Embed Size (px)
Citation preview

Available online at www.sciencedirect.com
www.elsevier.com/locate/brainres
b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 6
http://dx.doi.org/100006-8993/& 2014 El
Abbreviations: 4V
AIN, anterior inter
interpositus area; D
DLP, dorsolateral p
GL, granular layer
IO, inferior olive; I
subdivision of the l
MN-CM, caudome
fasciculus; MVN, m
cell; PCG, pontine
PIN-parvo, ventral
reticular nucleus; p
RPo, raphe pontis;
nucleus; TMB, 3,30
Y, nucleus YnCorresponding autE-mail address: i
Research Report
Cerebellar afferents originating from the medullaryreticular formation that are different from mossy,climbing or monoaminergic fibers in the rat
Yuanjun Luo, Izumi Sugiharan
Department of Systems Neurophysiology and Center for Brain Integration Research, Tokyo Medical and Dental UniversityGraduate School, 1-5-45 Yushima, Bunkyo-ku, Tokyo 113-8519, Japan
a r t i c l e i n f o
Article history:
Accepted 12 April 2014
Integration of cortical Purkinje cell inputs and brain stem inputs is essential in generating
cerebellar outputs to the cerebellar nuclei (CN). Currently, collaterals of climbing and
Available online 18 April 2014
Keywords:
Precerebellar nucleus
Cerebellar mossy fiber
Cerebellar afferent
Raphe interpositus
Biotinylated dextran amine
Single axon reconstruction
.1016/j.brainres.2014.04.02sevier B.V. All rights res
, forth ventricle; 5-HT,
posed nucleus; BDA, bi
, dorsal; DAB, 3,30-dia
rotuberance; DMC, dor
; GRN, gigantocellular r
N, interposed nucleus;
ateral nucleus; Lt, left;
dial subdivision of the m
edial vestibular nucleus
central gray; PCL, Purkin
parvocellular subdivisio
y, pyramidal tract; PN,
RPa, raphe pallidus; Rt
,5,50-tetramethylbenzidin
hor. Fax: þ81 3 5803 [email protected]
a b s t r a c t
mossy fiber axons, noradrenergic, serotoninergic and cholinergic axons, and collaterals of
rubrospinal axons are known to innervate the CN from the brain stem. We investigated
whether other afferents to the CN from the medulla exist in the rat. Retrograde labeling
revealed the presence of neurons that project to the CN but not to the cerebellar cortex in
the median reticular formation in the rostrodorsal medulla (tentatively named ‘caudal
raphe interpositus area’, CRI). Anterograde tracer injection into the CRI labeled abundant
axonal terminals in the CN, mainly in the ventral parvocellular part of the posterior
interposed and lateral nucleus. Axonal reconstruction showed that a single CRI axon
projected to the CN with 170–1086 varicosities, more broadly and densely than collaterals
of a mossy or climbing fiber axon. CRI axons had no or a few collaterals that projected to
the granular and Purkinje cell layers of the cerebellar cortex with some small terminals,
indicating that these axons are different from mossy fiber axons. CRI axons also had
0erved.
5-hydroxytriptamine; V, trigeminal nucleus; VI, abducens nucleus; VII, facial nucleus;
otinylated dextran amine; C, caudal; CN, cerebellar nuclei; CRI, caudal raphe
minobenzidine; DLH, dorsolateral hump of the anterior interposed nucleus;
somedial crest of the anterior interposed nucleus; gVII, genu of the facial nerve;
eticular nucleus; HRP, horseradish peroxidase; icp, inferior cerebellar peduncle;
L, lateral; L.II–VI, lobules II–VI; LN, lateral nucleus; LN-parvo, ventral parvocellular
LVN, lateral vestibular nucleus; M, medial; ML, molecular layer; MN, medial nucleus;
edial nucleus; MN-M, middle subdivision of the medial nucleus; mlf, medial longitudinal
; nVII, tract of the facial nerve; NRTP, nucleus reticularis tegmenti pontis; PC, Purkinje
je cell layer; PHA-L, Phaseolus vulgaris leucoagglutinin; PIN, posterior interposed nucleus;
n of the posterior interposed nucleus; PrH, prepositus hypoglossi nucleus; PRN, pontine
pontine nucleus; R, rostral; RM, nucleus raphe magnus; RO, raphe obscurus;
, right; scp, superior cerebellar peduncle; SO, superior olive; SVN, superior vestibular
e; Tz, trapezoid body; V, ventral; VN, vestibular nuclei; WM, white matter;
.(I. Sugihara).

b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 632
collaterals that projected to the medial vestibular nucleus and an ascending branch that
was not reconstructed. The location of the CRI, electron microscopic observations, and
immunostaining results all indicated that CRI axons are not monoaminergic. We conclude
that CRI axons form a type of afferent projection to the CN that is different from mossy,
climbing or monoaminergic fibers.
& 2014 Elsevier B.V. All rights reserved.
1. Introduction
Purkinje cell axons form the most dominant projection to theCN (Chan-Palay, 1973), and the CN, in turn, are the majorsource of outputs from the cerebellum. However, sincePurkinje cell axons are inhibitory, how their activity leads togeneration of cerebellar output is not simple (Person andRaman, 2012). Besides Purkinje cell axons, the CN receiveafferent projections from the brain stem and spinal cord.Thus, CN activity is likely to reflect the integration of Purkinjecell activity with the activity of these afferents.
The sources of brain stem projections to the CN, inparticular, have been collectively labeled by retrograde tra-cers injected into the CN (Eller and Chan-Palay, 1976; Okaet al., 1985; Newman and Ginsberg, 1992). Several brain stemnuclei (nucleus reticularis tegmenti pontis, pontine nuclei,inferior olive, trigeminal nuclear complex, lateral reticularnucleus and some other nuclei in the brain stem) have beenidentified as possible sources of afferents to the CN. Sincethese nuclei are the origins of mossy and climbing projec-tions to the cerebellar cortex as well, the possibility of traceruptake through passing axons that project to the cerebellarcortex, but not to the CN, is hard to exclude. Thus, electro-physiological recording of synaptic responses from CN neu-rons or anterograde tracing studies of CN projecting axonsare required for conclusive demonstration of afferent projec-tions to the CN. Such studies have shown that almost allclimbing fiber axons (Kitai et al., 1977; Shinoda et al., 1987;Van der Want et al., 1989; Sugihara et al., 2001) and somemossy fiber axons (Matsushita and Yaginuma, 1995; Wu et al.,1999; Shinoda et al., 1992; Quy et al., 2011) give rise tocollaterals that provide excitatory innervation of the CN.However, it seems that the frequency of occurrence of CNcollaterals differs among mossy fiber axons originating fromdifferent sources.
Besides collaterals of mossy and climbing fibers, mono-aminergic (serotoninergic and noradrenergic) projectionshave been reported in the CN (Moore and Bloom, 1979;Takeuchi et al., 1982; Bishop and Ho 1985) as well as in thecerebellar cortex, on the basis of specific labeling by immu-nostaining. These projections presumably exert a modulatoryaction on the CN neurons (Murano et al., 2011). It is not clearwhether the monoaminergic projections to the CN and thecerebellar cortex originate from the same set of axons.
Cholinergic projections in the CN appear as fine varicoseaxons following immunostaining of choline acetyltrans-ferase (Woolf and Butcher, 1989). The pedunculopontine andlaterodorsal tegmental nuclei, the major sources of choliner-gic fibers in the brain stem, project to the CN (Woolf andButcher, 1989); however, the major cholinergic innervationof the cerebellum arises from the medial vestibular nucleus
(Barmack et al., 1992), the lateral paragigantocellular nucleus,and the raphe obscurus nucleus (Jaarsma et al., 1997). Whilethe cholinergic projection from the vestibular nucleus takesthe form of mossy fibers, other cholinergic projections have avaricose termination pattern in the cortex and/or CN. It isgenerally not clear whether the cholinergic projections tothe CN and to the cerebellar cortex originate from the sameaxons.
Except for collaterals of mossy and climbing axons andmonoaminergic and cholinergic axons, information on thebrain stem afferents to the CN is relatively limited. However,one retrograde fluorescent labeling study that focused on CNafferents showed that some spinal-projecting neurons of themagnocellular red nucleus supply collaterals to the CN, butnot to the cerebellar cortex (Huisman et al., 1983). An inputthat projects primarily to the CN but not to the cerebellarcortex can directly modulate the cerebellar output withoutaffecting information processing in the cerebellar cortex.In order to look into presence of such an input from otherareas, we performed a combination of retrograde and ante-rograde labeling experiments on afferents to the CN from themedulla. We identified a new cerebellar afferent system thatpredominantly targets the CN rather than the cerebellarcortex. Single axonal morphology, electron microscopy, andimmunostaining evidence are presented to support the ideathat this is a distinct type of projection from that of themossy, climbing, or monoaminergic axons. The functionalsignificance of this newly found projection is discussed.
2. Results
2.1. Screening of CN-projecting neurons by retrogradelabeling experiments
To examine the possible presence of brain stem neurons thatproject to the CN but not to the cerebellar cortex, wecompared the distribution of neurons in the medulla andcaudal pons that were labeled by horseradish peroxidase(HRP) injection into multiple positions in the left CN (fivecases) and by HRP injection into multiple points in the leftcerebellar cortex (four cases). By carefully comparing the twodistributions of retrogradely labeled neurons, we noticed thata group of neurons in the median reticular formation in therostrodorsal medulla was consistently labeled by CN injec-tions but rarely labeled by cortical injections (Fig. 1). Wereconstructed the distribution of labeled neurons in themedian and paramedian medulla and caudal pons (within125 mm to the midline) on the sagittal plane from one CNinjection and one cortical injection experiment (#100 and#110, respectively, Fig. 2). The area in which the neurons were

