Embed Size (px)
Citation preview

DEVELOPMENTAL BIOLOGY 145,128-138 (19%)
Changes in Temporal and Spatial Patterns of Gi Protein Expression in Postimplantation Mouse Embryos
JOLYNDA JONES, CATRIONA Y. LOGAN, AND RICHARD M. SCHULTZ
We previously demonstrated the presence of GTP-binding proteins, G proteins, in the preimplantation mouse embryo (Jones and Schultz, 1990. Dm Bid. 139.250-262). These studies have been extended to the Day 6.5,7.5, and 8.5 gestation embryo by employing PT-catalyzed ADP-ribosylation and immunoblotting techniques. We report here that the amount of embryonic ai increases from Day 6.5 to Day 7.5 of gestation, and remains at about the same level at Day 8.5. In contrast, the extent of PT-catalyzed ADP-ribosylation of G,,, protein(s) decreases between Days 6.5 and 7.5-this de- crease is global and not restricted to a particular germ layer of the Day 7.5 embryo-and then dramatically increases by Day 8.5 of gestation. In the Day 8.5 gestation embryo, the extent of PT-catalyzed ADP-ribosglation of G, proteins increases along the anterior-posterior axis, whereas the amount of immunoreactive q subunit decreases along this axis. By using a combination of PT-catalyzed ADP-ribosylation and immunoprecipitation with antisera specific for ql, q2, or oi3, we report that all three oi subtypes are present in the Day 8.5 gestation mouse embryo. Results of these experiments suggest that an activation of Gi proteins occurs between Days 6.5 and 7.5 of gestation in the postimplantation embryo, a time during which the embryo is gastrulating, and that a decreasing gradient of activation exists along the anterior to posterior axis in the Day 8.5 gestation embryo. Last, we report that oocgtes, eggs, and preimplantation embryos possess all three subtypes of q. * 1991 Academic Press, Inc.
INTRODIJCTION ogenesis (Greene, 1989 and references therein; Haus- man and Velleman, 1981; Runyan et al., 1990; Zalin, 1977). Early postimplantation mouse development is char-
acterized by the sequential differentiation of the inner cell mass of the blastocyst first into a bilaminar embryo, then into a trilaminar embryo, and finally into an em- bryo undergoing organogenesis (Rugh, 1968; Theiler, 1989). This series of events marks a significant transi- tion from a preimplantation embryo that has undergone a series of reductional divisions in the absence of net growth (Hogan et ah, 1986; Brinster, 1967) to a postim- plantation embryo that will undergo rapid changes in growth, differentiation, and morphogenesis (Rugh, 1968; Theiler, 1989).
Signal transduction pathways are implicated in cell proliferation and differentiation, as well as differential gene expression, all of which are requisite for embryo- genesis. In many instances, guanine nucleotide-binding regulatory proteins (G proteins) mediate specific signal transduction pathways, which result in the production of second messengers (Gilman, 198’7, and references therein). In fact, the preimplantation mouse embryo possesses G proteins (Jones and Schultz, 1990; Allworth et al., 1990), and CAMP appears to regulate specific aspects of preimplantation embryogenesis in the mouse (Manejwala et al., 1986, 1989; Poueymirou and Schultz, 1989). Moreover, signal transduction pathways are likely to exist in the early postimplantation embryo, since such pathways are implicated in regulating organ-
G proteins are composed of 01, p, and y subunits and comprise a family of proteins, which vary at least in the nature of the 01 subunit. When activated by hormone/re- ceptor coupling, the LY subunit dissociates from the /3y complex and interacts with its target effector protein. The specificity of the interaction of the 01 subunit with the different effector proteins appears to be due to the different types of N subunits. For example, o(, activates adenylyl cyclase, whereas cyi can inhibit adenylyl cyclase (Gilman, 1987 and references therein; Casey et al., 1988). In addition, aj can activate K+ channels or a phospholi- pase C (Brass et ul., 1986; Ueda et al., 1989; Mattera et ab, 1989).
Three subtypes of ai-cyil, I+, and cY,-exist and have molecular weights in the range of 40,000-41,000 daltons (Gilman, 1987 and references therein; Casey et al., 1988). These Qli subunits, which are products of separate genes (Jones and Reed, 1987; Kaziro et al., 1988), are frequently identified by their ability to serve as substrates for ADP-ribosylation catalyzed by pertussis toxin (PT) in the presence of [32P]NAD; the inactive heterotrimeric form of Gi is the substrate for PT (Bokoch et ah, 1984; Katada et al., 1984).
Previously, we utilized the PT-catalyzed ADP-ribosy- lation of Gi, subunits, as well as immunoblotting, to identify changes in the expression of G,-like proteins
0012-1606/91 $3.00 Copyright IL 1991 by Academic Press, Inc. All rights of reproduction in any form reserved.
128

JONES,LOGAN,ANDSHULTZ G Protcirls in A-1o~t.w E~~~hr,qos 129
during preimplantation mouse development. We have used these methods to extend this study to the early postimplantation embryo and report here that Gi pro- teins are present in the Day 6.5, 7.5, and 8.5 gestation mouse embryo. The amount of Ni increases from Day 6.5 to Day 7.5 of gestation, and remains at about the same level at Day 8.5. In contrast, the extent of PT-catalyzed ADP-ribosylation of Gi, proteins decreases between Days 6.5 and 7.5-this decrease is global and not re- stricted to a particular Day 7.5 embryonic germ layer- and then dramatically increases by Day 8.5 of gestation. Along the anterior-posterior axis two gradients of Gi protein expression are present in the Day 8.5 gestation embryo. The extent of PT-catalyzed ADP-ribosylation of Gi, proteins increases along the anterior-posterior axis, whereas the amount of immunoreactive LYE subunit detected by immunoblotting decreases along this axis. Last, by using a combination of PT-catalyzed ADP-ribo- sylation and immunoprecipitation with antisera spe- cific for Nil, (Yi2, or Nj3, we report that all three ai sub- types are present in the Day 8.5 mouse embryo, as well as in oocytes, eggs, and preimplantation embryos.
MATERIALS AND METHODS
Embryo Collection
Anterior ectoderm, posterior ectoderm and mesoderm (i.e., primitive streak region), and visceral embryonic endoderm were isolated from Day 7.5 embryos by enzy- matic treatment with 0.5% trypsin and 0.25% pancrea- tin in CMF-PBS (Beddington, 1987). Enzymatic treat- ment was inhibited by placing isolated tissues in Dul- becco’s modified media (GIBCO) containing 20% fetal bovine serum (FBS), 0.1 mM sodium pyruvate, 50 pg/ml streptomycin, and 50 IU/ml penicillin. Isolated germ layers were washed extensively in CMF-PBS prior to storage at -70°C in CMF-PBS/PI.
