Embed Size (px)
Citation preview

Strata exposed along northern Puget Soundrecord 3 late Pleistocene glacial cycles, thegreatest ice advance occurring approximately15,000 yr BP during the Fraser Glaciation. By10,000 yr BP the Fraser ice sheet had retreatednorth of the 49th parallel, leaving extensivedeposits in its wake (Armstrong et al. 1965,Crandall et al. 1965, Easterbrook 1979, 1992,1994). Whidbey Island’s central isthmus is theonly location in Puget Sound that provides adetailed stratigraphic record of the late Pleis-tocene transition from glacial till to subaerialoutwash and glaciomarine drift (Carlstad 1992).The scarcity of fossils has limited our ability toreconstruct faunal changes that occurred fol-lowing the final retreat of the Cordilleran icesheet, and Cedar Hollow is the only knowncoastal site in the Pacific Northwest that pre-serves fossils from the early Holocene (approx-imately 10,000–7500 yr BP).
The purpose of this paper is to (1) provide ageologic and paleoenvironmental perspectiveon the fossil site and (2) provide a descriptionof vertebrate paleofauna, including relationshipsto the modern fauna of the Whidbey Islandregion. We have divided responsibility for thework roughly as follows: Mustoe—collectionof specimens and organization of backgroundinformation, tables, figures, most references,and authorship of the 1st draft; Harington—
identification, description, and discussion ofmost of the larger vertebrate fossils; Morlan—identification, description, illustration, and dis-cussion of rodent fossils.
SITE DESCRIPTION
Cedar Hollow is located near Point Partridge,the westernmost projection of Whidbey Island.Coastal bluffs lie adjacent to an upland area ofextensive kettle topography (Fig. 1). At CedarHollow coastal erosion has exposed a crosssection of a kettle that is partially filled withdune sands (Fig. 2). In 1991 a brown bear (Ursusarctos) cranium was found near the base ofthese sand beds. Mustoe and Carlstad (1995)erroneously assumed the age of the fossil to belate Pleistocene based on radiocarbon ages ofmollusk shells in glaciomarine drift underlyingthe dune sand (Easterbrook 1992). We nowrecognize that the fossiliferous strata weredeposited during the early Holocene, as evi-denced by dating of a deer vertebra (Odoco-ileus sp.) collected in 1999 from the bed thatcontained the bear skull. Collagen extractedfrom the specimen (Geochron GX-25892-AMS)yielded a radiocarbon age of 8840 ± 50 yr BP(10,009 ± 97 cal yr BP; CALIB v4.1, Stuiverand Reimer 1993). The early Holocene age has
Western North American Naturalist 65(4), © 2005, pp. 429–440
CEDAR HOLLOW, AN EARLY HOLOCENE FAUNAL SITE FROM WHIDBEY ISLAND, WASHINGTON
George E. Mustoe1, C. Richard Harington2, and Richard E. Morlan3
ABSTRACT.—Coastal erosion at central Whidbey Island reveals a cross-section view of dune sands that contain theonly known record of animals that inhabited the Pacific Northwest coast during the early Holocene. Mammal fossilsinclude bones and teeth of deer, wolf, brown bear, and several species of rodents. A deer vertebra from basal bedsyielded a 14C age of 8840 ± 50 yr BP. The faunal diversity suggests that the region was repopulated relatively quickly fol-lowing the final retreat of the Cordilleran sheet. This paleofauna was quite different from that of the late Pleistocene,when the region was inhabited by mammoth, musk oxen, giant sloth, and other large mammals. Avian remains includehawks, geese, and seabirds, 3 groups that occupied quite different ecological niches. Early Holocene bird fossils fromWhidbey Island are dissimilar to late Pleistocene avifaunas from inland sites in California and Oregon, evidence that thecoastal environment offered habitat conditions quite unlike those of interior wetlands.
Key words: Holocene, Pleistocene, paleontology, Puget Sound, Whidbey Island.
1Corresponding author: Geology Department, Western Washington University, Bellingham, WA 98225.2Canadian Museum of Nature, Paleobiology, Box 3443, Station D, Ottawa, Ontario, Canada K1P 6P4.3Canadian Museum of Civilization, Box 3100, Station B, Gatineau, Québec, Canada J8X 4H2.
429

been confirmed by a radiocarbon age of 8280± 40 yr BP (Beta Analytic, Inc. 199434) deter-mined for vole bones collected approximately1 m above the level of the deer vertebra(Polenz et al. 2005).
The basal sand bed at Cedar Hollow has amaximum thickness of 3 m on the southwestslope, decreasing to 0.5 m at the kettle’s lowestpoint, and pinching out on the northeast slope(Fig. 3). Overlying this stratum is a 0.5-m-thick
430 WESTERN NORTH AMERICAN NATURALIST [Volume 65
Fig. 1. Late Pleistocene kettles are a common landform at central Whidbey Island, but Cedar Hollow is uniquebecause it has been dissected by coastal erosion.

sandy paleosol. The remainder of the kettle fillconsists of well-sorted fine sand, separatedfrom the paleosol by a fairly sharp contact.Gradual migration of the dunes in response toprevailing southeasterly winds caused the topo-graphic low to shift northwest from the origi-nal kettle bottom. No samples suitable forradiometric dating have been obtained fromupper strata, and both the sedimentation rateand the age of the youngest beds are unknown.Cedar Hollow eolian sediments date from aperiod when the climate was warmer and drierthan today, a xerithermic interval that is welldocumented by macrofossil evidence fromlacustrine sediments at Marion Lake, Vancou-ver, British Columbia, 120 km to the northeast(Wainman and Mathewes 1987; Fig. 3).
