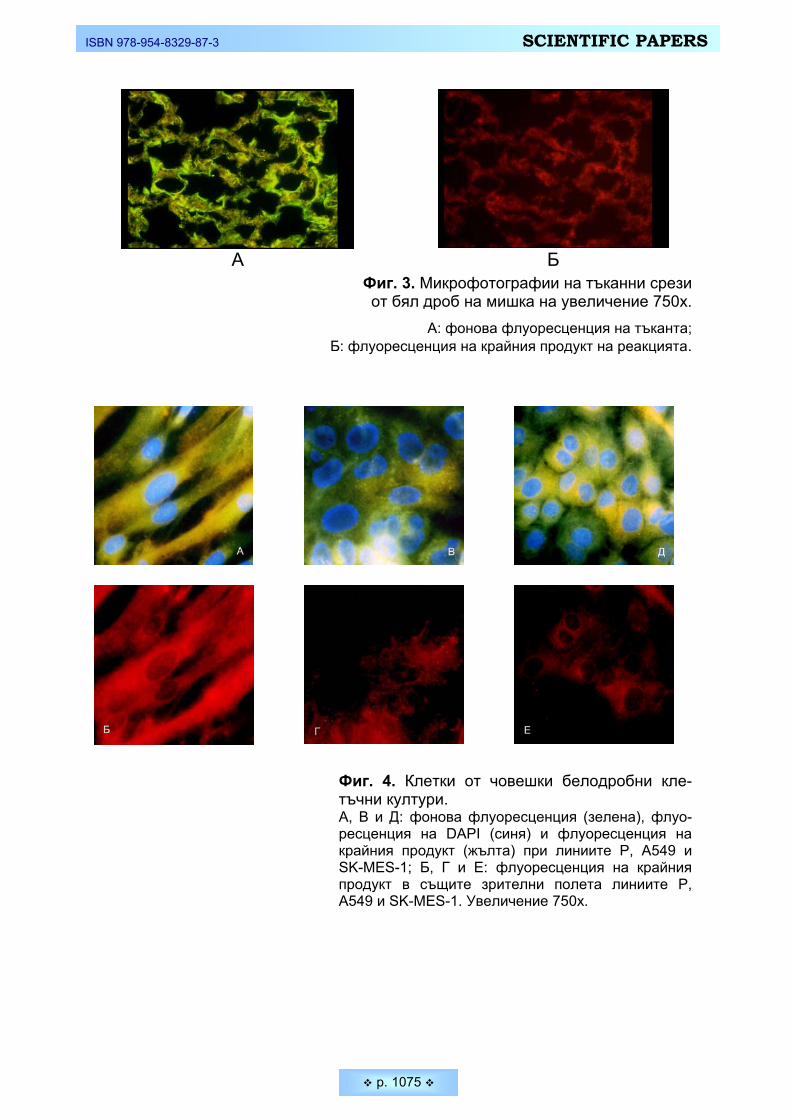
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1075
А Б
Фиг. 3. Микрофотографии на тъканни срези от бял дроб на мишка на увеличение 750х.
А: фонова флуоресценция на тъканта;Б: флуоресценция на крайния продукт на реакцията.
Фиг. 4. Клетки от човешки белодробни кле-тъчни култури. А, В и Д: фонова флуоресценция (зелена), флуо-ресценция на DAPI (синя) и флуоресценция на крайния продукт (жълта) при линиите Р, А549 и SK-MES-1; Б, Г и Е: флуоресценция на крайния продукт в същите зрителни полета линиите Р, А549 и SK-MES-1. Увеличение 750х.

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 1076
SCIENTIFIC PAPERS
СЕКУЛАРНИ ПРОМЕНИ ВЪВ ФИЗИЧЕСКОТО РАЗВИТИЕ НА СМОЛЯНСКИ ПОДРАСТВАЩИ
Рачо Стоев*, Силвия Младенова**
* ИЕМАМ – БАН, София **Филиал на ПУ “ Паисий Хилендарски”– гр. Смолян
SECULAR GROWTH CHANGES IN PHYSICAL DEVELOPMENT OF SMOLYAN ADOLESCENTS
Racho Stoev*, Silvia Mladenova**
* IEMAM-BAS, Sofia, Bulgaria ** Branch Smolyan at Plovdiv University, Smolyan, Bulgaria
ABSTRACT: This paper are present the results of comparison study of secular changes in physical development of schoolchildren of the town of Smolyan, Bulgaria in pe-riod 1986 – 2001. The sample included 169 boys and 192 girls, measured in 1986 (Stoev) and 465 boys and 480 girls, measured in 1997-2001 (Mladenova), all investigated by methods of Martin-Saller(1957). We could compare following somatic parameters: height, weight, BMI, biacromial, bicristal and bitrochanterial diameters, as well as transversal chest diameter. The results are shows secular changes in the physical development of the Smolyan schoolchildren for last 15 years. They аre connect with the stabilization in the growth of the height, biacromial and bitrochanterial diameters, also in transversal chest di-ameter; negative secular changes in weight and BMI, which tendency agree with the world process on gracilization; and positive secular changes in bicristal diameter, which are more pronounced in boys, than in the girls.
Key words: growth, secular changes, physical development, schoolchildren ВЪВЕДЕНИЕ Въпросът за секуларните промени във физическото развитие на децата и
подрастващите е обект на многобройни ауксологични изследвания [1]. У нас тези промени са проследени главно за София и Пловдив, и то предимно до 80-те години [12; 8; 9] на XX век. Откъслечни са изследванията на тези процеси в по-малките населени места, особено след 1990-та година [9].
Последните години на двадесетото столетие се характеризират с резки промени в социално-икономическите и демографски процеси в страната като цяло, а също и в отделните населени места. Наблюдава се спадане на дохо-дите за по-голямата част от населението, при рязко нарастване на социална-та диференциация. От друга страна, се наблюдава и рязко намаляване на раждаемостта. Едно такова намаляване, при общо влошаване на жизнените условия, може да има протективна функция върху физическото развитие на
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1077
децата и подрастващите, което да не се влоши, а напротив - дори да се по-добри [2; 11].
За Смолян специално, както по собствени наблюдения на авторите, така и според социологическите изследвания [5] през последните години е харак-терно значително повишаване на образователно-културното равнище на на-селението и рязко нарастване на грижите за децата и тяхното образование от страна на семейството. Тези социално-демографски промени също противос-тоят на влиянието на пониженото жизненото равнище върху растежа и разви-тието на младото поколение.
На фона на тези противоречиви социални влияния авторите си поставиха за цел да разгледат евентуалните промени във физическото развитие на под-растващите в град Смолян за последните две десетилетия.
МАТЕРИАЛ И МЕТОДИ Проведените по методиката на Martin-Saller [8] трансверзални изследва-
ния през 1986 година обхващат 169 момчета (провел Стоев), а в периода 1999-2001 г. 465 момчета и 480 момичета (провела Младенова). Изследвани-те са на възраст от 9 до 17 години, разпределени в 8 възрастови групи.
В представената работа физическото развитие на двете генерации деца е проследено чрез следните основни антропометрични признаци: ръст , тег-ло, индекса за телесна маса (ВМІ) и напречните костни диаметри на тя-лото (битрохантериален, бикристален, биакромиален и напречен диаме-тър на гръдния кош). Тези признаци са избрани във връзка с минималните различия в методиката на измерване от двамата автори.
Събраните данни са обработени чрез дескриптивен анализ. РЕЗУЛТАТИ И ОБСЪЖДАНЕ Съвременното поколение смолянски подрастващи се различава незначи-
телно по ръст (фиг. 1; 2) от своите връстници от 80-те години. Малко по-висок ръст и при двата пола се наблюдава към края на изследвания период, което би могло да се дължи на по-напредналия стадий на съзряване на съвремен-ните подрастващи [3; 12]. Ако това е причината, терминалният ръст на двете поколения би бил еднакъв. Малките и незначителни изменения в ръста съв-падат с литературните данни за затихването на акцелерационните процеси [1].
Фиг. 1: Ръст – момчета
130
140
150
160
170
180
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
Р(cm
)
X_1986 X_2001
Фиг. 2: Ръст – момичета
130
140
150
160
170
180
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
Р(cm
)
X_1986 X_2001
Основните различия между двете поколения са в телесната маса (фиг. 3; 4), която е намаляла значително в периода между двете изследвания. Най-съществени различия, достигащи до 10 кг и при двата пола се наблюдават
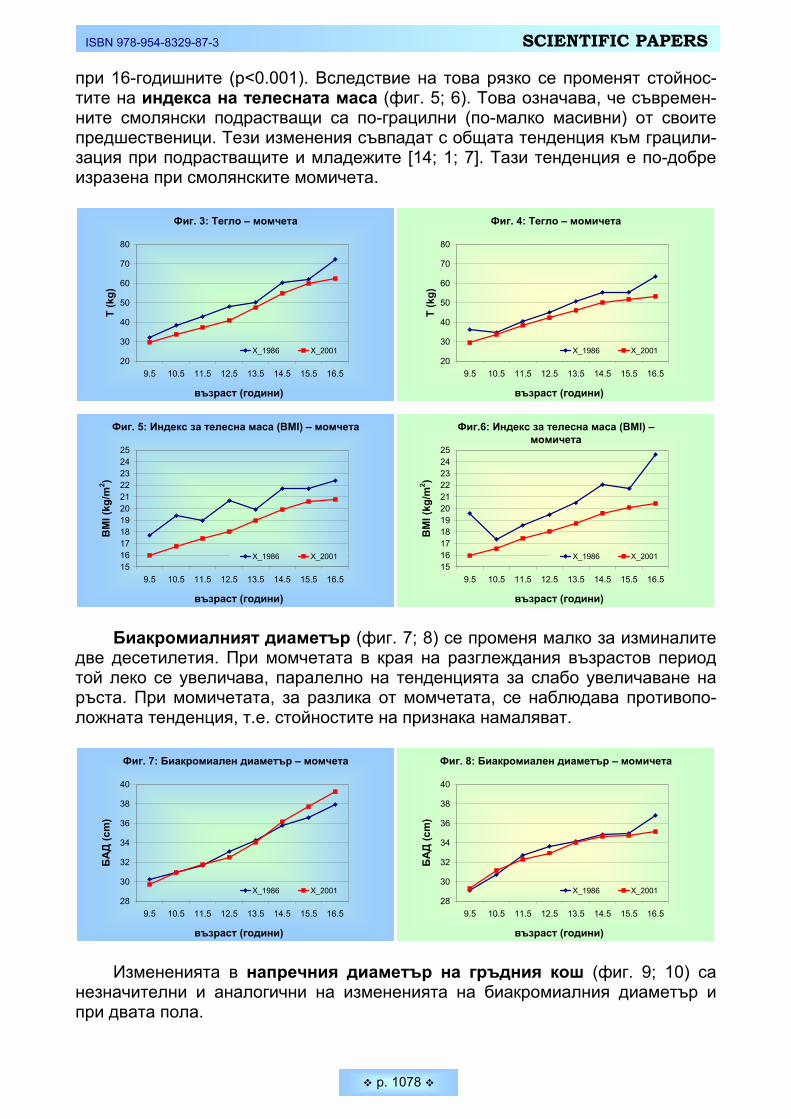
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1078
при 16-годишните (p<0.001). Вследствие на това рязко се променят стойнос-тите на индекса на телесната маса (фиг. 5; 6). Това означава, че съвремен-ните смолянски подрастващи са по-грацилни (по-малко масивни) от своите предшественици. Тези изменения съвпадат с общата тенденция към грацили-зация при подрастващите и младежите [14; 1; 7]. Тази тенденция е по-добре изразена при смолянските момичета.
Фиг. 3: Тегло – момчета
20
30
40
50
60
70
80
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
Т (k
g)
X_1986 X_2001
Фиг. 4: Тегло – момичета
20
30
40
50
60
70
80
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
Т (k
g)
X_1986 X_2001
Фиг. 5: Индекс за телесна маса (BMI) – момчета
1516171819202122232425
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
BM
I (kg
/m2 )
X_1986 X_2001
Фиг.6: Индекс за телесна маса (BMI) – момичета
1516171819202122232425
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
BM
I (kg
/m2 )
X_1986 X_2001
Биакромиалният диаметър (фиг. 7; 8) се променя малко за изминалите две десетилетия. При момчетата в края на разглеждания възрастов период той леко се увеличава, паралелно на тенденцията за слабо увеличаване на ръста. При момичетата, за разлика от момчетата, се наблюдава противопо-ложната тенденция, т.е. стойностите на признака намаляват.
Фиг. 7: Биакромиален диаметър – момчета
28
30
32
34
36
38
40
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
БАД
(сm
)
X_1986 X_2001
Фиг. 8: Биакромиален диаметър – момичета
28
30
32
34
36
38
40
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
БАД
(сm
)
X_1986 X_2001
Измененията в напречния диаметър на гръдния кош (фиг. 9; 10) са незначителни и аналогични на измененията на биакромиалния диаметър и при двата пола.
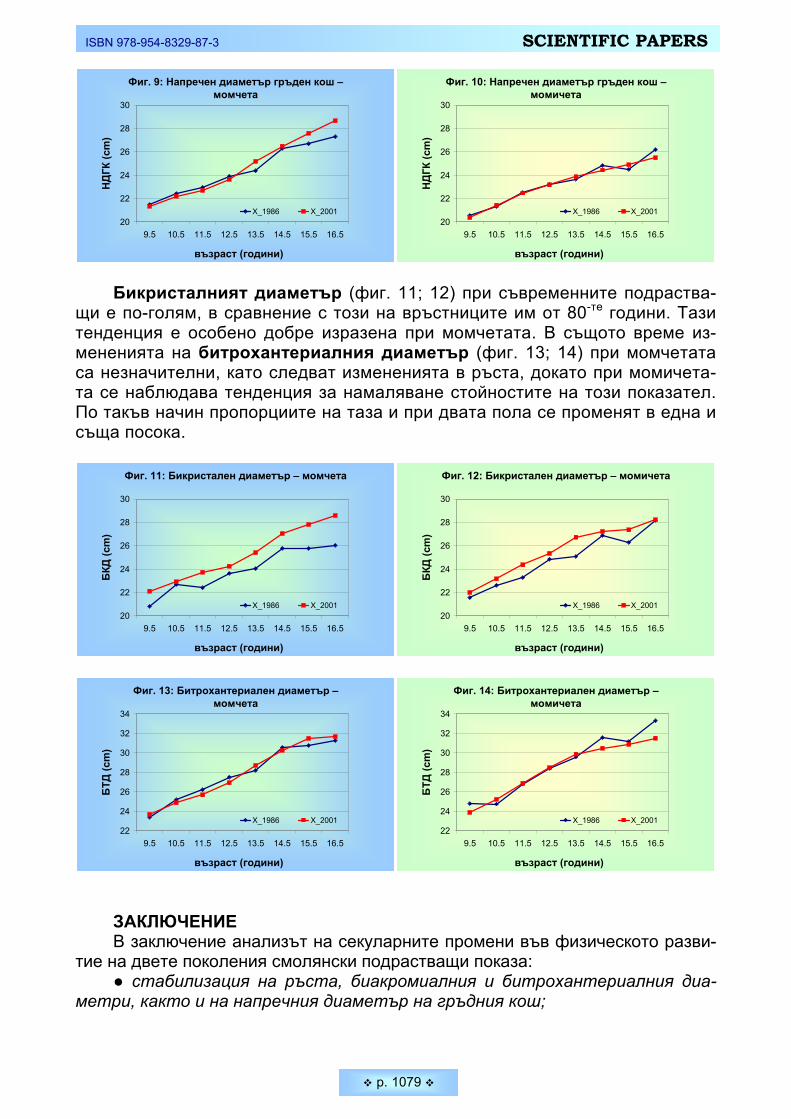
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1079
Фиг. 9: Напречен диаметър гръден кош – момчета
20
22
24
26
28
30
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
НДГК
(сm
)
X_1986 X_2001
Фиг. 10: Напречен диаметър гръден кош – момичета
20
22
24
26
28
30
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
НДГК
(сm
)
X_1986 X_2001
Бикристалният диаметър (фиг. 11; 12) при съвременните подраства-щи е по-голям, в сравнение с този на връстниците им от 80-те години. Тази тенденция е особено добре изразена при момчетата. В същото време из-мененията на битрохантериалния диаметър (фиг. 13; 14) при момчетата са незначителни, като следват измененията в ръста, докато при момичета-та се наблюдава тенденция за намаляване стойностите на този показател. По такъв начин пропорциите на таза и при двата пола се променят в една и съща посока.
Фиг. 11: Бикристален диаметър – момчета
20
22
24
26
28
30
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
БКД
(сm
)
X_1986 X_2001
Фиг. 12: Бикристален диаметър – момичета
20
22
24
26
28
30
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
БКД
(сm
)
X_1986 X_2001
Фиг. 13: Битрохантериален диаметър – момчета
22
24
26
28
30
32
34
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
БТД
(сm
)
X_1986 X_2001
Фиг. 14: Битрохантериален диаметър – момичета
22
24
26
28
30
32
34
9.5 10.5 11.5 12.5 13.5 14.5 15.5 16.5
възраст (години)
БТД
(сm
)
X_1986 X_2001
ЗАКЛЮЧЕНИЕ В заключение анализът на секуларните промени във физическото разви-
тие на двете поколения смолянски подрастващи показа: ● стабилизация на ръста, биакромиалния и битрохантериалния диа-
метри, както и на напречния диаметър на гръдния кош;

