Embed Size (px)
Citation preview

Bridging the gap: Transitive associations between items presented insimilar temporal contexts
Marc W. Howard, Bing Jing, Vinayak A. Rao, Jennifer P. Provyn, Aditya V. DateyDepartment of Psychology
Syracuse University
Re-submitted to JEP:LMC February 4, 2007, do not quote without permissionAssociations in episodic memory are formed between items presented close together in time.
The temporal context model (TCM) hypothesizes that this contiguity effect is a consequenceof shared temporal contexts rather than temporal proximity per se. Using double function listsof paired associates, which include chains of pairs (e.g. A-B, B-C), we examined associationsbetween items that were not presented close together in time but were presented in similartemporal contexts. For instance A and C do not appear together, but both occur in the contextof B. Although within-pair associations (e.g. A-B) were asymmetric, across-pair associations(e.g. A-C) showed no evidence for asymmetry. We attempted to describe these transitive as-sociations using two models. One was a heteroassociative model in which the A-C associa-tions resulted from mediated chaining as a result of “stepping through” the links in the chain.Although this heteroassociative model and TCM make identical predictions regarding simplecontiguity effects, the heteroassociative model had great difficulty accounting for the form oftransitive associations between items. TCM provided an excellent fit to the data. These dataraise the surprising possiblity that episodic contiguity effects do not reflect direct associationsbetween items but rather a process of binding, encoding and retrieval of a gradually-changingrepresentation of temporal context.
Episodic memory refers to the ability to remember eventsthat occur at a specific place and time (Tulving, 2002, 1983).Episodic memory consists of vivid recollection of stimulifrom various modalities that were experienced in a similartime and place. For instance, episodic memory for break-fast may bring to mind the crunch of a bagel, the smell of acup of tea and the sound and semantic content of a conver-sation. One can ask how strict the requirement of temporalco-occurrence is for the formation of an episodic memory.Recent work examining episodic memory performance in thelaboratory suggests that strict temporal co-occurrence is notrequired for items to become associated in episodic memory.The purpose of this paper is to ask how this is accomplishedand what this might tell us about the basis of episodic mem-ory.
One way to investigate episodic memory in the laboratoryis to use the free recall task. In free recall, subjects are pre-sented with a list of words one at a time. Their task is torecall as many words as they can from the list in the orderthat these words come to mind. Free recall is an episodicmemory task—the subject’s task is to recall the items that
Supported by MH069938-01 from the NIH. The authors ac-knowledge the generous support of Michael Hasselmo and Chan-tal Stern in developing pilot versions of this task when the seniorauthor was at Boston University. Thanks to Diana Hobbins, RadhaModi, and Madhura Phadke for helping collect and score data.
were experienced in a particular time and place. The orderin which subjects recall the items from the list reflects theorganization of the items that are part of that episode. Exam-ination of transitions in free recall demonstrates that itemspresented close together in time, but that are not presentedsimultaneously, become associated to each other. The condi-tional response probability as a function of lag (lag-CRP) de-veloped by Kahana and colleagues (e.g. Howard & Kahana,1999; Kahana, 1996; Kahana, Howard, Zaromb, & Wing-field, 2002; Klein, Addis, & Kahana, 2005) describes theprobabilty of making a recall transition from a just-recalledwords to other words in the list as a function of their distancein the list. For instance if the 10th word in a list of 20 hasjust been recalled in a delayed free recall experiment, then,all other things being equal, the next word the subject recallsis more likely to come from a nearby serial position (e.g.serial position 9 or 11) than a remote serial position (e.g. 5or 15). The lag-CRP falls off gradually with distance, orlag, between the items in the list. That is, the function doesnot fall abruptly, but decays gradually over several list posi-tions. In fact, associations between items presented in serieshave been observed when a distractor is presented betweeneach item in the list (Howard & Kahana, 1999) and even be-tween items presented in nearby lists (Howard, Youker, &Venkatadass, accepted pending minor revisions). In additionto this contiguity effect, lag-CRP functions in free recall alsoshow an asymmetry such that transitions forward in the list(e.g. a transition from item ten to item eleven) are more likelythan transitions backward in the list (e.g. a transition from

2 HOWARD ET AL.
item ten to item nine).The contiguity effect is not limited to the free recall
paradigm. Graded contiguity effects with both forward andbackward associations have also been observed in intrusionrates in serial recall (Kahana & Caplan, 2002; Klein et al.,2005; Raskin & Cook, 1937), intrusion rates in paired asso-ciates learning (Davis, Rizzuto, Geller, & Kahana, submit-ted) and hit rates in item recognition of pictures (Schwartz,Howard, Jing, & Kahana, 2005).1 Temporal contiguity ef-fects are observed in all of the major episodic memoryparadigms. They are observed whether memory is testedusing recall or recognition, whether the to-be-rememberedstimuli are pictures or words, and whether the associationshelp performance (free recall), are neutral with respect to per-formance (item recognition) or actively disrupt performance(intrusions in serial recall, across-pair associates in cued re-call). The contiguity effect is apparently a very general prop-erty of episodic memory (see Kahana, Howard, & Polyn, inpress, for a more thorough review of contiguity effects inepisodic memory).
Theoretical accounts of temporal contiguity effectin episodic memory tasks
Given the ubiquity of temporal contiguity effects inepisodic memory, it is clearly of great theoretical interest toidentify the mechanism (or perhaps mechanisms) that givesrise to them. Much of the work done on episodic memorytheory has been to describe and define the properties thatare identified as episodic memory (see Tulving, 2002, for areview). The progress that has been made on these fronts,however, does not provide a satisfactory mechanistic accountof how episodic recall—and in particular the contiguity ef-fect in episodic memory tasks—is accomplished. On theother hand, the mathematical modeling literature provides awealth of potential mechanisms to explain contiguity effectsin episodic memory tasks. These quantitative explanationscan be classified into two broad classes which we will referto here as heteroassociative and retrieved context accounts.
In heteroassociative models, representations of items pre-sented together are directly associated to one another. Theseitem-to-item associations are a common theme in distributedmemory models (e.g., Chappell & Humphreys, 1994; Mur-dock, 1982; Humphreys, Bain, & Pike, 1989). The idea be-hind these models is that to-be-remembered items evoke adistributed representation of activity in the brain. The pat-terns corresponding to the to-be-remembered items are thenassociated by means of some type of plasticity. Heteroas-sociative distributed memory models differ in the nature ofthe representation used to describe the items, the mechanismthat supports the association and the assumptions as to whichitems are associated but share the basic assumption that con-tiguity effects follow from direct item-to-item associations.For instance, Metcalfe and Murdock (1981) hypothesizedthat in free recall adjacent items are associated to each otherby means of convolution. A similar approach was used intreating serial recall by Lewandowsky and Murdock (1989).2
An important question in formulating a heteroassociative
model is how the item representations of items presentedat different times are simultaneously available so that theymight be bound together. Classic notions of a limited capac-ity short-term store serve just this function (Jensen & Lis-man, 2005; Raaijmakers & Shiffrin, 1980). The quantitativeimplementation of the buffer model that has been applied tothe most broad range of experimental data is the search of as-sociative memory (SAM) model (Gillund & Shiffrin, 1984;Kahana, 1996; Raaijmakers & Shiffrin, 1980). In SAM thestrength of the association between items in long-term mem-ory is incremented when they are co-active in short-termmemory. SAM uses this property to accurately describe con-tiguity effects in free recall when items are presented suc-cessively (Howard & Kahana, 1999; Kahana, 1996; Sirotinet al., 2005). In SAM, environmental context, perhaps gradu-ally changing context (Mensink & Raaijmakers, 1988), playsan important role in focusing the memory search on the ap-propriate temporal interval, for instance the most recentlystudied list (Sirotin et al., 2005), but does not account forassociations formed between items within a list unless ad-ditional assumptions are made (Howard & Kahana, 1999).Gradually-changing contextual states have also been usedin models of serial recall (Brown, Preece, & Hulme, 2000;Burgess & Hitch, 1999) to generate serial position and tem-poral grouping effects (see Burgess & Hitch, 2005, for a re-view).
It is possible to build on this idea of gradually-changingcontext to construct an alternative model of contiguity ef-fects. Dennis and Humphreys (2001) elaborated variablecontext models by postulating that items can recover previ-ous contextual states in the service of item recognition. Thetemporal context model (TCM, Howard & Kahana, 2002;Howard, Fotedar, Datey, & Hasselmo, 2005) proposes thata variable context signal is recovered as a means to generatecontiguity effects in free recall (Howard, Kahana, & Wing-field, 2006). Although TCM implements context using a for-malism similar to previous mathematical models of memory(Murdock, 1997; Murdock, Smith, & Bai, 2001), it offers aradical departure from previous models in its explanation ofthe origin of temporal contiguity effects.3 TCM does not pos-
1 While asymmetry was robustly observed in the serial recallstudies cited, neither the paired associates study nor the item recog-nition study showed statistically reliable asymmetry effects.
2 Although heteroassociative models of serial recall(Lewandowsky & Murdock, 1989) have been a subject ofsome controversy (Henson, Norris, Page, & Baddeley, 1996 but seeHulme, Stuart, Brown, & Morin, 2003), heteroassociation remainsinfluential for models of cued recall and paired associate learningas well as free recall (e.g. Kahana, Rizzuto, & Schneider, 2005;Sirotin, Kimball, & Kahana, 2005).
3 Although the equations appear at first blush to be more distant,conceptually, TCM is actually much closer to stimulus samplingtheory (Estes, 1955a) than more recent models of variable context.In stimulus sampling theory, the population of fluctuating elementsthat subsequent authors have identified as environmental context iscaused by the presentation of the stimulus itself (Estes, 1950). Instimulus sampling theory associations between a stimulus and re-sponse can be seen as the consequence of elements recovered by

TRANSITIVE ASSOCIATIONS 3
tulate the formation of direct heteroassiative connections atall during learning of episodic associations. Rather, temporalcontiguity effects are a consequence of items’ ability to bebound to and recover a gradually changing representation oftemporal context, which serves as the cue for episodic recall.Put another way, rather than relying on item-to-item associ-ations, as heteroassociative models do, TCM produces tem-poral contiguity effects as a consequence of item-to-context-to-item associations. For instance, suppose a pair of wordsABSENCE—HOLLOW is presented. During study, the encod-ing context for each pair is composed of elements retrievedby each member of the pair. During study, the item represen-tation for HOLLOW is associated with the encoding contextof the pair. When ABSENCE is repeated as a cue, part of theencoding context of the pair is recovered. Because the con-text retrieved by ABSENCE at test overlaps with the encodingcontext of HOLLOW, the result is a behaviorally observed as-sociation between ABSENCE and HOLLOW.
Heteroassociative and contextual recovery accounts ofepisodic association cannot be distinguished by their abilityto describe the form of temporal contiguity effects in stan-dard free recall tasks. That is, given the universality of conti-guity effects, a heteroassociative model could easily explaina broad class of contiguity effects if the heteroassociativemechanism is chosen to have an appropriate form. In fact,computational modeling has demonstrated that SAM can sat-isfactorily describe lag-CRP functions in free recall as longas the items in the list are not separated by a delay (Howard& Kahana, 1999; Kahana, 1996; Sirotin et al., 2005). TCMcan also provide a satisfactory account of temporally-definedassociations in free recall, including the hallmark asymme-try (Howard & Kahana, 2002; Howard et al., 2006). Al-though SAM cannot account for the temporally-defined as-sociations observed in continuous-distractor free recall inwhich a lengthy interitem distractor intervenes between eachitem (Howard & Kahana, 1999) while TCM can account forthis finding (Howard & Kahana, 2002; Sederberg, Howard,& Kahana, in preparation), this discrepancy is not a conse-quence of the basic assumptions about heteroassociation ver-sus contextual recovery. Instead, a version of SAM in whichitems did not drop out of the short-term buffer in an all-or-none fashion but rather decayed gradually would almostcertainly be capable of describing at least the existence ofthe temporally-defined associations observed in continuous-distractor free recall. In order to differentiate the predictionsof these accounts of contiguity effects in episodic memory,we need to go beyond simple temporal proximity.
Dissociating temporal context from temporal con-tiguity
According to TCM items presented close together in timebecome associated to each other not because of their tempo-ral proximity per se, but because of the similarity betweenthe encoding context of one item and the context recoveredby the other. As long as there is similarity between thesestates of context, items should become associated to eachother regardless of whether they were actually in close tem-
−1
−2POLICE
ABSENCE
FARMER
MONEY
DARLING
WINDOW
FOUNTAIN
SHIPPING
ABSENCE
DARLING
HOLLOW
HOLLOW
HOLLOW
PUPIL
PUPIL
PUPIL RIVERRIVER
RIVER
+1
+2
Figure 1. Schematic of presentation of double-function pairsand the corresponding linked-list. Colors are shown for explana-tory purposes and were not shown to the participants. Words werepresented one at a time on the screen. Participants were able todistinguish members of the same pair by means of a longer delaybetween pairs than within-pair. On the left are a sample of pairs thatthe subject might be shown in order. On the right is the linked listthat would be formed by stringing together pairs with overlapping.The numbers on the right indicate the linked-list lag associated withseveral recall transitions. For instance, given PUPIL as a cue, a tran-sition to the correct response, RIVER would have a linked-list lag of+1. Given PUPIL as a cue, a backward intrusion, HOLLOW in thisexample, would have a linked-list lag of −1. Remote intrusions, forinstance ABSENCE or DARLING would be associated with linked-list lags with absolute value greater than one.
poral proximity. What is needed is an experimental manip-ulation that allows for items that are presented in similartemporal contexts but not close together in time. Double-function lists of paired associates, in which pairs share over-lapping items (Primoff, 1938, see also Figure 1), provide asetting in which items do not co-occur but nonetheless appearin similar temporal contexts. A double-function list mightinclude the pairs ABSENCE–HOLLOW and HOLLOW–PUPIL.In learning these paris, ABSENCE and PUPIL are never pre-sented in the same pair, but are both presented in the con-text of HOLLOW. The contextual retrieval hypothesis pre-dicts that ABSENCE and PUPIL should become associated byvirtue of being encoded in similar contexts, even though theydo not co-occur.
Associations that bridge across double-function pairs havein fact been observed (Popper, 1959; Slamecka, 1976; Bun-sey & Eichenbaum, 1996). For example Slamecka (1976)presented subjects with double-function lists of paired as-sociates. After learning, Slamecka (1976) gave subjects arecognition test in which a cue item from one of the pairswas presented along with three alternatives. Subjects wereinstructed to choose the word that was the response to the cueitem during study. Of the two foils, one was always an un-related item from a different double-function list that did notoverlap with the double-function list that contained the cueitem. In an experimental condition, the other foil was an itemthat was a bridging associate of the cue item. In the controlcondition, both foils were from other double-function lists.Performance on the experimental condition was worse thanon the control condition. This suggests associative strengthbetween the cue item and the bridging associate was greaterthan that between the cue item and an unrelated item from a
the stimulus being conditioned to a response. The set of elementsrecovered by the stimulus gradually change over repeated presenta-tions of the item, analogous to Eqs. 3 and 4

