Embed Size (px)
Citation preview

A
vsdvwtw©
K
1
lsdCbATbaoWtTcst
1d
Available online at www.sciencedirect.com
Palaeoworld 20 (2011) 134–145
Biostratigraphy, phylogeny and paleobiogeography ofCarboniferous–Permian radiolarians in South China
Yu-Jing Wang, Qun Yang ∗Department of Micropaleontology and SKLPS, Nanjing Institute of Geology and Palaeontology, CAS, Nanjing 210008, China
Received 30 July 2010; received in revised form 17 February 2011; accepted 19 May 2011Available online 30 May 2011
bstract
The Carboniferous–Permian radiolarian faunas from the bedded chert strata of the open marine facies are characterized by superb preser-ation, higher abundance/diversity and vertical continuity and divided into 22 radiolarian zones. We found that in the Permian bedded chertytrata of the open ocean basinal facies, the genera Pseudoalbaillella and Follicucullus have undergone two distinct evolutionary pathwaysuring Guadalupian–Lopingian times: the Pseudoalbaillella ishigai–P. fusiformis–P. monacanthus lineage and the Follicucullus scholasticus–F.entricosus–F. orthogous/Follicucullus scholasticus–F. hamatus–F. bipartitus lineage. This interpretation of evolutionary pathways is in accordanceith stratigraphic relationship of the species in Pseudoalbaillella and Follicucullus. Through comparison of the faunal composition and zonation of
he Carboniferous and Permian radiolarians in South China with those of elsewhere in the world, we suggest that the Panthalassa and Paleo-Tehtysere probably merged without any distinct biotic differentiation in the open oceanic environment.2011 Elsevier Ltd and Nanjing Institute of Geology and Palaeontology, CAS. All rights reserved.
eywords: Radiolaria; Upper Paleozoic; Biostratigraphy; Morphological evolution; Paleobiogeography; China
pttdsirua
ic(die
. Introduction
Biostratigraphic studies on Carboniferous–Permian radio-arians have substantially advanced in China during the pasteveral years, with discoveries of continuous sequences of bed-ed chert formations in the provinces of South China. Thearboniferous–Permian radiolarian faunas are characterizedy preserving distinct species of the following four genera:lbaillella, Pseudoalbaillella, Follicucullus, and Neoalbaillella.hese distinct fossils are crucial to the Carboniferous–Permianiostratigraphic division and correlation for open marine faciesnd usually serve as the sole evidence for dating the ophi-lites within collision belts (Wang et al., 1994, 1998, 2006;u et al., 1994; Feng et al., 1997; Wang and Yang, 2007),
herefore, useful for determining the time of plate collision.he Carboniferous–Permian radiolarians discovered in China
ommonly occur in the chert strata of open marine facies,lope facies, and platform-basinal facies; some occur in tec-onic or sedimentary mélanges and ophiolitic sequences. As∗ Corresponding author. Tel.: +86 25 8328 2103.E-mail address: [email protected] (Q. Yang).
nGabbw
871-174X/$ – see front matter © 2011 Elsevier Ltd and Nanjing Institute of Geologoi:10.1016/j.palwor.2011.05.001
ointed out in Wang et al. (2006), the radiolarian faunas fromhe bedded chert strata of the open marine facies are charac-erized by their excellent preservation, higher abundance andiversity and vertical continuity when divided into biozones;uch radiolarian faunas are best in establishing the radiolar-an biostratigraphic zonation. Radiolarian faunas extracted fromocks of other sedimentary facies are normally less contin-ous in vertical distribution with relatively lower abundancend diversity.
This paper discusses the Carboniferous–Permian radiolar-an biostratigraphy in China and its international correlation,oncerning the 22 biozones recently published by Wang et al.2006: Lopingian radiolarian zones) and Wang and Yang (2007:escription of all Carboniferous and Permian radiolarian zonesn South China), with representative radiolarian taxa figured withlectron microphotographs. Secondly, we analyze the phyloge-etic relations among species of Genus Pseudoalbaillella andenus Follicucullus, according to their stratigraphic occurrences
nd morphological transition. Finally, we discuss the paleo-iogeographic features of the Carboniferous–Permian oceansased on radiolarian faunal characteristics and distributionorldwide.
y and Palaeontology, CAS. All rights reserved.

aeow
2b
ldC2Fd(ccsoC
F
bp12345678911113. P. ishigai Abundance Zone (P2 )14. P. globosa Assemblage Zone (P2−1
2 )2−2
Y.-J. Wang, Q. Yang / Pal
. Carboniferous and Permian radiolarianiostratigraphy in China
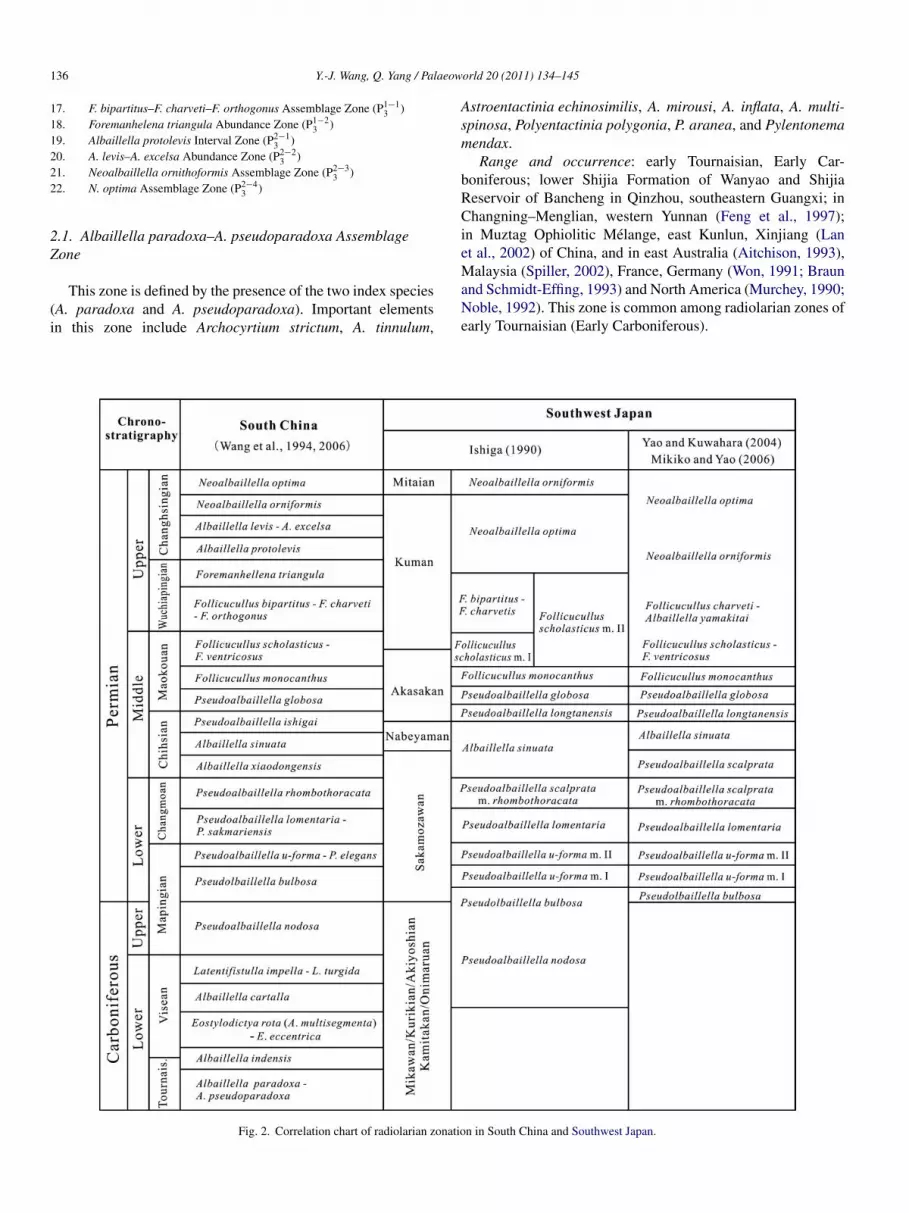
Recently, we established 22 Carboniferous–Permian radio-arian biozones primarily on the basis of abundant radiolarianata discovered in Qinzhou area, southeastern Guangxi, and inhangning–Menglian area, western Yunnan (Wang et al., 1994,006; Wang and Yang, 2007; Xie et al., 2011) (Fig. 1 herein).ollowing is a summary of the 22 radiolarian biozones and aiscussion of their age assignment and international correlationFig. 2). The 22 C–P radiolarian biozones with correspondinghronostratigraphic designation are listed below, where the ageonnotation refers to the regional chronostratigraphic sequencehown in Figs. 1 and 2 (e.g., C1−1
1 means “lower Carbonifer-1−1
us” [C1] and Tournaisian [C1 ], or ranging from the lowerarboniferous Stage 1 to Stage 1). Representative species of the
ig. 1. Radiolarian zonation in South China and ranges of zonal species. Thick vertic
11
orld 20 (2011) 134–145 135
iozones are figured in Figs. 3 and 4 (the electron micrographiclates).. Albaillella paradoxa–A. pseudoparadoxa Assemblage Zone (C1−1
1 ). Albaillella indensis Range Zone (C1−2
1 ). Eostylodictya rota–E. eccentrica Assemblage Zone (C2−1
1 ). Albaillella cartalla Assemblage Zone (C2−2
1 ). Latentifistula impella–L. turgida Abundance Zone (C2−3
1 ). Pseudoalbaillella nodosa Assemblage Zone (C2). P. bulbosa Assemblage Zone (P1−1
1 ). P. u-forma–P. elegans Abundance Zone (P1−2
1 ). P. lomentaria–P. sakmarensis Assemblage Zone (P1−3
1 )0. P. rhombothoracata Interval Zone (P1−4
1 )1. Albaillella xiaodongensis Interval Zone (P1−1
2 )2. A. sinuata Abundance Zone (P1−2
2 )1−3
al bars are abundant intervals; dotted vertical lines are rare range intervals.
