Embed Size (px)
Citation preview

sustainability
Article
Characterisation of Chamaecytisus tagasaste,Moringa oleifera and Vachellia karroo Vermicompostsand Their Potential to Improve Soil Fertility
Mahlare Mapula Mokgophi 1, Alen Manyevere 2,*, Kingsley Kwabena Ayisi 1 andLawrence Munjonji 1
1 Risk and Vulnerability Science Centre, University of Limpopo, Private Bag X 1106, Sovenga 0727,South Africa; [email protected] (M.M.M.); [email protected] (K.K.A.);[email protected] (L.M.)
2 Department of Agronomy, University of Fort Hare, Private Bag X1314, Alice 5700, South Africa* Correspondence: [email protected]
Received: 29 August 2020; Accepted: 15 October 2020; Published: 10 November 2020�����������������
Abstract: Poor soil fertility and land degradation limit crop production among smallholder farmers.The practice of agroforestry with leguminous trees has proven to be sustainable as it bolsters nutrientsupply through nitrogen fixation and nutrient cycling. The beneficiation of agroforestry species couldadd even more value by using tree based waste materials as mulch or vermicomposting. A study wasconducted to investigate the impact of vermicomposting on chemical and biological characteristics ofthree agroforestry species; Chamaecytisus tagasaste, Vachellia karroo and Moringa oleifera. Eisenia fetidaearthworms were added to the leaves and small twigs of the three trees in worm composting bins.The worms were allowed to feed on the feedstocks for six weeks under laboratory conditions.The results showed that vermicomposting significantly enhanced macronutrient nutrient content inall the three feedstocks. The findings also showed that the quality of the vermicompost depends on thefeedstock type. M. oleifera had the best quality vermicomposts with a significantly higher compositionof macronutrients which ranged between 50 and 170% higher for Ca, K, Mg and P. Vermicompostingincreased Mo while other micronutrients such as Zn, Mn, Fe and B significantly decreased withvermicomposting time. In addition, vermicomposting increased E. fetida reproduction with morethan a 450% increase in earthworm numbers in all three feedstocks. In conclusion, vermicomposthave potential to be used to improve soil fertility and thus reduce the use of synthetic fertilisers incrop production.
Keywords: agroforestry; macronutrients; micronutrients; soil fertility; vermicompost
1. Introduction
Agroforestry is a land use practice in which selected woody perennial species, mainly trees andshrubs are grown in association with herbaceous crops, pastures or livestock where both ecological andeconomic interactions occur between the woody species and the other components [1]. Agroforestrytrees have multiple benefits such as bolstering nutrient supply through nitrogen (N) fixation andnutrient cycling [2], improved soil health, structure and water infiltration [3] and enhanced carbon (C)storage both above-ground and below-ground [4]. The trees and shrubs contribute to food securitydirectly in the form of fruits, seeds and other edible parts or indirectly by maintaining and restoringsoil fertility and water resources, which subsequently increase agricultural production [5].
Though agroforestry is a resilient and sustainable system for resource-poor farmers, its adoptionas an intervention strategy is not widespread in South Africa [6]. The adoption of agroforestry infarming systems in conjunction with other conservation agriculture techniques is being promoted
Sustainability 2020, 12, 9305; doi:10.3390/su12229305 www.mdpi.com/journal/sustainability

Sustainability 2020, 12, 9305 2 of 18
as one of the location specific strategies, and an affordable science based approach to improve landcare among smallholder food production in Limpopo Province [6]. Reconnaissance surveys haveestablished that only passive agroforestry systems exist in Limpopo province [6]. The main agroforestryspecies that have been extensively utilised include tree legumes such as Acacia spp. ((Carl.) Martius),Faidherbia albida ((Delile) A. Chev.), Sesbania sesban ((Elmer) Merr.), Gliricidia sepium ((Jacq.) Steud) andChamaecytisus tagasaste ((L.). These species provide high quality biomass and residue that can be usedfor soil fertility improvement. The low C/N ratio of C. tagasaste foliage could be an indication that itsincorporation may improve soil N and increase yields of non-leguminous crops such as maize [7].The foliage of C. tagasaste contains 18–22% crude protein which provides a good source of calcium,vitamins and minerals [8]. Large volumes of organic materials generated could pose a problem forsafe disposal. Beneficiation of agroforestry species has added more value to the concept of agroforestryby introducing the use of tree based waste material as mulch or vermicompost among others [9].
Agroforestry tree species such as Vachellia karroo ((Hayne) Banfi & Galasso), Moringa oleifera (Lam.)and C. tagasaste could be used as feedstock in the process of vermicomposting. Vermicomposting isan environmentally friendly practice of using organic wastes and incorporating them with selectedearthworms in order to form a humus-like byproduct that is used as a source of nutrients for theplant growth [10]. Vermicomposting involves the use of earthworms to mix, fragment and aerateorganic waste material making it more conducive to microbial activity [10]. It reduces the harmfuleffects of waste material through biodegradation and releases nutrients that can be used for soilfertility improvement [11]. The mutual action of digestion in guts of earthworms and microbesfragment the waste into fine homogeneous humus manure, which is odor-free and rich in nutrients.Vermicomposting reduces the C:N ratio and retains more N than the traditional methods of preparingcomposts [12].
Vermicomposting could also be used as a management strategy where encroachment of the invasiveN-fixing Acacia species on farms is uncontrollable. The Vachellia spp., in particular are indiscriminateaggressive invasive species which generate large quantities of biomass and are widespread in thesavanna and grassland biomes of Limpopo Province [13]. M. oleifera, which is widely cultivatedaround farms in Limpopo Province has the potential to be a dominant agroforestry species due to itsfast growth rate, exceptionally high nutritional value of its leaves, fruit, flowers, and immature podscompared with other food crops and a range of medicinal uses [14–17].
Vermicomposting has the ability to biodegrade the toxic metabolites such as polyphenols, tannins,phytotoxins and saponin produced by M. oleifera [18,19] into harmless nutrients that could be usedfor plant nutrition. C. tagasaste is a legume plant that nodulates freely and can fix atmosphericN in association with the rhizobium bacteria, thereby reducing the requirements for additionalsoil N fertilisation [8]. The three species provide high quality biomass and residue feedstock forvermicomposting and for the improvement of soil fertility.
This study sought to (i) characterise the chemical composition of tree lucerne, Acacia and Moringavermicomposts and (ii) determine their potential to improve soil fertility. It was hypothesised thatvermicomposting could increases the nutrient content of the compostable feedstock of three agroforestrytree pruning residues and could potentially be used for soil fertility improvements.
2. Materials and Methods
2.1. Collection and Processing of Feedstock Materials
The V. karroo, which is vastly distributed in the province was collected from the University ofLimpopo farm. M. oleifera and C. tagataste were outsourced from neighbouring farms within a 5 kmdistance of the University farm. The species were all collected in summer 2015, dried and thenchipped to leaf-size pieces using a portable wood chipper (Portable Chipper Machine, HR 40, China)in preparation for vermicomposting. Only leaves and small branches were collected for use as

Sustainability 2020, 12, 9305 3 of 18
feedstock material. The earthworm species E. fetida ((Marie.) Savigny) used were obtained from theBertie van Zyl (Pty) farms in the Limpopo Province, South Africa.
2.2. Study Site and Experimental Design
The study was conducted under ambient conditions with a recommended moisture contentranging from 75–80%, with perforations on the lid and at the bottom of the vermicomposting boxes.The vermicomposting experiment was arranged as a Completely Randomised Design with threetreatments V. karroo, C. tagasaste and M. oleifera, replicated three times. Vermicomposting was carriedout in dark coloured ultraviolet resistant polyvinyl plastic boxes measuring 0.50 m × 0.40 m × 0.30 m(length ×width × depth), which provided a volume of 0.06 m3. Mature earthworms (E. fetida) wereintroduced at the recommended stocking rate of 1.6 kg earthworms m−2 into each box [20].
2.3. Sampling and Analyses of Vermicomposts
Sampling of the vermicomposts was done randomly in each box at two-week intervals.From each box, 200 g of the vermicompost were collected and then carefully turned for ensuringsample homogeneity and to preserve the existing earthworms. During sampling of the vermicompost,care was taken not to include any earthworms and where earthworms were sampled, they were countedand removed from the samples and then added to the final count. The samples collected during thevermicomposting were air-dried to constant weight and crushed into fine powder in preparationfor analyses.
The vermicompost samples were analysed for total N using a block digester (SC154 HotBlock digester, Environmental Express, UK) [21]. Soil organic carbon by was estimated by the loss onignition method [22]. Soil pH and EC were estimated using a pH meter (HI2002-01 edge, UK) and ECmeter (HI-2030 edge hybrid multi parameter, UK), respectively, in a mixture of compost to water ratioof 1:5 w/v. The UV/VIS spectrophotometer (Thermo Scientific Spectronic, Helide Gamma, NY, USA)was used to measure P after extraction from compost with a 0.5 M NaHCO3 solution at pH 8.5 [23].Potassium, Mg, Ca, Zn, Na, Fe, Mn, and Cu were extracted from 2 g of air-dried ground vermicompostusing 2 mL of Mehlich 3 extracting solution [24] and quantified with UV/VIS spectrophotometer.
The humic and fulvic acid fractions were extracted in 0.1 M NaOH at ratio of 1: 20 w/v [25].The supernatant was then divided into portions, with one half being analysed for total extractable C(TEC) fraction. The other half was acidified to pH 2 with H2SO4 and then centrifuged at 8000 rpmafter coagulation, with the supernatant being used for analysis of the fulvic acid (FA) portion. The Ccontents in the supernatants were determined using the dichromate oxidation method, with thecontent of the humic acid (HA) fraction being calculated as the difference between TEC and FA.Humification index (HI), humification ratio (HR) and polymerisation index (PI) which are indices usedfor evaluation of humification level in the vermicompost were calculated using the formula:
H1 = HA/C × 100 (1)
PI = HA/FA × 100 (2)
Phytochemicals testing of flavonoids was done following the method by Borokini and Omotayo [26],alkaloids using Dragendorff’s reagent [27], tannins following the method by Trease and Evans (1989);and saponins test using the method by Odebiyi and Sofowora [28]. Phytotoxicity tests were done atthe end of the composting cycle using lettuce and onion seeds because of their high sensitivity to toxicmaterials and rapid germination [29].