4 HOWARD ET AL.
Study Heteroassociation Contextual retrieval
Item
Context
A B C
a b c
A B CItem
Con
text
Test
C
Context
A BItem
Con
text
a b
A B CItem
Figure 2. Two accounts of bridging associations across pairs.The top row illustrates connections made at study; the bottom rowillustrates retrieval events at test. The left column describes the as-sumptions of a heteroassociative model. The right column describesthe assumptions of a contextual retrieval model, in particular TCM.(top left) At study, the heteroassociative account describes encod-ing of the pairs A−B and B−C as direct item-to-item associationsbetween each of the to-be-remembered items. (top right) At study,the contextual account describes encoding as binding of an itemrepresentation to a representation of temporal context. These repre-sentations of temporal context depend on the items being presented,so that the context for the A−B pair, denoted ab, overlaps to someextent with the context for the B−C pair, denoted bc. (lower left)At retrieval, associations between A and C are possible in the het-eroassociative account by means of activation spreading from A toB to C. (lower right) The contextual retrieval account enables as-sociations between A and C that do not depend on the intermediateactivation of B. The probe item A recovers the temporal contextfrom the encoding of A−B. Because this context overlaps with theencoding context for B−C, C is indirectly activated by the contextretrieved by A.
non-overlapping list.The mere existence of associations that bridge across pairs
in a double function list is not sufficient to rule out a het-eroassociative account of temporal contiguity effects. As-sociations among items that were never presented togethercan be explained by heteroassociative accounts using medi-ated chaining, the explanation that Slamecka (1976) favored.That is, one can explain a behaviorally-observed associationbetween ABSENCE and PUPIL if the item representation forHOLLOW is activated as an intermediate step (see Figure 2).This can be manifest as a retrieval effect or can give rise toan encoding effect.
Mediated chaining could result in a bridging associationat retrieval if the subject surreptitiously retrieves a link inthe chain but, for whatever reason, withholds the responsewaiting until the next link in the chain is traversed. Thiswould not be a particularly useful strategy in paired asso-ciate learning, but we will later examine final free recallof studied pairs, in which participants recall all the studyitems they can remember in any order after learning a set ofdouble-function items. In this case, participants may chooseto buffer their output and postpone speaking a response untilmultiple items are available. This retrieval chaining accountpredicts a strong dependence between bridging associations
and the links in the chain that must be traversed to bridgebetween the items. Data from a lesion study with rats byBunsey and Eichenbaum (1996) argue against the chainingretrieval explanation. Bunsey and Eichenbaum (1996) usedan analogue of paired associate learning using odors as stim-uli to teach animals chained pairs A−B and B−C.4 Intactanimals showed a generalization to bridging A−C associ-ations during probe trials. Although animals with lesionsto the hippocampus were able to learn the pairs—the linksin the chain—as well as intact animals, they did not showevidence for retrieval of bridging associations. This is notwhat one would expect given a mediated chaining retrievalexplanation, but there are a number of discrepancies betweenthe task the rats learned and standard episodic memory tasks(e.g. number of trials, differential availability of responses intraining vs probe trials, use of language) to leave the medi-ated chaining retrieval explanation a viable account of bridg-ing associations in episodic memory tasks.
In this paper, we present the results of a study in whichpariticpants learn a long list of double-function pairs. Wecompare the associative strength between the items that com-pose the pairs by examining transition probabilities from afinal free recall session after the participants have learned thepairs. We will use these data to constrain models of bridgingassociations. We will argue that the results of the experimentrule out retrieval variants of the mediated chaining account.We will explicitly simulate a more complex heteroassocia-tive model and compare it to the results from a simulation ofTCM. We briefly sketch the predictions of these models sothat the reader will be able to have a sense of the features ofthe data that constrain the models.
We will be able to rule out the mediated chaining surrep-titious retrieval explanation by noting a discrepancy betweenthe asymmetry between adjacent, A−B associations and thesymmetry of remote A−C associations in recall of doublefunction pairs. Heteroassociative models may either predictassociative symmetry between members of a pair, such thatafter learning a pair A−B, the backward association B−Ais just as strong as A−B. Associative symmetry is a predic-tion of models based on convolution (e.g. Murdock, 1982)and has been observed in a number of standard cued recallstudies (see Kahana, 2002, for a review). Other heteroasso-ciative models, particularly those not developed for describ-ing paired associate learning can predict an asymmetry be-tween items (Gillund & Shiffrin, 1984; Sirotin et al., 2005).The observation of asymmetry in the double function pairsin the present study rules out simple symmetric heteroasso-ciative models, but does not rule out asymmetric heteroasso-ciative models. However, if an asymmetric heteroassociativemodel is used to drive a mediated chaining account of bridg-ing associations that depends on surreptitious retrieval, this
4 The procedure was actually a conditional discrimination. In afirst stage of learning, rats recieved a cue item (either A or X) andthen had to select which of a pair of choice odors, B and Y wouldcontain reward. The identity of the cue odor predicted which of thechoice odors was correct. For instance, if the cue odor was A, thenthe rat would learn to approach B rather than Y .

TRANSITIVE ASSOCIATIONS 5
would predict that the asymmetry should persist, and in factbe amplified in stepping to more remote links in the chain.That is, if A−B associations are stronger than B−A asso-ciations, then a heteroassociative retrieval account of bridg-ing associations predicts that the asymmetry should be largerbetween A−C and C−A associations, since such A−C as-sociations depend on stepping through two asymmetric linksin the chain. We will report that although there was a strongasymmetry between adjacent pairs (e.g. A−B), there was noevidence for asymmetry at more remote pairs (e.g. A−C).A direct comparison of the degree of asymmetry will showthat the adjacent asymmetry is larger than the remote asym-metry (which doesn’t differ from zero), thus falsifying thesurreptitious retrieval mediated chaining account.
One can also imagine a mediated chaining account basedon encoding surreptitious retrievals (or perhaps intrusions).Suppose that during cued recall testing, or perhaps learningthe pairs on multiple trials, participants make multiple sur-reptitious responses to a probe. Given A as a cue, subjectsmight recall B and then use B as a cue to surreptitiously thinkof C. If participants are able to encode during these retrievalepisodes, then associations can be formed between A and Cbecause they are experienced in temporal proximity as a con-sequence of surreptitious retrieval. Because the predictionsof this model regarding symmetry and asymmetry are lessobvious (see below), we will evaluate this encoding variantof the mediated chaining hypothesis by explicitly simulatingit.
Unlike the surreptitious retrieval mediated chaining ac-count, TCM does not predict a strong dependence betweenbridging associations and the links in the chain connectingthe two items. According to TCM, there are two sourcesof contiguity effects that arise from the two components ofcontext retrieved by an item. One component, which is con-sistent across repeated presentations of the item, is sufficientfor simple forward associations. The other component, re-ferred to as newly-learned context, allows a repeated item torecover the states of context that were presented when it wasinitially presented. This component, which has been hypoth-esized to depend on the hippocampus proper (Howard et al.,2005), is solely responsible for backward associations be-tween items presented in series. In TCM, these two compo-nents both contribute to recall under normal circumstances,providing a principled account of the asymmetry observedin temporal contiguity effects under a wide variety of cir-cumstances (Howard & Kahana, 2002; Howard et al., 2006).According to TCM, bridging associations should depend ex-clusively on the newly-learned context component, whereasforward associations within pair should depend on both thenewly learned and consistent components. This propertyenables TCM to explain the dissociation observed betweenpairwise and bridging associations with hippocampal dam-age in a rodent analogue of the paired associates task (Bun-sey & Eichenbaum, 1996; Howard et al., 2005). This samedistinction will also allow TCM to model a strong asymme-try between adjacent pairs coupled with symmetry betweenremote bridging associations. In order to illustrate this pointmore concretely, we will explicitly simulate an extension of
TCM and fit it to the critical experimental data we reporthere.
Experiment
Subjects were presented with a list of paired associatescontaining both double-function and single-function pairs(see Figure 1). Single-function pairs, e.g. POLICE-WINDOW,are just standard, non-overlapping, paired associates. Thedouble-function pairs give rise to a linked-list (Figure 1,right). Linked-list lag provides a convenient means to de-scribe the distance between two items in the linked-list. Forinstance, consider the word PUPIL in the linked-list in Fig-ure 1. The correct response to the stimulus PUPIL is RIVER,which is one step forward in the chain at a linked-list lagof +1. If PUPIL was given as a probe and the subject re-sponded HOLLOW, this would be a backward intrusion as-sociated with a linked-list lag of −1. Items with a linked-list lag of ±1 were actually presented together as part of thesame pair. Larger absolute values of linked-list lag separatedouble-function items that were not presented together. Forinstance, the linked-list lag between PUPIL and DARLING is+2 and the linked-list lag from PUPIL to ABSENCE is −2.Linked-list lag allows us to compare the probability of intru-sions or recall transitions (see below) as a function of theirrecall direction by comparing linked-list lags with the sameabsolute value but different signs (e.g. comparing +1 to−1).We can also look for the presence of bridging associations bylooking for an effect of the absolute value of linked-list lag onassociations between pairs that were not presented together(absolute value of linked-list lag ≥ 2).
We will examine the associations formed between double-function items, both within and across pair, in two ways. Oneis to observe the intrusions subjects make during paired as-sociates testing. Subjects also performed surprise final freerecall (FFR) of all items at the end of the experimental ses-sion. Transitions in this final free recall period presumablyreflect the organization of information in memory in the rel-ative absence of strategies to edit out intrusions during cuedrecall testing. The CRP of recall transitions as a functionof linked-list lag during FFR provides a means to estimatethe strength of the associations between those items. Insofaras this measure reflects associations between items that werenot presented close together in time, this can be seen as ameasure of the extent to which participants’ memories wereable to extract the linked-list structure from the collection ofdouble function pairs.
Methods
Participants. Two-hundred-sixteen participants partici-pated for course credit in an introductory psychology classat Syracuse University.
Materials. Study words were chosen from the noun sub-set of the Toronto word pool (Friendly, Franklin, Hoffman,& Rubin, 1982). Study lists were composed of 35 double-function pairs consisting of 36 distinct words and 8 single-function pairs consisting of 16 distinct words. The double-
6 HOWARD ET AL.
function pairs were formed by choosing 36 words randomlywithout replacement from the pool for each subject. The firsttwo words were assigned to the first pair. The second andthird word were assigned to the second pair and so on.
Procedure. Participants learned the pairs over four study-test trials. On each study-test trial each pair was presentedfor study three times and then, after a delay, each pair wastested once. This makes a total of twelve presentations ofeach pair over the course of the experiment.
The order in which pairs were presented in each study-test trial was randomized separately for each participant sub-ject to the constraint that double-function pairs from succes-sive linked-list positions were never presented sequentially.This was done to ensure that any associations across double-function pairs “adjacent” in the linked-list could not be dueto temporal contiguity. Words were presented one at a timein an uppercase font for 1000 ms. Words were also presentedauditorially in a female voice. Presentations of items withina pair were separated by a delay of 100 ms, whereas pairswere separated by a delay of 1800 ms. Prior to each test,subjects performed a true-false arithmetic distractor for 30 s.This delay should be long enough to prevent a recency effecton the cued recall testing.
After the completion of the distractor task, each pair wastested one at a time. The order of tests was randomized, againsubject to the constraint that adjacent pairs in the linked-listwere never tested successively. On each test, the stimuluselement of each pair was presented visually and auditoriallyas before. Subjects were instructed to recall the word thatfollowed the probe in a study pair. Participants were given5 s to respond verbally to each probe.
Following the last study-test trial, participants were ad-ministered a surprise FFR test. The delay between the com-pletion of the last test item and the beginning of the FFRtest was typically a few minutes, which consisted of time tonotify the experimenter that the study-test cycles were com-plete, time for the experimenter to set up the FFR test andprovide the participant with instructions for the FFR period.The length of this delay was large relative to the spacingbetween the presentation of items—and each pair was pre-sented twelve times over several tens of minutes—so that re-cency effects in the FFR test would be expected to be negli-gible. Participants were given five minutes to recall as manystudy words as possible without regard to order. An experi-menter encouraged participants to continue attempting to re-call for the entire five minute period.
Analyses.We measured the proportion of remote (absolute value of
linked-list lag ≥ 2) double-function intrusions in response toa double-function probe item that came from each linked-listlag. Because pilot testing demonstrated edge effects in recallof the pairs such that the first and last pairs in the linked-listwere better recalled (presumably due to reduced associativeinterference from other double-function items), we omittedall intrusions in which the probe word or the remote intrusionwas from the first or last pair in the linked-list.
0 0.2 0.4 0.6 0.8 1P(R) Single Function Pairs
0
0.2
0.4
0.6
0.8
1
P(R
) Dou
ble
Func
tion
Pai
rs
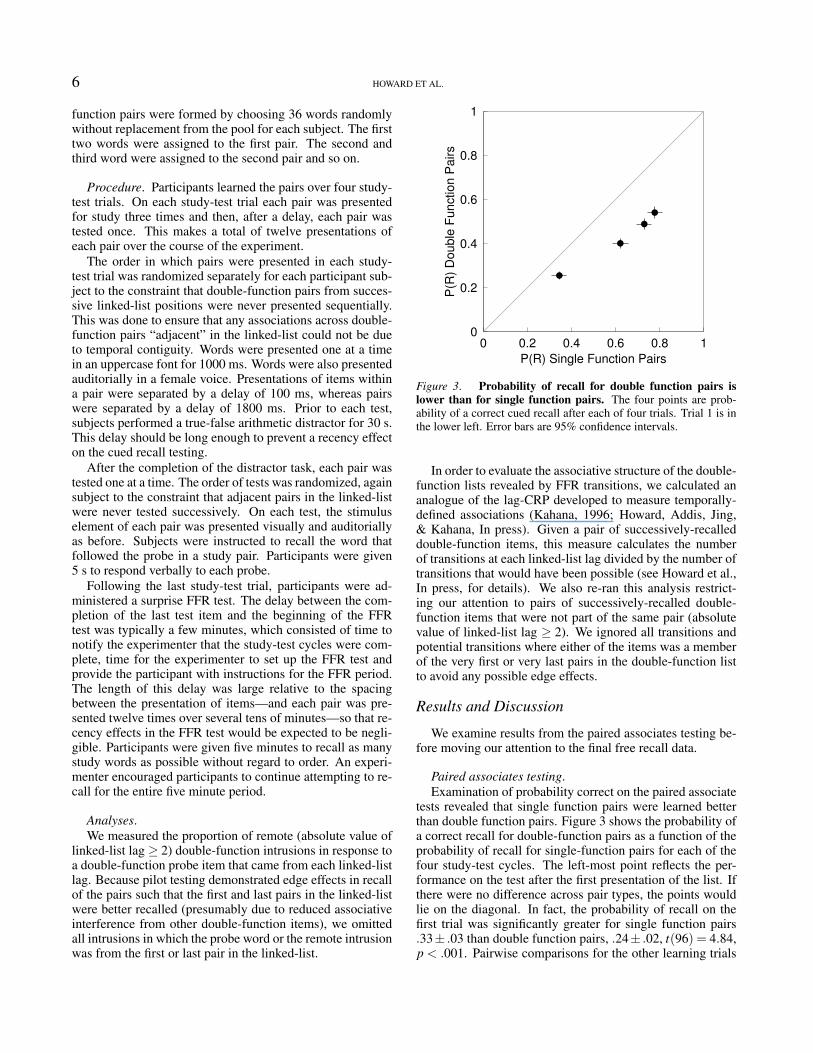
Figure 3. Probability of recall for double function pairs islower than for single function pairs. The four points are prob-ability of a correct cued recall after each of four trials. Trial 1 is inthe lower left. Error bars are 95% confidence intervals.
In order to evaluate the associative structure of the double-function lists revealed by FFR transitions, we calculated ananalogue of the lag-CRP developed to measure temporally-defined associations (Kahana, 1996; Howard, Addis, Jing,& Kahana, In press). Given a pair of successively-recalleddouble-function items, this measure calculates the numberof transitions at each linked-list lag divided by the number oftransitions that would have been possible (see Howard et al.,In press, for details). We also re-ran this analysis restrict-ing our attention to pairs of successively-recalled double-function items that were not part of the same pair (absolutevalue of linked-list lag ≥ 2). We ignored all transitions andpotential transitions where either of the items was a memberof the very first or very last pairs in the double-function listto avoid any possible edge effects.
Results and Discussion
We examine results from the paired associates testing be-fore moving our attention to the final free recall data.
Paired associates testing.Examination of probability correct on the paired associate
tests revealed that single function pairs were learned betterthan double function pairs. Figure 3 shows the probability ofa correct recall for double-function pairs as a function of theprobability of recall for single-function pairs for each of thefour study-test cycles. The left-most point reflects the per-formance on the test after the first presentation of the list. Ifthere were no difference across pair types, the points wouldlie on the diagonal. In fact, the probability of recall on thefirst trial was significantly greater for single function pairs.33± .03 than double function pairs, .24± .02, t(96) = 4.84,p < .001. Pairwise comparisons for the other learning trials

