Embed Size (px)
Citation preview

Chromosome Analysis of Parallel Short-Term Cultures from Four Testicular Germ-Cell Tumors
Jennifer M. Parrington, Lynne F. West, and Eadie Heyderman
ABSTRACT: Cytogenetic analysis has been carried out on 17 parallel short-term cultures from four malig- nant testicular teratomas of intermediate differentiation (MTI), two of them combined with seminomas. Clonal development was seen in three tumors, with most of the variation involving different rearrange- ments of chromosome 1. Two copies of the i(12p), characteristic of testicular germ-cell tumors, were pres- ent in the two tumors with yolk sac elements, a single i(12p) and a 12q- were found in an MTI that had metastasized, and a rearrangement of chromosome 12 containing centromeric chromatin from chromo- some 18 was found instead of the i(12p) in a mixed tumor. A 13p + marker containing Q-negative material was seen in two of the tumors, with a der(7)t(Y;7) in one. Chromosomes 1, 3, 6, 7, 8, 12, 17, and X were invariably overrepresented either as complete or partial trisomies, and chromosomes 4, 5, 13, and 18 were underrepresented in all four tumors, strengthening the idea that tumor suppressor genes on the latter four chromosomes, all of which show some loss of heterozygosity with DNA markers, may be important in tumor progression.
INTRODUCTION
Testicular germ-cell tumors (I~CT), although one of the rarer forms of cancer in the general population, are the most preva- lent malignancy among young men in the 20-40-year-old age group; in Northern European countries, the incidence is in- creasing. The life-time risk for men of developing this form of cancer is now about I in 500. The only consistent risk fac- tor is cryptorchidism, and individuals with this condition have a 1 in 40 chance of developing a TGCT (for review see [1]). Familial cases of TGCT have been reported in the litera- ture. A recent case-control study gives an incidence of 1.5%, with a I in 50 risk for brothers of affected cases [2]. The figures suggest an earlier age of onset in familial/bilateral cases, com- patible with the existence of a germ-line mutation in some cases predisposing carriers to the disease [3]. Although germ- cell tumors respond well to combined chemotherapy [4], the treatment required after they have metastasized may have to be very intensive. An understanding of initiating events at the cytogenetic and molecular level could lead to earlier di- agnosis at the primary stage, especially for those who are more at risk.
Germ-cell tumors of adults and young men can be divided into seminomas (about 50%), non-seminomas (40%), and
From the MRC Human Biochemical Genetics Unit (J. M. P., L. E W.), The Galton Laboratory, University College London; and the CRC Department of Clinical Oncology [E. H.), Royal Free Hos- pital, School of Medicine, London, U.K.
Address reprint requests to: Dr. Jennifer M. Parrington, MRC Hu- man Biochemical Genetics Unit, Wolfson House, 4 Stephenson Way, London NW1 2HE, U.K.
Received ]uly 23, 1993; accepted December 1Z 1993.
90 Cancer Genet Cytogenet 75 :90-102 [1994) 0165-4608/94/$07.00
combined tumors (10%), all of which are derived from car- cinoma in situ as a precursor [5]. There is now good evidence that seminomas with chromosome numbers in the triploid and tetraploid range, and non-seminomas with chromosome numbers in the hypotriploid range, both of which are charac- terized by the specific i(12p) marker, are stages in the same neoplastic pathway [6]. On this model, non-random chro- mosome losses, after the initiating process, could be impor- tant events in tumor progression, as is the case in colorectal tumor development [7].
We have studied parallel short-term cultures from malig- nant non-seminomatous germ-cell tumors (NSGCT) of inter- mediate differentiation, with and without a seminomatous component, at the chromosome and molecular level (DNA, RFLP analysis). The advantage of using short- and long-term cultures is that better chromosome morphology can be ob- tained and live cells can be stored in liquid nitrogen and grown when new techniques become available. The analy- sis of parallel cultures makes it possible to distinguish be- tween clonal abnormalities which must have been present in vivo and those which may have arisen during the culture period.
We here present the chromosome findings in 17 parallel cultures from four testicular germ-cell tumors.
