Embed Size (px)
Citation preview

Respiratory pathophysiologic responses
Bronchial epithelial cells of patients with asthma release chemoattractant factors for T lymphocytes
Albert0 Bellini, MD, Hiroshi Yoshimura, PhD, Enza Vittori, PhD, Maurizio Marini, MD, and Sabrina Mattoli, MD, PhD Milan, Italy
Background: T &mphoqtes may orchestrate the inflammatory response in atopic asthma, but the mechanisms that promote T-cell accumulation in asthmatic aitways are still unclear. In this study we tested the hypothesis that bronchial epithelial cells of patients with atopic asthma release chemoattractant factors for T lymphocytes. Methods: Sixteen patients with atopic asthma and eight healthy control subjects were selected for this study. Bronchial epithelial cells were isolated from biopsy specimens obtained by means of bronchoscopy and cultured for 48 hours in serum- and hormone-free medium, with or without 10e6 mol/L histamine. Results: On&v the supematants of cells from donors with asthma showed chemotactic activity for T lymphocytes, and this was significantly increased (p < 0.025) by exposure to histamine. Chemotactic activity was in part mediated by interleuhin-8 (IL-S), because an antibody against human IL-8 significantly reduced it (p < 0.05) and the cell supematants contained appreciable amounts of immunoreactive IL-8 (0.89 Jo 0.39 nglml). Both the residual chemotactic activity of unstimulated epithelial cells and the increased activity caused by histamine were mediated by a single protease-sensitive substance with an apparent molecular weight of 56,000 d and an extimated isoelectric point of 8.8 to 9.1. The partial& pun&d chemoattractant specifical& enhanced the migration of CD4+ T lymphocytes, and its activity was inhibited by the univalent Fab fragment of a monoclonal antibody against CD4. Conclusion: These results extend our previous observations, indicating an impartant effector role of bronchial epithelium in asthma. (JALLERGY CLIN IMMUNOL 1993;92:412-24.)
Key wora% Airway epithelium, T lymphocytes, interkukin-8, lymphocyte chemoattractant factor, asthma
InGltration of activated eosinophils and lym- phocytes and increased responsiveness of the air- ways are characteristic features of chronic symp- tomatic asthma.“’ Activated eosinophils can re- lease a number of mediators that are directly involved in the pathogenesis of airway hyperre- sponsiveness,9M’1 and there is a correlation be-
From the Diagnostic Center for Respiratory and Allergic Diseases, Institute of Experimental Medicine, Milan; and the Department of Pathology and Experimental Biology, University of Milan.
Supported by Grants from the National Research Council, the Italian Association for Cancer Research, and Casse di Risparmio delle Provincie Lombarde, Italy.
Received for publication Oct. 9, 1992; revised Feb. 17, 1993; accepted for publication Feb. 18, 1993.
Reprint requests: S. Mattoli, MD, PhD, Diagnostic Center for Respiratory and Allergic Diseases, P. Box 6, Via Alessan- dria 4, 20144 Milan, Italy.
Copyright 0 1993 by Mosby-Year Book, Inc. 0091-6749l93 $1.00 + .lO UU46W
412
tween the extent of eosinophilic inflammation in the airways and the clinical severity of asthma.l’
In atopic asthma, T lymphocytes may contrib- ute to airway hyperresponsiveness by promoting accumulation and local activation of eosinophils. This is because a particular subset of T helper lymphocytes (T,,-type) predominates in the asth- matic airways, and these cells express a limited number of cytokines that alter the motility, viabil- ity, and function of eosinophils in vitro.‘, 8, 13* l4 It is possible that CD4+ T lymphocytes are selec- tively recruited to the bronchial mucosa of pa- tients with atopic asthma and that the level of interaction between bronchial dendritic cells and antigenic stimuli can then favor the expansion of the T,-type cells under the regulation of factors released locally at the tissue site.” After their recruitment and activation, CD4+ T lymphocytes of the T,, subtype would produce interleukin-3 (IL-3), IL-5, and granulocyte-macrophage colony-

J ALLERGY CLIN IMMUNOL VOLUME 92, NUMEER 3
Abbreviahons used EC-CF: Chemotactic factor partially purified
from supematants of epithelial cells of asthmatic patients by size exclu- sion chromatography and Sephadex QAE anion exchange chroma- tography
EC-CM: Epithelial cell-conditioned medium (crude or concentrated by lyophiliza- tion after dialysis)
FEV,: Forced expiratory volume in 1 second
IL: Interleukin LCF: Lymphocyte chemoattractant factor PBS: Phosphate-buffered saline
PC20M: Concentration of inhaled methacho- line causing a 20% fall in FEV,
stimulating factor, and these lymphokines would promote eosinophilopoiesis and eosinophil migra- tion, prolong eosinophil survival, and enhance eosinophil functioning.‘5-20
The mechanisms by which CD4+ T lympho- cytes are selectively recruited to the bronchial mucosal in atopic asthma are still unclear.
In this report, we present data demonstrating that bronchial epithelial cells of patients with atopic asthma produce two lymphocyte chemotac- tic factors. One of these substances is IL-8. The second factor is distinct from IL-8, IL-l, and IL-2 and selectively alters the motility of CD4+ T lymphocytes. The release of this latter substance is upregulated by histamine. Thus the lymphocyte chemotactic activity of epithelial cells may repre- sent one of the factors involved in the compart- mentalization of helper T cells in the airways of patients with atopic asthma.
METHODS Subjects
Sixteen subjects with mild asthma (eight women) with a mean age of 29.9 years (range, 19 to 44 years) and eight healthy control subjects (three women) with a mean age of 27.7 years (range, 20 to 37 years) partici- pated in the study (Table I). All of the subjects were nonsmoke IS.
The subjects with asthma were atopic, with a skin wheal response of greater than 3 mm in diameter to one or m’ore of 12 common allergen extracts.21 The subjects who had pollen-related asthma were studied out of the hay fever season. Their asthma was con- trolled with inhaled &-agonists alone on an as-required basis, and none of them had experienced a respiratory infection or an acute exacerbation of asthma within 4
Bellini et al. 413
weeks of the day on which bronchoscopy was per- formed. The subjects with asthma had a mean (-+ SD) baseline forced expiratory volume in 1 second (FEV,) of 82.9% -t 9.4% of the predicted value, and all had hyperreactive airways to inhaled methacholine, with a geometric mean provocative concentration of inhaled agonist reducing FEV, by 20% of 0.63 mg/ml (range, 0.12 to 4.57 mg/ml) (Table I).
The healthy volunteers were all nonatopic and had normal baseline values of FEV, (103.3% * 8.1% of predicted value) and PC20M values greater than 32 mg/ml (Table I).