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1080
● отрицателни секуларни промени в теглото и индекса на телесната маса, които съвпадат със световната тенденция за грацилизация през последните десетилетия;
● положителни секуларни промени в бикристалния диаметър, особено добре подчертани при момчетата.
Тези разнопосочни промени се наблюдават на фона на ускоряване на половото съзряване с около 0.6 години, което говори за слабата корелация между половото съзряване, физическото развитие и отделните негови показатели, отбелязана и от други автори за последните десетилетия [6].
По такъв начин, въпреки влиянието на противоречивите социално-икономически и демографски фактори, сред подрастващото поколение на сравнително малкия и изолиран град Смолян се наблюдават някои съвремен-ни световни тенденции в растежа и развитието, а именно разнопосочни секу-ларни промени във физическото развитие.
ЛИТЕРАТУРА:
[1] Година, Е.З., 2003. Ауксология человека – наука ХХІ века: проблемы и перспективы. В: Алексеева, Т.И., Балановская, Е.В., Година, Е.З., Дубова, Н.А. (Ред.). Антропология на пороге ІІІ тысячелетия, т.2, Москва, Российское отделение ЕАА, стр. 529-566.
[2] Миронов, Б.И., 1999. Социальная история России. С. Петербург, “Дмит-рий Буланин”. Т. 2, стр.335-356.
[3] Младенова, 2003. Антропологична характеристика на растежа и разви-тието на деца и подрастващи от Смолянски регион в съвременните усло-вия на живот., Дис.труд, Пловдив, 2003, 81-92.
[4] Павлов, Сп. и кол., 1980. Върху промените в телесния растеж на цдецата от детските градини в гр. Пловдив за времето 1953/1956 -1977 г., В: Пе-диатрия , XIX, 1, 58-63.
[5] Томова, И., Янакиев, Я., 2002. Етническите отношения в армията. Меж-дунар. център по пробл. на малц. и култ. взаимод., С., стр. 20.
[6] Ямпольская, Ю.А., 2003. Физическое развитие школьников Москвы во второй половине ХХ века: состояние, тенденции, прогноз. В: Алексеева, Т.И., Балановская, Е.В., Година, Е.З., Дубова, Н.А. (Ред.). Антропология на пороге ІІІ тысячелетия, т.2, Москва, Российское отделение ЕАА, стр. 567-592.
[7] Atanasova-Timeva, N. , 2005. Anthropological characteristics of Bulgarian students at the beginning of 21st century. In: Gruev. B., Nikolova, M. , Donev, A. (Eds). Proceedinngs of the Balcan Sc. Conf. of Biology in Plovdiv (Bul-garia), 82-101.
[8] Martin R., Saller K. 1957. Lehrbuch der Anthropologie, Stuttgart: Custav Fisher Verlag, 1,661.
[9] Mladenova, S., M. Nikolova, 2001. Screening in secular growth processes of children from birth to three years of age (longitudinal study in three genera-tions children in the town of Smolyan)., XV National Congress of the Bulgarian Anatomical Society with International participation, Stara Zagora, Abstracts, 70-71.
[10] Nikolova M., Mladenova S., 2003. Morphological peculiarities in the cephalic and facial structure in children and adolescents from three generations from

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1081
the town of Plovdiv., 85th Aniversary of the Department of Anatomy and Histology, Anatomical collection, Medical University of Sofia, 64-66.
[11] Piasecki, E., 1990. Konsekwencje fenotypowe transformacji demograficznej, II. Planowanie rodziny a rozwoj fizyczny dzieci I mlodziezy. Mat.Pr.Antr., 111, 5-20.
[12] Stoev, R., 2004. Age at menarche in Smolyan region – 20th century. Acta morphologica et anthropologica, Sofia, 9, 144-147.
[13] Stoev, R., Yordanov, Y., 1998. Secular changes in Bulgaria. In: Bodzar, B.E., Susanne, C. /Eds/. Secular growth changes in Europe. Eotvos Univ. Press, Budapest, 65-73.
[14] Tomazo-Ravnik, Zerbo, 1996/97. Secular growth changes of Ljubljana schoolchildren in the period from 1958 to 1994 (longitudinal series)., An-throp.Kozl., 38, 61-68.

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 1082
SCIENTIFIC PAPERS
ХЕТЕРОГЕННА ТЪКАННА ЕКСПРЕСИЯ НА КРЪВНО-ГРУПОВИТЕ АНТИГЕНИ А И В ПРИ ГРИЗАЧИ
Елена Томова, Виктория Сарафян
ПУ “Паисий Хилендарски”, МУ – Пловдив
HETEROGENIC TISSUE EXPRESSION
OF BLOOD GROUP ANTIGENS A ANG B IN RODENTS
Elena Tomova, Victoria Sarafian Plovdiv University “Paisii Hilendarski”, Medical university of Рlovdiv
ABSTRACT: Paraffin embedded tissues from stomach, intestine and ureter of four species of free-living mice from order Rodentia, family Muridae were examined for the expression of blood group antigens A and B. The immunohistochemical study was performed with the biotin-streptavidin-peroxidase staining system. A differential expression of both antigens was detected in the mucosal layer of all organs studied. In three of the species blood group antigen B showed a stronger staining intensity. In Apodemus sylvaticus antigen A revealed stronger reactivity. The observed difference in the localization and intensity of the tissue expression of А and antigens might be due to a different genetic control and/or posttranscriptional regulation.
Key words: Blood group antigens A and B, heterogenic tissue expression, Rodents
ВЪВЕДЕНИЕ Антигените на АВН кръвно-груповата система са терминални гликозилни
остатъци на олигозахаридните гликоконюгати. А и В антигените се оформят под действие на специфични гликозилтрансферази, които пренасят съответно N-acetyl-D-galactosamin или D-galactosil към предварително изградената Н верига. Тези вериги се разполагат върху клетъчните повърхности, но също се откриват и като свободни олигозахариди в някои биологични секрети.
Изучаването на тъканните и молекулни механизми на А и В антигенната експресия при различни видове бозайници и човека може да осветли някои неясни аспекти в еволюцията на АВО системата и на нейните биологични функции. В изучените представители на бозайниците експресията на кръвно-груповите антигени А и В по еритроцитите е непостоянна. Сред приматите еритроцитната експресия се ограничава само в човека и човекоподобните маймуни (шимпанзе, горила, орангутан и гибон). В други бозайници, като някои прасета, тя е резултат от пасивна абсорбция на А гликолипиди върху еритроцитната клетъчна мембрана. Обратно на непостоянната експресия на АВН антигените по повърхността на червените кръвни клетки, тъканната им

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1083
експресия във всички изучени до момента бозайници е константен феномен. Експресията на АВН антигените се явява перманентна в мукозата на червото и на горните дихателни, както и крайните пикочополови пътища [13]. Вероятно това има отношение към граничното разположение на покривните епителни клетки и връзката им с външната среда. Предполага, че тези полиморфни антигени имат роля и при взаимодействието с патогени. В определени случаи е описана зависимост между АВО фенотипове и инфекциозни заболявания при човека. Установена е модулация в адхезионната способност на някои бактерии, при доказване на А и/или В детерминанти [2], [3], [9], [11], [15]. Вероятно АВО полиморфизма има грижата да осигури механизъм на протекция срещу патогени на ниво на популациите, като съществува селективен натиск за запазване разнообразието на АВО локуса [12].
Изключително ограничени са проучванията на АВН тъканната експресия в свободно живеещи диви видове, представители на разред Гризачи. Демонстрира се наличие на АВН антигени върху различни породи плъхове, мишки и хамстери, отглеждани лабораторно. Reibel J. и съавтори [14] ги идентифицират имунофлуоресцентно в епитела на устната мукоза и предстомаха на плъхове и мишки. Bry L. и съавтори [6] доказват глюкоконюгатна продукция в интерстициални епителни клетки у трансгенни мишки. В плъховете генетичния полиморфизъм е описан чрез инбредни видове, експресиращи А антиген по целия интестинум, други показват ограничена експресия по дългия интестинум, а трети видове, с липса или минимално експресиращи А антиген в червото [4], [5]. Не се откриват данни за диференцирана тъканна антигенна експресия на кръвно-груповите антигени А и В в диви видове от разред Гризачи.
ЦЕЛ Целта в настоящата работа е да се проучи тъканната експресия на
кръвно-груповите антигени А и В в някои органи у диви видове мишки от сем. Мишевидни на разред Гризачи.
МАТЕРИАЛ И МЕТОДИ В проучването са използвани свободно живеещи, полово зрели животни
представители на разред Гризачи (Rodentia). Видовете са уловени в различни места на Южна България (близо до гр. Пловдив, гр. Хасково, гр. Велинград и гр. Царево). Изследваните органи са:
- от храносмилателна система – стомах и черво (Изследвани са три части от червото – начало, среда и край)
- от отделителна система – уретер. Органите са получени от животни под етерна наркоза в съответствие с
нормативите за хуманно отношение при експерименти с лабораторни и диви животни. Обработени са от 3-5 индивида от всеки вид. Животните принадлежат към семейство Мишевидни (Muridae) и са: Домашна мишка (Mus musculus); Горска мишка (Apodemus sylvaticus); Жълтогърла горска мишка (Apodemus flavicollis);Подземна полевка (Pythimys subteraneus).
От органите са подготвени парафинови срези, върху които е приложена биотин-стрептавидин-перохсидазна система (DAKO LSAB кит). Като първични антитела за имунохистохимичното изследване са използвани моноклонални антитела срещу човешките А и В антигени, а като субстратен хромоген – 2%

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1084
разтвор на АЕС. Контраоцветяване на ядрата е извършено с хематоксилин по Mayer. За положителна реакция се приема наличието на червено-кафяви гранули в клетъчната цитоплазма или по плазмената мембрана. За отчитане на реакцията е използвана полуколичествена скала: (++) – 80-100% позитивни клетки; (+) – 40-50% позитивни клетки; (+/-) – 10-15%; (-) – няма позитивни клетки. За всеки срез са проведени и негативни контроли. За изключване на неспецифична активност на моноклоналните антитела в животинските тъкани са осъществени инхибиционни тестове.
РЕЗУЛТАТИ И ОБСЪЖДАНЕ От проведеното имунохистохимично проучване върху тъканни срези от
органи на храносмилателната система получихме следните резултати: Черво: Mus musculus. Кръвно-групов антиген А се позитивира само в
лимфоцитни клетки на мукозния слой. Епителните клетки както и жлезите в него са имунонегативни (Фиг.1). За разлика от това кръвно-групов антиген В се доказва със значително по-висока интензивност на имунопероксидазната реакция в tunica mucosa. Позитивират се почти 100% от епителните клетки на l. epithelialis, както и лимфоцитите и някои елементи на съединителната тъкан в l. propria.
Apodemus sylvaticus. Кръвно-групов антиген A е налице в епителните клетки в tunica mucosa (Фиг. 2). На места, по дължината на червото, броят на позитивните клетки в l. epithelialis намалява, като към края А антиген се демонстрира само в секреторните гранули и в цитоплазмата на жлезистите клетки.
Кръвно-групов антиген В има подобна локализация в l. epithelialis, но в чревните жлези секреторните гранули не се позитивират. По-висока е интензивността на реакция и броят на позитивиращите се клетки при кръвно-групов антиген А.
Apodemus flavicollis и Pythimys subterraneus. Кръвно-групов антиген А се доказва само в лимфоцити от l propria, докато В антиген – в базалната част на секреторните клетки в l. epithelialis, както и в лимфоцитите на l. propria. По-висока е интензивността на реакция и броят на позитивиращите се клетки за кръвно-групов антиген В.
Стомах: В парафиновите срези от стомах на M. musculus кръвно-групов антиген А се наблюдава само в шипчестия (спинозния) и гранулозния слой на многослойния епител (Фиг. 3). Клетките на базалния и повърхностен слой, както и стомашните жлезиса имунонегативни. Кръвно-групов антиген В е налице във всички слоеве на многослойния епител (Фиг. 4), както и в стомашните жлези от tunica mucosa.
В стомаха на A. flavicollis и P. subterraneus кръвно-групов антиген А се открива с подобна на M. musculus позитивна реакция – в спинозния и гранулозния слой на многослойния епител, но липсва в базалния слой и в първите един, два реда от шипчестия слой и в жлезите. При A. flavicollis на места по дължината на органа експресията се засилва, като може да обхване всички слоеве на епитела. За кръвно-групов антиген В се позитивират основно всички слоеве на многослойния епител, както и епителните клетки на жлезите. На места обаче експресията намалява и се ограничава само до базалния слой.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1085
Наблюдаваната от нас експресия в стомах на Apodemus sylvaticus показва кръвно-групов антиген А във всички слоеве на епитела, както и в стомашните жлези, а кръвно-групов антиген В – само в клетките на базалния слой.
Уретер: Реакцията за кръвно-групов антиген В е много по-интензивна и обхваща всички слоеве на преходния епител, докато за кръвно-групов антиген А се маркират единични клетки в повърхностния слой (Фиг. 6 и 7). Находката важи за всички изследвани видове гризачи.
Една от най-неочакваните и трудни за интерпретация находки е диференцираната експресия на двата гликопротеина А и В в тъканите на разред Гризачи.
Макар че гризачите са бозайници, принадлежността им към един и същи клас с човека, не е достатъчна причина за сходен тип тъканна експресия на кръвно-груповите антигени. Основната разлика е, че А и В епитопите при човека се локализират във всички органи и системи, докато при гризачите това очевидно не е така. Cailleau-Thomas и съавтори [7] съобщават за различна локализация на А и В кръвно-груповите антигени в парафинови и криостатни срези от лабораторни плъхове BDIX. Като цяло В антиген се открива в по-малък брой тъкани. При човека А и В антигените са гликопротеини или гликолипиди [8], докато при плъховете носещите им вериги са само гликолипиди [4]. В дебелото черво А антигенът се намира във всички слоеве на мукозата, а В антигенът само в повърхностния слой. В бъбрек находката е противоположна – открива се само В антиген [7]. Gil-Loyzaga и съавтори [10] и Astic и съавтори [1] описват само В, но не и А епитопи в олфакторни и кохлеарни клетки на плъх. Данните в тези три публикации са близки по смисъл с нашите, макар че те касаят инбредни линии лабораторни гризачи.
Turcot и съавтори [16] разширяват проучването като включват освен BDIX плъхове и породите DA и WKY, в които описват подобна находка – А антигенът се експресира в цялата мукоза на стомашно-чревния тракт, а В – в повърхностните клетки. Екипът изказва хипотеза, че плъховете притежават вариабилен брой АВО гени – един ген, кодиращ В антиген, който се открива във всички индивиди и определен брой гени, кодиращи А антиген. Френските учени успяват да клонира гена, кодиращ кръвно-груповия ензим В. Оказва се, че плъховете имат повече от един АВО ген [16]. Анализът на геномните последователности при плъха показва два АВО гена, локализирани в същия локус на хромозома 3, вероятно възникнали чрез дупликация. Дупликацията на А гена при лабораторните породи би могла да се дължи и на инбридинга, чрез който се поддържа популацията им.
Това е единственото съобщение, което дава възможност да се интерпретира нашето откритие. То обяснява защо двата антигена не се суперпозират напълно в тъканите.
Твърде е възможно при свободно живеещите гризачи също да има повече от един АВО ген. Наблюдаваната от нас експресия в стомах на A. sylvaticus показва кръвно-групов антиген А във всички слоеве на епитела, както и в стомашните жлези, а кръвно-групов антиген В – само в клетките на базалния слой. В червото на същия вид за кръвно-групов антиген А се маркират епителните клетки на жлезите, а в покривния епител се позитивират само секреторните гранули, за разлика от кръвно-групов антиген В, който не

