Embed Size (px)
Citation preview

Blockade of CD49d inhibits allergic airway pathologiesindependent of effects on leukocyte recruitment
MICHAEL T. BORCHERS,1 J. CROSBY,1 S. FARMER,1 J. SYPEK,2
T. ANSAY,1 N. A. LEE,1 AND J. J. LEE1
1Department of Biochemistry and Molecular Biology, Mayo Clinic Scottsdale,Scottsdale, Arizona 85259; and 2Genetics Institute, Incorporated,Cambridge, Massachusetts 02140Received 29 August 2000; accepted in final form 1 November 2000
Borchers, Michael T., J. Crosby, S. Farmer, J. Sypek,T. Ansay, N. A. Lee, and J. J. Lee. Blockade of CD49dinhibits allergic airway pathologies independent of effects onleukocyte recruitment. Am J Physiol Lung Cell Mol Physiol280: L813–L821, 2001.—Lymphocyte and/or eosinophil re-cruitment is dependent on the sequential interactions be-tween adhesion molecules expressed on activated endothelialcells and both leukocyte subtypes. Endothelial P- and E-selectins mediate tethering and rolling of leukocytes throughinteractions with P-selectin glycoprotein ligand-1 (PSGL-1),and diapedesis subsequently occurs by engagement of endo-thelial vascular cell adhesion molecule-1 and CD49d (a4-integrins). The anti-inflammatory potential of interferingwith these adhesive interactions was assessed with anovalbumin challenge mouse model of asthma. Administrationof a soluble form of PSGL-1 reduced eosinophils (80%) andlymphocytes (50%) in bronchoalveolar lavage fluid withoutaffecting epithelial changes or airway hyperreactivity (AHR).In contrast, although administration of anti-CD49d monoclo-nal antibodies (PS/2) resulted in similar reductions in eosin-ophils (75%) and lymphocytes (50%), PS/2 reduced andabolished mucous cell metaplasia and AHR, respectively.Administration of both PSGL-1 and PS/2 had the additiveeffect of eliminating eosinophils from the airways (96% de-crease), with few or no additional reductions (relative to PS/2administration alone) in lymphocyte recruitment, mucouscell metaplasia, or AHR. These data show that eosinophilsand lymphocytes differentially utilize adhesive interactionsduring recruitment and that the inhibition of AHR is inde-pendent of this recruitment.
airway hyperreactivity; a4-integrin; P-selectin glycoproteinligand-1
ALLERGIC AIRWAY INFLAMMATION is characterized by air-way and peribronchial leukocyte accumulation thatconsists of eosinophils and lymphocytes. It is widelybelieved that lymphocytes, through the production ofTh2 cytokines [i.e., interleukin (IL)-4, -5 and -13], me-diate the recruitment and activation of eosinophils(34). In allergic airway diseases such as asthma, eosin-ophils have been hypothesized to secrete toxic granuleproteins and other inflammatory mediators that con-
tribute to the hallmark features of the disease, such asexcess mucus production and airway dysfunction (8).Sensitization and aerosol challenge of mice withovalbumin (Ova) is a common and well-characterizedmethod used to produce phenotypic changes associatedwith human asthma, including mucous cell metapla-sia, airway hyperreactivity (AHR), and accumulationof eosinophils and lymphocytes in the lung (7, 10, 11,24). The protocols used to generate these phenotypesvary in timing and the number of sensitizations andchallenges required to elicit these phenotypes. Al-though specific studies may be limited because theypresent data from a single strain of mice under one setof conditions, these studies nonetheless display fea-tures of human asthma and thus constitute relevantmodels of allergic pulmonary inflammation.
The current paradigm of eosinophil-mediated in-flammation in response to allergen involves the in-creased production, maturation, and release of eosino-phils from the marrow into the blood and increasedadhesion to and migration through the capillary endo-thelium. The cell-cell interactions that mediate selec-tive eosinophil migration are complex, overlapping pro-cesses controlled by families of cytokines, adhesionmolecules, and chemoattractants (32). Leukocyte adhe-sion and migration through the vascular endotheliuminvolve the sequential tethering, rolling, and firm ad-hesion along the endothelium followed by diapedesisbetween endothelial cells and directed migration intothe tissue (27). The initial binding and subsequent firmadhesion of eosinophils have been shown to be medi-ated by binding to P-selectin and vascular cell adhesionmolecule-1 (VCAM-1) receptors on endothelial cells,respectively (14). Eosinophils bind to these receptors,predominantly through their cell surface ligands P-selectin glycoprotein ligand-1 (PSGL-1) (29) and het-erodideric integrins comprised of the a4-subunit(CD49) (33).
Previous studies with P-selectin-deficient mice havedemonstrated an important role for P-selectin in aller-gic pulmonary inflammation, including tissue eosino-
Address for reprint requests and other correspondence: J. J. Lee,Dept. of Biochemistry and Molecular Biology, SCJMRB-Research,Mayo Clinic Scottsdale, 13400 E. Shea Blvd., Scottsdale, AZ 85259(E-mail: [email protected]).
The costs of publication of this article were defrayed in part by thepayment of page charges. The article must therefore be herebymarked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
Am J Physiol Lung Cell Mol Physiol280: L813–L821, 2001.
1040-0605/01 $5.00 Copyright © 2001 the American Physiological Societyhttp://www.ajplung.org L813

philia (5, 7) and AHR (7). Similarly, studies have es-tablished a role for CD49d in pulmonary inflammationin several species including sheep (2), guinea pigs (23),and mice (19). Furthermore, CD49d-blocking antibod-ies have also been shown to attenuate AHR after aller-gen challenge in these species (2, 11, 23).