The study was approved by the institutional review boards, and informed consent was given by each subject.
Prebronchoscopy evaluation
All subjects underwent bronchial reactivity testing 4 days before bronchoscopy and in absence of any treat- ment for 24 hours. Spirometric measurements and a bronchoprovocation test with methacholine were per- formed according to a standard protocol?* Physiologic saline and doubling concentrations of methacholine ranging from 0.03 to 32 mg/ml were sequentially in- haled by means of a Wright nebulizer (Ferraris Medi- cal, Inc., Holland, N.Y.). FEV, was measured at 1 and 3 minutes after the inhalation with a Vitalograph spirometer (Vitalograph, Inc., Lenexa, Kan.). The best of three readings was recorded at each time point, and the lowest of the two values was plotted on a log concentration-FEV, graph. The inhalations were con- tinued until FEV, had fallen by at least 20% from the value after saline administration or until the highest concentration of methacholine had been administered. The noncumulative concentration of methacholine causing a 20% fall in FEV, (PC20M) was calculated by linear interpolation of the last two points on the log concentration-response curve if a 20% fall in FEV, had been achieved. Otherwise, it was recorded as being greater than 32 mglml.
Bronchoscopy
Subjects attended the laboratory after withholding their medications for 24 hours before the procedure. Fiberoptic bronchoscopy was undertaken according to our protoc01,23~ *4 which is based on international guide- lines.=
In all of the tested subjects, fiberoptic bronchoscopic examination was performed afer premeditation with atropine and diazepam, salbutamol administered through a nebulizer, and local anesthesia with 2% lidocaine. Ten endobronchial biopsy samples were ob- tained through the bronchoswpe in randomly selected lobar and segmental bronchi of the two lungs, with the use of Olympus FB-1SC model forceps (Olympus Corp., Lake Success, N.Y.), which allowed the operator to perform limited stripping of bronchial epithelium and caused minimal bleeding.

414 Bellini et al. J ALLERGY CLIN IMMUNOL SEPTEMBER 1993
TABLE I. Subject characteristics
Subject No.
Asthma 1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16
Normal 1 2 3 4 5 6 7 8
Age W)
31 42 26 35 32 21 19 39 25 28 33 44 27 36 21 20
24 26 20 28 31 22 37 34
Gender
F M F M M M F F M M F M F M F F
M F M M F F M M
FEV, PCZOM Atow (% pred) bw/mll (prick test)
93 0.68 GP 70 0.15 GP 72 0.21 GP 83 2.13 DP 95 4.57 DP 76 1.05 DP, GP 71 0.38 DP 75 0.52 GP, DP 85 0.93 GP 88 0.71 C 97 3.32 GP, DP 84 1.18 C 97 0.55 GP 72 0.12 DP 85 0.25 DP 83 0.34 DP
96 >32 - 115 >32 - 108 ~32 - 97 >3?+! -
104 >32 - 92 >32 -
112 ~32 - 103 >32 -
Pred, Predicted value; GP, grass pollen; DP, Dermtophagoiaks pteronyssinus; C, cat.
After this procedure, patients with asthma again received salbutamol through a nebulizer and 25 mg of prednisone, administered orally.
Sample processing The biopsy samples were processed for isolation of
epithelial cells, according io a standard procedure pre- viously described in detail.“, 24 Briefly, after enzymatic digestion of the tissue fragments, fibroblasts and mono- cytes were removed by adherence to a plastic surface for 2 hours. Residual monocyteslmacrophages, granu- locytes, lymphocytes, and dendritic cells were sequen- tially eliminated by treatment with the antibodies against the surface markers of these cells’, Z* 24 and appropriately diluted rabbit complement. The total numbers of cells that remained after depletion were 12.6 ? 2.2 x lo6 per donor, and these cells were more than 92% viable, as assessed by exclusion of trypan blue dye. The purity of epithelial cells was greater than 98.8%, as determined by immunospecific labeling with the anti-cytokeratin antibody DAKO-CKl (Dakopatts, Glostrup, Denmark) with 0% of the depleted cell populations present.
Isolation of T lymphocytes and subsets Human peripheral blood mononuclear cells were
isolated from heparinized venous blood of healthy volunteers by density centrifugation or Ficoll-Paque (Pharmacia, Uppsala, Sweden). Cells were recovered from the interface. Residual monocytes were elimi- nated by panning with the anti-CD14 monoclonal anti- body (Ortho Diagnostic Systems, .Milan, Italy). Result- ing cells were then washed and separated by density centrifugation on a discontinuous Percoll (Pharmacia) gradient to obtain resting T cells that were depleted of large granular lymphocytes and spontaneously prolifer- ating lymphocytes.z6 Finally, residual HLA-DR-positive cells and B lymphocytes were eliminated by treatment with the anti-HLA Ia and Tee pan B monoclonal antibodies (Technogenetics, Turin, Italy) and appropri- ately diluted rabbit complement in a standard two-step cytotoxicity assay. This protocol resulted in T lympho- cytes that were more than 98% pure, as assessed by immunospecific labeling with the OKT3 monoclonal antibody (Ortho Diagnostic Systems), and the cells were more than 95% viable according to the trypan blue exclusion test.
J ALLERGY CLIN IMMUNOL VOLUME 92, NUMBER 3
CD4+ and CD8+ lymphocyte populations were enriched with the use of magnetic beads attached to an anti-mouse :IgG antibody (Advanced Magnetics, Bos- ton, Mass.). Cells were first labeled with either anti- CD4 or anti-CD8 mouse monoclonal antibodies by incubating 1 x 10’ lymphocytes with 1 pg of the appro- priate antibody for 30 minutes at 4” C. The labeled cells were then washed and incubated with the magnetic beads coupled to the anti-mouse IgG for 30 minutes at 37” C. The bead-cell complex was then extracted from the unlabeled cells in a magnetic field. The purity of the T-lymphocyte subsets was greater than 92.8%, as de- fined by immunospecific labeling with either OKT4 or OKT8 monoclonal antibodies (Ortho Diagnostic Systems).
Epithelial cell cultures The bronchial epithelial cells isolated from each
donor were finally resuspended in Iscove’s modified Dulbecco’s medium supplemented with streptomycin (100 pg/ml) and penicillin (100 U/ml). This medium did not contain fetal calf serum or substances that can induce cell growth or activation. Epithelial cells can be cultured in this medium for 5 days without significant changes in their characteristics and viability.24 Epithe- lial cells were plated in collagen-coated 24-well culture plates at a density of 5 x l@ cells per well and incu- bated for 48 hours at 37” C, in 5% CO, and 100% humidity.
