Embed Size (px)
Citation preview

Characterization of macroaggregates and photodegradation of their
water soluble fraction
N. KOVAC1*, J. FAGANELI1, B. SKET2 and O. BAJT1
1Marine Biological Station, Fornace 41, 6330 Piran, Slovenia and 2Department of Organic Chemistry,University of Ljubljana, Askerceva 6, 1000 Ljubljana, Slovenia
AbstractÐThe analysis of macroaggregates of phytoplankton originating from the northern Adriaticusing 13C and 1H nuclear magnetic resonance (NMR) spectroscopy showed the presence of long poly-methylene chains, heteropolysaccharides and carboxyl groups. It is our belief that the polymethylenechains compose a part of bound carboxylic acids. Although it is possible that they are bonded in someother way, we favour this structure because it is commonly encountered in lipids. The water solublefraction of macroaggregates were photolyzed using a photochemical reactor and exposure to sunlight.The photochemical transformations of aqueous solutions of natural macroaggregates and model mono-(glucose), di- (maltose) and polysaccharides (pullulan, laminarin) were followed colorimetrically at regu-lar time intervals during light exposure using 3-methyl-2-benzothiazolinone hydrazone hydrochloride(MBTH) and also using 1H NMR spectroscopy. Concentrations of monosaccharides produced frommacroaggregates, pullulan and laminarin were increased under arti®cial light and sunlight. The photo-chemical degradation of macroaggregates in sunlight exhibits similar ®rst order degradation kinetics tothat under arti®cial light, but with a lower reaction rate. The 1H NMR studies of the irradiation pro-ducts of pullulan and laminarin revealed a decrease in the signal of polysaccharide anomeric protons,suggesting degradation of the glycoside linkage. The prolonged irradiation of macroaggregates leads todecreasing monomer concentration and also suggests the degradation of produced monomers. The ir-radiation experiments on glucose con®rmed this conclusion. Maltose, according to 1H NMR spectra,degrades to monomers which successively degrade. Photopolymerization can also occur after cleavageof glycoside linkages. These experiments showed that the northern Adriatic macroaggregates, mostlycomposed of heteropolysaccharides and polymethylene chains and originating from the cell wall, arephoto-labile and their degradation proceeds through the cleavage of glycoside linkage and the sub-sequent degradation of monosaccharides produced. This photochemical degradation is signi®cant andcompetitive with microbial degradation and could be important in reducing macroaggregate concen-trations in shallow water columns of the northern Adriatic during summertime. Moreover, the lowermolecular weight photoproducts can be subsequently used as substrates by microbes. # 1998 ElsevierScience Ltd. All rights reserved
Key wordsÐNMR, macroaggregates, heteropolysaccharides, algaenan, phototransformation
INTRODUCTION
Mucillaginous material is important in many marine
¯ux processes (Decho, 1990). Hyperproduction of
mucous macroaggregates in the northern Adriatic
occurred during the summers of 1988, 1989, 1991
and 1997 and has been the object of various eco-
logical (Malej and Harris, 1993; Decho and Herndl,
1995) and chemical (Posedel and Faganeli, 1991;
Faganeli et al., 1995; Kovac et al., 1995) studies.
These macroaggregates are found in a variety of
stages or forms: small ¯ocs, macro¯ocs, stringers,
tapes, clouds, creamy surface and gelatinous layers
(Stachowitsch et al., 1990). During the summer,
horizontal and vertical distribution and accumu-
lation of macroaggregates was heterogeneous and
time-dependent on di�erent factors such as con-
ditions of the water column, size, form and compo-
sition of aggregates (biological and chemical) andenvironmental conditions (Stachowitsch et al., 1990;Herndl, 1992; Degobbis et al., 1995; Mingazziniand Thake, 1995; Rinaldi et al., 1995).
Macroaggregates contain various entrapped phyto-plankters, microzooplankters, bacteria and detritaland mineral particles (Stachowitsch et al., 1990).
The majority of authors agree that mucilage pro-duction in the northern Adriatic is primarily associ-ated with phytoplankton, mostly with diatom
blooms (Fanuko and Turk, 1990; Herndl, 1992;Degobbis et al., 1995; Mingazzini and Thake, 1995;Myklestad, 1995). The ``new'' production occurringin late spring as a consequence of the massive river-
ine inputs of N, Si and P nutrients in the surfacelayer of the Gulf of Trieste produced high particu-late carbohydrate and protein concentrations
(Faganeli et al., 1995). Subsequent depletion ofintroduced nutrients caused a decrease of particu-late protein concentration but not of particulate
carbohydrate (Kiùrboe et al., 1990; Faganeli et al.,
Org. Geochem. Vol. 29, No. 5±7, pp. 1623±1634, 1998# 1998 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0146-6380/98 $ - see front matterPII: S0146-6380(98)00178-8
*To whom correspondence should be addressed. E-mail:[email protected]
1623

1995; Myklestad, 1995). Batch culture experiments
also showed that disappearance of ambient nitrate
slowed cell protein synthesis, but cells continued to
synthesize carbohydrate during a period of unba-
lanced growth (Richardson and Cullen, 1995).
Carbohydrates constituted a major portion of
particulate organic matter (POM) as well as dis-
solved organic matter (DOM) produced by phyto-
plankton (Biddanda and Benner, 1997). Most
mucilages are primarily composed of polysacchar-
ides and proteins (Decho and Herndl, 1995). While
polysaccharides make up the bulk of the biomass of
most mucilages, proteins usually occur in lesser
amounts and often in the form of glycoproteins
(Vollenweider et al., 1992; Decho and Herndl,
1995). Polysaccharides are also thought to comprise
the bulk of the dissolved organic matter (DOM)
pool in sea water (McCarthy et al., 1993) and to
play an important role in the formation of organic
particles in the sea (Mingazzini and Thake, 1995).
Aggregation proceeds through the formation of
more permanent chemical bonding between mol-
ecules of mucous polysaccharides which function as
a glue between planktonic cells (Azam and Smith,
1991). They might, however, also bind higher mol-
ecular weight organic compounds such as humic
acids.
The previous chemical study of macroaggregate
composition in the Gulf of Trieste (northern
Adriatic) showed that water-soluble carbohydrates
were a major component comprising 12 to 34% of
the total matter (Posedel and Faganeli, 1991) and
were composed in decreasing order, of the mono-
saccharides glucose, fructose, mannose, galactose,
fucose, arabinose, ribose and xylose (Faganeli et al.,
1995). These results suggest that the primary origin
of monosaccharides is from phytoplanktonic struc-
tural polysaccharides (Hama and Handa, 1992)
which are thought to be similar in composition to
phytoplanktonic mucous (Ittekkot et al., 1982). The
amino acid composition of macroaggregates was
similar to that of particulate organic matter (POM)
in the Gulf of Trieste of planktonic origin
(Faganeli, 1989), demonstrating that the amino
acids are mostly bonded in the proteins of
entrapped cells. Chemical and spectroscopic (FTIR,1H NMR) data (Kovac et al., 1995) suggested a
complex structure of polysaccharides composed of
di�erent monosaccharide units with free hydroxylic
groups and relatively long polymethylene chains
attached to the monosaccharide units which are free
or linked to other polysaccharide chains forming
crosslinked structures. The lack of acidic groups
was con®rmed by heavy metal analysis showing
rather low contents and indicating a weak heavy
metal bonding capacity.
Episodic and irregular appearance of marine
mucous macroaggregates in the northern Adriatic
has been observed for more than 250 years (Fonda-
Umani et al., 1989), but many details about theircauses and mechanisms of formation and detailed
chemical composition remain obscure. Moreover,their degradation pathways, comprising biologicaland photochemical processes, are also largely
unknown. Among possible reactions photodegrada-tion which is competitive with microbially mediatedreactions could be of special importance in shallow
summer strati®ed coastal waters. Light may act asthe energy source for priming refractory organicmaterials for further degradation (Zika, 1981;
Lindell et al., 1995; Wetzel et al., 1995; Miller andMoran, 1997; Moran and Zepp, 1997). The entirephotic zone represents a region of potential photo-chemical reactivity, and some 80±90% of visible
and ultraviolet radiation incident on the oceans isavailable to initiate photochemical reactions (Zika,1981). Photochemical reactions have already been
shown to play an important role in the chemistryand biology of the upper part of the water column(Za®riou et al., 1984).
