Embed Size (px)
Citation preview

ORIGINAL PAPER
Capsaicin Modulates Proliferation, Migration,and Activation of Hepatic Stellate Cells
Shanna Bitencourt • Fernanda Mesquita • Bruno Basso • Julia Schmid •
Gabriela Ferreira • Lucas Rizzo • Moises Bauer • Ramon Bartrons •
Francesc Ventura • Jose Luis Rosa • Inge Mannaerts •
Leo Adrianus van Grunsven • Jarbas Oliveira
� Springer Science+Business Media New York 2013
Abstract Capsaicin, the active component of chili pep-
per, has been reported to have antiproliferative and anti-
inflammatory effects on a variety of cell lines. In the cur-
rent study, we aimed to investigate the effects of capsaicin
during HSC activation and maintenance. Activated and
freshly isolated HSCs were treated with capsaicin. Prolif-
eration was measured by incorporation of EdU. Cell cycle
arrest and apoptosis were investigated using flow cytome-
try. The migratory response to chemotactic stimuli was
evaluated by a modified Boyden chamber assay. Activation
markers and inflammatory cytokines were determined by
qPCR, immunocytochemistry, and flow cytometry. Our
results show that capsaicin reduces HSC proliferation,
migration, and expression of profibrogenic markers of
activated and primary mouse HSCs. In conclusion, the
present study shows that capsaicin modulates proliferation,
migration, and activation of HSC in vitro.
Keywords Hepatic stellate cell � Capsaicin �Proliferation � Migration � Activation
Introduction
The recruitment of HSCs to sites of injury is thought to be
an early step in their function in repair and matrix
remodeling during liver fibrosis. These cells undergo a
process of activation headed for a phenotype characterized
by increased proliferation, motility, contractility, and syn-
thesis of ECM components. Stimulation of HSCs is regu-
lated by several soluble factors, including cytokines,
chemokines, and growth factors [1]. HSC activation is the
major contributor for the development of hepatic fibrosis.
Based on this knowledge, antifibrotic therapies for the liver
could be based on the inhibition or reversion of HSC
activation [2, 3]. Triggering the activity of interstitial col-
lagenases, such as matrix metalloproteinases (MMPs),
could be also an attractive strategy to deactivate HSC from
the view point that the imbalance between MMPs and
tissue inhibitors of metalloproteinases (TIMPs) is the main
obstacle to the reversion of activated HSC [4].
Previously published work demonstrates the efficacy of
natural products for the treatment of hepatic fibrosis. The
use of nutraceuticals that can inhibit the proliferation of
activated HSCs has a great potential in reversing fibrosis
[5]. Capsaicin, a naturally occurring phytochemical, is the
major ingredient of hot peppers and it is well known to have
anti-inflammatory and antiproliferative properties [6–8].
Leo Adrianus van Grunsven and Jarbas Oliveira have contributed
equally to this study.
S. Bitencourt � F. Mesquita � B. Basso � J. Schmid �G. Ferreira � J. Oliveira (&)
Laboratorio de Pesquisa em Biofısica Celular e Inflamacao,
Pontifıcia Universidade Catolica do Rio Grande do Sul
(PUCRS), Avenida Ipiranga 6681, predio 12, bloco C, sala 221,
Porto Alegre, RS CEP 90619-900, Brazil
e-mail: [email protected]
S. Bitencourt � I. Mannaerts � L. A. van Grunsven
Liver Cell Biology Lab, Department of Cell Biology,
Vrije Universiteit Brussel, Brussels, Belgium
L. Rizzo � M. Bauer
Laboratorio de Imunologia do Envelhecimento, Instituto de
Pesquisas Biomedicas, Pontifıcia Universidade Catolica do Rio
Grande do Sul, Porto Alegre, RS, Brazil
R. Bartrons � F. Ventura � J. L. Rosa
Departament de Ciencies Fisiologiques II, Campus de Bellvitge,
Universitat de Barcelona, L’Hospitalet de Llobregat, Barcelona,
Spain
123
Cell Biochem Biophys
DOI 10.1007/s12013-013-9719-0

In order to clarify the mechanism underlying the anti-
proliferative effects of capsaicin, we investigated whether
capsaicin alters cell cycle or induces programmed cell
death and examined its effect on cell migration and acti-
vation of HSCs, which play key roles in liver fibrogenesis.
Here, we show that capsaicin reduces cell proliferation
through cell cycle arrest, regulates profibrogenic/anti-
fibrogenic molecules balance, and inhibits activation and
cell migration in mouse HSCs.
Methods
Cell Culture and Treatment
The murine HSC line GRX was obtained from the Rio de
Janeiro Cell Bank (HUCFF, UFRJ, Rio de Janeiro, Brazil).
Cells were cultured in Dulbecco’s Modified Eagle’s Med-
ium supplemented with 5 % fetal bovine serum (Invitrogen,
Carlsbad, CA), 2 g/L HEPES buffer, 3.7 g/L NaHCO3, and
1 % penicillin and streptomycin (Invitrogen) in a humidi-
fied atmosphere of 5 % CO2 at 37 �C. Mouse HSCs were
isolated from normal livers of male Balb/c mice (aged
20–27 weeks) by the pronase-collagenase method followed
by a Nycodenz gradient as described elsewhere [9, 10].
After isolation, cells were plated on plastic for several days.
Animals were used in accordance with institutional ethical
guidelines. Cells were incubated with 100 lM capsaicin
(M2028, Sigma-Aldrich, St. Louis, MO) for different time
points as indicated in the figure legends.