In some experiments epithelial cells were incubated for 48 hou:rs in the presence or absence of histamine dihydrochloride, 10m6 mol/L (Sigma Chemical Co., St. Louis, MO.). The effective concentration of this com- pound was determined in preliminary experiments.
All culture reagents were purchased from Gibco Europe (Paiseley, Scotland) and were free of endotoxin contamination as assessed by limulus test.
Processing of epithelial cell supernatants At the end of the period of incubation, cell-free
supematants were harvested, centrifuged at 800 g for 10 minutes, stored in 100 ~1 aliquots, and frozen at - 70” C. After thawing, some aliquots from each sample were used for IL-8 assay, as reported below. The other sample fractions were centrifuged at 25,000 g for 30 minutes, dialyzed at 4” C against phosphate-buffered saline (PBS), pH 7.4, and concentrated to one twenti- eth of the original volume by lyophilization. These samples of concentrated epithelial cell-conditioned me- dium (EC,-CM) were tested (1: 10 dilution) for migra- tion enhancement activity with T lymphocytes as the target cell population.
In further experiments, the EC-CM from donors with asthma was pooled and applied to a Sephadex G-100 column (Pharmacia). Chromatography was per- formed at 4” C, and elution was done with PBS at 15 ml/hour. Two-milliliter fractions of the eluted material
Bellini et al. 415
NORMAL ASTHMA
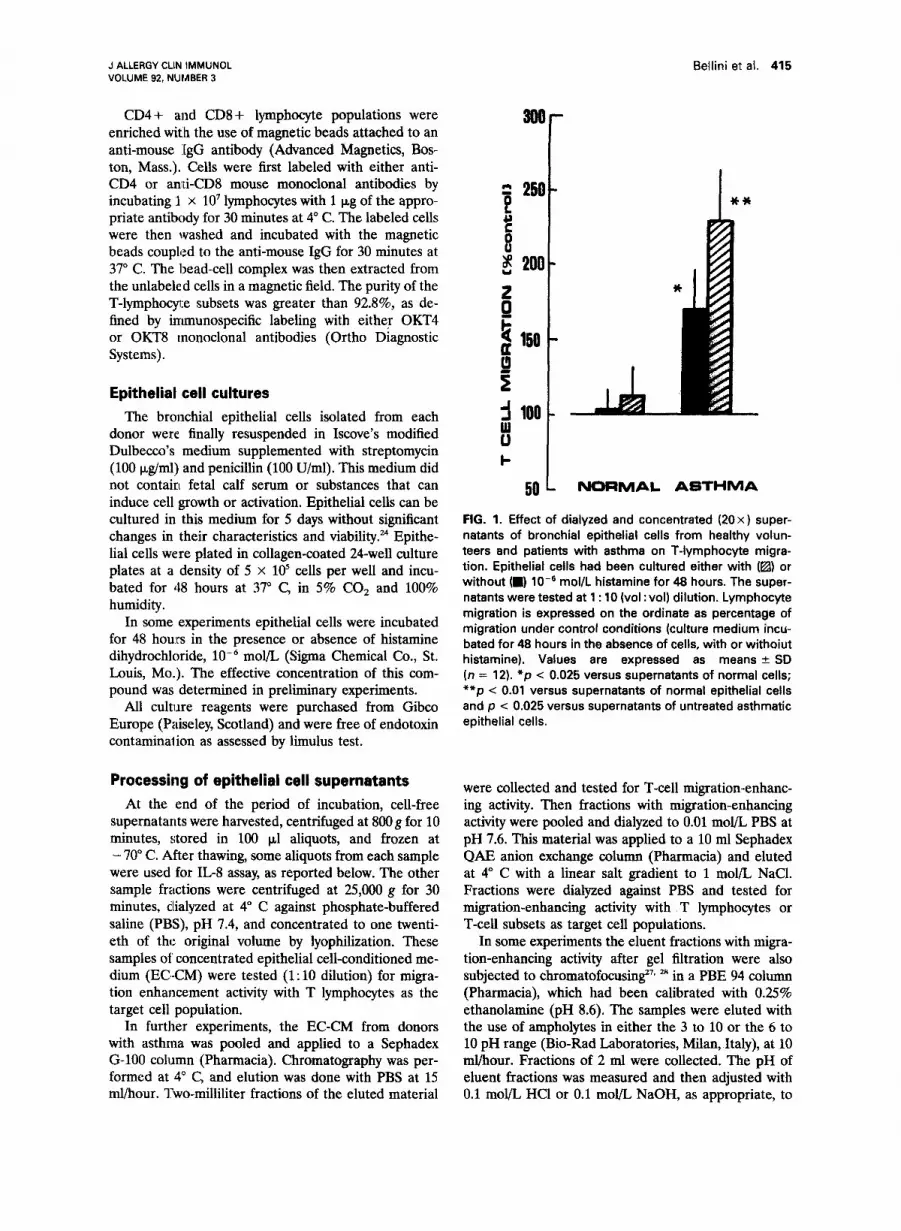
FIG. 1. Effect of dialyzed and concentrated (20x) super- natants of bronchial epithelial cells from healthy volun- teers and patients with asthma on T-lymphocyte migra- tion. Epithelial cells had been cultured either with (@) or without (m) lo-’ mol/L histamine for 48 hours. The super- natants were tested at 1 : 10 (vol : vol) dilution. Lymphocyte migration is expressed on the ordinate as percentage of migration under control conditions (culture medium incu- bated for 48 hours in the absence of cells, with or withoiut histamine). Values are expressed as means + SD (n = 12). *p < 0.025 versus supernatants of normal cells; **p < 0.01 versus supernatants of normal epithelial cells and p < 0.025 versus supernatants of untreated asthmatic epithelial cells.
were collected and tested for T-cell migration-enhanc- ing activity. Then fractions with migration-enhancing activity were pooled and dialyzed to 0.01 mol/L PBS at pH 7.6. This material was applied to a 10 ml Sephadex QAE anion exchange column (Pharmacia) and eluted at 4” C with a linear salt gradient to 1 mol/L NaCl. Fractions were dialyzed against PBS and tested for migration-enhancing activity with T lymphocytes or T-cell subsets as target cell populations.