Nulliparous CF-1 female mice (6-8 weeks of age, Har- lan, Sprague-Dawley), two per cage, were housed over- night with single B6D2F,/J males (Jackson Labora- tory). The next morning, females were examined for vag- inal plugs as evidence of mating, and this was designated Day 0.5 of gestation. On Days 6.5,7.5, and 8.5 of gestation, embryos were dissected from the decidua in bicarbonate-free minimal essential medium (Earle’s salts) containing pyruvate (100 pg/ml), gentamicin (10 pg/ml), polyvinylpyrrolidone (3 mg/ml), and 25 mM Hepes, pH 7.2 (MEM/PVP). Riechart’s membrane, the ectoplacental cone, and all extraembryonic membranes were removed from Day 6.5 (stage 9; Theiler, 1989), Day 7.5 (stage 10; Theiler, 1989), and Day 8.5 (stage 12; Theiler, 1989) embryos. All embryos were rinsed in cal- cium, magnesium-free phosphate-buffered saline (CMF-PBS), pH 7.2, and stored at -70°C in CMF-PBS containing 1% Lubrol-PX, leupeptin (10 /*g/ml), apro- tinin (10 pg/ml), and phenylmethylsulfonyl fluoride (10 PIM) (CMF-PBS/PI). Prior to being assayed by the methods described below, the embryos were thawed in the presence of CMF-PBS/PI and homogenized on ice with ten strokes in a Dounce homogenizer. The total amount of embryonic protein was determined using the bicinchoninic acid (BCA, Pierce, Inc.) protein assay (Smith et al., 1985). Mouse oocytes, eggs, and preimplan- tation mouse embryos were collected as previously de- scribed (Jones and Schultz, 1990).
Preparation r?f Plasma Membranes
Crude plasma membrane preparations were derived from normal rat kidney cells (NRK) (Woolkalis and Manning, 1987), platelets (PL) (Brass et uh, 1988), and rat brain (RB) (Sternweis and Robishaw, 1984) as de- scribed previously and were generously provided by Marilyn Woolkalis, Department of Pharmacology, Uni- versity of Pennsylvania.
Pertussis Toxin-Cutalyxed ADP-Ribosylation qf Mouse Embryos, NRK, PL, and RB Membranes
PT-catalyzed ADP-ribosylation of mouse embryonic homogenates and control cellular plasma membrane preparations was performed as previously described (Jones et t~l., 1989). [32P]NAD was synthesized according to the method of Cassel and Pfeuffer (1978), except that an ATP regenerating system, consisting of pyruvate ki- nase and phosphoenolpyruvate, was included. When PT was omitted from the reaction mixture, the radiolabeled ADP-ribosylated species of M, = 38,000 and 39,000 were not detected (Fig. 1). Equal amounts of embryonic pro- tein were radiolabeled and subjected to electrophoresis.
PT-catalyzed ADP-ribosylation of Q, was performed by adding 2 pmole each of purified recombinant rat CY, and purified bovine brain B-y to the reaction mixture at the time of addition of PT. Following the incubation, the
Where stated, female mice were injected with 1 I.U. of pregnant mare’s serum gonadotropin (PMSG), and 48 hr later injected with 1 I.U. of human chorionic gonadotro- pin (hCG); this treatment regime resulted in a higher percentage of females that mated but did not induce superovulation. Females were housed with males over- night and screened for evidence of mating as described above. Embryos collected from gonadotropin-primed mice developed at the same rate as embryos from natu- rally mated mice (data not shown).
Oocytes, eggs, and preimplantation embryos were col- lected as previously described (Jones and Schultz, 1990).
Germ Lager Isolat%on

130 DEVELOPMENTAL BIOLOGY VOLUME 145, 1991
1 2 3 1' 2' 3' Mrx 10-3
45-
29-
FIG. 1. Autoradiograms of day 6.5,7.5, and 8.5 embryo homogenates incubated with or without PT and with [32P]NAD. The experiments were performed as described under Materials and Methods, and 2.5 fig of protein was used for each sample. Lanes 1-3, PT present; lanes l’-3, PT absent. Lanes 1 and I’, Day 6.5 embryos; lanes 2 and 2, Day 7.5 embryos; lanes 3 and 3, Day 85 embryos. The experiment was per- formed three times with similar results. Shown is a representative example. The radiolabeled species in the Day 7.5 gestation embryo (lane 2) are present but did not reproduce well.
samples were alkylated with N-ethylmaleimide as previously described (Sternweis and Robishaw, 1984).
Polyacrylamide Gel Electrophoresis
Samples were subjected to electrophoresis in 10% polyacrylamide gels containing sodium dodecyl sulfate (SDS) according to the method of Laemmli (Laemmli, 1970). [32P]radiolabeled species were detected by autora- diography at -70°C using Kodak X AR-5 X-ray film and an intensifying screen. Exposure times ranged from 1 to 16 hr.
The SDS used in these studies is the 95% grade from Sigma. This SDS, which contains higher chain length homologs relative to more pure forms of SDS, results in resolving two ADP-ribosylated proteins of M, = 38,000 and 39,000. Pure forms of SDS only resolve a single spe- cies of M, = 41,000 (Jones et al., 1989; Jones and Schultz, 1990).
To compare the electrophoretic mobility of ADP-ribo- sylated cy, and embryonic protein that was alkylated, polyacrylamide gel electrophoresis was conducted in 11% gelan and a more pure form of SDS was used. Al- though this resulted in only the separation of a single species for the embryo-derived material, it afforded ample resolution of (Y~ from cy,.
Immunoblotting
Equal amounts of protein from Day 6.5, 7.5, and 8.5 gestation embryos were subjected to electrophoresis in either 10% polyacrylamide-SDS mini-gels (5 cm
length); in contrast to the large gels, the mini-gels do not resolve well the two species that are detected by PT-catalyzed ADP-ribosylation. Following electropho- resis, proteins were transferred to Immobilon-P (Milli- pore) or nitrocellulose (Schleicher and Schuell) as previously described (Jones and Schultz, 1990). Silver staining of mini-gels (Wray et al., 1981) or Coommassie staining of standard gels revealed that essentially com- plete transfer of proteins in the M, = 25,000-100,000 range occurred.
Antisera specific for olilm3, cyil, ai2, ai3, o(,, or cy,, as well as an antiserum that recognizes part of the GTP-bind- ing site common to all o( subunits were used; these anti- sera were generated against the peptides described in Table 1 and were generous gifts from Dave Manning, Department of Pharmacology, University of Pennsyl- vania. Binding of the primary antisera (1:lOO dilution used in all experiments) to specific LY subunits on Immo- bilon-P or nitrocellulose was visualized using either a Vectastain peroxidase kit (Vector) and diaminobenzi- dine (Sigma), or 1251-protein A (Amersham, sp. act. 100 &i/ml). When 1251-protein A was utilized the Immobi- lon-P or nitrocellulose membranes were incubated with the appropriate primary antisera and washed in Tris- HCl-buffered saline (TBS), pH 7.5, as described previously (Jones and Schultz, 1990). Following the wash, the membranes were incubated with 1251-protein A (1 pCi/ml) for 30 min. The membranes were then washed in TBS containing 0.05% Tween-20 and air dried. [1251]protein A labeled proteins were detected by autoradiography at -70°C using Kodak X AR-5 X-ray film.
Immunoprecipitution
Equal amounts of protein from Day 6.5, 7.5 and 8.5 gestation embryos were subjected to PT-catalyzed ADP-ribosylation in the presence of [32P]NAD. The ra- diolabeled proteins were then immunoprecipitated ac- cording to the method of Carlson et al. (1989) except that 1:lO dilutions of the antisera were used. In order to dem- onstrate antisera specificity, the specific synthetic pep- tides described in Table 1 for oil, ai2, or Lyi3 (200 pg/ml) were initially incubated for 8 hr at 4°C with antisera directed against ail, ai2, or ai3, respectively, prior to im- munoprecipitation. The synthetic peptides were a gift from Dave Manning, Department of Pharmacology, Uni- versity of Pennsylvania.