Viable Cell Counting
GRX cells were seeded into 24-well plates at a density of
5 9 105 cells/well and treated with capsaicin for 24 h. For
determination of cell number, cells were counted using a
hemocytometer. Trypan blue analyses were performed to
determine cellular viability.
Proliferation Assay
Cell proliferation was measured as active DNA synthesis
with the Click-iT EdU Cell Proliferation Assay Kit
(Invitrogen). GRX and freshly isolated mouse HSCs were
plated in the presence or absence of capsaicin and/or the
mitogen PDGF-bb (20 ng/mL; R&D Systems, Minneapo-
lis, MN). After 48 h, EdU labeling was initiated. After 24
or 48 h, for GRX and primary mouse HSC, respectively,
cells were formalin-fixed. Visualization of the EdU incor-
poration was obtained according to the manufacturer’s
instructions. The ratio of total cells and EdU incorporated
cells was calculated.
Cell Cycle Analysis
To determine the effect of capsaicin on the cell cycle,
defined as G0/G1, S, and G2/M phase, GRX cells were first
synchronized by 24 h serum starvation and then exposed to
capsaicin for 24 h. Cell cycle phase analysis was per-
formed using the FITC BrdU Flow Kit (BD Biosciences,
San Jose, CA). In brief, after treatment and BrdU labeling,
cells were harvested by trypsinization and adjusted to
1 9 106 cells/mL. Samples were fixed with BD Cytofix/
Cytoperm Buffer (BD Biosciences). Following fixation,
cells were treated with DNase for 20 min to expose BrdU
epitopes. Later on, DNA was stained for cell cycle analysis
using 7-AAD dye. The DNA profiles were determined by
FACSCanto II flow cytometer (BD Biosciences) and ana-
lyzed using the FlowJo 7.2.5 software (Tree Star Inc.).
Quantification of Apoptosis
Apoptosis was assessed using the FITC Annexin V
Apoptosis Detection Kit I (BD Bioscience). In brief, after
treating with capsaicin for 24 h, GRX cells were washed
twice with PBS and resuspended in binding buffer before
addition of annexin V-FITC and propidium iodide (PI).
Cells were vortexed and incubated for 15 min in the dark at
room temperature. A total of 10,000 events were acquired
for each assayed sample. All data were acquired with a
FACSCanto II flow cytometer (BD Biosciences). Data
were analyzed using the FlowJo 7.2.5 software (Tree Star
Inc., Ashland, OR). Results are displayed as scatter dots
allowing discrimination among viable cells, apoptotic cells
with an intact membrane, and cells undergoing secondary
necrosis.
Analysis of Mitochondrial Membrane Potential (DWm)
Breakdown of DWm was determined by FACS analysis
using the MitoScreen Kit (BD Biosciences). JC-1 (5,5,6,
6-tetra-chloro-1,1,3,3-tetraethylbenzimidazol-carbocyanine
iodide) dye, which is selectively incorporated into mito-
chondria, is a sensitive and reliable method to detect
changes of the mitochondrial membrane potential (DWm)
[11, 12]. After incubation for 24 h with capsaicin, cells
were stained with 0.5 ml JC-1 solution for 15 min at
37 �C. Stained GRX were washed twice in JC-1 Mito-
Screen wash buffer. A total of 10,000 events were acquired
for each assayed sample. All data were obtained immedi-
ately after staining on a FACSCanto II flow cytometer with
CellQuest PRO v4.0.2 software (BD Biosciences). Results
are displayed as scatter dots allowing discrimination
between polarized and depolarized cells.
Cell Biochem Biophys
123

Cell Migration
Cell motility was determined with a modified Boyden
chamber assay. In brief, cells were suspended in a medium
supplemented with 2.5 % serum (GRX, 2 9 105/well) or
serum-free medium (mouse HSCs, 5 9 104/well) in the
presence or absence of capsaicin. The cells were then
seeded into the upper chambers of transwell inserts
equipped with 8-lM pore polyethylene terephthalate filters
(Millipore, Billerica, MA) previously coated with type I
collagen (4 mg/mL; BD Biosciences). The lower chambers
were filled with medium supplemented with 2.5 % serum.
For the chemotaxis experiment, the lower chambers were
filled with serum-free medium supplemented with PDGF-
BB (20 ng/mL). After 24 h, non-migrated cells on the
upper side of the membrane were rubbed off with a cotton
swab. Migrated cells on the underside of the membrane
were fixed in 100 % methanol, stained with DAPI, and
counted in a fluorescent microscope.
Real Time Quantitative PCR (qPCR)
Total RNA of GRX and primary mouse HSC was extracted
using Reliaprep RNA Cell Miniprep System purification kit
(Promega, Madison, WI). RNA was reverse-transcribed
using RevertAidTM Premium Reverse Transcriptase kit
(Fermentas, St. Leon-Rot, Germany). For real-time PCR,
GoTaq qPCR Master Mix with BRYTE green was used
(Promega), subjected to qPCR in an ABI 7500 Real Time
PCR System and analyzed using System SDS software
(Applied Biosystems, Foster City, CA, USA). The gene-
specific primers used are listed in Table 1. GAPDH was
used as reference gene.