Manual earthworm counts were done at the start and final stages of the experiment. Microbiologicalanalyses of total bacteria, total fungi, total E. coli and total phosphate solubilising bacteria was doneevery two weeks since the beginning of the vermicomposting using the serial dilution and spreadplate counting [30]. For total bacteria, nutrient agar was used with a serial dilution of 105 and theplates were incubated at 37 ◦C for 24 h before counting the number of colony forming units (CFU). The

Sustainability 2020, 12, 9305 4 of 18
m-FC agar was used for total E. coli counts with a serial dilution of 103. The plates were incubated at44 ◦C for 24 h following which counts were made to determine the number of CFU. The Pikovskaya′sAgar was used for total phosphate solubilising bacteria (PSB) with a serial dilution of 104 being used.The plates were incubated for a period of 3–5 days at 30 ◦C and counts were made to determine thenumber of CFU. The Rose Bengal Chloramphenicol agar was used for the determination of total fungiwith a serial dilution of 103 being used. The plates were incubated at 30◦ C for a period of 3–5 days,following which counts were made to determine the number of CFU.
2.4. Statistical Analyses
The collected data were subjected to repeated measures analysis of variance (ANOVAR) using theStatistix software version 10.0 (Tallahassee, FL, USA). All assumptions, such as the equality of variancesand the normality of residuals were satisfied before running the ANOVA. Significant differencesbetween mean values were assessed using the Fisher’s least significant difference (LSD) test at aprobability level of 95% (p ≤ 0.05) [31]. Pearson correlations between the chemical and microbiologicalparameters were used to analyse the relationships between the analysed parameters. The PrincipalComponents Analysis was performed to create a minimum dataset for interpretation of results andonly highly weighted Principal Components (>10%) and the measured soil properties with a loadingmatrix >0.6 were retained in the dataset [32].
3. Results and Discussion
3.1. Nutrient Content of V. karroo, M. oleifera and C. tagasaste Vermicomposts
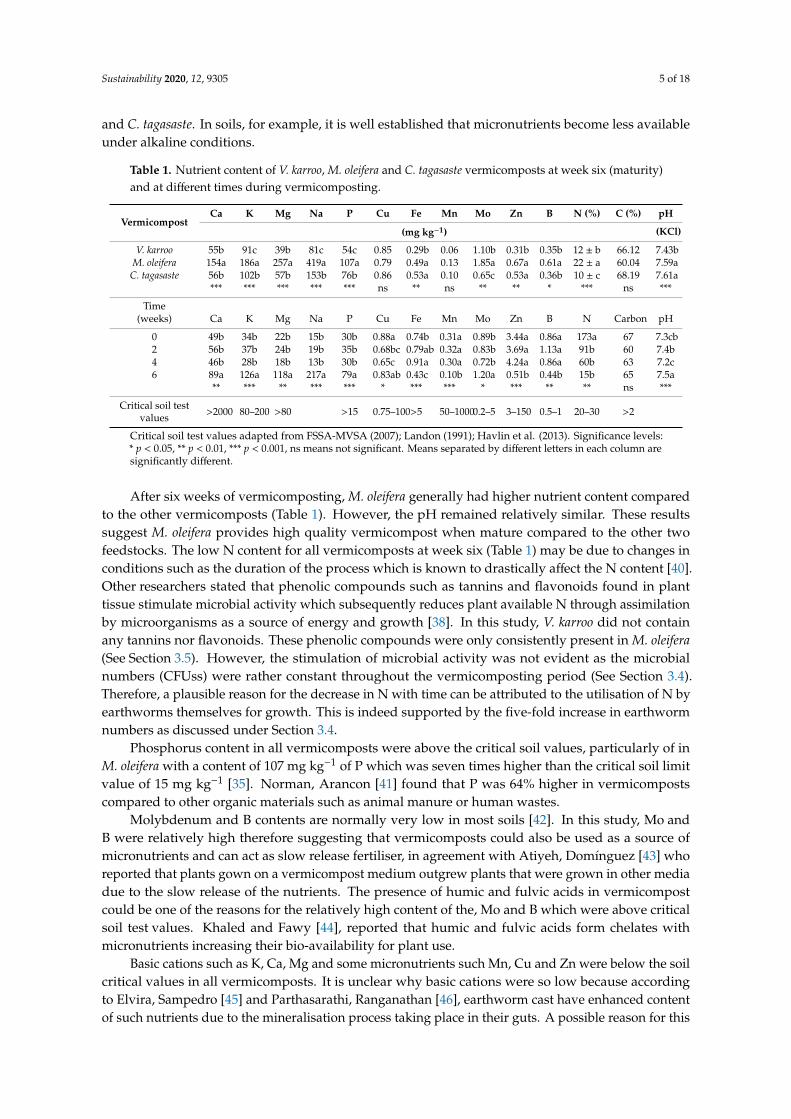
Table 1 shows the nutrient content of V. karroo, M. oleifera and C. tagasaste vermicomposts atthe final week of sampling compared with soil critical limits adopted by various researchers [33–35].The results show that nutrient contents in vermicomposts depended on the feedstock i.e., the materialbeing vermicomposted. These results were observed for all the nutrients determined except for Cu,Mn and C, which did not vary among the vermicomposts. Of the three feedstock materials, M. oleiferahad the highest content of most nutrients compared to V. karroo, and C. tagasaste. For instance, Cacontent in M. oleifera vermicompost was 154 mg kg−1, which was more than 170% higher than theother two vermicomposts. Similarly, such large differences in contents among M. oleifera and the othervermicomposts were also observed with other macronutrients (K, Mg, and P). Primary macronutrients(N, P and K) differed between the vermicomposts and followed the order M. oleifera > V. karroo > C.tagasaste for N. For P and K the trend was M. oleifera > C. tagasate > V. Karroo. Micronutrients suchas Fe and Zn were lower in C. tagasaste compared to the other two vermicomposts. The pH of thevermicomposts were slightly alkaline.
When compared over time, the macronutrients (Ca, Mg, K, P), with the exception of N, were higherat the later stage of vermicomposting during week 6. (Table 1) while no differences were observedafter zero-, two- and four-weeks. The general increase in nutrients particularly basic cations at the laterstages of vermicomposting could be a result of earthworm digestion. Several studies have also reportedhigh levels of exchangeable bases in vermicompost, agreeing with the findings of this study [36,37].Higher quantities of basic cations explain the slightly alkaline pH observed at week six.
Nitrogen content was higher at the initial vermicomposting stage and lower at the later stage,a similar trend observed also for some micronutrients such as Mn, Zn, and B. The low N content atlater stages of vermicomposting could be attributed to assimilation by microorganisms as a sourceof energy and growth [38]. The decrease in the micronutrient as the vermicompost matures arecontrary to the findings of some studies [39]. The contradiction observed in micronutrients maybe attributed to the differences in the vermicomposted material where Sharma and Garg [39] usedrice straw. The feedstocks used in this study are of relatively low C:N ratio compared to the rice straw.M. oleifera, V. karroo and C. tagasaste are all leguminous trees and hence have lower C:N ratio thanwheat straw. The observed results may partially be attributed to the slightly higher pH in M. oleifera
Sustainability 2020, 12, 9305 5 of 18
and C. tagasaste. In soils, for example, it is well established that micronutrients become less availableunder alkaline conditions.
Table 1. Nutrient content of V. karroo, M. oleifera and C. tagasaste vermicomposts at week six (maturity)and at different times during vermicomposting.
VermicompostCa K Mg Na P Cu Fe Mn Mo Zn B N (%) C (%) pH
(mg kg−1) (KCl)
V. karroo 55b 91c 39b 81c 54c 0.85 0.29b 0.06 1.10b 0.31b 0.35b 12 ± b 66.12 7.43bM. oleifera 154a 186a 257a 419a 107a 0.79 0.49a 0.13 1.85a 0.67a 0.61a 22 ± a 60.04 7.59aC. tagasaste 56b 102b 57b 153b 76b 0.86 0.53a 0.10 0.65c 0.53a 0.36b 10 ± c 68.19 7.61a
*** *** *** *** *** ns ** ns ** ** * *** ns ***
Time(weeks) Ca K Mg Na P Cu Fe Mn Mo Zn B N Carbon pH
0 49b 34b 22b 15b 30b 0.88a 0.74b 0.31a 0.89b 3.44a 0.86a 173a 67 7.3cb2 56b 37b 24b 19b 35b 0.68bc 0.79ab 0.32a 0.83b 3.69a 1.13a 91b 60 7.4b4 46b 28b 18b 13b 30b 0.65c 0.91a 0.30a 0.72b 4.24a 0.86a 60b 63 7.2c6 89a 126a 118a 217a 79a 0.83ab 0.43c 0.10b 1.20a 0.51b 0.44b 15b 65 7.5a
** *** ** *** *** * *** *** * *** ** ** ns ***
Critical soil testvalues >2000 80–200 >80 >15 0.75–100>5 50–10000.2–5 3–150 0.5–1 20–30 >2
Critical soil test values adapted from FSSA-MVSA (2007); Landon (1991); Havlin et al. (2013). Significance levels:* p < 0.05, ** p < 0.01, *** p < 0.001, ns means not significant. Means separated by different letters in each column aresignificantly different.
After six weeks of vermicomposting, M. oleifera generally had higher nutrient content comparedto the other vermicomposts (Table 1). However, the pH remained relatively similar. These resultssuggest M. oleifera provides high quality vermicompost when mature compared to the other twofeedstocks. The low N content for all vermicomposts at week six (Table 1) may be due to changes inconditions such as the duration of the process which is known to drastically affect the N content [40].Other researchers stated that phenolic compounds such as tannins and flavonoids found in planttissue stimulate microbial activity which subsequently reduces plant available N through assimilationby microorganisms as a source of energy and growth [38]. In this study, V. karroo did not containany tannins nor flavonoids. These phenolic compounds were only consistently present in M. oleifera(See Section 3.5). However, the stimulation of microbial activity was not evident as the microbialnumbers (CFUss) were rather constant throughout the vermicomposting period (See Section 3.4).Therefore, a plausible reason for the decrease in N with time can be attributed to the utilisation of N byearthworms themselves for growth. This is indeed supported by the five-fold increase in earthwormnumbers as discussed under Section 3.4.
