Embed Size (px)
Citation preview

Gene, 151 (1994) l-10
0 1994 Elsevier Science B.V. All rights reserved. 0378-l 119/94/$07.00
GENE 08442
Characterization of the Streptomycespeucetius ATCC 29050 genes encoding doxorubicin polyketide synthase
(Aklanonic acid; antitumor antibiotic; acyl carrier protein; acyltransferase; daunorubicin; c-rhodomycinone; ketoreductase; 3-oxoacyl : ACP synthase; polyketide cyclase; type II polyketide synthase)
Ann Grimma, Krishnamurthy Madduri”, Arif Ali” and C.R. Hutchinsonb,”
‘Department of Genetics, University of Wisconsin, Madison, WI 53706, USA. Tel. (I-608) 262-2459; bDepartment of Bacteriology, University of
Wisconsin, Madison, WI 53706, USA; and ‘School of Pharmaq University of Wisconsin, Madison, WI 53706, USA
Received by K.F. Chater: 1 July 1994; Revised/Accepted: 22 August/24 August 1994; Received at publishers: 22 September 1994
1
SUMMARY
The dps genes of Streptomyces peucetius, encoding daunorubicin (DNR)-doxorubicin (DXR) polyketide synthase (PKS), are largely within an 8.7-kb region of DNA that has been characterized by Southern analysis, and gene sequencing, mutagenesis and expression experiments. This region contains nine ORFs, many of whose predicted products are homologous to known PKS enzymes. Surprisingly, the gene encoding the DXR PKS acyl carrier protein is not in this region, but is located about 10 kb distant from the position it usually occupies in other gene clusters encoding type-II PKS. An in-frame deletion in the @sB gene, encoding a putative subunit of the DXR PKS, resulted in loss of production of DXR and the known intermediates of its biosynthetic pathway, confirming that this gene and, by implication, the adjacent dps genes are required for DXR biosynthesis. This was verified by expression of the dps genes in the heterologous host, Streptomyces lividans, which resulted in the production of aklanonic acid, an early intermediate of DXR biosynthesis.
INTRODUCTION
DXR and its precursor, DNR, are chemotherapeutic antitumor antibiotics produced by S. peucetius. Although DXR, in particular, is effective against many kinds of cancer it is very expensive to produce and DXR is cumu-
Correspondence to: Dr. C.R. Hutchinson, School of Pharmacy,
University of Wisconsin-Madison, 425 N. Charter Street, Madison, WI
53706, USA. Tel. (l-608) 262-7582; Fax (l-608) 262-3134;
e-mail: [email protected]
Abbreviations: aa, amino acid(s); ACP, acyl carrier protein; ACT, acti-
norhodin; calcd., calculated; Am, apramycin; Ap, ampicillin; CoA, coen-
zyme A; DNR, daunorubicin; dps, genes encoding daunorubicin-
doxorubicin polyketide synthase; DXR, doxorubicin; FAS, fatty acid
synthase(s); HPLC, high-performance liquid chromatography; kb, kilo-
base(s) or 1000 bp; m/z, mass-to-charge ratio; MTase, methyltransferase;
ORF (Orf), open reading frame; PKS, polyketide synthase(s); RHO,
c-rhodomycinone; S., Streptomyces; SNA, soft nutrient agar; TCM,
tetracenomycin C; Th, thiostrepton; TLC, thin-layer chromatography;
UV, ultraviolet.
SSDI 0378-1119(94)00653-9
latively cardiotoxic so that its dosage must be limited. Study of the genes involved in DXR biosynthesis might lead to cheaper production and to the development of less toxic drugs. In addition, a study of the DXR pro- duction genes should provide new information about Streptomyces genetics because DXR production appears to involve more complex regulatory and resistance mech- anisms than other model systems for the biosynthesis of aromatic polyketides, such as TCM C biosynthesis in S. glaucescens (Guilfoile and Hutchinson, 1992; Decker and Hutchinson, 1993) or ACT biosynthesis in S. coelico/or
(Hallam et al., 1988; Fernandez-Moreno et al., 1991; Gramajo et al., 1993).
We had located the most likely region containing the dps genes by Southern blot hybridization during the iso- lation of the DXR gene cluster (Stutzman-Engwall and Hutchinson, 1989; Otten et al., 1990). The products of these genes, which are likely to be similar to components of bacterial type-II FAS (Magnuson et al., 1993) have
2
homologs in other Streptomyces species that produce aro-
matic polyketide antibiotics by type-II PKS (Hopwood
and Sherman, 1990; Katz and Donadio, 1993). The DXR
PKS creates the carbon backbone of RHO, a key inter-
mediate of DXR biosynthesis (Strohl et al., 1989) by
selecting the starter unit, propionyl-CoA, then adding 9
malonyl-CoA carbon chain extending units to form a
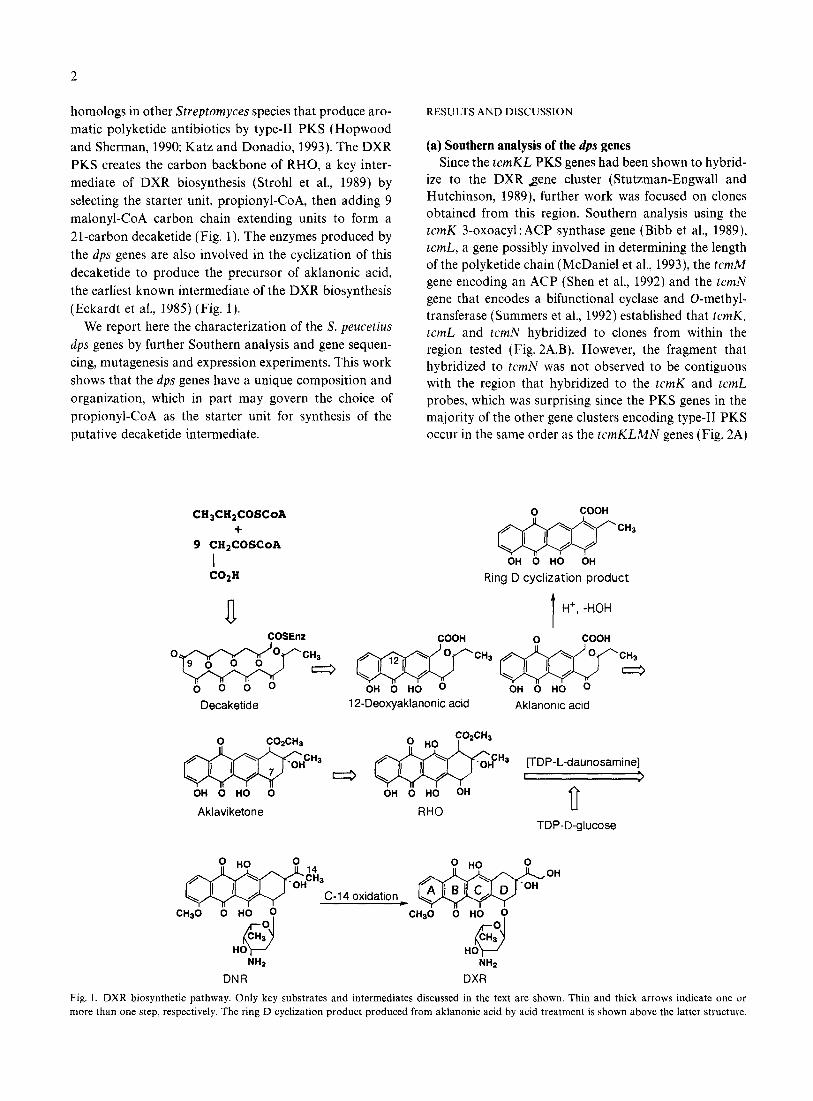
21-carbon decaketide (Fig. 1). The enzymes produced by
the dps genes are also involved in the cyclization of this
decaketide to produce the precursor of aklanonic acid,
the earliest known intermediate of the DXR biosynthesis
(Eckardt et al., 1985) (Fig. 1).