The tethering, rolling, and adhesion of leukocyteswere initially characterized as processes distinctly reg-ulated by unique families of endothelial cell receptors(i.e., selectins and immunoglobulin gene superfamilyadhesion receptors) (27); however, increasing evidencesuggests that there is an overlap in function of thesereceptors in the tethering and firm adhesion of eosin-ophils to the endothelium (12, 20). The objective of thisstudy was to determine whether simultaneously block-ing PSGL-1/P-selectin and CD49d/VCAM-1 receptor-
ligand interactions in a murine model of allergic pul-monary inflammation significantly reduces observedleukocyte recruitment, mucous cell metaplasia, andAHR compared with the disruption of either interac-tion individually. The data presented demonstrate thatintravenous coadministration of a soluble recombinantform of human PSGL-1 (rsPSGL-Ig) (28) and rat anti-mouse CD49d IgG (PS/2) before allergen challenge hadthe additive effect of eliminating airway and parenchy-mal eosinophilia, without additional reductions of re-cruited lymphocytes, compared with either antagonistadministered alone. Administration of rsPSGL-Ig hadno effect on mucous cell metaplasia or AHR. In con-trast, PS/2 significantly reduced mucous cell metapla-sia and abolished AHR. Coadministration of these an-tagonists further inhibited the development of mucouscells along the airways and also abolished AHR. These
Fig. 1. Human recombinant P-selectin glycoprotein ligand (rsPSGL-Ig) and rat anti-mouse CD49d monoclonal antibody (PS/2) equallyinhibit ovalbumin (Ova)-induced eosinophil (A) and lymphocyte (B)accumulation in the airway lumen but differentially affect leukocytetrafficking when used in combination. Bronchoalveolar lavage (BAL)was performed 48 h after the last of 3 Ova challenges, and totalrecovered eosinophils and lymphocytes were enumerated. Values aremeans 6 SE; n 5 8–12 mice/group. The administration of PS/2 aloneor PS/2 and rsPSGL-Ig together had no effect on leukocyte counts insaline-challenged mice (PS/2 5 0 eosinophils and 1.7 6 0.28 3 103
lymphocytes; PS/2 plus rsPSGL-Ig 5 0 eosinophils and 1.6 6 0.3 3103 lymphocytes). *Significantly different from BSA/Ova, P , 0.05.†Significantly different from PS/2/Ova, P , 0.05.
Fig. 2. rsPSGL-Ig and PS/2 inhibition of Ova-induced eosinophil (A)and lymphocyte (B) accumulation in the lung tissue parallels effectsobserved on BAL fluid accumulations. Collagenase digestion of per-fused lungs was performed 48 h after the last challenge, and totalrecovered eosinophils and lymphocytes were enumerated. Values aremeans 6 SE; n 5 8–12 mice/group. The administration of PS/2 aloneor PS/2 and rsPSGL-Ig together had no effect on leukocyte counts insaline-challenged mice (PS/2 5 0.20 6 0.04 3 106 eosinophils and0.84 6 0.11 3 107 lymphocytes; PS/2 plus rsPSGL-Ig 5 0.20 6 0.03 3106 eosinophils and 0.90 6 0.15 3 107 lymphocytes).*Significantlydifferent from BSA/Ova, P , 0.05. †Significantly different fromPS/2/Ova, P , 0.05.
L814 ANTI-CD49D AFFECTS AHR INDEPENDENT OF INFLAMMATION

observations suggest that blockade of CD49d reducesallergic pulmonary pathologies through a mecha-nism(s) independent of its effects on leukocyte recruit-ment.
MATERIALS AND METHODS
Experimental design. BALB/cJ mice (female, 6–8 wks;Jackson Laboratories, Bar Harbor, ME) were sensitized byan intraperitoneal injection (100 ml) of 20 mg of chicken Ova(Sigma, St. Louis, MO) emulsified in 2 mg of Imject Alum[Al(OH)3-Mg(OH)2] (Pierce, Rockford, IL) on days 0 and 14.Mice were subsequently challenged with an aerosol gener-ated from 1% Ova in saline or saline alone for 20 min byultrasonic nebulization (DeVilbiss, Somerset, PA) on days 24,25, and 26. Assessments of inflammation and pulmonarypathology, including mucous cell metaplasia and AHR, weremade on day 28 (n 5 8–12 mice/group). rsPSGL-Ig is arecombinant, chimeric construct containing the 47 NH2-ter-minal amino acid sequence of human PSGL-1 fused to amutated human IgG1 Fc heavy chain fragment (GeneticsInstitute, Cambridge, MA). The molecule is predominantly
dimeric and has been demonstrated to have binding activityto P-selectin in vitro (26). rsPSGL-Ig and/or PS/2 (AmericanType Culture Collection, Manassas, VA) was administeredvia the tail vein 3 h before each aerosol challenge. Theoptimal doses for in vivo neutralization of murine allergicinflammation (rsPSGL-Ig, 100 mg in 50 ml; PS/2, 200 mg in 50ml) were determined in earlier studies (11, 19, 28). All micewere maintained in microisolator cages housed in a specificpathogen-free animal facility. The sentinel cages within thisanimal colony were negative for viral antibodies and thepresence of known mouse pathogens. Protocols and studiesinvolving animals were conducted in accordance with Na-tional Institutes of Health and Mayo Clinic Foundationguidelines.