TRANSITIVE ASSOCIATIONS 7
were also highly significant. Moreover, the difference be-tween single and double-function pairs grew with learning.A repeated measures ANOVA showed significant main ef-fects of pair type (single vs double), F(1,672) = 391, MSe =6.83, p < .001, trial number, F(3,672) = 240, MSe = 4.20,p < .001, as well as a significant interaction of pair type andtrial F(3,672) = 11.5, MSe = .20, p < .001.
Most authors attribute the disadvantage for double func-tion pairs relative to single function pairs to associative inter-ference from competing responses, especially the backwardresponse (Primoff, 1938; Young, 1961). That is, if a subjectis given HOLLOW as a probe, the correct response is PUPIL.However, the backward association between HOLLOW andABSENCE will create a tendency to make a backward intru-sion (i.e. a response at linked-list lag −1) which could inter-fere with the correct response. Because single function pairsare not subject to interference from a backward association,this should create an advantage for performance on singlefunction pairs compared to double function pairs. Similarly,remote bridging associations compete with correct recalls fordouble function but not single function pairs.
Table 1 shows the proportion of responses of various typesto the double-function probes. As can be seen from inspec-tion of the table, there was a relatively large proportion ofbackward intrusions that persisted across trials. Nonetheless,the proportion of correct (forward) responses was larger ateach trial, indicating an asymmetry. At each of the four trialsin Table 1, there is a highly significant pairwise difference be-tween the probability of a correct response and the probabil-ity of a backward intrusion (paired t(215) from 9.87 to 19.7).To further quantify this apparent asymmetry, we ran a re-peated measures ANOVA on forward and backward responsetypes (the first two columns of Table 1) with trial as a factor.We found highly significant main effects of both response di-rection (forward vs backward), F(1,1505) = 1573.7, MSe =25.9, p < .001, and trial F(3,1505) = 100.1, MSe = 1.64,p < .001, as well as a highly significant interaction of trialand direction F(3,1505) = 106.93, MSe = 1.76, p < .001.5We conclude that there was a robust asymmetry between re-sponses of linked-list lags +1 and−1 during paired associatetesting.
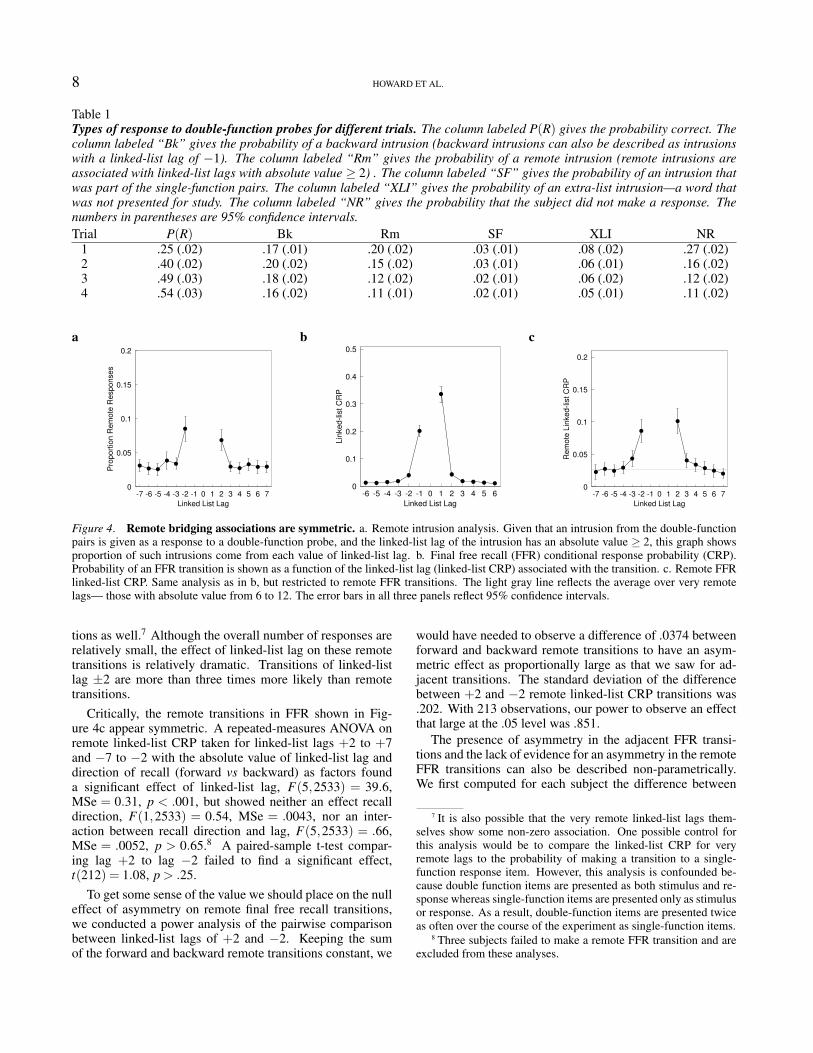
Table 1 also appears to show relatively large numbers ofremote linked-list intrusions. To address the critical questionof whether these remote bridging associations are symmetricor not we calculated the proportion of these remote intrusionsthat were observed at each linked-list lag. If the remote in-trusions were a consequence of bridging associations, thenwe would expect that linked-list lag would have a significanteffect on the probability of a remote intrusion probability.Figure 4a shows the result of these analysis collapsed overall test trials. First we note that there was a significant ef-fect of linked-list lag on remote intrusion proportion. Weconducted a repeated-measures ANOVA with absolute valueof linked-list lag (2 through 7 inclusive) and recall direction(forward vs backward) as factors. We found a main effectof linked-list lag, F(5,2567) = 24.8, MSe = 0.16, p < .001,providing unambiguous evidence for the presence of remotebridging associations during double-function learning. There
was no effect of direction, F(1,2567) = 1.56, MSe = 0.010,p > .2 nor an interaction of intrusion direction and linked-list lag F(5,2567) = 1.41, MSe = 0.009, p > .2. Despite thestrong asymmetry in adjacent responses (linked-list lags ±1,see Table 1) there was no corresponding asymmetry in theremote intrusions.
FFR transitions.Figure 4b shows the results of the linked-list CRP calcu-
lated from the FFR transitions. The first notable feature isa strong asymmetry between forward and backward adjacentlinked-list recall transitions. The linked-list CRP at a linked-list lag of +1 was significantly greater than that observed atlinked-list lag −1, paired-sample t(215) = 6.39, p < .001.The asymmetry was not only significant, but it was largewhen examined in terms of the size of the difference betweenthe means and also examined as an effect size. The differencebetween the linked-list CRPs at linked-list lags ±1 was .67the value at linked list lag −1. The difference between thelinked-list CRPs at linked-list lags ±1 was almost equal toone standard deviation of the linked-list CRP at lag −1 (.97).As with the responses to paired-associate testing, FFR tran-sitions showed a strong asymmetry among items that werepresented as part of the same pair, i.e. items at at adjacentlinked-list positions with lag ±1.
Figure 4c shows a version of the linked-list CRP thatonly considers remote transitions that bridge across double-function pairs. The curve appears peaked in the center, withtransitions to linked-list lags of ±2 more likely than moreremote linked-list lags and a gradual decline across severallinked-list lags. To confirm that the linked-list CRP waselevated for more than just the lags ±2, we compared thelinked-list CRP for small remote lags to those from more re-mote lags. To do so, we calculated the average over moreremote lags (absolute values from 6 to 12, inclusive). Themean value of the linked-list CRP for these more remotelinked-list lags was 0.027± .001 (see the thin gray line inFig. 4c). We then compared this value for more remotelinked-list lags to the linked-list CRP, averaged across for-ward and backward transitions, for specific values of linked-list lag. We found a significant difference for linked-list lags±2 (paired Wilcoxon V = 17382, p < .001) and for linked-list lags ±3 (paired Wilcoxon V = 11580, p < .05).6 Thisanalysis demonstrates that remote bridging associations ex-tend at least two pairs. That is, given B as a probe, thereare not only significant B-D associations, but B-E associa-
5 One might want to take the parametric statistics of this ANOVAwith a grain of salt due to the fact that the response values are notindependent of each other. If a response is correct, it is necessarilynot a backward intrusion. Similar concerns about a lack of indepen-dence can be raised for the intrusion analyses and final free recallCRP analyses that follow. For instance, if a response is an intrusionat linked-list lag −2, it cannot be an intrusion at linked-list lag −3.As a consequence, the proportion of intrusions at linked-list lag −2is not independent of the proportion of intrusions at linked-list lag−3.
6 Linked list lags ±2 and ±3 also differed from each other(paired Wilcoxon V = 8578, p < .001).
8 HOWARD ET AL.
Table 1Types of response to double-function probes for different trials. The column labeled P(R) gives the probability correct. Thecolumn labeled “Bk” gives the probability of a backward intrusion (backward intrusions can also be described as intrusionswith a linked-list lag of −1). The column labeled “Rm” gives the probability of a remote intrusion (remote intrusions areassociated with linked-list lags with absolute value ≥ 2) . The column labeled “SF” gives the probability of an intrusion thatwas part of the single-function pairs. The column labeled “XLI” gives the probability of an extra-list intrusion—a word thatwas not presented for study. The column labeled “NR” gives the probability that the subject did not make a response. Thenumbers in parentheses are 95% confidence intervals.Trial P(R) Bk Rm SF XLI NR
1 .25 (.02) .17 (.01) .20 (.02) .03 (.01) .08 (.02) .27 (.02)2 .40 (.02) .20 (.02) .15 (.02) .03 (.01) .06 (.01) .16 (.02)3 .49 (.03) .18 (.02) .12 (.02) .02 (.01) .06 (.02) .12 (.02)4 .54 (.03) .16 (.02) .11 (.01) .02 (.01) .05 (.01) .11 (.02)
a b c
-7 -6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6 7Linked List Lag
0
0.05
0.1
0.15
0.2
Pro
porti
on R
emot
e R
espo
nses
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6Linked List Lag
0
0.1
0.2
0.3
0.4
0.5Li
nked
-list
CR
P
-7 -6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6 7Linked List Lag
0
0.05
0.1
0.15
0.2
Rem
ote
Link
ed-li
st C
RP
Figure 4. Remote bridging associations are symmetric. a. Remote intrusion analysis. Given that an intrusion from the double-functionpairs is given as a response to a double-function probe, and the linked-list lag of the intrusion has an absolute value ≥ 2, this graph showsproportion of such intrusions come from each value of linked-list lag. b. Final free recall (FFR) conditional response probability (CRP).Probability of an FFR transition is shown as a function of the linked-list lag (linked-list CRP) associated with the transition. c. Remote FFRlinked-list CRP. Same analysis as in b, but restricted to remote FFR transitions. The light gray line reflects the average over very remotelags— those with absolute value from 6 to 12. The error bars in all three panels reflect 95% confidence intervals.
tions as well.7 Although the overall number of responses arerelatively small, the effect of linked-list lag on these remotetransitions is relatively dramatic. Transitions of linked-listlag ±2 are more than three times more likely than remotetransitions.
Critically, the remote transitions in FFR shown in Fig-ure 4c appear symmetric. A repeated-measures ANOVA onremote linked-list CRP taken for linked-list lags +2 to +7and −7 to −2 with the absolute value of linked-list lag anddirection of recall (forward vs backward) as factors founda significant effect of linked-list lag, F(5,2533) = 39.6,MSe = 0.31, p < .001, but showed neither an effect recalldirection, F(1,2533) = 0.54, MSe = .0043, nor an inter-action between recall direction and lag, F(5,2533) = .66,MSe = .0052, p > 0.65.8 A paired-sample t-test compar-ing lag +2 to lag −2 failed to find a significant effect,t(212) = 1.08, p > .25.
To get some sense of the value we should place on the nulleffect of asymmetry on remote final free recall transitions,we conducted a power analysis of the pairwise comparisonbetween linked-list lags of +2 and −2. Keeping the sumof the forward and backward remote transitions constant, we
would have needed to observe a difference of .0374 betweenforward and backward remote transitions to have an asym-metric effect as proportionally large as that we saw for ad-jacent transitions. The standard deviation of the differencebetween +2 and −2 remote linked-list CRP transitions was.202. With 213 observations, our power to observe an effectthat large at the .05 level was .851.
The presence of asymmetry in the adjacent FFR transi-tions and the lack of evidence for an asymmetry in the remoteFFR transitions can also be described non-parametrically.We first computed for each subject the difference between
7 It is also possible that the very remote linked-list lags them-selves show some non-zero association. One possible control forthis analysis would be to compare the linked-list CRP for veryremote lags to the probability of making a transition to a single-function response item. However, this analysis is confounded be-cause double function items are presented as both stimulus and re-sponse whereas single-function items are presented only as stimulusor response. As a result, double-function items are presented twiceas often over the course of the experiment as single-function items.
8 Three subjects failed to make a remote FFR transition and areexcluded from these analyses.