We report here the characterization of the S. peucetius
dps genes by further Southern analysis and gene sequen-
cing, mutagenesis and expression experiments. This work
shows that the dps genes have a unique composition and
organization, which in part may govern the choice of
propionyl-CoA as the starter unit for synthesis of the
putative decaketide intermediate.
RESULTS AND DISCUSSION
(a) Southern analysis of the dps genes Since the tcmKL PKS genes had been shown to hybrid-
ize to the DXR gene cluster (Stutzman-Engwall and Hutchinson, 1989), further work was focused on clones obtained from this region. Southern analysis using the tcmK 3-oxoacyl : ACP synthase gene (Bibb et al., 1989), tcmL, a gene possibly involved in determining the length of the polyketide chain (McDaniel et al., 1993), the tcmM
gene encoding an ACP (Shen et al., 1992) and the tcmN gene that encodes a bifunctional cyclase and O-methyl- transferase (Summers et al., 1992) established that tcmK, tcmL and tcmN hybridized to clones from within the region tested (Fig. 2A,B). However, the fragment that hybridized to tcmN was not observed to be contiguous with the region that hybridized to the tcmK and tcmL
probes, which was surprising since the PKS genes in the majority of the other gene clusters encoding type-II PKS occur in the same order as the tcmKLMN genes (Fig. 2A)
CH3CH2COSCoA +
9 CH2COSCoA
I
MC& OH 0 HO OH
cog Ring D cyclization product
a t H’, -HOH
COSEru
omCt& &C”a w%_
0 0 0 0 OH 0 HO o OH 0 HO o
Decaketide
MHW OH 0 HO 0
Aklaviketone
12-Deoxyaklanonic acid Aklanonic acid
!? HO 74W
I mH3 ,DP-L-;nosamine;
OH 0 HO OH
RHO IJ
TDP-D-glucose
Qy$$y ,, Oxidation, &G$O” CH30 0 HO 0
0
CH30 0 HO 0 0 0
CH3
HO icI
CH3
HO NH2 NH2
DNR DXR
Fig. 1. DXR biosynthetic pathway. Only key substrates and intermediates discussed in the text are shown. Thin and thick arrows indicate one or
more than one step, respectively. The ring D cyclization product produced from aklanonic acid by acid treatment is shown above the latter structure.

(Hopwood and Sherman, 1990; Katz and Donadio, 1993). Also surprising was the fact that the tcmM probe did not hybridize to any of the clones tested, although this could have been due to a greater degree of divergence between tcmM and its dps homolog.
(b) Sequence analysis of the dps genes Analysis of the 8.7 kb of DNA sequenced from the
~crn~~~-hyb~dizing region revealed the presence of nine complete ORFs (Figs. 2B and 3), many of which appear to encode proteins whose deduced sequence is similar to known PKS enzymes (Table I). These genes seem to be part of at least four transcription units on the basis of their relative orientation (Fig. 2B). Reading left to right, a transcription unit comprising three ORFs ends in dnrH
(the leftmost gene in the sequenced DNA). dnrH is a homolog of the ACT UC~~Z~ ketoreductase gene (Hallam et al., 1988) and presumably encodes one of the two keto- reductases that act on the 9 and 7 positions of the decake- tide (or a product of its cyclization) and aklaviketone, respectively, in the biosynthesis of RHO (Fig. 1).
The second putative operon, containing dpsF down- stream from dpsE, converges on the operon containing dnrH. The deduced DpsF protein is similar to TcmN and other polyketide cyclases, such as the ACT ActVII cyclase (Fernandez-Moreno et al., 1992), so it is probably involved in one or more of the early cyclization steps in aklanonic acid production. The location of dpsf; is unusual compared to most other gene clusters encoding type-II PKS, since these genes are normally downstream from the genes encoding a 3-oxoacyl : ACP synthase and an ACP (Hopwood and Sherman, 1990, Katz and Donadio, 1993). Although another polyketide cyclase in addition to DpsF is likely to be required for the biosyn- thesis of 12-deoxyaklanonic acid (Fig. l), the sequence data do not identify a dps homolog of actW, tcml or tcmJ, genes that perform such a function in other systems (Hopwood and Sherman, 1990; Katz and Donadio, 1993; Summers et al., 1993). Like dnrH, dpsE encodes an Act111 homolog, which could function in one of the two reduc- tion steps that occur during RHO formation (Fig. 1). The DpsE protein has a closer similarity to Act111 than DnrH does (Table I) and is located in the same relative position as actlll has in relation to the genes encoding a 3-oxoacyl : ACP synthase (Fernandez-Moreno et al., 1992). These facts suggest that the DpsE and Act111 enzymes have the same function, leaving DnrH as a can- didate for catalyzing the reduction of aklaviketone to aklavinone (Strohl et al., 1989). However, this assump- tion is tempered by the organization of the frenolicin (Bibb et al., 1994) and griseusin (Yu et al., 1994) gene clusters whose actIIZ homologs are downstream from the genes encoding type-II 3-oxoacyl : ACP synthases and
ACPs, in front of putative polyketide cyclase-encoding
genes. The third putative operon contains several genes, start-
ing with URF8. This small ORF encodes a deduced pro- tein similar to the putative products of genes recently discovered in the aclacinomycin-producing S. galitaeus
(I. Fujii, personal communication) and the DNR-produc- ing Streptomyces sp. C5 strain. (Ye et al., 1994). All three of these genes have the same location relative to the homologous 3-oxoacyl : ACP synthase encoding genes. About a third of the deduced sequence of ORF8 is homo- logous to the TcmH monooxygenase of TCM C synthesis (Fig. 4A) (Summers et al., 1993). Since TcmH catalyzes the oxidation of the naphthacene Tcm Fl to the naphtha- cenequinone Tcm D3 (Shen and Hutchinson, 1993a), the putative Orf8 protein could have an analogous function in DXR biosynthesis: the oxidation of the as yet uniden- tified 12-deoxyaklanonic acid to aklanonic acid (Fig. 1). The next gene, dpsA, should encode the 3-oxoacyl: ACP synthase that catalyzes the formation of the decaketide backbone of RHO. This gene is highly conserved among the known gene clusters encoding type-II PKS (Hopwood and Sherman, 1990; Katz and Donadio, 1993). Since the predicted translational start of dpsA
overlaps the 0RF8 stop codon (Fig. 3), these two genes may be translationally coupled. This does not appear to be the case for dpsA and dpsB (unlike their homologs in other type-II PKS systems) because the ATG for the putative translational start of dpsB is three codons down- stream from the TGA stop codon of dpsA (Fig. 3). dpsB-