Phosphorus content in all vermicomposts were above the critical soil values, particularly of inM. oleifera with a content of 107 mg kg−1 of P which was seven times higher than the critical soil limitvalue of 15 mg kg−1 [35]. Norman, Arancon [41] found that P was 64% higher in vermicompostscompared to other organic materials such as animal manure or human wastes.
Molybdenum and B contents are normally very low in most soils [42]. In this study, Mo andB were relatively high therefore suggesting that vermicomposts could also be used as a source ofmicronutrients and can act as slow release fertiliser, in agreement with Atiyeh, Domínguez [43] whoreported that plants gown on a vermicompost medium outgrew plants that were grown in other mediadue to the slow release of the nutrients. The presence of humic and fulvic acids in vermicompostcould be one of the reasons for the relatively high content of the, Mo and B which were above criticalsoil test values. Khaled and Fawy [44], reported that humic and fulvic acids form chelates withmicronutrients increasing their bio-availability for plant use.
Basic cations such as K, Ca, Mg and some micronutrients such Mn, Cu and Zn were below the soilcritical values in all vermicomposts. It is unclear why basic cations were so low because accordingto Elvira, Sampedro [45] and Parthasarathi, Ranganathan [46], earthworm cast have enhanced contentof such nutrients due to the mineralisation process taking place in their guts. A possible reason for this

Sustainability 2020, 12, 9305 6 of 18
could be that the earthworms are normally found in soils and they ingest soil material which generallyhave higher Ca, Mg, and K. In this study the earthworms mineralised nutrients that were in thefeedstocks. Therefore, the quantities of these nutrients should be a reflection of the amount of nutrientsin the feedstocks. However, the contents of basic cations were still rather too low than expected.A study by Mabapa, Ayisi [17] showed that M. oleifera dry biomass contained significantly higherquantities of basic cations than those found in the present study. The lower contents of micronutrientsin some of the vermicomposts could be attributed to the high pH which is known to reduce theiravailability [47].
The pH value of all the three vermicompost was found to be alkaline at 7.35. A good soilpH is essential in order to ensure the availability of nutrients because some nutrients are found tobecome unavailable at pH that is extremely alkaline or acidic [48]. The pH also controls enzymeactivity as well as biotic decomposition [49], hence the need for optimal soil pH. High pH levelsin vermicompost may result from the release of excess organic N released as ammonia that quicklydissolved in water producing ammonium and consequently increasing the pH [50]. It has alreadybeen observed that vermicompost tend to have higher N levels [51]. So, considering the leguminousnature of the feedstocks and the relatively high N content associated with them, ammonification couldhave occurred. Thus, vermicomposts have the potential to ameliorate soil acidity. However, somestudies have shown that in some instances pH may decrease during vermicomposting especiallytowards maturity [52,53].
3.2. Correlation Relationships and Principal Components Analysis
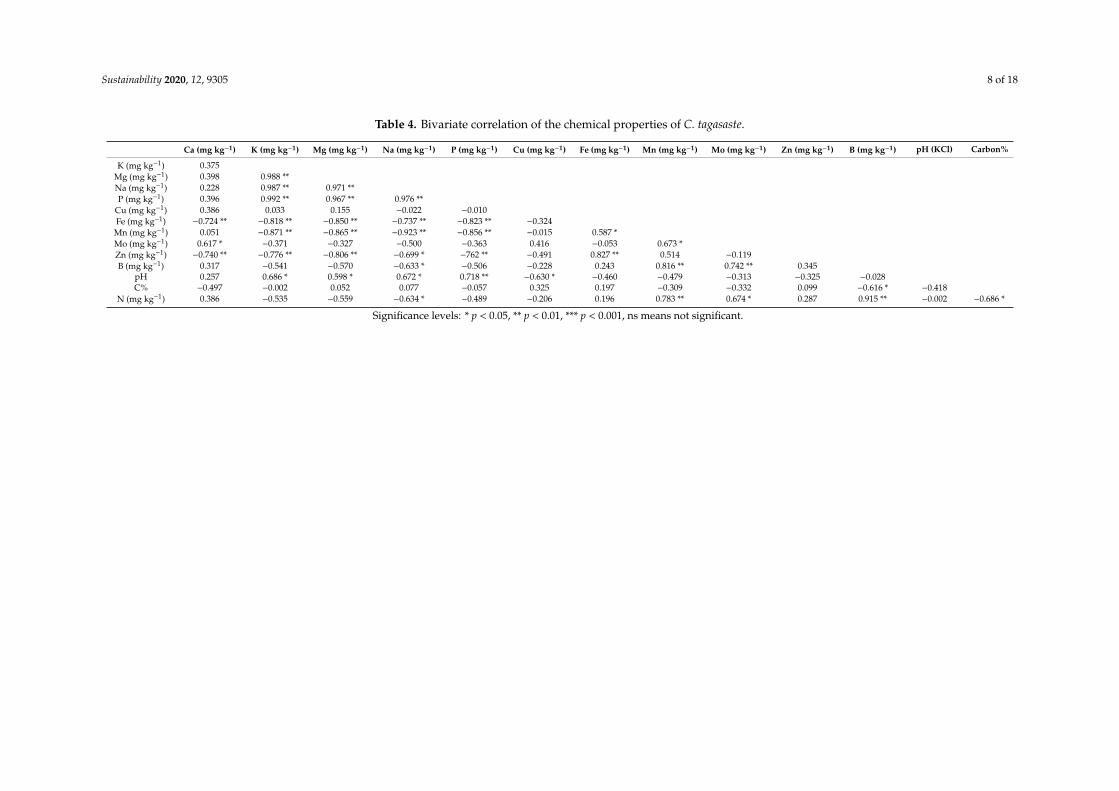
Bivariate correlation analyses for the different vermicomposts shown in Tables 2–4 for V. karroo,M. oleifera, and C. tagasaste, respectively, show that M. oleifera, and C. tagasaste had positive correlationsbetween pH and basic cations (Ca, Mg, K, and Na) and negative relationship with micronutrients(Cu, Fe, Mn, Zn, B). However, Mo was the only micronutrient positively related to the vermicompostpH. Such relationships confirm earlier discussions where pH had positive and negative relationshipswith bases and micronutrients, respectively. In contrast to the correlations shown by M. oleifera,and C. tagasaste, V. karroo showed a significant negative relationship between pH and Ca while theother basic cations had non-significant positive relationships with pH.
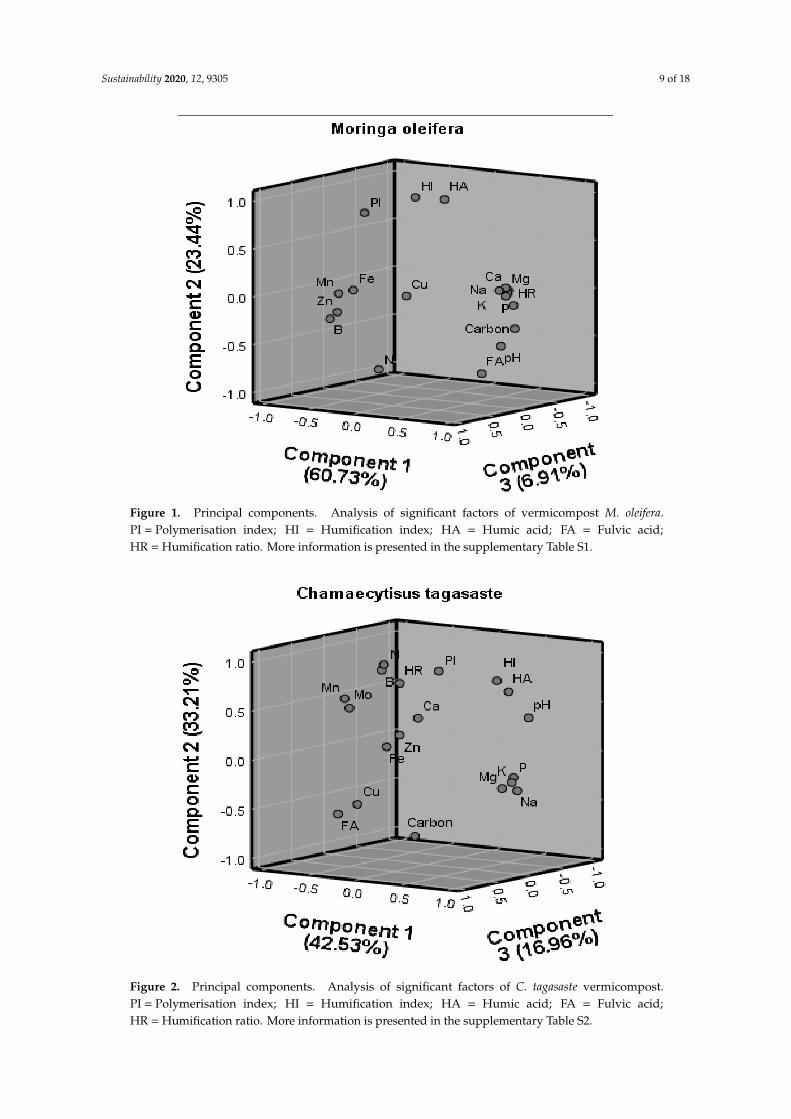
Principal components analysis for M. oleifera vermicompost showed that PC1 and PC2 exhibiteda variance of >10% and were therefore retained (Figure 1 and Table S1). The cumulative variancefor the two PCs was 84%. In the first PC, all the bases, P and HR showed a strong positive loading.In contrast, micronutrients Mn, Fe and B showed a strong negative loading. In PC2, only HI and Nloaded significantly.
Meanwhile, the principal components analysis for C. tagasaste vermicompost produced threesignificant PCs which accounted for 92% of variance (Figure 2 and Table S2). In PC1, Na, K, P, Mg,and pH exhibited significant positive loadings while micronutrients, Mn, Fe and Zn showed significantnegative loadings. Humification indices, N and HA showed positive loadings in PC2 with carbonand FA showing significant loadings. Bases, P, and HR showed a strong positive loading. ParametersCu, Ca and Mo loaded significantly in PC3. The strong loading of the above nutrients serves toconfirm their dominant occurrence in C. tagasaste. The negative loading of most micronutrients alsoconfirms the negative interactive effects between micronutrients and the PCA. In the PCA analysisnegative values of loadings of variable in the components of the PCA means the existence of an inversecorrelation between the factor PCA and the variables.