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1086
се открива в секреторните гранули. Нашето описание напълно съвпада с това на френския екип, изследвал лабораторни породи плъхове. Не е изключено в стомашно-чревния тракт на обикновената горска мишка да се експресират преимуществено А гените, а по-слабо единственият В ген. В тънкото черво на BDIX плъхове въпреки наличието на иРНК за АВО гена, не се открива В антиген, което предполага, че експресията на В антигена се регулира на посттранскрипционно ниво [16].
Находката в другите три вида гризачи – A. flavicollis, M. musculus и P. subterraneus е коренно различна по отношение интензитет и локализация на двата антигена спрямо тази, описана при лабораторните породи. Много е вероятно при свободно живеещите гризачи, в които експресията на В антигена е по-силна и повсеместна, да е на лице обратната зависимост – повече от един В ген и само един А ген. Достоверността на тази хипотеза предстои да бъде проверена чрез секвениране на изолираната от нас ДНК от изследваните гризачи.
ЗАКЛЮЧЕНИЕ Ние представяме първите данни за диференцирана тъканна експресия на
кръвно-групов антиген А и В при свободно живеещи гризачи. Предполагаме, че тя се дължи на различен генетичен контрол и/или посттранскрипционна регулация.
ЛИТЕРАТУРА [1] Astic L., Le Pendu., Mollicone R., Saucher D., and Oriol R., 1989, Cellular
expressionof H and B antigena in the rat olfactory system during development. J. Comp. Neurol., 289, 386-394.
[2] Black R. E., Levine M. M., Clements M. L., Hughes T, and O, Donell S., 1987, Association between O blood group and occurrence and severity of diarrhoea due Escherichia coli. Trans. R. Soc. Trop. Med. Hyg., 81, 120-123.
[3] Boren T., Falk P., Roth K. A., Larson G., and Normak S., 1993, Attachment of Helicobacter pylori to human gastric epithelium mediated by blood group antigens. Science, 262, 1892-1895.
[4] Bouhours D., Hansson G.C. and Bouhours J.F., 1995, Structure and genetic polymorphism of blood group A-active glycosfingolipids of the rat large intestine. J. Biol. Chem., 16370-16375.
[5] Breimer M. E., Hansson G.C., Karlsson K. A. and Leffler H., 1980, Human blood group A-positive and –negative strains of rats. Chemical basis as shown by fucolipids of small intestine. FEBS Lett., 1143, 51-56.
[6] Bry L., Falk P. G. and Gordon J. L., 1996, Genetic engineering of carbohydrate biosynthetic pathways in transgenic mice demonstrates cells cycle-associated regulation of glycoconjugate production in small intestinal epithelial cells. Proc Natl Sci USA, 93, 1161-1163.
[7] Cailleau-Thomas et al., 2002, Cloning of rat gene encoding the histo-blood group A enzyme. Tissue expression of the gene of the A and B antigens. Eur.J.Biochem., 269, 4040-4047.
[8] Clausen H. & Hakomori S. I., 1989, ABH and related histo-blood group antigens; immunochemical differences in carrier isotopes and their distribution. Vox Sang. 56, 1-20.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1087
[9] Geisel J., Steuer M. K., Ko H. L. and Beuth J., 1995, The role of ABO blood groups in infections induced by Staphylococcus saprophyticus and Pseudomonas aeruginosa. Zentralbl. Bacteriol., 282, 427-430.
[10] Gil-Loyzaga P., Pujol R., Mollicone R., Dalix A. M. & Oriol R., 1989, Appearance of B and H blood group antigens in the developing cochlear hair cells. Cell Tissue Res., 257, 17-21.
[11] Lindstedt R., Larson G., Faik.P., Jodal U., Leffler H., and Swanborg C.,1991, The receptor repertoire defines the host range for attaching Escherichia coli strains that recognize globo-A. Infect. Immun., 59,1086-1092.
[12] Merionneau S., Cailleau-Thomas A., Rocher J., Le Moullas-Vaidye B., Ruvoën-Clouet N., Clement M. &Le Pendu J., 2001, ABH and Lewis histo-blood group antigens, a model for the meaning of oligosaccharide diversity in the face of a changing world. Biochimie 83, 563-573.
[13] Oriol R., Mollicone R., Couillin P., Dalix A. M. and Candelier J. J., 1992, Genetic regulation of the expression of ABH and Lewis antigens in tissue. APMIS, 100 (Suppl 27). 28-38.
[14] Reibel J., Dabelsteen E., Hacomori S., Young W. and Makenzie I., 1984, The distribution of blood group antigens in rodent epithelia. Cell Tissue Res., 237, 111-116
[15] Swerdlow D. L., Mintz E. D., Rodriguez M., Tejada E., Ocampo C., Espejo L., Barett T. J., Petzelt J.,Bean N. H., and Seminario L., 1994, Severe life-threatening cholera associated with blood goup O in Peru: implications for the latin american epidemic. J. Infect Dis., 170, 468-472.
[16] Turcot A. L., Blancher A., Le Moullac-Vaidye B., Despiau S., Rocher J., Roubinet F., Szpirer C. and Le Pendu J., 2003, Cloning of rat gene encoding the histo-blood group B enzyme: rats more than one Abo gene. Glycobiology, Vol 13, 12, 919-928. гл. ас. д-р Елена Стоилова Томова Катедра “Биология на развитието”, ПУ”П. Хилендарски” Тел: +359 32 261 535 е-mail: е[email protected] доц. д-р Виктория Степан Сарафян катедра “Биология”, МУ – Пловдив тел: +359 32 602 531 е-mail: [email protected]
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1088
ПРИЛОЖЕНИЯ
фиг. 1: Кръвно-групов антиген А в черво от Mus musculus. Биотин-стрептавидин-пероксидазна реакция в лимфоцити от l. propria, х 400.
фиг. 2: Кръвно-групов антиген А в черво от A. sylvaticus. Биотин-стрептавидин-пероксидазна реакция в l. epithelialis, х 400.
фиг. 3: Кръвно-групов антиген А в стомах от Mus musculus. Биотин-стрептавидин-пероксидазна реакция в шипчестия
и гранулозен слой на многослойния епител, х 200.
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1089
фиг. 4: Кръвно-групов антиген В в стомах от Mus musculus. Биотин-стрептавидин-пероксидазна реакция в многослойния епител, х 200.
фиг. 5: Кръвно-групов антиген В в уретер от A. sylvaticus. Биотин-стрептавидин-пероксидазна реакция в слоевете
на преходния епител, х 400.
фиг. 6: Кръвно-групов антиген А в уретер от A. sylvaticus. Биотин-стрептавидин-пероксидазна реакция в единични
клетки на повърхностния слой от преходния епител, х 400.

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 1090
SCIENTIFIC PAPERS
CALCIUM OXALATE CRYSTALS IN VEGETATIVE AND FLORAL ORGANS
OF GALANTHUS SP. (AMARYLLIDACEAE)
Nuran Ekici & Feruzan Dane Trakya, University “Edirne, TURKEY”
ABSTRACT: In this study Ca-oxalate crystals crystals in vegetative and floral organs of Galanthus plicatus L., Galanthus gracilis L., Galanthus elwesii L. was investigated. Although crystals were seen in all vegetative organs as young adventative roots, bulb membrane, flower stalks, and leaves, they were seen only in tepal and ovary of floral organs. They were not seen in mature anthers and aged adventative roots. Calcium oxalate crystals were of the raphide type.
Key words: Amaryllidaceae, Ca-oxalate crystals, Galanthus sp., generative organs, vegetative organs
INTRODUCTION Druse and raphide crystals which are found in the structure of Ca-oxalate are
inclusions that are often seen in higher plants [1, 2, 3] and in fungi [4]. Their occurrence and abundance in specific tissues of various plants are so constant that they are used as a taxonomic tool [1, 2, 5]. The Ca-oxalate crystals in the epiderma, mesophyll, cortex, phloem, xylem parenchyma and pericarp of various plants [1, 2, 3, 6] were observed by plant anatomists. Presence of the crystals in various tissues of laticifers [7], corm [8] and seeds [9] is known. In recent years it is also notable that these crystals are present in transitory floral organs such as stamens, gynoecia and petals [10]. They are quite prevalent in floral organs of many taxa including Dilleniaceae, Liliaceae, Palmae, Malvaceae, Cunoniaceae, Euphorbiaceae [11], Solanaceae [12], Leguminosae [13]. The value of Ca-oxalate crystals to normal plant growth and development is largely unknown and probably variable [14]. Nowadays, detailed description of the calcium oxalate raphide crystal content of Leucojum aestivum vegetative and generative tissues in different phases of its life cycle were studied [15]. In this study Ca-oxalate crystals of some Galanthus species which is sister genus with Leucojum L. were researched.
AIM The aim of this study is both to contribute to the solution of the problems about
Ca-oxalate crystals and also contribute to taxonomical problems of Amaryllidaceae. MATERIAL AND METHODS Ca-oxalate crystals characters studies were made on plants from herbarium
materials of Galanthus plicatus L. (EDTU 2055), Galanthus gracilis L. (EDTU 4723), Galanthus elwesii L. (EDTU 3101). Vouchers are kept in the herbarium of Biology Department of Trakya University (EDTU). Clearing-squash technique which