In this paper we present the chemical elucidationof the composition of phytoplankton macroaggre-gates using 13C and 1H NMR techniques, as well as
a study of photochemical degradation of the watersoluble fraction of macroaggregates under exposureto sunlight and in photochemical reactors. In paral-lel, photochemical degradation studies of aqueous
solution of model carbohydrate compounds, namelypullulan (1,6 and 1,4 linkages), laminarin (1,3 lin-kages), maltose (1,4 linkages) and glucose, were per-
formed to obtain insight into the photochemicaldegradation pathways of macroaggregates sincealgal polysaccharides are synthesized with a wide
range of linkages including (1,2), (1,3), (1,4) and(1,6) for non-branched and (1,2,3), (1,2,4), (1,3,4),(1,3,6), (3,4,6) for branched sugars (Aluwihare etal., 1997).
MATERIALS AND METHODS
Samples
Macroaggregates were collected in July 1991 atthe sea surface using polyethylene bottles in thesouth eastern part of the Gulf of Trieste (northern
Adriatic). At that time in the investigated area themacroaggregates were well developed and gelati-nous surface layers covered large portions of the
sea surface. Hence, it was possible to sample theseaggregates with a minimal amount of surroundingwater.1 l of collected macroagreggates was centrifuged
at 15,000 rpm for 15 min at ambient (208C) tem-perature. Part of the gelatinous supernatant wasused for photochemical experiments. The gelatinous
sediment was rinsed with distilled water severaltimes to remove salt and then freeze-dried. Thefreeze-dried macroaggregates were extracted twice
with diethyl ether to remove lipids and pigments
N. Kovac et al.1624

and the residue gently evaporated to dryness. Drymacroaggregates were used for C and N elemental
and 13C and 1H NMR spectroscopic analyses.
Analyses
The organic C and (total) N content of thefreeze-dried and acid-washed sample of macroaggre-gates was determined using a Carlo Erba model1108 elemental analyzer.
The greatest part (080%) of the powderedmacroaggregates were solubilized using trimethylsi-lylchloride in the presence of triethylamine, or
acetic anhydride at room temperature. They weresubsequently dissolved in deuterated chloroformand tetramethyl silane (TMS) was added as an in-
ternal standard. 13C and 1H NMR spectra wererecorded on a Bruker VVX 300 spectrometer. Theexperimental conditions for 13C NMR analysis
were: temperature 300 K, spectrometer frequency75.5 MHz, number of scans 16±18,000, pulse width9 ms, spectral width 16,667 Hz, acquisition time1.38±1.97 s and pulse delay 1 s. The experimental
conditions for 1H NMR analyses were: temperature302 K, spectrometer frequency 300.1 MHz, numberof scans 16±18,000, pulse width 11±12.3 ms, spectralwidth 6173 Hz, acquisition time 2.65 s and pulsedelay 1 s.
Photochemical experiments
2 ml of gelatinous supernatant were diluted withdistilled water to obtain 100 ml of solution. In par-allel, 100 ml of 0.1 mM aqueous solutions of lami-
narin, maltose and glucose and 10 mM of pullulan(all from Sigma) were also used in photochemicalexperiments. The solutions were irradiated in an
immersion well photochemical reactor from AppliedPhotophysics (England), model RQ 125, equippedwith a 125 W medium pressure mercury lamp. This
lamp emits predominantly 365±366 nm radiation,with smaller amounts in UV at 265, 297, 303, 313and 334 nm, as well as a signi®cant amount in thevisible region at 404±408, 436±546 and 577±579 nm.
The temperature during the experiment was 258C.Photolysis in sunlight was performed in stopperedquartz tubes 30�2 cm exposed for 44 days at the
Meteorological Station in Piran in August andSeptember 1996. In parallel, measurements of totalirradiance (WMO) and UV-B light (MS-210 W UV-
B pyranometer, EKO-Japan) were performed. Darkcontrols were wrapped in aluminium foil.Concentrations of dissolved monosaccharides wereperiodically monitored during irradiation (after 2,
4, 6, 8, 11 and 30 h) using the procedure ofJohnson and Sieburth (1977) and the data obtainedwas used for kinetic studies. This method is based
on the colorimetric detection of formaldehyde pro-duced from the periodic acid oxidation of monosac-charide-derived alditols with 3-methyl-2-
benzothiazolinone hydrazone hydrochloride
(MBTH). Total dissolved carbohydrates were alsoanalyzed colorimetrically with MBTH (Burney and
Sieburth, 1977) after 12 M H2SO4 pretreatment andhydrolysis of the samples (Pakulski and Benner,1992). The isolation of reaction products was