Assessment of allergic inflammation. Allergic inflamma-tion was assessed on day 28 by enumerating peripheralblood, lung, and bronchoalveolar lavage (BAL) fluid leuko-cytes. Assessments of blood leukocytes were determined byrecovering blood from the tail vein and removing contami-nating red blood cells by hypotonic lysis. Total white bloodcell counts were quantified with a hemacytometer, and cell
Fig. 3. Localization of tissue eosinophilswithin compartments of the lung wereunaffected by either rsPSGL-Ig or PS/2.Eosinophils were identified in the peri-vascular and peribronchial regions ofthe lung by immunohistochemistry witha rabbit polyclonal antiserum againstmurine eosinophil major basic protein.A: BSA/saline. (agonist pretreatment/experimental provocation). B: BSA/Ova.C: rsPSGL-Ig/saline. D: rsPSGL-Ig/Ova.E: PS/2/Ova. F: rsPSGL-Ig plus PS/2/Ova. Each section is representative of 5mice/group. Arrows, eosinophils. Bar,100 mm.
L815ANTI-CD49D AFFECTS AHR INDEPENDENT OF INFLAMMATION

differentials were performed on Wright-stained blood filmsby counting $300 cells. Lungs were lavaged three times with0.5 ml of Hanks’ balanced salt solution (HBSS; GIBCO BRL,Life Technologies, Gaithersburg, MD) containing 2% FCS.Individual BAL fluid returns were pooled and stored at 4°Cuntil the cells were counted. Total cell counts were deter-mined with a hemacytometer, and cell differentials wereperformed on Wright-stained cytospin slides (Cytospin 3,Shandon Scientific, Pittsburgh, PA) by counting $300 cells.
Leukocytes within the lung parenchyma were assessed bycollagenase digestion of perfused lungs and immunohisto-chemistry to determine the localization of infiltrating eosin-ophils. Isolation of lung cells was performed as previouslydescribed (10). Briefly, perfused lungs were removed anddiced into ,300-mm pieces. Four milliliters of HBSS contain-ing 175 U/ml of collagenase (Sigma), 10% FCS, 100 U/ml ofpenicillin, and 100 mg/ml of streptomycin were added to thetissue and incubated for 60 min at 37°C in an orbital shaker.The digested lungs were sheared with a 20-gauge needle andfiltered through 45- and 20-mm filters. Cells were washed,resuspended in HBSS, and counted with a hemacytometer.Differential cell counts were performed on Wright-stainedcytospin slides ($300 cells). Immunohistochemistry was per-formed with a rabbit polyclonal antibody against murinemajor basic protein (15). Murine basic protein antigen-anti-body complexes were detected in 4-mm sections of formalin-fixed, paraffin-embedded sections of mouse lungs (n 55/group) with the VECTASTAIN ABC Elite goat IgG kit(Vector Laboratories, Burlingame, CA). Endogenous peroxi-dase was blocked with 3% hydrogen peroxide in methanol for20 min at 25°C. Sections were trypsin digested (0.1% trypsinin 0.02 M Tris zHCl, pH 7.8) for 30 min at 25°C and washedin PBS (GIBCO BRL) for 15 min. Sections were blocked in1.0% normal goat serum in PBS with 1% BSA for 30 min at25°C and subsequently incubated with primary antibody(1:2,000 dilution) for 60 min at 25°C. Antibody-bound slideswere washed in PBS-1.0% BSA and incubated (30 min at25°C) with biotinylated goat anti-rabbit antibody (1:500 di-
lution). Sections were incubated first with avidin-biotin-per-oxidase complex (30 min at 25°C; Vector) and then withdiaminobenzidine (3 min at 25°C) for the development of acolored reaction product before being counterstained withmethyl green.
Mucous cell metaplasia. Mucous cell development alongthe airway epithelium was quantified in paraffin-embeddedtissue sections (4 mm) stained with periodic acid-Schiff re-agent. Parasaggital sections (n 5 5 mice/group) were ana-lyzed by bright-field microscopy with an image analysis soft-ware program (ImagePro Plus, Media Cybernetics, SilverSpring, MD) to derive an airway mucus index that wasreflective of both the amount of mucus per airway and thenumber of airways affected. The airway mucus index wascalculated by summing the ratio of the periodic acid-Schiff-positive epithelial area to the total epithelial area per sectionand dividing by the number of airways per section.
Measurements of AHR. AHR was determined by inducingairflow obstruction with a methacholine aerosol. Total pul-monary airflow in unrestrained conscious mice was esti-mated with a whole body plethysmograph (Buxco Electron-ics, Troy, NY). Pressure differences between a chambercontaining the mice and a reference chamber were used toextrapolate minute volume, tidal volume, breathing fre-quency, and enhanced paused (Penh). Penh is a dimensionlessparameter that is a function of total pulmonary airflow inmice during the respiratory cycle. This parameter closelycorrelates with airway resistance in BALB/c mice as mea-sured by traditional invasive techniques performed with ven-tilated mice (10).
Statistical analysis. Data are means 6 SE. Statisticalanalysis was performed on parametric data with t-tests, withdifferences between means considered significant when P ,0.05.
RESULTS
rsPSGL-Ig and PS/2 administration differentiallyreduce leukocyte infiltration of the lung. Ova-chal-lenged mice exhibited an increase in both airway eo-sinophils and lymphocytes recovered in the BAL fluid48 h after the last challenge (Fig. 1, A and B, respec-tively). Administration of either rsPSGL-Ig or PS/2before the Ova challenges resulted in a significantreduction in airway eosinophils (80 and 76%, respec-tively) recovered in the BAL fluid (Fig. 1A). Moreover,administration of rsPSGL-Ig and PS/2 in combinationfurther reduced eosinophil accumulation (;95% reduc-tion) in the BAL fluid compared with PS/2 alone (Fig.1B). In contrast, although treatment with eitherrsPSGL-Ig or PS/2 alone significantly reduced (;50%)the number of lymphocytes in the BAL fluid (Fig. 1B),treatment with both rsPSGL-Ig and PS/2 did not fur-ther reduce the number of lymphocytes recovered inthe BAL fluid. The administration of either antagonistalone or in combination had no effect on BAL fluidleukocyte counts in saline-challenged mice.