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1091
developed by Herr (1971) is used to observe Ca-oxalate crystals in different tissues of Galanthus sp. [16]. Dry herbarium materials of Galanthus sp. were hydrated in warm water and then they were first pretreated in lactic acid (85 %) for 24 hours at room temperature. After this pretreatment materials were transferred directly to a recently developed clearing fluid composed of lactic acid (85 %), chloral hydrate, phenol, clove oil and xylene (2:2:2:2:1, by weight) for 48 hours at room temperature. Then they were taken on slides with a small amount of clearing fluid by means of a Pasteur pipette for microscopic examination [16]. Some of the slides were coloured with 2% aceto-orcein. All preparations were examined under phase contrast optics of Olympus BH-2 microscope. Photos were taken with Olympus C-5060 digital camera.
RESULTS AND DISCUSSION In this study is Ca-oxalate crystals in vegetative and floral organs of Galanthus
plicatus L., Galanthus gracilis L., Galanthus elwesii L. was investigated. Crystals were seen in three species of Galanthus in vegetative organs as young adventative roots, bulb membrane (Fig.1a,b,c), flower stalk (Fig.2 a,b,c) and leaf cells (Fig.3 a,b,c), they were only seen in ovary (Fig.4 a,b,c) and tepal cells (Fig.5 a,b,c) of floral organs. They were not seen in mature anthers and aged adventative roots. Crystal content are dense in bulb membrane, flower stalks, leaves, tepals, ovary and young adventative roots respectively. Calcium oxalate crystals were of the raphide type.
Physical and chemical conditions such as temperature, pressure, pH and ion concentration, may affect crystal growth, location and properties [17], however it is considered that crystal formation within the cell is under genetic control [18].
Although some species of monocotyledons have different crystal types in adjacent cells, a particular taxon can have specific crystal shape [14]. According to Watson and Dallwitz, (2005) mezophyll cells of leaves of Amaryllidaceae plants contain raphide crystals [19]. In this study, raphide crystals were observed in all vegetative and generative tissues except anthers and aged adventive roots of Galanthus sp. at their flowering phases like L. aestivum which sister genus with Galanthus [15]. Their existence in meristematic cells has been demonstrated in several studies [6, 15, 20, 21, 22] as they were seen in young adventative roots of Galanthus sp. These crystals were also seen in ovary wall of Triteleia (Themidaceae) [23] which is related taxon to Amaryllidaceae and Galanthus nivalis (Amaryllidaceae) [24], L. aestivum (Amaryllidaceae) [15] like G. plicatus, G. gracilis and G. elwesii.
CONCLUSION In conclusion, this study is determined CaOx crystals in G. plicatus, G. gracilis
and G. elwesii, being members of Amaryllidaceae in floral stages of its development. Although crystals were seen in connective tissue of anthers of L. aestivum in polen mitosis phase they were not seen in mature anthers of Galanthus species and L. aestivum. In further studies we will focus on crystal content of these taxa during their life cycles.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1092
REFERENCES [1] Esau, K. 1976. Anatomy of Seed Plant. California. [2] Fhan, A. 1982. Plant Anatomy. Pergamon Press. New York. [3] Yentür, S. 1984. Plant Anatomy. İst. Univ. Sci. Fac. Publications. No: 3283.
İstanbul. [4] Arnott, H. J. 1995. Calcium oxalate in fungi, in calcium oxalate in biological
systems, ed. S. Khan. CRC Press, Boca Raton, FL. 73-111. [5] Lersten, N. R. 1974. Morphology and distribution of colleters and crystals in
relation to the taxonomy and bacterial leaf nodule symbiosis of Psychotria (Rubiaceae). Amer. J. Bot. 61(9): 973-981.
[6] Dane, F., Meriç, Ç. & Huseyinova, G. 2000. Some Ultrastructural observations on calcium oxalate raphide crystal idioblasts and meristematic cells of the adventive root tips of Sternbergia lutea (L.) Ker-Gawl. ex Sprengel (Amaryllidaceae). – Turk. J. Bot., 24: 71-80.
[7] Dehgan, B. & Craig, M. E. 1978. Types of laticifers and crystals in Jatropha and their taxonomic implication. Amer. J. Bot. 65(3): 345-352.
[8] Sunell, L. A. & Healy, P. L. 1979. Distribution of calcium oxalate crystal idioblasts in corm of taro (Colocasia esculenta). Amer. J. of Bot. 66(9): 1029-1032.
[9] Ilarslan, H., Palmer, R. G., Imsande, J. & Horner, H. T. 1997. Quantitative determination of calcium oxalate and oxalate in developing seeds of soybean (Leguminosae). Amer. J. Bot. 84(8): 1042-1046.
[10] Meriç, Ç. & Dane, F. 2004. Calcium oxalate crystals in floral organs of Helianthus annuus L. (Asteraceae). – Acta Biologica Szegediensis, 48(1-4): 19-23.
[11] Tilton, V. R. & Horner, H. T. 1980. Calcium oxalate raphide crystals and crystalliferous idioblasts in the carpels of Ornithogalum caudatum. Ann. Bot. 46: 533-539.
[12] Horner, H. T. & Wagner, B. L. 1992. Association of four different calcium crystals in the anther connective tissue and hypodermal stomium of Capsicum annuum (Solanaceae) during microsporogenesis. Amer. J. Bot. 67: 1347-1360.
[13] Buss, P. A. & Lersten, N. R. 1972. Crystals in tapetal cells of the Leguminosae. Bot. J. Linn. Soc. 65: 81-85.
[14] Prychid, C. J. & Rudall, P. J. 1999. Calcium oxalate crystals in monocotyledons: a review of their structure and systematics. Ann. Bot. 84: 725-739.
[15] Ekici, N. & Dane F. 2006. Calcium oxalate crystals during development of male and female gametophyte in Leucojum aestivum L. (Amaryllidaceae). Book of abstracts. IV. Balkan Botanical Congress. Sofia, Bulgaria. Pp. 125.
[16] Herr, J.M. 1971. A new clearing-squash technique for the study of ovule development in angiosperms. Amer. J. Bot. 58(8): 785-790.
[17] Franceschi, V. R. & Horner, H. T. 1980. Calcium oxalate crystals in plants. Bot. Rev. 46: 361-427.
[18] Ilarslan, H., Palmer, R. G., & Horner, H. T. 2001. Calcium oxalate crystals in developing seeds of soybean. Ann. Bot. 88: 243-257.
[19] Watson, L. & Dallwitz, M. J. 2005. The Families of Flowering Plants. http://delta-intkey.com/angio/www/amaryllidaceae.html.
[20] Horner, H. T. & Wagner, B. L. 1980. Association of druse crystals with the developing stomium of Capsicum annuum (Solanaceae) anthers. Amer. J. Bot.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1093
67(9): 1347-1360. [21] Kausch, A. P. & Horner, H. T. 1983. The development of mucilaginous raphide
crystal idioblasts in young leaves of Typha angustifolia L. (Typhaceae). Amer. J. Bot. 70(5): 691-705.
[22] Seago, J. L. & Marsh, L. 1989. Adventitious root development in Typha glauca with emphasis on the cortex. Amer. J. Bot. 76(6): 909-923.
[23] Berg, R. Y. 2003. Development of ovule, embryo sac and endosperm in Triteleia (Themidaceae) relative to taxonomy. Amer. J. Bot. 90(6): 937-948.
[24] Chudzik, B. & Sniezko, R. 2003. Calcium ion presence as a trait of receptivity in tenuinucellar ovules of Galanthus nivalis L. Acta Biologica Cracoviensia Series Botanica 45(1): 133-141. Official address of autors: Trakya University, Faculty of Sciences &Arts, Department of Biology 22030, Edirne, Turkey. E-mails: [email protected], [email protected]
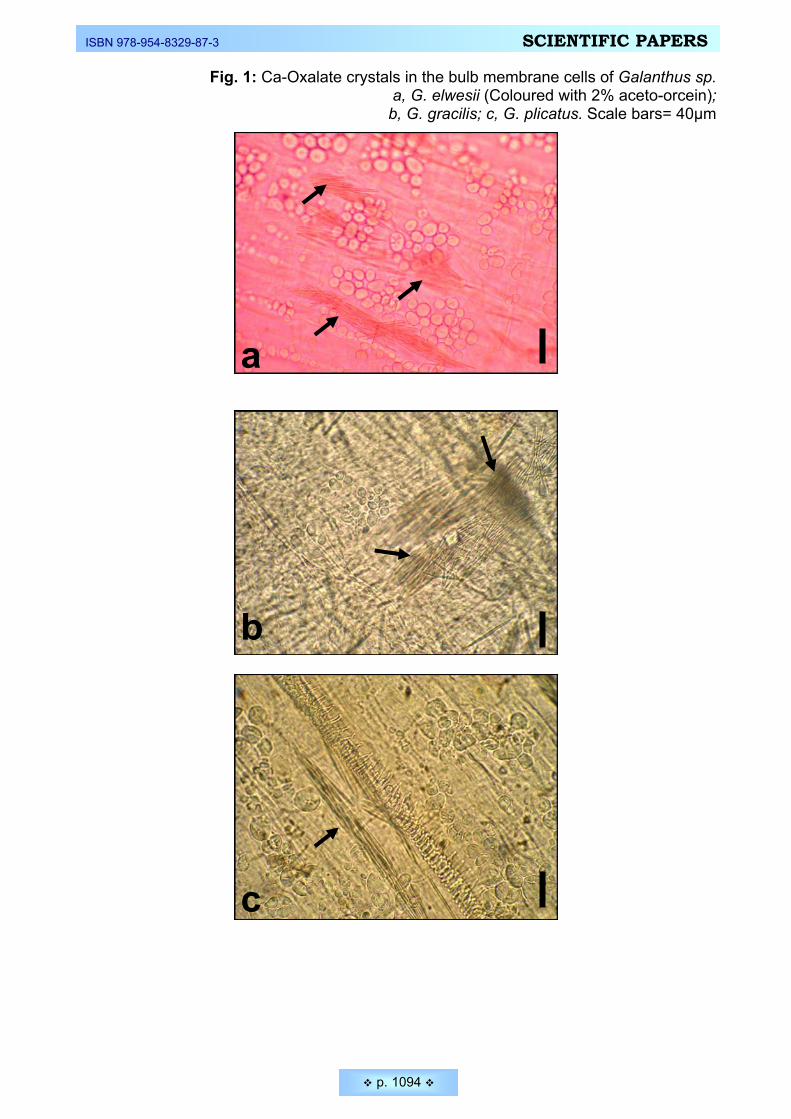
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1094
Fig. 1: Ca-Oxalate crystals in the bulb membrane cells of Galanthus sp. a, G. elwesii (Coloured with 2% aceto-orcein); b, G. gracilis; c, G. plicatus. Scale bars= 40µm
a
b
c

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1095
Fig. 2: Ca-Oxalate crystals in the flower stalk cells of Galanthus sp. a, G. elwesii; b, G. gracilis; c, G. plicatus. Scale bars= 40µm
a
b
c

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1096
Fig. 3: Ca-Oxalate crystals in the leaf cells of Galanthus sp. a, G. elwesii; b, G. gracilis; c, G. plicatus. Scale bars= 40µm
a
b
c
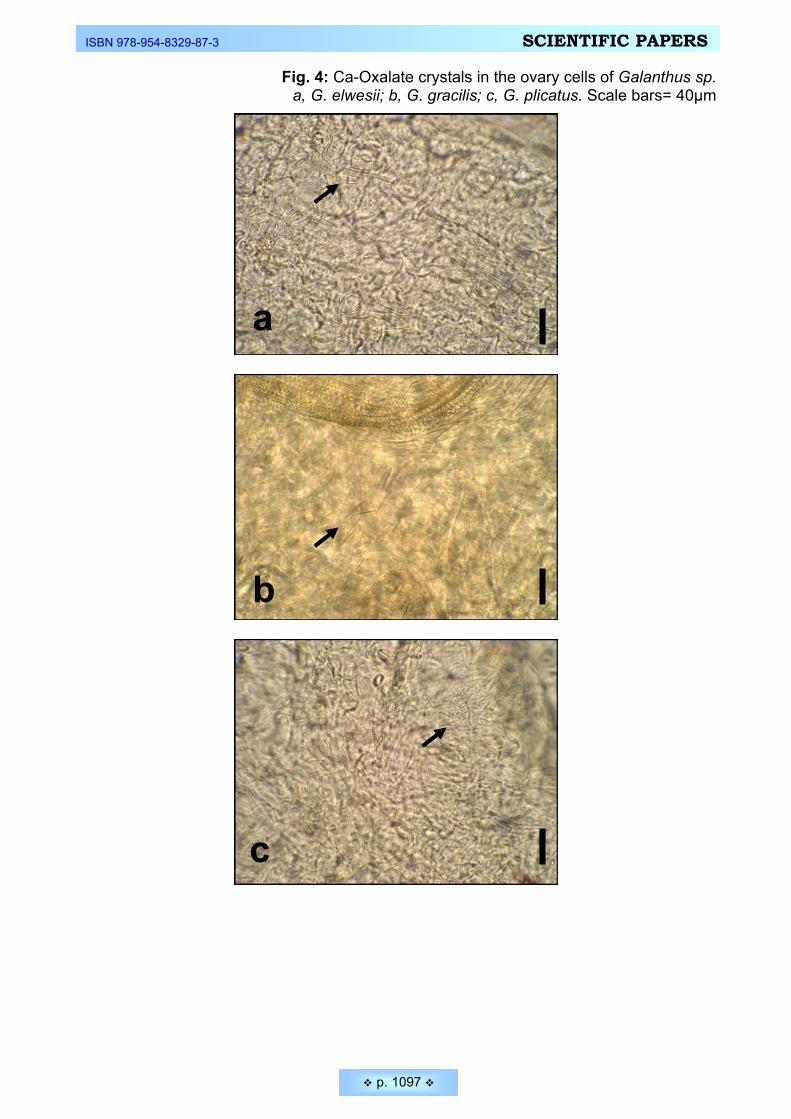
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1097
Fig. 4: Ca-Oxalate crystals in the ovary cells of Galanthus sp. a, G. elwesii; b, G. gracilis; c, G. plicatus. Scale bars= 40µm
c
b
a
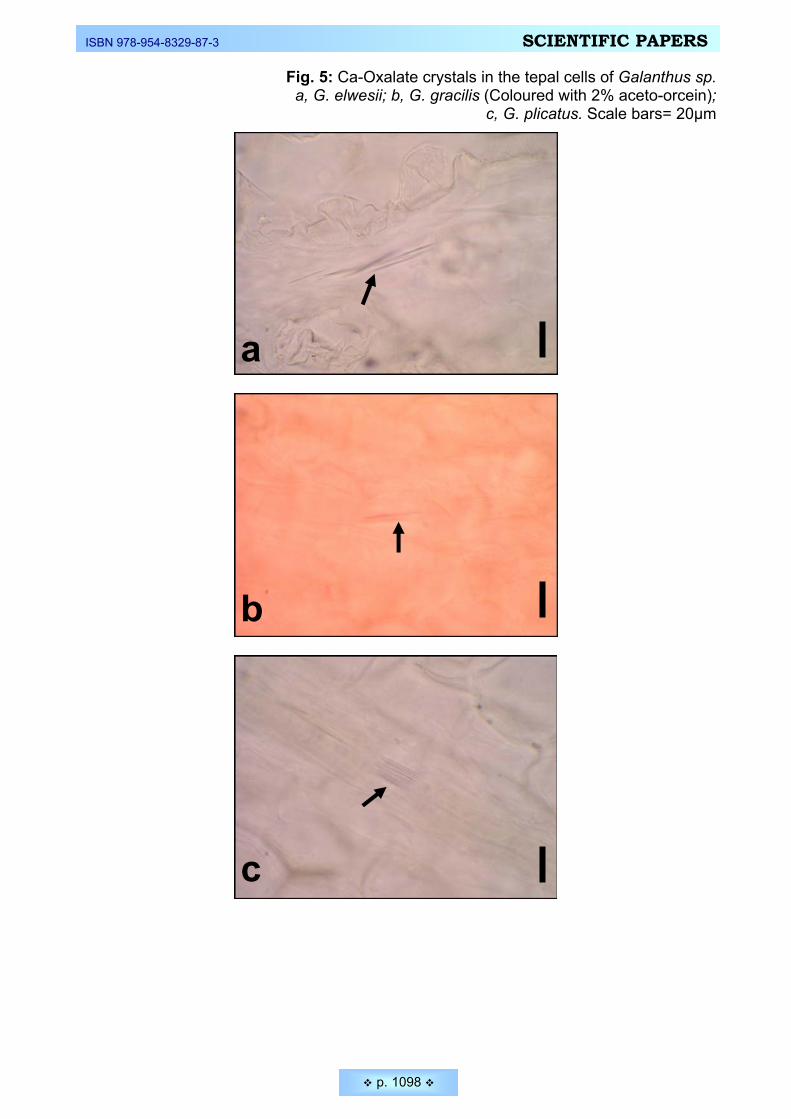
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1098
Fig. 5: Ca-Oxalate crystals in the tepal cells of Galanthus sp. a, G. elwesii; b, G. gracilis (Coloured with 2% aceto-orcein);
c, G. plicatus. Scale bars= 20µm
a
b
c

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 1099
SCIENTIFIC PAPERS
BIOLOGICAL DIVERSITY AND ECOLOGICAL EVALUATION OF THE FRESH WATER ECOSYSTEMS FROM THE ARDA RIVER, BULGARIA
Diana Kirin
Agricultural university – Plovdiv ABSTRACT: The purpose of the carried on analysis has been to investigate the bio-
logical variety of the freshwater ecosystems form the Arda river and to make an ecological evaluation of their condition. The analysis have been made in the region from the reservoir “Studen Kladenets” to the border area with Republic of Greece (region of village Slaveevo, Eastern Rhodopes) during the three seasons of the research period: 2002-2005 years. For an ecological evaluation of the situation of the analyzed freshwater ecosystems, principal biotic indexes have been fixed. The analysis of the dominant structure of the found taxons has been presented to the level of the component societies. These researches would allow to make a characterization of the situation and a prognostication of the changes in the studied freshwater biocenoses under the influence of the anthropogenous effects; to pre-sent recommendations for preservation of the genetic fund in them; to plan possibilities for preservation of the natural water resources in connection with the total decreasing of the water resources in Bulgaria and the possibility to use them for drinking water in the nearest future; to plan possibilities to prevent any transborder pollution and the consequences re-lated to it (Agreement between the Republic of Bulgaria and the Republic of Greece for collaboration at using the river waters that flow through the territories of the two countries ect.).
Key words: biodiversity, monitoring, Arda River INTRODUCTION The Arda River is related to the Aegean water collecting region. The valley of
the river from the sources to the State border is indicated as a region with middle and high degree of importance in respect of the date about the species richness, the endemic and rare taxons [14].
Macroinvertebrates, freshwater fishes and their helminthes are between the most frequently used bioindicating organisms to evaluate the situation of the fresh-water systems [15].
These researches would allow to make a characterization of the situation and a prognostication of the changes in the studied freshwater biocenoses under the influence of the anthropogenous effects; to present recommendations for preserva-tion of the genetic fund in them; to plan possibilities for preservation of the natural water resources in connection with the total decreasing of the water resources in Bulgaria and the possibility to use them for drinking water in the nearest future; to plan possibilities to prevent any transborder pollution and the consequences related to it etc. [18].