accomplished by gentle evaporation of the irra-diated solution to dryness. The products were dis-solved in deuterated water, TMS was added as an
internal standard and 1H NMR spectra wererecorded on Bruker VVX 300 spectrometer asdescribed before.
RESULTS AND DISCUSSION
Composition of macroaggregates
Macroaggregates from the gelatinous surfacelayer contained more than 90% water according totheir gel-like nature (Alldredge and Crocker, 1995).The organic C and total N contents averaged 18.2
and 1.8%, respectively. Macroaggregates were char-acaterized by rather a high C/N ratio (approxi-mately 12, atomic), due to their low protein content
(Faganeli et al., 1995) and their well developedstage (gelatinous surface layer) indicating thatrefractory substances accumulate in aging aggre-
gates (MuÈ ller-Niklas et al., 1994).In Fig. 1 the 13C NMR spectra of silylated and
acetylated macroaggregates from the northernAdriatic are presented. The region at d= 0±50 ppm
is assigned to para�nic carbons (Wilson et al.,1981; Poutanen, 1985; Kalinowski and Blondeau,1988; Guggenberger et al., 1994; Knicker et al.,
1996). Strong peaks observed in the spectra atd= 25 ppm in silylated and d = 40 ppm in acetyl-ated derivatives are due to derivatization. The res-
onance around d= 33 ppm is characteristic ofpolymethylene chains (Poutanen, 1985; Zelibor etal., 1988; Benner et al., 1990; Derenne et al., 1991;
Gelin et al., 1996; Knicker et al., 1996). Thed= 55 ppm peak may be due to methoxy carbon(Wilson et al., 1981; Benner et al., 1990; Gamble etal., 1994; Guggenberger et al., 1994; Sihombing et
al., 1996). The spectra show strong signals in therange d= 65±100 ppm which were assigned to car-bon atoms in the carbohydrates (Benner et al.,
1990; McCarthy et al., 1993; de Beer et al., 1994;Gamble et al., 1994; Guggenberger et al., 1994;Knicker et al., 1996; Sihombing et al., 1996), con-
sidering the fact that carbohydrates have beenidenti®ed as major components of macroaggregates(Faganeli et al., 1995; Baldi et al., 1997). The reson-ances at 50±110 ppm could be also due to carbons
bound to heteroatoms (Allard et al., 1997) mostlikely to oxygen. The resonances aroundd= 100 ppm can be assigned to anomeric C-1 car-
bon in carbohydrates (Benner et al., 1990;Guggenberger et al., 1994) and/or ±O±C±O± func-tionalities (Poutanen, 1985). This is also con®rmed
by previously obtained FTIR spectra showing
Characterization and photodegradation of macroaggregates 1625
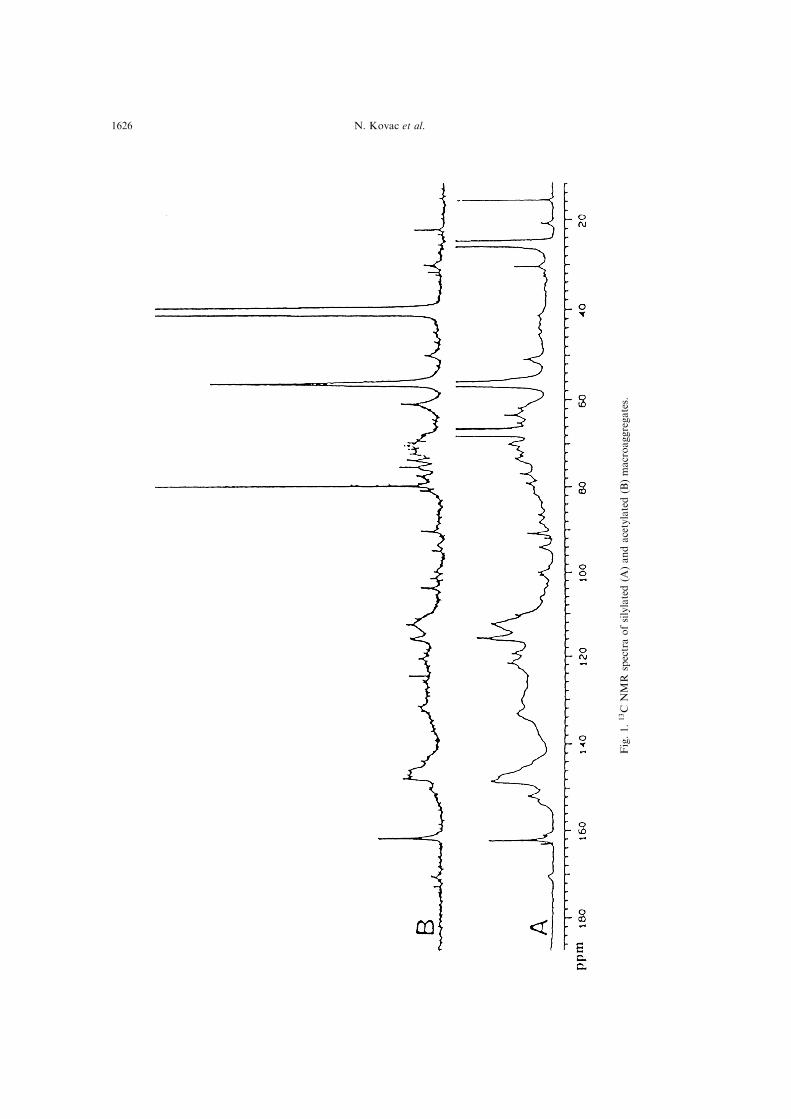
Fig.1.13C
NMR
spectraofsilylated(A
)andacetylated(B)macroaggregates.
N. Kovac et al.1626
intense bands at 1124, 1200 and 1096 cmÿ1 (Kovac
et al., 1995). The signals between d = 110±160 ppmare assignable to aromatic-C and ole®nic-C whichare not distinguishable (Wilson et al., 1981;
Poutanen, 1985). Signals at d = 140±160 ppm canbe assigned to aromatic esters and amides (Gelin etal., 1996). The aromatic region between d= 145±
160 ppm also contains signals from hetero-substi-tuted (O, N) aromatic carbons (Wilson et al., 1981;
Poutanen, 1985; Benner et al., 1990). A weak signalaround d = 170 ppm can be assigned to carbonyl-C(Wieruszeski et al., 1989) in carboxyl groups,
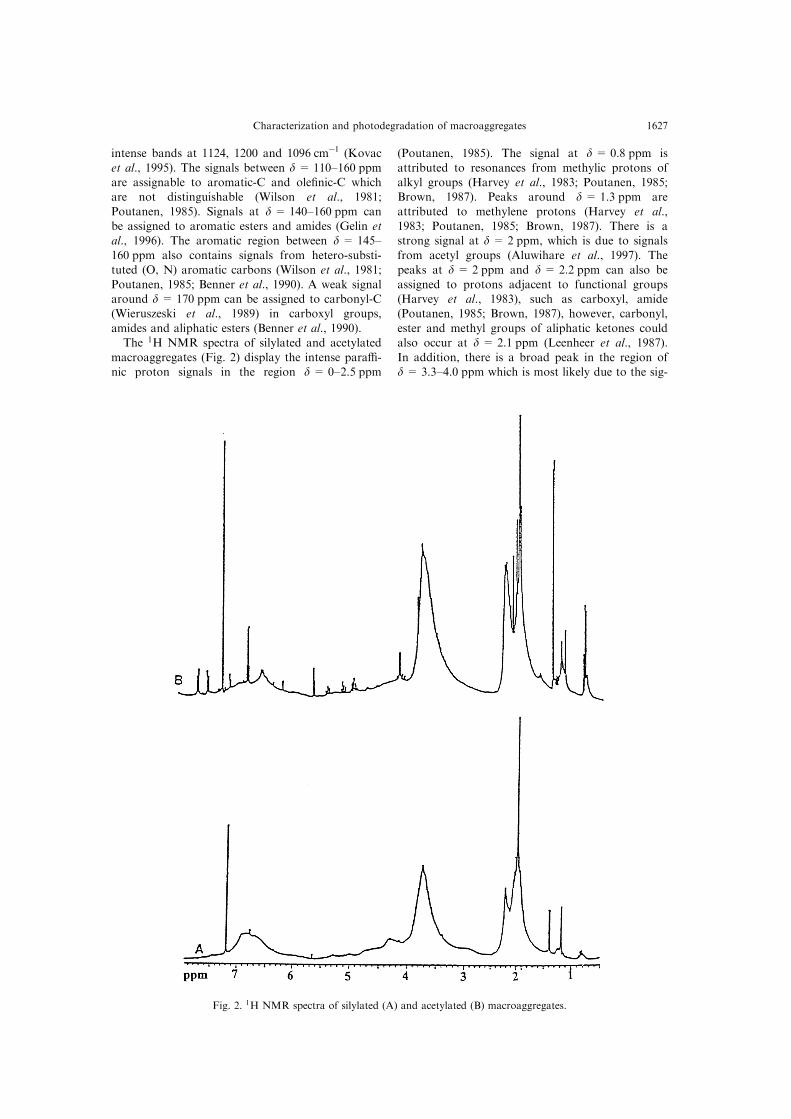
amides and aliphatic esters (Benner et al., 1990).The 1H NMR spectra of silylated and acetylated
macroaggregates (Fig. 2) display the intense para�-nic proton signals in the region d = 0±2.5 ppm
(Poutanen, 1985). The signal at d = 0.8 ppm is
attributed to resonances from methylic protons ofalkyl groups (Harvey et al., 1983; Poutanen, 1985;Brown, 1987). Peaks around d = 1.3 ppm are
attributed to methylene protons (Harvey et al.,1983; Poutanen, 1985; Brown, 1987). There is astrong signal at d = 2 ppm, which is due to signals
from acetyl groups (Aluwihare et al., 1997). Thepeaks at d = 2 ppm and d = 2.2 ppm can also be
assigned to protons adjacent to functional groups(Harvey et al., 1983), such as carboxyl, amide(Poutanen, 1985; Brown, 1987), however, carbonyl,
ester and methyl groups of aliphatic ketones couldalso occur at d = 2.1 ppm (Leenheer et al., 1987).
In addition, there is a broad peak in the region ofd= 3.3±4.0 ppm which is most likely due to the sig-
Fig. 2. 1H NMR spectra of silylated (A) and acetylated (B) macroaggregates.
Characterization and photodegradation of macroaggregates 1627
nals from protons in carbohydrates (Poutanen,
1985; Brown, 1987; Homans, 1992; Arnosti and
Repeta, 1994). The signals in the aromatic (Harvey
et al., 1983; Poutanen, 1985; Brown, 1987) and con-
jugated ole®nic region between d= 6±9 ppm are
also present (Poutanen, 1985).
