Embed Size (px)
Citation preview

Cdc2 phosphorylation of nucleolin demarcates mitotic stages andAlzheimer’s disease pathology<
Alex Dranovskya,b, Inez Vincentc, Luisa Gregorib, Alexander Schwarzmanb, David Colfleshd,Jan Enghilde, Warren Strittmattere, Peter Daviesc, Dmitry Goldgaberb,*
aMedical Scientist Training Program, State University of New York, Stony Brook, NY 11794, USAbDepartment of Psychiatry and Behavioral Science, State University of New York, Stony Brook, NY 11794, USA
cDepartment of Pathology, Albert Einstein College of Medicine, Bronx, NY 10461, USAdUniversity Microscopy Imaging Center, State University of New York, Stony Brook, NY 11794, USA
eDepartment of Neurology, Duke University, Durham, NC 27710, USA
Received 28 August 2000; received in revised form 2 November 2000; accepted 3 November 2000
Abstract
Nucleolin is a major multifunctional nuclear phosphoprotein that is phosphorylated by Cdc2 kinase in mitosis and that participates ina number of cellular processes. The monoclonal antibody TG-3 generated against neurofibrillary tangles (NFT) found in Alzheimer’s disease(AD) is highly specific for mitotic cells in culture. We here demonstrate that phosphorylation of nucleolin by Cdc2 kinase generates the TG-3epitope. The unique pool of TG-3 immunoreactive nucleolin appears abruptly during the prophase. It is associated with chromosomesthrough the metaphase and it gradually disappears during separation of chromosomes and exit from mitosis. In the brain, nucleolin waslocalized not only to nuclei but also to neuronal cytoplasm, and it is a marker for early NFT. In patients with AD, Cdc2 phosphorylatednucleolin was present in NFT. These findings suggest that phosphorylation of nucleolin by Cdc2 kinase is a critical event and the point ofconvergence of two distinct pathways, mitosis and neurodegeneration. © 2001 Elsevier Science Inc. All rights reserved.
Keywords:Nucleolin; Mitosis; Neurodegeneration; Alzheimer’s disease
1. Introduction
Several recent reports suggest that reactivation of certaincell cycle events in postmitotic neurons may underlie neu-rofibrillary degeneration in AD [6,25,28,29,43,44]. Evi-dence for involvement of cell cycle specific proteins inneurodegeneration stems from studies on the role of phos-phorylation in the formation of neurofibrillary tangles(NFT), one of the neuropathological hallmarks of AD. NFTconsist of paired helical filaments (PHF) and straight fila-ments. Phosphorylation by proline directed serine-threoninekinases results in PHF properties in tau (a structural com-ponent of PHF) [31]. Experiments in vitro have shown thatmany proline directed kinases phosphorylate recombinanttau [1,27,31,46]. However, the biologic relevance of such
experiments is not clear due to the substrate promiscuity ofthese kinases [30] and the ability of tau to form PHF in theabsence of phosphorylation in vitro [48].
In vivo support for tau phosphorylation by mitotic pro-line directed kinases was provided when a kinase related tothe major mitotic kinase Cdc2 was immunolocalized to NFTin the AD brain [26,50]. Moreover, Vincent and colleaguesrecently demonstrated that Cdc2 kinase was activated in ADneurons [43]. Furthermore, immunoreactivity of other cellcycle markers was detected in the brains of patients affectedby AD [6,28–29]. Aberrant activation of mitotic mecha-nisms in postmitotic neurons may facilitate cellular eventsleading to NFT formation and neurodegeneration. Evidencewas obtained for the involvement of mitotic mechanisms inNFT formation when monoclonal antibodies (TG/MC se-ries) raised against immunopurified NFT showed specificityfor both the AD brain in immunohistochemical studies, andfor mitotic cells in culture [44]. The same study providedprovocative evidence that monoclonal antibody MPM-2,which was raised against mitotic cells [10], was highlyimmunoreactive with AD, but not with control brains.
< This paper is part of the special issue ‘Cell Cycle’ (Guest Editor InezVincent).
* Corresponding author. Tel.:11-631-444-1369; fax:11-631-444-7534.
E-mail address:[email protected] (D. Gold-gaber).
www.elsevier.com/locate/neuagingNeurobiology of Aging 22 (2001) 517–528
0197-4580/01/$ – see front matter © 2001 Elsevier Science Inc. All rights reserved.PII: S0197-4580(00)00248-7

Moreover, MPM-2 recognized NFT, the neuritic compo-nents of plaques, and morphologically healthy neurons inthe AD brain, thus suggesting that it is a marker for the earlystages of neurodegeneration. Similar to the TG/MC series,MPM-2 recognized mitotic phosphoepitopes [10,16,39].The presence of mitotic phosphoepitopes in degeneratingneurons suggests that mitotic and neurodegenerative path-ways converge in AD. Therefore, antibodies that demon-strate high specificity for both AD pathology and mitoticevents can serve as ideal tools for identifying commonmolecules within the two pathways.
Of all the antibodies reported by Vincent and colleagues,TG-3 was the most specific for AD neurofibrillary pathol-ogy demonstrating no detectable immunoreactivity in con-trol brains. Therefore, identifying and characterizing theTG-3 antigen or antigens are essential to our understandingof the convergence of neurofibrillary degeneration and mi-tosis. The first TG-3 immunoreactive protein in the brains ofpatients with AD was recently identified as an abnormallyphosphorylated microtubule associated protein tau [20].Other potentially biologically relevant TG-3 antigens in thebrain remain unknown. However, in cultured cells synchro-nized in mitosis, TG-3 recognizes not tau but a 105 kDaprotein [44]. We here report that the cellular TG-3 antigenis the major nucleolar phosphoprotein nucleolin after phos-phorylation by Cdc2 kinase. We also report that in thehuman brain, nucleolin is present in neuronal cytoplasm andcolocalizes with NFT in AD.
2. Materials and methods
2.1. Antibodies
Monoclonal antibody TG-3 (hybridoma medium) wasdescribed in a previous publication [44] and used at a 1:10dilution for Western blotting. Anti-nucleolin monoclonalantibody MS3 (hybridoma medium) was kindly provided byDrs. H. Busch and L. Perlaky from Baylor College ofMedicine, and used at a dilution of 1:10 for immunocyto-chemistry and at a 1:1000 dilution for Western blotting [42].MS3 ascites fluid was used for immunoprecipitations at a1:10 dilution. Monoclonal antibody D3 (hybridoma medi-um), kindly provided by Dr. J.-S. Deng from the Universityof Pittsburgh, was used at a 1:10 dilution for immunocyto-chemistry [11]. Anti-nucleolin monoclonal antibody CC98(hybridoma medium), kindly provided by Dr. N.H. Yeh,National Yang Ming College, Taiwan, was used at a 1:5dilution for immunocytochemistry [8]. Anti-nucleolinmonoclonal antibody 4E2 (hybridoma medium) from Re-search Diagnostics (Flanders, NJ) was used at a 1:5 dilutionfor immunocytochemistry. Rabbit polyclonal antisera W15generated against human nucleolin and nucleolin-maltose-binding protein fusion protein were generously provided byDr. N. Maizels, Yale University [17]. Each polyclonal an-tiserum was used at a 1:1000 dilution. Non-immune rabbit
serum was used as a control. Monoclonal antibody AT180(Innogenetics, Belgium), which recognizes phospho-threo-nine 231 (Thr231) of tau [15], was used at a 1:10 dilutionfor the blocking experiments. Monoclonal antibodiesagainstb-tubulin (Sigma, St. Louis, MO) and histone pro-tein mAb052 (Chemicon, Temecula, CA) were used at a1:200 final dilution. The specificity of each primary mono-clonal and polyclonal antibody was characterized by theauthors who generated the antibody and was described inthe papers cited above. Secondary goat anti-mouse IgG1aand goat anti-mouse IgM antibodies conjugated to fluores-cein and rhodamine (Jackson Immunochemicals, WestGrove, PA) were used at a 1:200 final dilution for immu-nocytochemistry. Secondary antibodies conjugated to biotinalong with streptavidin conjugated to CY3 and CY5 fluo-rochromes (Southern Biotech, Birmingham, AL) were allused at a 1:500 dilution for the immunohistochemistry ofbrain sections. Secondary antibodies conjugated to horse-radish peroxidase (Amersham, Oakville, Ontario) were usedat a 1:3000 final dilution for Western blotting.