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1100
PURPOSE The purpose of the carried on analysis has been to investigate the biological
variety of the freshwater ecosystems form the Arda river and to make an ecological evaluation of their condition.
MATERIAL AND METHODS The analysis have been made in the region from the reservoir “Studen Klad-
enets” to border area with Republic of Greece (region of village Slaveevo, Eastern Rhodopes) during the three seasons of the research period (2002-2005 years): spring; summer and autumn. The region of village Slaveevo is a part of the National ecological net – object of NATURA 2000 as Slaveevo – Dolna Arda, as well as the valley of the Arda River, the dam Studen kladenets, the dam Ivaylovgrad, Madzharovo, etc.
Hydrobiological monitoring through analysis of the bioindicative macroinverte-brate taxa was carried out. The biotic index and saprobic index were determined [12,
19]. Freshwater fishes – hosts were examined for helminths using standard tech-
niques. Specimens were fixed and preserved in 70% ethanol. The trematodes and cestodes were stained with iron acetocarmine [3] and studied on Canada balsam whole mounts. The nematodes and acanthocephalans were studied on temporary mounts with 5% glycerol in 70% ethanol [11, 13].
For each one of the helminthes species found for each season, data were pre-sented as to the number of examined hosts (N), the number of infected hosts (n), number of parasites (p), prevalence (P%) and mean abundance (MA).
The ecological terms prevalence, mean abundance and mean intensity are used here based on the terminology of Bush et al. and Marcogliese [1, 10].
Analyses of helminth communities and of water macroinvertebrate communi-ties structure were carried out at thirty seasons and at both levels: infracommunity and component community [1]. The infracommunity data were used to calculate the total number of species, mean number of helminth worms, the Brilloun’s diversity index (HB), the evenness index of Pielou (E) and the Simpson’s index of domi-nance (C) [9].
The criterion of Bush et al. [2] is used for determining the dominant structure of the component helminths communities (Bush et all., 1997). The species are divided into accidental species (P%<10), component species (P%<20) and basic species (P%>20), based on their prevalence [8].
The semblance of the helminths communities in L. cephalus from the Arda River was determined according to quantitative data (total number of specimens, prevalence and mean intensity) by means of Jaccard’s coefficient (Ij) [17].
The similarity and diversity measures were calculated using the program BIO-DIV [1].
RESULTS AND DISCUSSION І. General characterization of the studied biotopes Studies have been performed of freshwater ecosystems from the Arda River in
the vicinity of the village of Rabovo, the town of Madzharovo and of the villages Huhla and Slaveevo, respectively marked as biotopes Rabovo, Madzharovo, Huhla, Slaveevo. Biotope Rabovo includes a section of the Arda River, starting after the

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1101
dam wall of the dam Studen kladenets. It begins with shallow, fast running water and continues with deep pools and slow running water. The bed is entirely stony.
The aquatic vegetation is abundantly represented by the dominant species: Marchantia polymorpha L., Fontinalis antipyretica L., Drepanocladus aduncus (Hedw.), Marsupella sphacelata (Gies.). The overgrowing of them periodically (dur-ing the night hours) are washed over by the water rising from the dam lake “Studen kladenets” when the turbines of the Water Electric Plant “Studen kladenets” are turned on, which fills almost entirely the deep river bed. The riverside vegetation is situated high above the river bed.
Biotope Madzharovo is situated at 30 km away from biotope Rabovo (before the dam lake Ivaylovgrad). It is distinguished with a less depth and slow running water, with rapids at some places. The waterside vegetation is represented mainly by Salix sp. and Alnus glutinosa, and the water vegetation – by the species Fon-tinalis antipyretica and Drepanocladus aduncus. The riverbed is considerably mote slanting with good formed sandy bottom.
The species Marchantia polymorpha, Fontinalis antipyretica, Drepanocladus aduncus, Marsupella sphacelata are bioindicators, Marchantia polymorpha and Marsupella sphacelata are indicators for o-saprobity, and the rest two species are developed in hydro biotopes, which are distinguished with o- to β-mesosaprobity, but with higher saprobity valences for o-saprobity [5].
Biotope Huhla includes a section of the Arda River, starting after the reservoir “Ivaylovgrad”. It begins with shallow, fast running water and continues with deep pools and slow running water. The bed is entirely gritty and clayey.
The riverside vegetation is situated high above the river bed. The riverside is absorbed in greenery, represented mainly by Quercus frainetto Tenore, Q. coccif-era L., Q. cerris L., Q. pubescens Willd., Q. thracica Stef. and Ned., Q. policarpa Schur., Acer tataricum L., A. campestre L., Ulmus minor Mill., Carpinus betulus L., C. orientalis Miller, Fraxinus ornus L., ect.
Biotope Slaveevo includes a section of the Arda River, before the border area with Republic of Greece, Eastern Rhodopes, Bulgaria. It begins with shallow, fast running water and continues with deep pools and slow running water. The bed is entirely gritty and clayey. The riverside vegetation is situated high above the river bed.
The riverside is absorbed in greenery, represented mainly by Quercus frainetto Tenore, Q. coccifera L., Q. cerris L., Q. pubescens Willd., Q. thracica Stef. and Ned., Q. policarpa Schur., Acer tataricum L., A. campestre L., Ulmus minor Mill., Carpinus betulus L., C. orientalis Miller, Fraxinus ornus L., Alnus glutinosa (L.), ect.
ІІ. Biological diversity and ecological estimation of the macroinvertebrate
fauna from the study of freshwater ecosystems of the Arda River In biotopes Rabovo and Madzharovo during the period of the study are regis-
tered generally 20 taxons of bioindicators macroinvertebrate organisms, 13 respec-tively for each of the two biotopes. In biotope Huhla are registered generally 21 tax-ons of bioindicators macroinvertebrate organisms (Table 1).
With highest numbers in biotopes Rabovo and madzharovo are distinguished the representatives of Ancylus fluiviatilis (respectively 42 and 65 specimens), of Nematoda sp. (respectively by 8 and 42 specimens) and so on. With highest num-bers in the biotope Huhla are distinguished the representatives of Nematoda sp.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1102
(25 specimens), Eylais meridionalis (18 specimens), Hydropsyche sp. (15 speci-mens) and Tabanus sp. (12 specimens) (Table 1).
The general number of the specimens in the biotope Madzharevo is higher (154 specimens) than this in the biotope Rabovo (105 specimens) and that this in the biotope Huhla (126 specimens) (Table 1).
The determined indexes of diversity HB (HB=1.84, HB=1.54 and HB=2.36, re-spectively) and determined indexes of evenness E (E=0.79, E=0.65 and HB=0.86, respectively) testify to originally higher diversity of species in the biotope Huhla. These results are opposites by the determined index of dominance C (C=0.20, C=0.27 and E=0.10, respectively) (Table 1, Fig. 1).
The saprobical degree of the studied biotopes of the Arda River is determined with the using of saprobical valences and indicatoring weight, as well as the num-ber of each one of the concrete found bioindicating taxons [12]. The saprobical va-lences of four taxons in the performed study are higher for o-saprobity. These are: Ancylus fluviatilis, Podura aquatica, Simulium sp. (larvae), and Chaoborus sp. (lar-vae).
P. aquatica, Simulium sp. (larvae), and Chaoborus sp. (larvae) are repre-sented only in the biotope Rabovo.
Ten taxons are distinguished with higher saprobical valences for β-mesosaprobity. Of them: 4 are common for both of the biotopes, 4 can be found only in the biotope Madzharovo, and 2 – only in the biotope Rabovo. 3 taxons of those found at this stage are distinguished with equal saprobical valences for o- and β-mesosaprobity. These are: Hyphydrus sp. (represented only in the biotope Rabovo); Ephemerella sp. (larvae) (found in both of the biotopes) and Stenophylax sp. (larvae) (registered only in the biotope Madzharovo). Two of the taxons (Asellus aquaticus and Tipula sp. (larvae)) have proved highest saprobical valences for α-mesosaprobity, and one taxon – for p-saprobity (Limnodrilus hoffmeisteri). The last three taxons are found only in the biotope Madzharovo and are represented with low numbers (Table 1).
The values of the saprobical degrees determine, according to the present study, the biotope Rabovo as o-saprobical, and the biotope Madzharovo – as β-mesosaprobity. The diversity of species of the determined taxons justifies that bio-tope Rabovo can be determined with biotical index 4, and biotope Madzharovo – with biotical index 3 [19].
The values of the determined index of equalization (E) confirm in a mathemati-cally authentic way the established degrees of o- and β-mesosaprobity in the stud-ied biotopes (E>0.7 – o-saprobical zone, E>0.6 – β-mesosaprobical zone, accord-ing to the criteria of Pielou [17]).
The saprobical valences, in the biotope Huhla, of one taxon in the performed study are higher for o-saprobity (Simulium sp. (larvae)), 19 taxons are distinguished with higher saprobical valences for β-mesosaprobity. 3 taxons of those found at this stage are distinguished with equal saprobical valences for o-saprobity and β-mesosaprobity. These are: Hyphydrus sp.; Ephemerella sp. (larvae) and Stenophy-lax sp. (larvae). Two of the taxons (Asellus aquaticus and Tipula sp. (larvae)) have proved highest saprobical valences for α-mesosaprobity, and one taxon – for p-saprobity (Limnodrilus hoffmeisteri) (Table 1).
The values of the saprobical degrees determine, according to the present study, the biotope Huhla as β-mesosaprobity. The diversity of species of the deter-

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1103
mined taxons justifies that biotope Huhla can be determined with biotical index 4,5 [12].
The values of the determined index of evenness (E) confirm in a mathemati-cally authentic way the established degrees of β-mesosaprobity in the studied bio-topes (E>0.6 – β-mesosaprobical zone, according to the criteria of Pielou [17]).
The authenticity of the obtained results is also confirmed by t – criterion (t=1.01; ν=25; q>0.3), guarantying in more than 90% of the cases, that the obtained results are not accidental.
In the biotope Slaveevo didn’t make hidrobiological monitoring through the higher level on the water during the period on these study.
The semblance of the macroinvertebrates in the Arda River is determined by comparing the total number of specimens of the species found, utilizing the sem-blance coefficient of Jaccard (Ij) (Table 3, Fig. 3).
Upon determining the Ij by comparing the total number of specimens of the helminthes species found were defined the following semblance groups: the macro-invertebrate communities are with highest semblance coefficient read in the bio-topes Madzharovo – Rabovo (Ij=0.349), following these in the biotopes Huhla – Madzharovo (Ij=0.285). These are connecting with consecutive proximity on the biotopes and the presence of one and three common species, respectively.
ІІІ. Biological diversity and ecological estimation of the helminthic com-
plexes of Leuciscus cephalus (L., 1758) of the studied freshwater ecosys-tems.
Fifteen species of fish on the territory of Bulgaria and their helminthic com-plexes are determined as bioindicators [15]. One of the bioindicating species of fish is L. cephalus. The chub is a predatory fish.
As a result of the performed for the first time in Bulgaria helminthic study of 208 specimens of Leuciscus cephalus (28 specimens of the biotope Rabovo, 95 specimens of biotope Madzharovo, 33 specimens of biotope Huhla, 52 specimens of biotope Slaveevo) of the Arda river, an invasion has been found with 11 species of helminthes: Ichthyocotylurus pileatus (metacercaria); Clinostomum complanatum (metacercaria); Posthodiplostomum cuticola (metacerc.); Caryophyllaeides fennica; Caryophyllaeus brachycollis; Bothriocephalus achielognatii; Ligula intestinalis (ple-rocercoid); Paradilepis scolecina (cysticerc); Acathocephalus anguillae; Rhab-docona denudate; Raphidascaris acus.
Common species for the four biotopes is C. fennica (Table 2). L. cephalus from biotope Slaveevo is distinguished with the biggest number of
species (10 species of helminthes, Table 2), followed by the number of species of helminthes in the helminthic communities of the studied hosts from the biotope Huhla (by 8 species), followed by the number of the helminthic species of L. cepha-lus from the biotope Madzharovo (3 species) and the number of the helminthic spe-cies from the biotope Rabovo (two species of helmithes).
With the greatest general number of specimens is looms L. cephalus from the biotope Slaveevo (348 specimens), followed by the biotope Madzharovo (206 specimens), biotope Huhla (111 specimens) and Rabovo (3 specimens). With the greatest density of populations is distinguished again I. pileatus (MA=7.00) and C. brachycollis (MA=5.00) from the biotope Huhla, followed by those of I. pileatus (MA=4.45) and C. brachycollis (MA=4.20) from the biotope Madzharovo. The mean abundance is under 4.00 for the rest species and biotopes.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1104
The dominant structure of the found helminth communities has been deter-mined. According to the criteria of Bush et al. [1] the basic species of helminthes are: I. pileatus (P%=21.05) and C. brachycollis (P%=26.32) from the biotope Madzharovo; Rh. denudata from (P%=51.52) the biotope Huhla; I. pileatus (P%=23.07), P. cuticola (P%=32.69), C. brachycollis (P%=21.15), L. intestinalis (P%=71.15), P. scolecina (P%=21.15), Ac. anguillae (P%=36.54), Rh. denudata (P%=92.31) found for L. cephalus from the biotope Slaveevo. Composing species of helminthes, according to the performed study are C. brachycollis (P%=15.15) from the biotope Huhla and C. fennica (P%=17.31) from the biotope Slaveevo (Ta-ble 2). The rest of all species from the four biotopes are accidental for the helminth communities of the L. cephalus.
Represented species of helminthes are developing with the presence of inter-mediate hosts:
1. Intermediate hosts for Ichthyocotylurus pileatus are Mollusca and fishes from Cyprinidae, Percidae, Esocidae, Siluridae, etc. Definitive hosts are Laridae.
2. Clinostomum complanatum 3. The first intermediate hosts for Posthodiplostomum citicola are Planorbis
planorbis and the second – fishes from Cyprinidae, Percidae, Esocidae, Siluridae, etc. Definitive hosts are: Ardea cinerea, A. purpurea, fish eating birds from Nicti-corax.
4. Intermediate host for Caryophyllaeides fennica is Stylaria lacustris. Tubifex tubifex is an intermediates host of Caryophyllaeus brachycollis.
5. Caryophyllaeus brachycollis 6. Intermediate hosts for Bothriocephalus acheilognathi are diferent species
of cyclops: Mesocyclops leucarti, Ectocyclops phaleratus, Tetmocyclops taihkonen-sis, Cyclops strenuus, Mesocyclops dybowskii, M. crassus, Acanthcyclops vernalis, A. bicuspidatum, ets.
7. The first intermediate hosts for Ligula intestinalis are diferent species of cyclops: Cyclops strenuus, Acanthocyclops bicuspidatus, A. viridis, Euciclops seru-latus, Diaptomus gracilis, etc. The second intermediate host are diferent species of fishes: Abramis brama, A. sapa, Scardinius erythrophthalmus, Alburnus alburnus, Alburnoides bipunctatus, Gobio gobio, Rutilus rutilus, Barbus barbus, B. m. petenyi, L. cephalus, L. idus, Phoxinus phoxinus. Definitive hosts are fish eating birds from Larus, Bucephalus, Mergus, Podiceps.
8. Definitive hosts for Paradilepis scolecina are the birds from genus Pha-lacrocorax.
9. Asellus aquaticus is intermediate host for Acanthocephalus anguillae. De-finitive hosts are diferent freshwater fishes from Salmonidae, Percidae, Anguillidae, Esocidae, etc.
10. Intermediate hosts for Rhabdochona denudata are representatives of Hydropsyche, Heptagenia, Ephemerella, etc. Definitive hosts are freshwater fishes from Cyprinidae.
11. Hosts for Raphidascaris acus are freshwater fishes [4, 11]. In the biotope Rabovo are represented species of helminthes, which are de-
veloping with the presence of intermediate hosts: cyclops and representatives of Hydropsyche, Heptagenia and Ephemerella. In the biotope Madzharovo dominate species of helminthes, in the cycle of development of which participate Oligohaeta and Gastropoda. In the biotope Huhla are represented species of helminthes, which are developing with the presence of intermediate hosts cyclops and representatives