TRANSITIVE ASSOCIATIONS 9
forward and backward transitions. The adjacent transitionstook the difference between linked-list lag +1 and linked-list lag −1. Of the 204 subjects who showed a non-zerodifference in adjacent transitions, 129 of these had a higherprobability of a transition in the forward direction. This pro-portion is significantly different from 0.5 (p < .001). Forthe remote FFR transitions we took the difference betweenlinked-list lag +2 and linked-list lag −2. Of the 148 subjectswho showed a non-zero difference in the probability of for-ward and backward remote transitions, 79 showed a higherprobability of a forward transition. This proportion was notdifferent from 0.5 (p > 0.45).
The preceding analyses illustrated that there is evidencefor asymmetry for adjacent FFR transitions but not for re-mote FFR transitions. We directly compared the degreeof asymmetry from adjacent transitions to the degree ofasymmetry for remote transitions. We also directly com-pared the effect of recall direction on adjacent vs remoteFFR transitions. To compare the difference between adja-cent and remote transitions on a comparable footing, we di-vided the difference score used above for each subject bythe sum of forward and backward transitions averaged oversubjects. For the adjacent transitions, this asymmetry indexwas .25± .04. For the remote transitions, the asymmetryindex was .03± .07. Because of deviations from normal-ity, we compared the asymmetry indices for adjacent andremote transitions using a paired Wilcoxon test, V = 8428,p < .001.9 This analysis provides positive evidence that thedegree of asymmetry observed in adjacent FFR transitionswas greater than the degree of asymmetry observed in re-mote FFR transitions. From this we conclude that in addi-tion to there being no evidence for an asymmetry in the re-mote recall transitions, there is positive evidence that degreeof asymmetry in the remote recall transitions is smaller thanthat observed for the adjacent recall transitions.
Even if mediated chaining could be reconciled to the datafrom linked-list lags ±2, it would predict that the asymme-try persist to some degree throughout more remote linked-list lags. Examination of the more remote linked-list lags inFigure 4c reveals no evidence for even a trend in the rightdirection. These findings lead us to conclude that remotebridging associations are very unlikely to be a consequenceof mediated chaining.
Modeling
In this we section compare the ability of two models todescribe the central results of this experiment. We wanted toequate the ability of the models to describe simple temporalcontiguity effects while varying their sensitivity to temporalcontext. We compared a reformulation of TCM to a heteroas-sociative analogue that implements mediated chaining by en-coding the results of surreptitious retrieval. We restricted ourattention to the CRP as a function of linked-list lag in finalfree recall. This frees us from having to explicitly modelthe recognition processes that presumably edit out intrusionsduring cued recall, which would itself be a formidable chal-lenge. Indeed, Slamecka (1976) showed that false alarm
rate in a variant of associative recognition is affected by thelinked-list lag of the lure items.
TCMIn TCM, the cue for episodic recall is the current state of
temporal context, ti, which changes from time step i− 1 totime step i according to
ti = ρiti−1 +βtINi (1)
That is, the current state of context is composed of two parts:the previous state of context ti−1 and an input pattern tIN
icaused by the current item presented. The previous state ofcontext persists, multiplied by a scalar ρi, chosen such thatthe length of ti is always unity. This is added to an inputpattern tIN
i weighted by a parameter β that, together with thesimilarity of the input pattern to the previous state of context,controls the rate at which ti varies from time-step to time-step. With a set of uncorrelated inputs, as might be expectedfrom an infinitely long list of random words that appear once,Eq. 1 describes a random walk and we find that the innerproduct between states of context varies exponentially
ti · t j = ρ|i− j|, (2)
where ρ =√
1−β2. The property that temporal contextchanges gradually is indispensible for the ability of TCM toexplain recency and temporal contiguity effects.
Extending TCM as a model of learning. In order to fitthese data, we used an extension of TCM. Previously pub-lished versions of TCM were designed to describe tempo-ral contiguity effects between items presented once in arandomly-assembled series of items, as would typically bethe case in single-trial free recall (Howard & Kahana, 2002;Howard et al., 2006). The model was not designed to ad-dress situations in which items are repeated multiple times.In fact, the previously published versions of TCM are subjectto collapse of the input space. In TCM, the space spannedby the context vectors t is determined by the space spannedby the input vectors tIN . In TCM, a process of contextuallearning allows input patterns to come to resemble the tem-poral contexts in which they were presented. This propertyis extremely useful in describing backward and transitive as-sociations (Howard et al., 2005, 2006). However, the preciseform of contextual learning in previously published versionsof TCM had the undesirable property that with a sufficientnumber of presentations, the input vectors can collapse to a
9 Note that this comparison is not subject to concerns regardingindependence. First, the remote FFR CRP excludes responses atlinked-list lags ±1. This means that the values of the remote FFRCRP are independent of the values obtained at lags ±1 for the stan-dard FFR CRP. Second, we are comparing one (normalized) dif-ference score to another. Although the values at various linked-listlags that go into the difference scores are dependent, the differencescores should be independent of each other. This is so because theeffect of the value of the FFR CRP at one lag should have the sameeffect on both values going into the difference score.

10 HOWARD ET AL.
point (this point was discussed in Howard et al., 2005). Thatis, with runaway contextual learning, the tINs correspondingto all the words in the experiment become identical.
Here we use an extension of TCM that avoids runawaycontextual learning. This extension retains all of the keyfeatures of previously published versions of TCM. In fact itis precisely isomorphic to the model used by Howard et al.(2006) to describe single-trial free recall if items are pre-sented only once. The important differences between theextended model and previous treatments come when itemsare repeated multiple times. In this extension contextuallearning and encoding do not continue without bound. Twochanges prevent the catastrophic collapse. One is the use oflocal Widrow-Hoff learning rather than simple Hebbian outerproducts. Widrow-Hoff learning is a gradient descent algo-rithm that enables the model to stop learning when the encod-ing context of an item is well-predicted. The other change isthat the input patterns tIN caused by an item are composedof a fast-changing component we will refer to as hIN anda slowly-changing component we will refer to as cIN . Theslowly-changing component “anchors” the space so that itdoes not collapse completely. This extended formulation hasmany implications that do not bear directly on the ability ofTCM to describe these data. Here, we simply provide a bare-bones description of the model in the level of detail necessaryto explain the fits presented later. Finally, we note that previ-ously published versions of the model capture the qualitativefeatures of the final free recall CRP data—asymmetric adja-cent and symmetric remote associations—but do not achievethe excellent quantitative fits reported here (Howard & Jing,2003).
In previously published versions of TCM, the input pat-tern caused by presentation of a specific item A changes fromone presentation of that item to the next according to10
tINAi+1
= αOtINAi
+αNtAi−1. (3)
With a single repetition, this retrieved context includes a termthat changes relative to the initial presentation (tAi−1) and acomponent that is constant in direction from the first to sec-ond presentation (tIN
Ai). Because the former component over-
laps with the encoding contexts of items that both followedand preceded item A, it provides a symmetric cue. BecausetINAi
is only part of the encoding context for items that fol-lowed presentation of A, it is only a good retrieval cue foritems that followed A.11 The distinction between these twocomponents enabled TCM to provide a principled account oftemporal asymmetry (Howard & Kahana, 2002), the distinc-tion between pairwise and transitive associations (Howardet al., 2005) and a concise description of the mnemonicdeficit in cognitive aging (Howard et al., 2006). In the exten-sion we use here, we maintain the distinction between a com-ponent that changes rapidly from presentation to presenta-tion and a more slowly-changing component, but keep thesecomponents from becoming too thoroughly mixed. On theith presentation of item A, the input pattern tIN
Aiis composed
of cINAi
and hINAi
. These components cIN and hIN play rolesanalogous to tIN
Aiand tAi−1 in Eq. 3. The weighting of these
components are controlled by a parameter γ and the result isnormalized:
tINAi
=γcIN
Ai+(1− γ)hIN
Ai
||γcINAi
+(1− γ)hINAi||
(4)
The “hippocampal” component hINAi
follows a Widrow-Hoffalgorithm attempting to predict the context in which item Ais presented, controlled by a learning rate αH :
hINAi+1
= hINAi
+αH(tAi−1−hIN
Ai
). (5)
Note that learning stops when hINAi
= tAi−1. Put another way,hIN
A stops changing when it successfully matches the contextin which the item is presented. If hIN
A has not successfullypredicted the context in which it is presented, it changes inthe direction of that context.
The “cortical” input cINAi
follows a similar learning rule,but now it is trying to predict hIN
Ai:
cINAi+1
= cINAi
+αC(hIN
Ai− cIN
Ai
)(6)
That is, cINAi
is a representation of the contexts in which itemA has been presented, but passed through a series of low-passfilters. We assume in general that αC � αH ; in the simula-tions reported here we will assume that αC = 0 so that the setof cINs are fixed throughout the experiment.
Viewed in the light of the mapping hypothesis betweenTCM and the anatomy of the medial temporal lobe proposedby (Howard et al., 2005), these two components cIN and hIN
could correspond to inputs to parahippocampal cortices fromcortical association areas and the hippocampus respectively.This is consistent with known properties of medial temporallobe anatomy (see Eichenbaum, 2000, for a review). In thepresent treatment, we will not concern ourselves with explic-itly modeling how Eqs. 5 and 6 are implemented. It is worthnoting, however, that the physical structure of the system (seeFigure 5) suggests that recovery of hIN depends on the corti-cal input cIN perhaps modulated by the preceding contextualstate (Norman & O’Reilly, 2003).
As in previous formulations of TCM, the current state ofcontext serves as a probe for recall of an item via an associa-tive matrix MT F . However, unlike previous formulations, thematrix MT F is not simply a Hebbian outer product matrix. Itretains the property that items are encoded in their encodingcontexts. However, in a way analogous to the rules for con-textual learning above, learning is gradual and stops when an
10 This equation differs slightly from the one used in Howard et al.(2006) in that it does not include retrieval noise, which was usedto account for the discrepancy between younger and older adults’temporal contiguity effects. Retrieval noise also proves useful in ac-counting for dissociations between immediate recency and delayedrecall (Sederberg et al., in preparation).
11 Put another way, the state of context that was present when theitem was initially presented includes input patterns from the itemsthat preceded it.