METHODS
Fossils were first discovered following the1990–91 winter after unusually severe stormseroded a channel in the face of the bluff. Forthe next several years weathering continued toexpose new specimens, but slumping of thesandy face eventually buried the fossiliferous
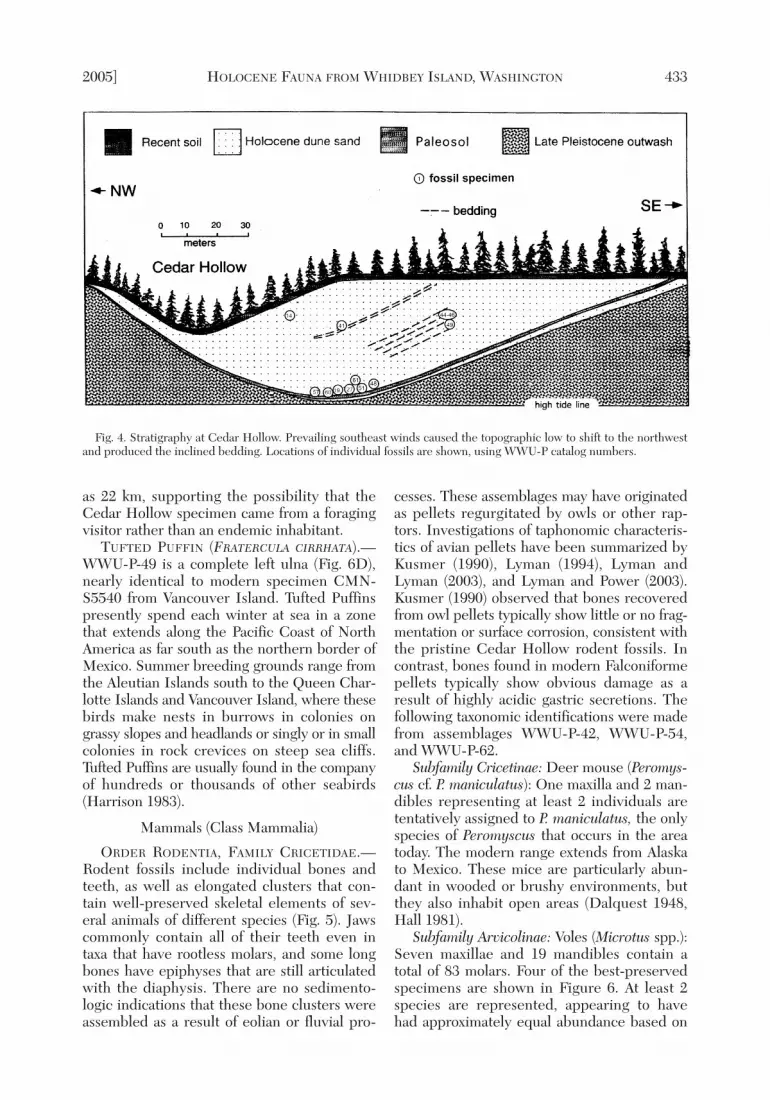
strata. Figure 4 shows stratigraphic locationsfor most specimens collected between 1992and 1995 and described in this report; notshown are a few additional fossils that werecollected from basal strata during preliminaryinvestigation. The seeming scarcity of speci-mens from middle and upper levels is partly aresult of the loose sand and vegetation thatobscures these beds. Specimens cataloged witha WWU-P prefix are part of the Western Wash-ington University paleontology research col-lection. Canada Goose and rodent specimenscollected in 1999 by Harington from the north-west flank of Cedar Hollow are labeled with aCR prefix. CMN catalog numbers indicatereference specimens in the collections of theCanadian Museum of Nature, Ottowa.
SYSTEMATIC PALEONTOLOGY
Birds (Class Aves)
CANADA GOOSE (BRANTA CANADENSIS).—CR-99-26 consists of 2 fragments of a right humerus:(1) a proximal portion running from near mid-shaft to a point just below the proximal arti-cular end showing, in anterior view, the
2005] HOLOCENE FAUNA FROM WHIDBEY ISLAND, WASHINGTON 431
Fig. 2. Cedar Hollow viewed from the intertidal zone, looking northeast.

lowermost end of the deltoid crest and the rel-atively broad convexity right of the lower 2/3of the crest, as well as the nutrient foramensome 35 mm below the deltoid crest; the speci-men compares closely in shape and size(within 0.2 mm in width and depth at midshaftand 1 mm in length, i.e., position of the nutri-ent foramen below the deltoid crest) to CMN55031, a modern Canada Goose humerus fromOntario; (2) a smaller portion corresponding tothe lower part of the shaft (with the posteriorsurface being slightly crushed), but whichdoes not quite articulate with the proximalfragment. The surface bone shows a fine net-work of rootlet tracks, suggesting that afterdeath the goose was buried in grassy surround-ings. This specimen probably represents anancestor of the dusky race that breeds alongthe southeastern coast of Alaska and makes
landfall in southwestern Washington, followingthe Willamette Valley of Oregon to its winter-ing grounds (Bellrose 1976).
