Embed Size (px)
Citation preview

Research in Microbiology 165 (2014) 175e189www.elsevier.com/locate/resmic
CadA of Mesorhizobium metallidurans isolated from a zinc-rich mining soilis a PIB-2-type ATPase involved in cadmium and zinc resistance
Geraldine Maynaud a,*, Brigitte Brunel b, Erika Yashiro b,1, Max Mergeay c,Jean-Claude Cleyet-Marel a, Antoine Le Quere d,2
a INRA USC1242, Universite Montpellier II, TA A-82/J, 34398 Montpellier Cedex 5, FrancebMontpellier SupAgro, Universite Montpellier II, TA A-82/J, 34398 Montpellier Cedex 5, France
cMolecular and Cellular Biology, Unit Microbiology, Belgian Nuclear Research Center (SCK CEN), Boeretang 200, B-2400 Mol, Belgiumd IRD, Laboratoire des Symbioses Tropicales et Mediterraneennes UMR113, IRD/INRA/CIRAD/Montpellier SupAgro/Universite Montpellier II, TA A-82/J, 34398
Montpellier Cedex 5, France
Received 21 May 2013; accepted 13 February 2014
Available online 5 March 2014
Abstract
Mesorhizobium metallidurans STM 2683T is a nitrogen-fixing bacterium that nodulates Anthyllis vulneraria in mine tailings highlycontaminated in zinc, lead and cadmium. To study the mechanisms whereby this bacterium copes with metals, we functionally screened acosmid genomic library of M. metallidurans for zinc or cadmium tolerance. A cosmid clone harbored a gene encoding PIB-type ATPase ho-mologous to CadA that leads to cadmium and zinc resistance in Escherichia coli. The CadA protein structure presents one duplication of the twoN-terminal metal binding domains (i.e. a heavy metal-associated domain followed by a histidine-rich domain) which allows specific binding tozinc and cadmium cations. A cadA-deleted strain of M. metallidurans failed to grow at high zinc concentrations (2 mM) and its growth wasdelayed at lower zinc concentrations. Expression studies using a transcriptional fusion of cadA promoter to gfp showed that cadA is specificallyinduced in a dose-dependent manner by zinc and cadmium in M. metallidurans in vitro conditions and into A. vulneraria nodules after Zn stress.Metal induction sensitivity was increased in the strain where cadA gene was deleted.
This study identified cadA as a first mesorhizobial resistance determinant involved in detoxification of cadmium and zinc and which confersupon M. metallidurans greater capacity for coping with high zinc concentrations. This improves the knowledge of this bacterium for potentialuse as a symbiotic inoculant of Anthyllis in phytostabilization strategies of metal-rich sites.� 2014 Institut Pasteur. Published by Elsevier Masson SAS. All rights reserved.
Keywords: Metal efflux transporter; PIB-type ATPase; cadA transcription; Metal tolerance; Mesorhizobium
* Corresponding author. Tel.: þ33 467593801; fax: þ33 467593802.
E-mail addresses: [email protected] (G. Maynaud), brunel@supagro.
inra.fr (B. Brunel), [email protected] (E. Yashiro), [email protected]
(M. Mergeay), [email protected] (J.-C. Cleyet-Marel), antoine.le-quere@
ird.fr (A. Le Quere).1 Present address: Universite de Lausanne, Departement de Microbiologie
Fondamentale, Batiment Biophore, CH-1015 Lausanne, Switzerland.2 Present address: Laboratoire Mixte International, Biotechnologie Micro-
bienne et Vegetale, Rabat, Morocco.
http://dx.doi.org/10.1016/j.resmic.2014.02.001
0923-2508/� 2014 Institut Pasteur. Published by Elsevier Masson SAS. All rights
1. Introduction
Metals released by anthropogenic activities are amajor causeof environmental pollution due to their toxicity [70]. Mininggenerates spoils that are extremely rich in toxic metals whichoften contaminate the surrounding soils and waters. Since theycannot be degraded or destroyed, they persist within naturalenvironments and have been shown to disturb ecosystem dy-namics. Metal pollution constitutes a significant threat to publichealth [33] and to natural environments [52]. Some metals (zinc
reserved.

176 G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
(Zn), iron (Fe) or cobalt (Co)) are essential for life at low con-centrations and play important roles in biological cell func-tioning [3]. High concentrations of these essential metals or oftoxic metals with unknown biological roles, such as cadmium(Cd), lead (Pb) or mercury (Hg), can damageDNA structure andcell membranes, alter enzymatic activity and disrupt cellularfunctions because of their interference with oxidative phos-phorylation and osmotic balance [15].
Bacteria use a variety of mechanisms to protect sensitivecellular components against metals, such as metal exclusion,thanks to a permeability barrier [71], efflux transport [50],intracellular sequestration [35], precipitation as insoluble saltsto reduce their bioavailability [1] or transformation of metalsinto less toxic forms by enzymatic detoxification [45]. Amongthe known metal resistance mechanisms, efflux systems are themost frequently represented; these include PIB-type ATPases,the family of resistant nodulation cell division (RND) proteinsspecialized in the efflux of heavy metal (HME-RND systems),and cation diffusion facilitator proteins (CDFs) [50]. The best-characterized Zn- and Cd-efflux systems include (i) the PIB-type ATPases CadA and ZntA (Cd, Zn and Pb resistance)identified in Staphylococcus aureus [51] and Escherichia coli[59], respectively, (ii) the HME-RND systems CzcCBA (Co,Ni, Zn and Cd resistance) from the pMOL30 of Cupriavidusmetallidurans CH34 [20] and CzrCBA (Cd and Zn resistance)from Pseudomonas aeruginosa [30] and (iii) the CDF CzcD(Zn, Co and Cd resistance) described in C. metallidurans CH34[4] and the Zn-specific CDF ZitB from E. coli [26].
PIB-type ATPase transporters belong to the large super-family of ATP-driven pumps involved in the transport ofmetals across cell membranes against their concentrationgradients to maintain constant and non-toxic levels of metalswithin the cytoplasm. They are grouped into four major sub-groups according to distinct metal binding and transportspecificities [5]: PIB-1 or Cuþ/Agþ-ATPases, PIB-2 or Zn2þ/Cd2þ/Pb2þ-ATPases, PIB-3 or Cuþ/Agþ/Cu2þ-ATPases andPIB-4 or Co
2þ-ATPases. The PIB-type ATPases have structuralfeatures that distinguish them from their P-type ATPasecounterparts such as (i) a topological arrangement of 6e8transmembrane domains (TMs) flanking the large cytoplasmicloop, (ii) several regulatory cytoplasmic metal binding do-mains located in the N-terminal region (N-MBDs) which areresponsible for metal recognition, regulation and movementsacross the membrane permeability barrier [5,8,39] and (iii) asignature sequence in the sixth TM containing a CPX(X¼ cysteine, histidine or serine), a SPC or a TPC motifdownstream of the largest cytoplasmic loop that might be partof the cation channel [5,6,41].
Saint-Laurent-le-Minier in the south of France was once animportant mining area for Zn exploitation (1872e1914).Among the former mining sites, that of Avinieres is a hotspotof high pollution by metals, particularly Zn, Cd and Pb [40].However, efficient nitrogen-fixing symbiosis between theleguminous plant Anthyllis vulneraria and the rhizobial spe-cies Mesorhizobium metallidurans was identified at the Avi-nieres site [68]. This Anthyllis-Mesorhizobium symbiotic pairhas recently been described as a key potential bioremediation
agent for stimulating sustainable plant cover which could thenlimit metal dispersion from the contaminated sites [24,40]. AsA. vulneraria and the microbiont M. metallidurans are able tosurvive and grow in such highly contaminated soils, they arelikely to have developed strategies for tolerating high con-centrations of metals such as Zn or Cd [68].
The aim of this work was to investigate possible moleculardeterminants involved in M. metallidurans Zn and Cd adap-tation in order to gain a better understanding of its adaptationin such highly polluted mining sites. Using a functionalscreening approach, we isolated a genomic region of the M.metallidurans type strain that responded to Zn and Cd stress.A putative PIB-type ATPase was identified and its role in metalresistance was further analyzed.
2. Materials and methods
2.1. Bacterial strains, cosmids, plasmids and growthconditions
The bacterial strains, plasmids and cosmids used in thisstudy are listed in Table S1. E. coli strains were cultured inLuria Bertani (LB) broth at 37 �C [61]. Mesorhizobium strainswere cultured at 28 �C in tryptone yeast extract (TY) mediumor TY medium supplemented with 0.5 mM Na2-EDTA whenPb was used [16]. Traces of Zn contained in the TY medium(9 mM) were measured by atomic absorption spectrometry(UMR 5569 Hydrosciences, Montpellier, France). Ampicillin(Ap, 50 mg ml�1), tetracycline (Tc, 15 mg ml�1), gentamicin(Gm, 10 mg ml�1), kanamycin (Km, 50 mg ml�1), chloram-phenicol (Cm, 50 mg ml�1), isopropyl-beta-D-thiogalactopyr-anoside (IPTG, 100 mM) and 5-bromo-4-chloro-3-indolyl-beta-D-galactosidase (X-gal, 80 mg ml�1) were added asrequired. Solid media contained 2% agar.
2.2. Molecular techniques
General DNAmanipulations were performed using standardtechniques [61]. Products from PCR and restriction digestionswere cleaned up using Illustra� GFX� PCR DNA and GelBand purification kits (GE Healthcare). All enzymes werepurchased from Promega and all primers from Eurofins. Theprimers used in this study are listed in Table S1. All con-structions were sequenced by Genoscreen Company (France).
2.3. Minimal inhibitory concentration (MIC) assays
MIC assays were determined in liquid medium. MIC wasdefined as the lowest concentration of metal that inhibitedvisible growth. E. coli strains were precultured overnight andMesorhizobium strains for 4 days, and each culture wasadjusted to OD600nm¼ 0.5 before inoculation. One hundredand fifty microliters of medium containing increasing con-centrations of metal (ZnSO4 or CdCl2 or NiSO4 or CoCl2 orPbCl2) or no metal (positive control) were inoculated with10 ml of preculture and incubated at 28 �C or 37 �C for Mes-orhizobium or E. coli strains, respectively, and at 150 rpm.