Quantification of Cytokines
To determine cytokine production, GRX cells were cul-
tured for 24 h and 10 days. The supernatants were col-
lected and stored at -20 �C for later analysis. Multiple
soluble cytokines (IL-12 (p70), IFN-c, IL-10 and MCP-1)
were simultaneously measured by flow cytometry using the
cytometric bead array (CBA) Mouse Inflammation Kit (BD
Biosciences). Acquisition was performed with a FACSC-
anto II flow cytometer (BD Biosciences). Quantitative
results were generated using FCAP Array v1.0.1 software
(Soft Flow Inc., Pecs, Hungary). The detection limit was
20–5,000 pg/mL.
Immunocytochemistry
Freshly isolated mouse HSCs were cultivated on glass
coverslips and formalin-fixed after 10 days of culture in the
presence or absence of capsaicin followed by overnight
incubation with primary antibody against a-smooth muscle
actin (a-SMA, 1/1000; Sigma). Antibody binding was
visualized using Alexa 488-labeled antibody (1/250;
Molecular Probes, Eugene, OR). Following mounting with
ProLong� Gold antifade reagent with DAPI (Invitrogen),
cells were analyzed by fluorescent microscope (Carl Zeiss,
Zaventem, Belgium).
Table 1 List of qPCR primersGene Genebank accession number Primers sequence (forward/reverse)
GAPDH NM_008084.2 50-cctgcttcaccaccttcttg-30/
50-tgtccgtcgtggatctgac-30
a-SMA NM_007392 50-ccagcaccatgaagatcaag-30/
50-tggaaggtagacagcgaagc-30
Lox NM_010728 50-tcactgcgctcgttctgat-30/
50-cgatcgaaagtatgagggatg-30
Col1a1 NM_007742 50-cctaagggtaccgctgga-30/
50-tccagcttctccatctttgc-30
Col3a1 NM_009930.1 50-tggtcctgctggaaaggat-30/
50-caggcagtccacgctctc-30
MMP-13 NM_008607.2 50-tgtttgcagagcactacttgaa-30/
50-cagtcacctctaagccaaagaaa-30
MMP-2 NM_008610.2 50-aactttgagaaggatggcaagt-30/
50-tgccacccatggtaaacaa-30
MMP-9 NM_013599.2 50-ctggacagccagacactaaag-30/
50-ctcgcggcaagtcttcagag-30
TIMP-1 NM_001044384.1 50-gcaaagagctttctcaaagacc-30/
50-aagggatagataaacagggaaaca-30
Cell Biochem Biophys
123

Statistical Analysis
Statistical analyses were performed using GraphPad Prism
5.0 (GraphPad Software, San Diego, CA). All experiments
were repeated at least three times and data expressed as
mean ± SEM. Comparisons were performed according to
Student’s t test or one-way ANOVA followed by Bonfer-
roni’s post-test wherever appropriate. A P value\0.05 was
considered statistically significant.
Results
To further characterize the antiproliferative action of cap-
saicin, we investigated the effect of this molecule on cell
cycle and programmed cell death of activated HSCs. GRX,
a murine cell line that presents a myofibroblast transitional
phenotype, was cultured with capsaicin for a 10-day per-
iod. The effect on cell migration and the expression and
release of pro- and anti-fibrogenic markers were also
assessed. To analyze the effects of capsaicin during HSC
activation, freshly isolated mouse HSCs were also cultured
with capsaicin. The dose and period of incubation of cap-
saicin were based on our previous work [8].
Capsaicin Inhibits Activated HSC Proliferation
and Migration
As shown in Fig. 1a, a 24 h capsaicin (100 lM) exposure
significantly reduced the number of viable GRX cells by
approximately 32 % compared with untreated control. In
order to analyze whether the above effect of capsaicin on
cell viability was accompanied by decreased entry into S
phase and DNA synthesis, incorporation of EdU, a thy-
midine analog, was assessed in serum-starved GRX cells.
EdU incorporation was measured after a 24 h treatment
with PDGF-BB in the presence or absence of capsaicin. As
shown in Fig. 1b, capsaicin led to a significant decrease in
the proportion of EdU? cells when compared with non-
treated cells. The effects of capsaicin on PDGF-BB-
induced mitogenesis were also evaluated. PDGF-induced
HSC proliferation was drastically reduced after capsaicin
administration. While PDGF-BB-treated cells showed
about 25 % DNA incorporation of EdU, the co-treatment
of PDGF-BB and capsaicin showed only 2 %, indicating
less proliferation (Fig. 1b). We also analyzed the cell cycle
by flow cytometry. The analysis showed that the antipro-
liferative effect of capsaicin was associated with a G0/G1
cell cycle arrest (Fig. 1c). The percentage of cells in phase
G0/G1 increased from 31.2 % in control cells to 44.7 % in
cells treated with capsaicin.
The above-mentioned findings raised the question
whether apoptosis was changed in capsaicin-treated cells.
The potential effect of capsaicin on apoptosis was analyzed
by two different methods, one detecting apoptotic cells by
measuring the translocation of phosphatidylserine to the
outer cell membrane surface and a second measuring the
impact on mitochondrial transmembrane potential (DWm).