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1105
of Hydropsyche, Heptagenia, Ephemerella, Oligohaeta, Gastropoda and Crustacea (Asellus aquaticus). In the biotope Slaveevo are represented species of helminthes, which are developing themselves with the presence of intermediate hosts too.
The obtained results for the helminthes complexes of L. cephalus correspond and are in close connection and dependence of the biology and ecology of the con-crete species of helminthes and the place of the intermediate hosts as bioindicators for the status of the studied natural hydroecosystems.
For the biotope Slaveevo higher values are found for HB, and the results are in close connection with the dominant structure of the found communities of helmin-thes (HB=1.95). For the biotope Rabovo higher values are found for C (C=0.50) and E (E=1.00). These biotic indices are concerning the fully counted collections and it is sensitive to data with low frequency [17].
These biotic indices are concerning the fully counted collections and it is sensi-tive to data with low frequency [17].
The semblance of the helminthes communities in L. cephalus in the Arda River is determined for the four biotopes of the period the research was carried out by comparing their prevalence and mean intensity utilizing the semblance coefficient of Jaccard (Ij) (Table 4; Fig. 4-5).
Upon determining the Ij by comparing the prevalence of each of them there were defined the following semblance groups: the helminths communities in the chubs are with highest semblance coefficient read in the Huhla – Slaveevo (Ij=0.436). Those higher semblance coefficient are most probably due to the pres-ence of the seven common species – I. pileatus, C. fennica, C. brachycollis, B. achielognatii, L. intestinalis, A. anguillae, R. denudata, of which R. denudata is a basic species for the helminths communities in the chub in the two biotops (Table 4; Fig. 5).
The helminthological communities in the chub in the biotopes Huhla – Slaveevo are distinguished with the highest coefficients based on the mean inten-sity (Ij=0.405) (Table 4, Fig. 4).
With a very low coefficient are included the helminthological communities of the biotopes Rabovo – Madzharovo (Table 4; Fig. 4-5).
CONCLUSIONS For the first time in Bulgaria systematical ecological monitoring researches
have been performed of the freshwater ecosystems of the Arda River (biotopes Rabovo, Madzharovo, Huhla, Slaveevo). Researches have been performed of the bioindicating aquatic vegetation, the bioindicating macroinvertebrate fauna, the ichtiofauna and the helminthic complexes of L. cephalus of the studied ecosystems. 4 types of dominant aquatic vegetation have been found, totally 25 taxons bio indi-cating macroinvertebrate fauna and 11 species of helminthes. The obtained results and the performed mathematical processing determine, according to the present study, the biotope Rabovo as o-saprobical, and biotopes Madzharovo—Huhla-Slaveevo – as β-mesosaprobical and indicate the favorable development of the studied ecosystems of the Arda river.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1106
REFERENCES [1] Baev, P., L. Penev, 1995. BIODIV. Program for calculating biological diversity
parameters, similarity, niche overlap and cluster analysis. Version 5.1. – Exeter Software, New York, 57.
[2] Bush, A. O., K. D. Lafferty, J. M. Lotz, A. W. Shostak. 1997. Parasitology meets ecology on its own terms: Margolis et al. revised. Journal of Parasitol-ogy 83: 575-583.
[3] Georgiev, B., V. Biserkov, T. Genov, 1986. In toto staining method for cesto-des with iron acetocarmine. – Helminthologia, 23, 279-281.
[4] Kakacheva – Avramova, D.,1983. Helminthes of the freshwater fish in Bulgaria – Bulgarian Academy of Science, Sofia, 261.
[5] Karapetkova, M., M. Zhivkov, 1995. The fish in Bulgaria – Gea Libris, Sofia, 248.
[6] Klemm, D. et all., 1990. Macroinvertebrate field and laboratory methods for evaluating the biological integrity of surface waters. EPA: Cincinnati, OH, 285.
[7] Кокin, К., 1982. Ecology highest water plants. – The publication house of the Moscov University, 158.
[8] Krebs C., 1989. Ecological Methodology. – Harper Collins Publishers, New York, 654.
[9] Maguran, A., 1988. Ecological diversity and its measurement. – Princeton Uni-versity Press, Princeton, 189.
[10] Marcogliese, D. M. 1999. Parasites of fishes in fresh water. Ecological Monitor-ing and Assessment Network 784.
[11] Margaritov, N., 1959. Parasites on some freshwater fish in the country. Scien-tific and Research Institute of Fish Industry, Varna, 21.
[12] Methods for hydrobiological monitoring, 1998 – MOSV, 16. [13] Moravec, F. et al. Endohelminth fauna of barbel, Barbus barbus (L.), under
ecological conditions of the Danube basin in Central Europe. – PHAS of the Czech Republic, Praha, 1997, 96.
[14] National strategy for preservation of the biological diversity, 1998. NBDCS, Sofia, т. І, 663.
[15] National program for biomonitoring of Bulgaria, 1999. – Gea Libris, Sofia, 240. [16] Official Newspaper 87/1964. [17] Pielou, Е., 1975. Ecological diversity. – Wiley, New York, 273. [18] Rosenberg, D. M. and V. H. Resh. 1997. Introduction to freshwater biomonitor-
ing and bentic macroinvertebrates, p. 1-9. In: M. D. Rosenberg and V. H. Resh (eds.) Freshwater biomonitoring and bentic macroinvertebrates. Chapman and Hall, New York.
[19] Russev B., 1993. Principles of the saprobiology – The publication house of the University “Sveti Kliment Ohridski”, 161. Diana Atanasova Kirin, Assoc. Prof. Dr. Department of Agroecology University of Agriculture 12 Mendeleev Street, 4000 Plovdiv Bulgaria` Tel./Fax: +358 32 685 855 E-mail: [email protected]
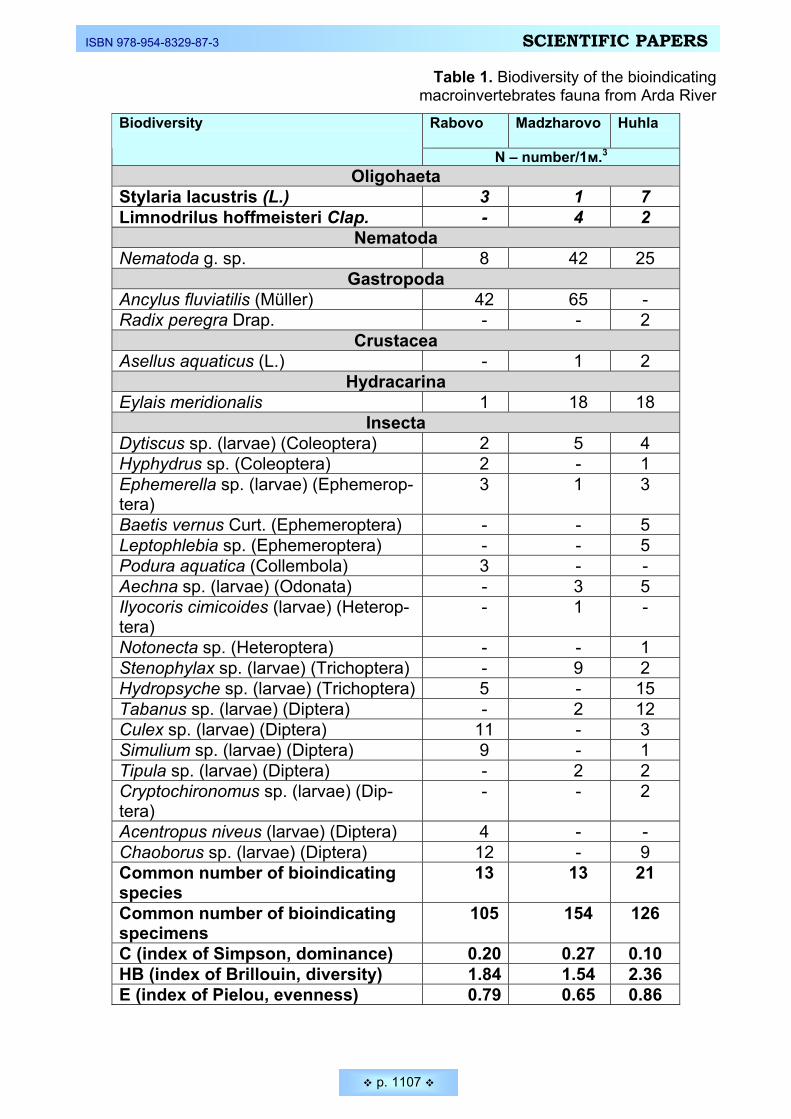
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1107
Table 1. Biodiversity of the bioindicating macroinvertebrates fauna from Arda River
Rabovo
Madzharovo Huhla Biodiversity
N – number/1м.3 Oligohaeta
Stylaria lacustris (L.) 3 1 7 Limnodrilus hoffmeisteri Clap. - 4 2
Nematoda Nematoda g. sp. 8 42 25
Gastropoda Ancylus fluviatilis (Müller) 42 65 - Radix peregra Drap. - - 2
Crustacea Asellus aquaticus (L.) - 1 2
Hydracarina Eylais meridionalis 1 18 18
Insecta Dytiscus sp. (larvae) (Coleoptera) 2 5 4 Hyphydrus sp. (Coleoptera) 2 - 1 Ephemerella sp. (larvae) (Ephemerop-tera)
3 1 3
Baetis vernus Curt. (Ephemeroptera) - - 5 Leptophlebia sp. (Ephemeroptera) - - 5 Podura aquatica (Collembola) 3 - - Aechna sp. (larvae) (Odonata) - 3 5 Ilyocoris cimicoides (larvae) (Heterop-tera)
- 1 -
Notonecta sp. (Heteroptera) - - 1 Stenophylax sp. (larvae) (Trichoptera) - 9 2 Hydropsyche sp. (larvae) (Trichoptera) 5 - 15 Tabanus sp. (larvae) (Diptera) - 2 12 Culex sp. (larvae) (Diptera) 11 - 3 Simulium sp. (larvae) (Diptera) 9 - 1 Tipula sp. (larvae) (Diptera) - 2 2 Cryptochironomus sp. (larvae) (Dip-tera)
- - 2
Acentropus niveus (larvae) (Diptera) 4 - - Chaoborus sp. (larvae) (Diptera) 12 - 9 Common number of bioindicating species
13 13 21
Common number of bioindicating specimens
105 154 126
C (index of Simpson, dominance) 0.20 0.27 0.10 HB (index of Brillouin, diversity) 1.84 1.54 2.36 Е (index of Pielou, evenness) 0.79 0.65 0.86
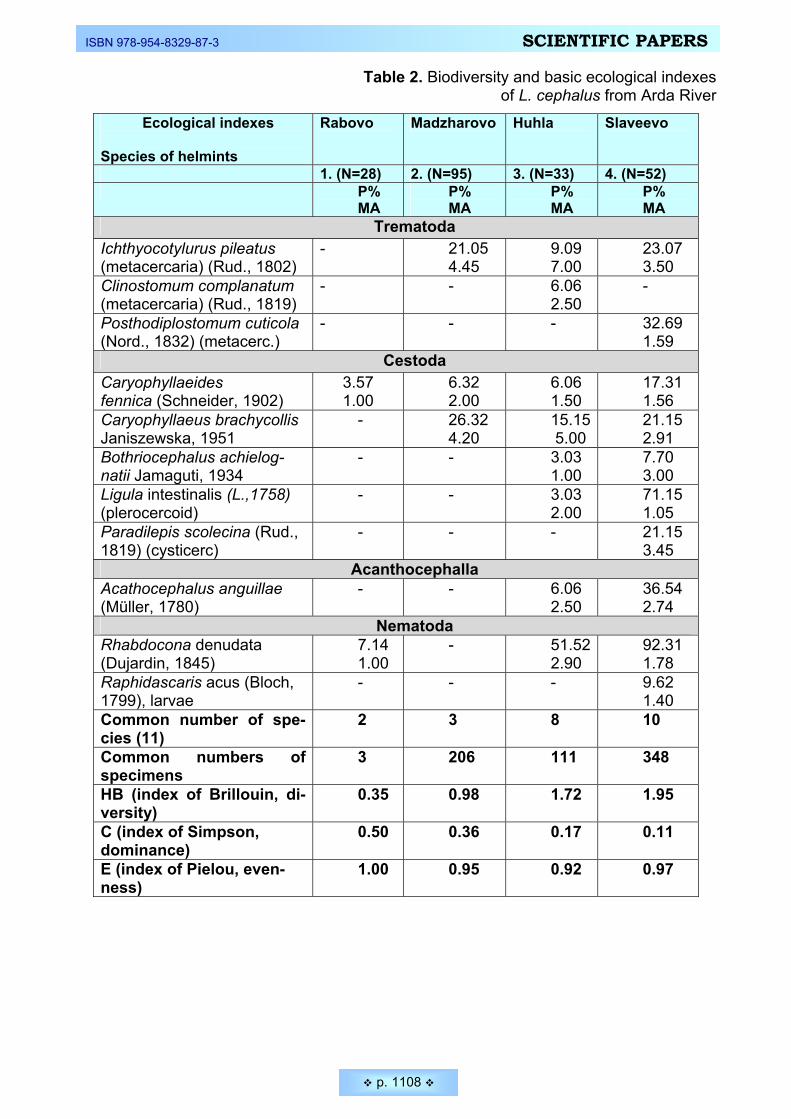
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1108
Table 2. Biodiversity and basic ecological indexes of L. cephalus from Arda River
Ecological indexes
Species of helmints
Rabovo Madzharovo Huhla Slaveevo
1. (N=28) 2. (N=95) 3. (N=33) 4. (N=52) P%
MA P% MA
P% MA
P% MA
Trematoda Ichthyocotylurus pileatus (metacercaria) (Rud., 1802)
-
21.05 4.45
9.09 7.00
23.07 3.50
Clinostomum complanatum (metacercaria) (Rud., 1819)
-
-
6.06 2.50
-
Posthodiplostomum cuticola (Nord., 1832) (metacerc.)
-
-
-
32.69 1.59
Cestoda Caryophyllaeides fennica (Schneider, 1902)
3.57 1.00
6.32 2.00
6.06 1.50
17.31 1.56
Caryophyllaeus brachycollis Janiszewska, 1951
-
26.32 4.20
15.15 5.00
21.15 2.91
Bothriocephalus achielog-natii Jamaguti, 1934
-
-
3.03 1.00
7.70 3.00
Ligula intestinalis (L.,1758) (plerocercoid)
-
-
3.03 2.00
71.15 1.05
Paradilepis scolecina (Rud., 1819) (cysticerc)
-
-
- 21.15 3.45
Acanthocephalla Acathocephalus anguillae (Müller, 1780)
-
-
6.06 2.50
36.54 2.74
Nematoda Rhabdocona denudata (Dujardin, 1845)
7.14 1.00
- 51.52 2.90
92.31 1.78
Raphidascaris acus (Bloch, 1799), larvae
- - - 9.62 1.40
Common number of spe-cies (11)
2 3 8 10
Common numbers of specimens
3 206 111 348
HB (index of Brillouin, di-versity)
0.35 0.98 1.72 1.95
C (index of Simpson, dominance)
0.50 0.36 0.17 0.11
Е (index of Pielou, even-ness)
1.00 0.95 0.92 0.97
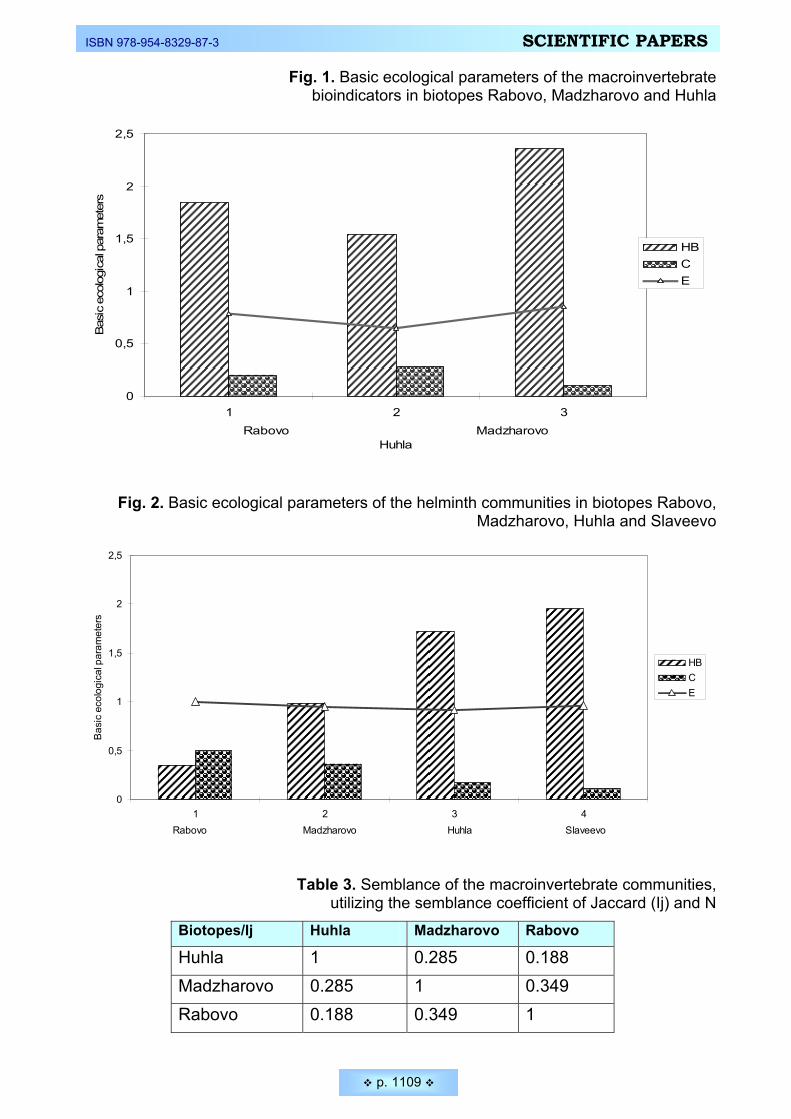
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1109
Fig. 1. Basic ecological parameters of the macroinvertebrate bioindicators in biotopes Rabovo, Madzharovo and Huhla
0
0,5
1
1,5
2
2,5
1 2 3Rabovo Madzharovo
Huhla
Bas
ic e
colo
gica
l par
amet
ers
HBCE
Fig. 2. Basic ecological parameters of the helminth communities in biotopes Rabovo, Madzharovo, Huhla and Slaveevo
0
0,5
1
1,5
2
2,5
1 2 3 4Rabovo Madzharovo Huhla Slaveevo
Bas
ic e
colo
gica
l par
amet
ers
HBCE
Table 3. Semblance of the macroinvertebrate communities, utilizing the semblance coefficient of Jaccard (Ij) and N
Biotopes/Ij Huhla Madzharovo Rabovo
Huhla 1 0.285 0.188 Madzharovo 0.285 1 0.349 Rabovo 0.188 0.349 1