Fig. 1 – Screening medullary neuronal populations that project to the CN but not to the cerebellar cortex. A and B, Cameralucida drawing of cerebellar HRP injection sites in two rats in which injection sites were centered in the left CN (A) and in thecerebellar cortex (B), respectively. C and D, Camera lucida drawing of the distribution of retrogradely labeled neurons in acoronal section of the rostral medulla in the rats with injections shown in A and B, respectively. HRP-labeled neurons werevisualized with the TMB reaction in these sections. Each dot indicates a labeled neuron. Dotted curve with arrowscircumscribes the CRI in which neurons were labeled with HRP injections into the CN but not with HRP injections into thecerebellar cortex. E and F, Photomicrographs of labeled neurons in the CRI in the rat shown in (A) and (C).
b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 6 33
labeled only by the CN injection was observed in the rostro-dorsal medulla (dotted curve with arrows in Fig. 2). Theextent of this area was as follows: rostral border, rostral edgeof the abducens nucleus; caudal border, 1 mm caudal to therostral edge of the abducens nucleus; ventral border, 2/5thickness of the medulla dorsal to the ventral surface in themidline; dorsal border, 1/10 thickness of the medulla ventralto the dorsal surface in the midline; lateral border, 125 mm tothe midline. Except for this area, retrogradely labeled neuronswere distributed similarly in other areas of the median andparamedian medulla and caudal pons: both CN and corticalinjections led to dense labeling of neurons in the nucleusreticularis tegmenti pontis, medial vestibular nucleus, andinferior olive, and to sparse labeling in other parts of thereticular formation (Fig. 2A and B).
The area of specific neuronal labeling following a CNinjection was located dorsal to the raphe magnus, ventral to
the medial longitudinal fasciculus, and caudal to the nucleusraphe interpositus. The raphe interpositus is the medianpontine reticular formation, 1–3 mm below the floor of thefourth ventricle, which contains pause neurons of the oculo-motor system (Büttner-Ennever et al., 1988; Ohgaki et al.,1987; Hittinger and Horn, 2012). The caudal boundary of theraphe interpositus nucleus is around the level of the abdu-cens nucleus (Ohgaki et al., 1987). Therefore, the distributionarea of CN-projecting neurons in the present study wasimmediately caudal to the raphe interpositus, with somepossible overlap. Therefore, we will refer to the area of thedistribution of the CN projecting neurons as the ‘caudal rapheinterpositus area (CRI)’.
We counted the number of retrogradely labeled neurons inthe CRI in the five CN injection cases and four corticalinjection cases. The number of retrogradely labeled neuronsin the raphe interpositus was generally larger for CN

Fig. 2 – Distribution of CN-projecting neurons in the median reticular formation in the rostral medulla that delineated the CRI.A and B, Summary of mapping of retrogradely labeled neurons within 125 lm from the midsagittal plane in experimentsshown in Fig. 1A (#100) and B (#119), respectively. Mapping was done in every other section that was stained with TMBreaction. Dotted circle (filled arrows) circumscribes the CRI in which neurons were labeled by HRP injections into the CN(A) but not by HRP injections into the cerebellar cortex (B). C–F, Camera lucida drawing of the distribution of retrogradelylabeled neurons in sections in experiment #100 as indicated by open arrowheads in A (caudal sections are to the left).Dotted circle (filled arrows) circumscribes the CRI in C–E.
b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 634