Flow cytometric analysis of the Annexin V labeling assay
Fig. 1 Effects of capsaicin on activated HSC proliferation. a GRX
cells were treated with capsaicin for 24 h and cell viability assessed
by direct cell counting. Results are expressed as cell number. b GRX
cells were cultured for 24 h in the presence or absence of CPS and/or
PDGF-BB. Proliferation rate was assessed by EdU incorporation
assays. Results are expressed as percentage of EdU? cells. c Effect of
24-h capsaicin treatment on the GRX cell cycle measured by flow
cytometry. Results are expressed as percentage of cell number. All
data represent the mean ± SEM (n = 3). *P \ 0.05, **P \ 0.01,
***P \ 0.001. CPS capsaicin
Cell Biochem Biophys
123

did not detect apoptosis in the capsaicin-treated cells. As
shown in Fig. 2a, the number of apoptotic cells did not
increase compared to control experiments. To assess the
effects of capsaicin on mitochondrial injury, we analyzed
the DWm of treated cells. Changes of DWm were deter-
mined by JC-1 staining of GRX cells treated with capsaicin
for 24 h. Capsaicin did not provoke significant loss of
DWm (Fig. 2b).
To examine the effects of capsaicin on the migration of
activated HSCs, a modified Boyden chamber migration
assay was performed. PDGF-BB was used as chemoat-
tractant. As shown in Fig. 3, GRX cells treated with cap-
saicin showed a reduced cell motility even in the presence
of PDGF-BB.
MMPs and Inflammatory Cytokines are Modulated
by Capsaicin
We had previously proposed that capsaicin might de-acti-
vate HSCs via decreasing profibrotic and proinflammatory
mediators. HSCs dedifferentiate when exposed to capsaicin
for 10 days [8]. To complement our studies, we investi-
gated the effect of capsaicin on MMP gene expression. As
shown in Fig. 4, capsaicin upregulates the expression of
MMP-13 after 10 days of capsaicin exposure, while the
expression of MMP-2 and MMP-9 is downregulated.
TIMP-1 was not affected by capsaicin.
We additionally investigated whether capsaicin could
also modulate soluble agents involved in HSC activation.
Multiple inflammatory cytokines (IL-12 (p70), IFN-c, IL-
10, and MCP-1) were assessed in GRX culture supernatants
by CBA. All cytokines were analyzed after 24 h and
10 days of 100 lM capsaicin exposure (Fig. 5). IL-12,
known to be the trigger of other cytokines (Kong et al.
[27]), had its bioactive form (p70) secretion increased only
after 10-day exposure to capsaicin. Consequently, the
release of IFN-c increased two times. In addition, anti-
inflammatory cytokine IL-10 levels were substantially
unaffected in the first 24 h. However, IL-10 release had a
4.3-fold increase after 10-day treatment with capsaicin.
Fig. 2 Effects of capsaicin on
programmed cell death. a Flow
cytometric scatter plot of FITC-
annexin V/PI stained GRX
control and capsaicin-treated
cells for 24 h. The lower left
quadrant shows the viable cells,
which are negative for annexin
V and PI. The lower right
quadrant represents the
apoptotic cells, annexin V
positive, and PI negative. The
upper right quadrant contains
the late apoptotic or dead cells
that are positive for annexin V
and PI. Representative
experiment out of three. b Flow
cytometric scatter plot of the
capsaicin impact on DWm. GRX
cells non-treated or treated with
capsaicin for 24 h. The upper
quadrant represents the
polarized cells and the lower
quadrant the depolarized cells.
Representative experiment out
of three. CPS capsaicin
Cell Biochem Biophys
123

MCP-1 levels were increased in the first 24 h and then,
drastically reduced up to ninefold in day 10.
Effects of Capsaicin During Activation of HSCs
Our results suggest that capsaicin is capable of reducing
proliferation, migration, and expression and release of
profibrogenic markers in activated HSCs. This prompted us
to investigate whether this molecule is able to impair the
activation of HSCs. To this end, we incubated freshly
isolated mouse HSCs with capsaicin. In vitro activation of
HSC is characterized by a phenotypic transdifferentiation
from a small star-shaped cell filled with lipid droplets at
day 1 toward a large myofibroblast-like cell depleted of
Fig. 3 Migratory response of GRX cells cultured for 24 h in the
presence or absence of CPS and/or PDGF-BB. a Representative
images out of three experiments showing migrated cells stained with
DAPI. b Direct cell counting of migrated cells in control conditions or
in the presence of PDGF-BB with or without CPS. Results are
presented as cell number. Data are expressed as mean ± S.E.M.
(n = 3). *P \ 0.05, **P \ 0.01, ***P \ 0.001. CPS capsaicin
Fig. 4 Timecourse of MMP
gene expression after capsaicin
exposure. mRNA levels of
MMP-13, TIMP-1, MMP-2, and
MMP-9 in capsaicin-treated
GRX cells were determined by
RT-qPCR. Results are presented
as relative expression of mRNA
levels at day 1. Data are
expressed as mean ± S.E.M.