The effects of rsPSGL-Ig and PS/2 on Ova-inducedlung inflammation was further examined by analysesof leukocytes recovered from collagenase-digestedlungs. These experiments demonstrated that treat-ment with either rsPSGL-Ig or PS/2 alone significantlyinhibited tissue eosinophil accumulation (76 and 70%,respectively), whereas treatment with rsPSGL-Ig incombination with PS/2 further inhibited the number of
Fig. 4. Peripheral blood eosinophil numbers increase as a conse-quence of adhesion molecule blockade. Peripheral blood was col-lected, contaminating red blood cells were removed by hypotoniclysis 48 h after the last of 3 Ova challenges, and total recoveredeosinophils were enumerated. Values are means 6 SE; n 5 8–12mice/group. The administration of PS/2 alone or PS/2 and rsPSGL-Igtogether had no effect on peripheral blood eosinophil counts insaline-challenged mice (PS/2 5 63 6 9 eosinophils/mm3 and PS/2plus rsPSGL-Ig 5 70 6 6 eosinophils/mm3). *Significantly differentfrom BSA/Ova, P , 0.05.
L816 ANTI-CD49D AFFECTS AHR INDEPENDENT OF INFLAMMATION
eosinophils recruited to the lung (Fig. 2A). Similar towhat was observed with BAL fluid lymphocytes, treat-ment with either rsPSGL-Ig or PS/2 alone significantlyreduced the number of lymphocytes in the lung (Fig.2B). However, treatment with both rsPSGL-Ig and
PS/2 did not further attenuate the number of lympho-cytes recovered from the lung.
The spatial distribution of eosinophils within thelung was determined by immunohistochemistry of par-affin-embedded sections (Fig. 3). These data demon-
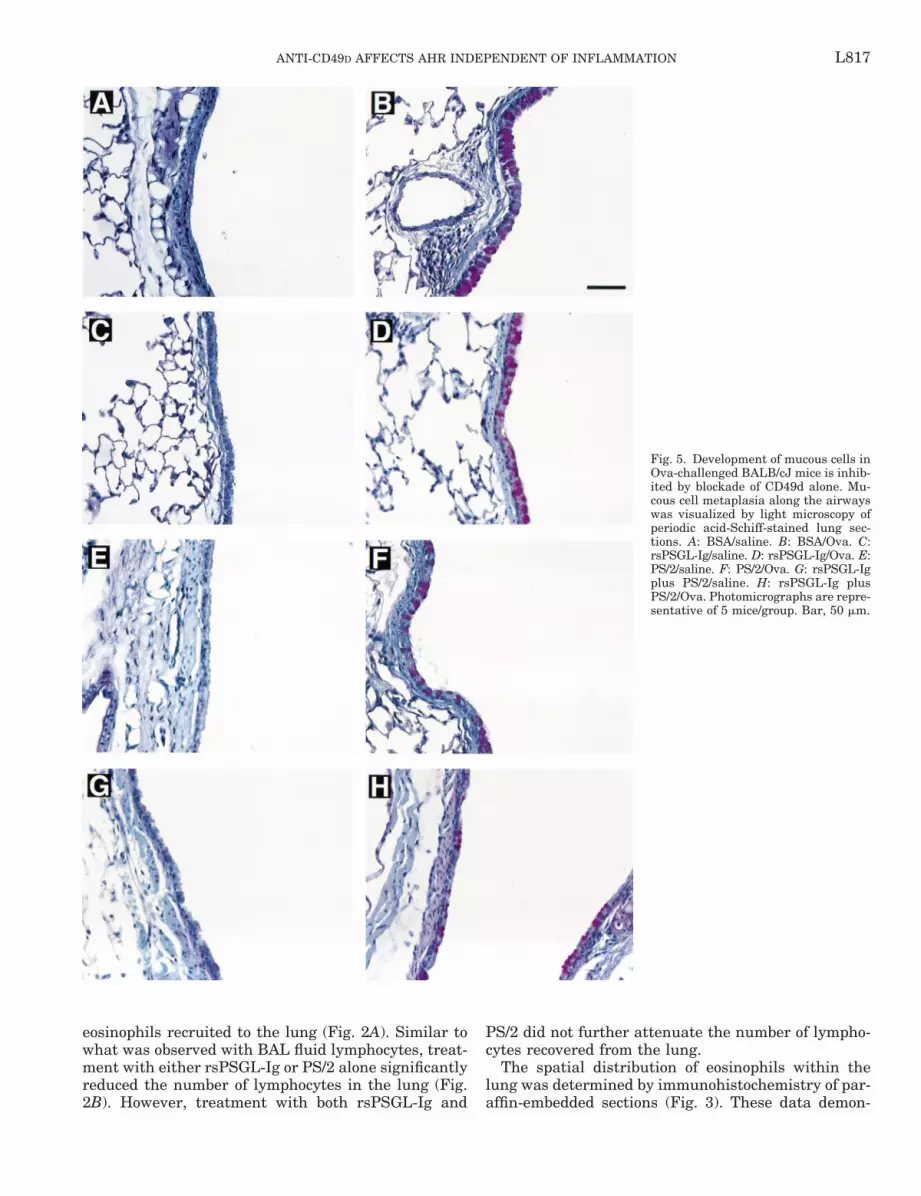
Fig. 5. Development of mucous cells inOva-challenged BALB/cJ mice is inhib-ited by blockade of CD49d alone. Mu-cous cell metaplasia along the airwayswas visualized by light microscopy ofperiodic acid-Schiff-stained lung sec-tions. A: BSA/saline. B: BSA/Ova. C:rsPSGL-Ig/saline. D: rsPSGL-Ig/Ova. E:PS/2/saline. F: PS/2/Ova. G: rsPSGL-Igplus PS/2/saline. H: rsPSGL-Ig plusPS/2/Ova. Photomicrographs are repre-sentative of 5 mice/group. Bar, 50 mm.