In some experiments the eluent fractions with migra- tion-enhancing activity after gel filtration were also subjected to chromatofocusingz7 28 in a PBE 94 column (Pharmacia), which had been calibrated with 0.25% ethanolamine (pH 8.6). The samples were eluted with the use of ampholytes in either the 3 to 10 or the 6 to 10 pH range (Bio-Rad Laboratories, Milan, Italy), at 10 ml/hour. Fractions of 2 ml were collected. The pH of eluent fractions was measured and then adjusted with 0.1 mol/L HCl or 0.1 mol/L NaOH, as appropriate, to

416 Bellini et al. J ALLERGY CLIN IMMUNOL SEPTEMBER 1993
TABLE II. Checkerboard analysis of T-cell migration-enhancing activity of supernatants of histamine-exposed epithelial cells from patients with asthma
Above the filter
Supematant dilutions (vol:vol)
Below the filter Control medium 1:loo 1:lSO 1:lO
Control medium 14 It 2 18 2 3 15 + 2 12 t 3 Supematant dilutions (~01: vol)
1:lOO 25 k 2 22 + 5 13 + 4 14 + 4 1:50 31 + 4* 28 + 3t 17 2 3 13 2 2 1:lO 49 k 8* 39 2 5* 28 f 7t 16 2 8
Bronchial epithelial cells of patients with asthma were exposed to low6 mol/L histamine for 48 hours and concentrated (20x) supematants were tested for T-cell chemotactic activity. Different dilutions of these supematants were seeded in the upper and/or lower compartment of the chemotaxis chamber. Control medium was the culture medium additioned with 10e6 mol/L histamine, incubated for 48 hours in the absence of the cells, and processed in parallel with the experimental samples. Results were mean numbers of migrated T lymphocytes per high-power field with two replicates. Values are means f SD from eight experiments.
*p < 0.01 versus control medium (above and below the filter). tp -C 0.025 versus control medium.
pH 7.4. Each eluate was tested for migration-enhancing activity.
To test sensitivity of the partially purified chemotac- tic factor (EC-CF) to proteolytic agents, the sample fractions with migration-enhancing activity were pooled, treated with 100 &ml pepsin (Sigma Chemical Co.) in 0.1 N HCl for 3 hours at 37” C, and then retested for chemotactic activity after neutralization with 50 mmol/L N-2-hydroxyethylpiperazine-N-2- ethanesulfonic acid, pH 7.4. To test sensitivity to heat- ing, the sample fractions with migration-enhancing ac- tivity were heated to 60” C for 30 minutes before migration experiments.
Assay of lymphocyte migration
Migration was assessed with standard blind-well chemotaxis chambers.29 To assess the effect of a given experimental sample on lymphocyte migration, 225 pl of the tested sample or the appropriate control medium was placed in duplicate bottom wells. An 8 km pore size polyvinylpyrrolidone-treated polycarbonate filter (Nucleopore, Milan, Italy) was positioned in each chamber assembly, and 333 ~1 of a lymphocyte suspen- sion (1 x 106 cells/ml in medium 199 supplemented with 25 mmol/L N-2-hydroxyethylpiperazine-N-2- ethanesulfonic acid buffer, 100 U/ml penicillin, 100 &ml streptomycin, and 0.4% bovine serum albumin) was seeded in each of the top wells. Appropriate controls were samples of culture medium of bronchial epithelial cells incubated for 48 hours in the absence of the cells, with or without the addition of 10e6 mol/L histamine, and processed in parallel with the experi- mental samples.
Chemotaxis chamber assemblies were incubated at 37” C in humidified air and 5% COZ for 3 hours. The filters were removed, fixed, and stained with Diff-Quick
(Baxter Healthcare Corp., Scientific Div., McGaw Park, Ill.). T cells that had migrated through the bottom of each filter were counted in 5 high-power fields (1000 x ), and the mean cell number per high-power field was calculated. In all experiments chemotactic activity was tested in duplicate and expressed as the percent of control.
Neutralization experiments
In these experiments we evaluated the possibility that the presence of cytokines with known chemotactic activity for T lymphocytes, particularly IL-l,M IL-2,31 and IL-8,32 could account for any biologic activity of the supematants of asthmatic bronchial epithelial cells. To do that, the EC-CMs were incubated with saturating concentrations of monospecific antisera against IL-l, IL-2, and IL-8 or with preimmune sera as controls for 30 minutes at 37” C and then reassayed for chemotactic activity. To neutralize the activity that is potentially due to IL-l and IL-2, we used three commercially available antibodies (polyclonal rabbit anti-human IL-la, 1: 10 dilution; polyclonal rabbit anti-human IL-lp, 1: 10 dilution; polyclonal rabbit anti-human IL-2, 1: 20 dilution; all from Genzyme Co., Boston, Mass.); the specificity and neutralizing properties of these antibod- ies had been tested in previous studies.6* 26 The polyclo- nal rabbit anti-human IL-8 antiserum was obtained as previously described in detail.*9 It reacts with recombi- nant human IL-8 in Western blot analysis and fails to cross-react with connective tissue-activating protein III or platelet factor 4. When used at a dilution of 1:50, as in this study, it neutralizes the biologic activity of 10 rig/ml recombinant human IG8,29 and this IL-8 concen- tration is 10 times higher than that required to induce maximal T-cell chemotactic activity.3”
Because another identified and recently cloned hu-

J ALLERGY CLIN IMMUNOL VOLUME 92, NUMBER 3
Bellini et al. 417
5OL UNTREATED HlETAMlNE ti66Ml
FIG. 2. Neutralizing effects of the antibodies against IL-la, IL-18 IL-2, and IL-8 on the chemotactic activity of dialyzed and concentrated (20 x) supernatants of bronchial epithelial cells from donors with asthma cultured with or without 10m6 mol/L histamine, as indicated. Supernatants were tested at 1 : 10 (vol:vol) dilution after preincubation with saturating concentrations of each antibody for 30 minutes at 37” C. Control samples were preincubated with rabbit preimmune serum (control serum) for the same period of time. T-lymphocyte migration is expressed on the ordinate as percentage of migration under control conditions (culture medium +- histamine). Values are expressed as means k SD (n = 12). l p < 0.05 versus supernatants preincubated for 30 minutes in absence of any antisera (non&
man T-lymphocyte chemoattractant cytokine possesses specific activity for CDCpositive T lymphocytes and because CD4 serves as the cellular receptor for this substance,3’“5 we next evaluated the possibility that the release of this cytokine could also account for any biologic activity of EC-CM. Thus we tested the blocking effect of the Fab fragment of an anti-CD4 monoclonal antibody (OKT4) by placing the partially purified chemoattractant and the antibody Fab fragments to- gether in the bottom wells of the chemotaxis chambers. We used the Fab fragments instead of the divalent antibody because the latter is known to induce specific locomotory responses in CD4+ T lymphocytes.M The Fab fragment of a monoclonal antibody against CD3 (OKT3, Ortho Diagnostic Systems) was used as a control.