Table 1 – Number of neurons in the CRI retrogradely labeled by HRP injections in the CN or cerebellar cortex. All injectionswere made in the left side.
Rat no. Injection centers Injectionvolume (ml)
No. of labeledneurons in the CRI
#100 Medial, interposed and lateral nuclei 2.0 201#158 Interposed nucleus 1.0 74#114 Interposed nucleus 0.5 40#115 Lateral nucleus 1.0 106#116 Medial nucleus 1.0 78#109 Vermis, hemisphere and paraflocculus 3.0 47#110 Vermis, hemisphere and paraflocculus 7.0 15#112 Vermis, hemisphere and paraflocculus 2.5 36#156 Rostral vermis and hemisphere (lobule II–V) 3.0 33
b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 6 35
injections than for cortical injections (Table 1). Case #114 hadnot as many as retrogradely labeled neurons in the rapheinterpositus as in other CN injection cases, which may be dueto smaller amount of injection volume of HRP in this case. Asa whole, the results suggested that there is a population ofneurons that projected predominantly to the CN in the CRI.
2.2. Anterograde tracing experiments
To confirm the projection of neurons in the CRI to the CN, weinjected anterograde axonal tracer (biotinylated dextranamine, BDA or Phaseolus vulgaris leucoagglutinin, PHA-L) intothe CRI or adjacent median areas (injection center, o0.2 mmto the midsagittal plane) in 17 rats. In the experiment (#129)shown in Fig. 3 the injection into the CRI labeled numerousaxonal terminals within the CN. The terminals were mainlydistributed in the ventral parts of the posterior interposedand lateral nuclei, but were also seen in other areas of the CNand in the vestibular nuclei (Fig. 3A–D). On the other hand,terminals were rarely observed in the cerebellar cortex;a small number of terminals (20 at most) were found in thegranular and Purkinje cell layers in a section, and manysections showed no terminals.
However, the distribution patterns of terminals werediffered significantly between injections; for example, manyrosette-type mossy fiber terminals were observed in thecerebellar cortex in some cases, and few terminals wereobserved either in the cortex or in CN in other cases. There-fore, we tried to relate the different terminal distributionpatterns to the location of the injection sites. By counting thenumber of terminals in the CN and cerebellar cortex in serialsections in each case, the 17 injection cases were classifiedinto one of three groups: (1) cases with labeled terminalspredominantly in the CN (41000 terminals in the CN and noor o200 terminals in the cerebellar cortex, 5 cases), (2) caseswith labeled terminals predominantly in the cerebellar cortex(41000 terminals in the cerebellar cortex, 7 cases), and (3)cases with few labeled terminals either in the CN or in thecerebellar cortex (o1000 terminals in the CN and o1000terminals in the cerebellar cortex, 5 cases). Injection sites ofonly the first group were located within the borders of the CRI(group 1, Fig. 4A). Axons and axonal terminals were denselyobserved in the CN but were not, or were only infrequently,observed in the cerebellar cortex in these cases. Injectionsthat were located outside of the CRI either produced
predominant terminal labeling in the cerebellar cortex(group 2, Fig. 4B), presumably due to labeling of mossy fiberaxons, or did not label many terminals in either the CN or thecerebellar cortex (group 3, Fig. 4C), presumably because theinjections were small or they did not hit a source of mossyfiber axons. These results confirm the presence of a neuronalpopulation in the CRI that projects to the CN, but not to thecerebellar cortex.
The area-dependency of the distribution of CN terminalsis summarized for five cases of CRI injection in Table 2.Terminals were densely distributed in the ventral parvocel-lular parts of the posterior interposed and lateral nuclei in allcases as seen in the mapping of #129 (Fig. 3). Terminals werealso observed less densely in most parts of the CN but werenot found in the dorsomedial crest or dorsolateral hump.
2.3. Anterograde single axon reconstruction
The most numerous afferent projections to the CN from thebrain stem are presumably the collaterals of climbing andmossy fibers. These afferent axons have morphologicallyspecialized terminations in the cerebellar cortex. Nuclearcollaterals of these axons also have a distinct morphology,which has been revealed by the reconstruction of individualaxons (Wu et al., 1999; Sugihara et al., 1999; Sugihara, 2011).In the present study, we reconstructed 12 axons in the fiverats, in which the anterograde tracer injection was localizedwithin the CRI (cases shown in Fig. 4A). We first located awell-labeled axon in the CN or in the white matter near theCN, and tried to trace the axon backward to its origin. Wethen tried to trace all branches in the cerebellum includingthe CN. Four out of 12 axons were traced down to a cell body.The neurons thus identified were located in the CRI, inside orin the close vicinity of the tracer injection site (Fig. 5A),thereby confirming the origin of the predominantly nuclearprojection from the CRI. Three axons out of 12 were com-pletely reconstructed in the cerebellum. The cell body wasidentified in one of these three.
The trajectory of the axon that was completely recon-structed and whose cell body was identified as being in theCRI is shown in Fig. 6. The cell body was relatively large(shown in Fig. 5A, long and short diameter, 45 and 12 mm,respectively) and was located nearly on the midline, 0.5 mmventral to the medullary floor of the fourth ventricle, and0.5 mm caudal to the center of the abducens nucleus. The
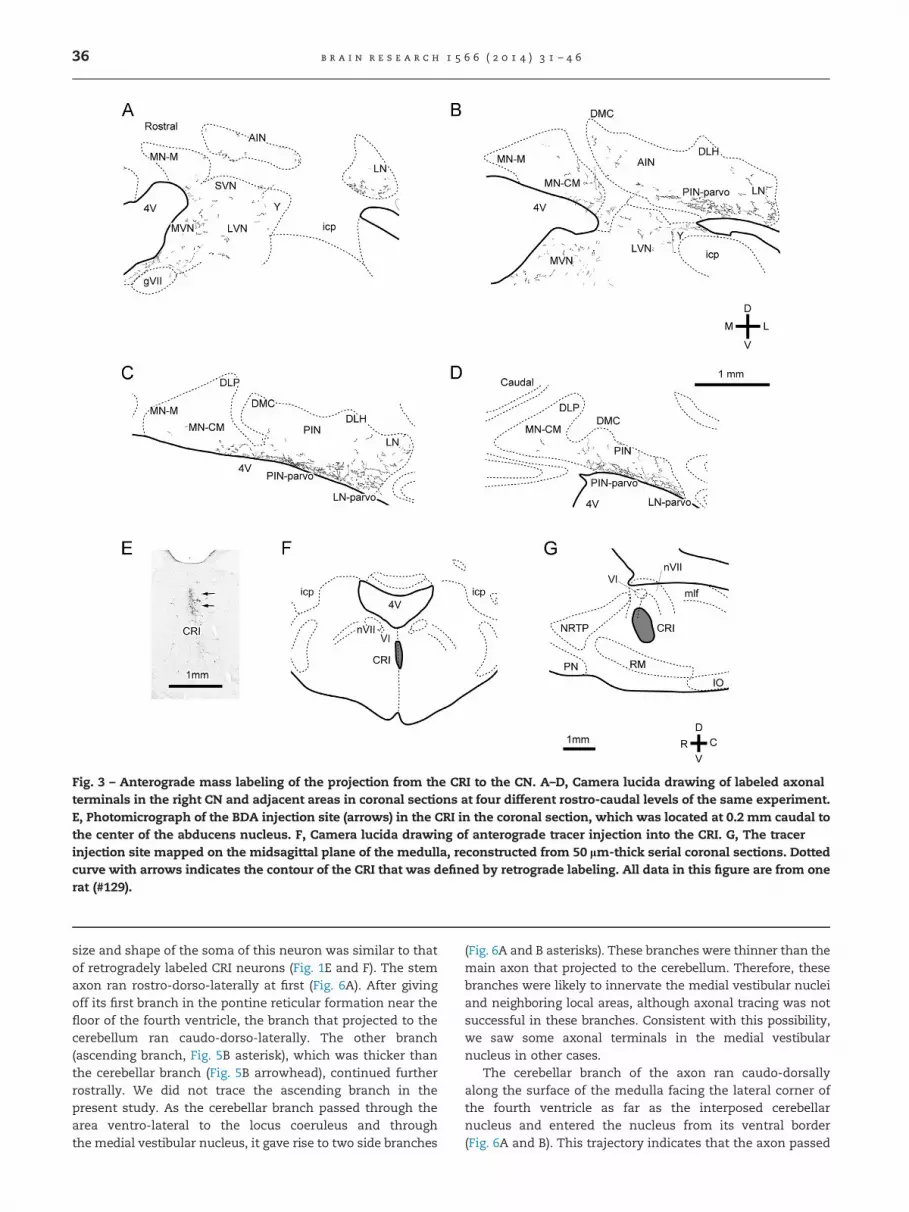
Fig. 3 – Anterograde mass labeling of the projection from the CRI to the CN. A–D, Camera lucida drawing of labeled axonalterminals in the right CN and adjacent areas in coronal sections at four different rostro-caudal levels of the same experiment.E, Photomicrograph of the BDA injection site (arrows) in the CRI in the coronal section, which was located at 0.2 mm caudal tothe center of the abducens nucleus. F, Camera lucida drawing of anterograde tracer injection into the CRI. G, The tracerinjection site mapped on the midsagittal plane of the medulla, reconstructed from 50 lm-thick serial coronal sections. Dottedcurve with arrows indicates the contour of the CRI that was defined by retrograde labeling. All data in this figure are from onerat (#129).
b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 636
size and shape of the soma of this neuron was similar to thatof retrogradely labeled CRI neurons (Fig. 1E and F). The stemaxon ran rostro-dorso-laterally at first (Fig. 6A). After givingoff its first branch in the pontine reticular formation near thefloor of the fourth ventricle, the branch that projected to thecerebellum ran caudo-dorso-laterally. The other branch(ascending branch, Fig. 5B asterisk), which was thicker thanthe cerebellar branch (Fig. 5B arrowhead), continued furtherrostrally. We did not trace the ascending branch in thepresent study. As the cerebellar branch passed through thearea ventro-lateral to the locus coeruleus and throughthe medial vestibular nucleus, it gave rise to two side branches
(Fig. 6A and B asterisks). These branches were thinner than themain axon that projected to the cerebellum. Therefore, thesebranches were likely to innervate the medial vestibular nucleiand neighboring local areas, although axonal tracing was notsuccessful in these branches. Consistent with this possibility,we saw some axonal terminals in the medial vestibularnucleus in other cases.
The cerebellar branch of the axon ran caudo-dorsallyalong the surface of the medulla facing the lateral corner ofthe fourth ventricle as far as the interposed cerebellarnucleus and entered the nucleus from its ventral border(Fig. 6A and B). This trajectory indicates that the axon passed