L817ANTI-CD49D AFFECTS AHR INDEPENDENT OF INFLAMMATION

strate that eosinophils were localized predominantly inthe peribronchial and perivascular regions of the lungafter Ova sensitization and challenge (Fig. 3, A and B).Administration of rsPSGL-Ig and PS/2, alone or incombination, inhibited the accumulation of eosinophilsin these regions of the lung without any noticeablechanges in their relative localization (i.e., eosinophilsremained predominantly in peribronchial and perivas-cular regions).
Peripheral blood eosinophilia is augmented as aconsequence of administering rsPSGL-Ig and/orPS/2. Relative to saline control mice, Ova-challengedmice exhibited a significant increase in peripheralblood eosinophils (73 6 17 vs. 262 6 25 cells/mm3) 48 hafter the last challenge (Fig. 4). Administration ofrsPSGL-Ig or PS/2 increased eosinophil accumulationin the blood of Ova-challenged mice (362 6 55 and400 6 27 cells/mm3, respectively), and treatment withboth antagonists together further increased blood eo-sinophil counts (539 6 56 cells/mm3). No effects onperipheral blood eosinophil counts were observed insaline-challenged mice treated with rsPSGL-Ig and/orPS/2.
Coadministration of rsPSGL-Ig and PS/2 had littleadditional effect on airway mucous cell developmentcompared with PS/2 alone. Mice sensitized and chal-lenged with Ova developed several histopathologies inthe lung, including increased epithelial cell thicknessand mucous cell metaplasia along the conducting air-ways (Figs. 5, A vs. B, and 6). Administration ofrsPSGL-Ig had no effect on the development of mucouscells along the airways (Figs. 5D and 6), whereas PS/2treatment alone inhibited the production of mucus bythe epithelium (Figs. 5F and 6). This effect was en-hanced by the administration of both antagonists incombination (Figs. 5H and 6).
Blockade of CD49d obviates antigen-induced AHR.Mice sensitized and challenged with Ova demonstratedan increased reactivity (i.e., AHR) to methacholineprovocation compared with saline-challenged mice.This AHR was unaffected by pretreatment withrsPSGL-Ig (Fig. 7A). In contrast, treatment with PS/2alone completely inhibited AHR in Ova-challengedmice (Fig. 7B). Furthermore, pretreatment with bothantagonists in combination also abolished AHR (Fig.7C).
DISCUSSION
The migration of leukocytes from the vasculatureinto the lung tissue in response to allergen provocationinvolves tethering, rolling, and firm adhesion to the
Fig. 7. PS/2 administration alone inhibits the development of airwayhyperreactivity in Ova-challenged BALB/cJ mice. Airway reactivity[enhanced pause (Penh)] to increasing doses of inhaled methacholinewas determined by whole body plethysmography 48 h after the lastof 3 Ova challenges. Values are means 6 SE; n 5 10–12 mice/group.
Fig. 6. Mucus production by airway epithelial cells in Ova-chal-lenged BALB/cJ mice is differentially attenuated by rsPSGL-Ig vs.PS/2 pretreatment. Values are means 6 SE of evaluations performedin duplicate as independent observer-blinded assessments; n 5 5–6mice/group. The administration of PS/2 alone or PS/2 and rsPSGL-Igtogether had no effect on mucus production in saline-challenged mice(PS/2 5 0.55 6 0.20 mucus index and PS/2 plus rsPSGL-Ig 5 0.43 60.06 mucus index).*Significantly different from BSA/Ova, P , 0.05.†Significantly different from PS/2/Ova, P , 0.05.
L818 ANTI-CD49D AFFECTS AHR INDEPENDENT OF INFLAMMATION

endothelial cell surface. These processes are controlledby cell surface receptor-ligand interactions that in-clude the binding of leukocyte-expressed PSGL-1 withendothelial cell P-selectin and the binding of leukocyteCD49d-containing integrins with VCAM-1 (27).PSGL-1 is a sialomucin consisting of two 120-kDasubunits expressed on the cell surface of most leuko-cytes, including eosinophils and T cells (18), and isthought to be the principal ligand responsible for eo-sinophil tethering on endothelial cell-expressed P-se-lectin (20, 29). The importance of P-selectin in eosino-phil and lymphocyte recruitment has been establishedwith P-selectin-deficient mice (5, 7, 9). These studiesshowed that in Ova-induced models of allergic airwayinflammation, the onset and magnitude of leukocyteaccumulation in the airways were inhibited in P-selec-tin-deficient compared with wild-type mice. This hasalso been demonstrated with P-selectin antibodies in invitro eosinophil adhesion assays (20, 30). CD49d (a4-integrin) is the common subunit of two cell surfaceheterodimers (a4b1 and a4b7) expressed on leukocytesthat mediate cell-cell adhesion to VCAM-1 and cell-matrix adhesion to fibronectin (17). The importance ofCD49d expression on eosinophils and lymphocytes andits role in Ova-induced allergic inflammation has pre-viously been established by antibody depletion studiesin several species (19, 23, 25). These studies demon-strated that inhibition of CD49d with blocking antibod-ies inhibited both eosinophil and lymphocyte accumu-lation in the lung, with a concomitant increase in thecirculating levels of these cells.