MATERIALS AND METHODS
Tumor tissue was obtained from four patients, all with malig- nant teratomas of intermediate differentiation (MTI). Two contained yolk sac elements and two areas of seminoma (Ta- ble 1). Fresh tumor samples were washed, minced, and in- cubated for 2-4 days in culture medium (Dulbecco's MEM
© 1994 Elsevier Science Inc. 655 Avenue of the Americas , New York, NY 10010

Karyotype of Four Test icular Teratomas 91
Table 1 Clinical detai ls of the four test icular tumor pat ients
Patient Age Diagnosis
TT16MU TT32GA
TT34DA
TT36ER
26 Malignant teratoma intermediate. 23 Malignant teratoma intermediate with some
trophoblast and yolk sac elements. 25 Combined malignant teratoma intermediate
and seminoma. 32 Malignant teratoma intermediate with yolk
sac tumor and focal seminoma.
or Leibovitz LI5 with 15% fetal bovine serum) with and with- out col lagenase (Worthington 200 units/ml). After collagen- ase treatment, cells were spun down and plated out in fresh med ium in 25-cm 2 flasks or Leighton tubes in an 8% CO2 incubator. Chromosome preparat ions from paral le l cul tures were made in most cases wi th in 1 month of setting up the sample using standard methods. G-, Q-, and C-bandings were carried out as previously descr ibed [8]. To establish the mo- dal chromosome number of each culture, 20-30 orcein- s tained metaphases were counted (or fewer [10 cells] where the chromosome number either exceeded 100 or showed lit- tle variation).
RESULTS
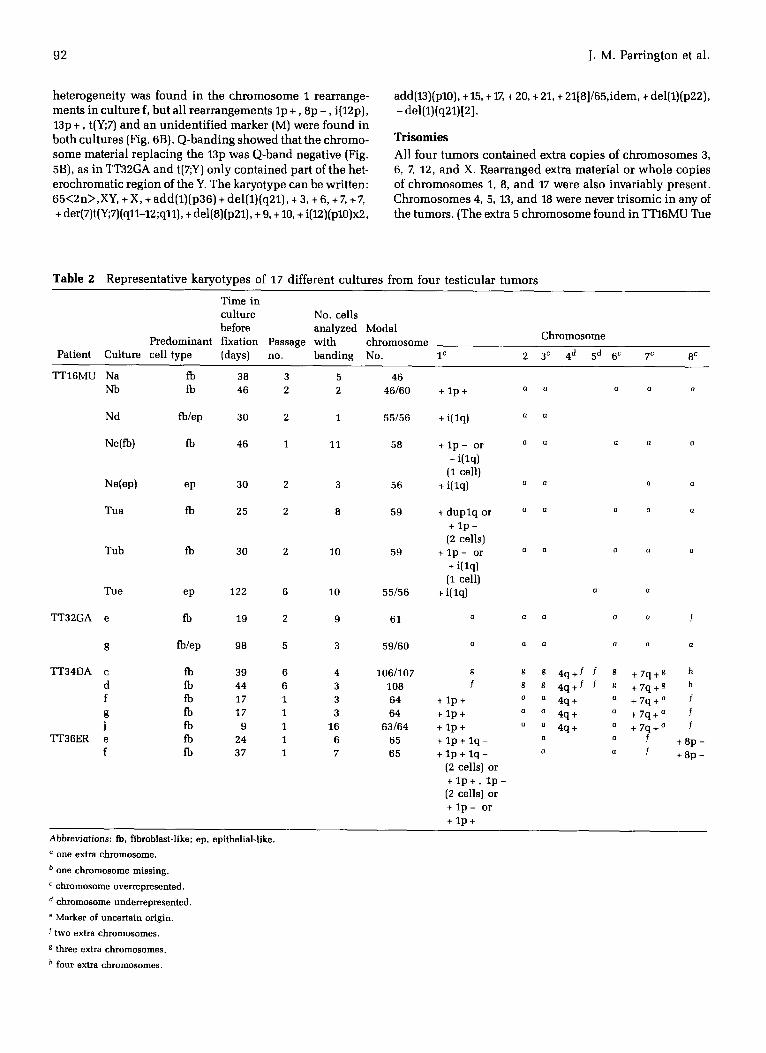
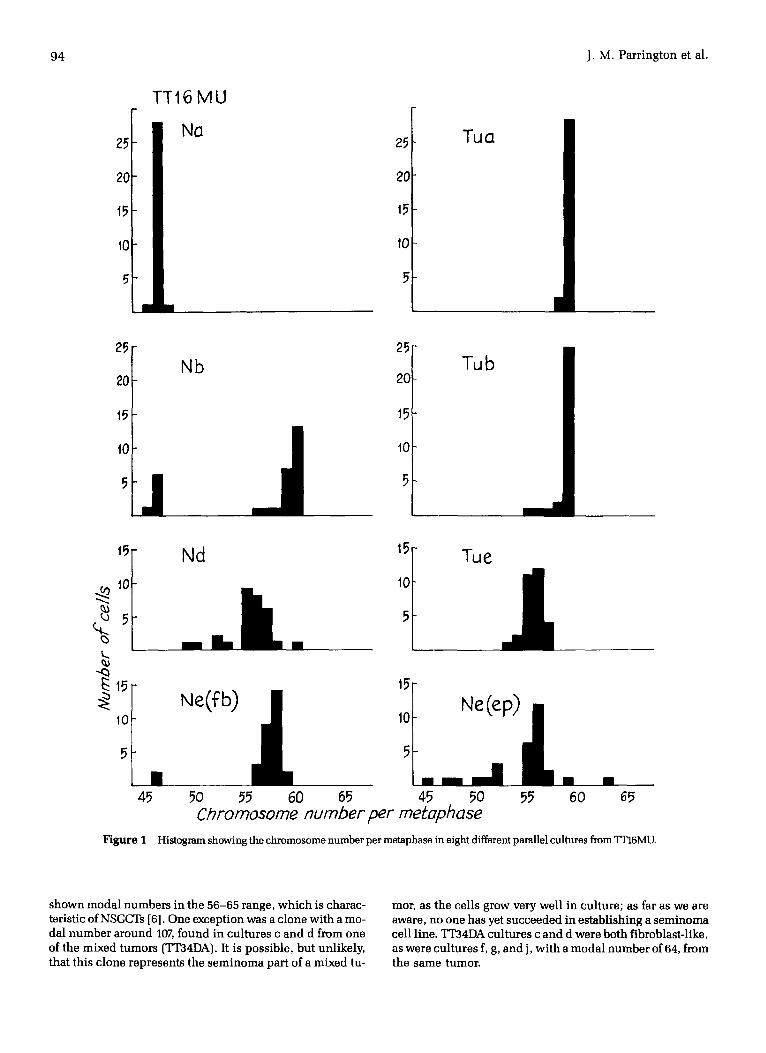