The Fab’ fragments were obtained by a modification of a previously described method.36 The OKT4 and OKT3 mo:noclonal antibodies (50 p,g) were digested with 2.5 U papain (Sigma Chemical Co.) in 0.1 mol/L PBS that contained 10 mmol/L 2-mercaptoethanol, 4 mmol/L ethylenediaminetetraacetic acid, and 1 mg/ml sodium azide, pH 7.4, at 37” C, for 12 hours. Intact IgG and Fc fragments were eliminated by repeated incuba- tions with Protein A-Sepharose (Sigma Chemical Co.),
and the resulting material was dialyzed to PBS. The purity of the fragments was then assessed by sodium dodecylsulfate-polyacrylamide gel electrophoresis. The concentrations of Fab fragments were determined by the Bio-Rad protein microassay (Bio-Rad Laborato- ries), and these were used at the concentration of 0.8 p&ml, according to the results of preliminary experi- ments.
IL-8 assay To confirm the results of neutralization experiments
with the antibody against IL-8, the contents of immu- noreactive IL8 in crude supernatants of untreated and histamine-exposed epithelial cells were assayed in du- plicate by ELISA. We used a double-ligand method, as previously described,29 with a rabbit anti-IL-8 antiserum as the capturing antibody and a biotinylated goat anti- IL8 antibody as the revealing antibody. In this assay recombinant human IL-8 (Genzyme Co.) was used as standard at 1:2 log dilutions. Fresh culture medium was used to dilute the standard IL-8 and as the blank. The assay consistently detected IL-8 concentrations greater than 0.35 q/ml in a linear fashion, with an inter- and intraassay variability of 12.5% and 6%, respectively.

418 Bellini et al. J ALLERGY CLIN IMMUNOL SEPTEMBER 1993
TABLE III. Assay of immunoreactive IL-B in crude supernatants of bronchial epithelial cells from normal and asthmatic subjects
IL-8 (rig/ml/5 x l@ cells)
Constitutive Histamine-induced Subject No. release release
Asthma 1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16
-c 0.35 < 0.35 1.15 1.21 0.93 0.87
< 0.35 < 0.35 < 0.35 < 0.35
0.80 0.35 0.56 0.72 1.32 1.23 0.76 0.41 0.44 0.62
< 0.35 c 0.35 < 0.35 < 0.35
0.44 0.38 1.62 1.75 1.21 1.50 0.58 0.92
< 0.35 < 0.35 < 0.35 < 0.35 < 0.35 < 0.35 < 0.35 c 0.35
c 0.35 < 0.35 < 0.35 < 0.35 < 0.35 c 0.35 < 0.35 c 0.35
Comparison of the chemotactic activities of partially purified EC-CF and recombinant human IL 1fI
To compare the activity of the partially purified EC-CF to that of an agent known to augment the migratory response of T cells, the chemotactic factor, which was obtained from cultures of asthmatic epithe- lial cells exposed to histamine for 48 hours and partially purified by gel filtration and anion exchange chroma- tography, was tested for migration-enhancing activity together with different concentrations of recombinant human IL-lp (Genzyme).%
Statistical analysis All values were expressed as means ? SD, with the
exception of methacholine responsiveness for the groups, which was expressed as the geometric mean PC20M.
Nonparametric tests were used to analyze results concerning the chemotactic activity. The Mann-Whit-
ney U test was used to compare data between the control and asthma groups, and Wilcoxon’s matched signed-rank test was applied for within-group analysis. A p value of less than 0.05 was taken as the level of significance.
RESULTS T-cell chemotactic activity of EC-CM and effect of exposure to histamine
The supernatants of bronchial epithelial cells from the healthy control subjects did not signifi- cantly alter T-lymphocyte migration (Fig. 1). Ex- posure to histamine, low6 mol/L elicited some migration-enhancing activity in these cells, but this effect did not reach statistical significance. By contrast, the supematants of epithelial cells from the donors with asthma enhanced lymphocyte migration to 165% f 26% of control value, and those dervied from the same cells stimulated with histamine-enhanced T-cell migration to 233% r 32% of control value (Fig. 1).
In subsequent experiments we determined whether the supematants of asthmatic bronchial epithelial cells elicited a gradient-dependent lo- comotory response in T cells (true chemotaxis) or an augmentation of gradient-independent motility (chemokinesis). To evaluate this response, we performed checkerboard experiments with the supematants obtained from epithelial cells of pa- tients with asthma who were exposed to histamine for 48 hours. Culture medium, supplemented with 10d6 mol/L histamine and incubated for the same period of time in the absence of cells, served as the control. As shown in Table II, maximal induc- tion of lymphocyte migration across the filters occurred in the presence of a positive concentra- tion gradient between lower and upper compart- ments of the chemotaxis chamber. In the presence of a negative gradient (higher concentrations in the top well), no enhanced migration of T cells occurred. With equal concentrations of supema- tants above and below the filter, little or no enhancement of migration was observed.
Neutralization experiments and IL-8 assay To determine whether the T-cell chemotactic
activity of asthmatic bronchial epithelial cells was due to the release of one or more cytokines known to alter the migration of T lymphocytes, some of the experiments reported above were repeated after incubation of EC-CM with saturat- ing concentrations of the antibodies against hu- man IL-la, IL-@, IL-2, and IL-8. As shown in Fig. 2, only the antibody against IL-8 partially

J ALLERGY CLIIU IMMUNOL VOLUME 92, NlJMBER 3
Bellini et al. 419
BSA OA
I I
Chym 1
*,I. 30 40 60 60 70 80 90 100
% BED VOLUME
FIG. 3. Size exclusion chromatography of concentrated (20x) and pooled supernatants from bronchial epithelial cells of patients with asthma, cultured for 48 hours in the presence (solid he) or absence (dashed line) of IO-’ mol/L histamine. IL-8 was removed before chromatography by immunoprecipitation. T-lymphocyte migratory responses are shown on the ordinate as percent- age of migration under control conditions (culture medium incubated for 48 hours in absence of cells, with or without histamine, and processed in parallel with the experimental samples). Molecular weight standards are indicated: Blue dextran (SDj, 2 x 10’ d; bovine serum albumin (BSA), 68,000 d; ovalbumin @A), 43,000 d; chymotrvpsinogen (Chym), 25,000 d.
reduced the T-cell chemotactic activity constitu- tively released from the bronchial epithelial cells of patients with asthma (p c 0.05). This antise- rum did not neutralize the increase in chemotac- tic activity produced by the same cells on expo- sure to h:istamine. Next, we assayed crude super- natants of bronchial epithelial cells of asthmatic and normal donors for the presence of IL-8 im- munoreactivity. The samples derived from the epithelial cells of 11 subjects with asthma con- tained appreciable amounts of immunoreactive IL-8 (0.89 + 0.39 r&ml/5 x 105 cells), but hista- mine did not increase IL-8 release from these cells (0.90 2 0.47 rig/ml/S x lo” cells,p > 0.05 vs constitutive release) (Table III). No significant IL-8 immunoreactivity was detected in superna-
tants from untreated or histamine-stimulated nor- mal epithelial cells.