The incomplete inhibition of pulmonary inflamma-tion commonly observed by blocking either P-selectinbinding (tethering) or VCAM-1 binding (adhesion) sug-gests that there are other mechanisms for tetheringand adhesion (i.e., E-selectin, alternate integrin-adhe-sion molecule interactions) and/or overlapping func-tions between P-selectin and VCAM-1 (i.e., VCAM-1can support initial attachment of leukocytes to theendothelium, and P-selectin is sufficient to mediatesome degree of adhesion to the endothelium). Previousevidence demonstrating that initial attachment andsubsequent firm adhesion are not independently con-trolled is extensive. 1) Eosinophil adhesion to nasalpolyp endothelium is P-selectin dependent and canoccur independent of VCAM-1 (30). Conversely, eosin-ophils can tether via CD49d to purified VCAM-1 un-der flow conditions (3), and CD49d was shown to me-diate the initial attachment of eosinophils to tumornecrosis factor-a (TNF-a)-activated endothelium underflow conditions (31). 2) Hickey et al. (12) showed thatresidual eosinophil tethering was completely abolishedby CD49d-blocking antibodies in P-selectin-deficientmice. 3) Overlapping and cooperative effects betweenthese two receptors have also been demonstrated indouble-knockout P- and E-selectin-deficient mice (13)and in in vitro assays with neutralization antibodiesand IL-4-stimulated endothelium (20). Collectively,these experiments do not exclude the contribution ofintercellular adhesion molecule (ICAM)/CD18 interac-tions, also known to mediate adhesion of eosinophils to
vascular endothelium (5); however, these studies sug-gest that interfering with both P-selectin and VCAM-1effectively eliminates eosinophilia because of thegreater specificity toward eosinophils (as opposed toneutrophils) displayed by these two receptors (21). Thedata presented here (BAL fluid and collagenase diges-tion of lung tissue) support the idea that P-selectininteractions are required for leukocyte accumulation inallergic airway inflammation and also demonstratethat administration of rsPSGL-Ig is equally effective asa blockade of CD49d. Interestingly, simultaneous inhi-bition of both interactions effectively eliminated (96%reduction) BAL fluid eosinophilia, likely due to theinability of circulating cells to interact with the endo-thelium. In contrast, a further inhibition of lympho-cytes was not observed after the administration of bothantagonists, suggesting that lymphocytes are less de-pendent on PSGL-1 and CD49d interactions with thevascular endothelium in order to accumulate in thelung during allergic inflammation. In addition to directeffects on relative adhesive interactions, the differen-tial accumulation of eosinophils and lymphocytes mayalso reflect differences in systemic recirculation (i.e.,lymphocytes are able to leave mucosal tissues andreturn to the circulation, whereas eosinophils cannotrecirculate) or in the local expansion of lymphocytesthat occurs within the lung parenchyma of allergen-exposed mice (6).
Mucous cell metaplasia and AHR are two primarypathological and physiological abnormalities associ-ated with allergic pulmonary inflammation. Our re-sults clearly demonstrate that inhibition of leukocyterecruitment does not correlate with the obviation ofthese allergic pathophysiologies. Equivalent inhibitionof eosinophil and lymphocyte accumulation by blockingdistinct receptor-ligand interactions produced differenteffects on mucous cell development and AHR. Further-more, only a nominal further decrease in mucous celldevelopment was observed when eosinophilia wasabolished with both antagonists in combination. Thesedifferential effects on pulmonary pathologies by inhi-bition of a4-receptor-ligand interactions alone suggestthat these interactions mediate a generalized effect onthe leukocyte activation state or perhaps mediate spe-cific effects on unique leukocyte subpopulations in thelungs. The role of P-selectin/PSGL-1 appears to belimited to mediating extravasation from the circulationbecause leukocytes entering the tissue under condi-tions in which PSGL-1 binding was limited maintainedthe capacity to become activated, as evidenced by thedevelopment of mucous cell metaplasia and AHR. Con-versely, the a4-integrin system appears to have dualroles in allergic airway disease. In addition to mediat-ing leukocyte recruitment from the circulation, thesedata demonstrate that a4-integrin activation is neces-sary for the development of mucous cell metaplasia andAHR. The mechanism by which and the extent towhich CD49d binding mediates activation and/or sur-vival of either eosinophils or lymphocytes remain prob-lematic. However, CD49d binding of extravascular leu-kocytes to other cells and/or extracellular matrix
L819ANTI-CD49D AFFECTS AHR INDEPENDENT OF INFLAMMATION

components may be required for the functional releaseof mediators that subsequently lead to AHR. This issupported by data that demonstrate that eosinophilsare activated, as assessed by increased survival invitro, after CD49d binding to fibronectin (4) and bystudies (2, 22) suggesting that eosinophil activation asdetermined by granule protein release rather than thenumber of cells recruited is associated with the devel-opment of bronchial hyperreactivity in sheep andguinea pigs. Furthermore, small-molecule peptide in-hibitors of CD49d, which selectively block the fibronec-tin binding domain without interfering with VCAMbinding, effectively inhibited the early- and late-phaseairway responses in allergic sheep (1, 16). In addition,Henderson et al. (11) have shown inhibition of Th2cytokines (IL-4 and IL-5) in the BAL fluid of allergicmice after intranasal administration of CD49d anti-body, suggesting that T-cell activation and associatedfunctions depend on CD49d interactions.
The different effects on eosinophil and lymphocyteinhibition observed in our study may also be a conse-quence of perturbing unique subsets of the heteroge-neous lymphocyte populations in the lung. Althoughthe total lymphocyte accumulation is equally reducedafter rsPSGL-Ig or PS/2 treatment, one or more sub-types that contribute to the observed pulmonary pa-thologies may have been preferentially inhibited byPS/2. For example, Nakajima and colleagues (19) dem-onstrated an equal reduction (;75%) in CD41 andCD81 T cells after PS/2 administration in Ova-chal-lenged mice, whereas De Sanctis et al. (7) reported a40% reduction in CD41 subsets without a change inCD81 T cells in Ova-challenged P-selectin-deficientmice. Whether these different responses can be attrib-uted to the selective inhibition of unique T-cell sub-populations remains to be determined.