2.2. Cell culture and immunocytochemistry
HEp-2 human epithelial cells (ATCC, Rockville, MD)were cultured in DME containing 10% bovine calf serum,and penicillin-streptomycin (Gibco, Burlington, Ontario)was cultured at 37°C in 5% CO2. Cells were washed withPBS, harvested by trypsinization, counted and plated at1.25 3 104 - 2.5 3 104 cells/cm2 on 8 well chamberedcoverslips (VWR, Piscataway, NJ). On the following day,cells were fixed in 2% paraformaldehyde/0.1% glutaralde-hyde for 40 min, permeabilized with 0.5% Triton X-100 for10 min and stored in PBS at 4°C for immunocytochemistry.Prior to immunocytochemistry, subconfluent monolayerswere blocked with 5% bovine serum albumin (BSA) in PBSfor 30 min and rinsed in 3 volumes of PBS 3 times followedby three 10 min washes in PBS. Immunostaining of mono-layers was performed in the chambered coverslips. Primaryantibodies were applied for 2 h at room temperature orovernight at 4°C in a high humidity incubation chamber.The monolayers were washed as described above and incu-bated with secondary antibodies in the presence of DAPI(Sigma, St. Louis, MO) for 30 min. The monolayers werewashed as described above and stored in the ANTI-FADEreagent (Molecular Probes, Eugene, OR) at 4°C. Rodamine.Fluorescein was visualized by the Noran Confocal Odysseysystem through a Nikon inverted Diaphot microscope, andimages were captured using Image One software.
2.3. Protein purification
HEp-2 cells were grown in suspension according to theconditions described above. Five liters of logarithmicallygrowing cells were synchronized in mitosis by treatmentwith 2 mg/ml of nocodazole (Sigma, St. Louis, MO) over-
518 D. Dranovsky et al. / Neurobiology of Aging 22 (2001) 517–528

night. Nocodazole was diluted from a DMSO solution sothat the final DMSO concentration was 0.05%. Cells wereharvested by centrifugation at 2503 g for 2 min, rinsedwith PBS, centrifuged, and cell pellets were stored at280°C. All extraction and purification procedures wereperformed at 4°C. Cells were extracted in 5 volumes ofbuffer A for 15 min, gently vortexed, and centrifuged at 4003 g for 10 min. Buffer A contained 0.015 M Tris-Cl pH 7.4,0.08 M KCl, 2 mM EDTA-KOH, 1% Nonidet P-40, 0.2 mMspermine, 0.5 mM spermidine, 1 mM phenylmethane-sul-phonyl fluoride, 10 nM Microsystin-LR, 1 mM Na-o-vana-date (all from Sigma), and protease inhibitor cocktail(Boehringer Mannheim, Indianapolis, IN). The supernatantwas chromatographed on DEAE-Sepharose (Pharmacia,Piscataway, NJ) preequilibrated with buffer A, washed, andeluted with a 0.1–0.5 M NaCl step gradient in buffer Ausing 0.1 M increments. TG-3 reactive fractions were iden-tified by Western blotting using the ECL detection system(Amersham, Oakville Ontario). The TG-3 reactive fractionwas diluted in 15 mM Tris, 2 mM EDTA, pH 7.4 (buffer B),chromatographed on HiTrap-Heparin mini column (Phar-macia, Piscataway, NJ) preequilibrated in buffer B, washed,and eluted with a 0.1–0.5 M NaCl step gradient using 0.1 Mincrements. TG-3 immunoreactivity was determined byWestern blotting. The TG-3 reactive fraction was separatedby SDS-PAGE and stained with Coomassie Brilliant Blue.The purified TG-3 reactive 105 kDa protein was subjectedto N-terminal sequencing and the obtained sequence wascompared by BLAST search against the Genbank database.
2.4. Immunoprecipitation
Nocodazole-treated HEp-2 cells from 50 ml of culturemedia were collected by centrifugation and washed twicewith PBS. The following procedures were performed at4°C. Cells were lysed with 1 ml of RIPA buffer [18] andcentrifuged at 80003 g for 30 s. 30ml of supernatant weremixed with 3ml of MS3 ascites fluid. The reaction mixtureswere incubated with rotation for 1 h, and 70ml of anti-mouse IgG agarose (Sigma, St. Louis, MO) were added foran additional 30 min. The resin was washed three times with1 ml of RIPA buffer, and the immunoprecipitated proteinswere eluted by the addition of SDS-PAGE sample buffercontaining 2-mercaptoethanol. The sample was divided intotwo equal parts and subjected to SDS-PAGE, transferredonto PVDF and probed using either MS3 antibodies orTG-3 antibodies.
2.5. “Mitotic shake”
HEp-2 cells were grown in 175 mm2 flasks as describedabove. Mitotic cells were mechanically dislodged from therest of the monolayer by dropping the flasks from three feet.The dislodged mitotic cells were harvested by centrifuga-tion of the supernatant at 10003 g for 5 min. The cell pellet
was lysed and analyzed by Western blotting with TG-3 asdescribed above.
2.6. Chemical cleavage
TG-3 positive nucleolin was purified from nocodazoletreated HEp-2 cells and subjected to SDS-PAGE. The TG-3positive band was cut out of the gel, and the gel slice wassubjected to chemical digestion with N-chlorosuccinimide,which specifically cleaves at tryptophane (positions 481 and641 in nucleolin), or with hydroxylamine, which specifi-cally cleaves between asparagine and glycine (positions136, 137 and 519, 520 in nucleolin). N-chlorosuccinimidewas used at 10 ml of 0.1 M in 2 M urea, and 50% acetic acidfor 30 min at room temperature with rotation.
Hydroxylamine was used at 2 M in 6 M guanidine-HClat pH 9.0 for 12 h at room temperature with rotation. Gelslices were washed with 15 mM Tris, pH 6.8, 0.1% SDS,and 2-mercaptoethanol for 20 min. After chemical cleavage,the gel slice was placed on top of a second SS-gel andelectrophoresed in a direction perpendicular with respect tothe first gel. The separated cleavage products were trans-ferred onto PVDF membrane and analyzed by Westernblotting. The membranes were first immunostained withTG-3, then stripped with SDS and 2b- mercaptoethanol [18]and reprobed with MS3 to detect nucleolin. In the controlexperiments the cleaving reagent was omitted.
2.7. Immunohistochemistry
All human subjects used in this study were evaluatedboth clinically and neuropathologically at the Albert Ein-stein College of Medicine as described elsewhere [44]. Forneuropathological evaluation, tissue was immersion-fixed in4% paraformaldehyde and independently evaluated withantibody Alz-50 and Thioflavin S. Brains from patients withclinical symptoms of AD showed marked neuropathologicallesions. No clinical diagnosis of AD was noted in the con-trol cases. Immunostaining was performed essentially asdescribed elsewhere by free floating 50mm vibratome sec-tions in 24 well culture dishes [44]. For the blocking ex-periment the tissue was incubated overnight at 4°C withmonoclonal antibody AT180 which recognized phosphory-lated threonine 231 (Thr231) on tau and does not recognizenucleolin. The tissue was then washed as described else-where [44], incubated with unlabeled anti-mouse IgG1overnight at 4°C, washed again, incubated with TG-3 andnucleolin rabbit antisera W15 overnight at 4°C, and washed.TG-3 was detected with TRITC conjugated anti-mouseIgM, while W15 was detected with biotinylated anti-rabbitantibodies followed by streptavidin conjugated CY5. Allsecondary incubations were performed as described above.Fluorochromes were visualized by the Bio-Rad Confocalsystem through a Nikon inverted Diaphot microscope.
519D. Dranovsky et al. / Neurobiology of Aging 22 (2001) 517–528

3. Results
3.1. The TG-3 antigen in mitotic cells is nucleolin
TG-3 consistently recognized a major 105 kDa band inextracts from HEp-2 cells that were synchronized in mitosisby treatment with nocodazole (Fig. 1B, lane 5). This bandwas also observed in extracts from synchronized MSN neu-roblastoma [44] and HeLa cells (data not shown). Theappearance of this band was not an artifact of nocodazoletreatment because an identical band was observed in ex-tracts from mitotic HEp-2 cells that were enriched in mitoticcells by mechanical dislodging from the flask (Fig. 1B, lane3). The TG-3 antigen was thus purified from nocodazolesynchronized HEp-2 cells by sequential column chromatog-raphy and subjected to N-terminal sequencing (Fig. 1A).The sequence of the first 18 amino acids of the TG-3 antigenwas identical to the N-terminal sequence of human nucleo-lin, a protein with a molecular mass of 105 kDa [37].
To exclude the possibility that a minor protein of asimilar size, copurifying with nucleolin, could account forthe TG-3 immunoreactivity, we immunoprecipitated celllysates with anti-nucleolin monoclonal antibody MS3 andanalyzed the immunoprecipitate with TG-3. A band with themolecular weight of 105 kDa was immunoprecipitated bythe MS3 antibody and detected by the TG-3 antibody (Fig.