TRANSITIVE ASSOCIATIONS 11
i
ci
f
t
h
h
Figure 5. Architecture of a reformulation of the temporal con-text model. In the model, an item layer (f), corresponding roughlyto a word recognition system, is connected to a temporal contextlayer (t). The temporal context layer is connected to a “hippocam-pal” layer that functions to recover prior states of temporal contextin which the item was previously encoded. When an item is pre-sented on f, it provides two inputs to the context layer in sequence:first a cortical input cIN that is assumed not to change over the scaleof minutes and then a hippocampal input hIN that rapidly changesto recover states of temporal context. Both components contributeto the state of t that cues recall of items on f.
item “fits well” into the context in which it is presented. LetwA be the row of MT F corresponding to item A:
w′A := f′AMT F ,
where fA is the pattern corresponding to item A and the primedenotes the transpose.12 Then, in our revised learning rule forMT F , wA is updated when item A is presented according to
∆wAi+1 = λ(tAi−1−wAi) . (7)
As with hIN above, learning stops when wA is perfectlyaligned to the context tAi−1 in which A is presented.
In TCM, context is used as a cue for recall of items. Givena particular context cue t, this activates the item layer accord-ing to
fIN = MT F t (8)
where t is the probe context at the time of test (see, e.g.Howard & Kahana, 2002, also Figure 5). This input ontothe item layer activates each item i to an extent ai, defined bythe inner product f′ifIN , where the definition of fIN is given inEq. 8 and the prime again denotes the transpose.13 This quan-tity is equal to the inner product between the probe contextand the vector wA describing the average context in whichitem A is presented. Given the set of activations, the prob-ability of recalling item i from the list is given by the Lucechoice rule:
PR(i) =exp2ai/τ
∑ j exp2a j/τ. (9)
The sum in the denominator runs over all potential recalls14
In Eq. 9, the parameter τ controls the sensitivity of the re-trieval rule. As τ → ∞, the probability of recalling each itembecomes equal. As τ → 0, the probability of recalling themost activated item becomes one. For this reason, Eq. 9 isreferred to as a softmax rule in the connectionist literature.Other schemes for mapping a set of activations onto recallof an item are certainly possible (e.g. Sederberg et al., inpreparation). The Luce choice rule should not be consid-ered central to the assumptions of TCM, but rather a min-imal implementation that is convenient to work with. Thekey requirements on this component of the model are that itdepend on the relative rather than absolute activation of anitem in order to explain the long-term recency effect and thatit be sufficiently non-linear.
Heteroassociation and mediated chaining
There are a great many differences between TCM and anyspecific extant heteroassociative model. Our goal in model-ing here is to build a heteroassociative analogue of TCM thatdiffers from TCM in as few respects as possible other thanthe contextual retrieval hypothesis that is central to TCM.This should allow us, as much as possible, to evaluate theimplications of the contextual retrieval vs heteroassociativeexplanations of temporal contiguity in relative isolation fromthe other features necessary to build and test a simulation.We thus built a heteroassociative model which shares withTCM exponential forgetting (Eq. 2) and a competitive non-linear retrieval rule (Eq. 9). We also endowed this model withthe ability to describe forward and backward asymmetry intemporal contiguity effects, although this was simply hard-coded and not a consequence of a principled mechanism asin TCM. The end product is a heteroassociative model thatis identical to TCM in its ability to describe temporal conti-guity effects between items presented once in a series suchas in single-trial free recall, but that differs in the structuralassumptions that lead to this contiguity effect.
In TCM, in a list of non-repeating items, context changesgradually from time step to time step such that the similar-ity between the encoding context associated with item i anditem j within the list falls off exponentially (Eq. 2). Let usdefine an associative matrix S that defines a heteroassocia-tive strength between each item in the list and all the otheritems in the list. Following the SAM model (Raaijmakers &Shiffrin, 1980), we initialize S with a residual value betweeneach item, b. On learning items presented in a series, weincrement the associative strength between item i and item j
12 Describing w as a row of MT F assumes that the fs are not onlyorthonormal, but that we have rotated the space so that the f corre-sponding to the words are the bases of the space.
13 Here, as in previous treatments of TCM, we assume that theitem patterns fi are orthonormal. Interestingly, it is possible tomodel semantic relationships betwen words even with orthonormalitem vectors if the overlap in the w’s and tINs, corresponding to thetemporal contexts in which the item is presented, overlap.
14 In calculating CRPs we omit the just-recalled word from thissum.

12 HOWARD ET AL.
according to
∆Si j =
{AF ρ
|i− j|F , i < j
ABρ|i− j|B , i > j
. (10)
That is, for items presented in series, the increment to theassociative strength between items in series is an exponentialfunction of the distance between them. 15 The heteroassocia-tive model implements asymmetry between items presentedin series because there are separate parameters A and ρ for as-sociations formed between forward and backward associates.
We need a means to map this associative matrix onto recallof a particular item. We used the same Luce choice rule usedto select an item in TCM (Eq. 9). Given an item i as a cue,the probability of recalling item j will be given by a Lucechoice rule:
P( j|i) =expSi j/τ
∑k expSik/τ. (11)
In single trial free recall, the predictions for the heteroas-sociative model regarding the CRP are identical to those ofTCM if b = 0,
AF/AB =1− γ+ γβ
1− γ
andρF = ρB =
√1−β2.
The As are specified up to a constant, which could be ab-sorbed by changing the values of τ in the models accordingly.
We also tried a variant of the Luce choice rule in which theexponential terms go like exp(Si j/τ−1). This adjustmentmeans that the asymptotic level of recall of a non-activateditem is zero rather than a non-zero value as in Eqs. 9 or 11.
Although the heteroassociative model does not provide aprincipled account of asymmetry or transitive associations,this model can make temporal contiguity functions indistin-guishable to those of TCM for free recall data, includingthose from continuous distractor free recall.
In order to make it possible for the heteroassociative to de-scribe remote bridging associations across pairs we alloweda phase where the model was given probe items from thelist and then generated a series of recalls. This sequence ofrecalls was then subject to learning according to Eq. 10. Theidea is that if the model is given A as a probe, this can resultin the recall of B. Given B as a probe, C may be recalled,resulting in A and C being experienced in close temporalproximity. This scheme may even be able to address thedifference in asymmetry between adjacent and remote recalltransitions. If A and C are experienced at a lag of 2, thenboth cue strengths, SAC and SCA, will be incremented. Al-though these may be asymmetric due to the asymmetry builtin to Eq. 10, the difference may become small relative to theresidual association b at more remote lags. That is, if A andB are experienced at lag 1 N times, the asymmetry in the cuestrength goes like
SAB
SBA=
NAF ρF +bNABρB +b
,
whereas if A and C are presented at lag 2 N times, the asym-metry is given by
SAB
SBA=
NAF ρ2F +b
NABρ2B +b
.
Assuming that the parameters have been chosen so that thereis asymmetry between forward and backward recalls,16 thislatter quantity is clearly smaller than the former because theeffect of b will be larger as the exponential terms decay withincreasing lag. The number of times a particular lag is expe-rienced is itself a function of the other model parameters andthe particular sequence of retrievals that have taken place forthe item. Because of the difficulty in solving for N we willexplicitly simulate the model.
Methods.In simulating TCM, we initialized the cIN to be mutually
orthonormal. The hINA and wA for each item were intial-
ized to be null vectors. The initializations of hIN and ware not essential to fit these data. We presented TCM witha list of double-function pairs repeated three times in a ran-domized order four times to correspond to the four study-test cycles. After learning, we estimated the final free recallCRP by probing with each item and calculating the proba-bility of recall of each item in the list and averaging overlinked-list lags. For simplicity, we did not allow learning totake place during recall. To avoid any possibility that asso-ciations across pairs could be attributable to simple tempo-ral contiguity during study, we put an “infinitely long dis-tractor interval” between each pair. That is, we presenteda distractor—an input pattern orthonormal to every previousinput pattern—with β 1 before the first member of each pair(we refer to this value as βI). From Equation , this means thatthe effect of the preceding pair on context is completely lostwhen the first member of the subsequent pair is presented.By presenting a different distractor between each pair, weensure that the last word of a pair, and the first word of thenext pair do not share any context.
We allowed five parameters to vary in fitting TCM. Onewas β, which controlled the degree of contextual overlapwithin pair (Eq. 1). Two other parameters, γ and αH con-trolled the degree to which the rapidly-changing “hippocam-pal” component contributed to retrieved context (Eq. 4) andchanged with repeated presentations (Eq. 5) respectively.The learning rate on the slowly-changing “cortical” compo-nent, αC was fixed at zero. One parameter, λ, controlled therate of learning on MT F (Eq. 7), and τ controlled the sensi-tivity of the Luce choice rule (Eq. 9).
In simulating the heteroassociative encoding model, wealso included four study-test cycles. Prior to the first studylist, the matrix S was initialized such that every value was setto the residual value b. On each study-test cycle, the within-pair associations were incremented according to Eq. 10 with
15 In this model the self-strength Sii does not play a role and willbe ignored.
16 This also assumes that the ρs are less than 1.