These data demonstrate that administration ofrsPSGL-Ig is as effective as CD49d blockade in reduc-ing eosinophil and lymphocyte accumulation in thelungs of Ova-challenged mice, whereas, in combina-tion, the antagonists completely block eosinophil accu-mulation without additional effects on the accumula-tion of lymphocytes. This suggests that leukocytesubtypes capable of utilizing both adhesive interac-tions nonetheless migrate to the lung via independentmechanisms. Moreover, blockade of these cell adhesioninteractions also has different effects on the inhibitionof airway mucous cell development and AHR, demon-strating that inhibition of these pathologies occursindependent of leukocyte recruitment. These findingsindicate that the extent of activation is critical to thedevelopment of mucous cell metaplasia and AHR andmay represent alternative therapeutic targets of pul-monary pathologies associated with chronic allergicinflammation.
We acknowledge the technical assistance of the Mayo ClinicScottsdale Histology Facility (Anita Jennings, Director) and theMayo Clinic Scottsdale Graphic Arts Department (Marv Ruona). Wealso thank Edith Hines and Katie O’Neill for excellent technicalassistance and Linda Mardel, research program assistant.
This study was supported by funds from the Mayo Clinic Foun-dation; National Heart, Lung, and Blood Institute (NHLBI) GrantHL-60793-01S (to N. A. Lee), NHLBI Training Grant HL-07897 (toM. T. Borchers); and an individual National Research Service Award(to J. Crosby).
REFERENCES
1. Abraham WM, Ahmed A, Sielczak MW, Narita M, Arrhe-nius T, and Elices MJ. Blockade of late-phase airway re-sponses and airway hyperresponsiveness in allergic sheep with asmall-molecule peptide inhibitor of VLA-4. Am J Respir CritCare Med 156: 696–703, 1997.
2. Abraham WM, Sielczak MW, Ahmed A, Cortes A, LauredoIT, Kim J, Pepinsky B, Benjamin CD, Leone DR, Lobb RR,and Weller PF. Alpha 4-integrins mediate antigen-induced latebronchial responses and prolonged airway hyperresponsivenessin sheep. J Clin Invest 93: 776–787, 1994.
3. Alon R, Kassner PD, Carr MW, Finger EB, Hemler ME, andSpringer TA. The integrin VLA-4 supports tethering and roll-ing in flow on VCAM-1. J Cell Biol 128: 1243–1253, 1995.
4. Anwar AR, Moqbel R, Walsh GM, Kay AB, and WardlawAJ. Adhesion to fibronectin prolongs eosinophil survival. J ExpMed 177: 839–843, 1993.
5. Broide DH, Sullivan S, Gifford T, and Sriramarao P. Inhi-bition of pulmonary eosinophilia in P-selectin- and ICAM-1-deficient mice. Am J Respir Cell Mol Biol 18: 218–225, 1998.
6. Chvatchko Y, Kosco-Vilbois MH, Herren S, Lefort J, andBonnefoy JY. Germinal center formation and local immuno-globulin E (IgE) production in the lung after an airway antigenicchallenge. J Exp Med 184: 2353–2360, 1996.
7. De Sanctis GT, Wolyniec WW, Green FH, Qin S, Jiao A,Finn PW, Noonan T, Joetham AA, Gelfand E, DoerschukCM, and Drazen JM. Reduction of allergic airway responses inP-selectin-deficient mice. J Appl Physiol 83: 681–687, 1997.
8. Gleich GJ. The eosinophil and bronchial asthma: current un-derstanding. J Allergy Clin Immunol 85: 422–436, 1990.
9. Gonzalo JA, Lloyd CM, Kremer L, Finger E, Martinez AC,Siegelman MH, Cybulsky M, and Gutierrez-Ramos JC.Eosinophil recruitment to the lung in a murine model of allergicinflammation. The role of T cells, chemokines, and adhesionreceptors. J Clin Invest 98: 2332–2345, 1996.
10. Hamelmann E, Schwarze J, Takeda K, Oshiba A, LarsenGL, Irvin CG, and Gelfand EW. Noninvasive measurement ofairway responsiveness in allergic mice using barometric ple-thysmography. Am J Respir Crit Care Med 156: 766–775, 1997.
11. Henderson WR Jr, Chi EY, Albert RK, Chu SJ, Lamm WJE,Rochon Y, Jonas M, Christie PE, and Harlan JM. Blockadeof CD49D (alpha4 integrin) on intrapulmonary but not circulat-ing leukocytes inhibits airway inflammation and hyperrespon-siveness in a mouse model of asthma. J Clin Invest 100: 3083–3092, 1997.
12. Hickey MJ, Granger DN, and Kubes P. Molecular mecha-nisms underlying IL-4-induced leukocyte recruitment in vivo: acritical role for the alpha 4 integrin. J Immunol 163: 3441–3448,1999.
13. Kanwar S, Bullard DC, Hickey MJ, Smith CW, BeaudetAL, Wolitzky BA, and Kubes P. The association betweenalpha4-integrin, P-selectin, and E-selectin in an allergic model ofinflammation. J Exp Med 185: 1077–1087, 1997.
14. Kitayama J, Fuhlbrigge RC, Puri KD, and Springer TA.P-selectin, L-selectin, and alpha 4 integrin have distinct roles ineosinophil tethering and arrest on vascular endothelial cellsunder physiological flow conditions. J Immunol 159: 3929–3939,1997.