5. P. monacanthus Interval Zone (P2 )6. Follicucullus scholasticus–F. ventricosus Assemblage Zone (P2−3
2 )
1 aeow
111222
2Z
(i
Asm
bRCieMalaysia (Spiller, 2002), France, Germany (Won, 1991; Braun
36 Y.-J. Wang, Q. Yang / Pal
7. F. bipartitus–F. charveti–F. orthogonus Assemblage Zone (P1−13 )
8. Foremanhelena triangula Abundance Zone (P1−23 )
9. Albaillella protolevis Interval Zone (P2−13 )
0. A. levis–A. excelsa Abundance Zone (P2−23 )
1. Neoalbaillella ornithoformis Assemblage Zone (P2−33 )
2. N. optima Assemblage Zone (P2−43 )
.1. Albaillella paradoxa–A. pseudoparadoxa Assemblageone
This zone is defined by the presence of the two index speciesA. paradoxa and A. pseudoparadoxa). Important elementsn this zone include Archocyrtium strictum, A. tinnulum,
aNe
Fig. 2. Correlation chart of radiolarian zonatio
orld 20 (2011) 134–145
stroentactinia echinosimilis, A. mirousi, A. inflata, A. multi-pinosa, Polyentactinia polygonia, P. aranea, and Pylentonemaendax.Range and occurrence: early Tournaisian, Early Car-
oniferous; lower Shijia Formation of Wanyao and Shijiaeservoir of Bancheng in Qinzhou, southeastern Guangxi; inhangning–Menglian, western Yunnan (Feng et al., 1997);
n Muztag Ophiolitic Mélange, east Kunlun, Xinjiang (Lant al., 2002) of China, and in east Australia (Aitchison, 1993),
nd Schmidt-Effing, 1993) and North America (Murchey, 1990;oble, 1992). This zone is common among radiolarian zones of
arly Tournaisian (Early Carboniferous).
n in South China and Southwest Japan.

Y.-J. Wang, Q. Yang / Palaeoworld 20 (2011) 134–145 137
Fig. 3. Micrographs of representative species from the Carboniferous and Permian radiolarian zones in South China. All scale bars = 100 �m. Sample localitiesinclude: Shi26 and Jb from the Shijiang Reservoir section near Bancheng Township in Qinzhou, southeastern Guangxi; Ta4, 94-111, 94-113, 94-119, 94-127, 94-131,94-147; 93-3, 93-4, 93-6, 93-7, 93-8, 93-11, 93-12, 93-15, 93-16, and 93-22 from the Tiache section near Bancheng; 96St236 from the Shiti Reservoir section nearBancheng (refer to Wang et al., 2006). (A) Albaillella shijiangensis Wang, Shi26; (B) A. pseudoparadoxa Won, Shi26; (C) A. paradoxa Deflandre, Shi26; (D) A.tuboforma Won, 96St236; (E) Archocyrtium ferreum Braun, 96St236; (F) Pylentonema mendax (Deflandre), 96St236; (G, U) Astroentactinia multispinosa (Won), G,96St236; U, 93-8; (H, Y) A. mirousi Gourmelon, H, 96St236; Y, 93-15; (I, J, AA) Albaillella indensis Won, I, J, Jb; AA, 93-16; (K, N), A. crenulata Won; K, Jh; N, 93-6;(L, M) A. cartalla Ormiston and Lane, 93-6; (O) Popofskyellum annulatum Deflandre, 93-6; (P) Polyentactinia arenea Gourmelon, 93-6; (Q) Astroentactinia formosaWang, 93-6; (R, CC) A. radiata Braun, R, 93-6; CC, 93-16; (S), Tetragregnon sycamorensis Ormiston and Lane, 93-6; (T, DD) Latentifistula impella (Ormiston andLane), T, 93-7; DD, Ta4; (V, W) Albaillella multisegmenta Wang, 93-11; (X) Latentifistula turgida (Ormiston and lane), 93-12; (Z) Ceratoikiscum umbraculum Won,93-16; (BB) Belowea variabilis (Ormiston and Lane), 93-16; (EE, GG) Pseudoalbaillella bulbosa Ishiga, EE, 94-147; GG, 94-127; (FF) P. sakmarensis (Kozur),94-131; (HH) Latentifistula mushroomformis Wang, 93-22; (II) Ruzhencevispongus uralicus Kozur, 93-3; (JJ) Pseudoalbaillella reflexa Ling and Forsythe, 94-113;(KK, SS) P. u-forma Holdsworth and Jones, KK, 94-113; SS, 94-119; (LL, MM) P. elegans Ishiga and Imoto, 94-113; (NN, OO) P. triangularis Wang, 94-113; (PP,Q rmist
2
sp
ml
Q) P. nodosa Ishiga, 94-111; (RR) Triactofenestrella nicolica (Nazarov and O
.2. Albaillella indensis Range Zone
This zone is defined by the abundant occurrence of the zonalpecies Albaillella indensis. Main associated species are com-osed of A. crenulata, A. tuboforma, A. ferreum, Pylentonema
A
l
on), 93-4.