Fig. 4 – Mapping the CRI by anterograde tracing. Seventeencases of injection of PHA-L and BDA into median areas in therostral medulla and caudal pons were sorted into threeclasses based on the type of terminal labeling in thecerebellum. The injection sites were then mapped on adrawing of the midsagittal section with median andparamedian structures. A, Injections that labeled nuclearterminals predominantly (group 1). B, Injections that labeledcortical terminals predominantly (group 2). Corticalterminals were mostly mossy fibers. C, Injections thatlabeled few nuclear or cortical terminals (group 3). Dottedcurve with arrows indicates the contour of the CRI that wasdefined by retrograde labeling.
b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 6 37
through the superior cerebellar peduncle. Several brancheswere given off from the parent axon in the interposed nucleus.The parent axon came out of the nucleus and reached thelobular white matter, where it gave rise to several branchesprojecting to lobules III and IV, 1.2–1.7 mm lateral to themidline. Then, the parent axon returned to the interposednucleus to produce several further branches, including onesinnervating the medial and lateral nuclei. Such a hairpin-shaped path of the parent axon in the lobular white matter
was observed only in this case. In the CN, some branches wereshort and ended in small terminal arborizations, whereasothers were long and gave rise to many terminal branches(Fig. 6C). The diameter of the axon was 1.1–1.6 mm at the stemaxon before the first bifurcation, 0.6–1.1 at the parent axoninside the CN, 0.2–0.4 at the primary collateral in the CN, andabout 0.2 mm at the terminal branches in the CN. En-passantand terminal varicosities in the CN were roughly round-shaped and 1.0–2.0 mm (mostly 1–1.5 mm) in diameters. Thetotal number of the varicosities or swellings in this neuronwas 1086 in the CN and 64 in the cerebellar cortex.
In the CN, terminal branches not only bore manyen-passant varicosities but also frequently gave rise to shortbranchlets bearing one to several varicosities. As a result,varicosities often had a clustered appearance on terminalbranches (Fig. 5C–F). Most of the cortical varicosities were inthe granular layer (Fig. 5G), but some occur in the Purkinje celllayer (Fig. 5H and I). The terminal arbor in the cerebellarcortex was much smaller than that in the CN, and had asmaller number of varicosities as well (Fig. 6D and E).Terminal branches, and most of the varicosities, were similarin size to those in the CN, although a few varicosities wereslightly larger than others (up to 3 mm for their shortestdiameter). The morphology of this axon in the cerebellarcortex clearly distinguished it from mossy fiber axons, whichhave large rosette-type terminals, as well as from climbingfibers, which have a dense ivy-shaped arborization in themolecular layer, and from monoaminergic axons, which havesmall beaded varicosities (Moore and Bloom, 1979).
The morphological details of the other 11 reconstructedfibers that projected to the CN were similar to the casedescribed above. The trajectories of two axons that werecompletely reconstructed in the cerebellum are shown inFig. 7A and B. Within the CN, all of the reconstructed fibershad wide-spread branches with densely distributed varicos-ities, as demonstrated by the two axons shown in Fig. 7C andD. In all cases, the sizes of the varicosities and axons werenearly the same as those of the cases shown in Figs. 5 and 6.Concerning the cerebellar cortical projection, six axons didnot provide collaterals to the cortex while the five otheraxons had cortical branches bearing a small number of smallvaricosities (o100), similar to the case shown in Fig. 6.Collaterals in the medial vestibular nucleus were seen in allcases. The morphology of the terminal arbor in the medialvestibular nucleus (Fig. 7A) was similar to that in the CN. Thepath of eight axons was peri-ventricular, as in the casesshown in Figs. 6A, 7A and 7B, but the path of three otheraxons was more lateral, coursing through the medial vestib-ular nucleus. The ascending branch was seen in all axonsthat could be traced up to the soma (n¼4).
2.4. Electron microscopic observations
We made electron microscopic observations of the BDA-labeled terminals in the ventral part of the interposednucleus to clarify further the presynaptic characteristics ofthe CRI axons in the CN. Labeled terminals of CRI axons wereidentified by the presence of electron-dense amorphousreaction product (Fig. 8). Although the DAB reaction productmade detailed observation of the presynaptic terminal

Table 2 – Distribution of axonal terminals labeled by anterograde tracer injections into the CRI.
Rat no. MN-CM MN-M DLP DMC AIN DLH PIN PIN-parvo LN LN-parvo
#94Left � þ þ � þþ � þ þþ þ þRight þ þ � � þþ � þþ þ � þþ
#107Left þ þ � � � � þ þþ � þþRight � þ � � � � þ þþ þ þþ
#129Left � þ þ � þ � þ þþ þ þþþRight þ þ � � þþ � þþ þþþ þþ þþþþ
#131Left þ þ � � � � þ þþ þ þþRight � � þ � � � þ þþ � þþ
#134Left þþ þ þ � � � þ þþ þ þþRight � þ � � � � þ þþ þ þ
Definition of symbols, � , þ, þþ, þþþ, and þþþþ, less than 0.5, 0.5–4.0, 4.0–20.0, 20.0–100.0, and 100.0o terminals, respectively, per 20� visualfield of the microscope. Average of 20 observations in multiple sections. Terminals were scarce in the cerebellar cortex in all cases. Tracers:#94: PHA-L, and #107–#134: BDA. See Fig. 4A for location of injection sites. The medial nucleus was divided into the MN-CM, MN-M and DLPaccording to Voogd (2004).
Fig. 5 – Photomicrographs of a labeled CRI neuron and its axon and axonal terminals. A, Identified cell body. Arrowheadsindicate the axon. B, Branching point near the floor of the fourth ventricle in the pons. Arrowhead indicates the branch thatprojects to the cerebellum. Single asterisk indicates the ascending branch. Double asterisk indicates the stem axon. C–F,Terminal arborization bearing en-passant and terminal varicosities in the posterior interposed cerebellar nucleus. G, Terminalarborization in the granular layer. H and I, Terminal arborization in the Purkinje cell layer. A–E, G and H are from the neuronand axon shown in Fig. 6. F and I are from other reconstructed axons. Scale bar in I applies to C–I.
b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 638

Fig. 6 – Reconstructed single axon of an identified CRI neuron. A, The axonal trajectory was drawn from a caudal perspectiveonto the outline of the CN in a coronal section. Reconstruction was made from 61 serial coronal sections. This axon wascompletely reconstructed from the soma to all of its terminals in the CN, except for an ascending branch (double asterisk) andthin collaterals in the medial vestibular nucleus (single asterisks). Inset: entire fiber path to the cell body shown under lowmagnification. B, The trajectory in lateral view of the same axon as in A, depicted on a montage of parasagittal sections of therat brain. C, Parts of terminal arborization in the posterior interposed nucleus in a single section shown under highmagnification. Broken lines indicate continuation of the axon to the consecutive sections. D, Cortical collateral and boutons inthe lobule IV shown under high magnification. Reconstructed from 11 coronal sections. In C–E, thickness of varicosities andfibers is approximately to scale. Shaded areas in A and B indicate the injection site. This axon had 1086 varicosities in the CNand 64 varicosities in the cerebellar cortex.
b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 6 39
difficult, we could observe some characteristics of the term-inals. Specifically, the axonal terminals contained densely-packed, pleomorphic but generally round, pale synapticvesicles, and made putative asymmetric synaptic contactsonto the dendrites of nuclear neurons (arrowheads, Fig. 8). Nodense-core vesicles indicative of a monoaminergic terminal(Richards and Tranzer, 1970; Muller et al., 2007) were observedin the 20 terminals that were examined. This morphology
suggested that these terminals make non-monoaminergicsynaptic connections.
2.5. Distinction between CRI axons and serotoninergicaxons
Serotoninergic fibers and terminals are known to be abundantin the CN (Takeuchi et al., 1982; Bishop and Ho 1985). The

Fig. 7 – Samples of reconstructed CRI axons in the CN. A, axon of a CRI neuron that was completely reconstructed in the CNand medial vestibular nucleus. This axon had 170 varicosities in the CN and 98 varicosities in the medial vestibular nucleus.The axonal trajectory (caudal view) was drawn on the brain outlines of two coronal sections (at the level of medial vestibularnucleus slightly caudal to the locus coeruleus, and at the level of the posterior interposed nucleus). B, axon of another CRIneuron that was completely reconstructed in the CN. This axon had 515 varicosities. The axonal trajectory (caudal view) wasalso drawn on brain outlines of two coronal sections (at the rostral and caudal levels of the CN). C and D, Local terminal arborin the CN of other partially reconstructed CRI axons. These axons had 404 (C) and 268 (D) varicosities, respectively. Asteriskindicates the stem of the axon. Scale bar in D applies to A–D.
b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 640
raphe nuclei, located in the median region of the reticularformation, contain serotoninergic neurons. In particular, theraphe magnus is located just ventral to the CRI (Figs. 2–4).Although, serotoninergic neurons are otherwise relativelyscarce in the dorsal part of the rostral medulla (Jacobs et al.,
1984), spread of the tracer may have caused labeling of someserotoninergic neurons in the raphe magnus. Therefore, tofurther test whether the cerebellum-projecting CRI axons wereserotoninergic or not, we injected PHA-L into the CRI and madeserial sections through the CN. Sections were alternately stained