Thus IL-8 was one of the factors with T-cell chemotactic activity produced by asthmatic bron- chial epithelial cells. Residual activity of un- treated cells and activity upregulated by histamine were due to one or more factors distinct from IL-l, IL-2, and IL-8.
Partial purification of the T-cell chemotactic activity distinct from IL-8
The concentrated supematants of epithelial cells obtained from the patients with asthma and cultured for 48 hours in the presence or absence of histamine were depleted of IL-8 by immuno- precipitation, pooled, and subjected to Sephadex

420 Bellini et al. J ALLERGY CLIN IMMUNOL SEPTEMBER 1993
TABLE IV. Target cell specificity of the chemotactic factor partially purified from supernatants of untreated and histamine-exposed bronchial epithelial cells from patients with asthma
Chemoattractant
No histamine Histamine (lo-’ mol/L)
Target cells Control EC-CF Control EC-CF
CD3 + T cells 12.5 + 2.5 22.6 k 4.8 17.3 _+ 3.8 38.5 k 5.9 p c 0.025 p c 0.01
CD4 + T cells 14.6 + 3.9 24.4 f 5.9 16.9 + 4.2 33.9 + 3.7 p < 0.025 p < 0.01
CD8+ T cells 13.5 + 2.2 14.8 k 3.7 12.4 +- 2.1 12.5 + 4.8 p > 0.05 p > 0.05
EC-CF was cultured for 48 hours in the presence or absence of 10m6 mol/L histamine. Controls were culture media of epithelial cells incubated for 48 hours in absence of cells, with or without histamine, and processed in parallel with the supernatants. Lymphocyte migration was expressed as the mean numbers of cells migrating per high-power field in two duplicate filters. valies are mea& 2 SD from six experiments.
G-100 gel filtration. As shown in Fig. 3, the samples derived from untreated or histamine- exposed cells produced two identical areas of lymphocyte migration-enhancing activity that were eluted after bovine serum albumin and be- fore ovalbumin and that peaked at approximately 56,000 d.
For each experimental condition, the fractions of eluted material that had demonstrated T-cell chemotactic activity were pooled and subjected to Sephadex QAE anion exchange column chroma- tography. Then, this material was tested for its ability to alter the migration of T-lymphocyte subsets (CD4 + or CD8 + T cells). The results are shown in Table IV. The partially purified T-cell chemotactic activities, derived from untreated or histamine-exposed asthmatic epithelial cells, stim- ulated the migration of CD4+ lymphocytes (167% and 222% of controls, respectively) but not that of CD8 + T lymphocytes (109% and 101% of controls).
Treatment of these EC-CFs with a proteolytic enzyme greatly reduced their T-cell migration- enhancing activity (p < O.Ol), but the EC-CFs were not inactivated by heating to 60” for 30 minutes (Fig. 4).
The Fab fragment of the anti-CD4 antibody completely inhibited the response of CD4+ T cells to the EC-CFs @ < O.Ol), whereas the OKT3 Fab fragment, which binds to another cell surface antigen of CD4+ T cells, was ineffective (Fig. 5).
In some experiments the active fractions of supernatants from asthmatic epithelial cells ex- posed to histamine (four donors), which had been
obtained after gel filtration, were chromatofo- cused on a PBE column with a pH gradient of 10 to 6. Migration-enhancing activity for CD4+ T lymphocytes ranged between pH 8.8 and 9.1 (Fig. 6).
All of these findings indicate that: (1) the T-cell chemotactic activities, which are expressed by bronchial epithelial cells of patients with asthma constitutively and on histamine exposure and are distinct from IL-8, were due to the production of a single cationic protein with a molecular weight of 56,000 d; ‘(2) this factor specifically enhanced the locomotory response of CD4 + T lymphocytes through some direct or indirect interaction with the CD4 molecule expressed on their cell sur- faces.
Comparison of the migration-enhancing activity of EC-CF and IL-lfi
In these experiments the partially purified EC- CF obtained from cultures of asthmatic epithelial cells exposed to histamine (four donors) was tested for T-cell migration-enhancing activity to- gether with different concentrations of recomi- nant human IGlB. The potency of partially puri- fied EC-CF was similar to that of 1 U/ml IL-18 (Fig. 7).
DISCUSSION
In this study we demonstrated that bronchial epithelial cells of patients with atopic asthma released factors that enhanced the migration of T lymphocytes, whereas the cells from healthy sub- jects were unable to produce substances with similar activity.

J ALLERGY CLIN IMMUNOL VOLUME 92, NUMBER 3
Bellini et al. 421
NONE PEPEIN HeAT
FIG. 4. Inhibitoryeffects of proteases and heating on the biologic activity of the chemotactic factor partially purified from the supernatants of bronchial epithelial cells of pa- tients with asthma. These cells had been cultured for 48 hours in the presence (a) or absence (m) of lo-’ mol/L histamine. The concentrated (20 x) supernatants obtained under each experimental condition from each donor were pooled and subjected to Sephadex G-100 and Sephadex GAE anion exchange chromatography after removal of IL-8 by immunoprecipitation. The fractions of the resulting material that showed chemotactic activity for CD4+ T lymphocytes were pooled and retested after preincuba- tion with 100 kg/ml pepsin in 0.1 N HCI for 3 hours or heating to E’O“ C for 30 minutes. CD4+ T-cell migration is expressed on the ordinate as percentage of migration observed under control conditions (culture medium incu- bated for 48 hours in absence of cells, with or without histamine, .snd processed in parallel with the superna- tants). Values are expressed as means f SD. *p < 0.05 versus untreated chemotactic factor (none).
An II.,&like substance mediated in part the chemotactic activity of asthmatic bronchial epi- thelial cells, as demonstrated by the ability of a monospecific antiserum against IL-8 to reduce it and by the presence of appreciable amounts of immunoreactive IL-8 in the crude EC-CM. This result was in agreement with our previous data,” demonstrating the increased expression of the IL-8 gene and protein in bronchial epithelial cells of patients with symptomatic asthma.