endax, P. racheboeufi, P. banchengensis, Stigmosphaerosty-us vulgaris, S. densa, Archocyrtium wonae, A. lagabriellei,
strocentactinia radiata, and Latentifistula impella.Range and occurrence: early-middle Early Carboniferous;ower–middle Shijia Formation of Wanyao, Shijia Reservoir,

138 Y.-J. Wang, Q. Yang / Palaeoworld 20 (2011) 134–145
Fig. 4. Micrographs of representative species from the Carboniferous and Permian radiolarian zones in South China. Scale bars = 50 �m for OO and PP; scalebars = 100 �m for all other SEM micrographs. Sample localities include: 94-73 and 94-74 from the Shijiang Reservoir section near Bancheng Township in Qinzhou,southeastern Guangxi; 94-17 and Che21-1 from the Jiazhiping section near Bancheng; 92B15, 92B27 and 92B42 from the Shiti Reservoir section near Bancheng;D6, D9, D22, 92 × 13, 92 × 16, 92 × 22, 92 × 23, 92 × 24, 92 × 45, 92 × 49, 92 × 51, 92 × 55, 92 × 57, 92 × 59, 92 × 62, 92 × 72 and 92 × 76 from the Sanpaolingsection near Xiaodong Village, Qinzhou; 92-147B and 92-165 from the Nanban section and 91LC111 from the Laochang section in Lancang County, western Yunnan(refer to Wang et al., 2006). (A) Pseudoalbaillella chilensis Ling and Forsythe, 94-116; (B) P. reflexa Ling and Forsythe, Che21-1; (C, D) P. ishigai Wang, 92B27;(E, F) Albaillella sinuata Ishiga and Watase, 92B-27; (G, L) Pseudoalbaillella sakmarensis (Kozur), G, 94-17; L, 92B-15; (H, K, X) Albaillella xiaodongensis Wang,H, 94-73; K, 94-74; X, 92B-10; (I, J) Pseudoalbaillella rhombothoracata Ishiga and Imoto, 94-74; (M, R) P. globosa Ishiga and Imoto, M, 92 × 13; R, 92 × 76; (N,O) P. fusiformis (Holdsworth and Jones), 92 × 13; (P, Q) P. internata Wang, 92 × 16; (S, T) P. longtanensis Sheng and Wang, 92 × 76; (U, SS) Hegleria mammilla(Sheng and Wang), U, D6; SS, 92 × 72; (V, W) P. monacanthus (Ishiga and Imoto), D22; (Y, Z, AA) Follicucullus scholasticus Ormiston and Babcock, Y, 92 × 51; Zand AA, 92B42; (BB) F. porrectus Rudenko, 92 × 45; (CC, DD) F. ventricosus Ormiston and Babcock, CC, 92 × 45; DD, 92 × 49; (EE, FF) F. charveti Caridroit andDe Wever, EE, 92 × 23; FF, 92 × 24; (GG, HH) F. orthogonus Caridroit and Dever, GG, 92 × 57; HH, 92 × 59; (II, JJ, KK) F. bipartitus Cardrot and De Wever, II,92 × 23; JJ, 92 × 22; KK, 92 × 24; (LL, MM) F. guangxiensis Wang, n. sp., LL, 92 × 62; MM, 92 × 55; (NN), Albaillella levis Ishiga, Kito and Imoto, 92-165B; (OO,PP) A. protolevis Kuwahara, 92 × 72; (QQ) A. yamakitai Kuwahara, 92 × 24; (RR) Copicyntra akikawaensis Sashida and Tonishi, 92 × 57; (TT) Foremanhelenatriangula De Wever and Caridroit, D9; (UU) F. circula Shang, Caridroit and Wang, 92-165; (VV) Triplanospongos musashiensis Sashida and Tonishi, 92-147B;(WW) Latentifistula similicutis Caridrot and De Wever, 92 × 57; (XX) Neoalbaillella ornithoformis Takemura and Nakaseko, 91LC111; (YY) N. optima Ishiga, Kitoand Imoto, 92LB165; (ZZ) N. pseudogrypa Sashida and Tonishi, 92-147B.

aeow
TCCB1iC
2
sSla
SeX(bC
2
siSiAm
bSCi(11z(
2
smoeCIv
oQw
((Adz
2
Pr
BeCFC
2
sfgLi
miwT1Ssbz
2
sfiuZP
PieCA1992), Canada (Harms and Murchey, 1992), Malaysia (Spiller,
Y.-J. Wang, Q. Yang / Pal
iache, Shiti Reservoir in Qinzhou, southeastern Guangxi;hangning–Menglian, western Yunnan; Hoh Xil, Qinghai ofhina; east Australia (Aitchison, 1993), Germany (Won, 1991;raun and Schmidt-Effing, 1993) and France (Gourmelon,987). This zone has been considered a common radiolar-an zone of middle–late Tournaisian–early Visean (Earlyarboniferous).
.3. Eostylodictya rota–E. eccentrica Assemblage Zone
This zone is characterized by the presence of the indexpecies Eostylodictya rota and E. eccentrica, associated withtigmosphaerostylus vulgaris, S. variospina, Albaillella crenu-ata, Latentifistula impella, L. turgida, Astroentactinia radiata,nd Belowea variabilis.
Range and occurrence: late Early Carboniferous; middlehijia Formation of Tiache, Bancheng in Qinzhou, southeast-rn Guangxi and in Gengma, Manxin, and Zhongzai, Banshun,iarao, Changning–Menglian, western Yunnan and in Germany
Won, 1991; Braun and Schmidt-Effing, 1993). This zone mayecome a standard radiolarian zone of middle Visean (Earlyarboniferous).
.4. Albaillella cartalla Assemblage Zone
This zone is defined by the presence of the indexpecies Albaillella cartalla, with primary associated radiolar-an taxa including Stigmosphaerostylus vulgaris, S. variospina,. gourmelonae, S. tortispina, Albaillella crenulata, A.nflate, A. spinosa, A. ouachitaensis, Protoalbaillella media,stroentactinia digitosa, A. inflata, A. spatiosa, and A.irousi.Range and occurrence: middle–late Visean (Early Car-
oniferous); middle–upper Shijia Formation of Wanyao,hijia Reservoir, Tiache and Shiti Reservoir of Bancheng,hengwang in Yulin, Qinzhou, southeastern Guangxi;
n Changning–Menglian, western Yunnan; east AustraliaAitchison, 1993), Malaysia (Spiller, 2002), Germany (Won,991; Braun and Schmidt-Effing, 1993), France (Gourmelon,987) and North America (Murchey, 1990; Noble, 1992). Thisone is a common radiolarian zone of middle–late ViseanEarly Carboniferous).
.5. Latentifistula impella–L. turgida Abundance Zone
This zone is defined by the abundant occurrence of the zonalpecies Latentifistula impella and L. turgida, accounting forore than 50% of the whole radiolarian fauna in terms of number
f specimens, which first appeared in the Eostylodictya rota–E.ccentrica Assemblage Zone and flourished in late Visean Earlyarboniferous age. Sometimes, they have formed Radiolarites.
n the associated species of this zone, only one species, Beloweaariabilis, exists.
Range and occurrence: late Visean (Early Carbonifer-us); upper Shijia Formation of Wanyao; Shijia Reservoir, ininzhou, southeastern Guangxi and in Changning–Menglian,estern Yunnan; east Australia (Aitchison, 1993), Malaysia
2Ei(
orld 20 (2011) 134–145 139
Spiller, 2002), North America (Noble, 1992) and GermanyWon, 1983; Braun and Schmidt-Effing, 1993). Besides, thelbaillella nazarovi zone from the Hagong Formation in Zhong-ian, northwestern Yunnan of China may correspond to thisone.
.6. Pseudoalbaillella nodosa Assemblage Zone
This zone is defined by the presence of the zonal index speciesseudoalbaillella nodosa, and by the absence of any associated
adiolarian species in our specimens.Range and occurrence: Late Carboniferous; base of
ancheng Formation of Wanyao and Tiache, in Qinzhou, south-astern Guangxi and in Southwest Japan (Ishiga, 1986, 1990),anada (Harms and Murchey, 1992) and South Chile (Ling andorsythe, 1987). This zone is the only radiolarian zone of Latearboniferous discovered in South China.
.7. Pseudoalbaillella bulbosa Assemblage Zone
This zone is defined by the presence of the zonal indexpecies Pseudoalbaillella bulbosa, associated mainly with theollowing radiolarian species: P. reflaxa, P. simplex, P. trian-ularis, P. chilensis, P. elegans, Latentifistula mushroomformis,atentidiota trigemena, L. fallacia, and Latentibifistula nicol-
ca.Range and occurrence: early Wolfcampian (Early Per-
ian); lower Bancheng Formation of Tiache, Dasi and Migong,n Qinzhou, southeastern Guangxi of China, and in South-est Japan (Ishiga, 1986, 1990), America (Murchey, 1990),hailand (Sashida et al., 1998) and Chile (Ling and Forsythe,987). In Chile, this zonal species is associated with fusulinidchwagerina aff. munaniensis, Chalaroschwagerina tarltonen-is, Triticites tarltonensis in same section. This zone marks theeginning of the Permian period and is a common radiolarianone of early Wolfcampian (Early Permian).