1B, lane 7). These results support our initial finding thatboth MS3 and TG-3 antibodies recognize nucleolin in mi-totic cells.
As a further confirmation, the protein purified from no-codazole synchronized cells was subjected to chemicalcleavage with either N-chlorosuccinimide or hydroxylamine(Fig. 2). Human nucleolin has two recognition sites for eachreagent, distributed in such a way that cleavage generatesthree fragments, two of which were predicted to be detect-able with the antibodies TG-3 and MS3 (see methods).Major bands migrating at 92 kDa and 74 kDa were observedafter cleavage with N-chlorosuccinimide (Fig. 2A) and hy-droxylamine (Fig. 2B), respectively. It should be noted thatalthough nucleolin contains two cleavage sites for eachreagent, under our experimental conditions we detected theproduct of only one major cleavage event. After each cleav-age reaction, both MS3 and TG-3 stained identical bands,suggesting that they recognized the same protein. Thus, theresults from the chemical cleavage experiments, in conjunc-tion with the sequencing and immunoprecipitation experi-ments, proved that the TG-3 antibody recognized nucleolinin mitotic cells.
3.2. TG-3 reactive nucleolin is phosphorylated by Cdc2
Nucleolin is a major cellular phosphoprotein, which hasnine putative phosphorylation sites for the major mitotic
Fig. 1. Purification and identification of the TG-3 antigen. (A) Purification and Western blotting of the TG-3 reactive 105 kDa protein. HEp-2 cells weresynchronized with nocodazole, detergent extracted, and fractionated by sequential DEAE and Heparin chromatography. TG-3 immunoreactivity eluted in asingle fraction from both columns with 0.4 M and 0.5 M NaCl respectively. (B) Western blot analysis (lanes 1–5) and immunoprecipitations withanti-nucleolin MS3 antibody (lanes 6, 7). Lysates of synchronized (lanes 4–7) and asynchronous (lanes 1–3) HEp-2 cells were analyzed with MS3 (lanes1, 4) or TG-3 (lanes 2, 3, 5). Lane 3, cells collected by the mitotic shake technique (see methods).
Fig. 2. Identification of the TG-3 antigen as mitotic nucleolin by chemical cleavage. The TG-3 antigen was partially purified from synchronized cells as inFig. 1, separated by SDS-PAGE, and subjected to partial chemical digestion with N-chlorosuccinimide (A) and hydroxylamine (B). The cleavage productsof each digest were separated by SDS-PAGE and transferred onto two membranes. Each membrane was probed with the TG-3 antibody (lanes 1,2), stripped,tested by ECL, and reprobed with the MS3 antibody (lanes 3,4). Controls contained no cleaving reagent (lanes 1, 3).
520 D. Dranovsky et al. / Neurobiology of Aging 22 (2001) 517–528

kinase Cdc2 and has been shown to be phosphorylated byCdc2 both in vitro and in vivo [2]. Previous studies dem-onstrated that TG-3 recognizes phosphoepitopes [44,20].
Since TG-3 recognizes mitotic and not interphase cells,the following experiments were performed in order to test ifTG-3 immunoreactivity is the result of phosphorylation ofnucleolin by Cdc2. Interphase nucleolin, which is not reac-tive with TG-3, was partially purified from asynchronousHEp-2 cells. Nucleolin enriched samples were phosphory-lated with [32P]ATP in the presence or absence of Cdc2kinase. After phosphorylation, nucleolin was immunopre-cipitated with the MS3 antibody, separated by SDS-PAGE,and analyzed by autoradiography and immunoblotting witheither MS3 or TG-3 antibodies (Fig. 3). Autoradiographyrevealed32P incorporation into numerous bands suggestingthat the nucleolin enriched fraction contained many proteinsthat could be phosphorylated by Cdc2 kinase (Fig. 3A, lane1). A significant incorporation of32P into the protein im-
munoprecipitable with MS3 in the absence of Cdc2 sug-gested that nucleolin was also phosphorylated by copurifiedkinases (Fig. 3A, lane 4).
While many bands showed Cdc2 dependent incorpora-tion of 32P, only a single band migrating at 105 kDa wasimmunoreactive with TG-3 (Fig. 3C, lane 1). The appear-ance of the single TG-3 immunoreactive band at 105 kDaafter phosphorylation with Cdc2 kinase demonstrated theantigenic specificity of the TG-3 phosphoepitope-specificantibody for the nucleolin protein in the relatively crudepreparation. Furthermore, immunoprecipitation of nucleolinfrom the enriched fraction with non-phosphospecific MS3(see Valdez et al., 1995 for epitope mapping) [42] revealedthat nucleolin became immunoreactive with TG-3 only afterphosphorylation by Cdc2 (Fig. 3C, lane 3). Phosphorylationreactions carried out in the absence of Cdc2 did not produceepitopes recognizable by TG-3 (Fig. 3C, lanes 2 and 4).Therefore, TG-3 is specific for phosphoepitopes producedby Cdc2 and not by copurifying endogenous kinases. Fi-nally, phosphorylation was time and kinase concentrationdependent, and TG-3 immunoreactivity was proportional tothe amount of32P incorporation (data not shown). Together,these data demonstrate the specificity of the TG-3 antibodyfor nucleolin phosphorylated by Cdc2.
3.3. TG-3 reactive nucleolin undergoes temporal andspatial changes during mitosis in culture
TG-3 immunoreactivity was examined in an asynchro-nous population of HEp-2 (Fig. 4) and HeLa cells (data notshown) by laser confocal microscopy. Dual labeling withmonoclonal antibodies to eitherh-tubulin or to histoneproteins was used to identify cells in different stages ofmitosis. TG-3 reactive nucleolin (TG-3 nucleolin) appearedabruptly in cells entering mitosis. Interphase cells were notreactive with TG-3 (Fig. 4). TG-3 nucleolin was localized tothe nuclei in cells in late prophase and early prometaphaseat the beginning of centrosomal segregation (Figs. 4A-C).TG-3 nucleolin was also localized to the mitotic centro-somes starting from the earliest stages of polar migration(Figs. 4C and 4F). TG-3 nucleolin became dispersedthroughout the mitotic cytosol as the prometaphase pro-gressed (Figs. 4E and 4F). Optical sectioning revealed thatthe condensed chromosomes were immunoreactive withTG-3 as judged by the overlap with immunostaining thatwas detected by anti-histone antibody mAb053 (Fig. 4L).
The chromosomal localization of TG-3 nucleolin becamemost apparent in metaphase when the majority of the im-munoreactivity was localized to the aligned chromosomes(Figs. 4H and 4K). While chromosomal TG-3 immunore-activity was most intense, some also localized to the mitoticcytosol. The intensity of cytosolic TG-3 immunoreactivityvaried between the metaphase cells of the different cell linesbut was always lower than immunoreactivity associatedwith chromosomes (data not shown). TG-3 immunoreactiv-ity began to decrease at the earliest stages of chromosomal
Fig. 3. Phosphorylation of nucleolin by cdc2 confers TG-3 immunoreac-tivity. Nucleolin partially purified from asynchronous cells was phosphor-ylated by Cdc2/cyclinB in the presence of32P labeled ATP. The phosphor-ylated nucleolin was further immunoprecipitated with anti-nucleolinmonoclonal antibody MS3 and Western-blotted with either MS3 (panel B)or TG-3 (panel C) antibodies. (A) Autoradiograph of samples separated bySDS-PAGE (Lanes 1 and 2). Partially purified preparations were incubatedwith [32P]ATP in the presence (lane 1) and the absence (lane 2) of thecdc2/cyclinB complex. The 105 kDa TG-3 immunoreactive band appearsonly in the sample where Cdc2 was present (lane 1, panel C). (Lanes 3–6)Immunoprecipitations of nucleolin with MS3 from samples prepared as inlanes 1 and 2.32P is incorporated into nucleolin both in the presence (lane3, panel A) and absence (lane 4, panel A) of Cdc2. The 105 kDa, TG-3immunoreactive protein is immunoprecipitated only in the sample wherecdc2 was present (lane 3, panel C). No staining was detected when nonucleolin (lane 5) or no primary antibody (lane 6) was used in the exper-iment. (1) and (-) above lanes indicate presence or absence of the reagent.