IL-8 is a recently described peptide with neu- trophil and T-lymphocyte chemotactic activity.293 32 T lymphocytes appear to be two to 10 times more sensitive to this cytokine than neutrophils.32 In vivo studies carried out in animals have demon-
NONE ANTI-Cd ANTIC04 %b Fob
FIG. 5. Inhibitory effect of the univalent Fab fragment of an anti-CD4 antibody on the biologic activity of the T-cell chemotactic factor partially purified from the supernatants of bronchial epithelial cells of patients with asthma. These cells had been cultured for 48 hours in the presence (a) or absence (a) of lo-’ mol/L histamine. The concentrated (20 x) supernatants collected under each experimental condition from each donor were pooled and subjected to Sephadex G-100 and Sephadex QAE anion exchange chromatography after removal of IL-8 by immunoprecipi- tation. The fractions of resulting material that had demon- strated chemotactic activity for CD4 + T cells were pooled and retested for migration-enhancing activity with or with- out the anti-CD4 Fab in the bottom wells of the chemotac- tic chambers. Control samples were tested in the presence of the Fab fragment of an anti-CD3 antibody. CD4+ T- lymphocye migration is expressed on the ordinate as percentage of migration under control conditions (culture medium incubated for 48 hours in absence of cells, with or without histamine, and processed in parallel with the supernatants). Values are means +- SD from six experi- ments. *p < 0.01 versus the chemotactic factor tested alone (nonej.
strated the predominant recruitment of T lym- phocytes at the site of IL-8 injection when low doses of this peptide are used and a predominant recruitment of neutrophils when high doses are used.32 Although it seems that only a small subset of T cells express IL-8 receptors, there is no .evidence that CD4+ and CD8+ T lymphocytes respond differently to the migration-enhancing activity of IL-g?’
In our study the residual T-cell chemotactic activity of unstimulated bronchial epithelial cells

422 Bellini et al. J ALLERGY CLIN IMMUNOL SEPTEMBER 1993
i
I-
8
+ B * so
10 20 80 40 so
FRACTION NUMBER
FIG. 6. Chromatofocusing of active fractions of concentrated (20 x) supernatants from asthmatic epithelial cells, exposed to histamine, after Sephadex G-100 chromatography. Samples were eluted with ampholytes in the 6 to 10 pH range (- -). Eluates from chromatofocusing were adjusted to pH 7.4 and then assessed for enhancement of CD4+ T-cell migration. The migratory responses of CD4+ T lymphocytes are shown on the ordinate as percentage of migration observed under control conditions (culture medium supplemented with histamine, incubated for 46 hours in absence of cells, and processed in parallel with the supernatants).
from donors with asthma, detected afer inhibition of IL8 activity, could not be neutralized by the antibodies against two other cytokines that alter the motility of T lymphocytes, IL-130 and IG2.31 In addition, T-cell chemotactic activity was en- hanced by stimulation with 10m6 mol/L histamine. This chemotactic activity was partially purified by Sephadex G-100 and QAE anion exchange chro- matography or chromatofocusing and identified as a cationic protein with an apparent molecular weight of 56,000 d. The EC-CF selectively in- duced chemokinetic responses in CD4+ T lym- phocytes, and CD8+ T cells were unresponsive. The CD4 antigen appeared to be the potential receptor for this factor, as suggested by the inhib- itory activity of the univalent Fab fragment of anti-CD4 antibody.
The biochemical and functional characteristics of this EC-CF are similar to those of a previously
described and recently cloned lymphocyte chemoattractant factor (LCF).33-37 This lympho- kine is a 14,000 d cationic glycoprotein derived from antigen-, mitogen-, or histamine-stimulated CD8 + T lymphocytes and produced as a 56,000 d tetramer in cell cultures. The activities of LCF are limited to cells that express the CD4 molecule, and CD4 represents the cellular receptor for LCF. In addition to CD4 + T lymphocytes, mono- cytes and eosinophils express the CD4 molecule and both of these cell populations also migrate in response to LCF.33P 37 Further studies are needed to test the identity of the histamine-induced T-cell chemotactic activity of asthmatic bronchial epi- thelial cells with LCF.
Some recent articles’, 13, I4 have shown that the bronchial mucosa of patients with atopic asthma is infiltrated by a particular subset of CD4f T lymphocytes (T&-type cells), and these cells and

J ALLERGY CLIN IMMUNOL VOLUME 92, NUMBER 3
Bellini et al. 423
a 250 F 2 13 200 f
I- +
i 100
50
Hi--induoad EC-CF
0.5 . I
10
IL& -
U/ml I
FIG. 7. Comparison of the biologic activities of the T-cell chemotactic factor partially purified from the supernatants of histamine-stimulated asthmatic epithelial cells and the indicated concentrations of recombinant human IL-l@. CD4+ T-lymphocyte migration is expressed on the ordinate as percentage of migration observed with control medium (culture medium supple- mented with histamine, incubated for 48 hours in absence of cells, and processed in parallel with the supernatants). The different concentrations of IL-lf3 tested here were prepared by using this control medium as diluent. Values are mean 2 SD from four experiments, performed in triplicate.
their products seem to play an important role in the pathogenesis of airway hyperresponsiveness by promoting eosinophil migration and activa- tion.‘3-u, Our results provide the first evidence that bronchial epithelial cells may contribute to the recruitment of CD4+ T lymphocytes to the airways of patients with atopic asthma and suggest that the mechanism potentially involved in this process mlay be triggered or potentiated in vivo by the mast ~cell-derived histamine, once it has been released during antigen exposure.
We thank all the patients and healthy subjects who agreed to participate in this study. We also thank Dr. A. Laurenti and Ms. C. Rovati for general laboratory assistance, Dr. S. Vincenzi, Department of Bio- chemistry, University of Milan, for assistance with chro- matofocusing, and Ms. A. Grossi for typing this manu- script.
REFERENCES
1. Dunnill MS. The pathology of asthma with special refer- ence to change in the bronchial mucosa. J Clin Path01 1960;13:27-33.
2. Dunnill MS, Massarella GR, Anderson JA. A comparison of the quantitative anatomy of the bronchi in normal subjects, in status asthmaticus, in chronic bronchitis, and in emphysema. Thorax 1968;23:168-73.
3. Salvato G. Some histological changes in chronic bronchitis and asthma. Thorax 196&23:168-73.
4. Glynn AA, Michaels L. Bronchial biopsy in chronic bron- chitis and asthma. Thorax 1960,15:142-53.
5. Azzawi M, Bradley B, Jeffery PK, et al. Identification of activated T lymphocytes and eosinophils in bronchial bi- opsies in stable atopic asthma. Am Rev Respir Dis 1990; 1421407-13.
6. Mattoli S, Mattoso VL, Soloperto M, AIIegra L, Fasoli A. Cellular and biochemical characteristics of bronchoalveo- lar lavage fluid in symptomatic nonallergic asthma. J ALLERGY CLIN IMMUN~L 1991;87:794-802.
7. Marini M, Avoni E, Hollemborg J, Mattoli S. Cytokine mRNA profile and cell activation in bronchoalveolar la- vage fluid fron nonatopic patients with symptomatic asthma. Chest 1992;102:661-9.