Considering the NMR evidence it emerges that
carbohydrates, polymethylene chains and carboxyl
groups are present and it is our belief that the poly-
methylene chains compose a part of bound car-
boxylic acids. Although it is possible that they are
bonded in some other way, we favour this structure
because it is commonly encountered in lipids (de
Leeuw and Largeau, 1993). The polymethylene
chains are rather long and either free or connecting
di�erent heteropolysaccharide chains. These rather
long polymethylene chains could contain up to 29
C atoms (Derenne et al., 1996) and represent,
according to integrals in our 13C NMR spectra,
nearly half of the C atoms in analyzed macroaggre-
gates. Various authors reported that these amides
and esters could be an integral part of the poly-
methylenic network of refractory algaenan
(Derenne et al., 1993; Knicker et al., 1996) and
hypothetize that some of them could be protein-
aceous or lipidic in nature (Knicker et al., 1996).
The described structures are commonly found in
the resistant biopolymers of algae (Zelibor et al.,
1988; Derenne et al., 1992) and originate, similarlyto structural polysaccharides, from the cell wall of
phytoplanktonic algae (Derenne et al., 1993, 1996).Moreover, a similar macromolecular structure, com-posed of carbohydrates, acetate and lipids, was
recently proposed for ultra®ltrated dissolved or-ganic matter (UDOM) in surface sea water(Aluwihare et al., 1997).
Photochemical degradation
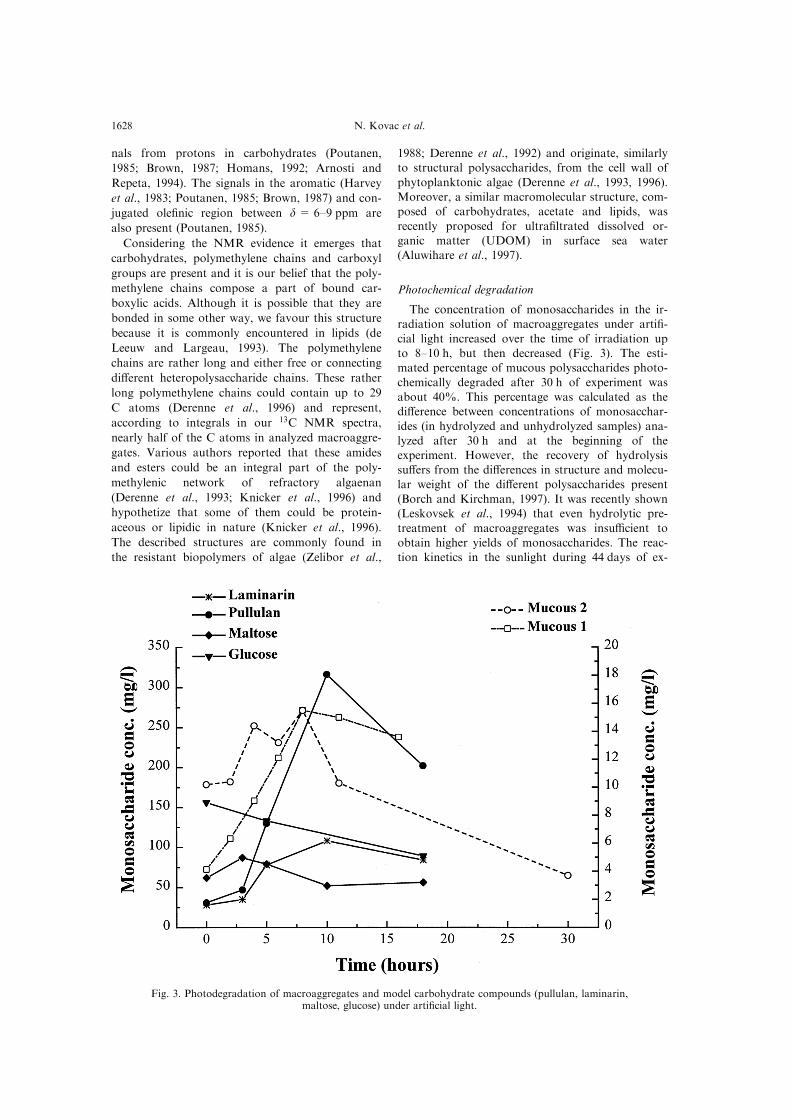
The concentration of monosaccharides in the ir-radiation solution of macroaggregates under arti®-
cial light increased over the time of irradiation upto 8±10 h, but then decreased (Fig. 3). The esti-mated percentage of mucous polysaccharides photo-
chemically degraded after 30 h of experiment wasabout 40%. This percentage was calculated as thedi�erence between concentrations of monosacchar-
ides (in hydrolyzed and unhydrolyzed samples) ana-lyzed after 30 h and at the beginning of theexperiment. However, the recovery of hydrolysissu�ers from the di�erences in structure and molecu-
lar weight of the di�erent polysaccharides present(Borch and Kirchman, 1997). It was recently shown(Leskovsek et al., 1994) that even hydrolytic pre-
treatment of macroaggregates was insu�cient toobtain higher yields of monosaccharides. The reac-tion kinetics in the sunlight during 44 days of ex-
Fig. 3. Photodegradation of macroaggregates and model carbohydrate compounds (pullulan, laminarin,maltose, glucose) under arti®cial light.
N. Kovac et al.1628
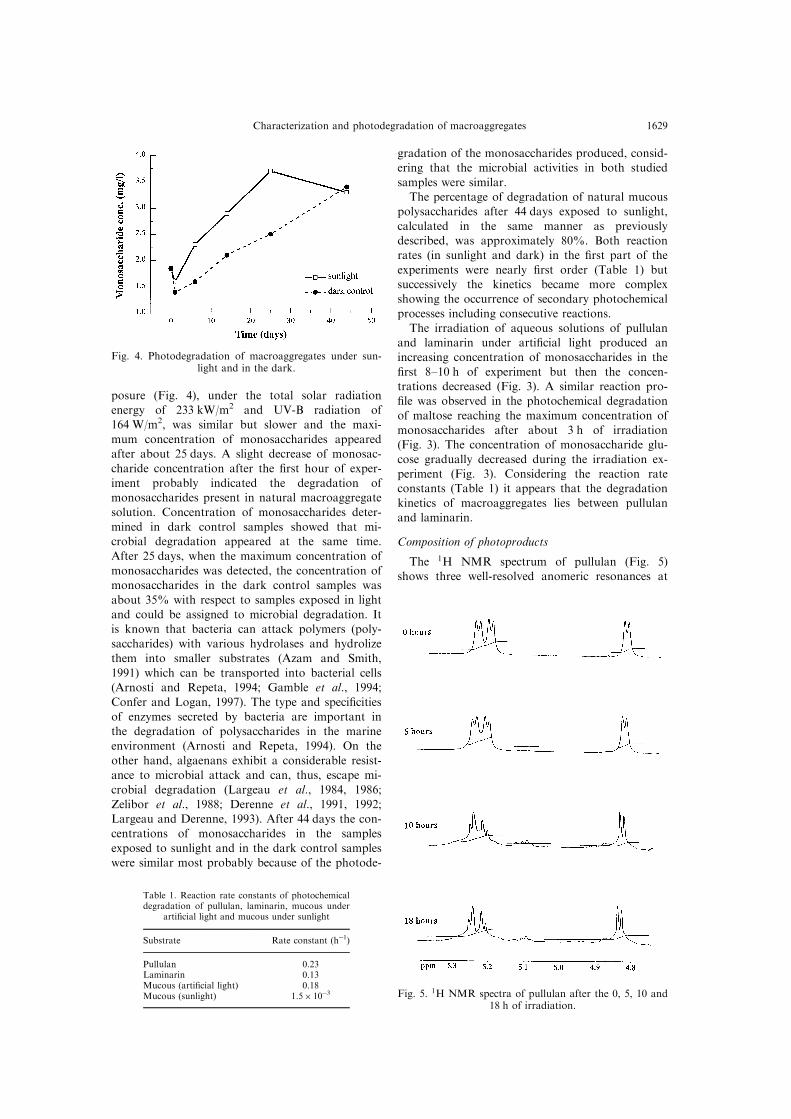
posure (Fig. 4), under the total solar radiationenergy of 233 kW/m2 and UV-B radiation of
164 W/m2, was similar but slower and the maxi-
mum concentration of monosaccharides appearedafter about 25 days. A slight decrease of monosac-
charide concentration after the ®rst hour of exper-
iment probably indicated the degradation ofmonosaccharides present in natural macroaggregate
solution. Concentration of monosaccharides deter-
mined in dark control samples showed that mi-crobial degradation appeared at the same time.