521D. Dranovsky et al. / Neurobiology of Aging 22 (2001) 517–528
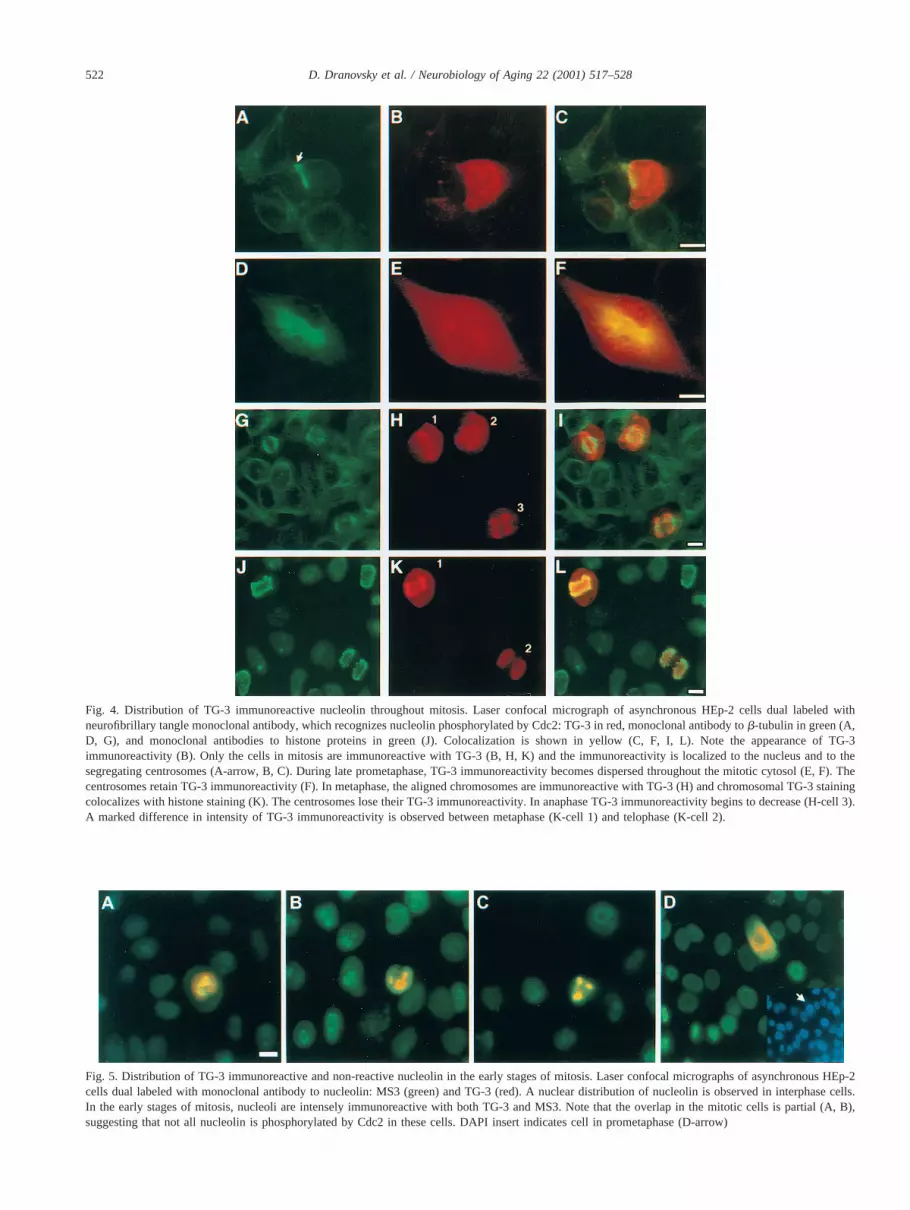
Fig. 4. Distribution of TG-3 immunoreactive nucleolin throughout mitosis. Laser confocal micrograph of asynchronous HEp-2 cells dual labeled withneurofibrillary tangle monoclonal antibody, which recognizes nucleolin phosphorylated by Cdc2: TG-3 in red, monoclonal antibody tob-tubulin in green (A,D, G), and monoclonal antibodies to histone proteins in green (J). Colocalization is shown in yellow (C, F, I, L). Note the appearance of TG-3immunoreactivity (B). Only the cells in mitosis are immunoreactive with TG-3 (B, H, K) and the immunoreactivity is localized to the nucleus and to thesegregating centrosomes (A-arrow, B, C). During late prometaphase, TG-3 immunoreactivity becomes dispersed throughout the mitotic cytosol (E, F). Thecentrosomes retain TG-3 immunoreactivity (F). In metaphase, the aligned chromosomes are immunoreactive with TG-3 (H) and chromosomal TG-3 stainingcolocalizes with histone staining (K). The centrosomes lose their TG-3 immunoreactivity. In anaphase TG-3 immunoreactivity begins to decrease (H-cell 3).A marked difference in intensity of TG-3 immunoreactivity is observed between metaphase (K-cell 1) and telophase (K-cell 2).
Fig. 5. Distribution of TG-3 immunoreactive and non-reactive nucleolin in the early stages of mitosis. Laser confocal micrographs of asynchronous HEp-2cells dual labeled with monoclonal antibody to nucleolin: MS3 (green) and TG-3 (red). A nuclear distribution of nucleolin is observed in interphase cells.In the early stages of mitosis, nucleoli are intensely immunoreactive with both TG-3 and MS3. Note that the overlap in the mitotic cells is partial (A, B),suggesting that not all nucleolin is phosphorylated by Cdc2 in these cells. DAPI insert indicates cell in prometaphase (D-arrow)
522 D. Dranovsky et al. / Neurobiology of Aging 22 (2001) 517–528

segregation, which was determined by histone specificmAb053 (Figs. 4J and 4L) and DAPI staining (data notshown). Whereas the intensity of TG-3 nucleolin immuno-reactivity proceeded to decrease from metaphase to telo-phase, its spatial distribution did not change and the chromo-somes persisted to be the predominant TG-3-immunoreactivestructures (Figs. 4H, cell 3; Fig. 4K, cell 2). Cells in cytokinesisand interphase showed only background levels of immuno-staining.
3.4. TG-3 reactive and non-reactive nucleolin aredifferentially distributed during mitosis
Asynchronous HEp-2 cells were dual-labeled with phos-phoepitope specific TG-3 and with MS3 which is specificfor the region of nucleolin that is devoid of phosphorylationsites [42]. Whereas TG-3 was highly specific for nucleolinin cells in mitosis, MS3 was immunoreactive with nucleolinin all phases of the cell cycle (Fig. 5). As expected, the mostprominent MS3 immunoreactive structures in interphasecells were nuclei and nucleoli. MS3 immunoreactivity wasmore dispersed throughout the nucleus in the early stages ofmitosis. In prophase, nuclear TG-3 immunoreactivity onlypartially colocalized with the MS3 immunoreactivity (Fig.5A). In metaphase, MS3 immunoreactivity was distributedthroughout the mitotic cell. MS3 and TG-3 immunoreactiv-ity colocalized primarily around the chromosomes sincechromosomal TG-3 immunoreactivity was most pro-nounced. MS3 immunoreactivity of cytosol persisted inanaphase, telophase, and cytokinesis, while TG-3 immuno-reactivity gradually became undetectable (Fig. 5C). Thepartial overlap of MS3 and TG-3 immunoreactivities in allstages of mitosis suggests that TG3 reactive nucleolin rep-resents a unique subpopulation of mitotic nucleolin.
3.5. Differential distribution of nucleolin in the normalhuman brain and in AD
The functions of nucleolin have been extensively studiedin cultured cells and in vitro [14,41]. Nucleolin has beenimmunocytochemically localized primarily to the nucleolusand the nucleus. However, the distribution and the role ofnucleolin in human brain cells remain unknown. Therefore,the distribution of nucleolin in the brain was examined infour individuals affected by AD and in eight age-matchedcontrols. Identical immunolocalization was obtained withall examined anti-nucleolin monoclonal and polyclonal an-tibodies, strongly suggesting that the distribution of immu-noreactivity reflected the true distribution of nucleolin andthat it was not the result of cross reactivity with otherproteins. The images on panels of Fig. 6 and 7 were ob-tained with anti-nucleolin monoclonal antibody MS3 andillustrate a typical distribution of nucleolin in the hippocam-pus. The specificity of TG-3 for AD brains has been previ-ously described [44], and it is here presented for illustrativevalue (Fig. 6A and 6F).