Modal Chromosome Numbers and Karyotypes Chromosome preparat ions were analyzed from at least two parallel cultures from each tumor. Representative karyotypes from each cul ture are shown in Table 2. Most had a modal chromosome number between 55 and 65. Some had a few normal cells, TT16MU Nb and Ne (Fig. 1), whi le one cul ture with all normal cells was found in each of two cases, TT16MU Na and TT36ER c (not shown in Table 2).
TT16MU
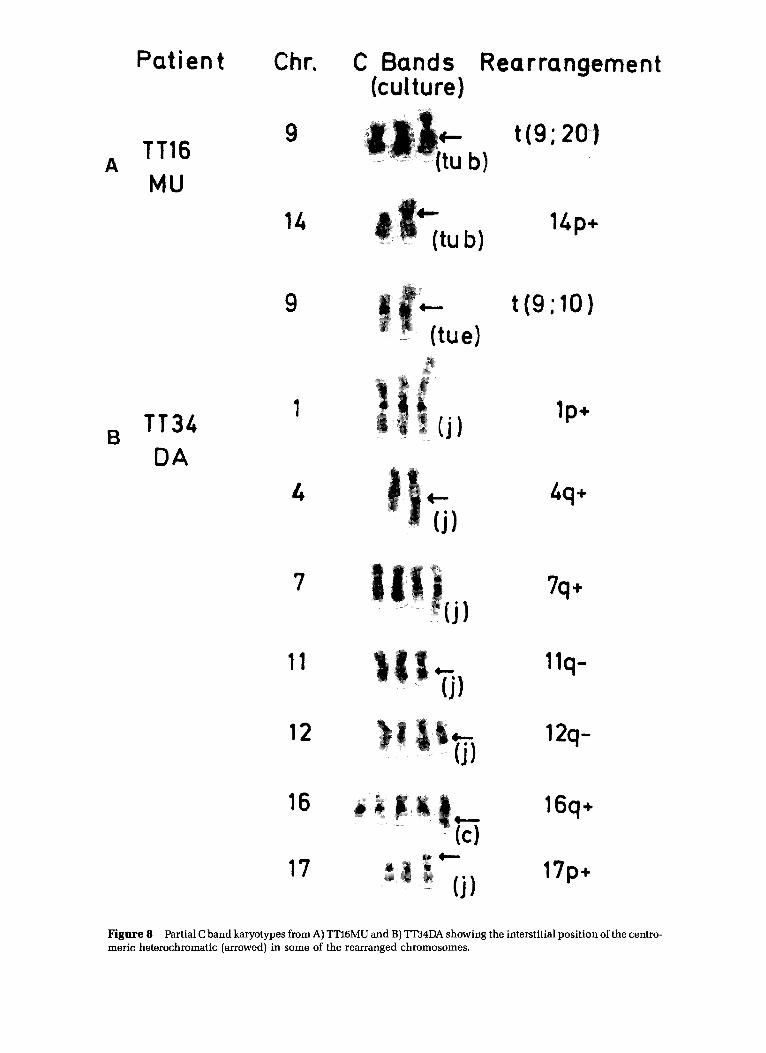
Two separate samples were provided from TT16MU. The one from which the five 'N' cul tures were developed (Table 2) was or iginal ly thought to be from an uninvolved region, but the pathology showed later that the whole testis was replaced by tumor. Seven separate cul tures from the two samples, T and N, both pa thologica l ly tumor, y ie lded analyzable metaphases. Culture Ne was separated at an early stage into fibroblast-like and epi thel ia l - l ike ceils by differential t ryp- sinization. Two dist inct modes, 55/56 and 59/60, were found (Fig. 1), which appeared to be correlated with a par t icular morphology (Fig. 3) and karyotype (Fig. 4), a l though some cul tures were clear ly mixtures of the two (Table 2). Mode 55/56 was associated wi th an epi thel ial- l ike morphology, an i(lq) and a t(9;10), (Fig. 4B), while cells with a mode of 59/60 had a fibroblast-like morphology, a l p - or alternative rear- rangement of chromosome 1, and a t(9;20) (Fig. 4A). Al l cul- tures with an abnormal karyotype had a characteristic i(12p), an extra 