Densitometry
Densitometry was performed on autoradiograms of samples subjected to either PT-catalyzed ADP-ribosy- lation or immunoblotting. For each case, the exposure time for the autoradiograms was such that the increase

131 JONES,LOGAN,ANDSHULTZ G Proff,itcs it/ Mo1r.w Ev~l~qos
TABLE 1 SPECIFICITY 0~ PEPTIDE ANTISERA TO (Y SIJBUNIT SUBTYPES
Antiserum Peptide Recognizes Reference
KNNLKDCGLF CDLDRIAQPNYI CDLERIAQSDYI RADDARAAEGFDIC CEYGDKERKADSK CTGPAESKGEITPELI, CGAGESGKTIVKQMK
Carlson vf trl.. 1989 Williams f’f ul.. 1990
Carlson c,f (I/. 1089 1 . LVilliams fat t/l.. 1990 Personal communication” Carlson c? trl.. 1989 Carlson vf trl., 1989
” This antisera recognizes LYE five times greater than (Y, polypeptides (personal communication, David Manning, Department of Phxrmacolo~~. Ilniwrsity of Pennsglvania).
in densitometer units was linear with respect to either the amount of PT-catalyzed ADP-ribosylation of (Y~ or immunoreactive (yi (data not shown).
RESULTS
Since we have previously demonstrated developmen- tal changes in the pattern of Gi protein expression dur- ing preimplantation development, we extended these studies by examining Gi protein expression during postimplantation development. As development pro- ceeds from Day 6.5 to Day 7.5 (primitive streak stage; stage 10 Theiler, 1989) a marked decrease in the extent of PT-catalyzed ADP-ribosylation was observed (Fig. 1); densitometry revealed an average decrease of about 5- fold (Fig. 2). A dramatic increase in the extent of PT-ca-
loooo l-----l
in LA 6.5 7.5 a.5
Age (days)
FIG. 2. Relative changes in the extent of PT-catalyzed AL)P-ribosy- lation in day 6.5, 7.5, and X.5 embryo homogenates. The autoradio- grams were subjected to densitometry and the data from three sepa- rate experiments have been pooled and are expressed as the mean k SEM relative to the extent of PT-catalyzed ADP-ribosylation in Day 6.5 embryo homogenates. Note that the ordinate is a log scale and that the intensity of the signal obtained for the Day 8.5 embryo mate- rial is beyond the linear range of the densitometer; thus the increase that occurs by Day 8.5 is an underestimate. The differences between any two groups is significant (I’ < 0.02, Student’s t test).
talyzed ADP-ribosylation was observed between Day 7.5 and Day 8.5 of development, at which time organogene- sis is occurring (Fig. 1); densitometry revealed an aver- age increase of about 22-fold relative to Day 6.5 (Fig. 2). The ADP-ribosylated species detected in the postim- plantation embryos had an M, = 38,000 and 39,000, which was very similar, if not identical to that present in preimplantation embryos (Jones and Schultz, 1990).
Results of two mixing experiments that used equal amounts of Day 7.5 and Day 8.5 embryonic protein re- vealed that the extent of PT-catalyzed ADP-ribosgla- tion was additive (data not shown). This result mini- mized the likelihood that an endogenous inhibitor was present in the Day 7.5 embryo homogenate and responsi- ble for the decrease in the extent of PT-catalyzed ADP- ribosylation observed in the Day 7.5 embryo.
Although there is no overt neuronal differentiation in the Day 8.5 embryo (Buse and Krisch, 1987), the increase in the extent of PT-catalyzed ADP-ribosylation could be due to an increase in the amount of G,, which is also a substrate for PT and is abundant in neuronal tissue (Sternweis and Robishaw, 1984). This possibility was most unlikely, since using an electrophoretic system that readily resolves (Y, from (yi (Sternweis and Robi- shaw, 1984), the ADP-ribosylated species present in the Day 8.5 embryo had an electrophoretic mobility that differed from that of recombinant CY, (Fig. 3).
1 2
FIG. 3. Region of an autoradiogram of Day 8.5 embrgonic material and recombinant q, subjected to PT-catalyzed ADP-ribosglation. The experiment was performed as described under Materials and Meth- ods: 1.5 PB of Daq’ 8.5 emhrvo protein and 2 pmole of recombinant o, were used. Lane 1, Day 8.5 embryo; lane 2, recombinant (Y,. The expo- sure times were adjusted to yield relatively similar signal levels. In this gel clectrophorctic system, q migrates slower than q, (Sternweis and Robishaw, 19841.

132 DEVELOPMENTAL BIOLOGY VOLUME 145, 1991
FIG. 4. Immunoblot of Day 6.5,7.5, and 8.5 gestation embryos using antiserum to Gi and the [‘=I]protein A detection method using serum 8730. The experiment was performed as described under Materials and Methods and 12.5 wcg of protein was used for each sample. Lane 1, NRK, 2 pg; lane 2, Day 6.5 embryos; lane 3, Day 7.5 embryos; lane 4, Day 8.5 embryos. Preimmune serum did not recognize the immunore- active q species (data not shown).
Since equal quantities of embryonic protein were sub- jected to PT-catalyzed ADP-ribosylation, the changes in the extent of labeling of 01~ could be attributed to either alterations in the state of dissociation of Gi-the heter- otrimeric, undissociated form of Gi is the substrate for PT (Bokoch et ul., 1984; Katada et al., 1984)-or changes in the amount of Gi, or both. Immunoblotting experi- ments were performed to discriminate between these possibilities.
Immunoblotting experiments were conducted with a polyclonal antibody 8730 (Table 1) that recognizes (Yil and cyiB equally, cyi3 at 50% the level of ail and Ni2, and cy, at 510% the level of ai1 and c+ (Carlson et ah, 1989). The amount of immunoreactive ai was greater in the Day 7.5 and Day 8.5 embryos when compared to the Day 6.5 em- bryo (Figs. 4 and 5). The electrophoretic mobility of this ai subunit (M,. = 38,000) was the same as that of cyi pres- ent in NRK membranes (Marilyn Woolkalis, personal communication; Jones et ul., 1989; Jones and Schultz, 1990).