(n = 3). *P \ 0.05,
**P \ 0.01, ***P \ 0.001.
CPS capsaicin
Cell Biochem Biophys
123

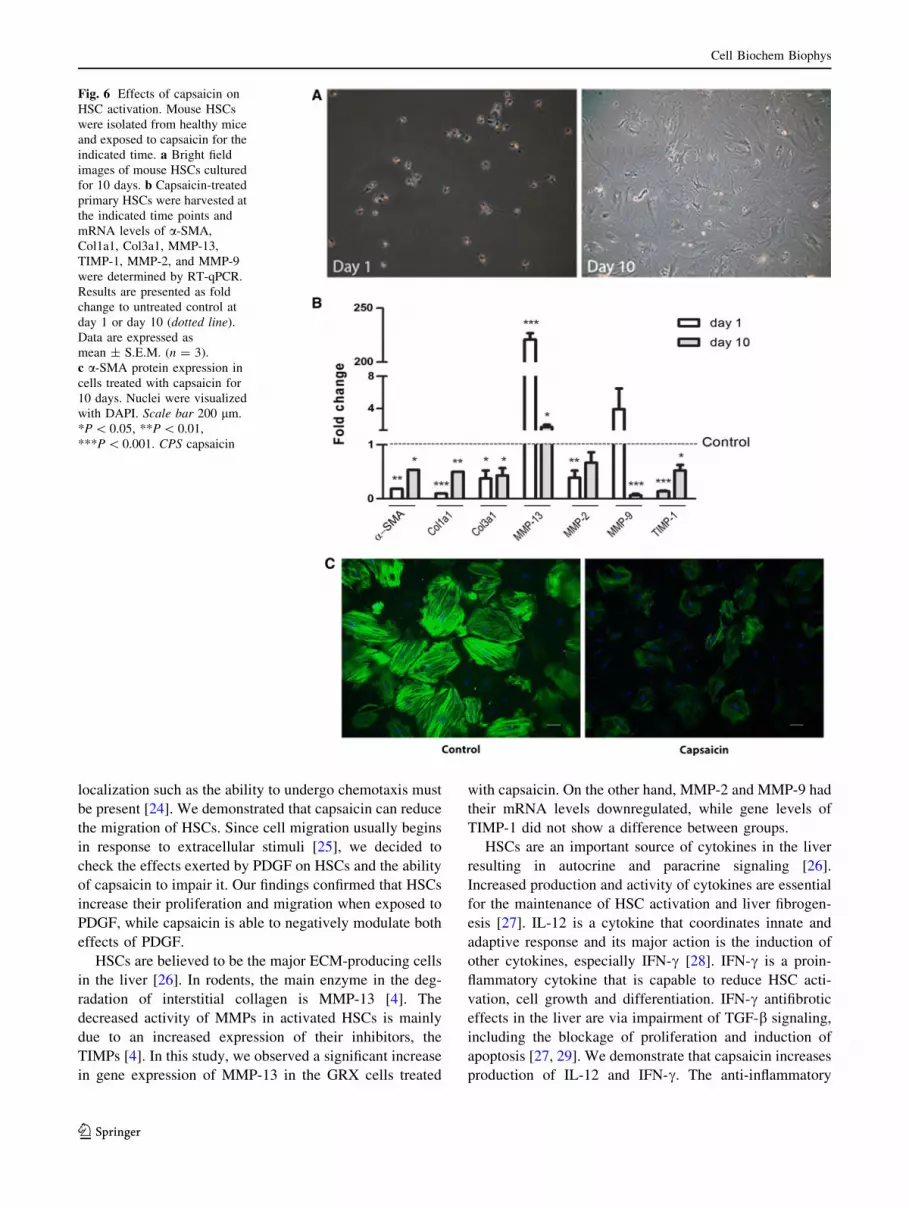
retinoid-containing lipid droplets at day 10 (Fig. 6a).
Therefore, we examined the short (24 h) and long term
(10 days) effects of capsaicin in primary mouse HSCs.
qPCR analysis shows the expression of a-SMA, Col1a1,
Col3a1 and TIMP-1 was downregulated in 24 h and
10 days (Fig. 6b). MMP-13 showed an upregulation in
both timepoints evaluated. MMP-2 was significantly
downregulated at day 1, while MMP-9 mRNA levels were
only decreased at day 10. Next, we evaluated by immu-
nocytochemistry whether a-SMA protein expression was
also influenced by capsaicin in primary cultures. As seen in
Fig. 6c, a-SMA protein expression is clearly downregu-
lated by capsaicin.
Proliferation and chemotaxis are also key features of
HSC activation. Hence, to complement our findings, we
investigated whether capsaicin could reduce the prolifera-
tion and migration of primary mouse HSCs. Figure 7a, b
shows that cells treated with capsaicin for 48 h have a
drastically decreased DNA incorporation of EdU. The
migratory capacity of primary HSCs was inhibited by
capsaicin, even in condition where the chemotactic stim-
ulation was enhanced with PDGF-BB (Fig. 7c).
Discussion
The murine GRX cell line presents several characteristics
of activated HSCs. They are in a transitional state between
quiescent lipocyte and fully activated myofibroblast [13]. It
has been previously demonstrated that GRX cells can be
induced to express the lipocyte phenotype with an overall
increase of lipid storage by a variety of phytochemicals
[8, 14–16]. Capsaicin, a naturally occurring alkaloid from
capsicum fruits, is known to have antiproliferative and anti-
inflammatory properties [17]. Several studies support that
capsaicin can inhibit proliferation by inducing apoptosis or
halting cell cycle in a variety of cell lines, especially tumor
cells [7, 18–20].
Our previous work showed that capsaicin deactivates
HSCs and leads to a more quiescent cell morphology by
suppressing fibrotic mediators probably via PPARc path-
way [8]. In this current study, we showed that capsaicin
treatment inhibited cell proliferation of activated HSCs.
Since PPARc is implicated in growth arrest and apoptosis
[21], we thought that capsaicin, as a PPARc-ligand, may
act via the apoptosis pathway. In the early stages of
apoptosis changes occur at the cell surface, one of these
alterations is the externalization of phosphatidylserine [22].
Furthermore, mitochondrial dysfunction has been shown to
participate and perhaps be central to the apoptotic pathway.