Cells corresponding to all major areas of the hippocam-pal formation, including the dentate gyrus, were positive fornucleolin. Nucleolin was detected in both large neuronalnuclei and smaller glial nuclei (Figs. 6C and 6D). Thedistribution of nucleolin in glia appeared primarily nuclear,while neuronal nucleolin was readily detected in both cy-toplasm and nuclei. Moreover, some neurons displayedmostly cytoplasmic distribution of nucleolin, while in otherneurons, nucleolin was primarily nuclear (Fig. 6B). In ad-dition to staining the neuronal cell bodies, nucleolin wasdetected in the apical dendrites of the pyramidal cells (Fig.6D). Neurons with predominantly nuclear, and neurons withpredominantly cytoplasmic nucleolin were observed in CA3and CA4, whereas more cytoplasmic and robust dendriticstaining was observed in CA2 and CA1 (Figs. 6C and 6D).The variability in the distribution of nucleolin between cellsis indicative of a dynamic function for the protein in neu-rons. Occasionally a small number of NFT were observed inthe hippocampus from individuals who showed no cognitiveimpairments prior to their death. The appearance of theseearly pathological changes in the brain prior to developmentof clinical symptoms is believed to characterize AD pro-gression. CA1 is the earliest region in the hippocampus tobe affected by neurodegenerative changes [5]. Interestingly,an intense nucleolin immunoreactivity in densely packedmaterial typical of NFT within some pyramidal cells in theCA 1 region was observed in four of our control cases (Fig.6E, representative example).
To further characterize the association of nucleolin withAD pathology, brains from 3 individuals affected by ADwere immunostained with mAb MS3 (Figs. 6G,H,I, and J).The distribution of nucleolin in the AD brains paralleled thedistribution of nucleolin in the control brains. However, theneurons in the AD brains exhibited a marked decrease innucleolin immunoreactivity throughout the hippocampalformation (Fig. 6G and 6H). Nucleolin immunoreactivity,shown in red, labeled small granular structures in many ofthe neurons in the CA3 and CA4 regions (Fig. 6H). Nucleo-lin localized to NFT in the CA1 and CA2 regions of thehippocampal formation (Figs. 6I and 6J). Using the classi-fication of NFT proposed by Wischik (1989) [49], we foundthat the NFT immunolabeled with MS3 had the appearanceof type 1 (early intracellular), type 2 (late intracellular) andtype 3 (extracellular) NFT. Type 1 NFT were observed asintensely fluorescent, densely packed material traversing thepyramidal neuron and surrounding the nucleus. Large swol-len pyramidal neurons with intensely labeled NFT and anucleus that was positioned near the plasma membraneresembled type 2 NFT. Finally, intense filament-like stain-ing of cell remnants with no noticeable nucleus was con-sistent with type 3 NFT.
3.6. TG-3 reactive nucleolin is present in NFT
A recent study demonstrated that in AD brains, TG-3antibody recognized microtubule associated protein tau
523D. Dranovsky et al. / Neurobiology of Aging 22 (2001) 517–528

phosphorylated on Thr231 [20]. It has not been established,however, if TG-3 recognizes other proteins in AD brains.After we established that TG-3 is highly specific for mitoticnucleolin in cell cultures, it became important to determineif TG-3 immunreactive nucleolin was present in the brainsof patients with AD and, if yes, to differentiate it from TG-3immunoreactive tau.
In order to identify a population of NFTs with TG-3
reactive nucleolin, dual-label confocal microscopy was firstperformed with TG-3 and MS3 antibodies on AD cases thatshowed abundant MS3 immunoreactive NFT (Fig. 7A).There were many TG-3 immunoreactive NFT and an in-tense neuritic staining by the TG-3 antibody. MS3 stainedNFT and nuclei, but there was no staining of neurites. Thus,there were many NFT that were positive with both antibod-ies (Fig. 7A).
Fig. 6. Distribution of nucleolin in control and AD brains. Light and immunofluorescent micrographs of control (B-E) and AD (G-J) hippocampal sectionsimmunostained with monoclonal antibody MS3. Control (A) and AD (F) brain sections immunostained with TG-3 were used as a reference for illustrativevalue. CA3-CA4 (B, C): Nucleolin is present in large neuronal nuclei and neuronal cytoplasm, and in small glial nuclei (B, bar5 0.05 mm; C, bar5 0.1mm). Nucleolin is primarily nuclear in some neurons, primarily cytoplasmic in others and was detected in both compartments in still other neurons (B). InCA1 (D, F), note the intense immunoreactivity in the apical dendrites (D, bar5 0.05 mm). Note the cytoplasmic distribution of nucleolin and presence ofnucleolin in early NFT in the control brain (E, bar5 0.05 mm). A decrease in nucleolin immunoreactivity in all regions of the AD hippocampus wasconsistently observed (G-J). In CA3-CA4 note infrequently observed immunoreactive plaque-like structures (G, arrows, bar5 0.2 mm), and the granularappearance of nucleolin staining (H, bar5 0.2 mm). CA1 (I, J) - note the abundance of nucleolin positive NFT. Type 1, 2, and 3 NFT were observed (bar50.05 mm).
524 D. Dranovsky et al. / Neurobiology of Aging 22 (2001) 517–528
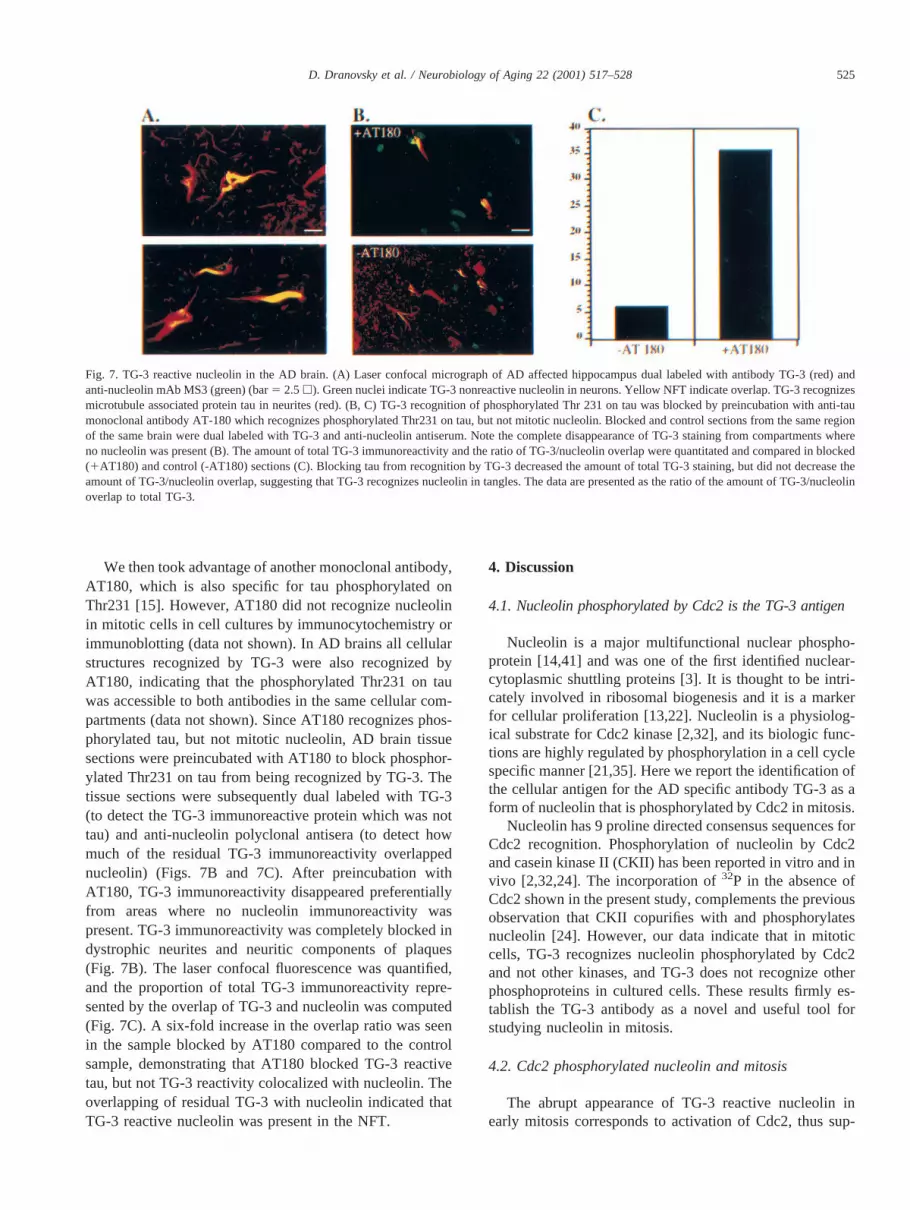
We then took advantage of another monoclonal antibody,AT180, which is also specific for tau phosphorylated onThr231 [15]. However, AT180 did not recognize nucleolinin mitotic cells in cell cultures by immunocytochemistry orimmunoblotting (data not shown). In AD brains all cellularstructures recognized by TG-3 were also recognized byAT180, indicating that the phosphorylated Thr231 on tauwas accessible to both antibodies in the same cellular com-partments (data not shown). Since AT180 recognizes phos-phorylated tau, but not mitotic nucleolin, AD brain tissuesections were preincubated with AT180 to block phosphor-ylated Thr231 on tau from being recognized by TG-3. Thetissue sections were subsequently dual labeled with TG-3(to detect the TG-3 immunoreactive protein which was nottau) and anti-nucleolin polyclonal antisera (to detect howmuch of the residual TG-3 immunoreactivity overlappednucleolin) (Figs. 7B and 7C). After preincubation withAT180, TG-3 immunoreactivity disappeared preferentiallyfrom areas where no nucleolin immunoreactivity waspresent. TG-3 immunoreactivity was completely blocked indystrophic neurites and neuritic components of plaques(Fig. 7B). The laser confocal fluorescence was quantified,and the proportion of total TG-3 immunoreactivity repre-sented by the overlap of TG-3 and nucleolin was computed(Fig. 7C). A six-fold increase in the overlap ratio was seenin the sample blocked by AT180 compared to the controlsample, demonstrating that AT180 blocked TG-3 reactivetau, but not TG-3 reactivity colocalized with nucleolin. Theoverlapping of residual TG-3 with nucleolin indicated thatTG-3 reactive nucleolin was present in the NFT.