On Day 7.5, primitive streak formation begins and results in the formation of a trilaminar embryo, which consists of embryonic ectoderm, embryonic mesoderm, and visceral endoderm. Thus, the decrease in the extent of PT-catalyzed ADP-ribosylation that was observed in the Day 7.5 embryo could have been due to a global de- crease or a decrease in a specific germ layer. In order to ascertain whether regional variations in the extent of PT-catalyzed ADP-ribosylation of Gi,, occur in specific Day 7.5 embryonic germ layers, the germ layers of the Day 7.5 embryo were enzymatically isolated (Bedding- tion, 1987), and equal amounts of protein were subjected to PT-catalyzed ADP-ribosylation. Since isolation of em-
Day 6.5 Day 7.5 Day 6.5
FIG. 5. Relative changes in the amount of immunoreactive q in Day 6.5, 7.5, and 8.5 embryo homogenates. The immunoblots were sub- jected to densitometry and the data from two experiments in which the [r”I] detection method was used and one experiment in which the peroxidase detection method was used have been pooled and are ex- pressed as the mean k SEM relative to the amount in the Day 6.5 embryo. When the peroxidase detection method was used, a positive negative was scanned. The increases observed at either Day 7.5 or Day 8.5 relative to Day 6.5 are significant (E < 0.01, Student’s t test) and the differences between Day 7.5 and Day 8.5 are significant (P < 0.02, Student’s t test ).
bryonic mesoderm from the posterior embryonic ecto- derm was difficult, these two tissues were combined. The extent of PT-catalyzed ADP-ribosylation was simi- lar in the embryonic anterior ectoderm, visceral embry- onic endoderm, and embryonic mesoderm plus posterior ectoderm (Figs. 6 and 7). Therefore, the decrease in ADP-ribosylation of ai in Day 7.5 embryos when com- pared to that in the Day 6.5 embryo was not due to a decrease in the extent of PT-catalyzed ADP-ribosyla- tion of Gi in one embryonic germ layer versus another. Also consistent with this interpretation was the obser- vation that immunoblots of protein isolated from Day 7.5 germ layers demonstrated that although the differ- ences in the amount of immunoreactive material in the three germ layers were significant, the three germ layers, nevertheless, contained roughly similar amounts of cyi (Fig. 8); densitometric analysis of these experi- ments is shown in Fig. 7.
123 45 6
&” 7”
FIG. 6. Region of an autoradiogram of Day 7.5 embryonic germ layers subjected to PT-catalyzed ADP-ribosylation. The experiment was performed as described under Materials and Methods. Lanes 1 and 4, 2 ).~cg protein of Day 7.5 embryonic visceral endoderm; lanes 2 and 5, 2 ~g protein of Day 7.5 embryonic anterior ectoderm, lanes 3 and 6, 2 (~g protein of Day 7.5 embryonic mesoderm and posterior ectoderm. The experiment was performed three times with similar results. Results of two experiments are shown.
I I--
Endo
,JONES,LOGAN,ANDSHULTZ
I AE 1
MPE
FIG. 7. Relative changes in the extent of PT-catalyzed ADP-ribosy- lation or amount of immunoreactive CY, in germ layers of Day 7.5 em- bryos Densitometry was performed and the data are expressed as the mean i SEM relative to the value obtained for the endoderm. The results of three PT-catalyzed ADP-ribosylation and four immunoblot- ting experiments were pooled. Solid bars, PT-catalyzed ADP-ribosyla- tion; open bars. immunoreactive q. For the PT-catalyzed ADP-ribo- sylation experiments, the decreases in the anterior ectoderm (AE) and mesoderm and posterior ectoderm (MPE) are significant (P < 0.05, Student’s f test) when compared to endodoerm (ENDO). The differ- ences between AE and MPE are not significantly different. For the amount of immunoreactive (yi, the difference between any two groups is significant (P < 0.001, Student’s t test).
Spafiul Chunges in the Pattern of G Protein Expression in the Day ti.5 Embryo
To ascertain if a gradient of Gi protein expression ex- isted along the anterior-posterior axis of the Day 8.5
1 2 3 1’ 2’ 3’
MI x 1C3
84-
27-
FIG. 8. Immunoblot of Day 7.5 embryonic germ layers using anti- serum to Gi. The experiment was performed as described under Mate- rials and Methods using the [lwI] detection method and serum 8730, and 5 pg of protein was used for each sample. Lanes 1-3, antiserum 8730; lanes l’-3’, normal rabbit serum. Lanes 1 and l’, Day 7.5 embry- onic visceral endoderm; lanes 2 and 2’, Day 7.5 embryonic anterior ectoderm; lanes 3 and 3, Day 7.5 embryonic mesoderm and posterior ectoderm. The chevron points to the region where q migrates in the mini-gel. The experiment was performed three times with similar re- sults. Shown is a representative example. The signal along the outer edges of the lanes is due to nonspecific binding to the prestained molec- ular weight standards.
133
12345678
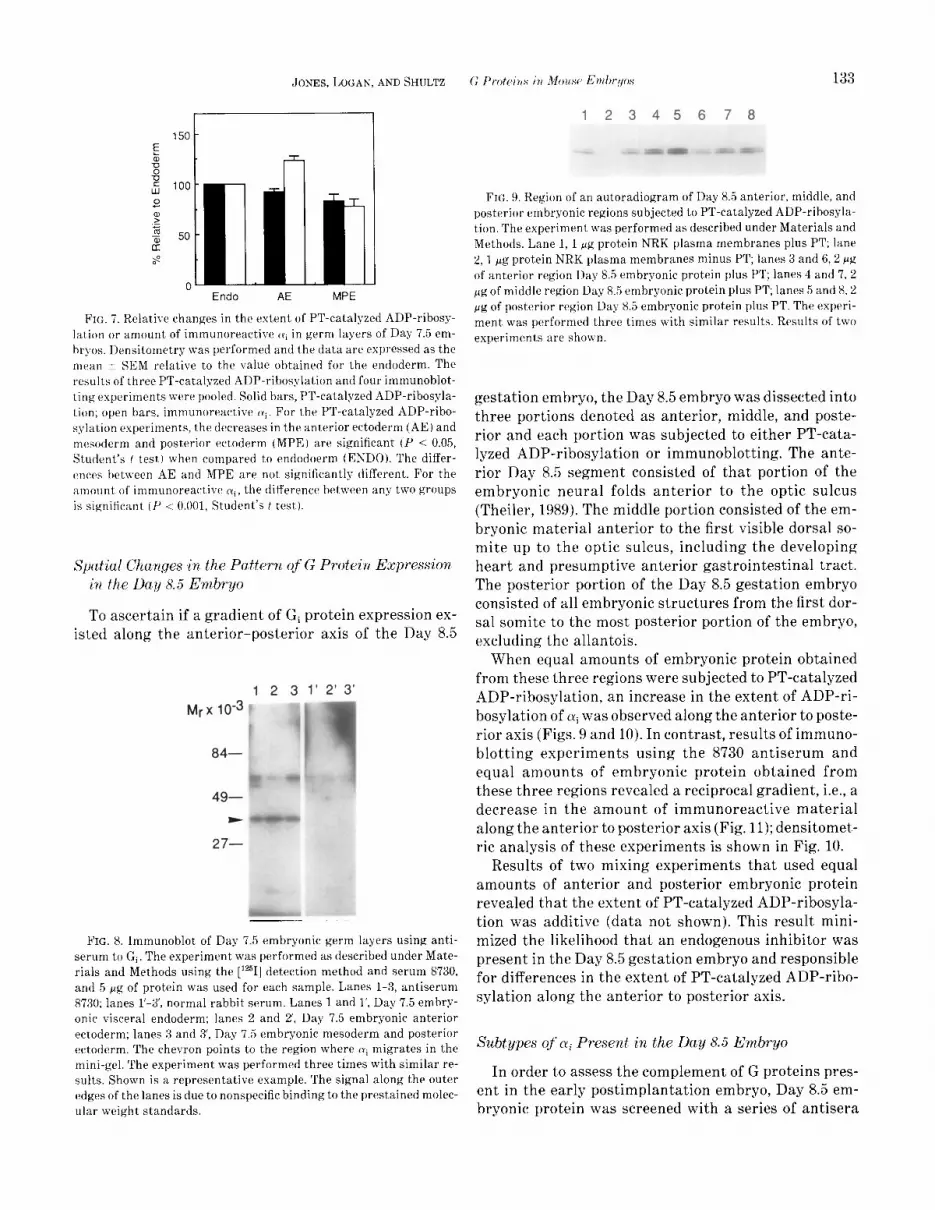
FIG. 9, Region of an autoradiogram of Day 85 anterior, middle, and posterior embryonic regions subjected to PT-catalyzed ADP-rihosyla- tion. The experiment was performed as described under Materials and Methods. Lane 1, 1 gg protein NRK plasma membranes plus PT; lane 8,l pg protein NRK plasma membranes minus PT; lanes 3 and 6,2 p’g of anterior region Day 8.5 embryonic protein plus PT; lanes 4 and 7, 2 pg of middle region Day 8.5 embryonic protein plus PT; lanes 5 and 8,2 fig of posterior region Day X.5 embryonic protein plus PT. The experi- ment was performed three times with similar results. Results of two experiments are shown.