Mitochondrial disintegration not only leads to a depolar-
ization of the transmembrane potential (DWm) but also
causes the release of proapoptotic factors. In some apop-
totic systems, loss of DWm may be an earlier event in the
apoptotic process [23]. According to our findings, capsaicin
does not induce change in plasma membrane structure or
disruption of mitochondrial membrane potential.
In established fibrosis, HSCs are not dispersed within
the parenchyma, but rather are present in very character-
istic areas of fibrosis. For this to take place, a mechanism of
Fig. 5 Capsaicin modulates the
release of inflammatory
cytokines. Flow cytometric
analyses of IL-12 (p70), IFN-c,
IL-10, and MCP-1 in GRX cells
supernatant of 24-h and 10-day
treatments. Cytokines levels are
expressed as picograms per
1 9 105 cells. Data represent
the mean ± S.E.M. (n = 3).
*P \ 0.05, ***P \ 0.001. CPS
capsaicin
Cell Biochem Biophys
123
localization such as the ability to undergo chemotaxis must
be present [24]. We demonstrated that capsaicin can reduce
the migration of HSCs. Since cell migration usually begins
in response to extracellular stimuli [25], we decided to
check the effects exerted by PDGF on HSCs and the ability
of capsaicin to impair it. Our findings confirmed that HSCs
increase their proliferation and migration when exposed to
PDGF, while capsaicin is able to negatively modulate both
effects of PDGF.
HSCs are believed to be the major ECM-producing cells
in the liver [26]. In rodents, the main enzyme in the deg-
radation of interstitial collagen is MMP-13 [4]. The
decreased activity of MMPs in activated HSCs is mainly
due to an increased expression of their inhibitors, the
TIMPs [4]. In this study, we observed a significant increase
in gene expression of MMP-13 in the GRX cells treated
with capsaicin. On the other hand, MMP-2 and MMP-9 had
their mRNA levels downregulated, while gene levels of
TIMP-1 did not show a difference between groups.
HSCs are an important source of cytokines in the liver
resulting in autocrine and paracrine signaling [26].
Increased production and activity of cytokines are essential
for the maintenance of HSC activation and liver fibrogen-
esis [27]. IL-12 is a cytokine that coordinates innate and
adaptive response and its major action is the induction of
other cytokines, especially IFN-c [28]. IFN-c is a proin-
flammatory cytokine that is capable to reduce HSC acti-
vation, cell growth and differentiation. IFN-c antifibrotic
effects in the liver are via impairment of TGF-b signaling,
including the blockage of proliferation and induction of
apoptosis [27, 29]. We demonstrate that capsaicin increases
production of IL-12 and IFN-c. The anti-inflammatory
Fig. 6 Effects of capsaicin on
HSC activation. Mouse HSCs
were isolated from healthy mice
and exposed to capsaicin for the
indicated time. a Bright field
images of mouse HSCs cultured
for 10 days. b Capsaicin-treated
primary HSCs were harvested at
the indicated time points and
mRNA levels of a-SMA,
Col1a1, Col3a1, MMP-13,
TIMP-1, MMP-2, and MMP-9
were determined by RT-qPCR.
Results are presented as fold
change to untreated control at
day 1 or day 10 (dotted line).
Data are expressed as
mean ± S.E.M. (n = 3).
c a-SMA protein expression in
cells treated with capsaicin for
10 days. Nuclei were visualized
with DAPI. Scale bar 200 lm.
*P \ 0.05, **P \ 0.01,
***P \ 0.001. CPS capsaicin
Cell Biochem Biophys
123

IL-10 that downregulates the immune response is also
synthesized by activated HSCs. As a key factor for reducing
perpetuation of fibrogenesis, it suppresses inflammation
through several mechanisms, including the reduction of
proinflammatory cytokines [4]. In this study, we showed
that IL-10 levels increased upon capsaicin treatment, sup-
porting the inhibitory effect of IL-10 on fibrogenesis. MCP-
1 is another chemoattractant implicated in direct migration
of activated HSCs to the site of injury. However, quiescent
cells do not express MCP-1, and responsiveness to it is an
indicator of at least a minimum level of HSC activation
[30]. Recent data indicate that MCP-1 expression can be
downregulated by agonists of PPARc [31]. We demon-
strated that capsaicin-treated HSCs had their secretion of
MCP-1 drastically decreased (*90 %).
To confirm that capsaicin could be a candidate to pre-
serve the quiescent HSC phenotype in vitro, we also tested
its effects on freshly isolated mouse HSCs. As expected,
we observed similar trends as in activated HSCs; capsaicin
inhibited the culture-induced activation of primary mouse
HSCs, prevents the upregulation of several activation
markers such as a-SMA, Col1a1, Col3a1, MMP-2, MMP-
9, and TIMP-1, and capsaicin inhibited the PDGF-induced
chemotaxis and proliferation of HSCs.
In conclusion, the present study indicates that capsaicin is
capable of modulating HSC activation. In addition to its capa-
city to induce a decrease in activation features in already
activated HSCs, capsaicin is able to inhibit the first steps of
activation. Further studies shall focus on examining the an-
tifibrotic effects of capsaicin using in vivo models.
Acknowledgments This work was supported by CAPES/DGU
(BEX 4426/10-0) Grant of the Brazilian Ministry of Science and
Technology and Secretaria de Estado de Universidades (PHB2008-
0080-PC) Grant of the Spanish Ministry of Science and Innovation.
SB is receipt of a fellowship from CAPES. LAvG is supported by the
Vrije Universiteit Brussels.