TRANSITIVE ASSOCIATIONS 13
the lag between the first member of a pair and the secondas +1 and the lag between the second and first membersof a pair as −1. That is, all forward within-pair strengthsSAB, SBC, etc were incremented by AF ρF and all backwardwithin-pair strengths were incremented by ABρB. After eachincrement corresponding to the study list, a round of test-phase learning was undertaken. As in the fits to TCM, wedid not try to fit the model to the probability of correct recallor any other behavioral measure collected from the cued re-call testing. However, because the heteroassociative modelrelies on encoding of surreptitous recalls to enable the for-mation of bridging associations, we explicitly simulated therecall process and allowed output encoding to operate duringthese recall attempts. Each double-function item was pre-sented as a probe. The cue strengths were then used to gen-erate a recall. This recalled item was then used as a probeto recall another item. A total of n recalls were made. Thevalue of n was fixed at five for these simulations.17 The asso-ciative strength between all of the n + 1 items including theprobe were incremented using Eq. 10 with the appropriatelag. This procedure was repeated with each double functionitem as a probe. For simplicity, the weights were updatedafter all probe items were presented and before the next pre-sentation of the study list. After all four study-test cycleswere complete, the linked-list CRP was calculated using thesame procedure as used for TCM. In an attempt to enhancethe heteroassociative model’s ability to fit the data, we alsotried a variant in which the exponential terms in the Lucechoice rule were altered as described above (see discussionfollowing Eq. 11).
In fitting the heteroassociative model we also allowed fiveparameters to vary freely. As in TCM, τ controls the sensitiv-ity of the Luce choice rule. The rate parameters controllingforward and backward decay in associative strength, ρF andρB were allowed to vary freely. The parameter b that con-trols the residual background strength across items was alsoallowed to vary freely. Not all of the parameters in the modelare identifiable. Accordingly, we allowed the forward coef-ficient AF to vary freely but fixed AB at one. As discussedabove, we fixed the number of retrieval attempts per probeat five. Because the heteroassociative model displayed someevidence for noise for some values of the parameters, we av-eraged across 100 replications of the model for each set ofparameter values.
For both models, we used a downhill simplex method(Nelder & Mead, 1965) to minimize the χ2 between the finalfree recall CRP from linked-list lags −6 to +6 (inclusive).We started the simplex at a variety of starting values to avoidlocal minima. The best-fitting parameters for all three mod-els are shown in Table 2.
Results and Discussion.Figure 6a shows the best-fitting solution of TCM to the fi-
nal free recall linked-list CRP. As can be seen from the figure,the fit is excellent. TCM described the qualitative featuresof the data—a larger asymmetry for the adjacent vs remoterecalls as well as robust transitive associations as manifestby the greater probability of recall for the items at ±2 than
Table 2Best-fitting parameter values for the three models tested.The column labeled “TCM” refers to the temporal contextmodel. The column labeled “het1” gives the best-fittingparameters for the heteroassociative encoding model. Thecolumn labeled “het2” gives the best-fitting parameters fora variant of the heteroassociative model in which the Lucechoice rule was altered. See text for details.
TCM het1 het2β .80 AF 1.08 AF .82αH .66 AB 1 AB 1γ .26 ρF .57 ρF .93λ .079 ρB .52 ρB .40τ .063 τ .18 τ 1.82βI 1 b .098 b .0098αC 0 n 5 n 5
for those at more remote linked-list lags. The asymmetry atlinked-list lags ±1 is possible because the fixed componentcIN
A contributes only to forward associations, which results ina boost to the CRP at linked-list lag +1. The CRP at linked-list lag−1, and at more remote linked-list lags, is not affectedby cIN
A as a probe, but is controlled by the similarity of hINA
to the w corresponding to the other recalled items. This isbecause cIN
A only reliably contributes to the w for words thatfollowed item A. TCM provided a outstanding quantitative fitto the data, χ2(6) = 4.36, n.s., indicating that deviations fromthe model’s values are not different than would be expectedby chance.
Although the reformulated TCM does well at describingthe large difference between adjacent and remote asymme-try, the best-fitting values did show a slight asymmetry in theremote bridging associations (.046 at lag +2 vs .041 at lag−2). Associations in TCM are a consequence of the degreeof overlap between the tIN for one item and the w for another.The fixed part of tIN , which we’ve been referring to as cIN ,does not contribute to transitive or backward associations.The rapidly-changing portion of tIN , hIN is solely responsi-ble for transitive associations. The vectors hIN and w bothtry to estimate the temporal contexts in which the item ispresented. However, the learning rates for these two vectorsare not in general the same. In the limit as the list is presentedan infinite number of times, wA and hIN approach the sameasymptotic value and symmetric remote associations result.The asymmetry comes from one lagging behind the other onthe way to this steady state solution. Perfect symmetry in theremote associations, as well as asymmetry in either direc-tion, are possible during learning in the reformulated TCMwith appropriate learning rates. All of these alternatives canbe simultaneously observed with asymmetric adjacent asso-ciations between linked-list lags ±1.
17 It may seem unreasonable that participants retrieve five itemsin response to each test probe. We set this value to a relatively highvalue reasoning that it gave the model the best chance to developrobust remote bridging associations.
14 HOWARD ET AL.
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6Linked list lag
0
0.1
0.2
0.3
0.4FF
R C
RP
Figure 6. TCM describes the form of transitive associationsobserved in final free recall. The symbols with error bars repre-sent the experimental data with 95% confidence intervals. The thickgrey lines that appear to connect the data points are the best-fittingvalues from TCM.
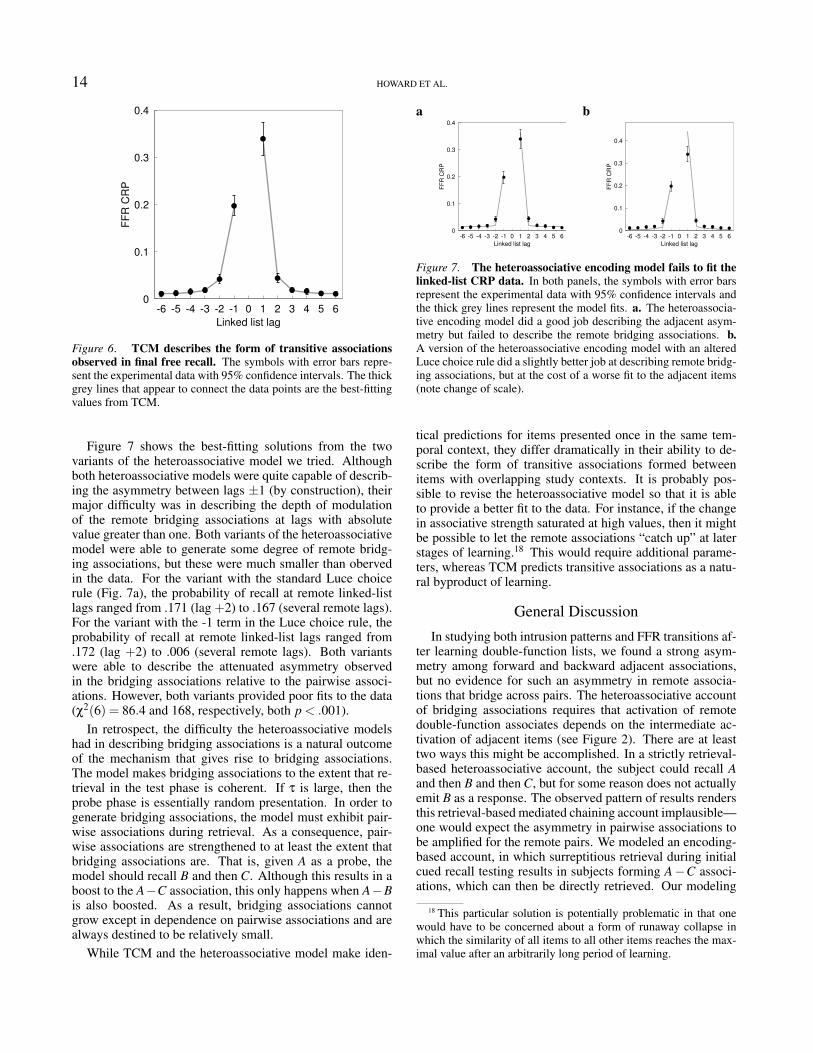
Figure 7 shows the best-fitting solutions from the twovariants of the heteroassociative model we tried. Althoughboth heteroassociative models were quite capable of describ-ing the asymmetry between lags ±1 (by construction), theirmajor difficulty was in describing the depth of modulationof the remote bridging associations at lags with absolutevalue greater than one. Both variants of the heteroassociativemodel were able to generate some degree of remote bridg-ing associations, but these were much smaller than obervedin the data. For the variant with the standard Luce choicerule (Fig. 7a), the probability of recall at remote linked-listlags ranged from .171 (lag +2) to .167 (several remote lags).For the variant with the -1 term in the Luce choice rule, theprobability of recall at remote linked-list lags ranged from.172 (lag +2) to .006 (several remote lags). Both variantswere able to describe the attenuated asymmetry observedin the bridging associations relative to the pairwise associ-ations. However, both variants provided poor fits to the data(χ2(6) = 86.4 and 168, respectively, both p < .001).
In retrospect, the difficulty the heteroassociative modelshad in describing bridging associations is a natural outcomeof the mechanism that gives rise to bridging associations.The model makes bridging associations to the extent that re-trieval in the test phase is coherent. If τ is large, then theprobe phase is essentially random presentation. In order togenerate bridging associations, the model must exhibit pair-wise associations during retrieval. As a consequence, pair-wise associations are strengthened to at least the extent thatbridging associations are. That is, given A as a probe, themodel should recall B and then C. Although this results in aboost to the A−C association, this only happens when A−Bis also boosted. As a result, bridging associations cannotgrow except in dependence on pairwise associations and arealways destined to be relatively small.
While TCM and the heteroassociative model make iden-
a b
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6Linked list lag
0
0.1
0.2
0.3
0.4
FFR
CR
P
-6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6Linked list lag
0
0.1
0.2
0.3
0.4
FFR
CR
P
Figure 7. The heteroassociative encoding model fails to fit thelinked-list CRP data. In both panels, the symbols with error barsrepresent the experimental data with 95% confidence intervals andthe thick grey lines represent the model fits. a. The heteroassocia-tive encoding model did a good job describing the adjacent asym-metry but failed to describe the remote bridging associations. b.A version of the heteroassociative encoding model with an alteredLuce choice rule did a slightly better job at describing remote bridg-ing associations, but at the cost of a worse fit to the adjacent items(note change of scale).
tical predictions for items presented once in the same tem-poral context, they differ dramatically in their ability to de-scribe the form of transitive associations formed betweenitems with overlapping study contexts. It is probably pos-sible to revise the heteroassociative model so that it is ableto provide a better fit to the data. For instance, if the changein associative strength saturated at high values, then it mightbe possible to let the remote associations “catch up” at laterstages of learning.18 This would require additional parame-ters, whereas TCM predicts transitive associations as a natu-ral byproduct of learning.
General Discussion
In studying both intrusion patterns and FFR transitions af-ter learning double-function lists, we found a strong asym-metry among forward and backward adjacent associations,but no evidence for such an asymmetry in remote associa-tions that bridge across pairs. The heteroassociative accountof bridging associations requires that activation of remotedouble-function associates depends on the intermediate ac-tivation of adjacent items (see Figure 2). There are at leasttwo ways this might be accomplished. In a strictly retrieval-based heteroassociative account, the subject could recall Aand then B and then C, but for some reason does not actuallyemit B as a response. The observed pattern of results rendersthis retrieval-based mediated chaining account implausible—one would expect the asymmetry in pairwise associations tobe amplified for the remote pairs. We modeled an encoding-based account, in which surreptitious retrieval during initialcued recall testing results in subjects forming A−C associ-ations, which can then be directly retrieved. Our modeling
18 This particular solution is potentially problematic in that onewould have to be concerned about a form of runaway collapse inwhich the similarity of all items to all other items reaches the max-imal value after an arbitrarily long period of learning.

TRANSITIVE ASSOCIATIONS 15
results suggest that this variation of the heteroassociative ac-count is not easily fit to the data. In contrast, TCM providesan excellent quantitative fit to the data.
It may be possible to reconcile the present results witha significantly elaborated heteroassociative model. For in-stance, if all one wanted to do was explain our findings ofremote bridging associations, a symmetric heteroassociativemodel would have no difficulty. The problem is in recon-ciling this with the robust asymmetry observed at linked-listlags±1. If one could somehow make a symmetric model ap-pear asymmetric, the heteroassociative model would do a finejob. As mentioned earlier, a broad class of distributed mem-ory models based on convolution have associative symmetryas a natural prediction. This is consistent with the associativesymmetry hypothesis (Asch & Ebenholtz, 1962), which pre-dicts that forward and backward associations should not onlybe equally strong on average (Murdock, 1966), but that thepair should either be encoded as a whole or not at all. (Ka-hana, 2002) showed there were indeed strong correlations be-tween forward and backward tests of pairs tested twice, con-sistent with the associative symmetry hypothesis (see alsoRizzuto & Kahana, 2001).
The robust asymmetry we observed in FFR transitions atlinked-list lags ±1 would seem to argue strongly against theassociative symmetry hypothesis. Although Kahana (2002)reviewed the evidence for the associative symmetry hypoth-esis and found the literature generally supportive, associativesymmetry is observed when subjects receive many trials on apair. The interaction of recall direction and trial we observedin comparing correct recall to backward intrusions (Table 1)is consistent with this finding. However, even after the firststudy-test cycle, when pairs were presented only three times,there was still a significant difference between correct recallsand backward intrusions. This could be a consequence of theinstructions to recall only in the forward direction—perhapswhen given B as a cue, participants generate A and C equallyoften but can identify A as a backward associate and edit itout of their recall protocols. Associative symmetry need notimply that participants retain no information about the orderin which items were presented. It is also possible that stan-dard paired associate learning data appears symmetric be-cause participants are aware that there will be both forwardand backward recall tests, which may induce participants toadopt an encoding strategy that is different from what theywould ordinarily engage in learning pairs of successively-presented items. It is clearly difficult to examine associativesymmetry without giving participants backward recall tests.
Heteroassociative accounts of contiguity effects inepisodic recall tasks are quite intuitive when viewed froma certain perspective. They have the advantage of parsimonyif the only goal to be accomplished is to provide a simple as-sociation between two stimuli, which is the minimum neces-sary to describe paired associate learning. It is probably pos-sible to construct a more elaborate heteroassociative modelthat would provide a better fit to the data. It is also possi-ble that one could construct a model that retains heteroas-sociative mechanisms to account for the asymmetry betweenlinked-list lags±1 but that utilizes contextual recovery to ac-
count for the symmetric across-pair associations. However,it is not clear that it is theoretically desirable to retain het-eroassociative mechanisms to account for temporal contigu-ity effects in episodic recall. The retrieved context accountmay be more intuitively appealing if one starts from a slightlydifferent perspective.
Perspectives on temporal contiguity
Tulving (1983, 2002) characterizes episodic memory as“mental time travel” in which one remembers not only“what” but also “when” and “where” of a memory. Two-process theorists of item recognition describe recollection asthe vivid recovery of contextual information associated witha probe item (e.g. Yonelinas, 2002). Starting from this per-spective, the idea that episodic recall, even paired associatelearning, is largely a consequence of binding and recoveryto a representation of spatio-temporal context seems muchmore natural than a simple heteroassociative account. Simi-lar arguments in favor of the intuitive appeal of a contextualrecovery account of temporal contiguity effects in episodicmemory can be made starting with considerations from neu-robiology and cognitive neuroscience. The hippocampus,a medial temporal lobe structure widely believed to be es-sential for episodic memory, also maintains a representationof location, or spatial context, as an animal moves throughits environment (e.g. O’Keefe & Dostrovsky, 1971; Wilson& McNaughton, 1993). Burgess and colleagues (Burgess,2002; Burgess, Maguire, & O’Keefe, 2002) have argued thatthe primary function of the hippocampus in spatial naviga-tion tasks is to enable binding of item representations to arepresentation of the spatial context in which they were en-countered (see also Howard et al., 2005; Smith & Mizumori,2006). From this perspective, the contextual recovery ac-count of temporal contiguity effects seems much more intu-itive than a heteroassociative account.
There is also evidence from animal learning paradigmsthat argues against a heteroassociative account of even sim-ple conditioning. We review three phenomena, secondarygeneralization, temporal encoding in trace conditioning, andtransitive learning in conditional discriminations. These phe-nomena share the property that at first glance the tasks wouldseem able to be accomplished by means of heteroassociation,but on closer examination display features that have led au-thors to eschew a simple heteroassociative account.
Secondary generalization, which has been experimentallyobserved in pigeons (e.g. Wasserman, DeVolder, & Cop-page, 1992), refers to the learning setting in which two ar-bitrary stimuli are associated to a common response. Sub-sequently, pairing one of the stimuli to another responseresults in generalization such that the other stimulus alsoevokes the new response without explicit pairing. That is,if stimulus S1 and stimulus S2 are separately paired withresponse R1 in an operant conditioning task, then if S1 ispaired with R2, S2 will generalize to R2 even if S2−R2 is nottrained. Secondary generalization is analogous to transitiveassociation—the secondary generalization develops after thestimuli are experienced in the temporal context of a particu-