gestation embryo, the Day 8.5 embryo was dissected into three portions denoted as anterior, middle, and poste- rior and each portion was subjected to either PT-cata- lyzed ADP-ribosylation or immunoblotting. The ante- rior Day 8.5 segment consisted of that portion of the embryonic neural folds anterior to the optic sulcus (Theiler, 1989). The middle portion consisted of the em- bryonic material anterior to the first visible dorsal so- mite up to the optic sulcus, including the developing heart and presumptive anterior gastrointestinal tract. The posterior portion of the Day 8.5 gestation embryo consisted of all embryonic structures from the first dor- sal somite to the most posterior portion of the embryo, excluding the allantois.
When equal amounts of embryonic protein obtained from these three regions were subjected to PT-catalyzed ADP-ribosylation, an increase in the extent of ADP-ri- bosylation of ai was observed along the anterior to poste- rior axis (Figs. 9 and 10). In contrast, results of immuno- blotting experiments using the 8730 antiserum and equal amounts of embryonic protein obtained from these three regions revealed a reciprocal gradient, i.e., a decrease in the amount of immunoreactive material along the anterior to posterior axis (Fig. 11); densitomet- ric analysis of these experiments is shown in Fig. 10.
Results of two mixing experiments that used equal amounts of anterior and posterior embryonic protein revealed that the extent of PT-catalyzed ADP-ribosyla- tion was additive (data not shown). This result mini- mized the likelihood that an endogenous inhibitor was present in the Day 8.5 gestation embryo and responsible for differences in the extent of PT-catalyzed ADP-ribo- sylation along the anterior to posterior axis.
Subtypes of q Present in the Day 8.5 Embryo
In order to assess the complement of G proteins pres- ent in the early postimplantation embryo, Day 8.5 em- bryonic protein was screened with a series of antisera

134 DEVELOPMENTAL BIOLOGY VOLUME 145, 1991
A M P
FIG. 10. Relative changes in the extent of PT-catalyzed ADP-ribosy- lation and amount of immunoreactive q in different regions of the Day 8.5 embryo. Densitometrp was performed and the data are ex- pressed as the mean i- SEM relative to the value obtained for the anterior region. The results of three PT-catalyzed ADP-ribosylation and four immunoblotting experiments were pooled. Solid bars, PT- catalyzed ADP-ribosylation; open bars, immunoreactive q. For the PT-catalyzed ADP-ribosylation experiments, the increases in the middle (M) and posterior (P) regions are significant (1’ < 0.02, Stu- dent’s t test) when compared to the anterior (A) region; the differ- ences between the M and P regions are also significant (P < 0.0.5, Student’s t test). Similarly, for the amount of immunoreactive (yi, the decreases in M and P are significant (P < 0.001, Student’s f test) when compared to A, and the difference between M and P are significant (P < 0.025, Student’s t test).
directed against the specific peptides listed in Table 1. Day 8.5 gestation embryos, which were from gonadotro- pin-primed mice, were used for two reasons. First, suffi- cient quantities of Day 8.5 embryonic protein could be obtained easily for the immunoblotting and immuno-
Mrx lo3 1 2 3 4 5 1' 2' 3' 4' 5'
84-
FIG. 11. Immunoblot of anterior, middle, and posterior Day 8.5 em- bryonic regions using antiserum to Gi. The experiment was performed as described under Materials and Methods using the peroxidase detec- tion method and serum 8’730, and 6 pg of protein was used for each sample. Lanes l-5, antiserum 8730; lanes l’-5’, normal rabbit serum. Lanes 1 and l’, 4 pg protein NRK plasma membranes; lanes 2 and 2’,2 pg protein NRK plasma membranes; lanes 3 and 3, anterior region Day 8.5 embryonic protein; lanes 4 and 4’, middle region Day 8.5 embry- onic protein; lanes 5 and 5, posterior region Day 8.5 embryonic pro- tein. The chevron points to the region where q migrates in the mini- gel. The experiment was performed three times with similar results. Shown is a representative example.
123456789 10 11
A
B
FIG. 12. Region of an autoradiogram of NRK plasma membranes and Day 8.5 gestation embryo homogenates subjected to PT-catalyzed ADP-ribosylation and immunoprecipitation with antisera listed in Ta- ble 1. The experiment was performed as described under Materials and Methods. (A) NRK plasma membranes (5 eg protein each lane); (B) Day 8.5 gestation embryo homogenates (5 pg protein each lane). Lane 1, normal rabbit serum; lane 2, 8730 antiserum; lane 3,3646 anti- serum; lane 4, 3646 antiserum initially incubated with 200 Kg/ml ql specific synthetic peptide; lane 5, 1521 antiserum; lane 6, 1521 anti- serum initially incubated with 200 @g/ml qZ specific synthetic pep- tide; lane 7,1518 antiserum; lane 8,1518 antiserum initially incubated with 200 pg/ml q3 specific synthetic peptide; lane 9, 2918 antiserum: lane lo,1398 antiserum; lane 11.2919 antiserum. The experiment was performed two times with similar results. Note that the two bands present in lane 3 did not reproduce well in the photograph. In addition, the two bands present in lanes 2 and 10 in each panel are not obvious, since the autoradiogram is overexposed for these lanes.
precipitation procedures. Second, Day 8.5 of gestation marks the beginning of organogenesis (Rugh, 1968; Theiler, 1989) and G proteins modulate a series of sec- ond messenger pathways known to regulate organ for- mation. For example, CAMP is implicated in palate (Greene, 1989 and references therein) and muscle (Hausman and Velleman, 1981; Zalin, 1977) develop- ment, while protein kinase C and G proteins are impli- cated in cardiac development (Runyan et al., 1990).
Results of immunoblotting experiments that used 100 pg of protein from Day 8.5 gestation embryos and the antisera listed in Table 1 revealed a consistent signal for ai (1521) and GTP-binding site common (1398) antisera when either the Vectastain or [lz51] protein A method of detection was used (data not shown). Only the Vecta- stain method of detection demonstrated a weak signal for cr, using the 2919 antiserum (data not shown).
Since efforts to increase the sensitivity of detection of (yil, (yi3, and 01, in the immunoblots were not successful, immunoprecipitation of the PT-catalyzed ADP-ribosy- lated species in the Day 8.5 gestation embryo was used. Antisera 8730 and 1398 (Table 1) immunoprecipitated species of M,. = 38,000 and 39,000 from Day 8.5 embryo homogenates, as well as from membrane preparations derived from platelets, NRK cells, or rat brain, which served as positive controls (Figs. 12 and 13).