LESSER SNOW GOOSE (CHEN HYPERBOREA).—Represented by 4 bones found together, allfrom the left front portion of the breast:WWU-P-43, proximal half of left scapula; P-44,proximal half of left furculum; P-45, completeleft coracoid; P-46, proximal half of left hum-erus. Size is close to that of an adult femalespecimen from Eskimo Point, Northwest Ter-ritories (CMN-ZIC-35). Whidbey Island iswithin the present wintering range of Chenhyperborea, its entire range extending fromarctic areas of Asia and North America in sum-mer, and south as far as California in winter.
HAWK (CF. ACCIPITER SP.).—WWU-P-27A isa left femur of a hawk similar in size and formto modern specimens of Sharp-shinned Hawk(Accipiter striatus) and Cooper’s Hawk (A.cooperi). Both raptors today inhabit the Whid-bey Island region, where they are usuallyfound in fields, open woodlands, and woodlandmargins. Ranges of these hawks extend fromAlaska to Mexico, where their habitats varyfrom tundra to freshwater marshes in desertregions (Clark and Wheeler 1987).
BLACK-LEGGED KITTIWAKE (RISSA TRIDAC- TYLA).—WWU-P-48 is a nearly complete lefthumerus of a gull, lacking the distal spur (Fig.6C). The maximum length of 87.0 mm is com-parable to Rissa tridactyla specimens CMN-S-6061 and CMN-S-5946 from Red Bay, Labra-dor. Members of this species presently winteroffshore along the Pacific Coast, includingWhidbey Island (Robbins et al. 1983), breed-ing on coastal cliffs and headlands of the Arc-tic Ocean in summer.
CLARK’S NUTCRACKER (NUCIFRAGA COLUM-BIANA).—WWU-P-27B is the distal 3rd of lefthumerus. The presence of Nucifraga columbianain an early Holocene coastal environment wasunexpected. The species presently winters inthe Puget Lowlands (Robbins et al. 1983). Todaythese birds inhabit coniferous forests fromsouthern British Columbia and Alberta to north-ern Baja California, most commonly being ob-served near timberline in mountain regions.They possess a sublingual pouch that allowsthem to transport seeds, an adaptation thatpermits nutcrackers to cache their entire foodsupply for their late winter nesting season.Heinrich (1989) reported that N. columbianahas been observed carrying pine seeds as far
432 WESTERN NORTH AMERICAN NATURALIST [Volume 65
Fig. 3. Late Pleistocene to modern climatic trends asindicated by plant microfossils from Marion Lake, BritishColumbia. Data from Wainman and Mathewes (1987).
as 22 km, supporting the possibility that theCedar Hollow specimen came from a foragingvisitor rather than an endemic inhabitant.
TUFTED PUFFIN (FRATERCULA CIRRHATA).—WWU-P-49 is a complete left ulna (Fig. 6D),nearly identical to modern specimen CMN-S5540 from Vancouver Island. Tufted Puffinspresently spend each winter at sea in a zonethat extends along the Pacific Coast of NorthAmerica as far south as the northern border ofMexico. Summer breeding grounds range fromthe Aleutian Islands south to the Queen Char-lotte Islands and Vancouver Island, where thesebirds make nests in burrows in colonies ongrassy slopes and headlands or singly or in smallcolonies in rock crevices on steep sea cliffs.Tufted Puffins are usually found in the companyof hundreds or thousands of other seabirds(Harrison 1983).
Mammals (Class Mammalia)
ORDER RODENTIA, FAMILY CRICETIDAE.—Rodent fossils include individual bones andteeth, as well as elongated clusters that con-tain well-preserved skeletal elements of sev-eral animals of different species (Fig. 5). Jawscommonly contain all of their teeth even intaxa that have rootless molars, and some longbones have epiphyses that are still articulatedwith the diaphysis. There are no sedimento-logic indications that these bone clusters wereassembled as a result of eolian or fluvial pro-
cesses. These assemblages may have originatedas pellets regurgitated by owls or other rap-tors. Investigations of taphonomic characteris-tics of avian pellets have been summarized byKusmer (1990), Lyman (1994), Lyman andLyman (2003), and Lyman and Power (2003).Kusmer (1990) observed that bones recoveredfrom owl pellets typically show little or no frag-mentation or surface corrosion, consistent withthe pristine Cedar Hollow rodent fossils. Incontrast, bones found in modern Falconiformepellets typically show obvious damage as aresult of highly acidic gastric secretions. Thefollowing taxonomic identifications were madefrom assemblages WWU-P-42, WWU-P-54,and WWU-P-62.
Subfamily Cricetinae: Deer mouse (Peromys-cus cf. P. maniculatus): One maxilla and 2 man-dibles representing at least 2 individuals aretentatively assigned to P. maniculatus, the onlyspecies of Peromyscus that occurs in the areatoday. The modern range extends from Alaskato Mexico. These mice are particularly abun-dant in wooded or brushy environments, butthey also inhabit open areas (Dalquest 1948,Hall 1981).
Subfamily Arvicolinae: Voles (Microtus spp.):Seven maxillae and 19 mandibles contain atotal of 83 molars. Four of the best-preservedspecimens are shown in Figure 6. At least 2species are represented, appearing to havehad approximately equal abundance based on
2005] HOLOCENE FAUNA FROM WHIDBEY ISLAND, WASHINGTON 433
Fig. 4. Stratigraphy at Cedar Hollow. Prevailing southeast winds caused the topographic low to shift to the northwestand produced the inclined bedding. Locations of individual fossils are shown, using WWU-P catalog numbers.