12, and a deleted 12, (12q - ) indicating that they must all have been present in the tumor in vivo. The 14p + marker and the t(9;20) in Tub both had two regions of centromeric heterochromatin (Fig. 8A) whereas the l p - and 12q- d id
not. Altogether, four different chromosome I rearrangements were found among the seven karyotypical ly abnormal cul- tures from this tumor. The karyotypes of the main clones can be descr ibed as follows: 59~2n~,XY, + X, + del(1)(p32), + 2, + 3, + 6, + 7, + 8, + der(9)t(9;?;20)(q11;?;p13), + 12, + del(12)(q21q24), + i(12)(p10),add(14)(p11), + 17, - 20, + 21, + 22111]/59,idem, - del (1), ÷ der(1)del(1)(p22)dup(1)(q12qter)[6]/55-56,XY, + X, + i(1) (ql0), + 2, + 3, + Z + 8,t(9;10)(p11;q11), + 12, + del(12)(q21q24), + i(12)(p10), + 1714]/55-56,XY, + X, + i(1)(ql0), + 5, ÷ 7, + ?del(8)
(q13),t(9;10)(p11;q11), ÷ 12, + del(12)(q21q24), + i(12)(p10), + 16, + 17, + ace[10].
TT32GA
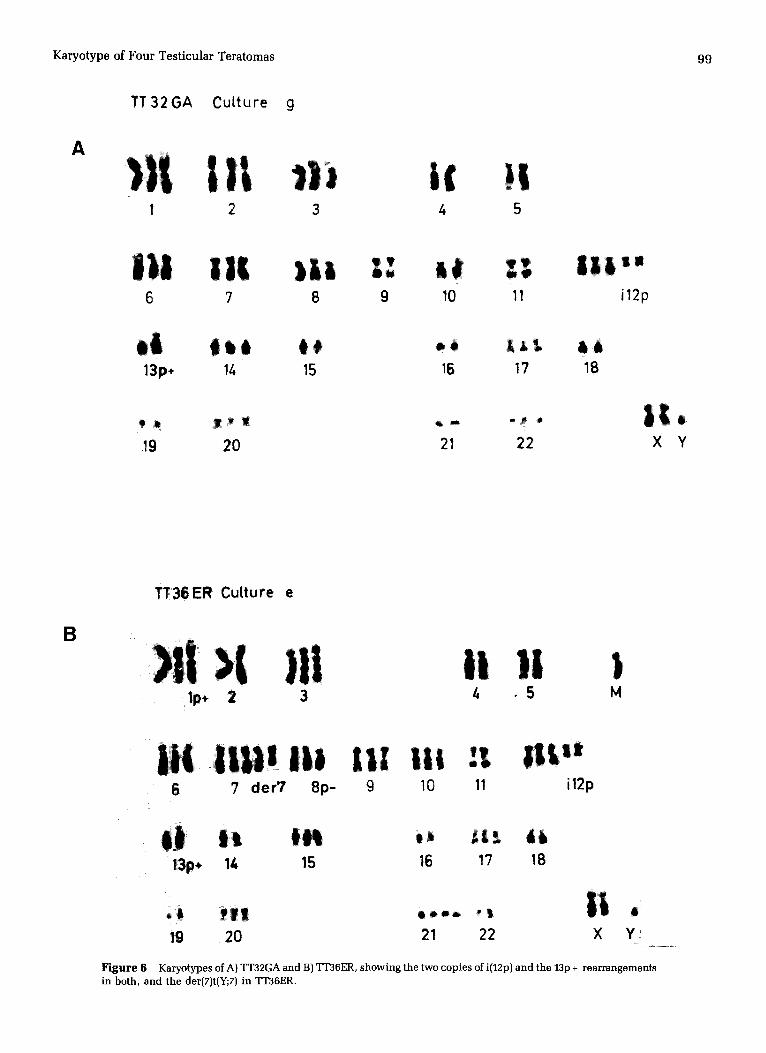
The two cul tures from this sample, a l though karyotyped at very different t imes (19 and 98 days after setting up), showed very s imilar modal numbers, 61 and 59/60 (Fig. 2A) and karyotypes (Table 2). The only structural rearrangements were the i(12p) and the 13p + (Fig. 6A). Chromosome material added to 13p was Q-band negative (Fig. 5A). It is clear that new rearrangements are not arising during the culture period and that the karyotype is very stable. It can be descr ibed as follows: 61~2n~,XY, + X, + 1, + 2, + 3, + 6, +7, + 8, + 8, + i(12) (p10),add(13)(p10), + 17, + 20, + 22.