177G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
Growth was monitored following the OD600nm by using aspectrofluorometer (TECAN Infinite M200) for 24 h for E.coli strains and for 1 week for Mesorhizobium strains.
2.4. Construction of a cosmid library of M.metallidurans STM 2683T and screening for Zn or Cdtolerance
Total DNA from M. metallidurans STM 2683T waspartially digested with Sau3AI and size-fractionated on a 0.3%agarose gel. The resulting DNA fragments (approximately40 kb) were ligated into the pCPP47 vector [10] that wasdigested with BamHI and treated with rapid alkaline phos-phatase [57]. The resulting cosmid library was constructedusing commercial E. coli XLI-Blue MR supercompetent cells(Stratagene). From this library, 2000 clones were individuallypicked and stored in twenty-two 96-well microtiter plates assingle clones while the remainder of the library was pooledand stored in cryotubes as clone pools containing approxi-mately 20,000 clones. Screening for Zn or Cd resistance wascarried out by plating an aliquot of the cosmid library on LBplates supplemented with 4 mM Zn or 1.6 mM Cd. Theseconcentrations inhibited the growth of the E. coli host con-taining the empty vector pCPP47. Selected cosmid cloneswere isolated using the alkaline lysis method followed bycosmid purification with phenolechloroform and isopropanolprecipitation (Wizard Plus SC Minipreps DNA PurificationSystem, Promega), and their DNA restriction digestion anal-ysis was carried out with EcoRI to verify the presence of aninsert in pCPP47. Five cosmids with inserts and the emptyvector pCPP47 were further transferred into the Zn-sensitiveE. coli strain GG48 (KmR and CmR) [26] by tri-parentalmating, using the helper plasmid pRK2013 [23]. Growth onLB agar plates supplemented with Km, Cm and Tc allowedselection of GG48 recombinants. To confirm the functionalrole of the cloned loci in metal tolerance, MIC assays in thepresence of increasing Zn or Cd concentrations were per-formed on the respective GG48 transconjugants.
2.5. Subcloning and identification of metal toleranceopen reading frames (ORFs)
The Cd-tolerant cosmid clone MPCdR2 was subcloned intothe mobilizable and medium copy number pBBR1MCS-5 vec-tor [36]. MPCdR2 was digested with EcoRI. The restrictionfragments were ligated into EcoRI-linearized pBBR1MCS-5.The ligation mixture was transferred into E. coli DH10B elec-trocompetent cells by electroporation [22,63] and transformantswere plated onto LB solid medium supplemented with Gm and1.6 mM Cd to select Cd-tolerant subclones. A selection ofplasmids from Cd-tolerant DH10B subclones were purified anddigested with EcoRI so as to confirm the presence of an insertand to assess its sizes. All the selected subclones contained aninsert of approximately 3 kbp. One DH10B subclone was cho-sen and the plasmid named pBBR1-CdE1 was sequenced usinguniversal T3 and T7 primers and custom-designed internalprimers (Table S1). DNA sequenceswere analyzedwith BioEdit
version.7.0.9.0 [28]. Putative open reading frames (ORFs) wereidentified with the BLASTX program [2]. The sequence of theinsert was mapped to the STM 2683T draft genome (EuropeanNucleotide Archive CAUM01000001-CAUM01000191) toidentify the DNA region involved in metal tolerance. Finally,pBBR1-CdE1 and the empty plasmid pBBR1MCS-5 weretransferred into the Zn-sensitive E. coli GG48 by tri-parentalmating as described previously. The GG48 recombinants wereselected on LB solid medium supplemented with Km, Cm andGm and subjected to MIC assays as described previously.
2.6. Transposon mutagenesis
Tn5-transposons containing the KmR gene were randomlyintroduced in vitro into MPCdR2 using the EZ-Tn5 KAN-1insertion kit (Epicentre) to generate MPCdR2::UKm. Recom-binant plasmids were transformed into the E. coli EC100 strainfollowing TransforMax� EC100 Electrocompetent E. coli in-structions (Epicentre) and clones were selected on Km. TheKmR clones were screened negatively on LB medium with1.5 mMCd andwere checked byMIC assayswith increasingCdconcentrations (1, 1.5 and 2 mM). The Zn tolerance phenotypewas investigated on liquid LB medium containing 1, 2 and2.5 mM Zn. KmR clones were sequenced bidirectionally usingthe KAN-2 FP-1 forward and KAN-2 RP-1 reverse transposon-specific-primers provided in the kit. DNA sequences wereanalyzed using BioEdit. ORFs were identified with theBLASTX program and the amino acid sequences obtained werecompared to the Genbank database using the BLAST Proteinprogram (BLASTP) in order to identify the most similar se-quences available. For complementation assays, pBBR1-CdE1was introduced into electrocompetent EC100 cells containingMPCdR2::UKm (described above) by electroporation andtransformants were selected with Tc and Gm. The restoration ofCd and Zn tolerance in Tn5-mutated E. coli EC100 was studiedby transforming the pBBR1-CdE1 plasmid in several clones ofEC100 containing MPCdR2::UKm. Metal sensitivity wasmeasured by MIC assays.
2.7. Phylogenetic analysis and structural description ofCadA
The sequence obtained from the Zn- and Cd-resistantDH10B subclone containing pBBR1-CdE1 matched with aCDS encoding PIB-type ATPase and theM. metallidurans genewas named cadA. cadA amino acid sequencewas blasted againstthe Genbank protein database in order to identify the mostsimilar sequences available. The amino acid sequence corre-sponding to the cadA gene was aligned with 15 bacterial PIB-type ATPase sequences identified by BLASTP, and a phyloge-netic tree was inferred by the maximum likelihood methodusing Mega 5 v5.1.1.0426 software [66]. The tree was rootedusing Cupriavidus metallidurans CH34 strain as an outgroup.For phylogenetic analysis, the sequences were aligned usingClustalX version.1.83 and optimized manually with GeneDocversion.2.7.00 [49]. The confidence level of the tree brancheswas estimated with 1000 bootstrap repetitions. The number of

178 G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
TMs and protein topology were predicted by HMMTOP serverversion 2.0 [67] and the conserved domains were definedmanually by comparing CadA with the closest protein se-quences retrieved from Genbank.
2.8. Construction of a cadA mutant andcomplementation experiments
The 2917-bp region carrying the cadA gene with its flankingregions including the partial 50 promoting region (104 bp)(accession number HE820903) was amplified by PCR withcustom-designed primers (STM 2849 forward and STM 2785reverse, Table S1) including XbaI sites. The PCR-amplifiedproduct was cloned as an XbaI fragment into a pGEM-TEasy Vector (Promega) to create pGEM-cadA. The 2.5-kbHindIII-flanked UKm resistance cassette (UKmR) wasexcised from pHP45 UKm and inserted in place of a 2.5-kbHindIII-deleted region carrying cadA. The resulting pGEM-cadA::UKm plasmid was digested with XbaI and the fragmentcontaining cadA::UKm was ligated into suicide vectorpJQ200SK to create pJQ-cadA::UKm. The latter was used totransform electrocompetentM. metallidurans STM 2683T cells[31]. Transformants were selected on TY plates containing Kmand 5% sucrose to allow selection of knockout clones that hadundergone a double cross-over. Double cross-over genereplacement in mutants was verified by screening for antibiotic-resistant phenotypes (KmR) and also by PCR using STM 2985forward and STM 2986 reverse primers to detect the insertedconstruction, STM 2612 forward and STM 2613 reverseprimers targeted to the deleted fragment, and STM 2849 for-ward and STM 2987 reverse primers to detect the presence ofthe UKmR cassette. To test for complementation of the mutatedstrain (named DcadA), a 3-kb PCR fragment containing thecadA gene and its promoter was PCR-amplified with STM 2849forward and STM 2785 reverse primers, then cloned into apGEM-T Easy vector before being sequenced and furthercloned into XbaI-linearized pBBR1MCS-5, generating thepBBR1-cadA plasmid. The latter construction was then used totransform electrocompetent cells of the DcadA strain. Com-plemented clones were confirmed by amplifying the cadA geneby PCR using STM 2245 forward and STM 2263 reverseprimers targeted to the mutated region, and one clone (namedDcadA-cadA) was chosen for phenotypic analysis.
2.9. Induction experiments using a promoter-gfp fusionreporter system on cell cultures
For testing the promoter activity of the cadA 50 region, apromoter/reporter-gene fusion was constructed in the broad-host-range promoter-probe vector pPROBE-GT [44], whichallows transcriptional fusions with the gfp gene. The 156-bpregion contained the 104-bp region upstream of the cadAstart codon harboring a regulatory site for metal-sensing reg-ulators of the MerR-family [34], and 52-bp downstream, thestart codon was PCR-amplified from the STM 2683T genomewith STM 2740 forward and STM 2244 reverse primers. ThePCR product was cloned into a pGEM-T Easy vector to create
pGEM-cadAp. It was digested with EcoRI and the fragmentwas ligated into the EcoRI site of the pPROBE-GT vector(renamed pGT), upstream of the gfp gene to create pGT-cadAp, and its sequence was determined by sequencing. Theplasmids pGT-cadAp and pGT were electroporated into STM2683T and into the DcadA strain, and transformants wereselected with Gm and GmþKm, respectively. The level ofcadA transcription, as estimated by the differential gfp-induced fluorescence obtained from the two strains containingpGT-cadAp or pGT, were calculated after incubation in TYmedia presenting increasing concentrations of Cd (0.00625,0.0125, 0.025, 0.05, 0.1, 0.2, 0.3, 0.4 and 0.8 mM), Zn(0.0125, 0.025, 0.05, 0.1, 0.2, 0.4, 0.5, 1, 1.5 and 2 mM), Ni(0.01, 0.02 and 0.03 mM), Co (0.01, 0.02 and 0.03 mM) andPb (0.05, 0.1 and 0.2 mM) and mixes of Zn and Cd. Theinducible activities of the cadA promoter upon metal exposurewere quantified by measuring cell fluorescence intensities witha TECAN microplate spectrofluorometer, using excitation andemission wavelengths of 485 nm and 535 nm, respectively.The relative expression values were obtained by subtractingthe fluorescence values of corresponding non-inoculated con-trols and by normalizing the fluorescence to cell density.Statistical analyses were performed using one-way ANOVAplus the Newman-Keuls post-hoc test, with p-values <0.01considered as significant. All assays were performed at leastthree times.