the fossils that we collected. Four maxillaehave dentitions characteristic of Microtus penn-sylvanicus (meadow vole). Three specimenshave dentitions similar to that of Microtustownsendii (Townsend’s vole). The mandibularmolars exhibit considerable morphologicalvariation that makes species assignment diffi-cult. Many of the M1 molars fall within therange of variation seen in M. pennsylvanicus, awidespread, highly variable species (Barnosky1990, 1994). Some M1s are most similar to M.townsendii.
The genus Microtus is listed among manyfaunal assemblages from archaeological sitesin Washington, but it is rarely identified tospecies (Morlan 2002). Microtus pennsylvanicusno longer occurs west of the Cascade Range,but this species is locally abundant on theColumbia Plateau of northeastern Washington,where individuals inhabit marshes and dampmeadows (Dalquest 1948, Maser and Storm1970). There is no obvious reason for the appar-ent contraction of its range during the Holocene.Microtus townsendii inhabits some Puget Soundislands where it lives in meadows and saltmarshes and often nests beneath driftwood(Dalquest 1948). The species has been reportedpreviously in late Holocene faunal assemblageson southernmost Vancouver Island (Blacklaws1979, Hanson 1991) and at the Umpqua-Edensite on the Oregon coast (Lyman 1991).
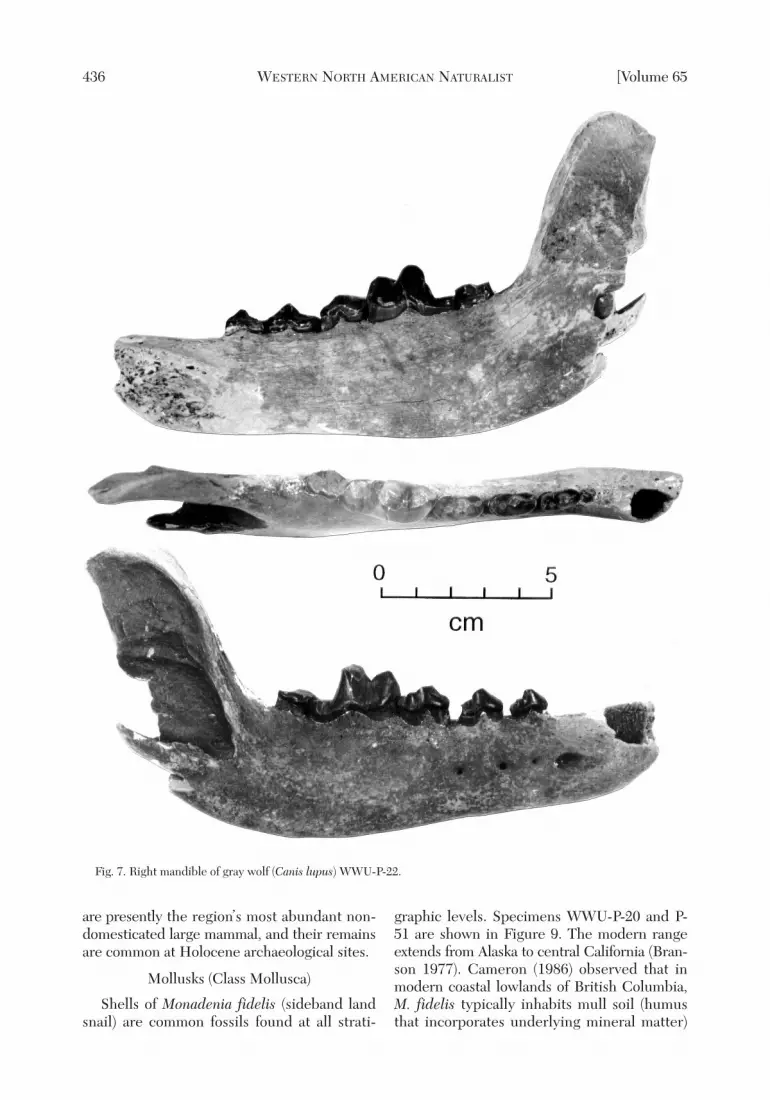
ORDER CARNIVORA.—Gray wolf (Canislupus): Specimen WWU-P-22 is a right man-
dible (Fig. 7) found on the upper beach at FortEbey picnic area near the trailhead to LakePondilla, at the base of eolian sand beds thatare correlative to Cedar Hollow, 2.5 km to thesoutheast. Five teeth (RP2-RM2) are preserved,with alveoli for M3, C1, and 2 incisors (Table 1).
Nowak (1983) suggested that wolves mi-grated to the Cascade Range from the south-ern Great Plains at the close of the Pleisto-cene, but bones from a variety of wolf speciesfound at Rancho La Brea and McKittrick, Cal-ifornia (Merriam 1912, Schultz 1938) and FossilLake, Oregon (Elftman 1931) indicate thatwolves were well established in the westernUnited States by this time. Prior to eradicationefforts that began in the late 19th century,wolves were the region’s most abundant carni-vore; by 1939 Washington’s C. lupus populationhad dwindled to only 10 individuals (Lauferand Jenkins 1989). The Whidbey Island mandi-ble most likely represents an ancestor of theCascade Mountain wolf, Canis lupus fuscusRichardson (1839).