TT34DA Two quite different modes, 64 and 106/107, were found in this tumor, each represented by two or more cul tures (Fig. 2B). All cul tures had a fibroblast-like morphology. They had six rearrangements in common, inc luding a 12q- (Fig. 7), but the clone with the larger mode was not s imply a doubl ing of the 64 clone, because most cells contained only one copy of each rearrangement. The 64 clone also had a distinctive chromosome I rearrangement (Fig. 7B). This tumor is nota- ble for the lack of a characteristic i(12p) marker chromosome. C-banding shows clearly that the 12q- marker is not s imply a dele t ion but has C-band material added to it (Fig. 8B), now shown by f luorescence in situ hybridizat ion (FISH) studies to come from chromosome 18 [9]. Interstitial C-banding is also found in the 4q+, 11q- , 16q +, and 17p + rearrange- ments. From the G-banding and init ial FISH studies, the karyotype of the two clones can be descr ibed as follows: 106-108 ~ 4n >,XXYY, + X, + 1, + 2, + 3,add(4)(q35), + 6, + 7, + add(7)(q32), + 8, + 8, + 9,del(11)(q22), + 12, + der(12)dic(12;18) (q11;q11)inv(18)(qllq22)add(18)(q10), + 14, + add(16)(q22), + add (17)(p13), - 18, + 19, + 2014]/63-64~2n~,XY, + X, + add(1)(p36), + 2, + 3, add(4)(q35), + 6, + 7, + add(7)(q32), + 8, + 8, + 9, + del(11) (q22), + 12, + der(12) (dic) (12 ;18) (q11; ql 1)inv(18) (qllq22 )add (18)(q10), + 14, + 15, + add(16)(q22), ÷ add(17)(p13), -18, + 19, + 20 [16].