The metal-dependent cadA transcriptional responses, asassessed by gfp expression under control of the cadA promoterwere subjected to non-linear regression analysis and fitted tothe sigmoidal doseeresponse curves with a variable slope(four parameters) using the Prism software package (version.5,GraphPad software) as described previously [27]. We usedobserved bottom (when no metal was added to the growthmedium) and top transcriptional relative expression levels tocalculate the background expression level and the maximum-fold induction levels for each strain within the concentrationrange that did not induce growth differences between thecompared strains. We also determined the effective concen-tration (EC50) corresponding to half the maximum transcrip-tional response (except for the wild-type response to Zn, forwhich we used the maximum relative expression), the Hillslope (or Hill coefficient), which describes the steepness of thecurve indicating ultrasensitivity of the response [56] andgoodness of fit as expressed by the R2 values.
2.10. cadA induction experiments on nodules of A.vulneraria
Seeds of A. vulneraria were surface-sterilized, planted andinoculated with M. metallidurans STM 2683T containing pGT-cadAp or empty pGT, or with the mutated cadA-deleted M.metallidurans strain containing pGT-cadAp or empty pGT, asdescribed previously [64]. Plants were pre-cultivated withoutZn 7 weeks post-inoculation in glass tubes containing calci-nated attapulgite (Oil Dri US Special, Damolin, Denmark;http://www.damolin.dk) in a greenhouse (for 14 h per day and10 h at night; day T�: 22 �C and night T�: 15 �C). Then, the
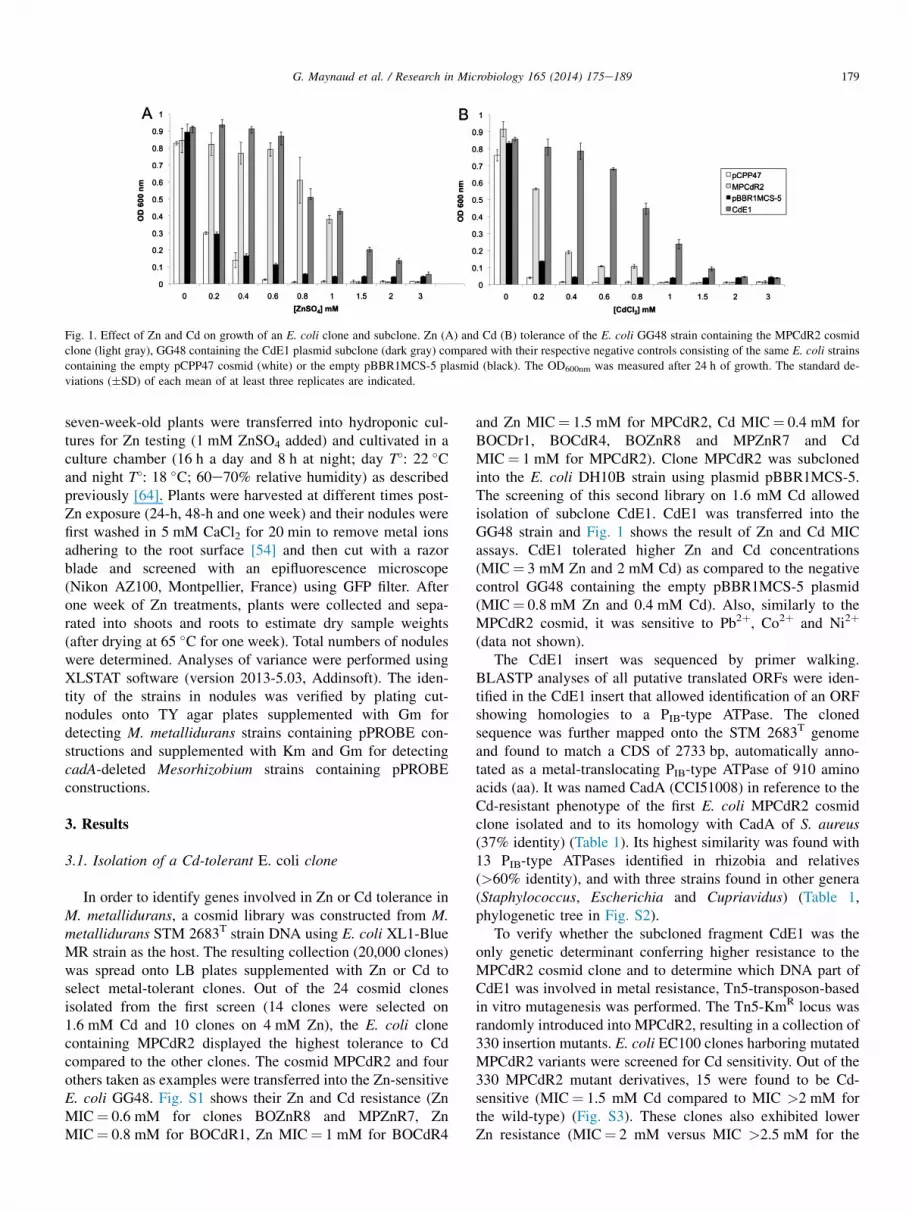
Fig. 1. Effect of Zn and Cd on growth of an E. coli clone and subclone. Zn (A) and Cd (B) tolerance of the E. coli GG48 strain containing the MPCdR2 cosmid
clone (light gray), GG48 containing the CdE1 plasmid subclone (dark gray) compared with their respective negative controls consisting of the same E. coli strains
containing the empty pCPP47 cosmid (white) or the empty pBBR1MCS-5 plasmid (black). The OD600nm was measured after 24 h of growth. The standard de-
viations (�SD) of each mean of at least three replicates are indicated.
179G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
seven-week-old plants were transferred into hydroponic cul-tures for Zn testing (1 mM ZnSO4 added) and cultivated in aculture chamber (16 h a day and 8 h at night; day T�: 22 �Cand night T�: 18 �C; 60e70% relative humidity) as describedpreviously [64]. Plants were harvested at different times post-Zn exposure (24-h, 48-h and one week) and their nodules werefirst washed in 5 mM CaCl2 for 20 min to remove metal ionsadhering to the root surface [54] and then cut with a razorblade and screened with an epifluorescence microscope(Nikon AZ100, Montpellier, France) using GFP filter. Afterone week of Zn treatments, plants were collected and sepa-rated into shoots and roots to estimate dry sample weights(after drying at 65 �C for one week). Total numbers of noduleswere determined. Analyses of variance were performed usingXLSTAT software (version 2013-5.03, Addinsoft). The iden-tity of the strains in nodules was verified by plating cut-nodules onto TY agar plates supplemented with Gm fordetecting M. metallidurans strains containing pPROBE con-structions and supplemented with Km and Gm for detectingcadA-deleted Mesorhizobium strains containing pPROBEconstructions.
3. Results
3.1. Isolation of a Cd-tolerant E. coli clone
In order to identify genes involved in Zn or Cd tolerance inM. metallidurans, a cosmid library was constructed from M.metallidurans STM 2683T strain DNA using E. coli XL1-BlueMR strain as the host. The resulting collection (20,000 clones)was spread onto LB plates supplemented with Zn or Cd toselect metal-tolerant clones. Out of the 24 cosmid clonesisolated from the first screen (14 clones were selected on1.6 mM Cd and 10 clones on 4 mM Zn), the E. coli clonecontaining MPCdR2 displayed the highest tolerance to Cdcompared to the other clones. The cosmid MPCdR2 and fourothers taken as examples were transferred into the Zn-sensitiveE. coli GG48. Fig. S1 shows their Zn and Cd resistance (ZnMIC¼ 0.6 mM for clones BOZnR8 and MPZnR7, ZnMIC¼ 0.8 mM for BOCdR1, Zn MIC¼ 1 mM for BOCdR4
and Zn MIC¼ 1.5 mM for MPCdR2, Cd MIC¼ 0.4 mM forBOCDr1, BOCdR4, BOZnR8 and MPZnR7 and CdMIC¼ 1 mM for MPCdR2). Clone MPCdR2 was subclonedinto the E. coli DH10B strain using plasmid pBBR1MCS-5.The screening of this second library on 1.6 mM Cd allowedisolation of subclone CdE1. CdE1 was transferred into theGG48 strain and Fig. 1 shows the result of Zn and Cd MICassays. CdE1 tolerated higher Zn and Cd concentrations(MIC¼ 3 mM Zn and 2 mM Cd) as compared to the negativecontrol GG48 containing the empty pBBR1MCS-5 plasmid(MIC¼ 0.8 mM Zn and 0.4 mM Cd). Also, similarly to theMPCdR2 cosmid, it was sensitive to Pb2þ, Co2þ and Ni2þ
(data not shown).The CdE1 insert was sequenced by primer walking.
BLASTP analyses of all putative translated ORFs were iden-tified in the CdE1 insert that allowed identification of an ORFshowing homologies to a PIB-type ATPase. The clonedsequence was further mapped onto the STM 2683T genomeand found to match a CDS of 2733 bp, automatically anno-tated as a metal-translocating PIB-type ATPase of 910 aminoacids (aa). It was named CadA (CCI51008) in reference to theCd-resistant phenotype of the first E. coli MPCdR2 cosmidclone isolated and to its homology with CadA of S. aureus(37% identity) (Table 1). Its highest similarity was found with13 PIB-type ATPases identified in rhizobia and relatives(>60% identity), and with three strains found in other genera(Staphylococcus, Escherichia and Cupriavidus) (Table 1,phylogenetic tree in Fig. S2).