Fig. 8 – Electron micrographs of BDA-labeled terminals in the ventral part of the posterior interposed nucleus. Two samples ofsections of labeled axonal terminals are shown (A and B). BDA injection was made into the CRI. Labeled axons werevisualized with DAB reaction. Arrowheads indicate putative asymmetric synapse formation.
b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 6 41
for 5-HT immunoreactivity and PHA-L. Camera-lucida drawingsof PHA-L labeled axons and 5-HT immunoreactive axons incontiguous sections were accurately superimposed blood vesselsas references. At the cut plane between the two sections, noPHA-L-labeled axons aligned with 5-HT-immunoreactive axons(e.g., red and black arrowheads in Fig. 9). This result indicatesthat the cerebellum-projecting axons originating from the CRIare not serotoninergic.
3. Discussion
The present study showed that the cerebellar projection fromthe CRI terminates mainly in the CN, and that only relativefew CRI fibers project to the cerebellar cortex. The morpho-logical characteristics of terminal arbor of the CRI axon in thecerebellar cortex and CN were clearly different from those ofmossy fibers, which originate from many precerebellar neu-rons in the brain stem, climbing fibers, and monoaminergicfibers. Thus, this study presented a detailed morphologicaldemonstration of a distinct type of the cerebellar afferentthat arises in the brain stem and predominantly targetsthe CN.
3.1. A cerebellar afferent system from the brain stem thatprojects predominantly to the CN
The CN are the major output source of the cerebellum, whichsends cerebellar output signals to various brainstem nucleiincluding the thalamus, red nucleus, vestibular nuclei and
reticular formation (Voogd, 2004). For the cerebellum toexecute its function in motor coordination, the CN wouldhave to change their activity in a timely manner in responseto changes in the activity of afferents to the cerebellum.In many behavioral experiments, including conditioning of theeye blink reflex, the CN increases its activity when the animalis required to generate modulation in its behavior. However,how the output signal is generated in the CN neurons by theirintrinsic electrophysiological properties and the synapticinputs to them from Purkinje cells and other afferents isnot well understood (Person and Raman, 2012). AlthoughPurkinje cell axonal terminals are the predominant inputsto the CN neurons (Chan-Palay, 1973), how this input isconverted into the activity of CN neurons is not straightfor-ward, since the Purkinje cell projection is inhibitory (Ito et al.,1970). Rebound excitation upon the temporal cessation ofPurkinje cell activity is the most likely cause, however,excitatory inputs from afferents other than Purkinje cellshave to be taken into account. In this sense it is important toclarify direct brain stem inputs to the CN.
Climbing fiber axons (or olivocerebellar axons) usuallyhave collaterals that project to the CN (Van der Want et al.,1989; Sugihara et al., 1999). The projection pattern of thesecollaterals has a precise topographic relationship to theirorigin (Ruigrok and Voogd, 2000; Pijpers et al., 2005; Sugiharaand Shinoda, 2007). Mossy fiber axons constitute the mostabundant afferent system in the cerebellar cortex (Voogd,2004), and thus their collaterals presumably form the mostabundant input to the CN from the brain stem. However, notall mossy fiber axons have collaterals that project to the CN.

Fig. 9 – Absence of 5-HT immunoreactivity in CRI axons in the CN. A and B, Camera lucida drawing of CRI axons and 5-HT-immunoreactive axons in consecutive sections of the CN. CRI axons were labeled by PHA-L injected into the CRI. Every othersection of the CN was immunostained for PHA-L and 5-HT. Black and red drawings are labeled terminal axons and bloodvessels (used as position landmarks) in consecutive sections stained for PHA-L (black) and 5-HT (red) in the ventral part of theposterior interposed nucleus. Cut ends of labeled axons at the boundary between the two sections are indicated by arrows. Nocut ends of black and red axons met with each other, indicating that they belong to distinct axonal populations. Note thatblood vessels (circles) met with each other nearly completely, showing the precise positional alignment of the drawings oftwo consecutive sections.
b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 642
For example, mossy fiber axons originating from the pontinenucleus and external cuneate nucleus have CN collateralsinfrequently (about 1:3 and 1:15, Shinoda et al., 1992; Quyet al., 2011). On the other hand, mossy fiber axons originatingfrom the lateral reticular nucleus always have CN collaterals(Wu et al., 1999). The topographic relationship between thecortical and nuclear projections is not as precise for mossyfiber systems as it is for the climbing fiber systems. Nora-drenergic and serotoninergic projections to the CN andcerebellar cortex have been well documented (Moore andBloom, 1979; Takeuchi et al., 1982). Whether these mono-aminergic projections to the CN arise as collaterals of themonoaminergic projection to the cerebellar cortex or not hasnot been clarified so far. However, it is possible that theseprojections originate from common axons since there are arelatively small number of monoaminergic neurons in thebrain, and since monoaminergic projections are generallydivergent (Moore and Bloom, 1979).
The present study has reported the presence of a distincttype of cerebellar afferent originating from the medulla:a population of CRI neurons that project predominantly to theCN and only sparsely to the cerebellar cortex. The innervationof the cortex by these neurons was very weak, consisting of asmall number of varicosities or swellings (short diameter,mostly 1–3 mm) that was much smaller than mossy fiberrosettes (short diameter, 4–8 mm). The diameter of CRI axonsat the entrance to the cerebellum (about 1 mm) was muchsmaller than that of mossy fibers (2–4 mm; Wu et al., 1999; Quyet al., 2011). Although the diameter of CRI axons was in thesame range as that of climbing fiber axons (0.7–1.4 mm;Sugihara et al., 1999), cortical termination of CRI axons
was quite dissimilar to that of climbing fibers. Therefore,the CRI axons were clearly different from mossy or climbingfiber axons.
Concerning the noradrenergic projection, locations ofnoradrenergic neurons (locus coeruleus near the lateral wallof the fourth ventricle in the rostral medulla, A5 cell group inthe ventrolateral medulla, and A7 cell group in the lateralpons; Dahlstrom and Fuxe, 1964; Moore and Bloom, 1979) areall far separated from the CRI, which is located in the medianreticular formation in the rostral medulla. Concerning theserotoninergic projection, the raphe magnus nucleus, one ofthe sources of the serotoninergic projection to the cerebellum(Hokfelt and Fuxe, 1969; Takeuchi et al., 1982; Bishop and Ho1985), is located just ventral to the CRI. However, the reporteddistribution of serotoninergic neurons does not extend to theCRI, which is located dorsal to the raphe magnus nucleus(Jacobs et al., 1984; Kitzman and Bishop, 1994; Bishop and Ho,1985; Harding et al., 2004). Moreover, the morphology of theCRI axons in the cerebellar cortex and CN does not fit wellwith immunostained noradrenergic or serotoninergic fibers,which are characterized as a varicose fine network (Hokfeltand Fuxe, 1969; Takeuchi et al., 1982; Bishop and Ho 1985).Furthermore, our immunostaining observations and electronmicroscopic observations did not support that the CRI axonswere serotoninergic. In sum, the present study indicated thatCRI axons were not monoaminergic.
Concerning cholinergic innervation of the CN, the CRI doesnot seem to overlap with the reported sources of the projectionsin the brain stem, such as the pedunculopontine and laterodor-sal tegmental nuclei (Woolf and Butcher, 1989), the medialvestibular nucleus (Barmack et al., 1992), lateral