Further FISH studies [10] have shown that two of the "normal-looking" chromosomes 1 in cul ture Tuc (Fig. 7A) have 12p regions inserted at lq42, i.e., der(1)t(1;12)(q42;p11). This rearranged chromosome 1 is also found in Tuj, along with another rearranged chromosome 1, add(1)(p36), which has 12p regions inserted in both arms, so that the rearrange- ment becomes der(1)t(1;12;12)(12pter-12p11::1p36-~cen -* lq42: :12 p11-~12pter).
TT36ER Karyotypes and modal numbers in cultures obtained from this tumor were vi r tual ly identical (Table 2, Fig. 2C). Some
92 J.M. Parrington et al.
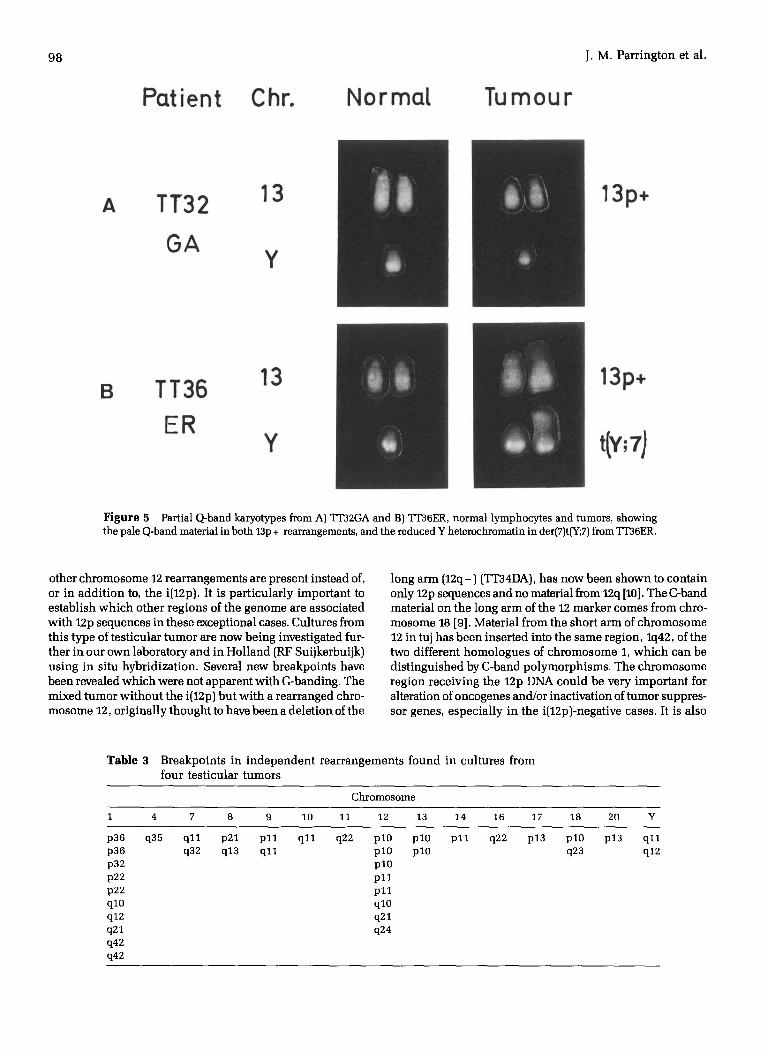
heterogeneity was found in the chromosome 1 rearrange- ments in cul ture f, but all rearrangements lp +, 8p - , i(12p), 13p +, t(Y;7) and an unident i f ied marker (M) were found in both cultures (Fig. 6B). Q-banding showed that the chromo- some material replacing the 13p was Q-band negative (Fig. 5B), as in TT32GA and t(7;Y) only contained part of the het- erochromatic region of the Y. The karyotype can be written: 65<2n>,XY, + X, + add(1)(p36) + del(1)(q21), + 3, + 6, + 7, + 7, + der(7)t(Y;7)(q11-12 ;q11), + del(8)(p21), + 9, + 10, + i(12)(p10)x2,
add(13)(p10), + 15, + 17, + 20, + 21, + 2118] /65 , idem, + del (1)(p22) ,
- del(1)(q21)[2] .
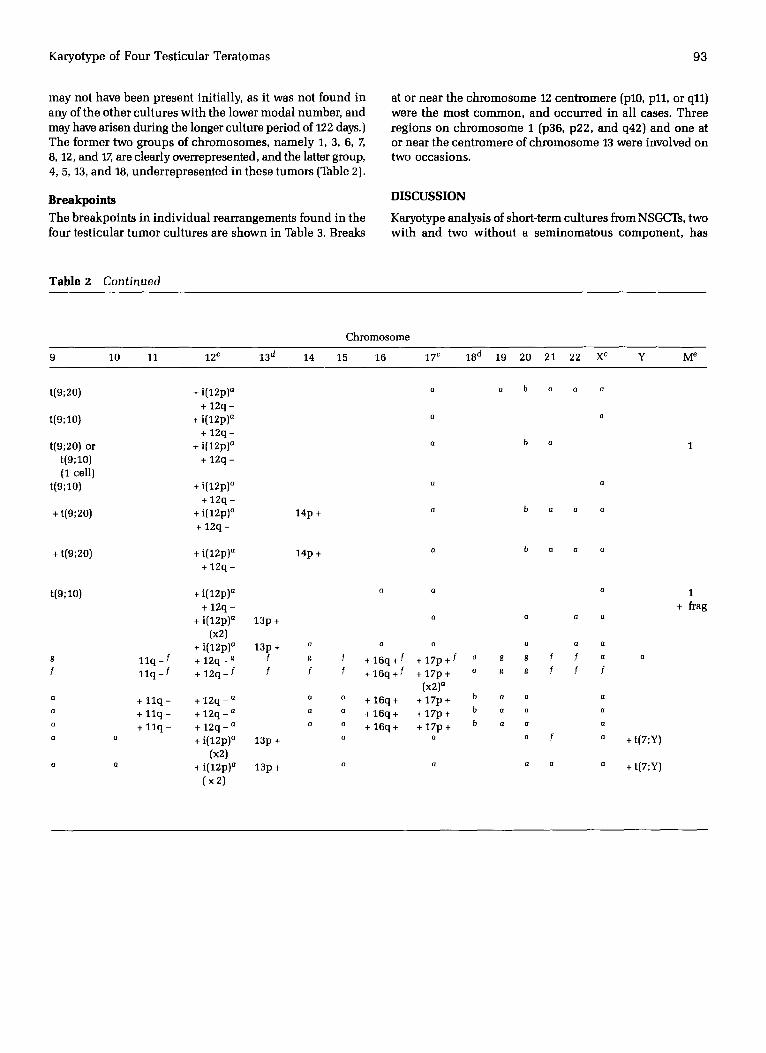
T r i s o m i e s
A l l f o u r t u m o r s c o n t a i n e d e x t r a c o p i e s o f c h r o m o s o m e s 3,
6, 7, 12, a n d X. R e a r r a n g e d e x t r a m a t e r i a l o r w h o l e c o p i e s
o f c h r o m o s o m e s 1, 8, a n d 17 w e r e a l s o i n v a r i a b l y p r e s e n t .