.8. Pseudoalbaillella u-forma–P. elegans Abundance Zone
This zone is defined by the abundant presence of the zonalpecies Pseudoalbaillella u-forma and P. elegans, which arerst found in the P. bulbosa Assemblage Zone and may extendpwards into the P. lomentaria–P. sakmarensis Assemblageone. Main associated radiolarian species include P. reflaxa,. triangularis, and P. chilensis.
Range and occurrence: early–middle Wolfcampian (Earlyermian); lower Bancheng Formation of Tiache and Migong
n Qinzhou, southeastern Guangxi; Changning–Menglian, west-rn Yunnan and in Huashixia Mélange of Maduo, Qinghai ofhina; Southwest Japan (Ishiga, 1986, 1990), West Coast ofmerica (Holdsworth and Jones, 1980; Murchey and Jones,
002), Thailand (Sashida et al., 1993, 1998) and the Farast of Russia (Rudenko and Panasenko, 1990). This zone
s a common radiolarian zone of early-middle WolfcampianEarly Permian).

1 aeow
2A
(sa
mSieSS1a(z
2
dl
lTQJ(c
2
Art
mSGJAl
2
zxPa
mRQ
eN1coo
2
dAPe
mSi(MCota
2
sflLir
mSKCJ1(OaPaetr
2
40 Y.-J. Wang, Q. Yang / Pal
.9. Pseudoalbaillella lomentaria–P. sakmarensisssemblage Zone
This zone is defined by the presence of the two index speciesP. lomentaria and P. sakmarensis). A few associated radiolarianpecies consist of P. longicornis, P. scalprata, P. postscalprata,nd Nazarovella sp.
Range and occurrence: middle–late Wolfcampian (Early Per-ian); lower Bancheng Formation of Shijia Reservoir, Tiache,hiti Reservoir and Migong in Qinzhou, southeastern Guangxi;
n Hoh Xil area of Qinghai; in Changning–Menglian, west-rn Yunnan; in Shuanghe of Qiangtan, Tibet of China, and inouthwest Japan (Ishiga, 1986, 1990), Oregon of the Unitedtates (Blome and Reed, 1992), Canada (Harms and Murchey,992), Ural and the Far East of Russia (Kozur, 1981; Rudenkond Panasenko, 1990), Malaysia (Spiller, 2002), and ThailandSpiller, 2002). This zone has become a common radiolarianone of middle-late Wolfcampian (Early Permian).
.10. Pseudoalbaillella rhombothoracata Interval Zone
This zone is defined by the presence of the index species Pseu-oalbaillella rhombothoracata, associated with P. scalprata, P.ongicornis, P. inflata, and P. postscalprata.
Range and occurrence: late Wolfcampian (Early Permian);ower Bancheng Formation of Shijia and Shiti Reservoirs,iache of Bancheng, Sanpaoling of Xiaodong and Migong ininzhou, southeastern Guangxi of China, and in Southwest
apan (Ishiga, 1986, 1990), Malaysia (Spiller, 2002), ThailandSashida et al., 1993), the Far East of Russia. This zone is aommon radiolarian zone of late Wolfcampian (Early Permian).
.11. Albaillella xiaodongensis Interval Zone
This zone is defined by the presence of the zonal index specieslbaillella xiaodongensis, associated mainly with the following
adiolarian species: P. scalprata, P. longicornis, Latentifistulaexana, Nazarovella scalae, and Albaillella sinuata.
Range and occurrence: early Leonardian (Middle Permian);iddle Bancheng Formation of Shiti Reservoir of Bancheng,hapaoling of Xiaodong and Migong in Qinzhou, southeasternuangxi; in Hoh Xil area of Qinghai of China and in Southwest
apan. Stratigraphically, this zone lies below the lower Chihsianlbaillella sinuata Abundance Zone, and may correspond to the
ower Leonardian of America.
.12. Albaillella sinuata Abundance Zone
This zone is defined by the abundant occurrence of theonal species Albaillella sinuate, which first appeared in the A.iaodongensis Interval Zone. Main associated species include. scalprata, P. longicornis, A. xiaodongensis, A. asymmetrica,nd Stigmosphaerostylus sp. B.
Range and occurrence: middle Leonardian (Middle Per-ian); middle Bancheng Formation of Tiache and Shitieservoir of Bancheng, Shapaoling of Xiaodong and Migong ininzhou, southeastern Guangxi; in Changning–Menglian, west-
d(m
orld 20 (2011) 134–145
rn Yunnan of China; in Southwest Japan (Ishiga, 1986, 1990),orth America (Blome and Reed, 1992; Murchey and Jones,992) and Malaysia (Spiller, 2002). Ishiga and Suzuki (1984)onsidered this zone to be of Leonardian age based on the con-dont data. This zone has become a common radiolarian zonef middle Leonardian (Middle Permian).
.13. Pseudoalbaillella ishigai Abundance Zone
This zone is defined by the abundant zonal species Pseu-oalbaillella ishigai, which first occurred in the A. sinuatabundance Zone; main associated species include P. scalprata,. longicornis, A. inflata, Stigmosphaerostylus sp. B, Nazarov-lla scalae.
Range and occurrence: late Leonardian Middle Permian;iddle Bancheng Formation of Shiti Reservoir of Bancheng andhapaoling of Xiaodong in Qinzhou, southeastern Guangxi; and
n Shuanghe of Qiangtan, Tibet of China; in Southwest JapanIshiga, 1986, 1990), North America (Blome and Reed, 1992),
alaysia (Spiller, 2002). This zone may compare with the upperhihsian stage of South China, and is also regarded to be a syn-nym of the P. sp. C zone (Ishiga et al., 1982; Ishiga, 1986) andhe P. longtanensis zone (Ishiga, 1990). This zone may becomestandard fossil zone in late Leonardian Middle Permian age.
.14. Pseudoalbaillella globosa Assemblage Zone
This zone is defined by the presence of the zonal indexpecies Pseudoalbaillella globosa, associated mainly with theollowing radiolarian species: P. fusiformis, P. yanaharensis, P.ongicornis, P. longtanensis, P. internata, Hegleria mammilla,atentifistula texana, Stigmosphaerostylus itsukachiensis, Cop-
cyntra akikawaensis, and Pseudotormentus monoporus. Theadiolarites commonly occur in this zone.
Range and occurrence: early Guadalupian (Middle Per-ian); upper Bancheng Formation of Shiwu of Bancheng andhapaoling of Xiaodong in Qinzhou, southeastern Guangxi;uhfeng Formation of South Jiangsu and Anhui, Fujian;hangning–Menglian, western Yunnan of China; Southwest
apan (Ishiga, 1986, 1990), North America (Blome and Reed,992; Murchey and Jones, 1992), North Island of New ZealandMurchey and Jones, 1992), Malaysia (Spiller, 2002), Thailand,man (De Wever et al., 1988), Crete Island of Greece (Kozur
nd Krahl, 1987) and the Far East of Russia (Rudenko andanasenko, 1990; Rudenko et al., 1997). Although Ishiga (1986)nd Blome and Reed (1992) considered this zone to be of the lat-st Leonardian to early early Guadalupian age, we suggest thathis zone is restricted to early early Guadalupian strata based onegional stratigraphy in South China.
.15. Pseudoalbaillella monacanthus Interval Zone
This zone is defined by the presence of the index species Pseu-oalbaillella monacanthus. It often contains only one speciesthe zonal species), sometimes associated with a few of Hegleriaammilla, P. yanaharensis, and Latentifistula texana.