After 25 days, when the maximum concentration of
monosaccharides was detected, the concentration ofmonosaccharides in the dark control samples was
about 35% with respect to samples exposed in light
and could be assigned to microbial degradation. Itis known that bacteria can attack polymers (poly-
saccharides) with various hydrolases and hydrolize
them into smaller substrates (Azam and Smith,
1991) which can be transported into bacterial cells(Arnosti and Repeta, 1994; Gamble et al., 1994;
Confer and Logan, 1997). The type and speci®cities
of enzymes secreted by bacteria are important inthe degradation of polysaccharides in the marine
environment (Arnosti and Repeta, 1994). On the
other hand, algaenans exhibit a considerable resist-ance to microbial attack and can, thus, escape mi-
crobial degradation (Largeau et al., 1984, 1986;
Zelibor et al., 1988; Derenne et al., 1991, 1992;Largeau and Derenne, 1993). After 44 days the con-
centrations of monosaccharides in the samples
exposed to sunlight and in the dark control sampleswere similar most probably because of the photode-
gradation of the monosaccharides produced, consid-ering that the microbial activities in both studied
samples were similar.The percentage of degradation of natural mucous
polysaccharides after 44 days exposed to sunlight,
calculated in the same manner as previouslydescribed, was approximately 80%. Both reactionrates (in sunlight and dark) in the ®rst part of the
experiments were nearly ®rst order (Table 1) butsuccessively the kinetics became more complexshowing the occurrence of secondary photochemical
processes including consecutive reactions.The irradiation of aqueous solutions of pullulan
and laminarin under arti®cial light produced anincreasing concentration of monosaccharides in the
®rst 8±10 h of experiment but then the concen-trations decreased (Fig. 3). A similar reaction pro-®le was observed in the photochemical degradation
of maltose reaching the maximum concentration ofmonosaccharides after about 3 h of irradiation(Fig. 3). The concentration of monosaccharide glu-
cose gradually decreased during the irradiation ex-periment (Fig. 3). Considering the reaction rateconstants (Table 1) it appears that the degradation
kinetics of macroaggregates lies between pullulanand laminarin.
Composition of photoproducts
The 1H NMR spectrum of pullulan (Fig. 5)shows three well-resolved anomeric resonances at
Fig. 4. Photodegradation of macroaggregates under sun-light and in the dark.
Table 1. Reaction rate constants of photochemicaldegradation of pullulan, laminarin, mucous under
arti®cial light and mucous under sunlight
Substrate Rate constant (hÿ1)
Pullulan 0.23Laminarin 0.13Mucous (arti®cial light) 0.18Mucous (sunlight) 1.5� 10ÿ3 Fig. 5. 1H NMR spectra of pullulan after the 0, 5, 10 and
18 h of irradiation.
Characterization and photodegradation of macroaggregates 1629
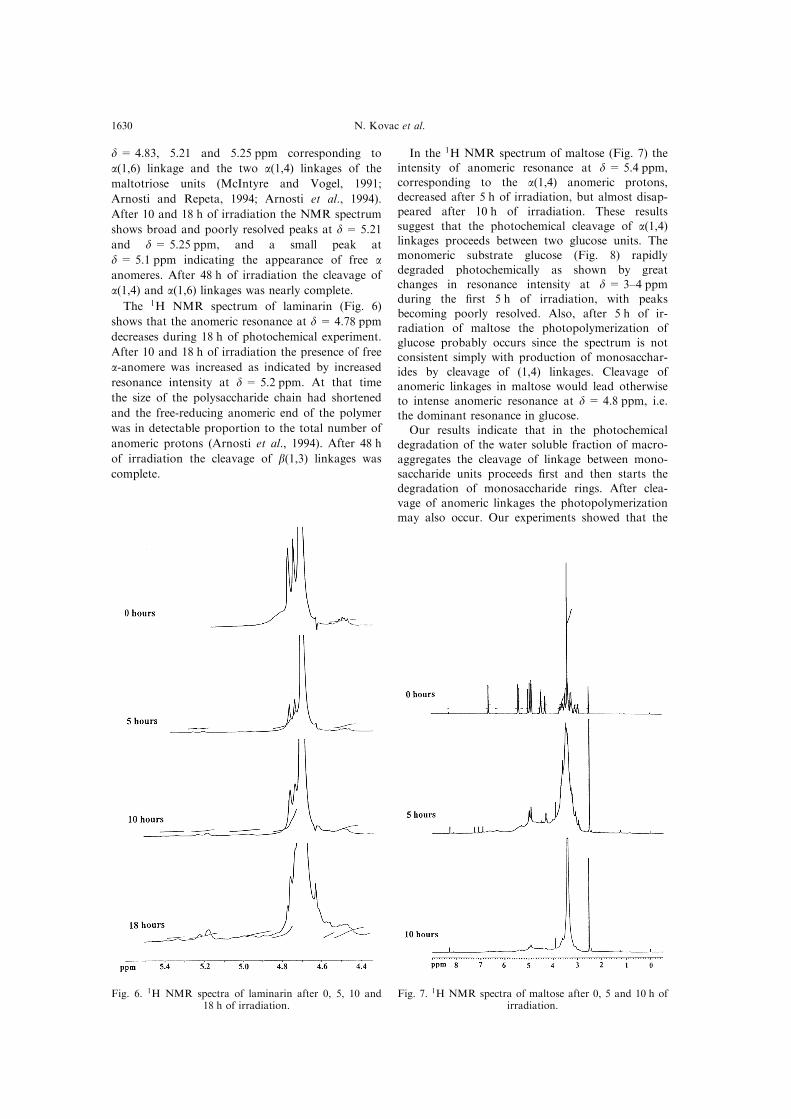
d = 4.83, 5.21 and 5.25 ppm corresponding to
a(1,6) linkage and the two a(1,4) linkages of the
maltotriose units (McIntyre and Vogel, 1991;
Arnosti and Repeta, 1994; Arnosti et al., 1994).
After 10 and 18 h of irradiation the NMR spectrum
shows broad and poorly resolved peaks at d= 5.21
and d= 5.25 ppm, and a small peak at
d = 5.1 ppm indicating the appearance of free aanomeres. After 48 h of irradiation the cleavage of
a(1,4) and a(1,6) linkages was nearly complete.
The 1H NMR spectrum of laminarin (Fig. 6)
shows that the anomeric resonance at d = 4.78 ppm
decreases during 18 h of photochemical experiment.
After 10 and 18 h of irradiation the presence of free
a-anomere was increased as indicated by increased
resonance intensity at d = 5.2 ppm. At that time
the size of the polysaccharide chain had shortened
and the free-reducing anomeric end of the polymer
was in detectable proportion to the total number of
anomeric protons (Arnosti et al., 1994). After 48 h
of irradiation the cleavage of b(1,3) linkages was
complete.