4. Discussion
4.1. Nucleolin phosphorylated by Cdc2 is the TG-3 antigen
Nucleolin is a major multifunctional nuclear phospho-protein [14,41] and was one of the first identified nuclear-cytoplasmic shuttling proteins [3]. It is thought to be intri-cately involved in ribosomal biogenesis and it is a markerfor cellular proliferation [13,22]. Nucleolin is a physiolog-ical substrate for Cdc2 kinase [2,32], and its biologic func-tions are highly regulated by phosphorylation in a cell cyclespecific manner [21,35]. Here we report the identification ofthe cellular antigen for the AD specific antibody TG-3 as aform of nucleolin that is phosphorylated by Cdc2 in mitosis.
Nucleolin has 9 proline directed consensus sequences forCdc2 recognition. Phosphorylation of nucleolin by Cdc2and casein kinase II (CKII) has been reported in vitro and invivo [2,32,24]. The incorporation of32P in the absence ofCdc2 shown in the present study, complements the previousobservation that CKII copurifies with and phosphorylatesnucleolin [24]. However, our data indicate that in mitoticcells, TG-3 recognizes nucleolin phosphorylated by Cdc2and not other kinases, and TG-3 does not recognize otherphosphoproteins in cultured cells. These results firmly es-tablish the TG-3 antibody as a novel and useful tool forstudying nucleolin in mitosis.
4.2. Cdc2 phosphorylated nucleolin and mitosis
The abrupt appearance of TG-3 reactive nucleolin inearly mitosis corresponds to activation of Cdc2, thus sup-
Fig. 7. TG-3 reactive nucleolin in the AD brain. (A) Laser confocal micrograph of AD affected hippocampus dual labeled with antibody TG-3 (red) andanti-nucleolin mAb MS3 (green) (bar5 2.5h). Green nuclei indicate TG-3 nonreactive nucleolin in neurons. Yellow NFT indicate overlap. TG-3 recognizesmicrotubule associated protein tau in neurites (red). (B, C) TG-3 recognition of phosphorylated Thr 231 on tau was blocked by preincubation with anti-taumonoclonal antibody AT-180 which recognizes phosphorylated Thr231 on tau, but not mitotic nucleolin. Blocked and control sections from the same regionof the same brain were dual labeled with TG-3 and anti-nucleolin antiserum. Note the complete disappearance of TG-3 staining from compartments whereno nucleolin was present (B). The amount of total TG-3 immunoreactivity and the ratio of TG-3/nucleolin overlap were quantitated and compared in blocked(1AT180) and control (-AT180) sections (C). Blocking tau from recognition by TG-3 decreased the amount of total TG-3 staining, but did not decrease theamount of TG-3/nucleolin overlap, suggesting that TG-3 recognizes nucleolin in tangles. The data are presented as the ratio of the amount of TG-3/nucleolinoverlap to total TG-3.
525D. Dranovsky et al. / Neurobiology of Aging 22 (2001) 517–528

porting the in vitro phosphorylation results discussed above.A recent report indicates that nucleolin is also phosphory-lated by cell cycle kinases that are active prior to mitosis[33]. The specificity of TG-3 for cells in mitosis presents insitu evidence that TG-3 is specific for a phosphoepitopegenerated by Cdc2 and not for phosphoepitopes generatedby other kinases. TG-3 immunoreactivity appears in thenucleus at the onset of mitosis. Here we report the translo-cation of nucleolin phosphorylated by Cdc2 to the cyto-plasm during late prophase and early prometaphase.Schwab and Dreyer (1997) [35] recently demonstrated thatCdc2 phosphorylation localizes microinjected X.laevisnucleolin to the cytoplasm. The redistribution of Cdc2 phos-phorylated nucleolin to the cytoplasm in late prophase pro-vides functional in situ support for the above result andraises the possibility that phosphorylation may play a role intargeting nucleolin outside the nucleus at this stage of thecell cycle. Interestingly, the in situ detection of nucleolin inthe cytoplasm has been elusive, suggesting that under nor-mal conditions the amount of nucleolin in the cytoplasm ofcultured cells is below the level detectable by immunocy-tochemical methods. However, in polio-virus infected cells[47] and in rat intestinal epithelial cells grown on a glasssurface [51], nucleolin was found in the cytoplasm.
The localization of Cdc2 phosphorylated nucleolin to thechromosomes is consistent with previously reported obser-vations [12]. Moreover, colocalization of MS3 and TG-3immunoreactivity in the perichromosomal region suggestedthat Cdc2 phosphorylation of nucleolin could be importantfor its chromosomal interaction. It has been hypothesizedthat interaction of Cdc2 phosphorylated nucleolin with his-tone H1 plays a role in chromatin condensation in mitosis[21].
While most of the nucleolin is located in the mitoticcytosol, most of the TG-3 reactive nucleolin is localized tomitotic chromosomes. In every stage of mitosis, MS3 im-munoreactivity only partially overlapped the TG-3 immu-noreactivity, suggesting heterogeneity of mitotic nucleolin.This partial overlap also establishes that TG-3 reactivenucleolin represents a unique subpopulation of mitoticnucleolin. Functional studies with nucleolin mutated in itsCdc2 phosphorylation domains will help to elucidate thebiologic role of phosphorylation in the interaction ofnucleolin with chromosomes and in the cytoplasmic target-ing of nucleolin.
We observed a decrease in TG-3 immunoreactivity start-ing at anaphase, and it became undetectable in cytokinesis.The decrease in TG-3 immunoreactivity during mitotic exitmay be due to dephosphorylation or degradation of nucleo-lin. Recent reports suggest that nucleolin is more suscepti-ble to degradation in nonproliferating cells [our unpublishedobservations; 8]. Our observation that TG-3 immunoreac-tivity decreases in late mitosis while MS3 immunoreactivityremains unchanged suggests the possibility that TG-3 reac-tive nucleolin is first dephosphorylated and then degraded.Interestingly, theSaccharomyces pombenucleolin homolog
gar2 is essential for cytokinesis and phosphorylated byCdc2 [23].
Perhaps dephosphorylation and/or degradation of chro-mosome associated nucleolin has functional significance forchromosomal segregation or cell division. Activation ofphosphatase Cdc14 triggers mitotic exit in budding yeast[36,45]. Similar events in mammalian cells may explain thedisappearance of TG-3 phosphoepitope, which parallels theseparation of chromosomes and mitotic exit. The recentlydemonstrated participation of nucleolin in special cases ofrecombination and replication may provide clues to the roleof Cdc2 phosphorylated TG-3 immunoreactive nucleolin inmitosis [17,40,4,9]. Thus, our finding of the dynamic asso-ciation of TG-3 reactive nucleolin with mitotic chromo-somes may represent the long sought but elusive direct linkbetween nucleolus and cell cycle regulation in higher or-ganisms [7].
4.3. Nucleolin in the brain
This study represents the first characterization of thedistribution of nucleolin in the human brain. The presenteddata were highly reproducible when several polyclonal an-tisera and monoclonal antibodies to nucleolin were used onsections of human brain tissues obtained from both archivalbiopsy and autopsy material.