Antiserum 3646, which is specific for Lyi1, also immu- noprecipitated both of these radiolabeled species, except that the species of AJ, = 39,000 was predominant in the Day 8.5 embryonic protein samples as compared to the M, = 38,000 species (Fig. 12). A similar situation was observed for rat brain, whereas an equal intensity of

JONES, LOGAN, AND SHULTZ G P,Y)l(Ji,,S in Mouse Etrtbr.,/ps 135
12345678 Subtypes of ai Presed in Oocytes, Eggs, U&
A **- Preimphltution Embryos
The amount of (yi is relatively constant from the GV- B intact oocyte to the two-cell embryo stage (Jones and
Schultz, 1990; Allworth et ub, 1990). Relative to the two-
FIG. 13. Region of an autoradiogram of rat brain plasma mem- cell embryo, a reduced amount of cyi is present in the
branes and platelet plasma membranes subjected to PT-catalyzed eight-cell and morula stages and this amount increases
ADP-ribosylation and immunoprecipitation with antisera listed in Ta- by the blastocyst stage (Jones and Schultz, 1990; All- hle 1. The experiment was performed as described under Materials worth ef al., 1990). In order to ascertain the different and Methods. (A) Rat brain plasma memhranes (5 kg protein each lane); (B) platelet plasma membranes (5 PK protein each lane). Lane 1,
types of cyi present and whether developmental changes
normal rabbit serum; lane 2, 8730 antiserum; lane 3, 3646 antiserum; in the types of ai expression occurred during this devel-
lane 4,1521 antiserum; lane .i, 1518 antiserum; lane 6,2918 antiserum; opmental period, a similar immunoprecipitation ap-
lane 7, 1398 antiserum; lane 8, 2919 antiserum. The experiment was preach to that described for the Day 8.5 embryo was performed two times with similar results. undertaken.
Similar to the Day 8.5 embryo, oocytes, ovulated eggs,
both M, = 38,000 and 39,000 species was present in plate- and preimplantation embryos all possessed CQ, Q, and
let plasma membranes, and 1M, = 38,000 species was (xi31 and cyi2 was apparently the most abundant species
more predominant in NRK plasma membranes (Figs. 12 (Fig. 14). In addition, the relative amounts of each of
and 13). These changes in intensity of the two immuno- these subtypes of Ni remained essentially constant.
precipitated species could reflect differences in either their relative amounts present in the homogenate or the efficiency of radiolabeling of the two species. In addi- tion, it may have been possible that these differences were due to other (Yi subtypes that may have been recog- nized by this antiserum.
DISClJSSION
Within the limits of detection, antiserum 1521, which is specific for Ni2, only precipitated the M,. = 38,000 spe- cies, and likewise antiserum 1518, which is specific for Q, only precipitated the M, = 39,000 species in the Day 8.5 embryo and the other cell types tested (Figs. 12 and 13). The immunoprecipitation was specific, since an ini- tial incubation of the antisera directed against Nil (3646), (yi2 (1521), or ai (1518) with the appropriate pep- tide used to generate each of these sera resulted in a dramatic reduction in the amount of immunoprecipi- tated material (Fig. 12).
In this report we extend the results of our previous study that documented developmental changes in the amount of ai and extent of PT-catalyzed ADP-ribosyla- tion of cyi in the mouse preimplantation embryo (Jones and Schultz, 1990) by performing similar experiments with the early postimplantation embryo. We observe that the amount of (yi increases almost 2-fold from Day 6.5 to Day 7.5 and then decreases in the Day 8.5 embryo
1 2 3 4 5
oocyte I”-... -
Little radiolabeled immunoprecipitate was observed when antiserum 2918, which is specific for LY,, was used and compared to normal rabbit serum (Figs. 12). Al- though this antiserum did not efficiently immunopre- cipitate radiolabeled proteins from rat brain membrane preparations (Fig. 13), which are known to contain G, (Marilyn Woolkalis, personal communication; Jones and Reed, 1987; Sternweis and Robishaw, 1984), our previous results which demonstrated that the electrophoretic mo- bility of the ADP-ribosylated species present in the em- bryo differed from that of CY, indicated that CY, was ei- ther not present or present at very low levels in the embryo. Since G,, does not contain the requisite cys- teine residue for PT-catalyzed ADP-ribosylation (Fong ef al., 1988), as expected, antiserum 2919, which is spe- cific for cy,, did not immunoprecipitate radiolabeled pro- teins in any instance (Figs. 12 and 13).
8-cell
morula
blastocyst
FIG. 14. Region of autoradiograms of oocytes, eggs, and preimplan- tation embryos subjected to PT-catalyzed ADP-ribosylation and im- munoprecipitation with antisera listed in Table 1. Lane 1, normal rab- bit serum; lane 2,873O antiserum; lane 3,3546 antiserum; lane 4,152l antiserum; lane 5, 1518 antiserum.

136 DEVELOPMENTALBIOLOGY VOLUME 145,lW
to a value about 50% greater than that observed in the Day 6.5 embryo. In contrast, the extent of PT-catalyzed ADP-ribosylation decreases about 5-fold between Day 6.5 and Day 7.5, and then increases about 22-fold from Day 7.5 to Day 8.5. The decrease in the extent of ADP-ri- bosylation that occurs between Day 6.5 and Day 7.5 ap- pears not to be restricted to a specific germ layer. In the Day 8.5 embryo, which contains all three ai subtypes, the amount of oli decreases along the anterior to poste- rior axis, whereas the extent of PT-catalyzed ADP-ribo- sylation increases along this axis. In addition, we docu- ment that oocytes, eggs, and preimplantation embryos contain all three subtypes of ai and that the relative amounts of each species remains relatively constant during this period of development; thus, the changes in expression of cui that occur during preimplantation de- velopment are coordinate.
The results of the immunoprecipitation experiments provide the likely explanation for resolving two ADP- ribosylated species under our electrophoretic conditions that employ 95% pure SDS (Jones et al., 1989). Both ai1 and oli3 have an electrophoretic mobility less than that of oli2. The apparent equal intensity of the signals is also accounted for by the observation that the intensity of the signal from ai is greater than that of either ai1 and ai3. The presence of a doublet for ai1 may represent some proteolysis or post-translational modification, e.g., phosphorylation (Carlson et al., 1989; Gundersen and Devreotes, 1990).
The decrease in the extent of PT-catalyzed ADP-ribo- sylation of cyi between Day 6.5 and Day 7.5 of gestation occurs at a time when the amount of cyi increases. Since the heterotrimeric form of Gi is the substrate for PT (Bokoch et al., 1984; Katada et ab, 1984), this decrease in the extent of PT-catalyzed ADP-ribosylation may re- flect an increase in the amount of dissociated Gi pro- teins that are present in the Day 7.5 embryo and are capable of interacting with their appropriate effector molecules.
The decrease in the extent of PT-catalyzed ADP-ribo- sylation is not restricted to a specific germ layer(s) in the Day 7.5 embryo. Consistent with this global decrease is that similar amounts of cyi, as detected by immuno- blotting, appear to be present in each of the germ layer dissections. It is possible, however, that the enzymatic method employed to isolate the germs layers may affect the ability of the Gi proteins, which are membrane- bound, to serve as substrates for PT-catalyzed ADP-ri- bosylation, and thus artifactually mask regional changes in the extent of PT-catalyzed ADP-ribosyla- tion.