References
1. Kershenobich Stalnikowitz, D., & Weissbrod, A. B. (2003). Liver
fibrosis and inflammation: A review. Annals of hepatology, 2(4),
159–163. PubMed PMID: 15115954.
2. Day, S. A., Lakner, A. M., Moore, C. C., Yen, M. H., Clemens,
M. G., Wu, E. S., et al. (2011). Opioid-like compound exerts anti-
fibrotic activity via decreased hepatic stellate cell activation and
inflammation. Biochemical Pharmacology, 81(8), 996–1003.
PubMed PMID: 21291870. Epub 2011/02/05. eng.
3. Van Beneden, K., Mannaerts, I., Pauwels, M., Van den Branden,
C., & van Grunsven, L. A. (2013). HDAC inhibitors in experi-
mental liver and kidney fibrosis. Fibrogenesis & Tissue Repair,
6(1), 1. PubMed PMID: 23281659. Pubmed Central PMCID:
3564760.
4. Iimuro, Y., & Brenner, D. A. (2008). Matrix metalloproteinase
gene delivery for liver fibrosis. Pharmaceutical Research, 25(2),
249–258. PubMed PMID: 17577645. Pubmed Central PMCID:
2245995.
5. Solıs-Herruzo, J., Solıs-Munoz, P., Munoz-Yague, T., & Garcıa-
Ruiz, I. (2011). Molecular targets in the design of antifibrotic
therapy in chronic liver disease. Revista Espanola de Enfermed-
ades Digestivas, 103(6), 310–323.
6. Kim, C. S., Park, W. H., Park, J. Y., Kang, J. H., Kim, M. O.,
Kawada, T., et al. (2004). Capsaicin, a spicy component of hot
pepper, induces apoptosis by activation of the peroxisome pro-
liferator-activated receptor gamma in HT-29 human colon cancer
cells. Journal of Medicinal Food, 7(3), 267–273. PubMed PMID:
15383218. Epub 2004/09/24. eng.
7. Lin, C. H., Lu, W. C., Wang, C. W., Chan, Y. C., & Chen, M. K.
(2013). Capsaicin induces cell cycle arrest and apoptosis in
human KB cancer cells. BMC Complementary and Alternative
Medicine, 13, 46. PubMed PMID: 23433093. Pubmed Central
PMCID: 3599796.
Fig. 7 Effects of capsaicin on
HSC activation and migration.
a Representative image of the
EdU-staining of freshly isolated
mouse HSCs that were cultured
for 48 h in the presence or
absence of CPS. b Proliferation
rate assessed by EdU
incorporation assay. Results are
expressed as percentage of
EdU? cells. c Direct cell
counting of migrated cells in the
presence of PDGF-BB with or
without CPS. Results are
presented as cell number. Data
are expressed as
mean ± S.E.M. (n = 3).
*P \ 0.05, **P \ 0.01. CPS
capsaicin
Cell Biochem Biophys
123

8. Bitencourt, S., de Mesquita, F. C., Caberlon, E., da Silva, G. V.,
Basso, B. S., Ferreira, G. A., et al. (2012). Capsaicin induces de-
differentiation of activated hepatic stellate cell. Biochemistry and
Cell Biology, 90(6), 683–690. PubMed PMID: 22905849. Epub
2012/08/22. eng.
9. Guimaraes, E. L., Empsen, C., Geerts, A., & van Grunsven, L. A.
(2010). Advanced glycation end products induce production of
reactive oxygen species via the activation of NADPH oxidase in
murine hepatic stellate cells. Journal of Hepatology, 52(3),
389–397.
10. Mannaerts, I., Nuytten, N. R., Rogiers, V., Vanderkerken, K., van
Grunsven, L. A., & Geerts, A. (2010). Chronic administration of
valproic acid inhibits activation of mouse hepatic stellate cells
in vitro and in vivo. Hepatology, 51(2), 603–614.
11. Cossarizza, A., Baccarani-Contri, M., Kalashnikova, G., &
Franceschi, C. (1993). A new method for the cytofluorimetric
analysis of mitochondrial membrane potential using the
J-aggregate forming lipophilic cation 5,50,6,60-tetrachloro-
1,10,3,30-tetraethylbenzimidazolcarbocyanine iodide (JC-1). Bio-
chemical and Biophysical Research Communications, 197(1),
40–45. PubMed PMID: 8250945. Epub 1993/11/30. eng.
12. Reers, M., Smiley, S. T., Mottola-Hartshorn, C., Chen, A., Lin,
M., & Chen, L. B. (1995). Mitochondrial membrane potential
monitored by JC-1 dye. Methods in Enzymology, 260, 406–417.
PubMed PMID: 8592463. Epub 1995/01/01. eng.
13. Borojevic, R., Guaragna, R. M., Margis, R., & Dutra, H. S.
(1990). In vitro induction of the fat-storing phenotype in a liver
connective tissue cell line-GRX. In Vitro Cellular & Develop-
mental Biology, 26(4), 361–368. PubMed PMID: 2188940. Epub
1990/04/01. eng.
14. Souza, I. C., Martins, L. A., Coelho, B. P., Grivicich, I., Guara-
gna, R. M., Gottfried, C., et al. (2008). Resveratrol inhibits cell
growth by inducing cell cycle arrest in activated hepatic stellate
cells. Molecular and Cellular Biochemistry, 315(1–2), 1–7.