In the 1H NMR spectrum of maltose (Fig. 7) theintensity of anomeric resonance at d = 5.4 ppm,
corresponding to the a(1,4) anomeric protons,decreased after 5 h of irradiation, but almost disap-peared after 10 h of irradiation. These results
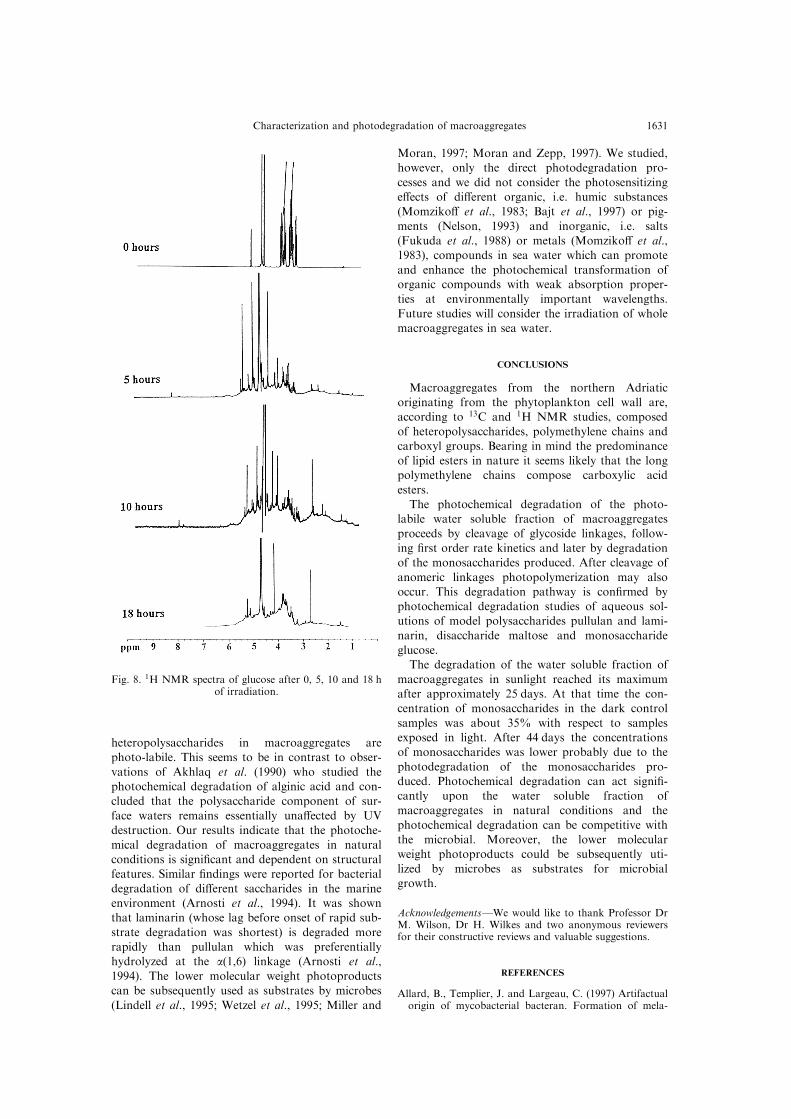
suggest that the photochemical cleavage of a(1,4)linkages proceeds between two glucose units. Themonomeric substrate glucose (Fig. 8) rapidly
degraded photochemically as shown by greatchanges in resonance intensity at d = 3±4 ppmduring the ®rst 5 h of irradiation, with peaks
becoming poorly resolved. Also, after 5 h of ir-radiation of maltose the photopolymerization ofglucose probably occurs since the spectrum is not
consistent simply with production of monosacchar-ides by cleavage of (1,4) linkages. Cleavage ofanomeric linkages in maltose would lead otherwiseto intense anomeric resonance at d= 4.8 ppm, i.e.
the dominant resonance in glucose.Our results indicate that in the photochemical
degradation of the water soluble fraction of macro-
aggregates the cleavage of linkage between mono-saccharide units proceeds ®rst and then starts thedegradation of monosaccharide rings. After clea-
vage of anomeric linkages the photopolymerizationmay also occur. Our experiments showed that the
Fig. 6. 1H NMR spectra of laminarin after 0, 5, 10 and18 h of irradiation.
Fig. 7. 1H NMR spectra of maltose after 0, 5 and 10 h ofirradiation.
N. Kovac et al.1630
heteropolysaccharides in macroaggregates arephoto-labile. This seems to be in contrast to obser-
vations of Akhlaq et al. (1990) who studied thephotochemical degradation of alginic acid and con-cluded that the polysaccharide component of sur-
face waters remains essentially una�ected by UVdestruction. Our results indicate that the photoche-
mical degradation of macroaggregates in naturalconditions is signi®cant and dependent on structuralfeatures. Similar ®ndings were reported for bacterial
degradation of di�erent saccharides in the marineenvironment (Arnosti et al., 1994). It was shownthat laminarin (whose lag before onset of rapid sub-
strate degradation was shortest) is degraded morerapidly than pullulan which was preferentiallyhydrolyzed at the a(1,6) linkage (Arnosti et al.,
1994). The lower molecular weight photoproductscan be subsequently used as substrates by microbes
(Lindell et al., 1995; Wetzel et al., 1995; Miller and
Moran, 1997; Moran and Zepp, 1997). We studied,however, only the direct photodegradation pro-
cesses and we did not consider the photosensitizinge�ects of di�erent organic, i.e. humic substances(Momziko� et al., 1983; Bajt et al., 1997) or pig-
ments (Nelson, 1993) and inorganic, i.e. salts(Fukuda et al., 1988) or metals (Momziko� et al.,1983), compounds in sea water which can promote
and enhance the photochemical transformation oforganic compounds with weak absorption proper-ties at environmentally important wavelengths.
Future studies will consider the irradiation of wholemacroaggregates in sea water.
CONCLUSIONS
Macroaggregates from the northern Adriaticoriginating from the phytoplankton cell wall are,according to 13C and 1H NMR studies, composed
of heteropolysaccharides, polymethylene chains andcarboxyl groups. Bearing in mind the predominanceof lipid esters in nature it seems likely that the longpolymethylene chains compose carboxylic acid
esters.The photochemical degradation of the photo-
labile water soluble fraction of macroaggregates
proceeds by cleavage of glycoside linkages, follow-ing ®rst order rate kinetics and later by degradationof the monosaccharides produced. After cleavage of
anomeric linkages photopolymerization may alsooccur. This degradation pathway is con®rmed byphotochemical degradation studies of aqueous sol-utions of model polysaccharides pullulan and lami-
narin, disaccharide maltose and monosaccharideglucose.The degradation of the water soluble fraction of
macroaggregates in sunlight reached its maximumafter approximately 25 days. At that time the con-centration of monosaccharides in the dark control
samples was about 35% with respect to samplesexposed in light. After 44 days the concentrationsof monosaccharides was lower probably due to the
photodegradation of the monosaccharides pro-duced. Photochemical degradation can act signi®-cantly upon the water soluble fraction ofmacroaggregates in natural conditions and the
photochemical degradation can be competitive withthe microbial. Moreover, the lower molecularweight photoproducts could be subsequently uti-
lized by microbes as substrates for microbialgrowth.
AcknowledgementsÐWe would like to thank Professor DrM. Wilson, Dr H. Wilkes and two anonymous reviewersfor their constructive reviews and valuable suggestions.
REFERENCES
Allard, B., Templier, J. and Largeau, C. (1997) Artifactualorigin of mycobacterial bacteran. Formation of mela-
Fig. 8. 1H NMR spectra of glucose after 0, 5, 10 and 18 hof irradiation.
Characterization and photodegradation of macroaggregates 1631

noidin-like artifact macromolecular material during theusual isolation process. Organic Geochemistry 26, 691±703.
Alldredge, A. L. and Crocker, K. M. (1995) Why do sink-ing mucilage aggregates accumulate in the water col-umn? The Science of the Total Environment 165, 15±22.
Aluwihare, L. I., Repeta, D. J. and Chen, R. F. (1997) Amajor biopolymeric component to dissolved organic car-bon in surface sea water. Nature 387, 166±169.
Akhlaq, M. S., Schuchmann, H.-P. and von Sonntag,C. (1990) Degradation of the polysaccharide alginicacid: A comparison of the e�ects of UV light andozone. Environmental Science and Technology 24, 379±383.
Arnosti, C. and Repeta, D. J. (1994) Extracellular enzymeactivity in anaerobic bacterial cultures: Evidence of pull-ulanase activity among mesophilic marine bacteria.Applied and Environmental Microbiology 60, 840±846.
Arnosti, C., Repeta, D. J. and Blough, N. V. (1994) Rapidbacterial degradation of polysaccharides in anoxic mar-ine systems. Geochimica et Cosmochimica Acta 58,2639±2652.