Sustainability 2020, 12, 9305 7 of 18
Table 2. Bivariate correlations of the chemical properties of V. karroo.
Ca (mg kg−1) K (mg kg−1) Mg (mg kg−1) Na (mg kg−1) P (mg kg−1) Cu (mg kg−1) Fe (mg kg−1) Mn (mg kg−1) Mo (mg kg−1) Zn (mg kg−1) B (mg kg−1) pH (KCl) Carbon%
K (mg kg−1) 0.546Mg (mg kg−1) 0.595 * 0.993 **Na (mg kg−1) 0.527 0.995 ** 0.984 **P (mg kg−1) 0.770 ** 0.931 ** 0.946 ** 0.924 **
Cu (mg kg−1) 0.474 0.876 ** 0.895 ** 0.850 ** 0.807 **Fe (mg kg−1) −0.223 −0.908 ** −0.877 ** −0.910 ** −0.782 ** −0.795 **Mn (mg kg−1) −0.531 −0.975 ** −0.958 ** −0.967 ** −0.916 ** −0.856 ** 0.914 **Mo (mg kg−1) 0.287 0.873 ** 0.862 ** 0.0871 ** 0.772 ** 0.841 ** −0.897 ** −0.848 **Zn (mg kg−1) −0.486 −0.861 ** −0.866 ** −0.880 ** −0.859 ** −710 ** 0.813 ** 0.838 ** −0.742 **B (mg kg−1) −0.239 −0.903 ** −0.866 ** −0.896 ** −0.768 ** −0.743 ** 0.954 ** 0.914 ** −0.810 ** 0.745 **
pH (KCl) −0.623 * 0.233 0.170 0.232 −0.109 0.222 −0.482 −0.224 0.351 −0.023 −0.502Carbon% −0.947 ** −0.446 −0.489 −0.428 −0.674 * −0.366 0.130 0.416 −0.189 0.299 0.179 0.587 *
N (mg kg−1) −0.848 ** −0.876 ** −0.895 ** −0.856 ** −0.952 ** −0.801 ** 0.643 * 0.874 ** −0.695 * 0.723 ** 0.665 * 0.197 0.761 **
Significance levels: * p < 0.05, ** p < 0.01, *** p < 0.001, ns means not significant.
Table 3. Bivariate correlation of the chemical properties of M. oleifera.
Ca (mg kg−1) K (mg kg−1) Mg (mg kg−1) Na (mg kg−1) P (mg kg−1) Cu (mg kg−1) Fe (mg kg−1) Mn (mg kg−1) Mo (mg kg−1) Zn (mg kg−1) B (mg kg−1) pH (KCl) Carbon%
K (mg kg−1) 0.996 **Mg (mg kg−1) 0.978 ** 0.990 **Na (mg kg−1) 0.997 ** 0.999 ** 0.989 **P (mg kg−1) 0.990 ** 0.994 ** 0.981 ** 0.994 **
Cu (mg kg−1) 0.365 0.363 0.391 0.352 0.329Fe (mg kg−1) 0.896 ** −0.903 ** −0.892 ** −0.901 ** −0.923 ** −0.154Mn (mg kg−1) −0.949 ** −0.939 ** −0.897 ** −0.942 ** −0.943 ** −0.21 0.825 **Mo (mg kg−1) 0.994 ** 0.990 ** 0.973 ** 0.986 ** 0.980 ** 0.437 −0.890 ** −0.924 **Zn (mg kg−1) −0.959 ** −0.957 ** −0.956 ** −0.962 ** −0.941 ** −0.264 0.889 ** 0.877 ** −0.945 **B (mg kg−1) −0.749 ** −0.756 ** −0.784 ** −0.774 ** −0.733 ** −0.116 0.553 0.727 ** −0.698 * 0.812 **
pH 0.781 ** 0.821 ** 0.806 ** 0.800 ** 0.821 ** −0.274 −0.744 ** −0.755 ** 0.787 ** −0.690 * −0.497Carbon% 0.579 * 0.621 * 0.617 * 0.616 * 0.606 * −0.045 −0.537 −0.569 0.539 −0.578 * −0.567 0.803 **
N (mg kg−1) −0.536 −0.502 −0.532 −0.534 −0.475 −0.238 0.402 0.449 −0.500 0.659 * 0.675 * 0.012 −0.130
Significance levels: * p < 0.05, ** p < 0.01, *** p < 0.001, ns means not significant.
Sustainability 2020, 12, 9305 8 of 18
Table 4. Bivariate correlation of the chemical properties of C. tagasaste.
Ca (mg kg−1) K (mg kg−1) Mg (mg kg−1) Na (mg kg−1) P (mg kg−1) Cu (mg kg−1) Fe (mg kg−1) Mn (mg kg−1) Mo (mg kg−1) Zn (mg kg−1) B (mg kg−1) pH (KCl) Carbon%
K (mg kg−1) 0.375Mg (mg kg−1) 0.398 0.988 **Na (mg kg−1) 0.228 0.987 ** 0.971 **P (mg kg−1) 0.396 0.992 ** 0.967 ** 0.976 **
Cu (mg kg−1) 0.386 0.033 0.155 −0.022 −0.010Fe (mg kg−1) −0.724 ** −0.818 ** −0.850 ** −0.737 ** −0.823 ** −0.324Mn (mg kg−1) 0.051 −0.871 ** −0.865 ** −0.923 ** −0.856 ** −0.015 0.587 *Mo (mg kg−1) 0.617 * −0.371 −0.327 −0.500 −0.363 0.416 −0.053 0.673 *Zn (mg kg−1) −0.740 ** −0.776 ** −0.806 ** −0.699 * −762 ** −0.491 0.827 ** 0.514 −0.119B (mg kg−1) 0.317 −0.541 −0.570 −0.633 * −0.506 −0.228 0.243 0.816 ** 0.742 ** 0.345
pH 0.257 0.686 * 0.598 * 0.672 * 0.718 ** −0.630 * −0.460 −0.479 −0.313 −0.325 −0.028C% −0.497 −0.002 0.052 0.077 −0.057 0.325 0.197 −0.309 −0.332 0.099 −0.616 * −0.418
N (mg kg−1) 0.386 −0.535 −0.559 −0.634 * −0.489 −0.206 0.196 0.783 ** 0.674 * 0.287 0.915 ** −0.002 −0.686 *
Significance levels: * p < 0.05, ** p < 0.01, *** p < 0.001, ns means not significant.
Sustainability 2020, 12, 9305 9 of 18
Figure 1. Principal components. Analysis of significant factors of vermicompost M. oleifera.PI = Polymerisation index; HI = Humification index; HA = Humic acid; FA = Fulvic acid;HR = Humification ratio. More information is presented in the supplementary Table S1.
Figure 2. Principal components. Analysis of significant factors of C. tagasaste vermicompost.PI = Polymerisation index; HI = Humification index; HA = Humic acid; FA = Fulvic acid;HR = Humification ratio. More information is presented in the supplementary Table S2.
Sustainability 2020, 12, 9305 10 of 18
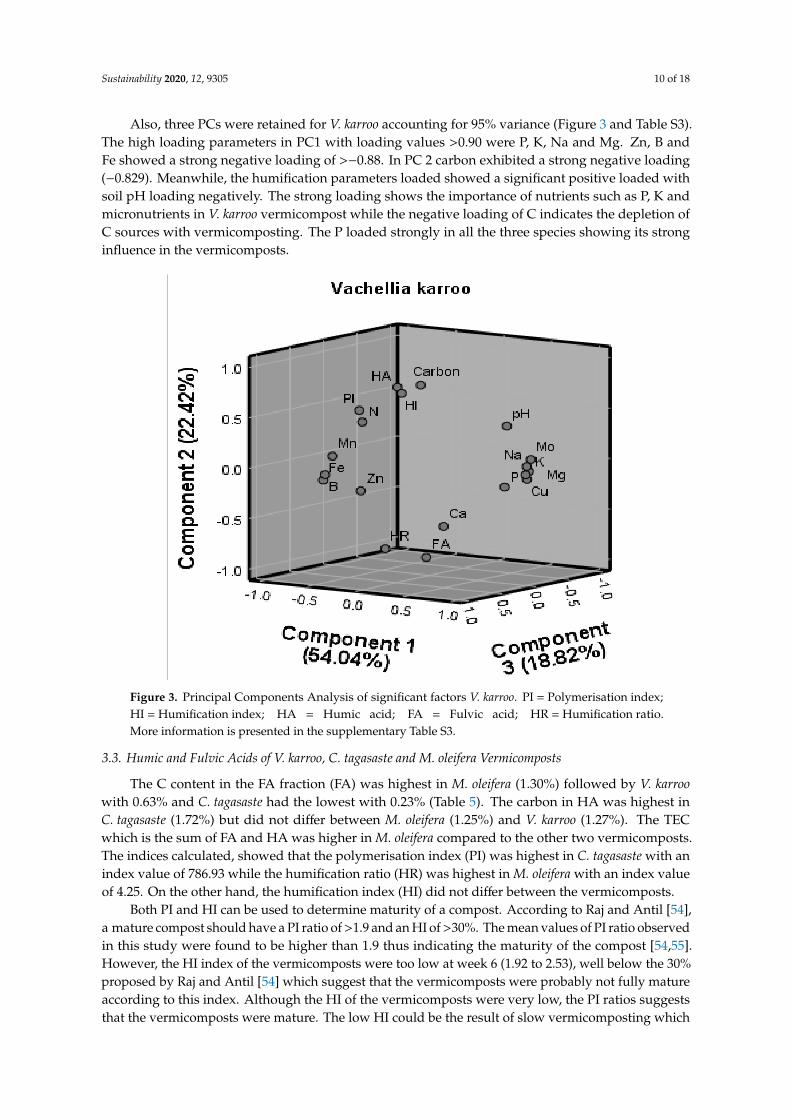
Also, three PCs were retained for V. karroo accounting for 95% variance (Figure 3 and Table S3).The high loading parameters in PC1 with loading values >0.90 were P, K, Na and Mg. Zn, B andFe showed a strong negative loading of >−0.88. In PC 2 carbon exhibited a strong negative loading(−0.829). Meanwhile, the humification parameters loaded showed a significant positive loaded withsoil pH loading negatively. The strong loading shows the importance of nutrients such as P, K andmicronutrients in V. karroo vermicompost while the negative loading of C indicates the depletion ofC sources with vermicomposting. The P loaded strongly in all the three species showing its stronginfluence in the vermicomposts.