15. Lee JJ, McGarry MP, Farmer SC, Denzler KL, Larson KA,Carrigan T, Brenneise IE, Horton MA, Haczku A, GelfandEW, Leikauf GD, and Lee NA. Interleukin-5 expression in thelung epithelium of transgenic mice leads to pulmonary changespathognomonic of asthma. J Exp Med 185: 2143–2156, 1997.
16. Lin K, Ateeq HS, Hsiung SH, Chong LT, Zimmerman CN,Castro A, Lee WC, Hammond CE, Kalkunte S, Chen LL,Pepinsky RB, Leone DR, Sprague AG, Abraham WM, GillA, Lobb RR, and Adams SP. Selective, tight-binding inhibitors
L820 ANTI-CD49D AFFECTS AHR INDEPENDENT OF INFLAMMATION

of integrin a4b1 that inhibit allergic airway responses. J MedChem 42: 920–934, 1999.
17. Lobb RR, Pepinsky B, Leone DR, and Abraham WM. Therole of alpha 4 integrins in lung pathophysiology. Eur Respir JSuppl 22: 104s–108s, 1996.
18. McEver RP and Cummings RD. Perspectives series: cell ad-hesion in vascular biology. Role of PSGL-1 binding to selectins inleukocyte recruitment. J Clin Invest 100: 485–491, 1997.
19. Nakajima H, Sano H, Nishimura T, Yoshida S, andIwamoto I. Role of vascular cell adhesion molecule 1/very lateactivation antigen 4 and intercellular adhesion molecule 1/lym-phocyte function-associated antigen 1 interactions in antigen-induced eosinophil and T cell recruitment into the tissue. J ExpMed 179: 1145–1154, 1994.
20. Patel KD. Eosinophil tethering to interleukin-4-activated endo-thelial cells requires both P-selectin and vascular cell adhesionmolecule-1. Blood 92: 3904–3911, 1998.
21. Patel KD and McEver RP. Comparison of tethering and roll-ing of eosinophils and neutrophils through selectins and P-selectin glycoprotein ligand-1. J Immunol 159: 4555–4565, 1997.
22. Pretolani M, Ruffie C, Joseph D, Campos MG, Church MK,Lefort J, and Vargaftig BB. Role of eosinophil activation inthe bronchial reactivity of allergic guinea pigs. Am J Respir CritCare Med 149: 1167–1174, 1994.
23. Pretolani M, Ruffie C, Lapa e Silva JR, Joseph D, LobbRR, and Vargaftig BB. Antibody to very late activation antigen4 prevents antigen-induced bronchial hyperreactivity and cellu-lar infiltration in the guinea pig airways. J Exp Med 180:795–805, 1994.
24. Renz H, Smith HR, Henson JE, Ray BS, Irvin CG, and Gel-fand EW. Aerosolized antigen exposure without adjuvant causesincreased IgE production and increased airway responsiveness inthe mouse. J Allergy Clin Immunol 89: 1127–1138, 1992.
25. Richards IM, Kolbasa KP, Hatfield CA, Winterrowd GE,Vonderfecht SL, Fidler SF, Griffin RL, Brashler JR, Krz-esicki RF, Sly LM, Ready KA, Staite ND, and Chin JE. Roleof very late activation antigen-4 in the antigen-induced accumu-
lation of eosinophils and lymphocytes in the lungs and airwaylumen of sensitized brown Norway rats. Am J Respir Cell MolBiol 15: 172–183, 1996.
26. Sako D, Chang XJ, Barone KM, Vachino G, White HM,Shaw G, Veldman GM, Bean KM, Ahern TJ, Furie B, Cum-ming DA, and Larson GR. Expression cloning of a functionalglycoprotein ligand for P-selectin. Cell 75: 1179–1186, 1993.
27. Springer TA. Traffic signals for lymphocyte recirculation andleukocyte emigration: the multistep paradigm. Cell 76: 301–314,1994.
28. Strauss EC, Larson KA, Brenneise I, Foster CS, LarsenGR, Lee NA, and Lee JJ. Soluble P-selectin glycoprotein ligand1 inhibits ocular inflammation in a murine model of allergy.Invest Ophthalmol Vis Sci 40: 1336–1342, 1999.
29. Symon FA, Lawrence MB, Williamson ML, Walsh GM,Watson SR, and Wardlaw AJ. Functional and structural char-acterization of the eosinophil P-selectin ligand. J Immunol 157:1711–1719, 1996.
30. Symon FA, Walsh GM, Watson SR, and Wardlaw AJ. Eo-sinophil adhesion to nasal polyp endothelium is P-selectin-de-pendent. J Exp Med 180: 371–376, 1994.
31. Ulfman LH, Kuijper PH, van der Linden JA, LammersJW, Zwaginga JJ, and Koenderman L. Characterization ofeosinophil adhesion to TNF-alpha-activated endothelium un-der flow conditions: alpha 4 integrins mediate initial attach-ment, and E-selectin mediates rolling. J Immunol 163: 343–350, 1999.
32. Wardlaw AJ, Walsh GM, and Symon FA. Adhesion interac-tions involved in eosinophil migration through vascular endothe-lium. Ann NY Acad Sci 796: 124–137, 1996.
33. Weller PF, Rand TH, Goelz SE, Chi-Rosso G, and Lobb RR.Human eosinophil adherence to vascular endothelium mediatedby binding to vascular cell adhesion molecule 1 and endothelialleukocyte adhesion molecule 1. Proc Natl Acad Sci USA 88:7430–7433, 1991.
34. Wills-Karp M. Immunologic basis of antigen-induced airwayhyperresponsiveness. Annu Rev Immunol 17: 255–281, 1999.
L821ANTI-CD49D AFFECTS AHR INDEPENDENT OF INFLAMMATION





