C h r o m o s o m e s 4, 5, 13, a n d 18 w e r e n e v e r t r i s o m i c i n a n y o f
t h e t u m o r s . ( T h e ex t r a 5 c h r o m o s o m e f o u n d i n T T 1 6 M U T u e
Table 2 Representative karyotypes of 17 different cultures from four testicular tumors
T i me in cu l ture No. cel ls before ana lyzed Modal
P r e d o m i n a n t f ixat ion Passage w i t h c h r o m o s o m e Pat ient Cul ture cell type (days) no. b a n d i n g No. i c
C h r o m o s o m e
2 3 c 4 d 5 d 6 c 7 c 8 c
TT16MU Na fb 38 3 5 46 Nb fb 46 2 2 46/60
Nd fb/ep 30 2 1 55/56
Ne(fb) fb 46 1 11 58
N e ( e p ) ep 30 2 3 56
Tua fb 25 2 8 59
Tub fb 30 2 10 59
Tue ep 122 6 10 55/56
TT32GA e
g
TT34DA c d f g
J TT36ER e
f
fb 19 2 9 61
fb/ep 98 5 3 59/60
fb 39 6 4 106/107 fb 44 6 3 108 fb 17 1 3 64 fb 17 1 3 64 fb 9 1 16 63/64 fb 24 1 6 65 fb 37 1 7 65
+ l p +
+ i( lq)
+ l p - or + i( lq)
(1 cell) + i( lq)
+ d u p l q or + l p -
(2 cells) + l p - or
+ i(lq) (1 cell)
+ i(lq)
g
f
+ l p + + l p + + l p + + l p + l q - + 1 p + 1 q -
(2 cells) or + l p + , l p - (2 cells) or + l p - or + l p +
G G 13 0 G
a 13
Q G G (] 0
0 0 Q Q
Q 0 G
(] Q G G G
a f
s g 4 q + f f s + 7 q + g h g g 4 q + f f g + 7 q + g h
a a 4q+ a + 7 q + a f a a 4 q + a + 7 q + a f
a a 4q+ a + 7 q + a f a a f + 8 p -
a a f + 8 p -
Abbreviat ions: fb, fibroblast-like; ep, epithelial-like.
° one extra chromosome.
b o n e c h r o m o s o m e m i s s i n g .
c chromosome overrepresented.
d chromosome underrepresented.
e Marker o f u n c e r t a i n or ig in .
f two extra chromosomes.
g three extra chromosomes.
h four extra chromosomes.
K a r y o t y p e o f F o u r T e s t i c u l a r T e r a t o m a s 93
m a y n o t have b e e n p r e s e n t in i t ia l ly , as it w a s n o t f o u n d in
a n y of t h e o t h e r c u l t u r e s w i t h t h e l o w e r m o d a l n u m b e r , a n d
m a y have a r i s e n d u r i n g the l onge r c u l t u r e p e r i o d of 122 days.)
T h e f o r m e r t w o g r o u p s of c h r o m o s o m e s , n a m e l y 1, 3, 6, 7,
8, 12, a n d 17, are c lea r ly ove r r ep re sen t ed , a n d the latter g roup ,
4, 5, 13, a n d 18, u n d e r r e p r e s e n t e d i n t h e s e t u m o r s (Table 2).
B r e a k p o i n t s
T h e b r e a k p o i n t s in i n d i v i d u a l r e a r r a n g e m e n t s f o u n d in t h e
f o u r t e s t i c u l a r t u m o r c u l t u r e s a r e s h o w n in Table 3. Breaks
at o r n e a r t h e c h r o m o s o m e 12 c e n t r o m e r e (pl0, p11, o r q11)
w e r e t h e m o s t c o m m o n , a n d o c c u r r e d in all cases . T h r e e
r e g i o n s o n c h r o m o s o m e I (p36, p 2 2 , a n d q42) a n d o n e at
o r n e a r t h e c e n t m m e r e of c h r o m o s o m e 13 w e r e i n v o l v e d o n
t w o o c c a s i o n s .