Figure 3. Principal Components Analysis of significant factors V. karroo. PI = Polymerisation index;HI = Humification index; HA = Humic acid; FA = Fulvic acid; HR = Humification ratio.More information is presented in the supplementary Table S3.
3.3. Humic and Fulvic Acids of V. karroo, C. tagasaste and M. oleifera Vermicomposts
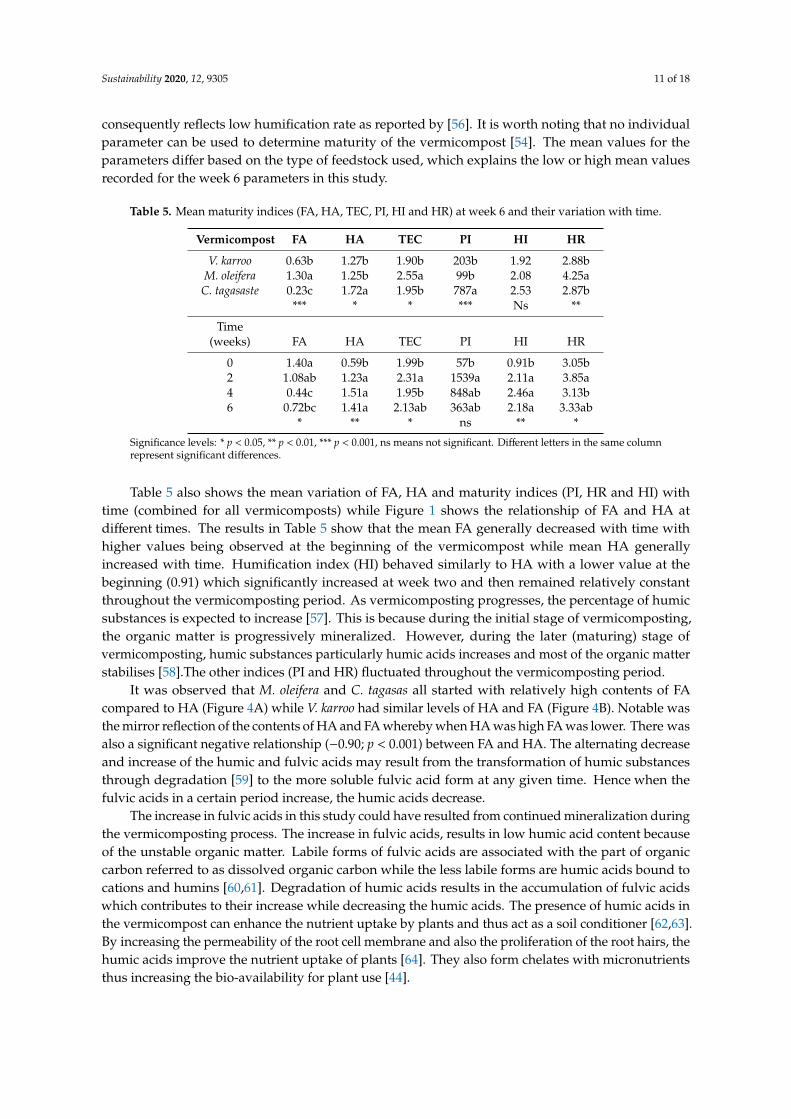
The C content in the FA fraction (FA) was highest in M. oleifera (1.30%) followed by V. karroowith 0.63% and C. tagasaste had the lowest with 0.23% (Table 5). The carbon in HA was highest inC. tagasaste (1.72%) but did not differ between M. oleifera (1.25%) and V. karroo (1.27%). The TECwhich is the sum of FA and HA was higher in M. oleifera compared to the other two vermicomposts.The indices calculated, showed that the polymerisation index (PI) was highest in C. tagasaste with anindex value of 786.93 while the humification ratio (HR) was highest in M. oleifera with an index valueof 4.25. On the other hand, the humification index (HI) did not differ between the vermicomposts.
Both PI and HI can be used to determine maturity of a compost. According to Raj and Antil [54],a mature compost should have a PI ratio of >1.9 and an HI of >30%. The mean values of PI ratio observedin this study were found to be higher than 1.9 thus indicating the maturity of the compost [54,55].However, the HI index of the vermicomposts were too low at week 6 (1.92 to 2.53), well below the 30%proposed by Raj and Antil [54] which suggest that the vermicomposts were probably not fully matureaccording to this index. Although the HI of the vermicomposts were very low, the PI ratios suggeststhat the vermicomposts were mature. The low HI could be the result of slow vermicomposting which
Sustainability 2020, 12, 9305 11 of 18
consequently reflects low humification rate as reported by [56]. It is worth noting that no individualparameter can be used to determine maturity of the vermicompost [54]. The mean values for theparameters differ based on the type of feedstock used, which explains the low or high mean valuesrecorded for the week 6 parameters in this study.
Table 5. Mean maturity indices (FA, HA, TEC, PI, HI and HR) at week 6 and their variation with time.
Vermicompost FA HA TEC PI HI HR
V. karroo 0.63b 1.27b 1.90b 203b 1.92 2.88bM. oleifera 1.30a 1.25b 2.55a 99b 2.08 4.25aC. tagasaste 0.23c 1.72a 1.95b 787a 2.53 2.87b
*** * * *** Ns **
Time(weeks) FA HA TEC PI HI HR
0 1.40a 0.59b 1.99b 57b 0.91b 3.05b2 1.08ab 1.23a 2.31a 1539a 2.11a 3.85a4 0.44c 1.51a 1.95b 848ab 2.46a 3.13b6 0.72bc 1.41a 2.13ab 363ab 2.18a 3.33ab
* ** * ns ** *
Significance levels: * p < 0.05, ** p < 0.01, *** p < 0.001, ns means not significant. Different letters in the same columnrepresent significant differences.
Table 5 also shows the mean variation of FA, HA and maturity indices (PI, HR and HI) withtime (combined for all vermicomposts) while Figure 1 shows the relationship of FA and HA atdifferent times. The results in Table 5 show that the mean FA generally decreased with time withhigher values being observed at the beginning of the vermicompost while mean HA generallyincreased with time. Humification index (HI) behaved similarly to HA with a lower value at thebeginning (0.91) which significantly increased at week two and then remained relatively constantthroughout the vermicomposting period. As vermicomposting progresses, the percentage of humicsubstances is expected to increase [57]. This is because during the initial stage of vermicomposting,the organic matter is progressively mineralized. However, during the later (maturing) stage ofvermicomposting, humic substances particularly humic acids increases and most of the organic matterstabilises [58].The other indices (PI and HR) fluctuated throughout the vermicomposting period.
It was observed that M. oleifera and C. tagasas all started with relatively high contents of FAcompared to HA (Figure 4A) while V. karroo had similar levels of HA and FA (Figure 4B). Notable wasthe mirror reflection of the contents of HA and FA whereby when HA was high FA was lower. There wasalso a significant negative relationship (−0.90; p < 0.001) between FA and HA. The alternating decreaseand increase of the humic and fulvic acids may result from the transformation of humic substancesthrough degradation [59] to the more soluble fulvic acid form at any given time. Hence when thefulvic acids in a certain period increase, the humic acids decrease.
The increase in fulvic acids in this study could have resulted from continued mineralization duringthe vermicomposting process. The increase in fulvic acids, results in low humic acid content becauseof the unstable organic matter. Labile forms of fulvic acids are associated with the part of organiccarbon referred to as dissolved organic carbon while the less labile forms are humic acids bound tocations and humins [60,61]. Degradation of humic acids results in the accumulation of fulvic acidswhich contributes to their increase while decreasing the humic acids. The presence of humic acids inthe vermicompost can enhance the nutrient uptake by plants and thus act as a soil conditioner [62,63].By increasing the permeability of the root cell membrane and also the proliferation of the root hairs, thehumic acids improve the nutrient uptake of plants [64]. They also form chelates with micronutrientsthus increasing the bio-availability for plant use [44].

Sustainability 2020, 12, 9305 12 of 18
Figure 4. Fulvic and humic acid contents in percentage of M. oleifera (A), V. karroo (B) and C. tagasaste(C) vermicomposts (p < 0.05).
3.4. Microbial and Earthworm Populations in Vermicomposts
The bacterial populations were found to be lower in V. karroo compared to other two vermicompost(Table 6). V. karroo had 9.00 colony-forming units (cfu) compared to 9.34 in both M. oleifera andC. tagasaste. Phosphate Solubilising Bacteria differed between all vermicompost at week six. Phosphatesolubilising bacteria followed the order M. oleifera > C. tagasaste > V. karroo. There were no E coli inV. karroo and C. tagasaste at week six. Only M. oleifera had E coliat week six with a cfu of 5.33. Fungion the hand was higher in M. oleifera (5.75 cfu) compared to V. karroo and C. tagasaste, which bothhad 5.49 CFU. Vermicompost are known to enhance soil biodiversity by promoting the beneficialmicrobes which in turn enhances plant growth directly by production of plant growth-regulatinghormones and enzymes and indirectly by controlling plant pathogens, nematodes and other pests,thereby enhancing plant health and minimizing the yield loss. In addition, earthworms also enhancethe beneficial microflora and suppress harmful pathogenic microbes [65]. However, the existence ofE. coli in M. oleifera was unexpected because the type of earthworms used in this study (E. fetida) areknown to completely destroy E. coli in their guts [65,66]. Therefore, the non-existence of any E. coli inV. karroo and C. tagasaste is in agreement with findings of Pathma and Sakthivel [65] and Emperor andKumar [66].
Table 6. Influence of six weeks of vermicomposting and time on bacteria, phosphate solubilisingbacteria, E. coli and fungi populations.