Three novel observations were made. First, nucleolinwas detected in neuronal cytoplasm. Second, nucleolin waspresent in NFTs, including early tangles, and NFT-associ-ated nucleolin was TG3-positive, suggesting that it wasphosphorylated by Cdc2 kinase. Third, in AD, the overalllevel of nucleolin was dramatically decreased in the tempo-ral cortex as well as in the CA1 and CA2 regions of thehippocampus. As reported by others in cultured cells [41,14,38] and as described here, nucleolin was found in nucleiand nucleoli of neurons and glia cells in the brain. Surpris-ingly, nucleolin was also present in the cytoplasm of gran-ular neurons of the dentate gyrus and pyramidal neurons ofthe hippocampus and of the lower neocortical layers. More-over, nucleolin was identified in the apical dendrites.
The finding of nucleolin in the cytoplasm was surprisingbecause it is generally believed that nucleolin, as the nameindicates, is present in the nucleolus and that it can also bedetected in the nucleus. Nucleolin has been found in thecytoplasm and plasma membrane by biochemical methods[41,14,38]. However, immunocytochemical detection visu-alized nucleolin normally in the nucleus and nucleolus ofcultured cells suggesting that the quantity of nucleolin out-side the nucleus is insufficient for its detection under normalconditions. Nucleolin can be detected in cytoplasm of cul-tured cells by immunocytochemical methods only undercertain conditions [51,47]. Yu et al. [51] detected nucleolinin the cytoplasm of rat intestinal epithelial IEC-6 cellsgrown on glass slides. However, in cells grown on laminin,nucleolin was found in the usual location of the nucleus andnucleolus. Waggoner and Sarnow (1998) [47] reported a
526 D. Dranovsky et al. / Neurobiology of Aging 22 (2001) 517–528

massive nucleolar-cytoplasmic relocalization of nucleolin inpoliovirus-infected cells. Recently a nucleolin-related pro-tein, probably nucleolin, was detected in the cytoplasm ofbovine photoreceptor cells. These studies suggest that inresponse to certain as yet unidentified signals, nucleolinmay relocalize to the cytoplasm. Thus, it is possible tospeculate that in the brain, nucleolin is localized to theneuronal cytoplam in response to signals that are absent incultured cells.
The association of nucleolin with neuronal degenerationstems from the localization of nucleolin to the NFT. It isimportant to stress that brain tissue from some of the socalled control age-matched cases, where no clinical symp-toms of AD were apparent, had a small number of nucleolinpositive, early (type 1) NFT in the CA1. Since CA1 isthought to be the earliest hippocampal region affected byNFT during the course of AD progression [5], nucleolin wasinferred to be a morphological marker for early neurofibril-lary changes.
Dual labeling experiments with antibodies to nucleolinand TG-3 yielded partial colocalization. Recognition ofhyperphosphorylated tau by TG-3 serves to explain theTG-3 immunoreactivity that does not overlap with nucleolinimmunoreactivity. Blocking of tau from being recognizedby TG-3 by preincubation with an excess of monoclonalantibody AT180 led to the disappearance of TG-3 immu-noreactivity in dystrophic neurites and neuritic componentsof plaques, thus confirming tau presence in these structures.Persistence of residual TG-3 immunoreactivity in NFT sug-gests that in NFT, TG-3 primarily recognized proteins otherthan tau. Inefficient blocking of Thr231 on tau in NFT is notlikely since AT180 is intensely immunoreactive with NFT.The colocalization of nucleolin immunoreactivity with theresidual TG-3 immunoreactivity strongly suggests thatCdc2 phosphorylated nucleolin is one of the major TG-3immunoreactive proteins in NFT.
Detection of TG-3 immunoreactive nucleolin in NFTmarks the first identification of a normal cellular substratefor mitotic kinases, which appears to undergo mitotic mod-ification in AD. Several recent reports indicating that mi-totic kinases are active in the AD brain contribute importantinformation to our understanding of mitotic mechanismsthat are reactivated in AD pathogenesis. However, in orderto understand the mechanisms of the resulting neuronaldegeneration, substrates for mitotic kinases in AD must beidentified. It is important to reemphasize that nucleolin is amarker for early (pre-clinical) neurofibrillary changes andthus its mitotic modification may prove important in theearly development of neuronal degeneration associated withAD.
The possibility that nucleolin is involved in both chro-mosomal segregation during mitosis as well as AD mayhelp to explain the observed co-occurrence of Down syn-drome (DS) and AD within certain families [19,34]. Themajority of DS cases are the result of non-dysjunction ofmaternal chromosome 21 in miosis, or a defect in chromo-
somal segregation. Therefore, the co-occurrence of DS andAD within specific families suggests a converging cellularmechanism between chromosomal segregation and neuronaldegeneration. Specifically, proteins and pathways involvedin chromosomal segregation and cell division in generalmay serve as signals for, or intermediates in, the degener-ation of post-mitotic neurons in AD. The possible involve-ment of nucleolin in mitosis and AD makes it a candidatefor such a protein.
Acknowledgments
This work was supported by NIA, Alzheimer’s Associ-ation, and Long Island Alzheimer Foundation (LIAF) toDG, AG/OD 12721 to IV and MSTP GM08444, AFAR, andLIAF fellowships to AD. The authors thank M. Cammer forassistance with confocal images; G. Jicha for assistancewith a phosphorylation experiment; Drs. C. Dingwall, andS. Strickland for critical reading of the manuscript; Drs. E.Bromet, P. Fisher, W. Quitschke, S. Simon, and W. VanNostrand for helpful comments on the manuscript.
References
[1] Baumann, K., E.M. Mandelkow, J. Biernat, H. Piwnica-Worms, andE. Mandelkow. Abnormal Alzheimer’-like phosphorylation of tau-protein by cyclin-dependent kinases cdk2 and cdk5. FEBS Lett.1993;336:417–424.
[2] Belenguer, P., M. Caizergues-Ferrer, J.C. Labbe, M. Doree, and F.Amalric. Mitosis-specific phosphorylation of nucleolin by p34Cdc2protein kinase. Mol Cell Biol. 1990;10:3607–3618.
[3] Borer, R.A., C.F. Lehner, H.M. Eppenberger, and E.A. Nigg. Majornucleolar proteins shuttle between nucleus and cytoplasm. Cell. 1989;56:379–390.
[4] Borggrefe, T., M. Wabl, A.T. Akhmedov, and R. Jessberger. AB-cell-specific DNA recombination complex. J Biol Chem. 1998;273:17025–17035.
[5] Braak H, Braak E. Staging of Alzheimer’s disease-related neurofi-brillary changes. Neurobiol Aging 1995;16:271–8.
[6] Busser, J., D.S. Geldmacher, and K. Herrup. Ectopic cell cycleproteins predict the sites of neuronal cell death in Alzheimers diseasebrain. J Neurosc. 1998;18:2801–2807.
[7] Carmo-Fonseca, M., L. Mendes-Soares, and I. Campos. To be or notto be in the nucleolus.Nature Cell Biol.2:E107–E112, 2000.
[8] Chen, C.M., S.Y. Chiang, and N.H. Yeh. Increased stability ofnucleolin in proliferating cells by inhibition of its self-cleaving ac-tivity. J Biol Chem. 1991;266:7754–7758.
[9] Daniely, Y, J.A. Borowiec. Formation of a complex between nucleo-lin and replication protein after cell stress prevents initiation of DNAreplication. J. Cell Biol. 149:799–809, 2000.
[10] Davis, F.M., T.Y. Tsao, S.K. Fowler, and P.N. Rao. Monoclonalantibodies to mitotic cells. Proc Nat Acad Sci. 1983;80:2926–2930.
[11] Deng, J.S., B. Ballou, and J.K. Hofmeister. Internalization of anti-nucleolin antibody into viable HEp-2 cells. Mol Biol Rep. 1996;23:191–195.
[12] Dundr, M., U.T. Meier, N. Lewis, D. Rekosh, M.L. Hammarskjold,and M.O.J. Olson. A class of nonribosomal nucleolar components islocated in chromosome periphery and in nucleolus-derived foci dur-ing anaphase and telophase. Chromos. 1997;105:407–417.
527D. Dranovsky et al. / Neurobiology of Aging 22 (2001) 517–528

[13] Ghisolfi, L., G. Joseph, M. Erard, J.M. Escoubas, C. Mathieu, and F.Amalric. Nucleolin–pre-rRNA interactions and preribosome assem-bly. Mol Biol Rep. 1990;14:113–114.
[14] Ginisty, H., H. Sicard, B. Roger, and P. Bouvet. Structure and func-tions of nucleolin. J Cell Sci. 1999;112:761–772.
[15] Goedert, M., R. Jakes, R.A, Crowther, P. Cohen, E. Vanmechelen, M.Vandermeeren, and P. Cras. Epitope mapping of monoclonal anti-bodies to the paired helical filaments of Alzheimer’s disease: identi-fication of phosphorylation sites in tau protein. Biochem J. 1994;301:871–877.