like genes are found in the same position in relation to many 3-oxoacyl : ACP synthase-encoding genes, and their products, which are very similar to DspA-like proteins but lack an identifiable active site with the critical Cys (Hopwood and Sherman, 1990; Katz and Donadio, 1993), are thought to form a complex with their respec- tive DpsA homologs and catalyze the condensation between the chain starter and extender units (Shen and Hutchinson, 1993~). McDaniel et al. (1993) have pro- posed that DpsB-like proteins determine the size of the resulting poly-&ketothioester intermediates. An ACP- encoding gene usually follows the dpsB homologs in clus- ters of genes encoding type-II PKS (Hopwood and Sherman, 1990; Katz and Donadio, 1993). However, in the present case the gene following dpsB appears to encode a protein of 309 to 353 aa, depending on the actual translational start codon, with a moderate resem- blance to FAS enzymes in S. glaucescens (FabB) (R. Summers, A.A., B. Shen, W. Wessel and C.R.H., unpub- lished results) (Table I) and E. coli (FabH) (Tsay et al., 1992) (Fig. 4B). Since dpsG, a homolog of the tcmM gene encoding an ACP (Shen et al., 1992), lies about 10 kb upstream from dpsC (Fig. 2B) (S.L. Otten, X.-C. Liu and

tunK tcmL tcmlw tcmff
m probes far Southern - anatysis d the cfps
w-
I A A I ORF7 ORF6
a dp$G
position d hybridizatkm to tcm PKS
ORFS characterized in this paper I
WI I- - /I'
92
pWHM75 i
pwHM7c3 I
pww.477
C
pwHM73
Fig. 2. Organization of the tcm and dps PKS genes. (A) The tcm PKS genes. The segments used for Southern analysis of the dps genes are shown below the four tcm ORFs. Methods: The LB (Sambrook et al. 1989), 2 x YT (Sambrook et al., 1989), SNA (Hopwood et al., 1986), seed (Guilfoile and Hutchinson, 19911, APM (Guilfoile and Hutchinson. 1991) and GPS growth media (Dekleva et al., 1985) were as described, except that distiiled water instead of tap water was used in making the APM medium. ISP4 medium was obtained from Difco Laboratories (Chicago, IL, USA). R2YE medium was the same as RZYE of Hopwood et al. (1985), except that 10 ml of 1 M NaOH instead of 5 mi was added per liter of medium. The S. peucetius ATCC 29050 wild-type strain was obtained from the American Type Culture Collection and used to construct the ddpsB mutant strain. S. liuidans TK24 was obtained from David Hopwood at the John Innes Institute (Norwich, UK). The E. coli strains DH5 and JM105 were as described by Sambrook et al. (1989). S. peucetius strains were grown on ISP4 (Difco, Detroit, MI, USA) plates for routine culturing, or from spore stocks in seed medium for 1 to 2 days, after which the cultures were transfered to the APM or GPS media and grown for 3 days in baffled Erlenmeyer flasks at 300 rpm and 30°C before metabolite extraction. S. hidans was grown on RZYE agar for propagation of spores or in liquid R2YE medium for preparation of mycelium for protoplast formation. p~EM7~(~) was obtained from Promega (Madison, WI, USA), pWHM3 was from Vara et al. (1989), pWHM601 was from Guilfoile and Hutchinson (1991), pDH5 was from Hillemann et al. (1991) and pGM160 was from Muth et al. (1989). Plasmids were maintained in Streptomyces transformants by selection with Th (5 pg/ml) or Am (10 pg/ml) in R2YE liquid media, after protoplast- mediated transformation as described by Hopwood et al. (1985) and Stutzman-Engwall and Hutchinson (1989). E. coli DH5 and JM10.5 were used for routine cloning and DNA sequencing, respectively, as described by Sambrook et al. (1989). (B) Organization of the dps genes (ORF6 and ORF7 are not discussed in the text). The regions that hybridize to specific tcm gene probes are indicated beneath the restriction map that shows only the sites of interest. The boundaries of the pWHM75-77 clones are indicated beneath the dps ORFs; the dashed line at the left hand end of pWHM77 indicates that the insert extends beyond the region shown, to near the 3’-end of dnrf. Southern blot hyb~d~zations were done at 40-65% stringency as defined by Hopwood et al. (1985) using the following 32P-labeled probes prepared by nick translation according to Sambrook et al. (1989): 0.8-kb StuI-ScaI fragment from tcmK, 0.65”kb NruI-MluI fragment from tcmL, 0.3-kb MroI-SstI fragment from tcmM and 0.6-kb SstI fragment from tcmN that contains the cyclase and part of the 0-MTase domain. Conditions of prehybridization and hybridization were as described by the manufacturer of the Zeta-Probe membranes (Bio-Rad, Richmond, CA, USA). Aklanonic acid was isolated as follows. The Streptomyces sp. C5 dauC mutant that accumuiates aklanonic acid (Bartel et al., 1990) and S. hiduns transformants containing the dps genes as pWHM75, pWHM76 and pWHM77 were grown in 1 litre of GPS medium at 300 rpm for 4 days at 30°C. Anthracycline metabolites were isolated from the culture supernatants by extraction with chloroform (1.5 1) after adjusting the pH to 1.5 with HCI. The crude mixture of metabolites resulting from evaporation of the solvent was applied to a potassium dihydrogen phosphate-buffered silica gel column prepared according to the protocol of Eckardt et al. (1985). The yellow band was eluted with chloroform/methanol (10: 1, v/v). Aklanonic acid from the Streptomyces sp. CS da& mutant strain could be purified in this way, but the aklanonic acid isolated from S. lividans could not be completely separated from two other yellow compounds. Therefore, this material was further

C.R.H., unpublished data), the &SC product is not likely to function as an ACP. Instead, it may play a role in
decaketide formation that possibly is analogous to the E.
coli FabH enzyme, which catalyzes the formation of the first 4-carbon intermediate of fatty acid biosynthesis, acetoacetyls-ACP, from acetyl-CoA and malonylS-ACP (Jackowski and Rock, 1987). This idea is tenuous, how- ever, in view of the limited sequence similarity between DpsC and FabH. The deduced product of the down- stream gene, dpsD, is very similar to acyltransferases like the propionyl-CoA acyltransferase domain of the DEBSl subunit of 6-deoxyerythronolide B synthase from Saccharapolyspora erythraea (Cortes et al., 1990; Donadio et al., 1991) and the malonyl-CoA acyltransferases of FAS from E. coli (Magnuson et al., 1992) and S. glaucescens (R. Summers, A.A., B. Shen, W. Wessel and C.R.H., unpublished results) (Fig. 4C). DpsD contains the aa sequence 98LGHSVGE’04, corresponding to the highly conserved active site region in acyltransferases (see, e.g., Schweizer et al., 1987). The central S”’ residue is thought to be the substrate attachment site in related acyltransfer- ases of FAS from bacteria, yeast and animals (Wakil et al., 1983). Thus, DpsD could be a propionyl-CoA acyltrans- ferase. An enzyme of this type has not been found in PKS that govern the formation of other aromatic polyketides, such as ACT or TCM C, suggesting that dpsD could
5
perform a function unique to DXR biosynthesis. Since the starter unit for formation of the decaketide precursor
of RHO is propionate, instead of acetate as in TCM C and ACT biosynthesis, it is sensible to propose that DpsD catalyzes the transfer of this 3-carbon starter unit to the DpsA subunit of the 3-oxoacyl:ACP synthase complex involved in decaketide formation. Alternatively, DpsC instead may catalyze assembly of the first 5-carbon inter- mediate, 3-oxopentanoylS-ACP, using propionyl-CoA and malonylS-ACP, leaving DpsD for some other role in DXR biosynthesis.