The potential global activation of Gi occurs between a time when the bilaminar embryo becomes trilaminar, i.e., the formation of mesoderm as a result of massive
cell migration in the gastrulating embryo. It will be of interest to ascertain if the decrease in the extent of PT- catalyzed ADP-ribosylation occurs prior to, concomi- tant with, or subsequent to the initiation of gastrula- tion. In this regard it will also be interesting to define more precisely when the increase occurs in the extent of PT-catalyzed ADP-ribosylation between Day 7.5 and Day 8.5; this increase occurs during a period of time in which the amount of ai remains essentially constant and is consistent with a functional inactivation of Gi. Again, results of such studies may indicate roles for Gi during this period of time in which organogenesis commences.
It should be noted, however, that the changes in ex- tent of PT-catalyzed ADP-ribosylation could be due to changes in the amount of 07 present in the developing embryo, since the heterotrimeric form of Gi is the sub- strate for PT. In fact, results of a recent study indicate that a preferential decrease in the amount of Pr relative to N occurs during mouse oocyte maturation; ,+y de- creases about 69% whereas 01 decreases about 40% (All- worth et al., 1990). The maturation-associated decrease in the extent of PT-catalyzed ADP-ribosylation of cyi cor- relates with these decreases in Gi protein subunits (Jones and Schultz, 1990; Allworth et al., 1990). If such changes in the ratio of o( to /3r are the molecular basis for the observed changes in the extent of PT-catalyzed ADP-ribosylation of 01~ that we report here, then the dramatic loo-fold increase in the extent of PT-catalyzed ADP-ribosylation that occurs between Day 7.5 and Day 8.5 may reflect a pronounced accumulation of 07. It should be noted that this accumulation of /3r would oc- cur during a period of time in which the results of immu- noblotting experiments indicate that the amount of (Y~ stays relatively constant. Whether such changes in p subunit expression occur during this period of develop- ment could be examined by immunoblotting experi- ments using antibodies to the /3 subunit.
The Day 8.5 embryo has several subtypes of Cyi present -ai1-3 -that are detected by antibodies raised to pep- tides specific for the different types of N subunits. The most prevalent form detected by either immunoprecipi- tation following ADP-ribosylation or immunoblotting is Lyiz. It is not known, however, whether this reflects the greater abundance of this type of LY protein, or if the antiserum that detects this subtype is better in both the immunoprecipitation and immunoblotting experiments. Regardless, it is clear that the Day 8.5 embryo contains aile3, and possibly N,. It is unlikely that (Y, is present in readily detectable amounts in the Day 8.5 embryo, since the electrophoretic mobility of the ADP-ribosylated species present in the embryo differs from that of recom- binant cy,. Results of immunocytochemical experiments using specific antisera or in situ hybridization experi- ments using specific riboprobes may reveal spatial pat-

JONES, LOGAN, AND SHULTZ 137
terns of expression that suggest possible functions for these different 01 subtypes.
Two opposing gradients along the anterior to poste- rior axis are present in the Day 8.5 gestation embryo; the amount of ai decreases along this axis, whereas the extent of PT-catalyzed ADP-ribosylation of q in- creases. Thus, when compared to the posterior region, the anterior region apparently has greater amounts of G, protein, and, furthermore, more of this protein is in a potentially activated form. It is interesting to note that this anterior to posterior gradient in the amount and level of potential activity of the Gi follows the anterior to posterior gradient of embryonic development (Rugh, 1968; Theiler, 1989). The anterior structures of the em- bryo develop prior to the posterior portions of the em- bryo as is evident in both somite and limb formation (Rugh, 1968; Theiler, 1989). This same anterior to poste- rior gradient is evident for the protein product of the oncogene id-1 (Wilkinson et al., 1987), terminal differ- entiation proteins such as muscle myosins (Sassoon et trl., 1989), and for transcripts of homeobox genes Hox 1.1 (McMahon ef ul., 1988), 1.3,1.4,1.5,3.1, and 6.1 (Gaunt ef (II., 1988). The pattern of Gi protein expression and acti- vation reported here may be another manifestation of this developmental gradient.
This research was supported by a grant from the NIH (HD 22681) to R.M.S. J.J. thanks the Manning lab for their )qnerous suppI>- of re- agents and constant advice on their use. We are very grateful to Drs. Maurine Linder and Patrick Cases forgenerouslg providing the recom- binant (Y, and [j-y, respectivclg. We also thank Greg Kopf, Dave Man- ning. and Marilyn Roolkalis for critically reading the manuscript and making useful sug,rgestions, as well as the reviewers for their construc- tivc, comments.
REFERENCES
ALLWORTH, A. E., HILDERBRANDT, J. D., and ZIOMEK, C. A. (1990). Differential regulation of G protein suhunit expression in mouse ooqtes, eggs, and earlg embryos. DC,,,. Biol. 142, 129-137.
BEDDINGTON, R. (1987). Isolation, culture and manipulation of post- implantation mouse embryos. 11, “Mammalian development” (M. Monk, Ed.), pp. 43-69. IRL Press, Oxford.
BOKOCH, G. M., KATADA, T., NORTHUP, J. K., UI, M., and GILMAN, A. G. (1984). Purification and properties of the inhihitorg guanine nucleo- tide-hindinK regulatory component of adenylate cq’clasc. tJ, Bid. (‘hc’m. 2.59, 3560-3567.
BRASS, I,. F., LAPOSATA, M., BANGA, H. S., and RITTENHOUSE, S. E. (1986). Regulation of phosphoinositide hgdrolysis pathway in thromhin-stimulated platelets by a pertussis toxin-sensitive gua- nine nucleotide-binding protein. Evaluation of its contribution to platelet activation and comparisons with the adenylate cyclase in- hibitory protein, Gr. J. Bird C//o,/. 261, 16X38-16847.
BRASS, L. F., WOOLKALIS, M. J., and MANNING, D. R. (19X8). Interac- tions in platelets between G proteins and the agonists that stimu- late phospholipase (: and inhibit adenylgl cgclase. ,I. Bid. Chon. 263, 534X-5‘355. . . < ?
BRINSTER, R. I,. (1967). Protein content of the mouse embryo during thr first ti\-cx days of development. Rc/~occ’. Fczrtil. 113, 413-420.
BUSE, E., and KIRSCH, B. (1987). The mouse neural plate as starting material for studying neuronal differentiation in r?tro. Amt. En- brqol. (Bf~rliw) 175, 331-340.
CARLSON, K. E., BRASS, L. F., and MANNING, D. R. (1989). Thrombin and phorbol esters cause the selective phosphorylation of a G pro- tein other than Gi in human platelets. J. Bid. Chem. 264, 1329% 13305.
CASEY, P. J., GRAZIANO, M. P., FREISSMUTH, M., and GILMAN, A. G. (1988). Role of G Proteins in Transmembrane Signaling. ColdSpri?lq Hurlxrr Sp~p Quo nt. Bid. 53, 203-208.
CASSEL, N., and PFEUFFER, T. (1978). Mechanism of cholera toxin ac- tion: Covalent modification of the guanyl nucleotide binding protein of the adenylate cyclase system. Pror. Nafl. ilccxd. Sci. USA 75,2669- 2673.