8. Bellini A, Vittori E, Marini M, Ackerman V, Mattoli S. Intraepithelial dendritic cells and selective activation of Th2-like lymphocytes in patients with atopic asthma. Chest 1993;103:997-1005.
9. Frigas E, Gleich GJ. The eosinophil and the pathology of asthma. J ALLERGY CLIN IMMUNOL 198677527-37.
10. Barnes PJ. New concepts in the pathogenesis of bronchial

424 Bellini et al. J ALLERGY CLIN IMMUNOL SEPTEMBER 1993
hyperresponsiveness and asthma. J ALLERGY CLIN IMMIJNOL 1989;87:1470-3.
11. Gundel RI-I, Letts LG, Gleich GJ. Human eosinophil major basic protein induces airway constriction and airway hyperresponsiveness in primates. J Clin Invest 1991;87: 1470-73.
12. Bousquet J, Chanez P, Lacoste JY, et al. Eosinophilic inflammation in asthma. New Engl J Med 1990;323:1033-9.
13. Hamid Q, Azzawi M, Ying S, et al. Expression of mRNA for interleukin 5 in mucosal biopsies from asthma. J Clin Invest 1991;87:1541-6.
14. Robinson DS, Hamid Q, Ying S, et al. Predominant ThZ-like bronchoalveolar T-lymphocyte population in atopic asthma. N Engl J Med 1992;326:298-304.
15. Lopez AF, To LB, Yang Y, et al. Stimulation of prolifer- ation, differentiation and function of human cells by primate interleukin 3. Proc Nat1 Acad Sci U S A 198784: 2761-5.
16. Yamaguchi Y, Hayashi Y, Sugama Y, et al. Highly purified murine interleukin 5 stimulates eosinophil function and prolongs in vitro survival. IL.5 as an eosinophil chemotac- tic factor. J Exp Med 1988;167:1737-42.
17. Wang JM, Rambaldi A, Biondi A, Chan ZG, Sanderson CJ, Mantovani A. Recombinant human interleukin 5 is a selective eosinophil chemoattractant. Eur J Immunol 1989;19:701-5.
18. Lopez AF, Sanderson CJ, Gamble JR, Campbell HD, Young IG, Vadas MA. Recombinant human interleukind is a selective activator of human eosinophil function. J Exp Med 1988;167:219-24.
19. Owen WF, Rothenberg ME, Silberstein AS, et al. Regu- lation of human eosinophil viability, density and function by granulocyte/macrophage colony-stimulating factor in the presence of 3T3 fibroblasts. J Exp Med 1987;166:129- 41.
20. Walker C, Virchow J-C Jr, Bruijnzeel PLB, Blaser K. T cell subsets and their soluble products regulate eosinophilia in allergic and non-allergic asthma. J Immunol 1991;146: 1829.35.
21. Corbo GM, Foresi A, Morandini S, Valente S, Mattoli S, Ciappi G. Probit analysis applied to the allergen dose- response curve: a method for epidemiologic surveys. J ALLERGY CLIN 1-0~ 1988,81:41-7.
22. Mattoli S, Rosati G, Mormile F, Ciappi G. The immediate and short-term effects of corticosteroids on cholinergic hyperreactivity and pulmonary function in subjects with well-controlled asthma. J ALLERGY CLIN IMMLJNOL 1985;76: 214-22.
23. Soloperto M, Mattoso VI, Fasoli A, Mattoli S. A bron- chial epithelial cell-derived factor in asthma which pro- motes eosinophil activation and survival as GM-CSF. Am J Physiol (Lung Cell Med Physiol, 4) 1991;26O:L530-L.538.
24. Marini M, Vittori E, Hollemborg J, Mattoli S. Expression of the potent inflamamtory cytokines, granulocyte-macro- phage-colony-stimulating factor and interleukin-6 and in- terleukin-8, in bronchial epithelial cells of asthmatic pa- tients. J ALLERGY CLIN IMMUNOL 1992;89:1001-9.
25. NHLBI Workshop Summaries. Summary and recormnen- dations of a workshop on the investigative use of fiberoptic bronchoscopy and bronchoalveolar lavage in asthmatics. Am Rev Respir Dis 1985;132:180-2.
26. Mattoli S, Colotta F, Fincato G, et al. Time course of IL1 and IL6 synthesis and release in human bronchial epithe- lial cell cultures exposed to toluene diisocyanate. J Cell Physiol 1991;149:260-8.
27. Sluyterman LAA, Elgersma 0. Chromatofocusing: isoelec- tric focusing on ion exchange columns. I. General princi- ples. J Chromatogr 1978;150:17-30.
28. Sluyterman LAA, Wijdenes J. Chromatofocusing: isoelec- tric focusing on ion exchange columns. II. Experimental verification. J Chromatogr 1978;150:31-44.
29. Marini M, Sciacca F, Colotta F, Mantovani A, Mattoli S. Protective effect of nedocromil sodium on the interleukin- 1 induced production of interleukin-8 in human bronchial epithelial cells. J ALLERGY CLIN IMMUNOL 1992;90:76-84.
30. Miossec P, Yu C, Ziff M. Lymphocyte chemotactic activity of human interleukin 1. J Immunol 1984;133:2007-13.
31. Komfeld H, Berman JS, Beer DJ, Center DM. Induction of human T lymphocyte motility by interleukin 2. J Immu- no1 1985;134:3887-90.
32. Larsen CG, Anderson AO, Appella E, Qppenheim JJ, Matsushima K. The neutrophil-activating protein (NAP-l) is also chemotactic for T lymphocytes. Science 1989;243: 1464-6.
33. Rand TH, Cruikshank W, Center DM, Weller PF. CD4- mediated stimulation of human eosinophils: lymphocyte chemoattractant factor and other CDCbinding ligands elicit eosinophil migration. J Exp Med 1991;173: 1521-8.
34. Berman JS, Cruikshank WW, Center DM, Theodore AC, Beer DJ. Chemoattractant lymphokines specific for the helper/inducer T-lymphocyte subsets. Cell Immunol 1985; 95:105-12.
35. Berman JS, Beer DJ, Theodore AC, Komfeld H, Bernard0 J, Center DM. State of the art: lymphocyte recruitment to the lung. Am Rev Respir Dis 1990;142:238-57.
36. Kaye J, Janeway CA. The Fab fragment of a directly activating monoclonal antibody that precipitates a disul- fide-linked heterodimer from a helper T cell clone blocks activation by either allogenic Ia or antigen and self-la. J Exp Med 1984;159:1397-1402.
37. Cruikshank WW, Berman JS, Theodore AC, Bernard0 J, Center DM. Lymphokine activation of T4+ T lympho- cytes and monocytes. J Immunol 1987;138:3817-23.





