aeow
moeCJ1aSm
2Z
wtOm
mBeiJ11Oa(o
2A
smitCgmSdI
afUZarPata
dtfs
up“fwwo
2
soiotsdSs
uQT
2
AgDms
miCn(At
2
bw
Y.-J. Wang, Q. Yang / Pal
Range and occurrence: middle Guadalupian (Middle Per-ian); upper Bancheng Formation of Tiache and Shiti Reservoir
f Bancheng and Shapaoling of Xiaodong in Qinzhou, south-astern Guangxi; Kuhfeng Formation in Chaohu of Anhui; inhangning–Menglian, western Yunnan of China; Southwest
apan (Ishiga, 1986, 1990), North America (Blome and Reed,992), Malaysia (Spiller, 2002), the Far East of Russia (Rudenkond Panasenko, 1990) and Sicily Island of Italy (Kozur, 1993).ometimes, this zone contains only the zonal species; it is a com-on radiolarian zone of middle Guadalupian (Middle Permian).
.16. Follicucullus scholasticus–F. ventricosus Assemblageone
This zone is defined by the presence of the two index species,ith primary associated radiolarian taxa including F. porrec-
us, Nazarovella gracilis, N. inflata, N. scalae, N. phlogidea,rmistonella robusta, Ishigaum obesum, and Pseudotormentusonoporus.Range and occurrence: late Guadalupian (Middle Per-
ian); upper Bancheng Formation of Tiache and Shiwu ofancheng and Shapaoling of Xiaodong in Qinzhou, southeast-rn Guangxi; Kuhfeng Formation of South Jiangsu and Anhui;n Changning–Menglian, western Yunnan of China; Southwestapan (Kuwahara et al., 1998), North America (Blome and Reed,992; Murchey and Jones, 1992), Canada, Philippines (Cheng,989), Malaysia (Spiller, 2002), Thailand (Sashida et al., 1997),man (De Wever et al., 1988), the Far East of Russia (Rudenko
nd Panasenko, 1990), and the North Island of New ZealandTakemura et al., 1999). This zone is a common radiolarian zonef late Guadalupian (Middle Permian).
.17. Follicucullus bipartitus–F. charveti–F. orthogonusssemblage Zone
This zone is defined by the presence of the three indexpecies, F. bipartitus, F. charveti, and F. orthogonus, whichay sometimes occur separately in the same stratigraphic
nterval. Important elements in this zone include Latentifis-ula similicultis, L. texana, Latentibifistula asperspongiosa,opicyntra akikawaensis, Hegleria mammilla, Nazarovellaracilis, N. sealae, N. inflata, Ormistonella robusta, Fore-anhelena triangular, Stigmosphaerostylus itsukachiensis,. tyrrelli, Pseudotormentus kamigoriensis, Trilonche pseu-ocimelia, Ruzhencevispongus uralicus, Quadricaulis femoris,shigaum trifustis, Follicucullus furca, and F. quadrataris.
Three recent studies (Nestell and Nestell, 2010; Maldonadond Noble, 2010; Noble and Jin, 2010) on radiolarian faunasrom the Middle Permian Guadalupian limestones in Texas,SA, indicate the presence of the F. bipartitus–F. ventricosusone, which is assigned to late Guadalupian in age. However,s shown in Fig. 2 herein, the similar radiolarian zones, occur-ing in chert facies, are assignable to latest Guadalupian (Middle
ermian) to earliest Lopingian (Late Permian) in South Chinand Southwest Japan (refer to Wang et al., 2006 for age con-rols using associated conodonts and fusulinid zones in the studyreas). More precise correlation of the radiolarian zones amongAamL
orld 20 (2011) 134–145 141
ifferent sedimentary facies will require further investigation onhe radiolarian assemblages assisted by associated age controlossils such as conodonts or other age control mechanisms (e.g.,tratigraphic context and/or radiometric dates).
Range and occurrence: early Wuchiapingian (Late Permian);pper Bancheng Formation of Shiwu of Bancheng and Sha-aoling of Xiaodong in Qinzhou of southeastern Guangxi;Kuhfeng Formation” of Tongtianyan and Sanmenjiang treearm in Liuzhou of Guangxi; Nanban and Nanpihe in Lancang,estern Yunnan; Lekang in Wangmo, Guizhou of China; South-est Japan and Philippines. This is a common radiolarian zonef early Wuchiapingian (Late Permian).
.18. Foremanhelena triangula Abundance Zone
This zone is defined by the abundant occurrence of the zonalpecies Foremanhelena triangula, accounting for more than 30%f the whole radiolarian fauna in terms of number of spec-mens, which first appears in the F. bipartitus–F. charveti–F.rthogonus Assemblage Zone and may extend upwards intohe Neoalbaillella optima Assemblage Zone. Main associatedpecies Foremanhelena circula, Quadricaulis femoris, Pseu-otormentus kamigoriensis, Latentibifistula asperspongiosa,tigmosphaerostylus itsukachiensis, and Copicyntra akikawaen-is.
Range and occurrence: late Wuchiapingian (Late Permian);pper Bancheng Formation of Shapaoling of Xiaodong ininzhou, southeastern Guangxi and “Kuhfeng Formation” ofongtianyuan in Liuzhou, Guangxi.
.19. Albaillella protolevis Interval Zone
This zone is defined by the presence of the index specieslbaillella protolevis, associated with Nazarovella inflata, Ishi-aum trifustis, Latentifistula similicultis, Albaillella yamakitai,eflandrella manica, Hegleria mammilla, Triplanospongosusashiensis, Copicyntra akikawaensis, and Trilonche cras-
ispinosa.Range and occurrence: early Changhsingian (Late Per-
ian); upper Bancheng Formation of Shapaoling of Xiaodongn Qinzhou, southeastern Guangxi; Longwangpo in Nandanounty, Guangxi; Laochang in Lancang County, western Yun-an; Hoh Xil area, North Tibet of China; Southwest JapanKuwahara et al., 1997; Kuwahara, 1999), West Coast of Northmerica (Noble and Renne, 1990), Philippines (Cheng, 1989),
he Far East of Russia (Rudenko et al., 1997).
.20. Albaillella levis–A. excelsa Abundance Zone
This zone is defined by the presence of the abundanted of the zonal species Albaillella levis and A. excelsa,hich may extend upwards into the Neoalbaillella optima
ssemblage Zone. Main associated species include A. tri-ngularis, A. lauta, Copicyntra akikawaensis, Deflandrellaanica, Foremanhelena triangula, Latentifistula similicultis,atentibifistula asperspongiosa, Pseudotormentus kamigorien-
1 aeow
si
Pa(1(
2
stdPcm
PaiS1(S1i(
2
sftchga
LGiCa1rg
3F
ogd
tim
(
(
bcacpvdp
Ftttt
42 Y.-J. Wang, Q. Yang / Pal
is, Spongosphaeradiscus shaiwaensis, Stigmosphaerostylustsukaichiensis, and Triplanospongos musashiensis.
Range and occurrence: early–middle Changhsingian (Lateermian); Longwangpo in Nandan County, Guangxi; Nanbannd Ali in Lancang County, western Yunnan; Southwest JapanSugamori, 2011), Philippines (Cheng, 1989; Tumanda et al.,990), Malaysia (Sashida et al., 1995) and the Far East of RussiaRudenko et al., 1997).
.21. Neoalbaillella ornithoformis Assemblage Zone
This zone is defined by the presence of the zonalpecies Neoalbaillella ornithoformis, associated with A.riangularis, A. lauta, Copicyntra akikawaensis, Deflan-rella manica, Neoalbaillella gracilis, N. pseudogrypa,seudotormentus kamigoriensis, Ruzhencevispongus urali-us, Spongosphaeradiscus shaiwaensis, and Triplanospongosusashiensis.Range and occurrence: middle–upper Changhsingian (Late
ermian); Longwangpo in Nandan County, Guangxi; Nanbannd Laochang in Lancang County, western Yunnan; Sidazhain Ziyun County and Mogang in Luodian County, Guizhou;huanghu in Qiangtan, Tibet of China; Japan (Kuwahara et al.,998; Nakae and Kurihara, 2011; Sugamori, 2011), MalaysiaSashida et al., 1995; Spiller, 2002), Thailand (Sashida andalyapongse, 2002), the Far East of Russia (Rudenko et al.,997), and North America (Blome and Reed, 1992, 1995). Thiss a common radiolarian zone of middle-upper ChanghsingianLate Permian).