Brown bear (Ursus arctos): Cedar Hollowprovides the 1st evidence of U. arctos in west-ern Washington from the early Holocene. Thespecimen (WWU-P-60; Fig. 8) consists of alarge cranial fragment, including 4 teeth. Threedetached incisors were found in the adjacentsediment. The dentition compares closely tomodern specimens of Ursus arctos horribilis(Mustoe and Carlstad 1995).
Harington (1989) noted that brown bearshad reached northwestern North America from
434 WESTERN NORTH AMERICAN NATURALIST [Volume 65
Fig. 5. These rodent remains (WWU-P-62) consist of a cluster of bones from several individuals distributed in an elon-gate pattern in sandy matrix, perhaps having originated as a regurgitated pellet from a raptor.

Asia by middle Wisconsinan time when loweredsea level created a land connection (BeringIsthmus) between Siberia and Alaska. This in-terpretation is based on 2 U. arctos specimensfrom the Yukon that yielded radiocarbon agesof 41,000 ± 1050 and 36,000 ± 1150 yr BP.Kurtén (1976) believed that brown bearsmigrated to the Pacific Northwest between12,000 and 11,000 yr BP. However, Churcherand Morgan (1976) identified a partial humerusfrom middle Wisconsinan till at Woodbridge,Ontario, as U. arctos, which they interpreted asevidence that the migration of this speciesfrom the Beringia refugium had begun by40,000 to 50,000 yr BP. Holocene history of U.arctos in the Pacific Northwest is poorly known.Of 110 archaeological sites in eastern and cen-
tral Washington that preserve faunal remains,only 5 contain brown bear bones (Lyman 1986).
ORDER ARTIODACTYLA.—(Columbian black-tailed deer (Odocoileus cf. O. hemionus colum-bianus): Specimen WWU-P-57 is an atlas verte-bra centrum collected approximately 1 mabove the kettle’s basal contact. Collagen ex-tracted from this bone yielded a radiocarbonage of 8840 ± 50 yr BP (GX-25892-AMS).WWU-P-41 is the distal half of a left humerus,and WWU-P-14 is the diaphysis and fused dis-tal epiphysis of a left tibia. The proximal endof the latter specimen has been crushed by acarnivore or omnivore. These deer bones arepresumed to represent O. hemionus columbi-anus because this is the only subspecies knownto have inhabited the Puget Lowlands. They
2005] HOLOCENE FAUNA FROM WHIDBEY ISLAND, WASHINGTON 435
Fig. 6. Dentition of vole specimens from Cedar Hollow: A, maxillary molars of Townsend’s vole (Microtus townsendii);B, C, maxillary molars of meadow vole (M. pennsylvanicus); D, E, mandibular molars presumed to be from M.townsendii.
are presently the region’s most abundant non-domesticated large mammal, and their remainsare common at Holocene archaeological sites.
Mollusks (Class Mollusca)
Shells of Monadenia fidelis (sideband landsnail) are common fossils found at all strati-
graphic levels. Specimens WWU-P-20 and P-51 are shown in Figure 9. The modern rangeextends from Alaska to central California (Bran-son 1977). Cameron (1986) observed that inmodern coastal lowlands of British Columbia,M. fidelis typically inhabits mull soil (humusthat incorporates underlying mineral matter)
436 WESTERN NORTH AMERICAN NATURALIST [Volume 65
Fig. 7. Right mandible of gray wolf (Canis lupus) WWU-P-22.

in forests of Thuja plicata (western red cedar)and deciduous hardwoods. Perhaps these fos-sils represent shells of dead snails that weretransported into the kettle by wind or rain, buttheir abundance suggests that they were partof a flourishing local population.
DISCUSSION
The early Holocene faunal history of westernNorth America is poorly known (Lyman andLivingston 1983, Graham and Mead 1987).Most early Holocene fossil sites in Washingtonand Oregon are located east of the Cascade
Range at latitudes well south of the maximumextent of the Cordilleran ice sheet, where theabsence of topographic and climatic barriersallowed a long history of faunal interchangewith the mid-continent region. The CedarHollow paleofauna is modern in character, verydifferent from the fauna of the late Pleistocenewhen the Puget Lowlands were inhabited byBison, Mammuthus (mammoth), Rangifer (cari-bou), Symbos (musk oxen), Megalonyx (giantsloth), and other large mammals (Harington1975, 1996, McDonald 1998). The Manis Masto-don site near Sequim on the northern OlympicPeninsula (Gustafson et al. 1979, Petersen etal. 1983, Gustafson 1985, Bergland and Marr1988) provides a useful comparison. Located50 km southwest of Cedar Hollow, bog depositsat Manis date from 11,000 to 12,000 yr BP andpreserve remains of mastodon, caribou, andmuskrat, as well as human artifacts. Mammothbones and teeth have been found in other latePleistocene deposits in the local area (Haring-ton 1975, Barton 1998). The final retreat of theCordilleran ice sheet was accompanied by theextinction of most large mammals, and theCedar Hollow paleofauna provides evidenceof repopulation of the region. Although Canislupus and Ursus arctos were driven to extinc-tion during the past century by humans, and
2005] HOLOCENE FAUNA FROM WHIDBEY ISLAND, WASHINGTON 437
Fig. 8. Cranium and teeth of brown bear (Ursus arctos) WWU-P-60.