Azam, F. and Smith, D. C. (1991) Bacterial in¯uence onthe variability in the ocean's biogeochemical state: Amechanistic view. In Particles Analysis in Oceanography,ed. S. Demers, NATO ASI Series, Series G, Vol. 27.Springer Verlag, Berlin, pp. 213±236.
Bajt, O., Sket, B. and Faganeli, J. (1997) The aqueousphotochemical transformation of acrylic acid. MarineChemistry 58, 255±259.
Baldi, F., Minacci, A., Saliot, A., Mejanelle, L., Mozetic,P., Turk, V. and Malej, A. (1997) Cell lysis and releaseof particulate polysaccharides in extensive marine muci-lage assessed by lipid biomarkers and molecular probes.Marine Ecology Progress Series 153, 45±57.
de Beer, T., van Zuylen, C. W. E. M., Hard, K., Boelens,R., Kaptein, R., Kamerling, J. P. and Vliegenthart, F.G. (1994) Rapid and simple approach for the NMR res-onance assignment of the carbohydrate chains of anintact glycoprotein. Application of gradient-enhancednatural abundance 1H±13C HSQC and HSQC-TOSCYto the a-subunit of human chorionic gonadotropin.Federation of European Biochemical Societies 348, 1±6.
Benner, R., Hatcher, P. G. and Hedges, J. I. (1990) Earlydiagenesis of mangrove leaves in tropical estuary: Bulkchemical characterization using solid-state 13C NMRand elemental analyses. Geochimica et CosmochimicaActa 54, 2003±2013.
Biddanda, B. and Benner, R. (1997) Carbon, nitrogen andcarbohydrate ¯uxes during the production of particulateand dissolved matter by marine phytoplankton.Limnology and Oceanography 42, 506±518.
Borch, N. H. and Kirchman, D. L. (1997) Concentrationand composition of dissolved combined neutral sugars(polysaccharides) in seawater determined by HPLC-PAD. Marine Chemistry 57, 85±95.
Brown, J. W. (1987) Studies of humic and fulvic aciddynamics in coastal marine water of south Florida.Marine Environmental Research 21, 163±174.
Burney, C. M. and Sieburth, J. McN. (1977) Dissolvedcarbohydrates in sea water. II. A spectrophotometricprocedure for total carbohydrate analysis and polysac-charide determination. Marine Chemistry 5, 15±28.
Confer, D. R. and Logan, B. E. (1997) Molecular weightdistribution of hydrolysis products during the biodegra-dation of model macromolecules in suspended and bio-®lm cultures: II. Dextran and dextrin. Water Research31, 2137±2145.
Decho, A. W. (1990) Microbial exopolymer secretions inocean environments: their role(s) in food webs and mar-ine processes. Oceanography and Marine Biology 28, 73±153.
Decho, A. W. and Herndl, G. J. (1995) Microbial activi-ties and the transformation of organic matter withinmucilaginous material. The Science of the TotalEnvironment 165, 33±42.
Degobbis, D., Fonda-Umani, S., Franco, P., Malej, A.,Precali, R. and Smodlaka, N. (1995) Changes in thenorthern Adriatic ecosystem and the hypertrophicappearance of gelatinous aggregates. The Science of theTotal Environment 165, 43±58.
Derenne, S., Largeau, C. and Berkalo�, C. (1996) Firstexample of an algaenan yielding an aromatic-rich pyro-lysate. Possible geochemical implications on marinekerogen formation. Organic Geochemistry 24, 617±627.
Derenne, S., Largeau, C. and Taulelle, F. (1993)Occurrence of non-hydrolysable amides in the macro-molecular constituent of Scenedesmus quadricauda cellwall as revealed by 15N NMR: Origin of N-alkylnitrilesin pyrolysates of ultralaminae-containing kerogens.Geochimica et Cosmochimica Acta 57, 851±857.
Derenne, S., Largeau, C., Casadevall, E., Berkalo�, C. andRousseau, B. (1991) Chemical evidence of kerogen for-mation in source rocks and oil shales via selective pres-ervation of thin resistant outer walls of microalgae:origin of ultralaminae. Geochimica et CosmochimicaActa 55, 1041±1050.
Derenne, S., Le Berre, F., Largeau, C., Hatcher, P. G.,Connan, J. and Raynaud, J. F. (1992) Formation ofultralaminae in marine kerogens via selective preser-vation of thin resistant outer walls of microalgae.Organic Geochemistry 19, 345±350.
Faganeli, J. (1989) Sedimentation of particulate nitrogenand amino acids in shallow coastal waters (Gulf ofTrieste, northern Adriatic). Marine Chemistry 26, 67±80.
Faganeli, J., Kovac, N., Leskovsek, H. and Pezdic,J. (1995) Sources and ¯uxes of particulate organic mat-ter in shallow waters characterized by summer macroag-gregate formation. Biogeochemistry 29, 71±88.
Fanuko, N. and Turk, V. (1990) The oceanologic proper-ties of the sea water in Gulf of Trieste before andduring the ``mare sporco'' phenomenon (summer 1988).Bolettino di Oceanologia Teorica ed Applicata 8, 3±11.
Fonda-Umani, S., Ghiarardelli, E. and Specchi, M. (1989)Gli episodi di ``mare sporco'' nell'Adriatico da 1729 ainostri giorni. Regione autonoma Friuli-Venezia Giulia,Trieste, 178 pp.
Fukuda, K., Inagaki, Y., Maruyama, T., Kojima, H.I. and Yoshida, T. (1988) On the photolysis of alkylatednaphthalenes in aquatic systems. Chemosphere 17, 651±659.
Gamble, G. R., Sethuraman, A., Akin, D. E. andEriksson, K.-E. L. (1994) Biodegradation of lignocellu-lose in bermuda grass by white rot fungi analyzed bysolid-state 13C nuclear magnetic resonance. Applied andEnvironmental Microbiology 60, 3138±3144.
Gelin, F., Boogers, A. A. M., Sinninghe Damaste , J. S.,Hatcher, P. G. and de Leeuw, J. (1996) Novel, resistantmicroalgal polyethers: An important sink of organic car-bon in the marine environment? Geochimica etCosmochimica Acta 60, 1275±1280.
Guggenberger, G., Zech, W. and Schulten, H. (1994)Formation and mobilization pathways of dissolved or-ganic matter: evidence from chemical structural studiesof organic matter fractions in acid forest ¯oor solutions.Organic Geochemistry 21, 51±66.
Hama, J. and Handa, N. (1992) Diel variations of water-extractable carbohydrate composition of natural phyto-plankton populations in Kinu-Ura Bay. Journal ofExperimental Marine Biology and Ecology 62, 159±176.
Harvey, G. R., Boran, D. A., Chesal, L. A. and Tokar,J. M. (1983) The structure of marine fulvic and humicacids. Marine Chemistry 12, 119±132.
N. Kovac et al.1632

Herndl, G. J. (1992) Marine snow in the northern AdriaticSea: possible causes and consequences for shallow eco-system. Marine Microbial Food Webs 6, 149±172.
Homans, S. W. (1992) Homonuclear three-dimensionalNMR methods for the complete assignment of protonNMR spectra of oligosaccharides: application toGalb1±4(Fuca1±3)GlcNAcb1±3Galb1±4Glc.Glycobiology 2, 153±159.
Ittekkot, V., Degens, E. T. and Brockmann, V. (1982)Monosaccharide composition of acid-hydrolyzablecarbohydrates in particulate matter during a planktonbloom. Limnology and Oceanography 27, 770±776.
Johnson, K. M. and Sieburth, J. McN. (1977) Dissolvedcarbohydrates in sea water. I. A precise spectrophoto-metric analysis for monosaccharides. Marine Chemistry5, 1±13.
Kalinowski, E. and Blondeau, R. (1988) Characterizationof sedimentary humic acids fractionated by hydrophobicinteraction chromatography. Marine Chemistry 24, 29±37.
Kiùrboe, T., Andersen, K. P. and Dam, H. G. (1990)Coagulation e�ciency and aggregate formation in mar-ine phytoplankton. Marine Biology 107, 235±243.