The final gene, ORF5, should be transcribed con- vergently to the putative ORF&dpsD operon. Several other DXR biosynthesis genes are upstream from ORF.5
(Madduri et al., 1993; KM. and S.L. Otten and C.R.H., unpublished work) but its function is unknown. By TFASTA analysis (Devereux et al., 1984) the deduced 0RF5 protein exhibits significant sequence similarity with the products of gerC2, a regulator of glucosyltrans- ferase activity in Lactococcus lactis (Henner et al., 1985) and of the 0RF7 gene of Tn4556 from S. fradiae
(Siemieniak et al., 1990) (data not shown). Although an 0RF5 homolog located at the same relative position in the DNR-producing Streptomyces C5 sp. encodes a pro- tein proposed to be similar to several MTases (Ye et al., 1994), we found only very low, and presumably insignifi-
an analytical reverse-phase HPLC column using a solvent consisting of 50% methanol in water (pH adjusted to 2.0 with concentrated phosphoric acid)
at flow rate of 3 ml/min. The fractions containing aklanonic acid were combined and the solvent was evaporated at 37°C. The yellow material extracted
with chloroform from the syrupy residue was analyzed by high resolution electron impact mass spectrometry to identify aklanonic acid and its aromatic
ring D cyclization product. (C) Construction of the AdpsB mutant. An in-frame deletion in dpsB was made by digesting a plasmid containing the dpsB
and dpsC genes in a 2.5-kb BamHI fragment (Fig. 2B) with SstII, treating the linearized DNA with T4 DNA polymerase to remove the 3’ overhangs,
and isolating the resulting fragment from an agarose gel with a DEAE membrane as described in Guilfoile and Hutchinson (1991). The DNA was
religated to give pWHM70 and the deletion was confirmed by sequencing. A 0.8-kb BamHI-SphI fragment from pWHM70 was moved into a pDH5
clone containing an adjacent 0.6-kb BamHI-SstI segment from dpsA to make pWHM71 so that the resulting AdpsB clone had 0.6-0.7 kb of flanking
sequence on either side of the deletion. A 1.4-kb EcoRI-Hind111 segment of pWHM71 was moved into pGEM7zf(+) at the corresponding sites to make
pWHM72. The resulting 1.4-kb segment was removed from pWHM72 by HindIII+ XbaI digestion and cloned between the Hind111 and XbaI sites of
the temperature-sensitve pGM160, replacing the gentamycin resistance gene and creating pWHM73 (Fig. 2C). pWHM73 was introduced into S.
peucetius 29050 by transformation with selection for Th resistance on R2YE plates, and the transformants were grown at 3O’C for a week, then
transferred to 39’C. After incubation for a further week, the colonies that had continued to grow were patched onto ISP4 plates containing Th (25 kg/ml)
and incubated at 39°C. After sporulation, the patches were streaked on ISP4 plates that lacked antibiotic and grown at 39°C to sporulation, then this
process was repeated once. Loss of the integrated vector during growth at 39°C was expected to result in Th sensitivity, but all of the colonies tested at
this point were Th resistant. Therefore, three sporulated patches were used to make protoplasts and 250 colonies were regenerated from one of these
strains by growth on ISP4 plates at 39’C. After sporulation, 8 of these clones were found to be sensitive to Th. Chromosomal DNA from each of these
colonies was digested with BamHI and probed with a digoxigenin-labeled 0.36-kb SphI-Sal1 fragment from the dpsC gene, made with the GeniusTM kit
(Boerhinger-Mannheim, Indianapolis, IN, USA). Hybridization was carried out according to the manufacturer’s instructions with DNA immobilized
on Hybond N membranes (Amersham, Chicago, IL, USA). One of the 8 clones, named WMH1535, showed the 1.3-kb band expected for the AdpsB
deletion and the others had a 2.5-kb band indicative of the wild-type genotype (data not shown). Complementation of the AdpsB mutation was carried
out as follows. A 1.3-kb XhoI-XmaI fragment containing the dpsB gene was cloned into pGEM7zf(+) to create pWHM78. This plasmid was digested
with EcoRI and HindIII, and the resulting fragment was gel purified with a DEAE membrane (Guilfoile and Hutchinson, 1991) and cloned into the
EcoRI-Hind111 sites of pWHM3 to make pWHM79 with dpsB oriented downstream from the lacZ promoter. pWHM79 was introduced into the
WMH1535 strain by transformation and the resulting Th-resistant colonies were patched onto ISP4 plates containing Th (25 pg/ml), sporulated and
used to start seed cultures. After two days of growth in 2.5 ml seed medium at 3o”C, the cultures were transferred to APM and GPS media, then grown
for 2-4 days at 3O’C. Complementation of the AdpsB mutation caused extracts of these cultures, prepared as described by Stutzman-Engwall and
Hutchinson (1989), to have an orange-red color indicating the presence of DXR or its intermediates. TLC analysis of these extracts was done as
described by Stutzman-Engwall and Hutchinson (1989), except that the solvent system was chloroform/methanol/acetic acid/water (15 : 5 : 0.3 : 1, v/v).
Complementation analysis was carried out with the group-I cosmid pWHM501 (Otten et al., 1990), group II cosmid pWHM313 (Stutzman-Engwall
and Hutchinson, 1989) and group III cosmid pWHM507 (Otten et al. 1990) in a similar manner, using Am (25 Kg/ml) instead of Th selection.

6
1 42 1%
% 142 45%
5251 cJzcmxxwvmB_ ~ACQXXGAl'ZGC0X!XACXXX~~~~ 294 LARAV ETALADAGLDGSD IAVVFADGAAV P ELDAAEAEALASVFG PRRVP
%" cxxcsxnw ~~~~~~~~~~~ A c
VTVPKTLTGRLYSGAG PL DVATmAbLALRD E D L P L DVVT ~5~~~~~A~~~~
GTVLSGRPGR' GAmma
8731 Mczmao3mmozh 7 KY S S DQM ?%%===A '.13'
Fig. 3. The nt sequences of the S.Fkb region shown in Fig. 2B that contains all of the dps genes, except dpsG, and deduced aa sequences. Sequencing was done as described in Summers et al. (1992). The dps ORFs and their products were identified by analysis of the sequence data with the Genetic Computer Group program CODON PREFERENCE (Devereux et al., 1984) and were further analyzed using the TRANSLATE and GAP programs (Devereux et al., 1984). The codon usage pattern of the S-end of ORF.5, which appears to extend approx. 80 nt beyond the EcoRI site of Fig. 2B. is unusual and may indicate that ORE3 is considerably shorter than indicated. Putative ribosomal-banding sites are underlined for some ORFs and translational ston codons are indicated bv asterisks (*). The seauence data have been deposited with GenBank under accession No. L35560.