b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 6 43
paragigantocellular nucleus, and the raphe obscurus nucleus(Jaarsma et al., 1997). This suggests that the CRI projection is notcholinergic. However, other raphe nucleus can also be a source ofcholinergic projection to the cerebellum to a lesser extent(Jaarsma et al., 1997). Moreover, the synaptic terminals of theCRI axons are not dissimilar to the cholinergic terminals in theCN when studied by electron microscopy (Jaarsma et al., 1997).Therefore, whether the CRI projection to the CN is cholinergicneeds further investigation. Neurons in the raphe obscurusnucleus were labeled retrogradely from both the CN and cortex(Fig. 1), suggesting the possibility that cholinergic neurons in thisnucleus project to both the CN and cortex.
Rubrospinal axons, which arise from the magnocellular rednucleus, give off collaterals to the CN, and these fibersconstitute another type of afferent to the cerebellum, onewhich does not supply terminals to the cerebellar cortex(Huisman et al., 1983). The cerebellar projection of CRI axonsis similar to this projection in that it is formed by thecollaterals of the main projection axons, and in that it mainlysupplies to the CN. Since no anterograde labeling study hasinvestigated the CN projection from the red nucleus, thepresent study is the first demonstration of the axonal mor-phology of CN projection from the brain stem except forclimbing and mossy fiber axons. Whether there are any otherbrain stem neurons that also send a similar new type ofcerebellar afferents is not clear.
3.2. Possible function of the CRI projection
The terminal arborization of single CRI axons in the CN wascharacterized by dense distributions of varicosities. Althoughthe CRI does not occupy a large volume and, consequently,the number of CRI neurons was not as large as the numberof neurons in major precerebellar nuclei, each CRI axon hada large number of varicosities that were distributed ina relatively wide area of the CN (�1000, Figs. 5 and 6).In comparison to a CRI axon, a single mossy fiber axon hasa much smaller number of varicosities in the CN (36–270 inthe lateral reticular nucleus axons in rat, Wu et al., 1999; 0–59in the dorsal column nucleus axons in rat, Quy et al., 2011).On the other hand, the cortical projection of the CRI neuronswas very weak. Therefore, the functional significance ofCRI axons would be very different from that of mossy fibersystems. Mossy fiber systems are supposed to supply massivesensory and efference copy signals to the cerebellum, whichare used in cortical and nuclear information processing togenerate appropriate cerebellar output. In contrast, the CNprojection from the CRI may be involved in modulating thecerebellar output directly.
The terminal arbor of a single CRI axon was more widely anddensely distributed than the collaterals of the climbing andmossy fiber axons in the CN. In this regard, activity of CRI axonsmay produce a particularly strong and broad effect on CNneurons in the ventral parvocellular parts of the posteriorinterposed and lateral nucleus. However, identification of theneurotransmitter and synaptic action of the CRI axon will berequired to further examine the function of CRI axons in the CN.
The function of the CRI is not well understood. As a part ofthe medial reticular formation, which is generally involved inmotor function (Rho et al., 1997), the CRI may also have some
function related to motor control. Indeed, eye-movement-related omnipause neurons were located in the raphe inter-positus nucleus, which is in the pontine median reticularformation immediately rostral to the CRI (Ohgaki et al., 1987;Hittinger and Horn, 2012). Omnipause neurons project to thearea around the abducens nucleus and other brain stemareas, but not to the cerebellum as demonstrated in anintracellular labeling study in cat (Ohgaki et al., 1987). Theaxonal trajectory of the CN-projecting CRI neurons in thepresent study was different from that of omnipause neuronsin the raphe interpositus. However, it is possible that CRIneurons may also be involved in eye movement control, since(1) the ventral part of the posterior interposed and lateral nuclei,where the CRI axons were found to project in the present study,is connected with the flocculus, paraflocculus, lateral uvula andcrus I (Sugihara and Shinoda, 2004; Sugihara et al., 2004, 2009),lobules that are involved in oculomotor control (Kheradmandand Zee, 2011); (2) the ventral part of the posterior interposedand lateral nuclei has also been shown to be involved in somekinds of eye movements in monkey (Zhang and Gamlin, 1998)and project to the superior colliculus (Teune et al., 2000); (3)injection of HRP into the H field of Forel labeled neurons in thearea that is equivalent to the CRI in the cat (Ohgaki et al., 1989);and (4) omnipause neurons send axonal branches to the areathat is equivalent to the CRI (Ohgaki et al., 1987). Although mostof studies on the oculomotor system in the brain stem weredone in cat and monkey, the oculomotor system is alsodeveloped in the brain stem in rat (Hittinger and Horn, 2012).CRI axons had an ascending branch that was not traced in thepresent study. Identifying the target of the ascending branch ofCRI axons will be needed to understand nature of CRI neuronsincluding the possibility that they are involved in eye move-ment control in rat.
4. Experimental procedure
4.1. Surgical procedures and tracer injection
Long-Evans male and female adult rats weighing 230–350 g(Kiwa Laboratory Animals, Wakayama, Japan) were used. Allanimal experiments were carried out in accordance with theguidelines of the animal welfare committee of the TokyoMedical and Dental University, subsequent to approval by theEthics Review Committee for Animal Experimentation of TokyoMedical and Dental University (approval numbers 0020238,0030129, 0040089, and 0130211A). Rats were anesthetized withintraperitoneal injection of ketamine (130mg/kg body weight),xylazine (8 mg/kg) and atropine (0.4 mg/kg). Supplementaldoses of ketamine (26 mg/kg) and xylazine (1.6 mg/kg) weregiven every 30min starting 1 h after the initial dose duringsurgery (less than three hours). A heating pad was used to keepthe rectal temperature between 35 and 37 1C.
To inject horseradish peroxidase (HRP) in the cerebellarcortex or CN to label projecting neurons retrogradely, the ratwas placed in a stereotaxic apparatus in a prone position, 451nose down. The bone covering the vermis and left hemi-sphere was removed. A micropipette filled with HRP (PEO-131,Toyobo, Osaka, Japan; 30% solution in saline) was insertedinto various positions in the left cerebellar cortex to inject

b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 644
HRP solution at depths of 100–500 mm at 20–30 points (vermis,pars intermedia, hemisphere, and paraflocculus–flocculus;total injection amount, 2.5–7 ml) in five rats. A stereotaxicatlas (Paxinos and Watson, 2007) was used to locate the CN. Infive other rats, a micropipette filled with HRP was inserted intothe left CN (medial, interposed and lateral nuclei, 0.5–4.0 mmlateral to the midline, through lobule VIc, crus I and crus IIa,4.5–3.5 mm deep; total injection amount, 0.5–2 ml). After asurvival period of 48 h, the animals were deeply anesthetizedwith ketamine (200mg/kg) and xylazine (12 mg/kg), and trans-cardially perfused with phosphate-buffered saline followed byfixative containing 2% paraformaldehyde, 0.6% glutaraldehydeand 4% sucrose in 0.05 M sodium phosphate buffer (pH 7.4).The brain was dissected and postfixed overnight in the samefixative and then soaked in 30% sucrose solution (pH 7.2 with10mM phosphate buffer).
To label projections of neurons in the CRI and nearbyareas anterogradely, rats were placed in a stereotaxic appa-ratus in a supine position. A small incision was made in theventral neck at the midline. Median neck muscles, thetrachea, esophagus, and pharynx were retracted to the leftto expose the clivus with a parapharyngeal approach. A holewas drilled in the right clivus under the rostral medulla(center: 2.8 mm rostral to the foramen magnum, 0.2 mm rightto the midline), and a small incision was made in the dura.A glass micropipette (tip diameter, 4 mm) filled with P. vulgarisleucoagglutinin (PHA-L, L-1110, Vector Labs, Burlingame, CA,U.S.A., 2.5% in sodium phosphate-buffered saline) or biotiny-lated dextran amine (BDA, D-1956, 10,000 MW, MolecularProbe, Eugene, Oregon, U.S.A.; 10% solution in saline) wasinserted into the CRI (depth, 2.3 mm, direction, 51 tilted in thecoronal plane toward the midline) and adjacent areas bychanging the rostrocaudal and dorsoventral coordinates.PHA-L was electrophoretically injected (2 mA positive currentpulses of 1 s duration at 0.5 Hz for 20 min), whereas BDA waspressure-injected (�0.05 ml). After a survival period of four orfive days, the animals were deeply anesthetized with keta-mine (200 mg/kg) and xylazine (12 mg/kg), and transcardiallyperfused with phosphate-buffered saline followed by fixativecontaining 2% paraformaldehyde, 0.6% glutaraldehyde and4% sucrose in 0.05 M sodium phosphate buffer (pH 7.4). Thebrain was dissected and postfixed overnight in the samefixative and then soaked in 30% sucrose solution (pH 7.4 with10 mM phosphate buffer).
4.2. Histological procedures
For brains where an HRP injection was made, the cerebellumand brain stem were dissected apart and processed sepa-rately. The cerebellum was cut into serial sagittal sectionswhereas the brain stem was cut into serial coronal sections(60 mm thick) on a freezing microtome. After washing for30 min, all the cerebellar sections and a set of every otherbrain stem section were treated with 3,30-diaminobenzidine(DAB, 0.5 mg/ml) plus 0.3% hydrogen peroxide in phosphatebuffer for 15 min. Sections were washed with PBS, mountedon glass slides, dried, dehydrated with ethanol and cover-slipped with Permount (Fisher Scientific, Fair Lawn, NJ). Theremaining set of brain stem sections was treated with3,30,5,50-tetramethylbenzidine (TMB), which is more sensitive
than DAB (Mesulam, 1982). TMB was dissolved first in ethanol(20 mg/10 ml ethanol) and then mixed with cooled solution ofsodium nitroprusside (1 g/l), sucrose (10 g/l) and sodiumacetate buffer (50 mM, pH 3.3) in distilled water to obtainfinal concentration of 40 mg/l. Hydrogen peroxide (0.3%) wasadded immediately before use. Sections were rinsed withacetate buffered saline (pH 3.3) and then incubated two timesin the TMB reaction solution at 4 1C for 30 min. Sections werewashed with acetate buffered saline, mounted on glass slides,dried, dehydrated with ethanol and coverslipped with Per-mount (Fisher Scientific, Fair Lawn, NJ). Retrogradely labeledneurons were counted in sections that underwent TMBtreatment, because of the more sensitive HRP visualizationby the TMB reaction than the DAB reaction.
For the PHA-L injections, each brain was embedded in agelatin block, which was fixed in a Formalin solution (For-malin 20%, sucrose 25%, 4 1C) for 2–4 days. The gelatin blockwas cut into serial coronal sections (30 mm thick) on a freezingmicrotome. After washing for 30 min, sections were incu-bated with 0.1% sodium azide plus 0.3% hydrogen peroxidefor quenching intrinsic peroxidase. Sections were incubatedwith goat anti-PHA(EþL) antibody (1:2000, AS-2224, Vector) inPBST plus 2% normal goat serum for 72 h, washed, thenincubated with rabbit anti-goat IgG (305-005-003, 1:200, Jack-son, West Grove, PA, U.S.A.), then goat peroxidase anti-peroxidase (PAP, 123-005-024, 1:400, Jackson). PHA-L was thenvisualized by modification of the cobalt-glucose oxidasemethod (Itoh et al., 1979; Van der Want et al., 1989). Sectionswere preincubated for 10 min in 0.5% cobalt acetate in Tris–HCl (50 mM, pH, 7.6). After being rinsed in Tris-HCl, and thenin PBS, sections were incubated with 0.05% DAB, beta-D-glucose (Sigma), and 0.4% ammonium chloride, glucose oxi-dase (0.5 mg/100ml) in PBS for 60 min. Sections were thenwashed with PBS, mounted on glass slides, dried, and cover-slipped with Permount (Fisher Scientific, Fair Lawn, NJ,U.S.A.).
For the BDA injections, each brain was embedded in a gelatinblock, which was fixed in a Formalin solution (Formalin 20%,sucrose 25%, 4 1C) for 2–4 days. Frozen coronal or parasagittalsections of 50 mm thickness were made and treated withbiotinylated HRP–avidin complex (Elite ABC kit KT-6100, Vector)in a sodium phosphate buffer (0.1 M, pH 7.4) overnight at 4 1C.Next, BDA was visualized by modification of the cobalt-glucoseoxidase method as described above. Then, the sections weremounted on chrome alum-gelatinized slides as described above.
Two brains with PHA-L injection were embedded in gelatinand cut into serial coronal sections (30 mm). A set of every othersection was processed for PHA-L labeling as above. The other setof every other section was processed for 5-hydroxytriptamine(serotonin, 5-HT) immunostaining. After quenching intrinsicperoxidase as above, sections were incubated with anti-serotonin rabbit polyclonal antibody (NT102, 1:1000, Eugene,Ridgefield Park, NJ, U.S.A.) with 2% normal rabbit serum for 72 h.This antibody specifically labels serotoninergic neurons (Ishidaet al., 1998; Michel et al., 2000). Then sections were incubatedwith biotinylated anti-rabbit IgG antibody (BA 1000, Vector),followed by biotinylated HRP-avidin complex (Elite ABC kit KT-6100, Vector) in a sodium phosphate buffer (0.1 M, pH 7.4)overnight at 4 1C. Next, BDA was visualized by modification ofthe cobalt-glucose oxidase method as described above. Lastly,