FONG, H. K. W., YOSHIMOTO, K. K., EVERSOLE-COLE, P., and SIMON, M. I. (1988). Identification of a GTP-binding protein o subunit that lacks an apparent ADP-ribosylation site for pertussis toxin. Proc Nutl. Artrtl. Sri. USA 85, 3066-3070.
GAUNT, S. J., SHARPE, P. T., and DUBOULE, D. (1988). Spatially re- stricted domains of homeo-gene transcripts in mouse embryos: Re- lation to a segmented body plan. &t>eloprnc~nf 104, (Suppl.), 169h 179.
GILMAN, A. G. (1987). G proteins: Transducers of receptor-generated signals. rlr~cr. Rw. Biochrnt. 56, 615-649.
GREENE, R. M. (1989). Signal transduction duringcraniofacial develop ment. (‘rif. RfJr. To.ricd. 20, 137-152.
GUNDERSEN, R. E., and DEVREOTES, P. N. (1990). In !liro reccptor-me- diated phosphorrlation of a G protein in Dictyostelium. Sric~ce 248, 591-593.
HAUSMAN, R., and VELLEMAN, S. (1981). Prostaglandin E, receptors on chick embryo myohlasts. Biochenc. Biophys. Kes. Cm muv. 103, 213- 218.
HOGAN, B., COSTANTINI, F., and LACY, E. (1986). “Manipulating the Mouse Embryo.” Cold Spring Harbor Lahoratory, Cold Spring Har- bor, NY.
JONES, D., and REED, R. R. (1987). Molecular cloning of live GTP-bind- ing protein cDNA species from rat olfactory neuroepithclium. J. Biol. C%o,r. 262, 14241-14249.
JONES, J., KOPF, G. S., and SCHULTZ, R. M. (1989). Variability in the electrophoretic mohilitr of G,-like proteins: Effects of sodium do- tlecvl sulfate. FEBS Mt. 243, 409-412.
JONES, J., and SCHULTZ, R. M. (1990). Pertussis toxin-catalyzed ADP- ribosylation of a G protein in mouse oocgtes, eggs, and preimplanta- tion embryos: Developmental changes and possible functional roles. Der~. Bid. 139 250-262.
KATADA, T., NO;T~UP, J. K., BOKOCH, G. M., UI, M., and GILMAN, A. G. (1984). The inhibitory guanine nucleotide-bindinaregulatorrcompo- nent of aden$ate qclase: Suhunit dissociation and guanine nucleo- tide-dependent hormonal inhibition. J. Bid. Cheat. 259, 357S3585.
KAZIRO Y., ITOH, H., KOZASA, T.. TOYAMA, R., TSUKAMOTO, T., MAT- SUOKA. M., NAKAFUKU, M., OBARA, T., TAKAGI, T., and HERNANDEZ, R. (198X). Structures of the genes coding for G-protein o subunits from mammalian and >Teast cells. Co/t1 S’l~i?ql Ho&or S!/n/p. @cutrt. Bid. 53 309-220. t,-.
LAEMMLI, U. K. (1970). Cleavage of structural proteins during the as- sembly of the head of bacteriophage T4. Nuture 227,680-685.
MAHON, K. A., WESTPHAL, H., and GRUSS, P. (1988). Expression of homeobox gene Ho.r 1.1 during mouse emhryogenesis. Df~cv,lol~tt/etct 104, (Suppl.), 187-195.
MANEJWALA, F., CRAGOE, E., JR., and SCHULTZ, R. M. (1989). Blasto- coel formation in the preimplantation mouse embryo: Role of extra- cellular sodium and chloride and possible apical routes of their entry. I&(~. Biol. 133, 210-220.
MANEJWALA. F., KAJI, E., and SCHULTZ, R. M. (1986). Development of

138 DEVELOPMENTAL BIOLOGY VOLUME 145, 1991
activatable adenylate cyclase in the preimplantation mouse embryo and a role for cyclic AMP in blastocoel formation. Cell 46, 95-103.
MATIERA, R., YATANI, A., KRISCH, G. E., GRAF, R., OKABE, K., OLATE, J., CODINA, J., BROWN, A. M., and BIRNBAUMER, L. (1989). Recombi- nant rui-3 subunit of G proteins activates G,-gated K’ channels. .J, Bid. Chem. 264, 465-471.
POLJEYMIROU, W. T., and SCHULTZ, R. M. (1989). Regulation of the mouse preimplantation development: Inhibition of synthesis of pro- teins in the two-ceil embryo that require transcription by inhibitors of CAMP-dependent protein kinase. Dev. Bid. 133,588-599.
RUGH, R. (1968). “The Mouse.” Burgess Publishing Co., Minneapolis. RUNYAN, R. B., Porrs, J. D., SHARMA, R. V., LOEBER, P. C., CHIANG,
J. J., and BHALLA, R. C. (1990). Signal transduction of a tissue inter- action during embryonic heart development. (%/I Requl. 1, 301-313.
SASSOON, D., LYONS, G., WRIGHT, W. E., LIN, V., LASSAR, A., WEIN- TRAUB, H., and BUCKINGHAM, M. (1989). Expression of two myogenic regulatory factors myogenin and MyoDl during mouse embryogen- esis. Nntnre 341, 303-307.
SMITH, P. K., KROHN, R. I., HERMANSON, G. T., MALLIA, A. K., GARTNER, R. H., PROVENZANO, M. D., FUJIMOTO, E. K., GOEKE, N. M., OLSON, B. J., and KLENK, D. C. (1985). Measurement of protein using bicinchoninic acid. Ancrl. Biochem 150, 76-85.
STERNWEIS, P. C., and ROBISHAW, J. D. (1984). Isolation of two pro-
teins with high affinity for guanine nucleotides from membranes of bovine brain. J. Biol. Chern. 259, 13,806-13,813.
THEILER, K. (1989). “The House Mouse.” Springer-Verlag, New York. UEDA, H., YOSHIHARA, Y., MISAWA, H., FUKUSHIMA, N., KATADA, T.,
UI, M., TAKAGI, H., and SATOH, M. (1989). The kyotorphin (tyrosine- arginine) receptor and a selective reconstitution with purified Gi, measured with GTPase and phospholipase C assays. J. Biol. Chew. 264, 3732-3741.
WILKINSON, D. G., BAILES, J. A., and MCMAHON, A. P. (1987). Expres- sion of the proto-oncogene inf-1 is restricted to specific neural cells in the developing mouse embryo. Cell 50,79-88.
WILLIAMS, A. G., WOOLKALIS, M. J., PONCZ, M., MANNING, D. R., GER- WIRTZ, A. M., and BRASS, L. F. (1990). Identification of the pertussis toxin-sensitive G proteins in platelets, megakaryocytes, and HEL cells. Blood 76, 721b730.
WOOLKALIS, M. J., and MANNING, D. R. (198’7). Structural characteris- tics of the 35. and 36-kDa forms of the 0 subunit common to GTP- binding regulatory proteins. Mol. Phan?Lacol. 32,1-6.
WRAY, W., BOULIKAS, T., WRAY, V. P., and HANCOCK, R. (1981). Silver staining of proteins in polyacryiamide gels. Ad. Biochem. 118, 197-203.
ZALIN, R. (1977). Prostaglandins and myoblast fusion. Dee Bid. 59, 241-248.





