To verify whether the subcloned fragment CdE1 was theonly genetic determinant conferring higher resistance to theMPCdR2 cosmid clone and to determine which DNA part ofCdE1 was involved in metal resistance, Tn5-transposon-basedin vitro mutagenesis was performed. The Tn5-KmR locus wasrandomly introduced into MPCdR2, resulting in a collection of330 insertion mutants. E. coli EC100 clones harboring mutatedMPCdR2 variants were screened for Cd sensitivity. Out of the330 MPCdR2 mutant derivatives, 15 were found to be Cd-sensitive (MIC¼ 1.5 mM Cd compared to MIC >2 mM forthe wild-type) (Fig. S3). These clones also exhibited lowerZn resistance (MIC¼ 2 mM versus MIC >2.5 mM for the

Table 1
Comparison of PIB-type ATPase sequences. Comparison of CadA from M. metallidurans STM 2683T with 13 PIB-type ATPases of rhizobia and relative strains and
three reference strains (E. coli, S. aureus and C. metallidurans).
PIB-type ATPase
accession number
PIB-type ATPase name Bacteria source % Identity (over amino
acid range indicated)
CAAF010000000 MESS4_360013 Mesorhizobium sp. STM 4661 97% (916)
YP665761 Meso_4129 Chelativorans multitrophicus BNC1 61% (972)
YP002544256 Arad_2044 Agrobacterium radiobacter K84 65% (775)
YP002977578 Rleg_3796 Rhizobium leguminosarum sv. trifolii WSM1325 67% (761)
YP001243187 Bbta_7421 Bradyrhizobium sp. BTAi1 67% (756)
YP769837 RL4262 Rhizobium leguminosarum sv. viciae 3841 67% (754)
YP002282994 Rleg2_83501 Rhizobium leguminosarum sv. trifolii WSM2304 66% (752)
YP001980107 RHECIAT_CH0003992 Rhizobium etli CIAT 652 67% (751)
YP471201 RHE_CH03719 Rhizobium etli CFN 42 68% (751)
NP103812 mll_2475 Mesorhizobium huakuii MAFF303099 65% (754)
NP384230 Smc_04128 Ensifer meliloti 1021 63% (752)
YP001328992 Smed_3336 Ensifer medicae WSM419 63% (763)
YP570767 RDP_3643 Rhodopseudomonas palustris BisB5 64% (747)
P37617 ECK3453 Escherichia coli K12 51% (636)
P37386 CADA2_STAAU Staphylococcus aureus 1280 37% (760)
CP000353 Rmet_4594 Cupriavidus metallidurans CH34 35% (826)
The results are from BLASTP searches of CadA from M. metallidurans with Genbank’s protein database. PIB-type ATPases are listed by their accession number
(first column) and their name (second column). The percent identities and the lengths of the homologous regions from BLASTP are shown. The closest match
found was PIB-type ATPase MESS4_360013 of Mesorhizobium sp. STM 4661 (97% identity).
180 G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
wild-type) (Fig. S3). Insertion sites of 11 MPCdR2 mutantssensitive to both Zn and Cd were characterized. Sequenceanalysis showed that they were all mutated in the domainsrequired for PIB-type ATPase functioning (phosphorylation,phosphatase activity, ATP-binding domain) and metal recog-nition and movement across the membrane permeability barrier(TM-MBD containing the CPX motif). After complementationof the Tn5 mutants by insertion of the CdE1 insert containingthe cadA gene, the Cd- and Zn-resistant phenotypes wererestored, confirming that cadA played a key role in theincreased tolerance to these metals (Fig. S3).
3.2. CadA belongs to the PIB-2-type ATPase subgroup
The HMMTOP transmembrane protein prediction tool pre-dicted that CadA contained eight potential TMs with a largecytoplasmic loop between TM6 and TM7. Moreover, it showedthree regions whose domains presented homologies with otherPIB-type ATPases, all of them involved in metal transport: (i) aduplicated 61-aa HMA domain in the N-terminal region con-taining one GXXCXXC motif each, (ii) the 225-aa PIB-typeATPase region containing the DKTGT motif with the phos-phorylation site of an aspartate residue, the phosphatase domain(TGES motif) and the ATP-binding domain (GDGXNDXPmotif), and (iii) the 125-aa C-terminal region which containsthe halo acid dehalogenase (HAD) domain (hydrolase-likedomain). The CadA sequence and 15 selected PIB-type ATPases(Table 1) were aligned and compared using Arguello’s PIB-typeATPase database [5] (Fig. 2). CadA exhibited all the charac-teristics of the PIB-2-family, with the TM-MBD containing theCPC motif in TM6 associated with a lysine (K) in TM7 and anaspartate (D), an asparagine (N) and a glycine (G) in TM8(signature sequence N(X)7K(X)20DXG(X)7N) (Fig. 2B and C).In addition, CadA had a duplicated N-MBD containing a
duplicated GMDCAAC motif of the GXXCXXC-type (alsonamed the HMA domain) (Fig. 2A) and another duplicatedN-MBD containing histidine-rich residues (HX)n. Finally,CadA did not contain the four additional cysteine residues asCCCDGAC (position 36e42 aa on Fig. 2A) located 25-aa up-stream of the GXXCXXC motif compared to the ZntA proteinsequence from M. loti K12 (Fig. 2A).
3.3. Mutation of cadA reduces M. metallidurans Znresistance
The function of cadA was investigated by directed muta-genesis through partial deletion and replacement of the M.metallidurans cadA gene by a UKmR cassette. The construc-tion containing the partially deleted cadA gene with the UKmR
interposon was subcloned into suicide vector pJQ200SK andthe resulting plasmid was used to transform wild-type strainSTM 2683T. The deletion mutant (named DcadA) was ob-tained upon double homologous recombination and verified toconfirm proper mutagenesis. The growth of the DcadA strainon TY medium supplemented with Zn or Cd was compared tothat of the STM 2683T wild-type and the sensitive M. tian-shanense ORS 2740 strains as positive and negative controls,respectively. DcadA growth was inhibited in the presence of2 mM Zn (Fig. 3A). In the presence of 1.5 mM Zn, DcadAgrowth was reduced and delayed compared to the wild-type(Fig. 3C). The higher Zn sensitivity of the DcadA strain wasabolished when complemented by the full copy of the cadAgene under the control of the native promoter region (inpBBR1MCS-5), demonstrating that the disruption of cadAwasresponsible for reduced Zn resistance at higher concentrations(Fig. 3A) and for the delayed growth at 1.5 mM Zn (Fig. 3C).In contrast, DcadA growth was not affected by increasing Cdconcentrations (Fig. 3B).

Fig. 2. Description of CadA MBDs. Alignment ofM. metallidurans STM 2683T CadA MBDs with 13 PIB-type ATPases of rhizobia and relative strains, ZntA of E.
coli and CadA of S. aureus using ClustalX. The PIB-type ATPases sequences are named by their strain name: M. metallidurans STM 2683T; Mesorhizobium sp.
STM 4661; Chelativorans multitrophicus BNC1; Rhizobium leguminosarum sv. trifoliiWSM1325; R. leguminosarum sv. viciae 3841; R. leguminosarum sv. trifolii
WSM2304; R. etli CIAT 652; R. etli CFN 42; Agrobacterium radiobacter K84; Bradyrhizobium sp. BTAi1; Rhodopseudomonas palustris BisB5; M. huakuii
MAFF303099; E. coli K12 and S. aureus MRSA252.MBD motifs are framed, and conserved amino acid residues of interest are highlighted by a dark gray
background: (A) N-MBD in the N-terminal regions containing the GXXCXXC motif, (B) TM-MBD in the TM6 regions containing the CP(X)5STP motif and (C)
the signature sequence N(X)7K(X)20DXG(X)7N containing a lysine (K) in TM7 and an asparagine (N) and a glycine (G) located in TM8. Asterisks indicate
conserved amino acids and the consensus sequences are given.
181G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
3.4. cadA is specifically upregulated by Zn and Cd in adose-dependent manner and its mutation increasestranscriptional sensitivity to these metals
The cadA promoter region of STM 2683T was cloned up-stream of the gfp reporter gene in pPROBE-GT. The gfp-fusedcadA promoter (pGT-cadAp) as well as the promoter-lessvector (pGT) was electroporated into STM 2683T and intothe DcadA strain by electroporation. The transcriptional ac-tivity of cadA as assessed by gfp-induced fluorescence wasexamined in the presence of Zn, Cd, Co, Ni, Pb and Zn/Cdmixes. No fluorescence was detected in the presence of Ni(0.01, 0.02 and 0.03 mM), Co (0.01, 0.02 and 0.03 mM) or Pb(0.05, 0.1 and 0.2 mM) (data not shown). In contrast, thefluorescence levels obtained with pGT-cadAp evidenced
statistically significant induction levels as a result of exposureto increasing Zn or Cd concentrations (Fig. 4A and B). Basedon the observation that the expression level of cadAwas metal-specific and dose-dependent, the effect of the cadA mutationwas tested for its transcriptional activity. To do so, pGT-cadApwas transferred to DcadA and its transcriptional responsesupon Zn or Cd exposure were analyzed. The relative expres-sion of cadA in these strains and in media containing variousZn or Cd concentrations was recorded and the sigmoidaldoseeresponse curve we obtained was fitted to a non-linearregression model that allowed us to estimate the slope pa-rameters (Fig. 4C and D). Interestingly, in the DcadA strain,higher cadA expression was observed when compared with theSTM 2683T wild-type even when the bacteria were subjectedto low metal doses (Fig. 4C and D). Indeed, even without

Fig. 3. Effect of cadA deletion on Zn and Cd resistance inM. metallidurans. OD600 nm was measured after 1 week of growth to determine Zn (A) and Cd (B) MICs,
after 2, 3, 6 and 7 days of growth for the time-course assays (C). The data obtained for the M. metallidurans STM 2683T wild-type strain (black for A and B, filled
diamonds for C), the DcadA mutant (light gray for A and B, empty triangles for C), the DcadA mutant complemented with the original copy under the control of its
own promoter (DcadA-cadA strain, dark gray for A and B, filled triangles for C) and sensitive strain M. tianshanense (white for A and B, filled squares for C). The
means� SD of at least three replicates are indicated.