b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 6 45
the sections were mounted on chrome alum-gelatinized slidesas described above.
We mapped labeled neurons and reconstructed singleaxons under a microscope with a camera lucida apparatusby using a computer-aided system (Sugihara and Fujita, 2010)as described before (Quy et al., 2011). We referred to samplesagittal sections of the rat brain atlas (Paxinos and Watson,2007) to summarize mapping from serial coronal sectionsonto a single sagittal plane.
The number of varicosities were counted manually under20� objective while adjusting the focus of the microscope tovisualize all levels throughout the thickness of each section.
4.3. Electron microscopy
BDA was injected into the median area of the medianreticular formation in the rostral medulla in three rats asdescribed above. After seven days the rat was perfused withfixative containing 2% paraformaldehyde and 3% glutaralde-hyde in 0.10 M phosphate buffer (pH¼7.4). The brain wasdissected and postfixed in the same fixative overnight. Eightymicrometer sections of the brainstem and the cerebellumwere cut with a microslicer. BDA was visualized with DAB-labeling as described above except that no cobalt preincuba-tion was done. After confirming that the injection site was inthe intended area and that there were labeled axons in theCN, the ventral part of the posterior interposed nucleus wastrimmed from the section and postfixed for 1 h with 1%osmium tetroxide in phosphate buffer, dehydrated with100% ethanol, infiltrated with propylene oxide and epoxyresin (Epon 812, TAAB, Berks, England), embedded, andpolymerized in a 60 1C oven for 48 h. Coronal sections of theventral part of the posterior interposed nucleus were cut at athickness of 60–90 nm, mounted on copper–rhodium meshgrids and stained with 3% aqueous uranyl acetate for 10 min.The sections were examined and photographed on a HitachiH-600 electron microscope (75 kV acceleration).
Acknowledgments
This study was supported by Grants-in-Aid for ScientificResearch from the Japan Society for the Promotion of Scienceto I.S. (25430032). The authors thank Dr. Sizuko Ichinose fortechnical assistance on electron microscopy and Dr. Eric J.Lang for comments to the manuscript. A student, with whomwe have lost contact, helped during the initial part ofthe study.
r e f e r e n c e s
Barmack, N., Baughman, R., Eckenstein, F., 1992. Cholinergicinnervation of the cerebellum of rat, rabbit, cat, and monkeyas revealed by choline acetyltransferase activity andimmunohistochemistry. J. Comp. Neurol. 317, 233–249.
Bishop, G.A., Ho, R.H., 1985. The distribution and origin ofserotonin immunoreactivity in the rat cerebellum. Brain Res.331, 195–207.
Buttner-Ennever, J.A., Cohen, B., Pause, M., Fries, W., 1988. Raphenucleus of the pons containing omnipause neurons of the
oculomotor system in the monkey, and its homologue in man.J. Comp. Neurol. 267, 307–321.
Chan-Palay, V., 1973. On the identification of the afferent axonterminals in the nucleus lateralis of the cerebellum. Anelectron microscope study. Z. Anat. Entwickl. 142, 49–186.
Dahlstrom, A., Fuxe, K., 1964. Evidence for the existence ofmonoamine-containing neurons in the central nervoussystem. Acta Physiol. Scand. 62 (Suppl. 232), S1–S55.
Eller, T., Chan-Palay, V., 1976. Afferents to the cerebellar lateralnucleus. Evidence from retrograde transport of horseradishperoxidase after pressure injections through micropipettes.J. Comp. Neurol. 166, 285–302.
Harding, A., Paxinos, G., Halliday, G., 2004. The Serotonin andTachykinin Systems. In: Paxinos, G. (Ed.), The Rat Nervous System3rd edition Elsevier Academic Press, Amsterdam, pp. 1205–1256.
Hittinger, M., Horn, A.K., 2012. The anatomical identification ofsaccadic omnipause neurons in the rat brainstem.Neuroscience 210, 191–199.
Hokfelt, T., Fuxe, K., 1969. Cerebellar monoamine nerveterminals, a new type of afferent fibers to the cortex cerebelli.Exp. Brain Res. 9, 63–72.
Huisman, A.M., Kuypers, H.G.J.M., Conde, F., Keizer, K., 1983.Collaterals of rubrospinal neurons to the cerebellum in rat.A retrograde fluorescent double labeling study. Brain Res. 264,181–196.
Ishida, Y., Hashiguchi, H., Todaka, K., Kuwahara, I., Ishizuka, Y.,Nakane, H., Uchimura, D., Nishimori, T., Mitsuyama, Y., 1998.Serotonergic activity in the rat striatum after intrastriataltransplantation of fetal nigra as measured by microdialysis.Brain Res. 788, 207–214.
Ito, M., Yoshida, M., Obata, K., Kawai, N., Udo, M., 1970. Inhibitorycontrol of intracerebellar nuclei by the purkinje cell axons.Exp. Brain Res. 10, 64–80.
Itoh, K., Konishi, A., Nomura, S., Mizuno, N., Nakamura, Y.,Sugimoto, T., 1979. Application of coupled oxidation reactionto electron microscopic demonstration of horseradishperoxidase: cobalt-glucose oxidase method. Brain Res. 175,341–346.
Jaarsma, D., Ruigrok, T.J.H., Caffe, R., Cozzari, C., Levey, A.I.,Mugnaini, E., Voogd, J., 1997. Cholinergic Innervation andReceptors in the Cerebellum. In: De Zeeuw, C.I., Strata, P.,Voogd, J. (Eds.), The Cerebellum: From Structure to Control.Elsevier, Amsterdam, pp. 67–96.
Jacobs, B.L., Gannon, P.J., Azimitia, E.C., 1984. Atlas of serotonergiccell bodies in the cat brain stem: an immunocytochemicalanalysis. Brain Res. Bull. 13, 1–31.
Kheradmand, A., Zee, D.S., 2011. Cerebellum and ocular motorcontrol. Front. Neurol. 2, 53.
Kitai, S.T., McCrea, R.A., Preston, R.J., Bishop, G.A., 1977.Electrophysiological and horseradish peroxidase studies ofprecerebellar afferents to the nucleus interpositus anterior.I. Climbing fiber system. Brain Res. 122, 197–214.
Kitzman, P.H., Bishop, G.A., 1994. The origin of serotoninergicafferents to the cat’s cerebellar nuclei. J. Comp. Neurol. 340,541–550.
Matsushita, M., Yaginuma, H., 1995. Projections from thecentral cervical nucleus to the cerebellar nuclei in the rat,studied by anterograde axonal tracing. J. Comp. Neurol. 353,234–246.
Mesulam, M.-M., 1982. Principles of Horseradish PeroxidaseNeurochemistry and their Application for Tracing NeuralPathways – Axonal Transport, Enzyme Histochemistry andLight Microscopic Analysis. In: Mesulam, M.-M. (Ed.), TracingNeural Connections with Horseradish Peroxidase. Wiley, NewYork, pp. 1–151.
Michel, S., Schoch, K., Stevenson, P.A., 2000. Amine and aminoacid transmitters in the eye of the mollusc Bulla gouldiana: animmunocytochemical study. J. Comp. Neurol. 425, 244–256.