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1110
Table 4. Semblance of the helminthe communities, utilizing the semblance coefficient of Ij, MA and P%
Biotopes/Ij MA Huhla Slaveevo Rabovo Madzharovo Huhla 1 0.405 0.404 0.197 Slaveevo 0.436 1 0.197 0.102 Rabovo 0.099 0.078 1 0.062 Madzharovo 0.382 0.210 0.104 1 Biotopes/Ij P% Huhla Slaveevo Rabovo Madzharovo
Fig. 3. Semblance of macro invertebrate communities, util-izing the semblance coefficient of Ij and N
Fig. 4. Semblance of the helminth communities, utiliz-ing the semblance coefficient of Ij and MA
Fig. 5. Semblance of the helminth communi-ties, utilizing the sem-blance coefficient of Ij and P%

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 1111
SCIENTIFIC PAPERS
STUDYING THE EFFECTS OF UITRAPHONOPHORESIS WITH “VURDON” – GEL AND “FELORAN” – ZEL
IN DISEASES WITH DEGENERATIVE DYSTROPHY O1 SPINAL CORD
Ivan St. Stanoev – MD.PHD
City Hospital “prof. K. Tchylov” – Madan, Bulgaria
Diana Iv. Stanoeva – MD.PHD Higher Medical Institute – Plovdiv, Bulgaria
ABSTRACT The studyis to establish the effect from application of “Vurdon” – gel and “Feloran” –
zel byindependently applied uitraphonophoresis on patients with osteochondrosis, spondy-losis and spondyloarthrosis.
Material – Method: The first group comprises 14 patients treated with ultraphono-phoresis with “Vurdon” – gel. The second group comprises 19 patients treated with uitra-phonophoresis with “Feloran”–zel.
The effect of the therapy – by objective and subjective indices. All data have been statistically processed by methods of alternative and variation analysis.
Results: From the first group 64.28 % have significant improvement, 21.42 % have just improved, and 14.28 % didn't change their condition. From the second group 63.14 % have significant improvment, 21,06 % have just improved.
Conclusions: These results show that the effect from the treatment is due to the ef-fect from the healing agents of Vurdon and Feloran gel as well as to the positive curative impact of local application of ultrasonic.
Key words: Medicina ultraphonophoresis with “Vurdon” – gel. INTRODUCTION AND OBJECT The subject of our study is the wide – spread degenerative dystrophy of spinal
cord with the subsequent temporary or continuous immobilization. Investigation is aiming at specifying the effectiveness of application of ultra-
phonophoresis with “Vurdon”–gel and “Feloran”–zel (independently applied) in pa-tients suffering from degenerative dystrophy of spinal cord (osteochondrosis, spondylosis, spondylo-artrosis)
(1,2,3,4,5,6,7) Material and methods “Vurdon”–gel is represents by “Help” Ltd. – hundred grams of it contain 1.16 g.
Diclofenac as an active substance under new pharmaceutical from (Diethylammo-nium) equivalent to 1 g of Diclofenac Sodium. Basis of gel is butyraceous non oily and non – staining emulsion that contains isopropranol and propylenglicol.

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1112
Diclofenac is a well – known composition with non – steroid structure and defi-nite anti – inflammatory and anti – rheumatic, pain relief and decrease of tempera-ture features, which are due to inhabitance of prostaglandin biosynthesis.
“Feloran” zel (TroyaPharm) Composition : Natrium-(0-)2.6 dichlor phenyl – amino-phenyl –aceticum. (Diclofenac Natrium), 1 % zel. Effects; “Feloran”–zel represent a non – steroid anti- inflamatorial remedy con-
taining phenyl aceticum acid. It excerts anti – inflammatory and anti – rheumatic pain relief and decrease of temperature features, which are due to inhabitance of prostaglandin biosynthesis.
Objects of our research are the following two groups of patients independently treated with ultraphonophoresis with “Vurdon”–gel and “Feloran”–zel.
The first group compresis 14 patients with degenerative dystrophy of spinal cord osteochondrosis, spondylosis and spondyloarthrosis treated with ultraphono-phoresis with “Vurdon”–gel. The second group comprises 19 patients treated with ultraphonophoresis with “Feloran”–zel. To obtain an impartial assessment, to com-pare results and to prove the effect of treatment we have included two other control groups that were independently treated with individual contacts substance – un-guent of Vaseline albi 30.0 g and paraffinic liquid 30.0 g.
The effectiveness of the carried out treatment is assessed by objective and subjective criteria using the following tests: Intensity of pain- strong, moderate and weak (Melzack – Canada) and relief of pain, visual examination, palpation and per-cussion to determine the inflammatory process, start inelasticity measured in min-utes, pathologic gait, X – rays to stablish degree of degenerative changes, high spirit.
Volume of movement – centimetry. Decrease of total index with more than 50.00 % shows sighificant improve-
ment, between 50.00% and 20.00 % decrease stands for improvement, whereas less than 20.00 % decrease shows lack of significant dynamics in patients status.
The whole course treatment with “Vurdon”-gel (“Feloran”–zel) comprises aver-age 10 procedures (one every day). It is carried out by the help of ultrasonic appa-ratus TUR-US-6 at impulse refimen. The procedures are accomplished by the method of labile sound after previous local skin aqpplication of “Vurdon”-gel in quantity 2-4 on the paravertebral zone.
The power of ultrasonic energy for paravertebral application is 0.2 –0.4 W/cm2 for 5–10 minutes.
Results and Discussion All data (intensity of pain, start gait in min., centimetry performed and bending
changes in gait, high spirit) have been statistically processed by alternative and variational analyses as shown in table N 1.
X–rays examination before and after 10 days treatment doesn’t show any sig-nificant difference. Favourable results are evident after 4th or 5th day of treatment with “Verdon”–gel containing 1 % Diclofenac aspecially formulated for external ap-plication. Results show that 64.28 % from the first group have significant improve-ment, 21.44 % have just improved, 14.28 % haven’t changed their condition. 63.16 % from the second group have significant improvement, 21.06% have just improved and 15.78 % didn’t change their condition. The results in control groups

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1113
are the following: 62.50 % are with significant improvement, 25.00 have improved and 12.50 % have no improvement .
The effectiveness of ultraphonophoresis is shown at table 2. The creamy form allows easier penetration in skin, while the alcohol In basis
exerts cooling and calming effect. After the local skin application of “Vurdon”–gel it Is easily absorbed and it's active substance – Diclofenac reaches the tissues un-derneath and attacks the inflammatory process with different etiology in higher plasma concentrations than those after single oral application of film – tablettes (in-formation from the manufacturer). “Feloran”–zel is resorbed in ultraphonophoresis and It's active substance excerts anti-inflammatory and pain relief effects.
The favourable effect of ultrasound is connected with it's influence over neuro –muscle apparatus thus stimulating the blood and lymph circulation, changes in membrane permeability, better trophic processes from one side and activation of the chemical compound of “Vurdon”–gel and “Feloran”–zel from other.
No local skin irritation from allergic origin is observed. It should not be applied upon skin scratches, nor any dressing after the procedure is suitable. Avoid contact with mucosa.
Conclusions 1. These results show that the effect from the treatment is due to the healing
agents of “Vurdon”–gel and “Feloran”–zel as well as to the positive curative impact of local application of ultrasonic.
2. It is evident that ultrasonic phonophoresis with “Vurdon”–gel and “Feloran”–zel can be successfully applied for diminishing and relief of pain as well as improv-ing the objective symptoms In such cases.
3. Comparative studies point out the priority of this method for maximal thera-peutic effect.
LITERATURE
[1] Dietze Christine, R. Callies and Marion Hippins: “Salizylsaurephonophorese und Salizylatnachweis” “Zeitschrift fur Physiotherapie” VEB, Georg Thieme Leipzig 1987, 2:61 – 128, 99 – 102.
[2] Smolenski U., Callies R., Manthey J. Marion Hippins “Untersuchunden zur Slizylesaure Phonophorese anhand eines Membranmodells”. “Zeitschift fur physiotheraple” VEB, Georg Thieme Leipzig 40 Jahrgan Helft 3 Mai/Juni 1988: 153-212, 171-175.
[3] Uibe P. Fritzsche G., Runge A., “Modlichkeiten der nauslischen Anwendung Phsiotherapeutischer Massnahmen bei Stotungen destutz und Bewegungsap-parates”. Zeitschrift fur Physiotherapie”. VEB Georg Thieme Leipzig, Heft L. Junuar/Februar 1990: 1-72, 45-59
[4] Unlemann G., Callies R., Hippins M. “Mehrmals tadliche salizylsaurephono-phorese”. “ Zeitschrift fur Physiotheraple”. VEB Georg Thieme, Leipzig, 40/1988/6: 365-452, 441-444.
[5] Stanoev Uvan “Ultraphonophoresis with “Ostochont” – salbe preparation in de-generative dystrophic lesions of the backbone and gonarthosis” Sofia, “Voenna Medicina” 1994 22-25

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1114
[6] Stanoev Ivan, Nikiforov N. “Ultaphonophoresis with “Velinol” preparation at some locomotion system diseases”. J. “Phys. Health Resort Rehad Med” 1994, 2&44-49
[7] Stanoev Ivan, D. Stanoeva “Studying the effect of Dynamic Thermostimulation and Ultraphonophoresis with “Kaminol” applied to patients suffering from de-generative diseases of the spinal cord”.
“Kaminol” applied to patients suffering from degenerative diseases of the spi-nal cord”.
“Balkan Military Medical Review” <BMMR> 1/1998 72-74 401 General Army Hospital – ATHENS-GREECE BETA medical publishers Ltd. – ATHENS-GREECE Dr. Ivan St. Stanoev 4004, Plovdiv Blvd. “Nikola Vapzarov” № 122 Phone : 032 / 69 71 49
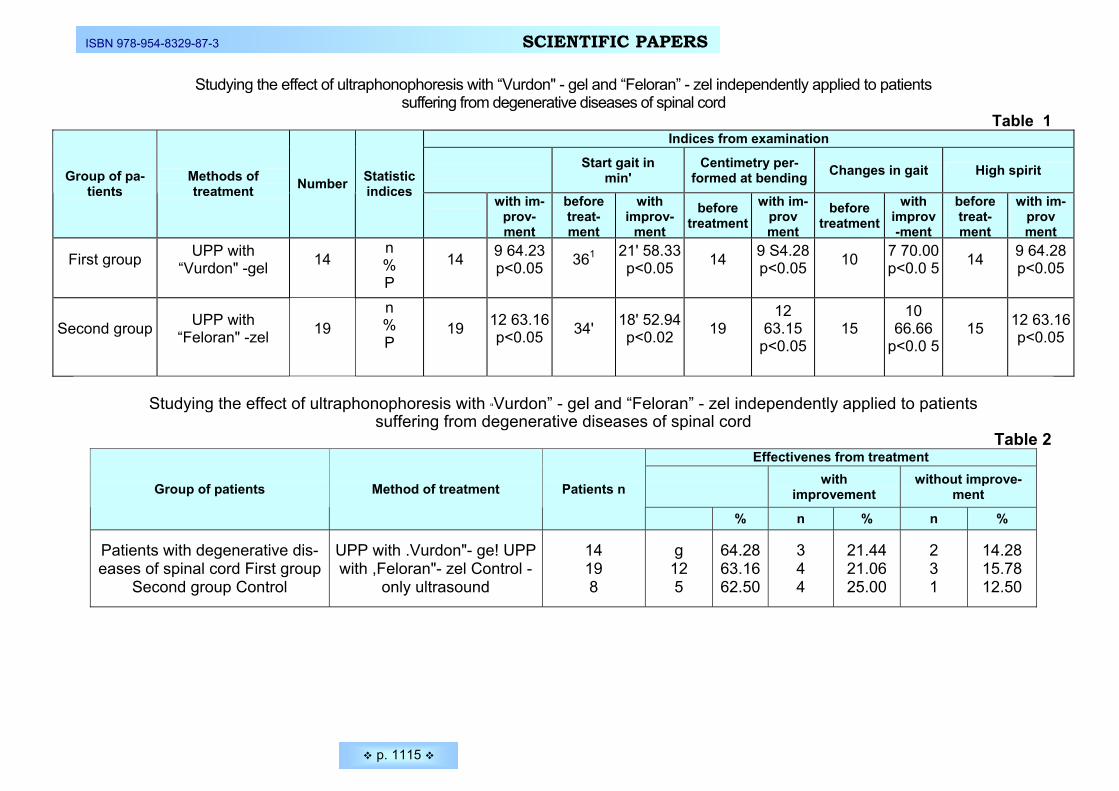
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1115
Studying the effect of ultraphonophoresis with “Vurdon" - gel and “Feloran” - zel independently applied to patients suffering from degenerative diseases of spinal cord
Table 1 Indices from examination
Start gait in
min' Centimetry per-
formed at bending Changes in gait High spirit Group of pa-tients
Methods of treatment Number Statistic
indices
with im-prov-ment
before treat-ment
with improv-
ment
before treatment
with im-prov ment
before treatment
with improv-ment
before treat-ment
with im-prov ment
First group
UPP with “Vurdon" -gel
14
n % P
14
9 64.23p<0.05
361
21' 58.33p<0.05
14
9 S4.28 p<0.05
10
7 70.00 p<0.0 5
14
9 64.28 p<0.05
Second group
UPP with “Feloran" -zel
19
n % P
19
12 63.16p<0.05
34'
18' 52.94 p<0.02
19
12 63.15
p<0.05
15
10 66.66
p<0.0 5
15
12 63.16 p<0.05
Studying the effect of ultraphonophoresis with “Vurdon” - gel and “Feloran” - zel independently applied to patients
suffering from degenerative diseases of spinal cord Table 2
Effectivenes from treatment
with improvement
without improve-ment Group of patients Method of treatment Patients n
% n % n %
Patients with degenerative dis-eases of spinal cord First group
Second group Control
UPP with .Vurdon"- ge! UPP with ,Feloran"- zel Control -
only ultrasound
14 19 8
g 12 5
64.28 63.16 62.50
3 4 4
21.44 21.06 25.00
2 3 1
14.28 15.78 12.50