Bacteria Phosphate Solubilising Bacteria E. coli Fungi
Vermicompost colony-forming unit (cfu/mL)
V. karroo 9.0b 6.4c 0.0b 5.5M. oleifera 9.3a 7.1a 5.3a 5.8aC. tagasaste 9.3a 6.7b 0.0b 5.5b
* *** *** ***
Time in weeks
0 9.2a 7.0a 4.5 5.72 8.9b 6.8b 4.0 5.54 8.9b 7.0ab 4.2 5.66 9.2a 6.8b 1.8 5.6
* * ns ns
Significance levels: * p < 0.05, ** p < 0.01, *** p < 0.001, ns means not significant. Different letters in the same columnrepresent significant differences.
Bacterial population were higher at the beginning, decreased at week two and four beforeincreasing again at week six (Table 6) while PSB fluctuated with time. The mean cfu of E. coli and fungidid not vary with time.
Sustainability 2020, 12, 9305 13 of 18
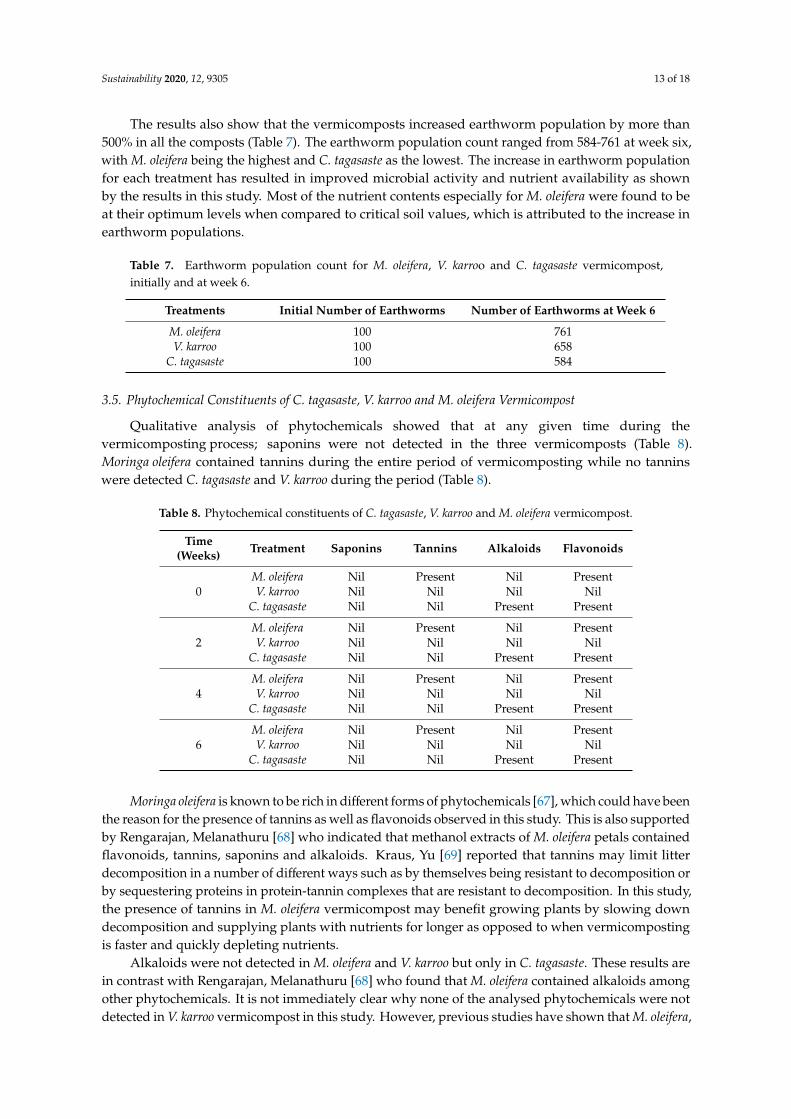
The results also show that the vermicomposts increased earthworm population by more than500% in all the composts (Table 7). The earthworm population count ranged from 584-761 at week six,with M. oleifera being the highest and C. tagasaste as the lowest. The increase in earthworm populationfor each treatment has resulted in improved microbial activity and nutrient availability as shownby the results in this study. Most of the nutrient contents especially for M. oleifera were found to beat their optimum levels when compared to critical soil values, which is attributed to the increase inearthworm populations.
Table 7. Earthworm population count for M. oleifera, V. karroo and C. tagasaste vermicompost,initially and at week 6.
Treatments Initial Number of Earthworms Number of Earthworms at Week 6
M. oleifera 100 761V. karroo 100 658
C. tagasaste 100 584
3.5. Phytochemical Constituents of C. tagasaste, V. karroo and M. oleifera Vermicompost
Qualitative analysis of phytochemicals showed that at any given time during thevermicomposting process; saponins were not detected in the three vermicomposts (Table 8).Moringa oleifera contained tannins during the entire period of vermicomposting while no tanninswere detected C. tagasaste and V. karroo during the period (Table 8).
Table 8. Phytochemical constituents of C. tagasaste, V. karroo and M. oleifera vermicompost.
Time(Weeks) Treatment Saponins Tannins Alkaloids Flavonoids
M. oleifera Nil Present Nil Present0 V. karroo Nil Nil Nil Nil
C. tagasaste Nil Nil Present Present
M. oleifera Nil Present Nil Present2 V. karroo Nil Nil Nil Nil
C. tagasaste Nil Nil Present Present
M. oleifera Nil Present Nil Present4 V. karroo Nil Nil Nil Nil
C. tagasaste Nil Nil Present Present
M. oleifera Nil Present Nil Present6 V. karroo Nil Nil Nil Nil
C. tagasaste Nil Nil Present Present
Moringa oleifera is known to be rich in different forms of phytochemicals [67], which could have beenthe reason for the presence of tannins as well as flavonoids observed in this study. This is also supportedby Rengarajan, Melanathuru [68] who indicated that methanol extracts of M. oleifera petals containedflavonoids, tannins, saponins and alkaloids. Kraus, Yu [69] reported that tannins may limit litterdecomposition in a number of different ways such as by themselves being resistant to decomposition orby sequestering proteins in protein-tannin complexes that are resistant to decomposition. In this study,the presence of tannins in M. oleifera vermicompost may benefit growing plants by slowing downdecomposition and supplying plants with nutrients for longer as opposed to when vermicompostingis faster and quickly depleting nutrients.
Alkaloids were not detected in M. oleifera and V. karroo but only in C. tagasaste. These results arein contrast with Rengarajan, Melanathuru [68] who found that M. oleifera contained alkaloids amongother phytochemicals. It is not immediately clear why none of the analysed phytochemicals were notdetected in V. karroo vermicompost in this study. However, previous studies have shown that M. oleifera,

Sustainability 2020, 12, 9305 14 of 18
V. karroo and C. tagasaste contain secondary metabolites [67,70,71]. The absence could be due to the factthat phytochemicals do not automatically accumulate at their site of synthesis and as a result of geneticvariability or leaf age development [72] may have been translocated to other parts of the plant thatwere not analysed. Reports also indicate that external stimuli can modulate the synthesis and changethe composition or quantities of the phytochemicals in plants [73,74]. The quantities are also affectedby environmental factors, such as soil composition, temperature, rainfall and ultraviolet radiationincidence [75,76]. Research has certified that flavonoid contents in some leaf exudates can be enhancedby ultra-violet radiation induction or by drought [77]. These findings indicate that the phytochemicalcontents of the tree species in this study may have possibly been affected by the above-mentionedfactors resulting in the absence of saponins and alkaloids.
Flavonoids were detected in M. oleifera and C. tagasaste vermicompost throughout thevermicomposting period (Table 8). These results are also in agreement with Rengarajan,Melanathuru [68] who noted that M. oleifera contains flavonoids among other phytochemicals.Legumes exude specific flavonoids that act as signalling molecules to attract N-fixing bacteria [78],which is essential in making N available in the soil for plant use. Flavonoids are produced whenplants are infected or injured [79] or when there are low nutrients [80]. This implies that their presencein the vermicompost is essential to plant health. This is supported by the fact that among others,phytochemicals are a source of carbon or energy for microorganisms which are consequently beneficialfor plant health and they also release hormone effectors of cell differentiation in plants [81]. Flavonoidsgive ultra violet protection to plant tissues, and their accumulation due to ultra-violet exposure is alsowell documented [82].
The absence of some phytochemicals such as saponins and tannins in some tree species indicatesthat they are naturally unavailable [83], or present in undetectable amounts. This supports the resultsobtained in this study, since the phytochemicals were not detected in some of the vermicomposttreatments. The presence of phytochemicals in the vermicompost indicate possible defence againstharmful bacteria and pests [29]. Therefore, this shows that the vermicomposts have potential to be usedfor soil fertility enhancement. Seed germination (phytotoxicity) was also above 70%, which indicatesthat the compost could not have been toxic to plants [29].
4. Conclusions
This Vermicomposting of the agroforestry tree species V. karroo, M. oleifera and C. tagasaste hasproven to be very beneficial given the chemical and biological characteristics observed in this study.The chemical composition of the vermicomposts were characterised by a significantly high nutrientssuch as P, N, K, Mg and Ca that are essential for plant growth and soil fertility enhancement.Although the nutrient content of each vermicompost varied during the vermicomposting period,the quantities of the nutrients found in the vermicomposts indicated that they have potential to be usedas for soil fertility improvements and thus reduce the use of synthetic fertilisers in crop production.
Further studies need to evaluate the potential of the vermicomposts as mediums for plant growthand its effect on soil properties and plant growth when applied as soil amendment.
Supplementary Materials: The following are available online at http://www.mdpi.com/2071-1050/12/22/9305/s1.
Author Contributions: Conceptualization, K.K.A. and A.M.; methodology, A.M.; formal analysis, A.M. and L.M.;investigation, M.M.M.; resources, A.M. and K.K.A.; data curation, M.M.M., A.M. and L.M.; writing—originaldraft preparation, M.M.M.; writing—review and editing, L.M. and A.M.; visualization, L.M.; supervision, A.M.and K.K.A. All authors have read and agreed to the published version of the manuscript.
Funding: This research received no external funding.
Acknowledgments: The authors would like to acknowledge the Risk and Vulnerability Science Centre,University of Limpopo for funding this research.