.22. Neoalbaillella optima Assemblage Zone
This zone is defined by the presence of the zonal indexpecies Neoalbaillella optima, associated mainly with theollowing radiolarian species: A. levis, A. triangularis, Copicyn-ra shaiwaensis, Stigmosphaerostylus itsukaichiensis, Triloncherassispinosa, Follicucullus porrectus, F. charveti, Foreman-elena triangula, F. circula, Neoalbaillella gracilis, Spon-osphaeradiscus shaiwaensis, Triplanospongos musashiensis,nd Triaenosphaera minuta.
Range and occurrence: late Changhsingian (Late Permian);ongwangpo in Nandan County and Liuqiao in Fusui County,uangxi; Nanban in Lancang County, western Yunnan; Lekang
n Wangmo County, and Sanchahe and Moyang south in Luodianounty, Guizhou of China; Japan (Kuwahara et al., 1998; Nakaend Kurihara, 2011), Philippines (Cheng, 1989; Tumanda et al.,990), Thailand (Sashida and Salyapongse, 2002). This zone isegarded as the uppermost radiolarian zone of late Changhsin-ian (latest Permian).
. Phylogenic relationship of Pseudoalbaillella andollicucullus in Middle–Late Permian
We found that in the Permian bedded cherty strata of the opencean basinal facies in South China (Wang et al., 2006), theenera Pseudoalbaillella and Follicucullus have undergone twoistinct evolutionary pathways during Guadalupian-Lopingian
cicr
orld 20 (2011) 134–145
imes (Fig. 5). The evolutionary lineages of Pseudoalbaillellashigai–P. fusiformis–P. monacanthus include the following
orphological transitions:
1) The P. ishigai–P. fusiformis lineage involves transitions ofthe decrease of the number of pseudoabdominal segments.P. ishigai of late Leonardian is characterized by having anapical cone, a pseudothorax with two lateral wings and pseu-doabdominal segments, of which the first segment is smalland short and the fifth one is skirt-like. In latest Leonardianto early Guadalupian, the descent species P. longtanensishas four pseudoabdominal segments, which in turn evolvedinto the further descent species P. fusiformis and P. globosa,both species with three pseudoabdominal segments and astrongly inflated globose pseudothorax.
2) The P. fusiformis–P. monacanthus lineage involves transi-tions of decreasing and disappearance of the lateral ventralwing on the pseudothorax. P. fusiformis appears in the upper-most Leonardian and flourishes in the lower Guadalupian.In middle Guadalupian, a transitional form (a new speciesto be named), characterized by the presence of a shrunklateral ventral wing (a slightly convex-like structure) onthe pseudothorax and a blade-like dorsal wing, links P.fusiformis and P. monacanthus, the latter species possessingno structure at the position of the ventral wing but with adorsal wing. We re-assign here the above-mentioned species(monacathus) to Genus Pseudoalbaillella instead of GenusFollicucullus as originally proposed because of the appar-ent evolutionary transitions as discussed above. Apparently,the two genera are closely related, transitional or insepara-ble (De Wever et al., 2001). The whole Pseudoalbaillellafauna, dominating the cherty strata of Upper Carboniferousthrough Guandalupian (Middle Permian), terminates at theposition where P. monacanthus disappears near the top ofGuadalupian Series.
The evolution of Follicucullus of late Guadalupian seems toegin at the most primitive species F. scholasticus, which isharacterized by having a simple straight conical shell withoutnnulations. F. scholasticus gave rise to F. porrestus throughonical shell weakly differentiating into the apical cone and theseudothorax with small apertural spines. On the other hand, F.entricosus was derived from F. scholasticus through markedifferentiation of the shell composed of the apical cone, theseudothorax, and the skirt-like apertural expansion.
F. scholasticus could have given rise to the F. hamatus and the. bipartitus through increased curvature of the apical shell ando F. burca through the bifurcating of the dorsal spine at the aper-ure. F. porrectus gave rise to F. quadrataris and F. quangxiensishrough changes of the pseudothoracic shape and the orienta-ion of the ventral apertural spine. F. ventricosus gave rise to F.
harveti and to F. orthogonus through the morphological mod-fication of the skirt-like apertural structure and orientationalhanges of the ventral spine. F. ventricosus may also have givenise to F. falx through morphological changes of the ventral spine.
Y.-J. Wang, Q. Yang / Palaeoworld 20 (2011) 134–145 143
Fig. 5. Phylogenetic relationship based on interpretation of morphological transitions in the stratigraphic context for some Pseudoalbaillella and Follicucullus inG
ad
4r
tdoTtfCKoGndt
nWaCeC
eT
LoaCisassPfbri
oCcffTtC
uadalupian and Lopingian strata in South China.
The above interpretation of evolutionary pathways is inccordance with stratigraphic relationship of the species in Pseu-oalbaillella and Follicucullus.
. Paleobiogeography of Carboniferous–Permianadiolarians
The Carboniferous–Permian radiolarian faunas are dis-ributed worldwide. Ross and Ross (1990) suggested thaturing Carboniferous–Permian time, there are two majorceanic basins on the Earth: the Panthalassa and the Paleo-ethys; the Paleo-Tethyan faunas migrated eastwards through
he Panthalassa Ocean. The Early Carboniferous radiolarianauna is found mainly in Qinzhou, southeastern Guangxi; inhangning–Menglian and Zhongdian, western Yunnan; in eastunlun and south Tianshan, Xinjiang; in Hoh Xil, Qinghaif China; and in east Australia, Malaysia, Thailand, Turkey,ermany, France and the West Coast of North America;evertheless, Early Carboniferous radiolarians have not beeniscovered in Southwest Japan and Philippines, which belongedo the Panthalassa Ocean.
On the other hand, the Early Carboniferous radiolarian fau-as found in the New England Orogen, east Australia, and theest Coast of North America are obviously derived from the
ccretionary complexes of the Mesozoic. Because the Earlyarboniferous radiolarian faunas found in Qinzhou, southeast-rn Guangxi and in Changning–Menglian, western Yunnan ofhina, show strong affinities with those of west Europe (Feng
TJC
t al., 1997), we suggest that west Europe belonged to the Paleo-ethys Ocean during Early Carboniferous.
Because the widespread glacial-marine event during theate Carboniferous and Early Permian caused the extinctionf the Early Carboniferous radiolarian fauna, no fossilsre found in the grayish yellow mudstone of lower Upperarboniferous in Qinzhou, southeastern Guangxi. However,
n the thinner cherty strata of upper Upper Carboniferoustrata, a new radiolarian species, Pseudoalbaillella nodosa,ppears. This species, which has been found in Qinzhou,outheastern Guangxi and in Southwest Japan, Canada andouth Chile, led to a newly emerging radiolarian fauna (theseudoalbaillella–Follicucullus–Albaillella–Neoalbaillella
aunas) of the Permian. Almost no Permian radiolarians haveeen found in west Europe, except for sporadic Middle Permianadiolarians discovered on islands (e.g., Crete and Sicilyslands) (Kozur and Krahl, 1987; Kozur, 1993).