7
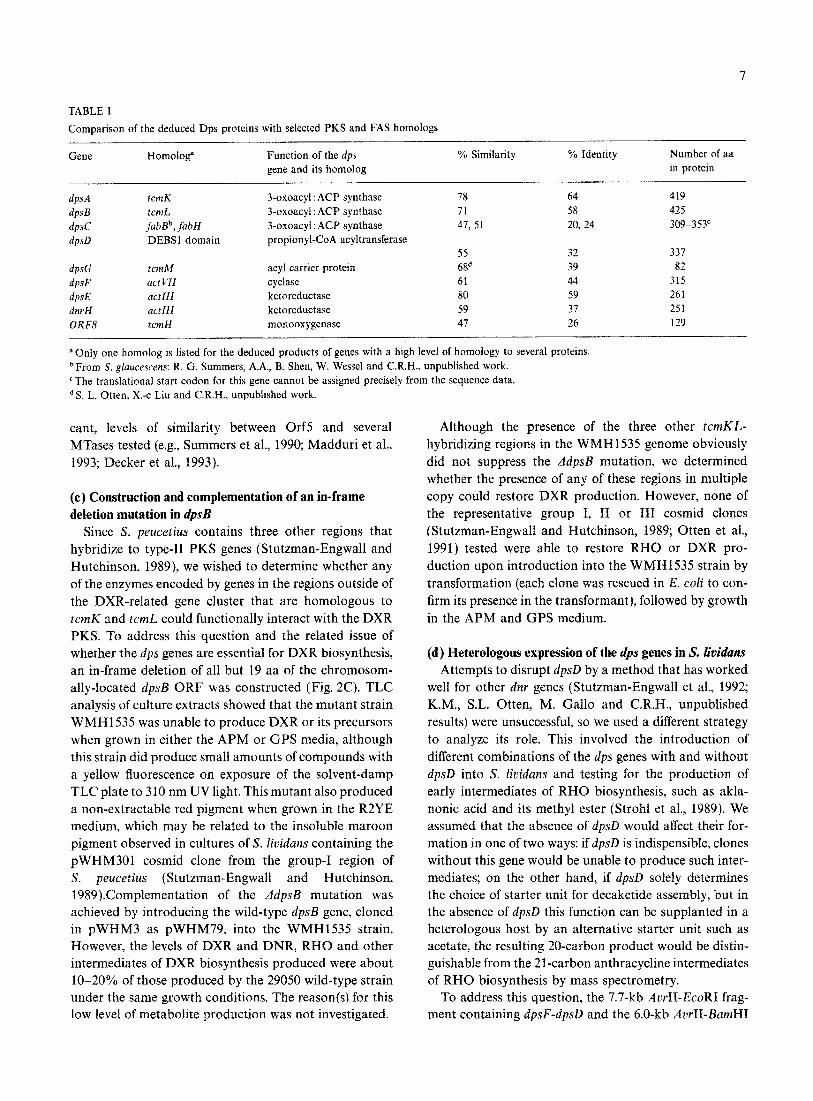
TABLE 1
Comparison of the deduced Dps proteins with selected PKS and FAS homologs
Gene Homolog” Function of the dps % Similarity % Identity gene and its homolog
dpsA tcmK 3-oxoacyl : ACP synthase 78 64
ilpSB tcmL 3-oxoacyl : ACP synthase 71 58
dpsC fablP, fubH 3-oxoacyl : ACP synthase 47, 51 20, 24
dpsD DEBSl domain propionyl-CoA acyltransferase 55 32
dpsG rcmM acyl carrier protein 68d 39
dpsF UCt Vfi cyclase 61 44
dpsE actlll ketoreductase 80 59
dnrH act111 ketoreductase 59 37
ORF8 tcmH monooxygenase 47 26
aOnly one homolog is listed for the deduced products of genes with a high level of homology to several proteins. b From S. glaucescens: R. G. Summers, A.A., B. Shen, W. Wessel and C.R.H., unpublished work. ‘The translational start codon for this gene cannot be assigned precisely from the sequence data.
Number of aa in protein
419 425 309-353°
337 82
315 261 251 129
d S. L. Otten, X.-c Liu and C.R.H., unpubIished work.
cant, levels of similarity between Orf5 and several MTases tested (e.g., Summers et al., 1990; Madduri et al., 1993; Decker et al., 1993).
(c) Construction and complementation of an in-frame deletion mutation in dpsB
Since S. peucetius contains three other regions that hybridize to type-11 PKS genes (Stutzman-Engwall and Hutchinson, 1989), we wished to determine whether any of the enzymes encoded by genes in the regions outside of the DXR-related gene cluster that are homologous to tcmK and tcmL could functionally interact with the DXR PKS. To address this question and the related issue of whether the dps genes are essential for DXR biosynthesis, an in-frame deletion of all but 19 aa of the chromosom- ally-located dpsB ORF was constructed (Fig. 2C). TLC analysis of culture extracts showed that the mutant strain WMH1535 was unable to produce DXR or its precursors when grown in either the APM or GPS media, although this strain did produce small amounts of compounds with a yellow fluorescence on exposure of the solvent-damp TLC plate to 310 nm UV light. This mutant also produced a non-extractable red pigment when grown in the R2YE medium, which may be related to the insoluble maroon pigment observed in cultures of S. Eividans containing the pWHM301 cosmid clone from the group-I region of S. ~e~ce~j~s (Stutzman-Engwall and Hutchinson, 1989).Complementation of the ddpsB mutation was achieved by introducing the wild-type dpsB gene, cloned in pWHM3 as pWHM79, into the WMH1535 strain. However, the levels of DXR and DNR, RHO and other intermediates of DXR biosynthesis produced were about lo-20% of those produced by the 29050 wild-type strain under the same growth conditions. The reason(s) for this low level of metabolite production was not investigated.
Although the presence of the three other tcmKL-
hybridizing regions in the WMH1535 genome obviously did not suppress the AdpsB mutation, we determined whether the presence of any of these regions in multiple copy could restore DXR production. However, none of the representative group I, II or III cosmid clones (Stutzman-Engwall and Hutchinson, 1989; Otten et al., 1991) tested were able to restore RHO or DXR pro- duction upon introduction into the WMH1535 strain by transformation (each clone was rescued in E. coEi to con- firm its presence in the transformant), followed by growth in the APM and GPS medium.
(d) Reterologous expression of the dps genes in S. lividans Attempts to disrupt dpsD by a method that has worked
well for other dnr genes (Stutzman-Engwall et al., 1992; K.M., S.L. Otten, M. Gallo and C.R.H., unpublished results) were unsuccessful, so we used a different strategy to analyze its role. This involved the introduction of different combinations of the dps genes with and without dpsD into S. tividans and testing for the production of early intermediates of RHO biosynthesis, such as akla- nonic acid and its methyl ester (Strohl et al., 1989). We assumed that the absence of dpsD would affect their for- mation in one of two ways: if dpsD is indispensible, clones without this gene would be unable to produce such inter- mediates; on the other hand, if dpsD solely determines the choice of starter unit for decaketide assembly, but in the absence of dpsD this function can be supplanted in a heterologous host by an alternative starter unit such as acetate, the resulting 20-carbon product would be distin- guishable from the %l-carbon anthracycline intermediates of RHO biosynthesis by mass spectrometry.