16 HOWARD ET AL.
lar response. Rather than appealing to a heteroassociative ac-count based on mediated chaining (Hull, 1947), Wassermanet al. (1992) attributed the finding of secondary generaliza-tion in pigeons to the development of conceptual categories.
Trace conditioning refers to a classical conditioningparadigm in which the offset of the conditioned stimulus(CS) and the onset of the unconditioned stimulus (UCS) areseparated by a temporal delay. Trace conditioning is sensi-tive to hippocampal damage, whereas delay conditioning (inwhich the offset of the CS coincides with the UCS) is un-affected by hippocampal lesions (Solomon, Vander Schaaf,Thompson, & Weisz, 1986). Cole, Barnet, and Miller (1995)studied trace conditioning in the rat and concluded that aheteroassociative account of associations between stimulusand response was insufficient. In their Experiment 2, Coleet al. (1995) paired two auditory stimuli CS1 and CS2, each5 s in duration, such that presentation of CS2 immediatelyfollowed the offset of CS1. Later CS1 was paired with anunconditioned stimulus (foot shock) such that presentationof the unconditioned stimulus followed the offset of CS1 byeither zero or five seconds. The choice of delays and theduration of CS2 was such that in the five second condition,the onset of the UCS corresponded to the time at which CS2would have terminated had it been presented as during theinitial pairing of CS1 and CS2. Although conditioned re-sponding to CS1 was greater after training in the zero secondcondition, the conditioned responding to CS2 via second or-der transfer was greater in the five second condition.
Cole et al. (1995) interpreted their remarkable results asevidence that CS1 and CS2 could not be connected by asimple, direct associative link between the two stimuli, butrather reflected information about the temporal relationshipsimplied by their pairing (see also Gallistel & Gibbon, 2000).Moreover, Cole et al. (1995) emphasize that in order to con-nect the CS2 to the unconditioned stimulus, it is necessaryto integrate information about the relationship between CS1and CS2 and the relationship between CS1 and the UCS tocreate information about the relationship between CS2. Thisis analogous to the need to integrate information about A−Band B−C to create information about the A−C associationin the present experiment.
Bunsey and Eichenbaum (1996) examined contextuallearning in an analogue of the paired associates task appro-priate for use with rodents. In this task, rats performed aconditional discrimination. In the sample phase of the condi-tional discrimination they sampled a cue odor, either A or X .In a choice phase, they had to approach one of a pair of odorsB or Y to receive a reward. The identity of the cue odor deter-mined which of the choice odors were rewarded. In this waythe rats learned associations A−B and X −Y . After learningthe first conditional discrimination the choice odors becamecue odors for a subsequent pair, so that the animals learnedoverlapping pairs A−B and B−C. In probe trials, animalswere presented the cue odors from the first pairs and given achoice between the choice odors from the second pair. Thisprovides a means to examine bridging A−C associations.While animals with hippocampal lesions were able to learnA−B and B−C as well as sham-lesioned animals, they dif-
fered from the sham-lesioned animals in that they showed noevidence for bridging A−C associations. Dusek and Eichen-baum (1997) showed evidence for a similar dissociation aftertraining animals on a so-called transitive inference task (re-viewed below). Other evidence suggests that similar deficitsin forming and/or utilizing transitive associations may be as-sociated with aging (Ohta et al., 2002; Provyn, Sliwinski,& Howard, submitted), suggesting a hippocampal locus foraging deficits (Small, Chawla, Buonocore, Rapp, & Barnes,2004; Wilson, Gallagher, Eichenbaum, & Tanila, 2006).
To Eichenbaum (2001), the effect of hippocampal lesionon learning of overlapping pairs of odors (Bunsey & Eichen-baum, 1996; Dusek & Eichenbaum, 1997) suggests that thehippocampus is responsible for maintaining and organizinginformation about the spatial and temporal relationships be-tween items. In this view, the ability to organize stimuli into a“memory space” is considered to be a central property of thehippocampus’ function in support of declarative and episodicmemory. This conception can be seen as integrating the posi-tions of Wasserman et al. (1992) and Cole et al. (1995). TCMcan be seen as a quantitative implementation of a memoryspace in which the patterns tIN evoked by the items reflect thetemporal relationships between those items. These includenot only temporal relationships that are explicitly trained, i.e.A−B pairs, but also latent temporal relationships that are notexplicitly presented.
Transitive inference and the hippocampus
Building on the findings of Bunsey and Eichenbaum(1996), Dusek and Eichenbaum (1997) examined the effectsof hippocampal lesion on a task that may exploit transitiveassociations between items. In this task, rats are trained ona series of conditional discriminations between odor pairs.These odor pairs are arranged in a hierarchy, expressed asA > B > C > D > E. When presented with a pair, the rat isrewarded for selecting the odor that has a greater value in thehierarchy. For instance, given a pair of odors, B and C, therat is rewarded for approaching B and avoiding C. However,given a pair C and D, the rat is rewarded for approaching Cand avoiding D. Odor A is always rewarded and odor E isnever rewarded. After learning adjacent pairs to a criterion,one can then ask whether the animal successfully generalizesto novel pairs that have not been presented together. The end-anchored pair, A−E can be solved simply because A wasalways rewarded during learning and E was never rewardedduring learning. Of particular interest is whether the animalselects B over D when this novel pair is presented. In order todo so, the rat must somehow integrate the knowledge aboutthe separate training pairs.
Dusek and Eichenbaum (1997) showed that rats with hip-pocampal lesions learned the adjacent pairs as well as intactanimals, and were equally good at generalizing to the end-anchored pairs. However, lesioned animals were impairedon generalizing to the critical B−D pairing. The findings ofDusek and Eichenbaum (1997) led to a relatively large lit-erature studying this “transitive inference” task and the roleof the hippocampus in supporting this behavior. Smith and

TRANSITIVE ASSOCIATIONS 17
Squire (2005) replicated the basic effect in human amnesiapatients. Neuroimaging studies have argued for hippocam-pal activation associated with learning of transitive inference(Greene, Gross, Elsinger, & Rao, 2006; Heckers, Zalesak,Weiss, Ditman, & Titone, 2004). Titone, Ditman, Holzman,Eichenbaum, and Levy (2004) argued for a similar patternof results with schizophrenia patients, noting that this sug-gests altered hippocampal-dependent cognitive function inschizophrenia.
There has also been a very active debate about whetherit is appopriate to think of performance in the transitive in-ference task as a true logical inference or not. One couldsolve the novel B−D pairing in the same way that if onelearns that Alice is taller than Ben, and Ben is taller thanCatherine, then one can infer that Alice is taller than Cather-ine without explicitly being told. In this view, the stimulithat enter into the training pairs acquire a unidimensionalattribute analogous to height—call it “goodness.” The par-ticipant then consciously reasons with this property to solvenovel pairings. It is also possible that the problem is solvedwithout conscious reasoning by means of as associative pro-cess binding the items differentially to reward (Frank, Rudy,& O’Reilly, 2003; Van Elzakker, O’Reilly, & Rudy, 2003).Logical reasoning would seem to depend on awareness of thehierarchical relationship, which can be assessed in studieswith humans by asking participants about their knowledgeof the relationship after learning. Greene, Spellman, Dusek,Eichenbaum, and Levy (2001) argued that successful gener-alization on B−D pairings was not dependent on awarenessof the hierarchical relationship (see also Frank, Rudy, Levy,& O’Reilly, 2005). This view has been challenged by Smithand Squire (2005) who observed robust performance on thenovel pairings only by participants who reported awarenessof the organization of the pairs (see also Moses, Villate, &Ryan, 2006). The distinction has theoretical implications forconcepts that describe declarative memory as dependent onthe integrity of the hippocampus and necessarily accompa-nied by awareness (Clark & Squire, 1998).
Regardless of whether performance on the transitive in-ference task depends on conscious logic or not, it certainlywould seem to depend on the development of bridging asso-ciations across premise pairs. Connectionist and neural net-work models of hippocampal function have taken the find-ings of the role of the hippocampus in relational memory(Bunsey & Eichenbaum, 1996; Dusek & Eichenbaum, 1997)as a benchmark property characteristic of hippocampal func-tion (e.g. O’Reilly & Rudy, 2001; Wu & Levy, 1998). Thesemodels solve the transitive association problem by postu-lating that a function of the hippocampus is to constructcompact codes for stimuli that efficiently extract informa-tion about the relationships between stimuli (Gluck & My-ers, 1993; Gluck, Myers, & Meeter, 2005). TCM’s accountof transitive associations shares much with these models. InTCM transitive associations are a consequence of an overlapin the contextual representations tIN retrieved by items as aconsequence of learning. This similarity can be seen as arepresentation that expresses latent contextual relationshipsacross items—a point that will be elaborated in subsequent
work. The fact that TCM has been shown to account forthe Bunsey and Eichenbaum (1996) effect by impairing newcontextual learning (Howard et al., 2005) and also providesa quantitative fit to human verbal recall data from a similarparadigm (Figure 6) suggests that it could provide a usefulbridge between a neurophysiological description of the func-tion of the medial temporal lobe and human episodic memory(see also Gluck, Meeter, & Myers, 2003; Meeter, Myers, &Gluck, 2005).
Transitive association, contextual recovery and se-mantic spaces
The contextual retrieval approach to temporal contiguityin episodic memory association bears a striking similarityto several computational models of semantic memory (e.g.Landauer & Dumais, 1997; Griffiths & Steyvers, 2002; Jones& Mewhort, in press). Such models predict transitive asso-ciations as a natural consequence of the algorithms used toconstruct semantic representations. These models exploit thecontextual relationships among words in naturally-occurringtext to estimate the similarity of words. For instance, in la-tent semantic analysis (LSA, Landauer & Dumais, 1997),the input to the algorithm is a item-by-context co-occurrencematrix. In this setting, context is functionally defined as adocument of text in a larger corpus composed of many suchdocuments. The item-by-context matrix is subsequently sub-jected to a mathematical transformation called singular valuedecomposition that compresses the information contained inthe matrix.
In LSA, two words are judged to have similar meaningsby virtue of having occurred in the same context. This prop-erty is analogous to pairwise A− B associations. In addi-tion, as a result of the compression of the information in theitem-by-context matrix that results from singular value de-composition, words are also judged to have similar meaningsby virtue of having occurred in similar contexts, that is con-texts that contain similar words (see for instance, Landauer& Dumais, 1997, Figure A4). This latter property is closelyanalogous to the transitive A−C associations observed here.Other computational models of semantic memory, includingthe probabilistic topics model (Griffiths & Steyvers, 2002,2003), the hyperspace analogue of language (HAL, Lund& Burgess, 1996) and the BEAGLE model (Jones & Me-whort, in press) also naturally predict transitive associations.This striking similarity between computational models of se-mantic learning and contextual retrieval accounts of temporalcontiguity effects in episodic recall suggests a deep relation-ship between semantic and episodic memory.
It is striking that these computational models are able toextract semantic structure so effectively simply by observingthe temporal contexts in which words are presented. How-ever, in many of these models, the notion of what constitutesa context is provided as an input to the model—collections ofwords are tagged as members of a single document. More-over, temporal context in these models is often discrete—words either co-occur in a particular document or not. Oneof the central ideas of TCM is that temporal context changes

18 HOWARD ET AL.
gradually over time. It is intriguing to wonder what proper-ties a semantic space based on a continuously-changing tem-poral context in which words recover and recombine theircontextual states might exhibit.
Conclusions
We studied the associative structure induced by learningdouble-function lists of paired associates. We found evi-dence for associations between words that were never pre-sented together but that were presented in similar temporalcontexts by examining both intrusion probabilities and FFRtransition probabilities. Whereas associations between ad-jacent members of the linked-list were strongly asymmet-ric, we observed no asymmetry in the remote bridging as-sociations, This finding cannot be simply reconciled with aheteroassociative account of temporal contiguity effects inpaired associate learning. These transitive associations are,however, a perfectly natural consequence of accounts of con-tiguity effects in episodic memory that depend on contextualretrieval. TCM, a formal model of episodic association basedon contextual retrieval, provided an outstanding quantitativefit to the key data describing pairwise and transitive associa-tions. The analogy between contextual retrieval accounts oftemporal contiguity effects in episodic memory and compu-tational models of semantic memory suggests that perhapstransitive associations will prove to be critical in understand-ing the transition between episodic and semantic memory.
References
Asch, S. E., & Ebenholtz, S. M. (1962). The principle of associativesymmetry. Proceedings of the American Philosophical Society,106, 135-163.
Brown, G. D. A., Preece, T., & Hulme, C. (2000). Oscillator-basedmemory for serial order. Psychological Review, 107(1), 127-181.
Bunsey, M., & Eichenbaum, H. B. (1996). Conservation ofhippocampal memory function in rats and humans. Nature,379(6562), 255-257.
Burgess, N. (2002). The hippocampus, space, and viewpoints inepisodic memory. Quarterly Journal of Experimental Psychol-ogy, 55(4), 1057-80.
Burgess, N., & Hitch, G. (2005). Computational models of work-ing memory: putting long-term memory into context. Trends inCognitive Science, 9(11), 535-41.
Burgess, N., & Hitch, G. J. (1999). Memory for serial order: Anetwork model of the phonological loop and its timing. Psycho-logical Review, 106(3), 551-581.
Burgess, N., Maguire, E. A., & O’Keefe, J. (2002). The humanhippocampus and spatial and episodic memory. Neuron, 35(4),625-41.
Chappell, M., & Humphreys, M. (1994). An autoassociative neuralnetwork for sparse representations: Analysis and application tomodels of recognition and cued recall. Psychological Review,101, 103-128.
Clark, R. E., & Squire, L. R. (1998). Classical conditioning andbrain systems: the role of awareness. Science, 280(5360), 77-81.
Cole, R. P., Barnet, R. C., & Miller, R. R. (1995). Temporal encod-ing in trace conditioning. Animal Learning & Behavior, 23(2),144-153.
Davis, O., Rizzuto, D., Geller, A. S., & Kahana, M. J. (submitted).Temporal associative processes revealed by intrusions in paired-associate recall.
Dennis, S., & Humphreys, M. S. (2001). A context noise model ofepisodic word recognition. Psychological Review, 108(2), 452-78.
Dusek, J. A., & Eichenbaum, H. (1997). The hippocampus andmemory for orderly stimulus relations. Proceedings of the Na-tional Academy of Science, USA, 94(13), 7109-7114.
Eichenbaum, H. (2000). A cortical-hippocampal system for declar-ative memory. Nature Reviews, Neuroscience, 1(1), 41-50.
Eichenbaum, H. (2001). The hippocampus and declarative mem-ory: cognitive mechanisms and neural codes. Behavioural BrainResearch, 127(1-2), 199-207.
Estes, W. K. (1950). Toward a statistical theory of learning. Psy-chological Review, 57, 94-107.
Estes, W. K. (1955). Statistical theory of spontaneous recovery andregression. Psychological Review, 62, 145-154.
Frank, M. J., Rudy, J. W., Levy, W. B., & O’Reilly, R. C. (2005).When logic fails: implicit transitive inference in humans. Mem-ory & Cognition, 33(4), 742-50.
Frank, M. J., Rudy, J. W., & O’Reilly, R. C. (2003). Transitivity,flexibility, conjunctive representations, and the hippocampus. II.a computational analysis. Hippocampus, 13(3), 341-54.
Friendly, M., Franklin, P. E., Hoffman, D., & Rubin, D. C. (1982).The Toronto Word Pool: Norms for imagery, concreteness, or-thographic variables, and grammatical usage for 1,080 words.Behavior Research Methods and Instrumentation, 14, 375-399.
Gallistel, C. R., & Gibbon, J. (2000). Time, rate, and conditioning.Psychological Review, 107(2), 289-344.
Gillund, G., & Shiffrin, R. M. (1984). A retrieval model for bothrecognition and recall. Psychological Review, 91, 1-67.
Gluck, M. A., Meeter, M., & Myers, C. E. (2003). Computationalmodels of the hippocampal region: linking incremental learningand episodic memory. 7(6), 269-276.
Gluck, M. A., & Myers, C. E. (1993). Hippocampal mediation ofstimulus representation: a computational theory. Hippocampus,3(4), 491-516.
Gluck, M. A., Myers, C. E., & Meeter, M. (2005). Cortico-hippocampal interaction and adaptive stimulus representation: Aneurocomputational theory of associative learning and memory.Neural Networks, 18, 1265-1279.
Greene, A. J., Gross, W. L., Elsinger, C. L., & Rao, S. M. (2006).An FMRI analysis of the human hippocampus: inference, con-text, and task awareness. Journal of Cognitive Neuroscience,18(7), 1156-73.
Greene, A. J., Spellman, B. A., Dusek, J. A., Eichenbaum, H. B.,& Levy, W. B. (2001). Relational learning with and withoutawareness: transitive inference using nonverbal stimuli in hu-mans. Memory & Cognition, 29(6), 893-902.
Griffiths, T., & Steyvers, M. (2002). A probabilistic approach tosemantic representation. In W. Gray & C. Schunn (Eds.), Pro-ceedings of the 24th annual conference of the cognitive sciencesociety (p. 381-386). Austin, TX: Cognitive Science Society.
Griffiths, T., & Steyvers, M. (2003). Prediction and semantic as-sociation. Advances in Neural Information Processing Systems,15.