Catalano et al. (1991) suggested that the mass occurrencesf radiolarians (with dominating Albaillellacea and missingopicyntrinae) are characteristic of deep-sea deposits of theircumpacific realm, but a large number of Copicyntrinae and aew of Albaillellacea are characteristic for the Ural radiolarianauna of the shallower marine environment in Early Permian.he 15 radiolarian zones of Southwest Japan are equivalent
o the seventeen zones of China (Fig. 2), ranging from Latearboniferous to late Changhsingian (Late Permian) in age.
he elements of the Copicyntrinae, which occur in Southwestapan representing the Panthalassa Ocean, also occur in Southhina belonging to the Paleo-Tethys. Apparently, for this time
1 aeow
iCnz
s(bAtgitpdm
A
LnLfooprmnN
R
A
B
B
B
C
C
D
D
F
G
H
H
I
I
I
I
K
K
K
K
K
K
L
L
M
M
M
M
N
44 Y.-J. Wang, Q. Yang / Pal
nterval (Late Carboniferous to the end of Permian), Southhina and Southwest Japan share identical radiolarian faunasot only in the faunal component but also in the radiolarianonation, indicating coeval faunal transitions.
After studying a number of radiolarian-bearing stratigraphicections in Southwest Japan and South China, Kuwahara et al.1997) and Kuwahara (1999) suggested that the faunal differenceetween the Panthalassa and the Paleo-Tethys was not detected.s Scotese and Langford (1995) suggested little is known about
he supposedly huge oceanic Basin—Panthalassa. The biostrati-raphic data of the Carboniferous–Permian radiolarian faunandicate that the Late Carboniferous–Permian radiolarian bios-ratigraphy is characterized by probably a single Paleo-Tethyslanktonic faunal realm with little biogeographic differentiationetected. This implies that the Late Carboniferous to Permianajor ocean basins were probably unified.
cknowledgements
We thank Mr. Yin Bao’an (Guangxi Regional Geological Mr.i Jiaxiang (Guangxi Geological Institute, Nanning), Dr. Yen-ien Cheng (National Museum of Natural History, Taiwan), Dr.uo Hui (Nanjing Institute of Geology and Palaeontology, CAS)
or participating in part of the field collections and for discussionn Paleozoic radiolarians, and Dr. Ma Junye (Nanjing Institutef Geology and Palaeontology, CAS) for assistance in graphicrocessing. Paula Noble and Kuei-yu Yeh are thanked for theireview and constructive comments that surely have improved theanuscript. This study has received financial support from Chi-
ese Academy of Sciences (Project No. KZCX2-SW-129) andatural Science Foundation of China (Grant No. 40172004).
eferences
itchison, J., 1993. Albaillellaria from the New England Orogen, eastern NSW,Australia. Marine Micropaleontology 21, 353–367.
lome, C.D., Reed, K.M., 1992. Permian and Early Triassic radiolarian faunasfrom the Grindstone Terrane, central Oregon. Journal of Paleontology 66,351–383.
lome, C.D., Reed, K.M., 1995. Radiolarian biostratigraphy of the Quinn RiverFormation, Black Rock terrane, north-central Nevada: Correlations witheastern Klamath terrane geology. Micropaleontology 41, 49–68.
raun, A., Schmidt-Effing, R., 1993. Biozonation, diagensis and evolution ofradiolarians in the Lower Carboniferous of Germany. Marine Micropaleon-tology 21, 369–383.
atalano, R., Stephano, R.D., Kozur, H., 1991. Permian Circum-Pacific deep-water faunas from the western Tethys (Sicily, Italy)—new evidences fromthe position of the Permian Tethys. Palaeogeography, Palaeoclimatology,Palaeoecology 87, 74–108.
heng, Y.N., 1989. Upper Paleozoic and Lower Mesozoic radiolarian assem-blages from the Busuanga Islands, North Palawan Block, Philippines.Bulletins of National Museum of Natural Science 1, 129–175.
e Wever, P., Bourdillon-de Grissac, C., Bechennec, F., 1988. Permian agefrom radiolarites of the Hawasina nappes, Oman Mountains. Geology 16,912–914.
e Wever, P., Dumitrica, P., Caulet, J.P., Nigrini, C., Caridroit, M., 2001. Radio-
larians in the Sedimentary Record. Gordon and Breach Science Publishers,Amsterdam, 527 pp.eng, Q.L., Ye, M., Zhang, Z.J., 1997. Early Carboniferous radiolarians fromwestern Yunnan. Acta Micropalaeontologica Sinica 14 (1), 79–92.
N
orld 20 (2011) 134–145
ourmelon, F., 1987. Les Radiolaires Tournaisiens des Nodules phosphatés de laMontagne Noire et des Pyrénées Centrales. Biostratigraphie du Paléozoïque6, 1–194 (in French).
arms, T.A., Murchey, B.L., 1992. Setting and occurrence of Late Paleozoicradiolarians in the Sylvester allochthon, part of a proto-Pacific ocean floorterrane in the Canadian Cordillera. Palaeogeography, Palaeoclimatology,Palaeoecology 96, 127–139.
oldsworth, B.K., Jones, D.L., 1980. Preliminary radiolarian zonation for LateDevonian through Permian time. Geology 8 (6), 281–285.
shiga, H., 1986. Late Carboniferous and Permian radiolarian biostratigraphy ofSouthwest Japan. Journal of Geoscience, Osaka City University 29, 89–100.
shiga, H., 1990. Paleozoic radiolarians. In: Ichikawa, K., Mizutani, S., Hara, I.,Hada, S., Yao, A. (Eds.), Pre-Cretaceous Terranes of Japan. Nippon Insatsu,Osaka, pp. 285–295.
shiga, H., Kito, Y., Imoto, N., 1982. Permian radiolarian biostratigraphy. Newsof Osaka Micropaleontologists 5, 17–26.
shiga, H., Suzuki, S., 1984. Discovery of Permian radiolarians and conodontsfrom the Shimomidani Formation in the “Maizuru Belt” south-west Japanand its significance. Earth Science (Chikyu Kagaku) 38, 197–206.
ozur, H., 1981. Albaillellidea (Radiolaria) aus dem Unterperm des Vorurals.Geologisch-Paläontogishe Mitteilungen Innsbruck 10, 263–274.
ozur, H., 1993. Upper Permian radiolarians from the Sosio Valley Area, westernSicily (Italy) and from the Uppermost Lamar Limestone of West Texas.Jahrbuch der Geologischen Bundesanstalt Wien 136 (1), 99–123.
ozur, H., Krahl, J., 1987. Erster Nachweis von Radiolarien im tethyalen PermEuropas. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen174, 357–372.
uwahara, K., 1999. Middle and Late Permian radiolarian assemblages fromChina and Japan. In: Yao, A., Ezaki, Y., Hao, W.C., Wang, X.P. (Eds.),Biotic and Geological Development of the Paleo-Tethys in China. PekingUniversity Press, Beijing, pp. 43–54.
uwahara, K., Yao, A., An, T.X., 1997. Paleozoic and Mesozoic complexes inthe Yunnan area, China (Part 1): preliminary report of Middle–Late Permianradiolarian assemblages. Journal of Geoscience, Osaka City University 40,37–49.
uwahara, K., Yao, A., Yamakita, S., 1998. Reexamination of Upper Permianradiolarian biostratigraphy. Earth Science (Chikyu Kagaku) 52, 391–404.
an, C.L., Wu, J., Li, J.L., Yu, L.J., Li, H.S., Wang, Y.T., 2002. Discovery ofEarly Carboniferous radiolarians in Muztag ophiolitic mélange, East Kun-lun, Xinjiang. Scientia Geologica Sinica 37 (1), 104–106 (in Chinese, withEnglish abstract).
ing, H.Y., Forsythe, R.D., 1987. Late Paleozoic pseudoalbaillellid radiolar-ians from southernmost Chile and their geologic significance. In: Elliot,D.H., Collinson, J.W., Mckenzie, G.D. (Eds.), Gondwana Six. AmericanGeophysical Union, Geophysical Monograph Series, pp. 253–260.
aldonado, A.L., Noble, P.J., 2010. Radiolarians from the upper Guadalu-pian (Middle Permian) reef trail member of the Bell Canyon Formation,West Texas and their biostratigraphic implications. Micropaleontology 56,69–115.
ikiko, S., Yao, A., 2006. Lower–Middle Permian radiolarian biostratigraphy inthe Qinzhou area, South China. Journal of Geoscience, Osaka City University49 (3), 31–47.
urchey, B.L., 1990. Age and depositional setting of siliceous sediments in theupper Paleozoic Havallah sequence near Battle Mountain, Nevada; implica-tions for the paleogeography and structural evolution of the western marginof North America. In: Harwood, D.S., Miller, M. (Eds.), Paleozoic and EarlyMesozoic Paleogeographic Relations: Sierra Nevada, Klamath Mountains,and Related Terranes. Geological Society of America Special Paper 255, pp.137–155.
urchey, B.L., Jones, D.L., 1992. A mid-Permian chert event: widespread depo-sition of biogenic siliceous sediments in coastal, island arc and oceanicbasins. Palaeogeography, Palaeoclimatology, Palaeoecology 96, 161–174.
akae, S., Kurihara, T., 2011. Direct age determination of an Upper Permianaccretionary complex (Kirinai Formation), Kitakami Mountains, Northeast
Japan. Palaeoworld 20, doi:10.1016/j.palwor.2010.12.003.estell, G.P., Nestell, M.K., 2010. Late Capitanian (latest Guadalupian, MiddlePermian) radiolarians from the Apache Mountains, West Texas. Micropale-ontology 56, 7–68.