To address this question, the 7.7-kb AurII-EcoRI frag- ment containing dpsF-dpsD and the 6.0-kb AvrII-BamHI

50 100
/
(
Orf8
/’
/
/.
/ . I *
- 100
- 50
( ,.o
300 ,
/ - 3oo
,’
- 200
/
/’
/’ 100
c DpsD Debsl SgFabA Consens
DpsD grL...rAdW IsdhpAVp‘iD hVtrsQPLLF AiidyaLgrMv mAttgrPwA1 Debsl ipFlraeAAR reqG?AlstE rVdwQPvMF AVmvsLaSMw rAhgVePaAV SgFabA ahYgtkadAd eirGtAVa.. . . . ..QPLLv Aa..gLlSaa algd:aPgAV consens ___-_.._k\_ ___D_AV___ _V___QPLLF A"___L_SM_ _A____P_AV
DpsD 1GHSVGEmAA AaLAGVFtLr DAAgLlLdRv RrLAqAPp.. ..g.GMlA'JA Debsl iGHSqGElAA AcvAGaLsLD DAArwalRs RviAtmPg.. ..nkGMasIA SgFabA aGHSVGEItA ArFAGV..LD DtAaLtLvRk RgLIunAeaaa itetGElsAl1 Consens m=EIAA A--AGV--LD DAR-L-L-R- R-LA-AP--- ----GM-A-A
DpsD AaEdv?gpyi gnGEvVvVAri VNsPRQti1,A GpvasLAgva ralrEag?tc Debsl ApagEVrari ..GDrVeIAA VNGPRsWvA GdsDeLdrLv asctteclra SgFabA ggDpEVsvah 1eklgltpAn VNGagQIVa A GtlEqLAaLn edkpEgvrki Consens A---ET&-- -_-G--V--AA V)jGPRQ_"-_A G____LA_L_ ____E_____
DpsD revpsltAFii TpmLAPIaag aAErFAst.. rvaPsIPLrS gyrpGavTgq Debsl krLas'dyAsH SsHVetIrDa LhaeLgeDFh PlpgfVPFfS tv.CGRwTqp SgFabA vpik','agAFt? Th~MAPa~Dk LAEa.ArELs PadPeVPYvS ".kdGRtvaT COnSeWS --L-';__AFx T-H-API-D- LAE__$,____ F__P_"P__S ___~G&'__
DpsD 1aaDpsYWaG hpleTVRFwp ALDAM..LGs RT........ . . ..gVvcLe Debsl dElDagYWyr nlrrTVRFad AvrALaEqGy RTFLEVsahp iltaaIeeig SgFabA gDevvarlvG qvanpVRWd1 cMEtFkELGv taLLEVcp.. ..ggtltgLa ConSenS ---D__Y,&G --__TVRF-- A__A__ELG_ RT_LE\'____ ___~_~--~_
@SD tGpGqGLST1 ArrhPtvRfG knSvv....p LLppQAgtae iiDRtSVaaa1 Debsl dGsGadLSa1 . . ..hslRrG dgSlaDfgea LsrafAagva VDWeSVhlgr. SgFabA kralpGvkT1 AlktP..... ..ddlDaare LFaeQA... . . . . . . . . ConSenS -G-G_GLSTI A-_-P--R-G _-S--D-___ L___Qj,--a_ ,JD-es”--_-
DpsD Debsl SgFabA Consens
1 50
mdapailLmF PGQGsQhArM AVGLygsePa FtAAMDEVED AYDDAgDsEg . VFVF PGQGwQwAgM AVdLLdtsPv FaAALrEcAD AlEphlDFEv
..vrvlVLVa PGQGaQ.... tpGFLtpwle LpgAaDrVA. AWsDAigLDl --mm--VLVF PGQG-Q-A-M AVGLL---P- F-AA-DEVAD A--DA-D-E-
100
259
367 aAvReqgtv1 sgrpgr gArR...... .r..... . . . . . . . . . . . . . . . . _A-R_____- ____---
Fig. 4. Comparisons of deduced protein sequences. (A) Sequence comparison of the S. glaucescens TcmH and OrfS proteins by COMPARE and DOTPLOT analysis (Devereux et al., 1984) using a window of 30 and a stringency of 1.5. (B) Sequence comparison of the E. coli FabH and DpsC proteins by COMPARE and DOTPLOT analysis (Devereux et al., 1984) using a window of 30 and a stringency of 15. (C) Sequence comparison of DpsD with aa 104881295 of the propionyl-boa acyltransferase domain (Donadio and Katz, 1992) of the S. erythrnea DEB.81 protein (Debsl) and S. giaucescens FabA (SgFabA) by PILEUP analysis (Devereux et al., 1984) using the default settings. The ‘consens’ line shows a capital letter wherever two or more identical residues are in the vertical column. The conserved active site GHSXG region is underlined.
fragment containing dpsF-dpsC were cloned in pWHM3 tance, and these strains were tested for metabolite pro-
separately to yield pWHM75 and pWHM76, respectively duction as before. Both strains were able to make a bright (Fig. 2B). No new metabolites were observed when S. yellow material, shown to be the same in each case by
lividans transformants carrying either of these plasmids TLC analysis. This material was not produced when S.
were grown in the GPS medium and culture extracts were lividans contained only pWHM75 and pWHM601, as its
analyzed by TLC. Since neither of these plasmids con- production depended on genes contained in pWHM77,
tains the dpsG gene that encodes an ACP (Fig. 2B), the presumably including dpsG.
lack of new metabolite production could have been due When the yellow material was purified by column
to its absence. Therefore, we cloned in pWHM601 a chromatography and analyzed by HPLC, it was found
10.3-kb BglII-AorII fragment (Fig. 2B) containing dpsG to consist of three components, The major compound
plus the dnrl transcriptional regulator (Stutzman- had the same retention time on HPLC analysis as authen-
Engwail et al., 1992; KM. and C.R.H., data not shown) tic aklanonic acid isolated from the S~~ep~~~_vces C.5 and six other genes to yield pWHM77. Plasmid dauC mutant (Bartel et al., 1990). The acidic conditions
pWHM77 was introduced along with either pWHM75 produced during evaporation of the chromatographic (with dpsD) or pWHM76 (without dpsl)) into S. lividans solvent from the HPLC column fractions apparently
by transformation with selection for Th and Am resis- resulted in the conversion of this aklanonic acid to the

9
aromatic tetracycle shown in Fig. 1 by an intramolecular aldol reaction analogous to the conversion of Tcm F2 to Tcm Fl in S. glaucescens (Shen and Hutchinson, 1993b). The structure of this compound is based on the fact that its high resolution electron impact mass spectrogram exhibited a base peak at m/z 334.082 (C,,Hr,O,, calcd. 334.084), due to the loss of CO, from the parent ion, instead of the ions at m/z 352.096 (CZoH,,06) and 337.076 (C,gH,,Os) produced by aklanonic acid (Eckardt et al., 1985). Both of these compounds were converted to a methyl ester upon treatment with CH2N,.