UNION OF SCIENTISTS IN BULGARIA National Scientific Conference with International Participation under the heading “20 Years Union of Scientists in Bulgaria – Branch Smolyan” October, 20th–21st, 2006, Smolyan, Bulgaria ISBN 978-954-8329-87-3
p. 1116
SCIENTIFIC PAPERS
STYDING THE EFFECTIVENESS OF CLIMATICTHERAPY AND AEROSOLOTHERAPY
WITH MINERAL WATER IN PATIENTS WITH CHRONIC BRONCHITIS I N CONDITION OF VELINGRAD
Ivan St. Stanoev – MD.PHD
City Hospital “prof. K. Tchylov” – Madan, Bulgaria
ABSTRACT Velingrad is a low-mountain resort (760 – 800 m above the sea level) with moderate
continental climate and a Mediterranian influence. Mineral water from spring №10 in Velingrad is characterised as hypertermal, hydro-
carbonate –sulphatic,potassium content with fluor and variety of microelements. With a view to the actuality of the problem of this disease and qualities of the resort
Velingrad the present study depicts the medical effectiveness of physical factors (low-mountain climate, mineral water ) in 30 patients with light and moderate degree of com-mon chronic bronchitis as well to 20patients completed aerosolotherapy in out –of-a resort conditions (in industrial center).
The following clinical methods were examined: clinical symptoms, percussion, aus-cultation, functional examination of breathing on Bodyscreen and Oxyspirometer.
All data was processed by alteration and variative analysis . Clinical symptoms are best affected in patients undergoing aerosolo and climatic
therapy (84,50% of this first group) in comparison with the patients put in the conditions of the industrial center (the second group).
Functional examination of breathing on Oxyspirometer shows significant im-provement for patients from the first group and is statistically reliable.
From the second group patients 60,00% were discharged with improvement and 40,00% had none.
Aerosolo and climatric therapy with mineral water from-spring NslO in Velingrad have positive effects on patients with common chronic bronchitis – light and moderate degree and when is properly applied has no side effects.
INTRODUCTION Chronic bronchitis is of great medical and social significance (2, 10, 12, 14, 15). Climatic therapy is a contemporary method for the treatment of chronic bron-
chitis (1, 3, 4, 5, 9, 12, 13, 16, 17, 18). Velingrad is a low-mountain resort (760–800 m above the sea level) with mod-
erate continental climate and a Mediterranian influence (1, 6, 7, 8). Mineral water from spring №10 in Velingrad is characterised as hypertermal,
hydrocarbonate – sulphatic, potassium content with fluor and variety of microele-ments (1, 7).
With a view to the actuality of the problem of this disease and qualities of the resort Velingrad the present study depicts the medical effectiveness of physical fac-

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1117
tors (lowmountain climate, mineral water) in patients with light and moderate de-gree of common chronic bronchitis as well to patients completed aerosolotherapy in out –of-a resort conditions (in industrial center).
MATERIALS AND METHODS OF TREATMENT AND INVESTIGATION We have studied 50 patients with common chronic bronchitis divided into 2
groups. The first group – 30 patients treated with aerosolotherapy with mineral water
from spring NslO in Velingrad . The following scheme was applied – temperature of water – 37 °C, duration of procedure from 5-15-20 min in increasing order, 5 times a week (Hirana apparatus INH – 21) Patients are undergoing'common sanatoria! and dietetic regimen with no drugs application.
Second group – 20 patients with the same aerosolotherapy without me-diearnental application but put in the conditions of the industrial center.
The foolowing clinical methods were examined: clinical symptoms, percussion, auscultation, functional examination of breathing on Bodyscreen and Oxyspirometer.
All data was processed by alteration and variative analysis. RESULTS AND DISCUSSION Best improved are the following clinical symptoms: the symptom cough dis-
appears in 80,00 % for the first group, less present in 6,67% and no improvement in 13,33 %.
Expectoration disappears in 80,00 % for the first group, is less present in 6,67% and is not affected in 13,33 % .
Impaired breathing in physical efforts disappears in 70,59 % for this group, is less present in 11,76 % and is not affected in 17,65 %.
Statoacoustic findings disappears in 69,24 % and diminishes in 15,33 %. The second group shows the following results: symptom cough is improved in
66,67 %, expectoration in 55,56 %, impaired breathing in 62,50 %, statoacousric findings in 55,56 %.
Clinical symptoms are best affected in patients undergoing climatic therapy in comparison with the patients put in the conditions of the industrial center.
Functional examination of breathing on Oxyspirometer shows significant im-provement for patients from the first group and is statistically reliable. Best were af-fected the following indices: for VCIN p < 0,01, for MMV p < 0,01, FEV1 p < 0,05, apnoic test in inhale p < 0,05, apnoic test in exhale p < 0,05.
Functional examination of breathing for the second group showes lower levels compared to first group. Good statistical reliability show VC, MMV, ERV, FEV1, PIF, RAW kPa (Table 1).
From the first group patients 84,50 % were discharged with improvement and 15,50 % had none.
From the second group patients 60,00 % were discharged with improvement and 40,00 % had none (Table 2).
The achieved positive data from the aerosoltherapy with mineral water from spring N210 in first group patients could be explained with the climatic and sanato-rial regimen of the resort Velingrad.
Therapeutic effect in second group patients is only due to aerosolotherapy with mineral water and its chemical compound.
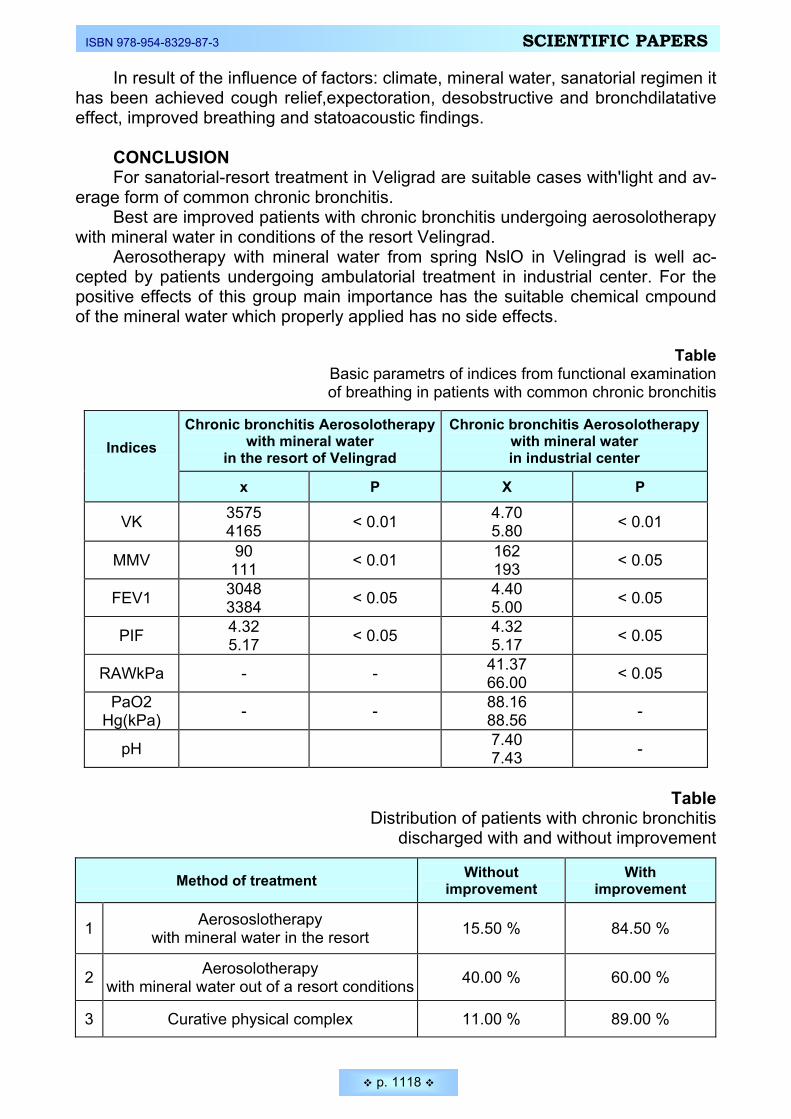
ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1118
In result of the influence of factors: climate, mineral water, sanatorial regimen it has been achieved cough relief,expectoration, desobstructive and bronchdilatative effect, improved breathing and statoacoustic findings.
CONCLUSION For sanatorial-resort treatment in Veligrad are suitable cases with'light and av-
erage form of common chronic bronchitis. Best are improved patients with chronic bronchitis undergoing aerosolotherapy
with mineral water in conditions of the resort Velingrad. Aerosotherapy with mineral water from spring NslO in Velingrad is well ac-
cepted by patients undergoing ambulatorial treatment in industrial center. For the positive effects of this group main importance has the suitable chemical cmpound of the mineral water which properly applied has no side effects.
Table Basic parametrs of indices from functional examination of breathing in patients with common chronic bronchitis
Chronic bronchitis Aerosolotherapy with mineral water
in the resort of Velingrad
Chronic bronchitis Aerosolotherapy with mineral water in industrial center
Indices
х P X P
VK 3575 4165 < 0.01 4.70
5.80 < 0.01
MMV 90 111 < 0.01 162
193 < 0.05
FEV1 3048 3384 < 0.05 4.40
5.00 < 0.05
PIF 4.32 5.17 < 0.05 4.32
5.17 < 0.05
RAWkPa - - 41.37 66.00 < 0.05
PaO2 Hg(kPa) - - 88.16
88.56 -
pH 7.40 7.43 -
Table
Distribution of patients with chronic bronchitis discharged with and without improvement
Method of treatment Without improvement
With improvement
1 Aerososlotherapy with mineral water in the resort 15.50 % 84.50 %
2 Aerosolotherapy with mineral water out of a resort conditions 40.00 % 60.00 %
3 Curative physical complex 11.00 % 89.00 %

ISBN 978-954-8329-87-3 SCIENTIFIC PAPERS
p. 1119
LITERATURE [1] Костадинов, Д. и Д. Караколев. Главни балнеологични курорти. С., Мед. и
физк., 1983, 38-53. [2] Костурков, Г., Н. Алексиев и П. Добрев. Бронхиална астма. С., Мед. и
физк., 1990, 9-19, 233-237, с246. [3] Пилипчук, Н.С. и Р.Г. Прощюк. Азр.озоль-терапия при заболеваниях
органов дыхание. Киев, Здоровья, 1988, 139-145. [4] Симеонов, Цвятко, Ст. Йонков и Гр. Найденов, Н. Никифоров. Терапия на
вътрешните заболявания с български минерални води. С., Мед. и физк., 1984, 48-65.
[5] Стаматов, Ст., Д. Караколев и Л. Цветкова. Клинична курортология. С., Мед. и физк., 1990, 130-136.
[6] Станоев, И. – М е д. д е л о, 1979, ,Р 1, 47-51. [7] Станоев, И. Резултати от някои клинично-лабораторни из-следвания при
болни от бронхиална астма след инхалационно приложение на минерална зода в условията на курорта Велинград. С., ВМА, Военна медицина, 1992, № 3, 15-20.
[8] Станчев, К. Велинград С., 1975, с. 3, 12. [9] Тодоров, Ненчо. Инхалационно лечение. С., Мед. и физк., 1987, 166-168,
с.189. [10] Федосеев, Г.Б. и А.М. Убайдуллаев. Ранная диагностика и первичная
профилактика бронхиальной астмн. Н., Медицина, 1989, 77-87, 112-120. [11] Федосеев, Г.Б. и Г.П. Хлопотова. Бронхиальная астма. М., Медицина,
1988, 5-11, 237-239. [12] Buhlo/Buhlo. Fibel der Inhalationsbebanl mit Aerosolen. Urban & Schuarzen-
berg, Munchen-Berlin-Wien , 1971. [13] Carlo Braga, Pier. Luigi Allegra, MD. Methods in Bronchial Muclology. Raven
Press – New York, 1988, 367-375. [14] Findeisen, D.G.R. Asthma bronchiale, VEB Verlag Volg und (Gesundheit, Ber-
lin, 1971, p. 297, 213-221. [15] Gershwin, M. Eric. Bronchial asthma. Principles of Diagnosis and Treatment.
Grun & Stratton. New York, London, Toronto,Sydney, San Francisco, 1981. [16] Jordan, K. Kurorttherapie. Aufl Jena. G.Fischer Verlag, 1980. [17] Jordan, K. Balneobiioklimatologie. Ein praktisches Bex-spiel ganzheiteich ori-
entierter Therapie. Zeitschrift fur Physiotherapie Georg Thieme Leipzig GmBK. 42.,1990. Heft 5. September/Oktomber, 1990, s. 277-340, 297-300.
[18] Reinhold, D. Bedentung und medizinischer inhaet der Kurorttheiapie. Zeitschrift fur Physiotherapie Georg Thieme Leipzig GmbK Heft 5 September-Oktober, 1990 . s.277-340, 287-290. Dr. Ivan St. Stanoev 4004, Plovdiv Blvd. “Nikola Vapzarov” № 122 Phone : 032 / 69 71 49





