aeow
N
N
N
R
R
R
S
S
S
S
S
S
S
S
T
T
W
W
W
W
W
W
W
X
Y.-J. Wang, Q. Yang / Pal
oble, P.J., 1992. Biostratigraphy of the Caballos Novaculite-Tesnus forma-tion boundary, Marathon Basin, Texas. Palaeogeography, Palaeoclimatology,Palaeoecology 96, 141–151.
oble, P.J., Jin, Y., 2010. Radiolarians from the Lamar Limestone, GuadalupeMountains, West Texas. Micropaleontology 56, 117–147.
oble, P.J., Renne, P., 1990. Paleoenvironmental and biostratigraphic signif-icance of siliceous microfossils of the Permo-Triassic bedding section,eastern Klamath Mountains, California. Marine Micropaleontology 15,379–391.
oss, J.R.P., Ross, C.A., 1990. Late Paleozoic bryozoan biogeography. In:McKerrow, W.S., Scotese, C.R. (Eds.), Paleozoic Paleogeography and Bio-geography. Geological Society of London Memoirs 12, 353–362.
udenko, B.C., Panasenko, E.C., 1990. Permian Albaillellaria (Radiolaria) ofthe Pantovyi Creek sequence in Primorye. In: Tisarena, G.I. (Ed.), New Dataon Paleozoic and Mesozoic Biostratigraphy of the South Far East. USSRAcademy of Sciences, Far-eastern Branch, Vladivostock, pp. 181–193 (inRussian).
udenko, V.S., Panasenko, E.S., Rybalka, S.K., 1997. Radiolaria fromPermian–Triassic boundary beds in cherty deposits of primorye (Sikhote-Alin). In: Dickins, J.M., Yang, Z.Y., Yin, H.F., Lucas, S.S., Acharyya, S.K.(Eds.), Late Palaeozoic and Early Mesozoic Circum-Pacific Events and TheirGlobal Correlation. Cambridge University Press, Cambridge, pp. 147–151.
ashida, K., Salyapongse, S., 2002. Permian radiolarian faunas from Thailandand their paleogeographic significance. Journal of Asian Earth Sciences 20(6), 691–701.
ashida, K., Igo, H., Hisada, K., Nakornsri, N., Ampornmaha, A., 1993. Occur-rence of Paleozoic and Early Mesozoic Radiolaria in Thailand (preliminaryreport). Journal of Southeast Asia Earth Science 8, 97–108.
ashida, K., Adachi, S., Igo, H., Koike, T., Ibrahim, A.B., 1995. Middle and LatePermian radiolarians from the Semanggol Formation, northwest PeninsularMalaysia. Transactions and Proceedings of the Palaeontological Society ofJapan, New Series 177, 43–58.
ashida, K., Adachi, S., Igo, H., Nokornsri, N., Ampornmaha, A., 1997. Middleto Upper Permian and Middle Triassic radiolarians from eastern Thailand.Science Reports of Institute of Geosciences, University of Tsukuba, SeriesB 18, 1–17.
ashida, K., Igo, H., Adachi, S., Ueno, K., Nokornsri, N., Sardsud, A., 1998.Late Paleozoic radiolarian faunas from northern and northeastern Thailand.
Science Reports of Institute of Geosciences, University of Tsukuba, SeriesB 19, 1–27.cotese, C.R., Langford, R.P., 1995. Pangea and the paleogeography of the Per-mian. In: Scholle, P.A., Peryt, T.M., Ulmer-Scholle, D.S. (Eds.), The Permian
Y
orld 20 (2011) 134–145 145
of Northern Pangea Volume 1: Paleogeography, Paleoclimates, Stratigraphy.Springer-Verlag, pp. 3–19.
piller, F.C.P., 2002. Radiolarian biostratigraphy of Peninsular Malaysia andimplications for regional palaeotectonics and palaeogeography. Palaeonto-graphica Abt. A 266 (1–3), 1–91.
ugamori, Y., 2011. Late Permian radiolarians from the Ajima Formation of theUltra-Tamba Terrane in the Sasayama area, Southwest Japan. Palaeoworld20, doi:10.1016/j.palwor.2011.05.004.
akemura, A., Morimoto, T., Aita, Y., Hori, R.S., Higuchi, Y., Spörli, K.B.,Campbell, H., Kodama, K., Sakai, T., 1999. Permian Albaillellaria (Radi-olaria) from a limestone lens at the Arrow Rocks in the Waipapa terrane,Northland, New Zealand. Geodiversitas 2 (4), 751–765.
umanda, F.P., Sato, T., Sashida, K., 1990. Preliminary Late Permian radiolarianbiostratigraphy of Busuanga Island, Palawan, Philippines. Annual Reportsof the Institute of Geosciences, University of Tsukuba 16, 39–45.
ang, Y.J., Yang, Q., 2007. Carboniferous–Permian radiolarian biozones ofChina and the palaeobiogeographic implication. Acta MicropalaeontologiaSinica 24 (4), 338–346 (in Chinese, with English abstract).
ang, Y.J., Cheng, Y.N., Yang, Q., 1994. Biostratigraphy and systematics ofPermian radiolarians in China. Palaeoworld 4, 172–202.
ang, Y.J., Luo, H., Kuang, G.D., Li, J.X., 1998. Late Devonian–Late Per-mian strata of cherty facies at Xiaodong and Bancheng countries of theQinzhou area, SE Guangxi. Acta Micropalaeontologica Sinica 15, 351–366(in Chinese, with English abstract).
ang, Y.J., Yang, Q., Cheng, Y.N., Li, J.X., 2006. Lopingian (Upper Permian)radiolarian biostratigraphy of South China. Palaeoworld 15, 31–53.
on, M.Z., 1983. Radiolarien aus dem Unterkarbon des Rheinischen Schiefer-gebirges (Deutschland). Palaeontographica Abt. A 182, 116–175.
on, M.Z., 1991. Phylogenetic study of some species of Genus AlbaillellaDeflandre 1952 and a radiolarian zonation in the Rheinische Schiefer-geirge, West Germany. Journal of the Paleontological Society of Korea 1,13–25.
u, H.R., Xian, X.Y., Kuang, G.D., 1994. Late Paleozoic radiolarian assem-blages of southern Guangxi. Scientia Geologica Sinica 29 (4), 339–345 (inChinese, with English abstract).
ie, L., Yang, W.Q., Liu, G.C., Feng, Q.L., 2011. Late Paleozoic Radi-olaria from the Upper Triassic sedimentary mélange in Shangrila,Southwest China and its geological significance. Palaeoworld 20,
doi:10.1016/j.palwor.2011.05.003.ao, A., Kuwahara, K., 2004. Radiolarian fossils from the Permian–Triassicof China. News of Osaka Micropaleontologists, Special Volume 13,29–45.





