Conflicts of Interest: The authors declare no conflict of interest.

Sustainability 2020, 12, 9305 15 of 18
References
1. Alao, J.; Shuaibu, R. Agroforestry practices and concepts in sustainable land use systems in Nigeria.J. Hortic. For. 2013, 5, 156–159.
2. Barnes, R.; Fagg, C.W. Faidherbia Albida: Monograph and Annotated Bibliography; Oxford Forestry Institute:Brighton, UK, 2003.
3. Chirwa, P.W.; Ong, C.K.; Maghembe, J.; Black, C.R. Soil water dynamics in cropping systems containingGliricidia sepium, pigeonpea and maize in southern Malawi. Agrofor. Syst. 2007, 69, 29–43. [CrossRef]
4. Makumba, W.; Janssen, B.; Oenema, O.; Akinnifesi, F.; Mweta, D.; Kwesiga, F. The long-term effects of agliricidia–maize intercropping system in Southern Malawi, on gliricidia and maize yields, and soil properties.Agric. Ecosyst. Environ. 2006, 116, 85–92. [CrossRef]
5. Sène, E.H. Forests and food security in Africa: The place of forestry in FAO’s Special Programme for FoodSecurity. Unasylva 2000, 51, 13–18.
6. Ayisi, K.K. Building a Large Evergreen Agriculture Network for Southern Africa (BLEANSA): Report on LimpopoProvince of South Africa; Risk and Vulnerability Science Centre: Eastern Cape, South Africa, 2014.
7. Emmanuel, Y. The Potential of Tagasaste (Chamaecytisus Palmensis) in Maintaining Fertility in a SustainableCropping System. Master’s Thesis, Lincoln University, Lincoln, New Zealand, 1999.
8. O’Donoghue, B. Tastee Tagasaste. Available online: http://www.victoriaparkalpacastud.com.au/Tastee_Tagasastee.pdf (accessed on 5 December 2019).
9. Mbah, B.; Eme, P.; Mbah, B.; Paul, A. Effect of Drying Techniques on the Proximate and Other NutrientComposition of Moringa oleifera Leaves from Two Areas in Eastern Nigeria. Pak. J. Nutr. 2012, 11, 1044–1048.[CrossRef]
10. Mupondi, L.T. Improving Sanitization and Fertiliser Value of Dairy Manure and Waste Paper MixturesEnriched with Rock Phosphate through Combined Thermophilic Composting and Vermicomposting.Ph.D. Thesis, University of Fort Hare, Alice, South Africa, 2010.
11. Mupambwa, H.A.; Mnkeni, P.N.S. Optimization of Fly Ash Incorporation into Cow Dung-Waste PaperMixtures for Enhanced Vermidegradation and Nutrient Release. J. Environ. Qual. 2015, 44, 972–981.[CrossRef]
12. Vermicomposting: Recycling Wastes into Valuable Organic Fertilizer. Available online: http://oar.icrisat.org/
3677/ (accessed on 15 October 2019).13. Dingaan, M.; Du Preez, P.J. Vachellia (Acacia) karroo Communities in South Africa: An Overview. In Pure
and Applied Biogeography; IntechOpen: London, UK, 2017.14. William, J.A.; Iddrisu, L.N.; Damian, T.-D.; Kwame, O.-B.; Kwami, B.K.; Asante, W.J.; Nasare, I.L.;
Tom-Dery, D.; Ochire-Boadu, K.; Kentil, K.B. Nutrient composition of Moringa oleifera leaves from two agroecological zones in Ghana. Afr. J. Plant Sci. 2014, 8, 65–71. [CrossRef]
15. Mendieta-Araica, B.; Spörndly, E.; Reyes-Sánchez, N.; Salmerón-Miranda, F.; Halling, M. Biomass productionand chemical composition of Moringa oleifera under different planting densities and levels of nitrogenfertilization. Agrofor. Syst. 2013, 87, 81–92. [CrossRef]
16. Anwar, F.; Latif, S.; Ashraf, M.; Gilani, A.H. Moringa oleifera: A food plant with multiple medicinal uses.Phytother. Res. 2007, 21, 17–25. [CrossRef]
17. Mabapa, M.P.; Ayisi, K.K.; Mariga, I.K. Effect of Planting Density and Harvest Interval on the Leaf Yield andQuality of Moringa (Moringa oleifera) under Diverse Agroecological Conditions of Northern South Africa.Int. J. Agron. 2017, 2017, 1–9. [CrossRef]
18. Moyo, B.; Oyedemi, S.; Masika, P.J.; Muchenje, V. Polyphenolic content and antioxidant properties ofMoringa oleifera leaf extracts and enzymatic activity of liver from goats supplemented with Moringa oleiferaleaves/sunflower seed cake. Meat Sci. 2012, 91, 441–447. [CrossRef] [PubMed]
19. Pakade, V.E.; Cukrowska, E.; Chimuka, L. Metal and flavonol contents of Moringa oleifera grown in SouthAfrica. S. Afr. J. Sci. 2013, 109, 1–7. [CrossRef]
20. Ndegwa, P.; Thompson, S.; Das, K. Effects of stocking density and feeding rate on vermicomposting ofbiosolids. Bioresour. Technol. 2000, 71, 5–12. [CrossRef]
21. Okalebo, J.R.; Gathua, K.W.; Woomer, P.L. Laboratory Methods of Soil and Plant Analysis: A Working Manual,2nd ed.; Sacred Africa: Nairobi, Kenya, 2002.

Sustainability 2020, 12, 9305 16 of 18
22. Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis: Part3 Chemical methods; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA,1996; Volume 5, pp. 961–1010.
23. Olsen, S.; Sommers, L.; Page, A. Methods of soil analysis. Part 2 Chemical and Microbiological Properties.Am. Soc. Agron. Soil Sci. Soc. Am. 1982, 2, 403–430.
24. Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci.Plant. Anal. 1984, 15, 1409–1416. [CrossRef]
25. Sánchez-Monedero, M.A.; Roig, A.; Martínez-Pardo, C.; Cegarra, J.; Paredes, C. A microanalysis methodfor determining total organic carbon in extracts of humic substances. Relationships between total organiccarbon and oxidable carbon. Bioresour. Technol. 1996, 57, 291–295. [CrossRef]
26. Borokini, T.I. Phytochemical and ethnobotanical study of some selected medicinal plants from Nigeria.J. Med. Plants Res. 2012, 6, 1106–1118. [CrossRef]
27. Harborne, A. Phytochemical Methods: A Guide to Modern Techniques of Plant. Analysis; Springer Science &Business Media: Berlin, Germany, 1998.
28. Odebiyi, O.O.; Sofowora, E.A. Phytochemical screening of Nigerian medicinal plants II. Lloydia 1978, 41, 234.29. Brielmann, H.L.; Setzer, W.N.; Kaufman, P.B.; Kirakosyan, A.; Cseke, L.J. Phytochemicals: The Chemical
Components of Plants. In Natural Products from Plants, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2006.30. Germida, J.; De Freitas, J. Cultural Methods for Soil and Root-Associated Microorganisms. Soil Sampl. Meth.
Anal. Sec. Ed. 2007, 341–353. [CrossRef]31. Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons:
Hoboken, NJ, USA, 1984.32. Manyevere, A.; Chikuvire, T.J.; Mnkeni, P.N.; Dhau, I. Spatial variability of selected soil micronutrients under
smallholder crop production in Zanyokwe, Eastern Cape, South Africa. S. Afr. J. Plant. Soil 2017, 34, 339–349.[CrossRef]
33. Halvin, J.L.; Tisdale, S.L.; Nelson, W.L.; Beaton, J.D. (Eds.) Soil Fertility and Fertilizers: An Introduction toNutrient Management, 8th ed.; Pearson Education Inc.: Upper Saddle River, NJ, USA, 2013.
34. Landon, J. Booker Tropical Soil Manual: A Handbook for Soil Survey and Agricultural Land Evaluation in the Tropicsand Subtropics; John Wiley & Sons Inc.: New York, NY, USA, 1991.
35. FSSA. Fertilizer Handbook, 6th ed.; Lynwood Ridge, Fertilizer Society of South Africa: Pretoria, South Africa, 2007.36. Oo, A.N.; Iwai, C.B.; Saenjan, P. Soil Properties and Maize Growth in Saline and Nonsaline Soils using
Cassava-Industrial Waste Compost and Vermicompost with or Without Earthworms. Land Degrad. Dev.2015, 26, 300–310. [CrossRef]
37. Nada, W.M.; Van Rensburg, L.; Claassens, S.; Blumenstein, O. Effect of Vermicompost on Soil and PlantProperties of Coal Spoil in the Lusatian Region (Eastern Germany). Commun. Soil Sci. Plant. Anal. 2011, 42,1945–1957. [CrossRef]
38. Lin, Y.M.; Liu, J.W.; Xiang, P.; Lin, P.; Gongfu, Y.; Da Sternberg, L.S.L. Tannin Dynamics of Propagulesand Leaves of Kandelia candel and Bruguiera gymnorrhiza in the Jiulong River Estuary, Fujian, China.Biogeochemistry 2006, 78, 343–359. [CrossRef]
39. Sharma, K.; Garg, V. Comparative analysis of vermicompost quality produced from rice straw and paperwaste employing earthworm Eisenia fetida (Sav.). Bioresour. Technol. 2018, 250, 708–715. [CrossRef]
40. Masunga, R.H.; Uzokwe, V.N.; Mlay, P.D.; Odeh, I.; Singh, A.; Buchan, D.; De Neve, S. Nitrogen mineralizationdynamics of different valuable organic amendments commonly used in agriculture. Appl. Soil Ecol. 2016,101, 185–193. [CrossRef]
41. Norman, Q.; Arancon, A.; Edwards Clive, A. Effects of vermicomposts on plant growth. Soil EcologyLaboratory, The Ohio State University, Columbus. In Proceedings of the International Symposium Workshopon Vermi Technologies for Developing Countries, Los Banos, Philippines, 16–18 November 2005.
42. Horneck, D.A.; Sullivan, D.M.; Owen, J.; Hart, J.M. Soil Test Interpretation Guide; Oregon State University:Corvallis, OR, USA, 2011.