These data are consistent with the formation of akla- nonic acid from the normal propionate starter unit in the two types of S. lividans transformants. Consequently, the dps genes in pWHM75 or pWHM76 plus pWHM77 are sufficient to produce this intermediate of DXR biosynthe- sis (Eckardt et al., 1985) but not its methyl ester, which can be cyclized to aklaviketone in vitro (Connors et al., 1990). Since these data also show that dpsD is not essen- tial for the biosynthesis of aklanonic acid, either a gene provided by S. lividans is able to compensate for the absence of dpsD or this gene has no role in aklanonic acid biosynthesis,
(e) Conclusions
(I) The organization of the dps genes is significantly different from the other clusters of bacterial genes encod- ing type-II PKS characterized to date because the dpsG and dpsF genes encoding an ACP and a polyketide cyclase, respectively, have unusual locations relative to the dpsA and dpsB genes, which specify the putative con- densing enzyme subunits, and the dpsE ketoreductase gene.
(2) A known intermediate of DXR biosynthesis, akla- nonic acid, is formed in S. lividans containing the dps genes we designate as forming the DXR PKS (dpsA-dpsF and dpsG) plus seven other genes in pWHM77 (whose functions are not specified here, except for dnrH, but on the basis of sequence analysis and gene disruption and expression experiments are largely known not to produce PKS-like enzymes; Stutzman-Engwall et al., 1992; K.M. and C.R.H., data not shown; S.L. Otten, X.-C. Liu, C. Scotti and C.R.H., unpublished results).
(3) The unique dpsC and dpsD genes downstream from dpsAB encode enzymes that may specify the use of propionate as the starter unit for assembly of the DXR decaketide. Nonetheless, since the putative DpsD acyltransferase seems not to be essential for this process in S. lividans, we emphasize that inclusion of dpsC and dpsD as part of the DXR PKS genes is quite tentative.
NOTE ADDED IN PROOF
Introduction of the dpsG gene cloned in pWHM3 into a S. liuidans (pWHM75) transformant resulted in the pro-
duction of aklanonic acid, confirming that the DXR PKS
consists of only the dpsA-dpsF and dpsG genes.
ACKNOWLEDGEMENTS
We thank William R. Strohl for providing the da& mutant and for a preprint of the paper by Ye et al. We also thank Dr. Ben Shen for assistance in the isolation of aklanonic acid and its mass spectroscopic analysis. This work was supported by grants from the American Cancer Society (CH-524), a traineeship to A.G. from the National Institutes of Health (TG32 GM07133) and a fel- lowship to A.A. from the Department of Biotechnology, Government of India.
REFERENCES
Bartel, P.L., Connors, NC. and Strohl, W.R.: Biosynthesis of anthracy-
clines: analysis of mutants of Srreptomyces sp. strain C5 blocked in
daunomycin biosynthesis. J. Gen. Microbial. 136 (1990) 187771886.
Bibb, M.J., Biro, S., Motamedi, H., Collins, J.F. and Hutchinson, C.R.:
Analysis of the nucleotide sequence of the S. glaucescens tcml genes
provide key information about the enzymology of polyketide anti-
biotic biosynthesis. EMBO J. 8 (1989) 2727-2736.
Bibb, M.J., Findlay, P.R. and Johnson, M.W.: The relationship between
base composition and codon usage in bacterial genes and its use for
simple and reliable identification of protein-coding sequences. Gene
30 (1984) 157-166.
Bibb, M.J., Sherman, D.H., Omura, S. and Hopwood, D.H.: Cloning,
sequencing and deduced functions of a cluster of Streptomyces genes
encoding the biosynthesis of the polyketide antibiotic frenolicin.
Gene 142 (1994) 31-39.
Connors, N.C., Bartel, P.L. and Strohl, W.R.: Biosymthesis of anthracy-
clines: Enzymatic conversion of aklanonic acid to aklavinone and
s-rhodomycinone by anthracycline-producing streptomycetes.
J. Gen. Microbial. 136 (1990) 188771894.
Cortes, J., Haydock, S.H., Roberts, G.A., Bevitt, D.J. and Leadlay, P.F.:
An unusually large multifunctional polypeptide in the erythromycin-
producing polyketide synthase of Saccharopolyspora erythraea.
Nature 348 (1990) 176-178.
Decker, H., Motamedi, H. and Hutchinson, CR.: Nucleotide sequences
and heterologous expression of tcmC and tcmP, biosynthetic genes
for TCM C in Streptomyces glaucescens. J. Bacterial. 175 (1993)
3876-3886.
Decker, H. and Hutchinson, C.R.: Transcriptional analysis of the
Streptomyces glaucescens tetracenomycin C biosynthesis gene clus-
ter. J. Bacterial. 175 (1993) 3887-3892.
Devereux, J. Haeberli, P. and Smithies, 0.: A comprehensive set of
sequence analysis programs for the VAX. Nucleic Acids Res. 12
(1984) 3877395.
Dekleva, M.L., Titus, J.A. and Strohl, W.R.: Nutrient effects on anthra-
cycline production by Streptomyces peucetius in a defined medium.
Can. J. Microbial. 31 (1985) 287-294.
Donadio, S., Staver, M.J., McAlpine, J.B., Swanson, S.J. and Katz, L.:
Modular organization of genes required for complex polyketide bio-
synthesis. Science 252 (1991) 6755679.
Donadio, S. and Katz, L.: Organization of the enzymatic domains in
the multifunctional polyketide synthase involved in erythromycin
formation in Saccharopolyspora erythraea. Gene 111 (1992) 51-60.
Eckardt, K., Tresselt, D., Schumann, G., Ihn, W. and Wagner, C.:
Isolation and chemical structure of aklanonic acid, an early interme-
diate in the biosynthesis of anthracyclines. J. Antibiotics 38 (1985)
1034-1039.

10
Fernandez-Moreno, M.A., Caballero, J.L., Hopwood, D.A. and Malpartida, F.: The act cluster contains regulatory and antibiotic export genes, direct targets for translational control by b&l tRNA gene of Streptomyces. Cell 66 (1991) 769-780.
Fernandez-Moreno, M.A., Martinez, E., Boto, L., Hopwood, D.A. and Malpartida, F.: Nucleotide sequence and deduced functions of a set of cotranscribed genes of Streptomyces coelicolor A3(2) including the polyketide synthase for the antibiotic ACT. J. Biol. Chem. 267 (1992) 19278819290.
Gramajo, H., Takano, E. and Bibb, M.J.: Stationary-phase production of the antibiotic actinorhodin in Streptomyces coeIicolor A3(2) is transcriptionally regulated. Mol. Microbial. 7 (1993) 837-845.
Guilfoile, P.G. and Hutchinson, CR.: A bacterial analog of the mdr
gene of mammalian tumor cells is present in Streptomyces peucetius. the producer of daunorubicin and doxorubicin. Proc. Natl. Acad. Sci. USA 88 (1991) 8553-8557.
Guilfoile. P.G. and Hutchinson, C.R.: The Streptomyces glaucescens
tcmR protein represses transcription of the divergently oriented tcmR and tcmA genes by binding to an intergenic operator region. J. Bacterial. 174 (1992) 3659-3666.
Hallam, SE., Malpartida, F. and Hopwood, D.A.: Nucleotide sequence. transcription and deduced function of a gene involved in polyketide antibiotic synthesis in Streptomyees coe~ica~ar. Gene 74 (1988) 305-320.
Henner, D.J., Band, L. and Shimotsu, H.: Nucleotide sequence of the Bacillus subtilis tryptophan operon. Gene 34 (1985) 169-177.