TABLE 1. Tooth dimensions of a gray wolf (Canis lupus)right mandible, WWU P-22.
Maximum Maximumantero-posterior transverse
Tooth diameter (mm) diameter (mm)
P2 12.5 6.0P3 14.5 7.5P4 15.0 8.5M1 28.0 13.0M2 12.0 9.0
P = premolar, M = molar, mandible length = 180 mm.

Microtus pennsylvanicus no longer resides westof the Cascade Range, all other Cedar Hollowfossils are from species that still inhabit cen-tral Whidbey Island.
The abundance and diversity of bird bonesat Cedar Hollow is noteworthy. The avifaunaincludes 3 types of birds: raptors that probablyfed on small rodents that inhabited the tree-less coastal plain, geese that consumed low-land vegetation, and marine birds that primar-ily fed on fish. Early Holocene avifaunas arescarce and clues for interpreting these fossilscome from late Pleistocene sites at Rancho LaBrea and McKittrick, California and FossilLake, Oregon. These deposits all demonstratethe close relationship between avian remainsand local environmental conditions. Asphalt de-posits at Rancho La Brea and McKittrick pre-serve abundant avian remains of scavengersand raptors that were attracted by creaturestrapped in the sticky tar. These deposits com-monly preserve entire bird skeletons, in con-trast to the disarticulated bones found at mostother sites. Rancho La Brea has also yieldedremains of a diverse array of passerines (perch-ing birds) that indicate open meadows, brush,and woodlands surrounding the asphalt seeps.Bones of ducks, geese, grebes, herons, andother birds constitute only a minor part of theavifauna (Howard and Miller 1939, Stock 1958).McKittrick beds are also rich in predators andscavengers, but 67% of the bird remains comefrom water birds, representing a site whereasphalt pools were part of a much larger wet-land environment (Miller 1935). Fossil Lake,
Oregon, is similar in age to the California asphaltbeds, but the avifauna of this large inland lakeis very different: 91% of the specimens are fromwater birds, and raptor remains comprise lessthan 1% (Miller 1911, Howard 1946, Allison1966).
Raptors may have been year-round residents,but the other birds whose remains are pre-served at Cedar Hollow presumably migratedgreat distances between their winter rangesand their summer breeding grounds. Presently,Tufted Puffins, Canada Geese, Lesser SnowGeese, and Black-legged Kittiwake breed atnorthern latitudes ranging from British Colum-bia to Arctic Alaska and travel as far south asCalifornia during the winter. Do avian remainsat Cedar Hollow, Rancho La Brea, McKittrick,and Fossil Lake represent birds that were win-ter residents, or summer breeding populationsthat come from a time when migratory speciesnested far to the south of their present nestinggrounds? The presence of continental glaciersmust have been a major influence on flightpaths and nesting areas, but no research hasbeen conducted on avian migration patternsduring the Pleistocene/Holocene transition.Cedar Hollow fossils show that several generaof migratory birds inhabited the Puget Soundregion during the early Holocene, but evidencefrom other locations will be needed before theseasonal cycle and the travel routes of thesebirds can be determined.
CONCLUSIONS
With the exception of Ursus americanus(black bear) skeletons discovered in a cave onVancouver Island (Nagorsen et al. 1995), CedarHollow contains the only known early Holocenepaleofauna west of the Cascade Range. Theseremains provide evidence of the faunal transi-tions that occurred following the final retreatof continental glaciation at the close of thePleistocene. Cedar Hollow is bordered on thewest by the Pacific Ocean, on the east by theCascade Mountains, and to the north by theCordilleran ice sheet that persisted untilapproximately 10,000 yr BP (Armstrong et al.1965, Easterbrook 1992). Despite these obstruc-tions to faunal migration, the Cedar Hollowfossils show that the ecosystem that developedwithin 2000 years following deglaciation waspopulated by many of the species that pres-ently inhabit the region.
438 WESTERN NORTH AMERICAN NATURALIST [Volume 65
Fig. 9. Shells of the land snail (Monadenia fidelis) areamong the most common fossils found at Cedar Hollow.Specimens WWW-P-20 and P-51.

ACKNOWLEDGMENTS
Cynthia Carlstad and Don Easterbrook(Western Washington University) discoveredthe fossil site. Bax Barton (University of Wash-ington Quaternary Research Center) identi-fied specimen WWU-P14 as Odocoileus cf. O.hemionus columbianus, and Alicia M. Corderoand Barry Roth (Department of IntegrativeBiology, University of California at Berkeley)confirmed the identification of land snails asMonadenia fidelis. Elaine Mustoe and SamuelGirouard, Jr., helped with fieldwork. MichaelPolenz provided additional radiocarbon agedata. The following Canadian Museum of Na-ture staff members deserve credit: DarleneBalkwill and Michel Gosselin (for the loan ofcomparative specimens of modern vertebrates)and Donna Naughton and Clayton Kennedy(fossil preparation). Gail Harington word pro-cessed the initial manuscript. R.A.D. Cameron,R.L. Lyman, and J.M. Rensberger providedhelpful reviews.
LITERATURE CITED
ALLISON, I.S. 1966. Fossil Lake, Oregon: its geology andfossil faunas. Oregon State University Monographs,Studies in Geology 9.