182 G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
adding metals, the basal cadA transcription level (corre-sponding to the background expression) in the DcadA strainwas more than sevenfold higher than in the wild-type (Table2). The mutant was more sensitive to both metals than thewild-type, as indicated by the lower EC50 values obtained inthe mutant (Table 2). The EC50 went down from 1.2 mM in thewild-type to 0.2 mM in the mutant for Zn and from 0.12 mM(wild-type) to 0.043 mM (mutant) for Cd (Table 2). Theseshifts corresponded to six and threefold increased transcrip-tional sensitivity of the mutant to Zn and Cd, respectively. Znions induced a higher (45-fold) maximal induction than Cd(20-fold) in the wild-type genomic background, whilemaximum induction was, respectively, eight and sixfold in themutant (Table 2). The maximum induction observed in themutant at the highest Zn and Cd concentrations may resultfrom saturated cadA transcriptional activity (Fig. 4C and D).Using the slope value fitted from the non-linear regressionmodel mentioned above, the Hill coefficient was found torange from 1.4 to 1.6 (Table 2) and was not affected either bystrain or metal, suggesting that the transcriptional responsewas not hypersensitive under the conditions tested. To bettercharacterize the regulatory effects of the inducing metals,cadA expression was monitored in the wild-type and DcadAstrains when exposed to various Zn and Cd mixes (Fig. 5). Bycomparing cadA relative expression in the mixes to those
obtained after summing up the corresponding metal concen-trations taken separately (Fig. 5), high correlation factors (R2)of over 0.99 and 0.94 for the wild-type and DcadA strainsrespectively, were found. This suggests that, despite differentinducing levels triggered by Zn or Cd, cadA transcriptionalinduction is cumulative.
3.5. cadA is induced in A. vulneraria nodules after Znstress
Roots of A. vulneraria were inoculated with M. metal-lidurans or the cadA-deleted derivative carrying the cadApromoter cloned upstream of gfp (into pGT) or the emptyvector. Nodulated plants were then subjected to Zn stress andcompared to control plants (without Zn). The mutated strain ofM. metallidurans was able to nodulate and fix nitrogen (basedon the leaf greenness) on A. vulneraria like the wild-type(Table S2). Neither the presence of pPROBE constructionsin Mesorhizobium nor Zn treatment had a significant effect onAnthyllis nodulation or shoot and root biomasses (Table S2).Nodules observed were pink-pigmented in all conditions,indicative of maturity and nitrogen fixation. One week of Znexposure time showed the highest intensity of fluorescencecompared to 24 or 48 h of contact. After a week of Znexposure, nodules formed by strains carrying plasmids as

Fig. 4. Effect of Zn and Cd on the transcriptional activity of cadA using the cadA promoter fused to a gfp construct (pGT-cadAp). cadA transcriptional levels (black)
induced by various concentrations of Zn (A) or Cd (B) in theM. metallidurans STM 2683Twild-type were compared to intensities of the promoter-less vector pGT
(white). Zn (C) and Cd (D) transcriptional response curves obtained for the wild-type strain (filled circles) and the DcadA strain (filled squares) are shown. The
parameters of the best-fitted curves (solid lines in C and D) are shown in Table 2. The relative expression values were obtained by subtracting the fluorescence
values of the non-inoculated control and by normalizing the fluorescence to the OD600nm. The means� SD of at least three replicates are indicated. Different letters
indicate statistically significant ( p-value <0.01) differences, while identical letters indicate no statistical difference (A, B).
183G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
described above were screened by epifluorescence micro-scopy. When M. metallidurans or the mutated strain containedthe cadAp::gfp fusion, more fluorescence was observed in theZn-supplemented condition compared to control Zn-freetreatment or to nodules produced by Mesorhizobium whichcarried the empty vector (Fig. 6). Furthermore, the fluores-cence intensity was relatively higher in nodules produced bythe cadA mutant when compared to the wild-type strain(Fig. 6C and G), which corroborates the in vitro inductionexperiment described above. This result suggests that at leastsome Zn enters into the nodule cells and 1 mM is sufficient toinduce cadA expression.
Table 2
Characteristics of cadA transcriptional activity. Characteristics of the response of
estimated from the best-fitted sigmoidal doseeresponse curve using non-linear reg
Strains Metal response EC50 (M)a Background fluores
STM 2683T Zn 1.20E�03 3511� 596
STM 2683T Cd 1.20E�04 3511� 596
DcadA Zn 2.00E�04 25642� 2130
DcadA Cd 4.30E�05 25642� 2130
a EC50 values were deduced from the best-fitted curves except for STM 2683T ab The mean background fluorescence corresponds to relative expression obtainedc Maximum-fold induction was calculated by dividing the mean of maximal relat
independent replications.d Hill coefficients were obtained from the best-fitted curves.
3.6. Metal resistance and metal efflux systems in thegenome of M. metallidurans STM 2683T
The STM 2683T genome contained six genes annotated asencoding PIB-type ATPases (Table 3). The PIB-type ATPasesclustered into two groups based on rather high identities toknown genes: a group containing cadA and zntA and a groupcontaining the MESS2_p120012 (the only plasmid-bornegene) and MESS2_1650024 genes. The two remaining geneswere actP and fixI, which were similar (33.3% identity). Thelatter appeared as the only Cu-efflux PIB-type ATPases of M.metallidurans. They were both located among a cluster of
STM 2683T and of the DcadA mutant to Zn and Cd exposure and parameters
ression analysis [56].
cence� SDb Max-fold inductionc Hill coefficientd R2
45.8 1.51 0.98
20.6 1.66 0.97
8.8 1.43 0.93
5.8 1.62 0.97
fter Zn exposure, for which we used the maximal relative expression.
in basal medium without addition of metals.
ive expression by the mean of minimal relative expression obtained from three

Fig. 5. Effect of Zn and Cd mixes on the transcriptional activity of cadA. The mean relative expression of cadA in STM 2683T (black) and in the DcadA strain
(gray) grown on various Zn and Cd mixes (A) was calculated from three replicates. Bars indicate means� SD. The relative expression values observed from the
mixed metal solutions (X axis) were plotted against the relative expression values expected by adding the corresponding relative expression values obtained from
single metal solutions (Y axis). They are shown for STM 2683T (B) and for the DcadA strain (C). OD600nm and fluorescence intensity were measured after 2 days of
growth. The relative expression values were obtained by subtracting the normalized data corresponding to the pGT-cadAp constructs from those obtained with the
promoter-less pGT vectors. Finally, to test whether the relative expression in Zn and Cd mixes was correlated with the sum of the relative expression value in single
metal assays (B and C), normalized relative expression values observed in the presence of metal were corrected by expression values obtained in TY control
medium.
184 G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
housekeeping genes. Among these six PIB-type ATPase genes,only zntA displayed one upstream regulatory gene (calledzntR) belonging to the MerR-family transcriptional regulator.For cadA, no synteny was observed with any regulatory gene,nor with a gene involved in gene mobility. Yet cadA was insynteny with a ccdA-like gene (MESS2_740029) that wasannotated as involved in the biogenesis of cytochrome c-type.When looking for other putative metal-processing genes,copCD genes (MESS2_740005-06) and HME-RND cusBAgenes (MESS2_730254 and MESS2_730253) putativelyinvolved in Cu homeostasis were identified. The cus operonlacked a czcC/cusC-like gene that was replaced by a smallgene named cusG (MESS2_730255). Additionally, a CDFfamily czcD-like gene (MESS2_730268-69) involved in Co,Zn and Cd export was identified in the STM 2683T genome
near the cus genes and the ccdA-cadA operon. Finally an Hgresistance locus containing five genes with the structure merR/merTPCA (MESS2_30049-53) was identified not far from thetraG/fixIS cluster.
4. Discussion
We report here the first mesorhizobial PIB-type ATPaseinvolved in Zn and Cd transport in M. metallidurans STM2683T, a nitrogen-fixing bacterium isolated from root nodulesof the metallicolous legume A. vulneraria [68]. The proteinsequence of this enzyme, named CadA, is similar to the best-characterized PIB-type ATPase efflux transporters, i.e. ZntA[11] and CadA [51] which promote Zn and Cd detoxificationvia cytoplasmic efflux. Such metal efflux proteins have been

Fig. 6. Expression of cadA in A. vulneraria nodules after Zn stress. Comparison of A. vulneraria nodule sections infected with STM 2683 pGT (A, B), STM 2683
pGT-cadAp (C, D), DcadA pGT (E, F) and DcadA pGT-cadAp (G, H), under Zn-supplemented conditions (A, C, E, G) and without Zn supplement (B, D, F, H).
Fluorescence obtained with an epifluorescence microscope using GFP filter after one week of Zn exposure. Bars correspond to 0.5 mm.