Hillemann, D., Puhler, A. and Wohllenben, W.: Gene disruption and gene replacement in Streptomyces via single stranded DNA trans- formation of integration vectors. Nucleic Acids Res. 19 (1991) 727-731.
Hopwood, D.A., Bibb, M.J., Chater, K.F., Kieser, T., Bruton, C.J., Kieser, H.M., Lydiate, D.J., Smith, C.P., Ward. J.M. and Schrempf, H.: Genetic Manipulation of Streptomyces, A Laboratory Manual, The John Innes Foundation, Norwich, 1985.
Hopwood, D.A. and Sherman, D.H.: Molecular genetics of polyketides and its comparison to fatty acid biosynthesis. Annu. Rev. Genet. 24 (1990) 37-66.
Jackowski, S. and Rock, CO.: Acetoacetyl-acyl carrier protein synthase, a potential regulator of fatty acid biosynthesis in bacteria. J. Biol. Chem. 262 (1987) 7927-7931.
Katz. L. and Donadio, S.: Polyketide synthesis: prospects for hybrid antibiotics. Annu. Rev. Mi~robiol. 47 (1993) 875-P 12.
Madduri, K., Torti, F., Colombo, A.L. and Hutchinson, C.R.: Cloning and sequencing of a gene encoding carminomycin 4-O-methyl- transferase from Streptomyces peucetius and its expression in Escherichia cob. J. Bacterial. 175 ( 1993) 3900-3904.
Magnuson, K., Oh, W., Larson, T.J. and Cronan Jr., J.E.: Cloning and nucleotide sequence of the _fabD gene encoding malonyl coenzyme A-acyl carrier protein transacyiase of Esc~ler~c~~a cofi. FEBS L&t. 299 ( 1992) 262-266.
Magnuson, K., Jackowski, S., Rock, C.O. and Cronan Jr., J.E.: Regulation of fatty acid biosynthesis in Escherichia coli. Microbial. Rev. 57 (1993) 5222542.
McDaniel, R., Ebert-Khosla, S.. Hopwood, D.A. and Khosla, C.: Engineered biosynthesis of novel polyketides. Science 262 (1993) 15461550.
Muth, G., Nussbaumer, B., Wohlleben, W. and Puhler, A.: A vector system with temperature-sensitive replication for gene disruption and mutational cloning in streptomyces. Mol. Gen. Genet. 219 (1989) 341-348.
Otten, SL, Stutzman-Engwall, K.J. and Hutchinson, CR.: Cloning and expression of dauno~bicin biosynthesis genes from Stre~tom~~ces peucetius and S. pence&s subsp. caesius. J. Bacterial. 172 (1990) 3427-3434.
Sambrook, J., Fritsch, E.F. and Maniatis, T.: Molecular Cloning. A Laboratory Manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989.
Schweizer, M., Roberts, L.M., Holtke, H.-J., Takabayashi, K., Hollerer. E., Hoffmann, B., Muller, G., Kottig, H. and Schweizer, E.: The pentafunctional FASl gene of yeast: its nucleotide sequence and order of the catalytic domains, Mol. Gen. Genet. 203 (1987) 479-486.
Siemieniak, D.R., Slightom, J.L. and Chung, S.-T.: Nuclcotide sequence of Streptomyces fradiae transposable element Tn4.556: a class-II transposon related to Tn3. Gene 86 (1990) l-9.
Shen, B., Summers, R.G., Gramajo, H., Bibb, M.J. and Hutchinson, C.R.: Purification and characterization of the acyl carrier protein of the Streptomyces glaucescens tetracenomycin C polyketide syn- thase. J. Bacterial. 174 (1992) 3818-3821.
Shen, B. and Hutchinson, CR.: Tetracenomy~in Fl monooxygenase: oxidation of a naphthacenone to a naphthacenequinone in the bio- synthesis of tetracenomycin C in Streptomyces glaucescens.
Biochemistry 32 (1993a) 66566663. Shen, B. and Hutchinson, C.R.: Tetracenomycin F2 cyclase: intramolec-
ular condensation in the biosynthesis of tetracenomycin C in Stre~tomyces g~au~esceF~s. Biochemistry 32 (1993b) 11149-I 1154.
Shen, B. and Hutchinson, CR.: Enzymatic synthesis of a bacterial polyketide from acetyl and malonyl coenzyme A. Science 262 (1993~) 1535-1540.
Strohl, W.R., Bartel, P.L., Conner% NC., Zhu. C.-B., Dosch, D.C.. Beale Jr., J.M.. Floss, H.G., Stutzman-Engwall, K., Otten, S.L. and Hutchinson. C.R.: Biosynthesis of natural and hybrid polyketides by anthracycline-produ~ng streptomyces. In: Hershberger. CL.. Queener, S.W. and Hegeman, G. (Eds.), Genetics and Molecular Biology of Industrial Microorganisms. American Society for Microbiology. Washington, DC, 1989, pp. 68-84.
Stutzman-Engwall, K.J. and Hutchinson, CR.: Multigene families for anthracycline antibiotic production in Streptomyces peucetius. Proc. Natl. Acad. Sci. USA 86 (1989) 31353139.
Stutzman-Engwall, K.J., Otten, S.L. and Hutchinson, C.R.: Regulation of secondary metabolism in Streptomyces spp. and overproduction of daunorubicin in Streptomyces peucetius. J. Bacterial. 174 (1992) 144- 154.
Summers, R.G., Wendt-Pienkowski, E., Motamedi, H. and Hutchinson, CR.: Nucleotide sequence of the t~F?~~J-~c~J~ region of the tetracen- omycin C biosynthesis gene cfuster of Streptamy~~s gluucescens and evidence that the tcmN gene encodes a multifunctional cyclase- dehydratase-O-methyl transferase. J. Bacterial. 174 (1992) 1810-1820.
Tsay. J.-T., Oh, W., Larson, T.J., Jackowski, S. and Rock, CO.: Isolation and characterization of the B-ketoacyl-acyl carrier protein synthase III gene (f&H) from ~.~c~~r~e~j~ co&. J. Biol. Chem. 267 (1992) 6807-6814.
Vara, J., Lewandowska-Skarbek, M., Wang, Y.-G., Donadio, S. and Hutchinson, C.R.: Cloning of gene governing the deoxysugar por- tion of the erythromycin biosynthesis pathway in Saccharopolyspora erythraeu. J. Bacterial. 171 (1989) 5872-5881.
Wakil, S.J.. Stoops, J.K. and Joshi, V.C.: Fatty acid synthesis and its regulation. Annu. Rev. Biochem. 52 ( 1983) 537-579.
Ye, J., Dickens, M.L.. Plater, R., Li, Y., Lawrence, J. and Stroht, W.R.: Isolation and sequence analysis of polyketide sythase genes from the daunomycin-producing Srreptomyces sp. strain C5. J. Bacterial. 176 (1994) 6270-6280.
Yu, T.-W., Bibb, M.J., Revill, P.W. and Hopwood, D.H.: Cloning. sequencing, and analysis of the griseusin polyketide synthase gene cluster from Streptomyces griseus. J. Bacterial. 176 (1994) 2627-2634.





