ARMSTRONG, J.E., D.R. CRANDELL, D.J. EASTERBROOK, AND
J.B. NOBLE. 1965. Late Pleistocene stratigraphy andchronology in southwestern British Columbia andnorthwestern Washington. Geological Society ofAmerica Bulletin 76:321–330.
BARNOSKY, A.D. 1990. Evolution of dental traits since lat-est Pleistocene in meadow voles (Microtus pennsyl-vanicus) from Virginia. Paleobiology 16:370–383.
______. 1994. Defining climate’s role in ecosystem evolu-tion: clues from late Quaternary mammals. Histori-cal Biology 8:173–190.
BARTON, B.R. 1998. Notes on the new Washington Statefossil, Mammuthus columbi. Washington Geology26(2/3):68.
BELLROSE, F.C. 1976. Ducks, geese, and swans of NorthAmerica. Stackpole Books, Harrisburg, PA. 540 pp.
BERGLAND, E.O., AND J. MARR. 1988. Prehistoric life onthe Olympic Peninsula: the first inhabitants of agreat American wilderness. Pacific Northwest NationalParks and Forests Association, Seattle, WA. 88 pp.
BLACKLAWS, R.W. 1979. Excavations at Esquimalt Lagoon:a contribution to Straits Salish prehistory. Master’sthesis, Simon Fraser University, Burnaby, BritishColumbia.
BRANSON, B.A. 1977. Freshwater and terrestrial Molluscaof the Olympic Peninsula, Washington. Veliger 19:310–330.
CAMERON, R.A.D. 1986. Environment and diversities offorest snail faunas from central British Columbia.Malacologia 27:342–355.
CARLSTAD, C.A. 1992. Late Pleistocene deglaciation his-tory at Point Partridge, central Whidbey Island, Wash-
ington. Master’s thesis, Western Washington Univer-sity, Bellingham.
CHURCHER, C.S., AND A.V. MORGAN. 1976. A grizzly bearfrom the middle Wisconsin of Woodbridge, Ontario.Canadian Journal of Earth Sciences 13:341–347.
CLARK, W.S., AND B.K. WHEELER. 1987. Field guide tohawks of North America. Houghton Mifflin, Boston,MA. 316 pp.
CRANDALL, D.R., D.J. EASTERBROOK, AND J.B. NOBLE. 1965.Late Pleistocene stratigraphy and chronology in south-western British Columbia and northwestern Wash-ington. Geological Society of America Bulletin 76:321–330.
DALQUEST, W.W. 1948. Mammals of Washington. Univer-sity of Kansas Publications, Museum of Natural His-tory 2:1–444.
EASTERBROOK, D.J. 1979. The last glaciation of northwestWashington. Pages 177–189 in J.M. Armentrout, M.R.Cole, and H. Terbest, editors, Cenozoic paleogeog-raphy of the western United States. Pacific CoastPaleogeography Symposium 3, Pacific Coast Section,Society of Economic Paleontologists and Mineralo-gists, Los Angeles, CA.
______. 1992. Advance and retreat of Cordilleran ice sheetsin Washington, U.S.A. Géographie Physique et Qua-ternaire 46:51–66.
______. 1994. Stratigraphy and chronology of early to latePleistocene glacial and interglacial sediments in thePuget Lowland, Washington. Pages 1–38 in D.A.Swanson, and R.A. Haugerud, editors, Geologic fieldtrips in the Pacific Northwest. 1994 Geological Soci-ety of America Annual Meeting, Seattle, WA.
ELFTMAN, H.O. 1931. Pleistocene mammals of Fossil Lake,Oregon. American Museum Novitiates, New York.21 pp.
GRAHAM, R.W., AND J.I. MEAD. 1987. Environmental fluc-tuations and evolution of mammalian faunas duringthe last deglaciation in North America. Pages 371–402 in W.F. Ruddiman and H.E. Wright, editors, Thegeology of North America, volume K-3. North Amer-ica and adjacent oceans during the last deglaciation.Geological Society of America, Boulder, CO.
GUSTAFSON, C.E. 1985. The Manis Mastodon site. NationalGeographic Research Reports 20:283–295.
GUSTAFSON, C.E., D. GILBOW, AND R.D. DAUGHERTY. 1979.The mastodon site: early man on the Olympic Penin-sula. Canadian Journal of Archaeology 3:157–164.
HALL, E.R. 1981. The mammals of North America. JohnWiley and Sons, New York.
HANSON, D.K. 1991. Late prehistoric subsistence in theStrait of Georgia region of the Northwest Coast.Doctoral dissertation, Simon Fraser University,Burnaby, BC.
HARINGTON, C.R. 1975. Pleistocene muskoxen (Symbos)from Alberta and British Columbia. Canadian Jour-nal of Earth Sciences 12:903–919.
______. 1989. Pleistocene vertebrate localities in the Yukon.Pages 93–98 in L.D. Carter, T.D. Hamilton, and J.P.Galloway, editors, Late Cenozoic history of the inte-rior basins of Alaska and the Yukon. U.S. GeologicalSurvey Circular 1026, Washington, DC.
______. 1996. Quaternary animals: vertebrates of the iceage. Pages 259–273 in R. Ludvigsen, editor, Life instone: a natural history of British Columbia’s fossils.University of British Columbia Press, Vancouver.
HARRISON, P. 1983. Seabirds: an identification guide.Houghton Mifflin, Boston, MA. 48 pp.