185G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
identified in C. metallidurans strain CH34 that is a multiple-metal-resistant bacterium isolated from the sludge of a Zndecantation tank in Belgium [43]. Moreover, CadA displayshigh identity with other rhizobial PIB-type ATPases such asPIB-type ATPase of the metallicolous Mesorhizobium sp. STM4661 (97% identity on the full sequence length) that wasisolated from a ZnePb mine [42] and the PIB-type ATPase of anon-metallicolous Ensifer meliloti (63% identity) which isshorter than CadA and that was functionally shown to play acrucial role in defense against Cd and Zn stress [60]. CadA isalso closely related to the PIB-type ATPase of Chelativoransmultitrophicus BNC1, an EDTA-degrading strain [21]. CadA
Table 3
Genomic information from M. metallidurans STM 2683T about the six genes anno
Gene labela
(accession number)
Gene productb Protein
size (aa)
Identity/
CadA (%)
MESS2_740030
(HE820903)
Heavy metal P-type ATPase/CadA 910 100
MESS2_10039
(HE965426)
Copper transport ATPase/FixI 760 31
MESS2_1050036
(HE965427)
Copper-transporting P-type
ATPase/ActP
834 40
MESS2_980073
(HE965428)
Zinc, cobalt and lead efflux/ZntA 748 67.15
MESS2_1650024
(HE965430)
Cadmium-exporting ATPase 822 38.01
MESS2_p120012
(HE965429)
Heavy metal P-type ATPase 842 35.29
The PIB-type ATPases listed in the table shared at least 30% identity with CadA.a Genes named by gene labels from MaGe.b Gene products deduced from automatic or manual (only cadA) annotations.c Not detected.
sequence structure indicates that CadA is involved in metalresistance and it conferred higher resistance to M. metal-lidurans during its growth at high Zn concentrations. Inaddition, cadA was specifically induced by Zn and Cd in vitroconditions and in A. vulneraria nodules after Zn stress, sup-porting a metal resistance role in free-living and symbiotic M.metallidurans. CadA displays all the key features of the PIB-2subgroup of ATPases involved in pump functioning and metalresistance activities [5,7]. Like ZntA of E. coli, the N-MBD ofCadA contains motifs involved in the binding of divalent ions,but not of monovalent ions such as Cuþ or Agþ [9]. However,CadA is structurally different from ZntA because it does not
tated as encoding PIB-type ATPases.
Closely linked Other linked genes of interest
NDc ccdA (MESS2_740029) Putative Zn transporter
(MESS2_740027)
ND 25-gene block glmS/traG/copG/trbBCDEJLFGI/
gir/hemN/fnrN/fix
NOPGHIS are conserved in five Mesorhizobium
genomes
ND Five genes mutS/yeaE/glnD/mviN/trpS are
conserved in five Mesorhizobium genomes
zntR (MESS2_980074) Unknown
ND Possible deleted remnant of a MerR regulator
seen in H. arsenicooxydans (MESS2_1650027)
deleted insertion sequence (IS) (MESS2_1650029)
ND Fragment of dsbA reductase (MESS2_120011)
fragment of transposase (MESS2_120014)

186 G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
contain the four additional cysteine residues upstream of theGXXCXXC motif which have been described as necessary forbinding Pb2þ in addition to Zn2þ and Cd2þ [38]. In our study,CadA seemed to bind Zn2þ/Cd2þ but not Pb2þ. Moreover, thelatter result was in line with the Pb-sensitive phenotypeidentified in the E. coli subclone CdE1. CadA from M. met-allidurans showed the special feature of containing oneduplication of the two N-MBDs (i.e. a GXXCXX motif (HMAdomain) followed by a (HX)n domain), whereas most of thePIB-2-type ATPases contain a unique GXXCXXC motif and/ora unique (HX)n domain. This characteristic could only bedetected in three previously described metal-resistant bacterialtypes: a Zn-resistant Agrobacterium tumefaciens strain [29],the multi-resistant strains of Brucella melitensis [19] andYersinia pestis, both isolated from clinical environments [48].These more numerous duplications of N-MBDs suggestsspecific adaptation to metal-contaminated environments.
According to functional analyses, STM 2683T cadA geneconfers Zn and Cd resistance upon E. coli. InM. metallidurans,cadA was proven to be important for tolerance to high Znconcentrations, whereas its involvement in Cd resistance couldnot be detected during growth. Since Cd is more toxic than Zn,we hypothesize that the metallicolous bacteria were able tomobilize other genes involved in the efflux or sequestration ofmetals, such as those identified using a transcriptome approach[42]. Indeed, the phenotypic differences between the two cadAtransformants can be explained by the different genetic back-grounds of the two host strains, which show contrasting prop-erties as far as metal resistance is concerned. Unlike thelaboratory strain of E. coli, STM 2683T is well adapted to sur-viving and thriving in a soil environment contaminated withhigh levels of metals [40] and may contain a larger geneticarsenal for tolerating toxic metals. Such genes involved in metaladaptation can belong to efflux systems or to metal sequestra-tion and detoxification processes. In the automatically anno-tated genome of STM 2683T [42] and in addition to CadA, fiveother PIB-type ATPases were identified and shared between 31and 67% identity with CadA, including a ZntA homologinvolved in Zn export [59], ActP and FixI proteins involved inCu export [25,58] and metal-exporting ATPases for whichsubstrates are not specified. Only cadA and zntA were signifi-cantly induced by Zn and Cd according to the transcriptionalresponses of M. metallidurans to metal exposure [42]. Along-side these PIB-type ATPases, putative CDF family genes (czcD-like), as previously described in C. metallidurans [4], and oneHME-RND-type representative with homologies to CusGBA,which is conserved in various Alpha-Proteobacteria, wereregulated in the STM 2683T upon metal stress. By comparingthree available metallicolous genomes of Mesorhizobium spe-cies (M. metallidurans STM 2683T, Mesorhizobium sp. STM4661 and M. amorphae CCNWGS0123) to four non-metallicolous genomes (M. ciceri sv. biserrulae WSM 1271,M. opportunistum WSM 2075, M. huakuii MAFF303099 andM. australicum WSM 2073), a higher number of genesencoding PIB-type ATPases was present in metallicolous Mes-orhizobium (6e8 genes) than in non-metallicolous Meso-rhizobium (2e4 genes). However, the overrepresentation of
PIB-type ATPases is not only described in metallicolous Mes-orhizobium species. As shown in C. metallidurans [69] andBurkholderia species [62], the PIB-type ATPase system seemsto be a major mechanism used by metal-resistant bacteria tocounteract metal toxicity. Our results indicate that cadA alonedoes not confer complete metal resistance upon M. metal-lidurans. Indeed, in addition to efflux systems, other mecha-nisms allowing bacteria to tolerate high Zn or Cd levels couldparticipate in bacterial tolerance to these metals. Transcrip-tional analyses of M. metallidurans after metal exposureshowed that several genes involved in the biosynthesis ofvarious intracellular, periplasmic and cell-wall proteins orpolysaccharides were differentially regulated and couldpossibly be involved inmetal sequestration or reduction in orderto limit metal bioavaibility [42]. Complete resistance probablyrelies on several systems that interact in an as-yet-undefinedway. The combination of these metal tolerance systems withina single strain was previously described in C. metalliduransCH34 [46,65] and altogether guarantees maximal metal resis-tance. Nevertheless, cadA was one of the top-induced genesafter Cd and Zn stresses ofM.metallidurans [42], confirming itskey role in the metal response of M. metallidurans.
The analysis of the promoter region upstream of the cadAgene revealed a regulatory consensus site previously describedin other rhizobia [34], which is recognized by a MerR-familytranscriptional regulator. MerR-family proteins regulatedetoxification systems of several metals, including Zn and Cd[14]. This suggests that cadA is also regulated by a MerR-family transcriptional regulator. Transcriptional analyses ofcadA revealed that it was specifically induced by Zn2þ andCd2þ. Our data showed that Cd activated cadA transcription atlower concentrations when compared to Zn. This could beacceptable for a toxic and non-essential metal like Cd, whichis toxic even at low concentrations. The higher sensitivity tometals observed in the DcadA mutant, as estimated by EC50
shifts, suggests that in the DcadA strain, higher intracellularmetal concentrations accumulate and could result from lowerefflux efficiency caused by cadA deficiency. This hypothesissupports transcriptional analyses of M. metallidurans to Znand Cd exposure [42]. The latter study showed that theznuABC homologs (MESS2_300037-39) encoding the high-affinity ABC-type Zn-import system were strongly repressedby Zn in the cadA-deleted strain, suggesting the key role ofcadA in Zn efflux. In the absence of cadA, metals present inthe culture media were sufficient to induce its transcription,suggesting that this transporter is functional in metal export, asdescribed previously for ZntA of E. coli [12]. Additionally,when we compared the expression levels obtained when bothinducing metals were present in the growth medium to thoseobtained in silico (by summing up the relative expressionsinduced by the same amounts of metals when added separatelyto the medium), the rate of cadA induction was found to becumulative for the metal concentrations tested. Such experi-ments indicate that Zn- and Cd-mediated cadA transcriptionmight be under the control of specific regulatory complexeswhich do not compete, at least not under the conditions tested.Complex regulation involving two distinct Mer regulators

187G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
(MerR/MerD) which specifically bind to the same DNAoperator site upstream of the mer operon has been described[47]. In the genome of STM 2683T, no regulatory gene wasidentified in synteny with the cadA gene, in contrast to zntA(MESS2_980073) for which a gene encoding the merR-typeregulator (zntR, MESS2_980074) was identified. Only a ccdA-like gene encoding cytochrome c-type biogenesis can beidentified in synteny with cadA. This ccdA-like gene displayssome similarities with C. metallidurans copT and a part ofpbrT that is adjacent to pbrA which codes an efflux Pb-effluxPIB-type ATPase [13] and was renamed cadT. In fact, bothcadT and cadA orthologs are observed in a four- or five-genestructures in different Methylobacter strains, Actinobacteriaand in Arthrobacter sp. strain FP24 (which contained threecopies of this structure and which also possesses various genesinvolved in resistance to heavy metals, especially chromate).The Methylobacter and Arthrobacter genes that are closelylinked to M. metallidurans cadT and cadA orthologs includedat least a regulatory gene (family ArsR or MerR), a peptidasegene and a transporter. This suggests interactions and syn-ergies between PIB-type ATPases and other systems, as alreadydescribed in C. metallidurans CH34 [37]. The authorsdemonstrated interplay of the Czc system and two PIB-typeATPases (ZntA and CadA) that could allow C. metalliduransto survive in highly metal-contaminated environments. TwoPIB-type ATPases seem to be necessary to Czc system functionin order to establish full resistance to metals. CadA regulationexerted via unlinked genes remains to be investigated.