Knicker, H., Scaroni, A. W. and Hatcher, P. G. (1996)13C and 15N NMR spectroscopic investigation on theformation of fossil algal residues. Organic Geochemistry24, 661±669.
Kovac, N., Faganeli, J., Bajt, O. and Leskovsek, H. (1995)Chemical characterization of macroaggregates from theNorthern Adriatic. In Organic Geochemistry:Developments and Applications to Energy, Climate,Environment and Human History, eds. J. O. Grimalt andC. Dorronsoro. A.I.G.O.A., Donostia-San Sebastian,Spain, pp. 1153±1155.
Largeau, C. and Derenne, S. (1993) Relative e�ciency ofthe selective preservation and degradation recondensa-tion pathways in kerogen formation. Source and en-vironment in¯uence of their contributions to type I andII kerogens. Organic Geochemistry 20, 611±615.
Largeau, C., Casadevall, E., Kadouri, A. and Metzger,P. (1984) Formation of Botryococcus braunii kerogens.Comparative study of immature Torbanite and of theextant alga Botryococcus braunii. Organic Geochemistry6, 327±332.
Largeau, C., Derenne, S., Casadevall, E., Kadouri, A. andSellier, N. (1986) Pyrolysis of immature Torbanite andof the resistant biopolymer (PRBA) isolated from extantalga Botryococcus braunii. Mechanism of formation andstructure of Torbanite. Organic Geochemistry 10, 1023±1032.
Leenheer, J. A., Wilson, M. A. and Malcom, R. L. (1987)Presence and potential signi®cance of aromatic-ketonegroups in aquatic humic substances. OrganicGeochemistry 11, 273±280.
de Leeuw, J. W. and Largeau, C. (1993) A review ofmacromolecular organic compounds that comprise liv-ing organisms and their role in kerogen, coal, and pet-roleum formation. In Organic Geochemistry, eds. M. H.Engel and S. A. Macko. Plenum Press, New York, pp.23±72.
Leskovsek, H., Perko, S., Zigon, D. and Faganeli, J. (1994)Analysis of carbohydrates in marine particulates by gaschromatography and tandem mass spectrometry.Analyst 119, 1125±1128.
Lindell, M. J., Grane li, W. and Tranvik, L. J. (1995)Enhanced bacterial growth in response to photochemi-cal transformation of dissolved organic matter.Limnology and Oceanography 40, 195±199.
Malej, A. and Harris, R. P. (1993) Inhibition of copepodgrazing by diatom exudates: a factor in the developmentof mucus aggregates? Marine Ecology Progress Series96, 33±42.
McCarthy, M. D., Hedges, J. I. and Benner, R. (1993)The chemical composition of dissolved organic matterin seawater. Chemical Geology 107, 503±507.
McIntyre, D. D. and Vogel, H. J. (1991) Nuclear magneticresonances studies of homopolysaccharides related tostarch. Starch 43, 69±76.
Miller, W. L. and Moran, M. A. (1997) Interaction ofphotochemical and microbial processes in the degra-dation of refractory dissolved organic matter from acoastal marine environment. Limnology andOceanography 42, 1317±1324.
Mingazzini, M. and Thake, B. (1995) Summary and con-clusions of the workshop on marine mucilages in theAdriatic Sea and elsewhere. The Science of the TotalEnvironment 165, 9±14.
Momziko�, A., Santus, R. and Giraud, M. (1983) A studyof the photosensitizing properties of seawater. MarineChemistry 12, 1±14.
Moran, M. A. and Zepp, R. G. (1997) Role of photoreac-tions in the formation of biologically labile compoundsfrom dissolved organic matter. Limnology andOceanography 42, 1307±1316.
MuÈ ller-Niklas, G., Schuster, S., KaltenboÈ ck, E. andHerndl, G. J. (1994) Organic content and bacterialmetabolism in amorphous aggregations of the northernAdriatic Sea. Limnology and Oceanography 39, 58±68.
Myklestad, S. M. (1995) Release of extracellular productsby phytoplankton with special emphasis on polysacchar-ides. The Science of the Total Environment 165, 155±164.
Nelson, J. R. (1993) Rates and possible mechanism oflight-dependent degradation of pigments in detritus de-rived from phytoplankton. Journal of Marine Research51, 155±179.
Pakulski, J. D. and Benner, R. (1992) An improvedmethod for the hydrolysis and MBTH analysis of dis-solved and particulate carbohydrates in seawater.Marine Chemistry 40, 143±160.
Posedel, N. and Faganeli, J. (1991) Nature and sedimen-tation of suspended particulate matter during densitystrati®cation in shallow coastal waters (Gulf of Trieste,northern Adriatic). Marine Ecology Progress Series 77,135±145.
Poutanen, E.-L. (1985) Comparison of the structures ofhumic acids from marine sediments and degraded ®elddiatoms by 13C- and 1H-NMR spectroscopy. MarineChemistry 17, 115±126.
Richardson, T. L. and Cullen, J. J. (1995) Changes inbuoyancy and chemical composition during growth of acoastal marine diatom: ecological and biogeochemicalconsequences. Marine Ecology Progress Series 128, 77±90.
Rinaldi, A., Vollenweider, R. A., Montanari, G., Ferrari,C. R. and Ghetti, A. (1995) Mucilages in Italian seas:the Adriatic and Tyrrhenian Seas, 1988±1991. TheScience of the Total Environment 165, 165±183.
Sihombing, R., Greenwood, P. F., Wilson, M. A. andHanna, J. V. (1996) Composition of size exclusion frac-tions of swamp water humic and fulvic acids asmeasured by solid state NMR and pyrolysis±gas chro-matography±mass spectrometry. Organic Geochemistry24, 859±873.
Stachowitsch, M., Fanuko, N. and Richter, M. (1990)Mucus aggregates in the Adriatic sea: An overview ofstages and occurrences. P.S.Z.N. I. Marine Ecology 11,327±350.
Vollenweider, R. A., Rinaldi, A. and Montanari, G.(1992) Eutrophication, structure and dynamics of mar-ine coastal system: results of ten-year monitoring alongthe Emilie-Romagna coast (Northwest Adriatic Sea). InMarine Coastal Eutro®cation, eds. R. A. Vollenweider,R. Marchetti and R. Viviani. Elsevier, Amsterdam, pp.63±106.
Characterization and photodegradation of macroaggregates 1633

Wetzel, R. G., Hatcher, P. G. and Bianchi, T. S. (1995)Natural photolysis by ultraviolet irradiance of recalci-trant dissolved organic matter to simple substrates forrapid bacterial metabolism. Limnology andOceanography 40, 1369±1380.
Wieruszeski, J.-M., Michalski, J.-C., Montreuil, J. andStrecker, G. (1989) Sequential 1H and 13C resonanceassignments for an octa- and decasaccharide of N-acet-yllactosamine type by multiple-step relayed correlationand heteronuclear correlation nuclear magnetic reson-ance. Glycoconjugate Journal 6, 183±194.
Wilson, M. A., Barron, P. F. and Gillam, A. H. (1981)The structure of freshwater humic substances asrevealed by 13C-NMR spectroscopy. Geochimica etCosmochimica Acta 45, 1743±1750.
Za®riou, O. C., Joussot-Dubien, J., Zepp, R. G. and Zika,R. G. (1984) Photochemistry of natural waters.Environmental Science and Technology 18, 358A±371A.
Zelibor, J. L. Jr., Romankiw, L., Hatcher, P. G. andCowell, R. R. (1988) Comparative analysis of the chemi-cal composition of mixed and pure cultures of greenalgae and their decomposed residues by 13C nuclear res-onance spectroscopy. Applied and EnvironmentalMicrobiology 56, 1051±1060.
Zika, R. G. (1981) Marine organic photochemistry. InMarine Organic Chemistry, eds. E. K. Duursma and R.Dawson. Elsevier Scienti®c Publishing Company,Amsterdam, pp. 299±326.
N. Kovac et al.1634





