TRANSITIVE ASSOCIATIONS 19
Heckers, S., Zalesak, M., Weiss, A. P., Ditman, T., & Titone, D.(2004). Hippocampal activation during transitive inference inhumans. Hippocampus, 14(2), 153-62.
Henson, R. N. A., Norris, D. G., Page, M. P. A., & Baddeley, A. D.(1996). Unchained memory: Error patterns rule out chainingmodels of immediate serial recall. Quarterly Journal of Experi-mental Psychology, 49A, 80-115.
Howard, M. W., Addis, K. A., Jing, B., & Kahana, M. J. (In press).Semantic structure and episodic recall. In T. L. Landauer, D. S.McNamara, S. Dennis, & W. Kintsch (Eds.), LSA: A road to-wards meaning. Hillsdale, NJ: Laurence Erlbaum Associates.
Howard, M. W., Fotedar, M. S., Datey, A. V., & Hasselmo, M. E.(2005). The temporal context model in spatial navigation andrelational learning: Toward a common explanation of medialtemporal lobe function across domains. Psychological Review,112(1), 75-116.
Howard, M. W., & Jing, B. (2003). Mediated associations in pairedassociate learning. Poster presented at 44th annual meeting ofthe Psychonomic Society.
Howard, M. W., & Kahana, M. J. (1999). Contextual variabilityand serial position effects in free recall. Journal of ExperimentalPsychology : Learning, Memory, and Cognition, 25, 923-941.
Howard, M. W., & Kahana, M. J. (2002). A distributed representa-tion of temporal context. Journal of Mathematical Psychology,46(3), 269-299.
Howard, M. W., Kahana, M. J., & Wingfield, A. (2006). Agingand contextual binding: Modeling recency and lag-recency ef-fects with the temporal context model. Psychonomic Bulletin &Review, 13, 439-445.
Howard, M. W., Youker, T. E., & Venkatadass, V. (accepted pendingminor revisions). The persistence of memory: Contiguity effectsacross several minutes. Psychonomic Bulletin & Review.
Hull, C. L. (1947). The problem of primary stimulus generalization.Psychological Review, 54, 120-134.
Hulme, C., Stuart, G., Brown, G. D. A., & Morin, C. (2003). High-and low-frequency words are recalled equally well in alternatinglists: Evidence for associative effects in serial recall. Journal ofMemory and Language, 49(4), 500-518.
Humphreys, M. S., Bain, J. D., & Pike, R. (1989). Different ways tocue a coherent memory system: A theory for episodic, semantic,and procedural tasks. Psychological Review, 96, 208-233.
Jensen, O., & Lisman, J. E. (2005). Hippocampal sequence-encoding driven by a cortical multi-item working memorybuffer. Trends in Neuroscience, 28(2), 67-72.
Jones, M. N., & Mewhort, D. J. K. (in press). Representing wordmeaning and order information composite holographic lexicon.Psychological Review.
Kahana, M. J. (1996). Associative retrieval processes in free recall.Memory & Cognition, 24, 103-109.
Kahana, M. J. (2002). Associative symmetry and memory theory.Memory & Cognition., 30(6), 823-40.
Kahana, M. J., & Caplan, J. B. (2002). Associative asymmetry inprobed recall of serial lists. Memory & Cognition, 30(6), 841-9.
Kahana, M. J., Howard, M., & Polyn, S. (in press). Associative pro-cesses in episodic memory. In H. L. Roediger (Ed.), Learningand memory - a comprehensive reference. Elsevier.
Kahana, M. J., Howard, M. W., Zaromb, F., & Wingfield, A. (2002).Age dissociates recency and lag-recency effects in free recall.Journal of Experimental Psychology : Learning, Memory, andCognition, 28, 530-540.
Kahana, M. J., Rizzuto, D. S., & Schneider, A. R. (2005). Theoret-ical correlations and measured correlations: relating recognitionand recall in four distributed memory models. Journal of Exper-imental Psychology : Learning, Memory, and Cognition, 31(5),933-53.
Klein, K. A., Addis, K. M., & Kahana, M. J. (2005). A comparativeanalysis of serial and free recall. Memory & Cognition, 33(5),833-839.
Landauer, T. K., & Dumais, S. T. (1997). Solution to Plato’s prob-lem : The latent semantic analysis theory of acquisition, induc-tion, and representation of knowledge. Psychological Review,104, 211-240.
Lewandowsky, S., & Murdock, B. B. (1989). Memory for serialorder. Psychological Review, 96, 25-57.
Lund, K., & Burgess, C. (1996). Producing high-dimensional se-mantic spaces from lexical co-occurrence. Behavior ResearchMethods, Instruments & Computers, 28(2), 203-208.
Meeter, M., Myers, C. E., & Gluck, M. A. (2005). Integratingincremental learning and episodic memory models of the hip-pocampal region. 112(3), 560-85.
Mensink, G.-J. M., & Raaijmakers, J. G. W. (1988). A model forinterference and forgetting. Psychological Review, 95, 434-55.
Metcalfe, J., & Murdock, B. B. (1981). An encoding and retrievalmodel of single-trial free recall. Journal of Verbal Learning andVerbal Behavior, 20, 161-189.
Moses, S. N., Villate, C., & Ryan, J. D. (2006). An investigationof learning strategy supporting transitive inference performancein humans compared to other species. Neuropsychologia, 44(8),1370-87.
Murdock, B., Smith, D., & Bai, J. (2001). Judgments of frequencyand recency in a distributed memory model. Journal of Mathe-matical Psychology, 45(4), 564-602.
Murdock, B. B. (1966). Forward and backward associations inpaired associates. Journal of Experimental Psychology, 71, 732-737.
Murdock, B. B. (1982). A theory for the storage and retrievalof item and associative information. Psychological Review, 89,609-626.
Murdock, B. B. (1997). Context and mediators in a theory ofdistributed associative memory (TODAM2). Psychological Re-view, 1997, 839-862.
Nelder, J. A., & Mead, R. (1965). A simplex method for functionminimization. Computer Journal, 7, 308-313.
Norman, K. A., & O’Reilly, R. C. (2003). Modeling hippocam-pal and neocortical contributions to recognition memory: acomplementary-learning-systems approach. Psychological Re-view, 110(4), 611-46.
Ohta, A., Akiguchi, I., Seriu, N., Ohnishi, K., Yagi, H., Higuchi, K.,& Hosokawa, M. (2002). Deterioration in learning and memoryof inferential tasks for evaluation of transitivity and symmetry inaged samp8 mice. Hippocampus, 12(6), 803-10.
O’Keefe, J., & Dostrovsky, J. (1971). The hippocampus as a spa-tial map. preliminary evidence from unit activity in the freely-moving rat. Brain Research, 34, 171-175.
O’Reilly, R. C., & Rudy, J. W. (2001). Conjunctive representationsin learning and memory: principles of cortical and hippocampalfunction. Psychological Review, 108(2), 311-345.
Popper, J. (1959). Mediated generalization. In R. R. Bush &W. K. Estes (Eds.), Studies in mathematical learning theory (p.94-108). Stanford, CA: Stanford University Press.

20 HOWARD ET AL.
Primoff, E. (1938). Backward and forward associations as an orga-nizing act in serial and in paired-associate learning. Journal ofPsychology, 5, 375-395.
Provyn, J. P., Sliwinski, M. J., & Howard, M. W. (submitted).Effects of age on contextually mediated associations in pairedassociate learning.
Raaijmakers, J. G. W., & Shiffrin, R. M. (1980). SAM: A theoryof probabilistic search of associative memory. In G. H. Bower(Ed.), The psychology of learning and motivation: Advances inresearch and theory (Vol. 14, p. 207-262). New York: AcademicPress.
Raskin, E., & Cook, S. W. (1937). The strength and direction of as-sociations formed in the learning of nonsense syllables. Journalof Experimental Psychology, 20, 381-395.
Rizzuto, D. S., & Kahana, M. J. (2001). A neural network model ofpaired associate learning. Neural Computation, 13, 1835-1852.
Schwartz, G., Howard, M. W., Jing, B., & Kahana, M. J. (2005).Shadows of the past: Temporal retrieval effects in recognitionmemory. Psychological Science, 16(11), 898-904.
Sederberg, P. S., Howard, M. W., & Kahana, M. J. (in preparation).Short- and long-term recency in the temporal context model: Re-ply to Davelaar et al. (2005).
Sirotin, Y. B., Kimball, D. R., & Kahana, M. J. (2005). Goingbeyond a single list: Modeling the effects of prior experience onepisodic free recall. Psychonomic Bulletin & Review, 12, 787-805.
Slamecka, N. J. (1976). An analysis of double-function lists. Mem-ory & Cognition, 4, 581-585.
Small, S. A., Chawla, M. K., Buonocore, M., Rapp, P. R., & Barnes,C. A. (2004). Imaging correlates of brain function in monkeysand rats isolates a hippocampal subregion differentially vulner-able to aging. Proceedings of the National Academy of Science,USA, 101(18), 7181-6.
Smith, C., & Squire, L. R. (2005). Declarative memory, aware-ness, and transitive inference. Journal of Neuroscience, 25(44),10138-46.
Smith, D. M., & Mizumori, S. J. (2006). Hippocampal place cells,context, and episodic memory. Hippocampus, 16(9), 716-29.
Solomon, P. R., Vander Schaaf, E. R., Thompson, R. F., & Weisz,D. J. (1986). Hippocampus and trace conditioning of the rabbit’sclassically conditioned nictitating membrane response. Behav-ioral Neuroscience, 100(5), 729-744.
Titone, D., Ditman, T., Holzman, P. S., Eichenbaum, H., & Levy,D. L. (2004). Transitive inference in schizophrenia: impair-ments in relational memory organization. Schizophrenia Re-search, 68(2-3), 235-47.
Tulving, E. (1983). Elements of episodic memory. New York:Oxford.
Tulving, E. (2002). Episodic memory: from mind to brain. AnnualReview of Psychology, 53, 1-25.
Van Elzakker, M., O’Reilly, R. C., & Rudy, J. W. (2003). Transi-tivity, flexibility, conjunctive representations, and the hippocam-pus. I. an empirical analysis. Hippocampus, 13(3), 334-40.
Wasserman, E. A., DeVolder, C. L., & Coppage, D. J. (1992). Non-similarity-based conceptualization in pigeons via secondary ormediated generalization. Psychological Science, 3(6), 374-378.
Wilson, I. A., Gallagher, M., Eichenbaum, H., & Tanila, H. (2006).Neurocognitive aging: prior memories hinder new hippocampalencoding. Trends in Neuroscience, 29(12), 662-70.
Wilson, M. A., & McNaughton, B. L. (1993). Dynamics of thehippocampal ensemble code for space. Science, 261, 1055-8.
Wu, X. B., & Levy, W. B. (1998). A hippocampal-like neuralnetwork model solves the transitive inference problem. In J. M.Bower (Ed.), Computational neuroscience: Trends in research(p. 567-572). New York: Plenum Press.
Yonelinas, A. P. (2002). The nature of recollection and familiar-ity: A review of 30 years of research. Journal of Memory andLanguage, 46(3), 441-517.
Young, R. K. (1961). Paired-associate learning when the sameitems occur as stimuli and responses. Journal of ExperimentalPsychology, 61, 315-8.





