To gain insight into the role of cadA during symbiosis,plants of A. vulneraria nodulated by wild-type strain STM2683T as well as the cadA mutant that was carrying thecadAp::gfp reporter system were subjected to Zn stress. Ourresults showed that cadA is expressed in infected nodule cellsafter Zn stress, suggesting that Zn could enter into the plantcells, which corroborates a recent study where higher Zncontents were measured in nodules of Anthyllis plants exposedto 1 mM compared to plants exposed to 0 or 2.5 mM Zn [64].Chen et al. [17] demonstrated that after nodulation by a metal-resistant Cupriavidus strain, the metal uptake abilities ofMimosa pudica plants markedly increased and were 12, 70 and86 % higher for Cu, Cd and Pb, respectively, than those ofnodule-free plants. Metal accumulation in the nodulated plantsoccurred mainly in the roots (65e95% of total metal uptake),whereas the shoots contributed to only 2e23% of total metaluptake. Also, Soussou et al. [64] showed a reduction of 2.5times in Zn concentration in shoots of Anthyllis when inocu-lated with M. metallidurans after 1 mM Zn stress. These re-sults suggest that nodules play an important role in metalbalance in plant roots and its transfer to aerial tissues. Chenet al. [17] argued that such accumulation of metals byCupriavidus strains within nodules of M. pudica could resultin (i) an increase in metal binding sites through physiologicalchanges of bacteroid cells or increased release in extracellularproducts like exopolysaccharides, or (ii) an alteration of metalmobility in nodulated plants, as metals can be sequestrated byplant-specific mechanisms [18]. Additional studies are neededto elucidate the speciation and localization of Zn within
nodules in order to better understand the role played by nod-ules in plant-metal adaptation.
In conclusion, we have identified the first mesorhizobialgene coding PIB-2-type ATPase, named CadA. The gene cadAof the tolerant M. metallidurans type strain improved Zn andCd tolerance in E. coli. Although CadA itself is not sufficientto explain how M. metallidurans can be adapted and evenenter into symbiosis in highly contaminated soils such as thosefrom which it originates, mutagenesis analyses showed thatCadA is functional and plays a key role in metal tolerance ofthis bacterium, especially in the presence of high Zn con-centrations. This is especially of interest from the perspectiveof plantemicrobe interactions and phytoremediation of highlycontaminated mining sites in the south of France. In this re-gard, M. metallidurans promotes the growth of the metal-tolerant A. vulneraria ecotype, which is currently beingstudied to assess its potential as a key phytostabilization agentin the Avinieres mining site. CadA is present in anothermetallicolous Mesorhizobium strain (STM 4661) associatedwith A. vulneraria which was also isolated from a ZnePbmine (Eylie mine, Pyrenees ariegeoises), supporting its role inmetal resistance. cadA is regulated specifically and in a dose-dependent manner by Zn and Cd divalent cations in M. met-allidurans in pure cultures and is also induced by Zn in nod-ules of A. vulneraria after Zn stress. In the cadA-deleted strain,metal induction sensitivity was probably increased because ofintracellular metal accumulation resulting from lower effluxefficiency, which supports the function of CadA as a metalexport system. The atypical feature of CadA and its involve-ment in metal resistance may enable the definition of abiomarker which could lead to the study of the distribution ofcadA among metal-tolerant Anthyllis mesorhizobia pop-ulations. Finally, cadA could also be used to follow the fate ofintroduced metal-resistant strains used as inocula to improvephytostabilization strategies initiated by Anthyllis settlementin association with non-fixing plant species.
Acknowledgments
This work was supported by the Agence Nationale de laRecherche Programme “Contaminants, Ecosystemes et Sante”(CES 2008-12, APTITUDE). G.M. received a PhD grant fromthe Ministere de la Recherche et de l’Education Nationale. Wethank Dr. C. Rensing for sending the GG48 strain and Dr. S.Soussou for advice on Anthyllis cultures.
Appendix A. Supplementary data
Supplementary data related to this article can be found athttp://dx.doi.org/10.1016/j.resmic.2014.02.001.
References
[1] AikingH,KokK, vanHeerikhuizenH, van ’tRiet J. Adaptation to cadmium
byKlebsiella aerogenes growing in continuous culture proceedsmainly via
formation of cadmium sulfide. Appl Environ Microbiol 1982;44:938e44.

188 G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
[2] Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local
alignment search tool. J Mol Biol 1990;215:403e10.
[3] Andreini C, Bertini I, Cavallaro G, Holliday G, Thornton J. Metal ions in
biological catalysis: from enzyme databases to general principles. J Biol
Inorg Chem 2008;13:1205e18.[4] Anton A, Grobe C, Reibmann J, Pribyl T, Nies DH. CzcD is a heavy
metal ion transporter involved in regulation of heavy metal resistance in
Ralstonia sp. strain CH34. J Bacteriol 1999;181:6876e81.[5] Arguello JM. Identification of ion-selectivity determinants in heavy-
metal transport P1B-type ATPases. J Membr Biol 2003;195:93e108.
[6] Arguello J, Eren E, Gonzalez-Guerrero M. The structure and function of
heavy metal transport P1B-ATPases. BioMetals 2007;20:233e48.[7] Axelsen KB, Palmgren MG. Evolution of substrate specificities in the P-
type ATPase superfamily. J Mol Evol 1998;46:84e101.
[8] Bal N, Mintz E, Guillain F, Catty P. A possible regulatory role for the
metal-binding domain of CadA, the Listeria monocytogenes Cd2þ-ATPase. FEBS Lett 2001;506:249e52.
[9] Banci L, Bertini I, Ciofi-Baffoni S, Su XC,Miras R, Bal N, et al. Structural
basis for metal binding specificity: the N-terminal cadmium binding
domain of the P1-type ATPase CadA. J Mol Biol 2006;356:638e50.
[10] Bauer DW, Collmer A. Molecular cloning, characterization and muta-
genesis of a pel gene from Pseudomonas syringae pv. lachrymans
encoding a member of the Erwinia chrysanthemi PelADE family of
pectate lyases. Mol Plant Microbe Interact 1997;10:369e79.
[11] Beard SJ, Hashim R, Membrillo-Hernandez J, Hughes MN, Poole RK.
Zinc(II) tolerance in Escherichia coli K-12: evidence that the zntA gene
(o732) encodes a cation transportATPase.MolMicrobiol 1997;25:883e91.[12] Binet MRB, Poole RK. Cd(II), Pb(II) and Zn(II) ions regulate expression
of the metal-transporting P-type ATPase ZntA in Escherichia coli. FEBS
Lett 2000;473:67e70.[13] Borremans B, Hobman JL, Provoost A, Brown N, van der Lelie D.
Cloning and functional analysis of the pbr lead resistance determinant of
Ralstonia metallidurans CH34. J Bacteriol 2001;183:5651e8.
[14] Brown NL, Stoyanov JV, Kidd SP, Hobman JL. The MerR family of
transcriptional regulators. FEMS Microbiol Rev 2003;27:145e63.
[15] Bruins MR, Kapil S, Oehme FW. Microbial resistance to metals in the
environment. Ecotoxicol Environ Saf 2000;45:198e207.
[16] Carrasco JA, Armarioc P, Pajueloa E, Burgosa A, Caviedesc MA,
Lopez R, et al. Isolation and characterization of symbiotically effective
Rhizobium resistant to arsenic and heavy metals after the toxic spill at the
Aznalcollar pyrite mine. Soil Biol Biochem 2005;37:1131e40.
[17] Chen W-M, Wu C-H, James EK, Chang J-S. Metal biosorption capability
of Cupriavidus taiwanensis and its effects on heavy metal removal by
nodulated Mimosa pudica. J Hazard Mater 2008;151:364e71.
[18] Clemens S. Toxic metal accumulation, responses to exposure and
mechanisms of tolerance in plants. Biochimie 2006;88:1707e19.
[19] DelVecchio VG, Kapatral V, Redkar RJ, Patra G, Mujer C, Los T, et al.
The genome sequence of the facultative intracellular pathogen Brucella
melitensis. Proc Natl Acad Sci U S A 2002;99:443e8.[20] Diels L, Dong Q, van der Lelie D, Baeyens W, Mergeay M. The czc
operon of Alcaligenes eutrophus CH34: from resistance mechanism to
the removal of heavy metals. J Ind Microbiol 1995;14:142e53.
[21] Doronina NV, Kaparullina EN, Trotsenko YA, Nortemann B, Bucheli-
Witschel M, Weilenmann HU, et al. Chelativorans multitrophicus gen.
nov., sp. nov. and Chelativorans oligotrophicus sp. nov., aerobic, EDTA-
degrading bacteria. Int J Syst Evol Microbiol 2010;60:1044e51.[22] Durfee T, Nelson R, Baldwin S, Plunkett G, Burland V, Mau B, et al. The
complete genome sequence of Escherichia coli DH10B: insights into the
biology of a laboratory workhorse. J Bacteriol 2008;190:2597e606.
[23] Figurski DH, Helinski DR. Replication of an origin-containing derivative
of plasmid RK2 dependent on a plasmid function provided in trans. Proc
Natl Acad Sci U S A 1979;78:1648e52.
[24] Frerot H, Lefebvre C, Gruber W, Collin C, Dos Santos A, Escarre J.
Specific interaction between local metallicolous plants improve the
phytostabilization of mine soils. Plant Soil 2006;282:53e65.
[25] Gonzalez-Guerrero M, Raimunda D, Cheng X, Arguello JM. Distinct
functional roles of homologous Cuþ efflux ATPases in Pseudomonas
aeruginosa. Mol Microbiol 2010;78:1246e58.