[16] Gorbsky GJ. Cell cycle progression and chromosome segregation inmammalian cells cultured in the presence of the topoisomerase IIinhibitors ICRF-187 [(1)-1,2-bis(3,5-dioxopiperazinyl-1-yl)propane;ADR-529] and ICRF-159 (Razoxane). Cancer Res 1994;54:1042–8.
[17] Hanakahi, L.A., L.A. Dempsey, M.J. Li, and N. Maizels. Nucleolin isone component of the B cell-specific transcription factor and switchregion binding protein, Lr1. Proc Natl Acad Sci. 1997;94:3605–3610.
[18] Harlow, E, D. Lane. Antibodies (Cold Spring Harbor: Cold SpringHarbor Laboratory), 1988.
[19] Heston LL, Mastri AR. The genetics of Alzheimer’s disease: associ-ations with hematologic malignancy and Down’s syndrome. ArchGen Psychiatry 1977;34:976–81.
[20] Jicha, G.A., E. Lane, I. Vincent, L. Otvos, Jr., R. Hoffmann, and P.Davies. A conformation- and phosphorylation-dependent antibodyrecognizing the paired helical filaments of Alzheimer’s disease.J Neurochem. 1997;69:2087–2095.
[21] Kharrat, A., J. Derancourt, M. Doree, F. Amalric, and M. Erard, M.Synergistic effect of histone H1 and nucleolin on chromatin conden-sation in mitosis: role of a phosphorylated heteromer. Biochem.1991;30:10329–10336.
[22] Lapeyre, B., H. Bourbon, and F. Amalric. Nucleolin, the majornucleolar protein of growing eukaryotic cells: an unusual proteinstructure revealed by the nucleotide sequence. Proc Natl Acad SciUSA. 1987;84:1472–1476.
[23] Leger-Silvestre, I., M.P. Gulli, J. Noaillac-Depeyre, M. Faubladier,H. Sicard, M. Caizergues-Ferrer, and N. Gas. Ultrastructural changesin the Schizosaccharomyces pombe nucleolus following the disrup-tion of the gar21 gene, which encodes a nucleolar protein structurallyrelated to nucleolin. Chromosoma. 1997;105:542–552.
[24] Li, D., G. Dobrowolska, and E.G. Krebs. The physical association ofcasein kinase 2 with nucleolin. J Biol Chem. 1996;271:15662–15668.
[25] Li, J.H., M. Xu, H. Zhou, J.Y. Ma, and H Potter. Alzheimer prese-nilins in the nuclear membrane, interphase kinetochores, and centro-somes suggest a role iIn chromosome segregation. Cell. 1997;90:917–927.
[26] Liu, W.K., R.T. Williams, F.L. Hall, D.W. Dickson, and S.H. Yen.Detection of a cdc2-related kinase associated with Alzheimer pairedhelical filaments. Am J Path. 1995;146:228–238.
[27] Mawal-Dewan, M., C.S. Parimal, M. Abdel-Ghani, D. Shalloway,and E. Racker. Phosphorylation of tau protein by purified p34 cdc28and a related protein kinase. J Biol Chem. 1992;267:19705–19709.
[28] McShea, A., P.L.R. Harris, K.R. Webster, A.F. Wahl, and M.A.Smith. Abnormal expression of the cell cycle regulators P16 andCdk4 in Alzheimerı´s disease. Am J Path. 1997;150:1933–1939.
[29] Nagy, Z., M.M. Esiri, and A.D. Smith. Expression of cell divisionmarkers in the hippocampus in Alzheimers disease and other neuro-degenerative conditions. Acta Neuropath. 1997;93:294–300.
[30] Nigg EA. Cellular substrates of p34cdc2 and its companion cyclin-dependent kinases. Trends Cell Biol 1993;3:296–301.
[31] Paudel, H., J. Lew, Z. Ali, and J.H. Wang. Brain proline-directedkinase phosphorylates tau on sites that are abnormally phosphorylatedin tau associated with Alzheimer’s disease helical filaments. J BiolChem. 1993;268:23512–23518.
[32] Peter, M., J. Nakagawa, M. Doree, J.C. Labbe, and E.A. Nigg, E.A.Identification of major nucleolar proteins as candidate mitotic sub-strates of cdc2 kinase. Cell. 1990;60:791–801.
[33] Sarcevic, B., R. Lilischkis, and R.L. Sutherland. Differential phos-phorylation of T-47d human breast cancer cell substrates by D1-, D3-,E-, and a-type cyclin-Cdk complexes. J Biol Chem. 1997;272:33327–33337.
[34] Schupf, N., D. Kapell, J.H. Lee, R. Ottman, and R. Mayeux. In-creased risk of Alzheimer’s disease in mothers of adults with Down’ssyndrome. Lancet. 1994;344:353–356.
[35] Schwab MS, Dreyer C. Protein phosphorylation sites regulate thefunction of the bipartite NLS of nucleolin. Eur J Cell Biol 1997;73:287–97.
[36] Shou, W., J.H. Seol, A. Shevchenko, C. Baskerville, D. Moazed,Z.W.S. Chen, J. Jang, A. Shevchenko, H. Charbonneau, and R.J.Deshaies. Exit from mitosis is triggered by Tem1-dependent releaseof the protein phosphatase Cdc14 from nucleolar RENT complex.Cell. 1999;97:233–244.
[37] Srivastava, M., P.J. Fleming, H.B. Pollard, and A.L. Burns. Cloningand sequencing of the human nucleolin cDNA. FEBS Lett. 1989;250:99–105.
[38] Srivastava, M, H.B. Pollard. Molecular dissection of nucleolin’s rolein growth and cell proliferation: new insights. FASEB J. 1999;13:1911–1922.
[39] Taagepera, S., M.S. Campbell, and G.J. Gorbsky. Cell-cycle-regu-lated localization of tyrosine and threonine phosphoepitopes at thekinetochores of mitotic chromosomes. Exp Cell Res. 1995;221:249–260.
[40] Thyagarajan, B., R. Lundberg, M.C. Rafferty, and C. Campbell.Nucleolin promotes homologous DNA pairing in vitro. Somatic CellMol Genet. 1998;24:263–272.
[41] Tuteja, R, N. Tuteja. Nucleolin: a multifunctional major nucleolarphosphoprotein. Crit Rev Biochem Mol Biol. 1999;33:407–436.
[42] Valdez, B. C., D. Henning, R.K. Busch, M. Srivastava, and H. Busch.Immunodominant RNA recognition motifs of human nucleolin/C23.Mol Immunol. 1995;32:1207–1213.
[43] Vincent, I., G. Jicha, M. Rosado, and D.W. Dickson. Aberrant ex-pression of mitotic cdc2/cyclin B1 kinase in degenerating neurons ofAlzheimers disease brain. J Neurosci. 1997;17:3588–3598.
[44] Vincent, I., M. Rosado, and P. Davies. Mitotic mechanisms in Alz-heimer’s disease? J Cell Biol. 1996;132:4134–4125.
[45] Visinti, R., E.S. Hwang, and A. Amon. Cfi1 prevents premature exitfrom mitosis by anchoring Cdc14 phosphatase in the nucleolus. Na-ture. 1999;398:818–823.
[46] Vulliet, R., S.M. Halloran, R.B., Braun, A.J. Smith, and G. Lee.Proline-directed phosphorylation of human tau protein. J Biol Chem.1992;267:22570–22574.
[47] Waggoner, S, P. Sarnow, P. Viral ribonucleoprotein complex forma-tion and nucleolar cytoplasmic relocalization of nucleolin in poliovi-rus-infected cells. J Virol. 1998;72:6699–6709.
[48] Wille, H., G. Drewes, J. Biernat, E.M. Mandelkow, and E. Man-delkow. Alzheimer-like paired helical filaments and antiparalleldimers formed from microtubule-associated protein tau in vitro. J CellBiol. 1992;118:573–584.
[49] Wischik CM. Cell biology of the Alzheimer tangle. Cur Op Cell Biol1989;1:115–22.
[50] Yen, S.H., A. Kenessey, S.C. Lee, and D.W. Dickson. The distribu-tion and biochemical properties of a Cdc2-related kinase, KKIALRE,in normal and Alzheimer brains. J Neurochem. 1995;65:2577–2584.
[51] Yu, D.H., M.Z. Schwartz, and R. Petryshyn. Effect of laminin on thenuclear localization of nucleolin in rat intestinal epithelial Iec-6 cells.Biochem. Biophys. Res. Comm. 247:186–192;1998.
528 D. Dranovsky et al. / Neurobiology of Aging 22 (2001) 517–528





