43. Atiyeh, R.M.; Domínguez, J.; Subler, S.; Edwards, C.A. Changes in biochemical properties of cow manureduring processing by earthworms (Eisenia andrei, Bouché) and the effects on seedling growth. Pedobiologia2000, 44, 709–724. [CrossRef]
44. Khaled, H.; Fawy, H. Effect of different levels of humic acids on the nutrient content, plant growth, and soilproperties under conditions of salinity. Soil Water Res. 2011, 6, 21–29. [CrossRef]

Sustainability 2020, 12, 9305 17 of 18
45. Elvira, C.; Sampedro, L.; Benítez, E.; Nogales, R. Vermicomposting of sludges from paper mill and dairyindustries with Eisenia andrei: A pilot-scale study. Bioresour. Technol. 1998, 63, 205–211. [CrossRef]
46. Parthasarathi, K.; Ranganathan, L.S.; Anandi, V.; Zeyer, J. Diversity of microflora in the gut and casts oftropical composting earthworms reared on different substrates. J. Environ. Biol. 2007, 28, 87–97.
47. Sharma, R.; Singh, M.; Sharma, J. Correlation studies on micronutrients vis-à-vis soil properties in some soilsof Nagaur district in semi-arid region of Rajasthan. J. Indian Soc. Soil Sci. 2003, 51, 522–527.
48. Chaudhari, P.; Ahire, D.; Ahire, V.D. Correlation between Physico-chemical properties and available nutrientsin sandy loam soils of Haridwar. J. Chem. Biol. Phys. Sci. 2012, 2, 1493.
49. Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.;Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008,11, 1252–1264. [CrossRef] [PubMed]
50. Rynk, R.; van de Kamp, M.; Willson, G.B.; Singley, M.E.; Richard, T.L.; Kolega, J.J.; Gouin, F.R.; Laliberty, L.;Kay, D.; Murphy, D.W.; et al. On-Farm Composting Handbook (NRAES 54); Northeast Regional AgriculturalEngineering Service (NRAES): New York, NY, USA, 1992.
51. Musaida, M.; Manyuchi, M.; Chitambwe, T.; Phiri, A.; Muredzi, P.; Kanhukamwe, Q. Effect of vermicompost,vermiwash and application time on soil physicochemical properties. Int. J. Chem. Environ. Eng. 2013, 4,216–220.
52. Mitchell, A. Production of Eisenia fetida and vermicompost from feed-lot cattle manure. Soil Biol. Biochem.1997, 29, 763–766. [CrossRef]
53. Ndegwa, P.M.; Thompson, S. Effects of C-to-N ratio on vermicomposting of biosolids. Bioresour. Technol.2000, 75, 7–12. [CrossRef]
54. Raj, D.; Antil, R. Evaluation of maturity and stability parameters of composts prepared from agro-industrialwastes. Bioresour. Technol. 2011, 102, 2868–2873. [CrossRef]
55. Jiménez, E.I.; García, V.P. Determination of maturity indices for city refuse composts. Agric. Ecosyst. Environ.1992, 38, 331–343. [CrossRef]
56. Unuofin, F.; Siswana, M.; Cishe, E.N. Enhancing rock phosphate integration rate for fast bio-transformationof cow-dung waste-paper mixtures to organic fertilizer. SpringerPlus 2016, 5, 1–17. [CrossRef]
57. Wu, L.; Ma, L.Q. Relationship between Compost Stability and Extractable Organic Carbon. J. Environ. Qual.2002, 31, 1323–1328. [CrossRef]
58. Veeken, A.; Nierop, K.; De Wilde, V.; Hamelers, H.V. Characterisation of NaOH-extracted humic acids duringcomposting of a biowaste. Bioresour. Technol. 2000, 72, 33–41. [CrossRef]
59. Sutton, R.; Sposito, G. Molecular Structure in Soil Humic Substances: The New View. Environ. Sci. Technol.2005, 39, 9009–9015. [CrossRef] [PubMed]
60. Andriulo, A.; Galantini, J.; Pecorari, C.; Torioni, E. Soil organic matter in the pampas region. I. A method offractionation using sieves. Agrochimica 1990, 34, 475–489.
61. Six, J.; Callewaert, P.; Lenders, S.; De Gryze, S.; Morris, S.J.; Gregorich, E.G.; Paul, E.A.; Paustian, K. Measuringand Understanding Carbon Storage in Afforested Soils by Physical Fractionation. Soil Sci. Soc. Am. J. 2002,66, 1981–1987. [CrossRef]
62. Valdrighi, M.M.; Pera, A.; Agnolucci, M.; Frassinetti, S.; Lunardi, D.; Vallini, G. Effects of compost-derivedhumic acids on vegetable biomass production and microbial growth within a plant (Cichorium intybus)-soilsystem: A comparative study. Agric. Ecosyst. Environ. 1996, 58, 133–144. [CrossRef]
63. Atiyeh, R.; Lee, S.; Edwards, C.; Arancon, N.; Metzger, J. The influence of humic acids derived fromearthworm-processed organic wastes on plant growth. Bioresour. Technol. 2002, 84, 7–14. [CrossRef]
64. Pramanik, P.; Ghosh, G.; Ghosal, P.; Banik, P. Changes in organic—C, N, P and K and enzyme activities invermicompost of biodegradable organic wastes under liming and microbial inoculants. Bioresour. Technol.2007, 98, 2485–2494. [CrossRef]
65. Pathma, J.; Sakthivel, N. Microbial diversity of vermicompost bacteria that exhibit useful agricultural traitsand waste management potential. SpringerPlus 2012, 1, 1–19. [CrossRef]
66. Emperor, G.; Kumar, K. Microbial population and activity on vermicompost of Eudrilus eugeniae and Eiseniafetida in different contents of tea waste with cow dung and kitchen waste mixture. Int. J. Curr. Microbiol.Appl. Sci. 2015, 4, 496–507.

Sustainability 2020, 12, 9305 18 of 18
67. Olabode, Z.A.I.N.A.B.; Akanbi, C.T.; Olunlade, B.; Adeola, A.A. Effects of drying temperature on the nutrientsof Moringa (Moringa oleifera) leaves and sensory attributes of dried leaves infusion. Dir. Res. J. Agric.Food Sci. 2015, 3, 117–122.
68. Rengarajan, S.; Melanathuru, V.; Munuswamy, D.; Sundaram, S.; Selvaraj, S.T. A comparative study ofin vitro antimicrobial activity and tlc studies of petals of selected indian medicinal plants. Asian J. Pharm.Clin. Res. 2016, 9, 259–263. [CrossRef]
69. Kraus, T.E.; Yu, Z.; Preston, C.M.; Dahlgren, R.A.; Zasoski, R.J. Linking chemical reactivity and proteinprecipitation to structural characteristics of foliar tannins. J. Chem. Ecol. 2003, 29, 703–730. [CrossRef]
70. Brown, D.; Ng’Ambi, J.; Norris, D. Voluntary Intake and Palatability Indices of Pedi Goats fed DifferentLevels of Acacia karroo Leaf Meal by Cafeteria Method. Indian J. Anim. Res. 2016, 50, 41–47. [CrossRef]
71. Edwards, N. A Review of Tannins and Other Secondary Metabolites in the Fodder Shrub Tagasaste (ChamaecytisusProliferus); Australian Centre for International Agricultural Research: Canberra, Australia, 2000.
72. Siddhuraju, P.; Becker, K. Antioxidant Properties of Various Solvent Extracts of Total Phenolic Constituentsfrom Three Different Agroclimatic Origins of Drumstick Tree (Moringa oleiferaLam.) Leaves. J. Agric.Food Chem. 2003, 51, 2144–2155. [CrossRef]
73. Kuc, J. Phytoalexins, Stress Metabolism, and Disease Resistance in Plants. Annu. Rev. Phytopathol. 1995, 33,275–297. [CrossRef]
74. Matsuki, M. Regulation of Plant Phenolic Synthesis: From Biochemistry to Ecology and Evolution. Aust. J. Bot.1996, 44, 613–634. [CrossRef]
75. Kouki, M.; Manetas, Y. Resource availability affects differentially the levels of gallotannins and condensedtannins in Ceratonia siliqua. Biochem. Syst. Ecol. 2002, 30, 631–639. [CrossRef]
76. Monteiro, J.M.; Albuquerque, U.P.; Neto, E.M.L.; Araújo, E.D.L.; Albuquerque, M.M.; Amorim, E.L. Theeffects of seasonal climate changes in the caatinga on tannin levels in Myracrodruon urundeuva (Engl.) Fr.All. and Anadenanthera colubrina (Vell.) Brenan. Rev. Bras. Farm. 2006, 16, 338–344. [CrossRef]
77. Chaves, N.; Escudero, J.C.; Gutiérrez-Merino, C. Role of Ecological Variables in the Seasonal Variation ofFlavonoid Content of Cistus ladanifer Exudate. J. Chem. Ecol. 1997, 23, 579–603. [CrossRef]
78. Broughton, W.J.; Hernández, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)—Modelfood legumes. Plant Soil 2003, 252, 55–128. [CrossRef]
79. Takahama, U.; Oniki, T. Flavonoids and Some Other Phenolics as Substrates of Peroxidase: PhysiologicalSignificance of the Redox Reactions. J. Plant Res. 2000, 113, 301–309. [CrossRef]
80. Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress.Polish J. Environ. Stud. 2006, 15, 523–530.
81. Demain, A.L.; Fang, A. The Natural Functions of Secondary Metabolites. Adv. Biochem. Eng. Biotechnol. 2000,69, 1–39.
82. Pavarini, D.P.; Pavarini, S.P.; Niehues, M.; Lopes, N.P. Exogenous influences on plant secondary metabolitelevels. Anim. Feed. Sci. Technol. 2012, 176, 5–16. [CrossRef]
83. Omokhua, A.G.; McGaw, L.J.; Finnie, J.F.; Van Staden, J. Chromolaena odorata (L.) R.M. King & H. Rob.(Asteraceae) in sub-Saharan Africa: A synthesis and review of its medicinal potential. J. Ethnopharmacol.2016, 183, 112–122. [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutionalaffiliations.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (http://creativecommons.org/licenses/by/4.0/).





