[26] Grass G, Fan B, Rosen BP, Franke S, Nies DH, Rensing C. ZitB (YbgR),
a member of the cation diffusion facilitator family, is an additional zinc
transporter in Escherichia coli. J Bacteriol 2001;183:4664e7.
[27] Hakkila KM, Nikander PA, Junttila SM, Lamminmaki UJ, Virta MP. Cd-
specific mutants of mercury-sensing regulatory protein MerR, generated
by directed evolution. Appl Environ Microbiol 2011;77:6215e24.
[28] Hall T. BioEdit: biological sequence alignment editor for Win95/98/NT/
2K/XP. Web site last modified on June 27, 2007. Available at: www.
mbio.ncsu.edu/BioEdit/bioedit.html; 2007 [accessed 13.09.11].
[29] Hao X, Xie P, Johnstone L, Miller SJ, Rensing C, Wei G. Genome
sequence and mutational analysis of plant-growth-promoting bacterium
Agrobacterium tumefaciens CCNWGS0286 isolated from a zinc-lead
mine tailing. Appl Environ Microbiol 2012;78:5384e94.
[30] Hassan M-e-T, van der Lelie D, Springael D, Romling U, Ahmed N,
Mergeay M. Identification of a gene cluster, czr, involved in cadmium and
zinc resistance in Pseudomonas aeruginosa. Gene 1999;238:417e25.[31] Hayashi M, Maeda Y, Hashimoto Y, Murooka Y. Efficient transformation
of Mesorhizobium huakuii subsp. rengei and Rhizobium species. J Biosci
Bioeng 2000;89:550e3.[33] Kampa M, Castanas E. Human health effects of air pollution. Environ
Pollut 2008;151:362e7.
[34] Kazakov AE, Kalinina OV, Permina EA, Gelfand MS. Bacterial metal
resistance systems regulated by transcription regulators of the merR
family. In: Proceedings of the fourth international conference on bioin-
formatics genome regulation structure, 25e30 July 2004; Novosibirsk,
Russia; 2004.
[35] Klaassen CD, Liu J, Choudhuri S. Metallothionein: an intracellular
protein to protect against cadmium toxicity. Annu Rev Pharmacol Tox-
icol 1999;39:267e94.
[36] Kovach ME, Elzer PH, Hill DS, Robertson GT, Farris MA, Roop RM,
et al. Four new derivatives of the broad-host-range cloning vector
pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene
1995;166:175e6.
[37] Legatzki A, Grass G, Anton A, Rensing C, Nies DH. Interplay of the Czc
system and two P-type ATPases in conferring metal resistance to Ral-
stonia metallidurans. J Bacteriol 2003;185:4354e61.
[38] Liu J, Stemmler AJ, Fatima J, Mitra B. Metal-binding characteristics of
the amino terminal domain of ZntA binding of lead is different compared
to cadmium and zinc. Biochemistry 2005;44:5159e67.
[39] Mana-Capelli S, Mandal AK, Arguello J. Archaeoglobus fulgidus CopB
is a thermophilic Cu2þ-ATPase. J Biol Chem 2003;278:40534e41.
[40] Mahieu S, Frerot H, Vidal C, Galiana A, Heulin K, Maure L, et al.
Anthyllis vulneraria/Mesorhizobium metallidurans, an efficient symbi-
otic nitrogen fixing association able to grow in mine tailings highly
contaminated by Zn, Pb and Cd. Plant Soil 2011;342:405e17.
[41] Mandal AK, Arguello JM. Functional roles of metal binding domains of
the Archaeoglobus fulgidus Cuþ-ATPase CopA. Biochemistry
2003;42:11040e7.
[42] Maynaud G, Brunel B, Mornico D, Durot M, Severac D, Dubois E, et al.
Genome-wide transcriptional responses of two metal-tolerant symbiotic
Mesorhizobium isolates to zinc and cadmium exposure. BMC Genomics
2013;14:292.
[43] Mergeay M, Nies D, Schlegel HG, Gerits J, Charles P, Van Gijsegem F.
Alcaligenes eutrophus CH34 is a facultative chemolithotroph with
plasmid-bound resistance to heavy metals. J Bacteriol 1985;162:328e34.
[44] MillerWG,Leveau JHJ,LindowSE. Improvedgfpand inaZbroad-host-range
promoter-probe vectors. Mol Plant Microbe Interact 2000;13:1243e50.
[45] Misra TK. Bacterial resistances to inorganic mercury salts and organo-
mercurials. Plasmids 1992;27:4e16.
[46] Monchy S, Benotmane MA, Janssen P, Vallaeys T, Taghavi S, van der
Lelie D, et al. Plasmids pMOL28 and pMOL30 of Cupriavidus metal-
lidurans are specialized in the maximal viable response to heavy metals.
J Bacteriol 2007;189:7417e25.
[47] Mukhopadhyay D, Yu HR, Nucifora G, Misra TK. Purification and
functional characterization of MerD. A coregulator of the mercury resis-
tance operon in gram-negative bacteria. J Biol Chem 1991;266:18538e42.
[48] Neyt C, Iriarte M, Thi VH, Cornelis GR. Virulence and arsenic resistance
in Yersiniae. J Bacteriol 1997;179:612e9.

189G. Maynaud et al. / Research in Microbiology 165 (2014) 175e189
[49] Nicholas KB, Nicholas HB. GeneDoc: a tool for editing and annotating
multiple sequence alignments. Distributed by the author, http://www.psc.
edu/biomed/genedoc; 1997.
[50] Nies DH. Efflux-mediated heavy metal resistance in prokaryotes. FEMS
Microbiol Rev 2003;27:313e39.[51] Nucifora G, Chu L, Misra TK, Silver S. Cadmium resistance from
Staphylococcus aureus plasmid pI258 cadA gene results from a
cadmium-efflux ATPase. Proc Natl Acad Sci U S A 1989;86:3544e8.[52] Nwuche CO, Ugoji EO. Effects of metal pollution on the soil microbial
activity. Int J Environ Sci Technol 2008;5:409e14.
[54] Papoyan A, Pineros M, Kochian LV. Plant Cd2þ and Zn2þ status effects
on root and shoot heavy metal accumulation in Thlaspi caerulescens.
New Phytol 2007;175:51e8.
[56] Ralston DM, O’Halloran TV. Ultrasensitivity and heavy-metal selectivity
of the allosterically modulated MerR transcription complex. Proc Natl
Acad Sci U S A 1999;87:3846e50.[57] Rapisarda VA, Chehın RN, De Las Rivas J, Rodrıguez-Montelongo L,
Farıas RN, Massa EM. Evidence for Cu(I)-thiolate ligation and predic-
tion of a putative copper-binding site in the Escherichia coli NADH
dehydrogenase-2. Arch Biochem Biophys 2002;405:87e94.
[58] Reeve WG, Tiwari RP, Kale NB, Dilworth MJ, Glenn AR. ActP controls
copper homeostasis in Rhizobium leguminosarum bv. viciae and Sino-
rhizobium meliloti preventing low pH-induced copper toxicity. Mol
Microbiol 2002;43:981e91.
[59] Rensing C, Mitra B, Rosen BP. The zntA gene of Escherichia coli en-
codes a Zn(II)-translocating P-type ATPase. Proc Natl Acad Sci U S A
1997;94:14326e31.[60] Rossbach S, Mai DJ, Carter EL, Sauviac L, Capela D, Bruand C, et al.
Response of Sinorhizobium meliloti to elevated concentrations of cad-
mium and zinc. Appl Environ Microbiol 2008;74:4218e21.[61] Sambrook J, Fritschi EF, Maniatis T. Molecular cloning: a laboratory
manual. New York: Cold Spring Harbor Laboratory Press; 1989.
[62] Schwager S, Lumjiaktase P, Stockli M, Weisskopf L, Eberl L. The ge-
netic basis of cadmium resistance of Burkholderia cenocepacia. Environ
Microbiol Rep 2012;4:562e8.
[63] Sharma R, Schimke R. Preparation of electrocompetent E. coli using salt-
free growth medium. BioTechniques 1996;20:42e4.
[64] Soussou S, Mahieu S, Brunel B, Escarre J, Lebrun M, Banni M, et al.
Zinc accumulation patterns in four Anthyllis vulneraria subspecies sup-
plemented with mineral nitrogen or grown in the presence of their
symbiotic bacteria. Plant Soil 2013;371:423e34.
[65] Taghavi S, Lesaulnier C, Monchy S, Wattiez R, Mergeay M, van der
Lelie D. Lead(II) resistance in Cupriavidus metallidurans CH34: inter-
play between plasmid and chromosomally-located functions. Antonie
Van Leeuwenhook 2009;96:71e182.
[66] Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S.
MEGA5: molecular evolutionary genetics analysis using maximum
likelihood, evolutionary distance, and maximum parsimony methods.
Mol Biol Evol 2011;28:2731e9.
[67] Tusnady GE, Simon I. The HMMTOP transmembrane topology predic-
tion server. Bioinformatics 2001;17:849e50.[68] Vidal C, Chantreuil C, Berge O, Maure L, Escarre J, Bena G, et al.
Mesorhizobium metallidurans sp. nov., a metal-resistant symbiont of
Anthyllis vulneraria growing on metallicolous soil in Languedoc, France.
Int J Syst Evol Microbiol 2009;59:850e5.[69] von Rozycki T, Nies D. Cupriavidus metallidurans: evolution of a metal-
resistant bacterium. Antonie Van Leeuwenhook 2009;96:115e39.
[70] Wuana RA, Okieimen FE. Heavy metals in contaminated soils: a review
of sources, chemistry, risks and best available strategies for remediation.
ISRN Ecol Art 2011;2011:1e20.
[71] Zhang JH, Min H. Characterization of a multimetal resistant Bur-
kholderia fungorum isolated from an e-waste recycling area for its po-
tential in Cd sequestration. World J Microb Biotechnol 2010;26:371e4.





