PubMed PMID: 18454344. Epub 2008/05/06. eng.
15. Teodoro, A. J., Perrone, D., Martucci, R. B., & Borojevic, R.
(2009). Lycopene isomerisation and storage in an in vitro model
of murine hepatic stellate cells. European Journal of Nutrition,
48(5), 261–268. PubMed PMID: 19533199. Epub 2009/06/18.
eng.
16. Braganca de Moraes, C. M., Melo, D. A., Santos, R. C., Biten-
court, S., Mesquita, F. C., Santos de Oliveira, F. D., et al. (2012).
Antiproliferative effect of catechin in GRX cells. Biochemistry
and Cell Biology, 90(4), 575–584. PubMed PMID: 22574829.
Epub 2012/05/12. eng.
17. Gupta, S. C., Kim, J. H., Prasad, S., & Aggarwal, B. B. (2010).
Regulation of survival, proliferation, invasion, angiogenesis, and
metastasis of tumor cells through modulation of inflammatory
pathways by nutraceuticals. Cancer and Metastasis Reviews,
29(3), 405–434. PubMed PMID: 20737283. Pubmed Central
PMCID: 2996866. Epub 2010/08/26. eng.
18. Surh, Y. J. (2002). Anti-tumor promoting potential of selected
spice ingredients with antioxidative and anti-inflammatory
activities: a short review. Food and Chemical Toxicology, 40(8),
1091–1097. PubMed PMID: 12067569. Epub 2002/06/18. eng.
19. Takahata, K., Chen, X., Monobe, K., & Tada, M. (1999). Growth
inhibition of capsaicin on HeLa cells is not mediated by intra-
cellular calcium mobilization. Life Sciences, 64(13), PL165–
PL171. PubMed PMID: 10210280. Epub 1999/04/21. eng.
20. Hsu, C. L., & Yen, G. C. (2007). Effects of capsaicin on induc-
tion of apoptosis and inhibition of adipogenesis in 3T3-L1 cells.
Journal of Agriculture and Food Chemistry, 55(5), 1730–1736.
PubMed PMID: 17295509. Epub 2007/02/14. eng.
21. Zhang, F., Kong, D., Lu, Y., & Zheng, S. (2012). Peroxisome
proliferator-activated receptor-gamma as a therapeutic target for
hepatic fibrosis: From bench to bedside. Cellular and Molecular
Life Sciences, 70(2), 259–276. PubMed PMID: 22699820. Epub
2012/06/16. Eng.
22. van Engeland, M., Nieland, L. J., Ramaekers, F. C., Schutte, B.,
& Reutelingsperger, C. P. (1998). Annexin V-affinity assay: a
review on an apoptosis detection system based on phosphatidyl-
serine exposure. Cytometry, 31(1), 1–9. PubMed PMID:
9450519. Epub 1998/02/05. eng.
23. Ly, J. D., Grubb, D. R., & Lawen, A. (2003). The mitochondrial
membrane potential (deltapsi(m)) in apoptosis; an update.
Apoptosis, 8(2), 115–128. PubMed PMID: 12766472. Epub
2003/05/27. eng.
24. Friedman, S. L. (2008). Mechanisms of hepatic fibrogenesis.
Gastroenterology, 134(6), 1655–1669. PubMed PMID:
WOS:000255771200003. English.
25. Yang, C., Zeisberg, M., Mosterman, B., Sudhakar, A., Yerra-
malla, U., Holthaus, K., et al. (2003). Liver fibrosis: insights into
migration of hepatic stellate cells in response to extracellular
matrix and growth factors. Gastroenterology, 124(1), 147–159.
26. Friedman, S. L. (2008). Hepatic stellate cells: protean, multi-
functional, and enigmatic cells of the liver. Physiological
Reviews, 88(1), 125–172. PubMed PMID: 18195085. Pubmed
Central PMCID: 2888531.
27. Kong, X., Horiguchi, N., Mori, M., & Gao, B. (2012). Cytokines
and STATs in Liver Fibrosis. Frontiers in physiology, 3, 69.
PubMed PMID: 22493582. Pubmed Central PMCID: 3318231.
28. Watford, W. T., Moriguchi, M., Morinobu, A., & O’Shea, J. J.
(2003). The biology of IL-12: coordinating innate and adaptive
immune responses. Cytokine & Growth Factor Reviews, 14(5),
361–368. PubMed PMID: 12948519. Epub 2003/09/02. eng.
29. Saile, B., Eisenbach, C., Dudas, J., El-Armouche, H., & Rama-
dori, G. (2004). Interferon-gamma acts proapoptotic on hepatic
stellate cells (HSC) and abrogates the antiapoptotic effect of
interferon-alpha by an HSP70-dependant pathway. European
Journal of Cell Biology, 83(9), 469–476. PubMed PMID:
15540463. Epub 2004/11/16. eng.
30. Li, J. T., Liao, Z. X., Ping, J., Xu, D., & Wang, H. (2008).
Molecular mechanism of hepatic stellate cell activation and an-
tifibrotic therapeutic strategies. Journal of Gastroenterology,
43(6), 419–428. PubMed PMID: 18600385. Epub 2008/07/05.
eng.
31. Marra, F. (2002) Chemokines in liver inflammation and fibrosis.
Frontiers in Bioscience, 7, d1899–d1914. PubMed PMID:
12161342. Epub 2002/08/06. eng.
Cell Biochem Biophys
123





