2005] HOLOCENE FAUNA FROM WHIDBEY ISLAND, WASHINGTON 439

HEINRICH, B. 1989. Ravens in winter. Vintage Books, NewYork. 379 pp.
HOWARD, H. 1946. A review of the Pleistocene birds ofFossil Lake, Oregon. Carnegie Institution of Wash-ington Publication 551(8):141–195.
HOWARD, H., AND A.H. MILLER. 1939. The avifauna asso-ciated with human remains at Rancho La Brea, Cali-fornia. Carnegie Institution of Washington Publica-tion 514(3):39–48.
KURTÉN, B. 1976. The cave bear story. Columbia Univer-sity Press, New York. 163 pp.
KUSMER, K.D. 1990. Taphonomy of owl pellet deposition.Journal of Paleontology 64:629–637.
LAUFER, J.R., AND P.T. JENKINS. 1989. A preliminary studyof gray wolf history and status in the region of theCascade Mountains of Washington State. Unpub-lished report, Washington Wolf Project, Wolf HavenAmerica, Tenino, WA.
LYMAN, R.L. 1986. On the Holocene history of Ursus ineastern Washington. Northwest Science 60:67–72.
______. 1991. Prehistory of the Oregon coast: the effectsof excavation strategies and assemblage size on archae-ological inquiry. Academic Press, Inc., San Diego,CA. 391 pp.
______. 1994. Relative abundance of skeletal specimensand taphonomic analysis of vertebrate remains. Palaios9:288–298.
LYMAN, R.L., AND S.D. LIVINGSTON. 1983. Late Quaternarymammalian zoogeography of eastern Washington.Quaternary Research 20:360–373.
LYMAN, R.L., AND R.J. LYMAN. 2003. Lessons from tempo-ral variation in the mammalian faunas from two col-lections of owl pellets in Columbia County, Washing-ton. International Journal of Osteoarchaeology 13:150–156.
LYMAN, R.L., AND E. POWER. 2003. Quantification andsampling of faunal remains in owl pellets. Journal ofTaphonomy 1:3–14.
MASER, C., AND R.M. STORM. 1970. A key to Microtinae ofthe Pacific Northwest (Oregon, Washington, Idaho).Oregon State University, Corvallis. 162 pp.
MCDONALD, H.G. 1998. The sloth, the president, and theairport. Washington Geology 26(1):40–42.
MERRIAM, J.C. 1912. The fauna of Rancho La Brea. PartII. Canidae. Memoirs of the University of California1:215–272.
MILLER, L.H. 1911. A synopsis of our knowledge concern-ing the fossil birds of the Pacific Coast of NorthAmerica. Condor 13:117–118.
______. 1935. A second avifauna from the McKittrick Plei-stocene. Condor 37:72–79.
MORLAN, R.E. 2002. Canadian archaeological radiocarbondatabase. Available online at www.canadianarchaeol-ogy.com/card.
MUSTOE, G.E., AND C.A. CARLSTAD. 1995. A late Pleisto-cene brown bear (Ursus arctos) from northwestWashington. Northwest Science 9:106–113.
NAGORSEN, D.W., G. KEDDIE, AND R.J. HEBDA. 1995. EarlyHolocene black bears (Ursus americanus) from Van-couver Island. Canadian Field-Naturalist 109:11–18.
NOWAK, R.M. 1983. A perspective on the taxonomy of wolvesin North America: wolves in Canada and Alaska.Canadian Wildlife Service Report 45:10–29.
PETERSEN, K.L., P.J. MEHRINGER, JR., AND C.E. GUSTAF-SON. 1983. Late-glacial vegetation and climate at theManis Mastadon site, Olympic Peninsula, Washing-ton. Quaternary Research 20:215–231.
POLENZ, M., S.L. SLAUGHTER, AND G. W. THORSEN. 2005.Geologic map of the Coupeville and part of the Port Townsend North 7.5-minute quadrangle, IslandCounty, Washington. Washington Division of Geol-ogy and Earth Resources Map GM-58, Olympia, WA.
RICHARDSON, J. 1839. The zoology of Captain Beechey’sVoyage of the Blossom, 1825–28. H.G. Bonn, Lon-don. 186 pp.
ROBBINS, C.S., B. BRUUN, H.S. ZIM, AND A. SINGER. 1983.Birds of North America. Golden Press, New York.360 pp.
SCHULTZ, J.R. 1938. A late Quaternary mammal faunafrom the tar seeps of McKittrick, California. CarnegieInstitution of Washington Publication 487(4):118–161.
STOCK, C. 1958. Rancho La Brea: a record of Pleistocenelife in California. 7th edition. Science Series 37, Nat-ural History Museum of Los Angeles County, LosAngeles, CA. 83 pp.
STUIVER, M., AND P.J. REIMER. 1993. Extended 14C data-base and revised CALIB radiocarbon calibrationprogram. Radiocarbon 35:215–230.
WAINMAN, N., AND R.W. MATHEWES. 1987. Forest historyof the last 12,000 years based on plant macrofossilanalysis of sediment from Marion Lake, southwest-ern British Columbia. Canadian Journal of Botany65:2179–2187.
Received 8 March 2005Accepted 24 January 2005
440 WESTERN NORTH AMERICAN NATURALIST [Volume 65





