b r a i n r e s e a r c h 1 5 6 6 ( 2 0 1 4 ) 3 1 – 4 646
Moore, R.Y., Bloom, F.E., 1979. Central catecholamine neuronsystems: anatomy and physiology of the norepinephrine andepinephrine systems. Annu. Rev. Neurosci. 2, 113–168.
Muller, J.F., Mascagni, F., McDonald, A.J., 2007. Serotonin-immunoreactive axon terminals innervate pyramidal cellsand interneurons in the rat basolateral amygdala. J. Comp.Neurol. 505, 314–335.
Murano, M., Saitow, F., Suzuki, H., 2011. Modulatory effects ofserotonin on glutamatergic synaptic transmission and long-term depression in the deep cerebellar nuclei. Neuroscience172, 118–128.
Newman, D.B., Ginsberg, C.Y., 1992. Brainstem reticular nucleithat project to the cerebellum in rats: a retrograde tracerstudy. Brain Behav. Evol. 39, 24–68.
Ohgaki, T., Curthoys, I.S., Markham, C.H., 1987. Anatomy ofphysiologically identified eye-movement-related pause neuronsin the cat: pontomedullary region. J. Comp. Neurol. 266, 56–72.
Ohgaki, T., Markham, C.H., Schneider, J.S., Curthoys, I.S., 1989.Anatomical evidence of the projection of pontine omnipauseneurons to midbrain regions controlling vertical eyemovements. J. Comp. Neurol. 289, 610–625.
Oka, H., Yoshida, K., Yamamoto, T., Samejima, A., 1985. Organizationof afferents connections to the lateral and interposed cerebellarnuclei from the brainstem relay nuclei: a horseradish peroxidasestudy in the cat. Neurosci. Res. 2, 321–333.
Paxinos, G., Watson, C., 2007. The Rat Brain in StereotaxicCoordinates, Sixth Ed. Academic Press Elsevier, Amsterdam.
Person, A.L., Raman, I.M., 2012. Synchrony and neural coding incerebellar circuits. Front. Neural Circuits 6, 97.
Pijpers, A., Voogd, J., Ruigrok, T.J., 2005. Topography of olivo-cortico-nuclear modules in the intermediate cerebellum ofthe rat. J. Comp. Neurol. 492, 193–213.
Quy, P.N., Fujita, H., Sakamoto, Y., Na, J., Sugihara, I., 2011.Projection patterns of single mossy fiber axons originatingfrom the dorsal column nuclei mapped on the aldolase Ccompartments in the rat cerebellar cortex. J. Comp. Neurol.519, 874–899.
Rho, M.J., Cabana, T., Drew, T., 1997. Organization of theprojections from the pericruciate cortex to thepontomedullary reticular formation of the cat: a quantitativeretrograde tracing study. J. Comp. Neurol. 388, 228–249.
Richards, J.G., Tranzer, J.P., 1970. The ultrastructural localisationof amine storage sites in the central nervous system with theaid of a specific marker, 5-hydroxydopamine. Brain Res. 17,463–469.
Ruigrok, T.J., Voogd, J., 2000. Organization of projections from theinferior olive to the cerebellar nuclei in the rat. J. Comp.Neurol. 426, 209–228.
Shinoda, Y., Sugiuchi, Y., Futami, T., 1987. Excitatory inputs tocerebellar dentate nucleus neurons from the cerebral cortexin the cat. Exp. Brain Res. 67, 299–315.
Shinoda, Y., Sugiuchi, Y., Futami, T., Izawa, R., 1992. Axoncollaterals of mossy fibers from the pontine nucleus in thecerebellar dentate nucleus. J. Neurophysiol. 67, 547–560.
Sugihara, I., 2011. Compartmentalization of the deep cerebellarnuclei based on afferent projections and aldolase Cexpression. Cerebellum 10, 449–463.
Sugihara, I., Shinoda, Y., 2004. Molecular, topographic, and
functional organization of the cerebellar cortex: a study with
combined aldolase C and olivocerebellar labeling. J. Neurosci.
24, 8771–8785.Sugihara, I., Shinoda, Y., 2007. Molecular, topographic, and
functional organization of the cerebellar nuclei: analysis by
three-dimensional mapping of the olivonuclear projection
and aldolase C labeling. J. Neurosci. 27, 9696–9710.Sugihara, I., Fujita, H., 2010. A computer-aided light microscopy
system for three-dimensional reconstruction of axonal
projections. In: Mendez-Vilas, A., Dıaz, J. (Eds.), Microscopy:
Science, Technology, Applications and Education. Formatex,
Badajoz, pp. 813–819.Sugihara, I., Wu, H.-S., Shinoda, Y., 1999. Morphology of single
olivocerebellar axons labeled with biotinylated dextran amine
in the rat. J. Comp. Neurol. 414, 131–148.Sugihara, I., Wu, H.-S., Shinoda, Y., 2001. The entire trajectories of
single olivocerebellar axons in the cerebellar cortex and their
contribution to cerebellar compartmentalization. J. Neurosci.
21, 7715–7723.Sugihara, I., Ebata, S., Shinoda, Y., 2004. Functional
compartmentalization in the flocculus and the ventral
dentate and dorsal group y nuclei: an analysis of single
olivocerebellar axonal morphology. J. Comp. Neurol. 470,
113–133.Sugihara, I., Fujita, H., Na, J., Quy, P.N., Li, B.Y., Ikeda, D., 2009.
Projection of reconstructed single Purkinje cell axons in
relation to the cortical and nuclear aldolase C compartments
of the rat cerebellum. J. Comp. Neurol. 512, 282–304.Takeuchi, Y., Kimura, H., Sano, Y., 1982. Immunohistochemical
demonstration of serotonin-containing nerve fibers in the
cerebellum. Cell Tissue Res. 226, 1–12.Teune, T.M., van der Burg, J., van der Moer, J., Voogd, J.,
Ruigrok, T.J. H., 2000. Topography of cerebellar nuclear
projections to the brain stem in the rat. Prog. Brain Res. 124,
141–172.Van der Want, J.J. L., Wiklund, L., Guegan, M., Ruigrok, T.J., Voogd,
J., 1989. Anterograde tracing of the rat olivocerebellar system
with Phaseolus vulgaris leucoagglutinin (PHA-L).
Demonstration of climbing fiber collateral innervation of the
cerebellar nuclei. J. Comp. Neurol. 288, 1–18.Voogd, J., 2004. Cerebellum. In: Paxinos, G. (Ed.), The Rat
Nervous System 3rd edition Elsevier Academic Press,
Amsterdam, pp. 205–242.Woolf, N.J., Butcher, L.L., 1989. Cholinergic systems in the rat
brain: IV. Descending projections of the pontomesencephalic
tegmentum. Brain Res. Bull. 23, 519–540.Wu, H.-S., Sugihara, I., Shinoda, Y., 1999. Projection patterns of
single mossy fibers originating from the lateral reticular
nucleus in the rat cerebellar cortex and nuclei. J. Comp.
Neurol. 411, 97–118.Zhang, H., Gamlin, P.D., 1998. Neurons in the posterior interposed
nucleus of the cerebellum related to vergence and
accommodation. I. Steady-state characteristics.
J. Neurophysiol. 79, 1255–1269.





