D I S C U S S I O N
K a r y o t y p e a n a l y s i s of s h o r t - t e r m c u l t u r e s f r o m NSGCTs, t w o
w i t h a n d t w o w i t h o u t a s e m i n o m a t o u s c o m p o n e n t , h a s
T a b l e 2 Continued
Chromosome
9 10 11 12 c 13 d 14 15 16 17 c 18 d 19 20 21 22 X c Y M e
t(9;20) + i(12p)a o o b a a a
+ 12q - t(9;10) + i(12p)a a a
+ 1 2 q - t(9;20) or + i(12p)a a b a 1
t(9;10) + 12q - (1 cell)
t(9;10) + i(12p)a a a + 1 2 q -
+ t(9;20) + i(12p) a 14p + a b a a a + 1 2 q -
+ t(9;20) + i(12p) a 14p + + 1 2 q -
a b a a a
t(9;10) + i(12p) . . . . + 12q -
+ i(12p) a 13p + a a a a (x2)
+ i(12p) a 13p + a a a a a a g 1 1 q - / +12q -~ f g ~ + 1 6 q + f + 1 7 p + / a g g f f a f 1 1 q - f +12q - f f f f + 1 6 q + f + 1 7 p + o s g f f f
(x2) a a + 1 1 q - +12q - a a o + 1 6 q + + 1 7 p + b a ~ a o + 1 1 q - +12q - a o a + 1 6 q + + 1 7 p + b a a a o + 1 1 q - + 1 2 q - a o o + 1 6 q + + 1 7 p + b o a o
o a + i(12p) ° 13p + o o o f o
(x2) a Q + i(12p) a 13p + a a a a a
(×2)
+ t(7;Y)
+ t(7;Y)
1
+ frag
94 J .M. Parrington et al.
25
20
15
t0
TT16 MU
Nd 25
20
10
Tua
2~ I Nb 2O
IP t°LL j P
2P
20
15
10
5
Tub
~b
t5
10
5
Nd 15
10
5
Tue
lo
5
45
Figure I
Ne(m) 101~fl5 Ne
50 ~ 60 65 45 PO Chromosome number per me~aphase
I I
55 60 65
Histogram showing the chromosome number per metaphase in eight different parallel cultures from TI~6MU.
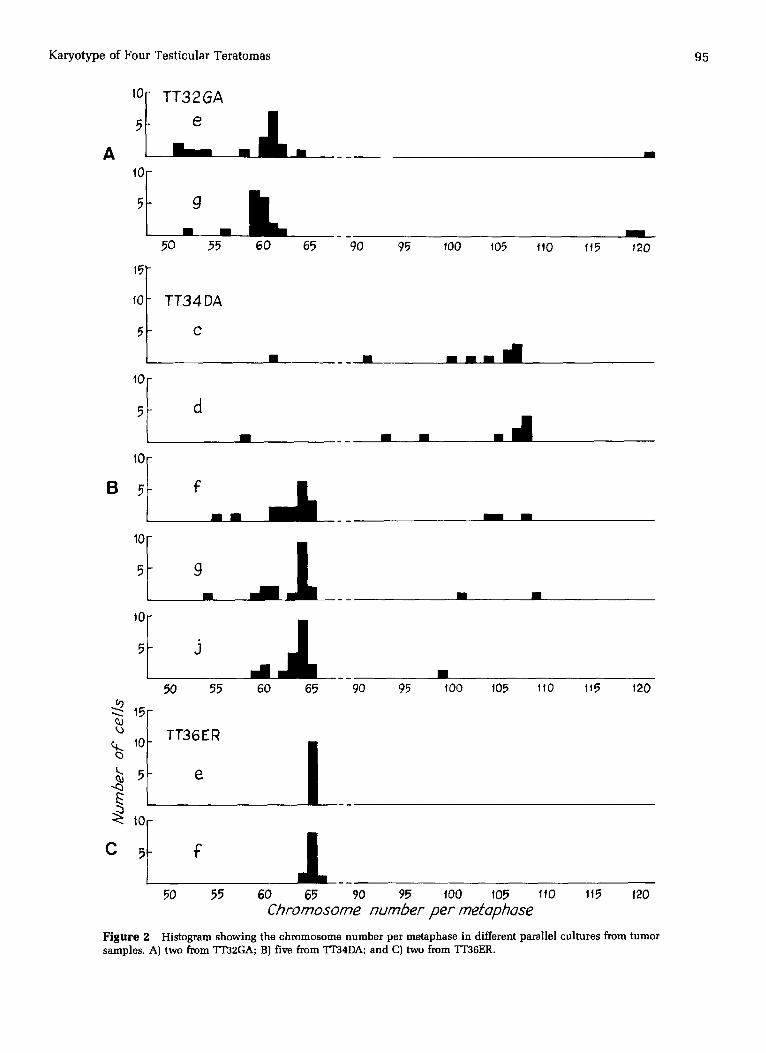
shown modal numbers in the 56-65 range, which is charac- teristic of NSGCTs [6]. One exception was a clone with a mo- dal number around 107, found in cultures c and d from one of the mixed tumors (TT34DA). It is possible, but unlikely, that this clone represents the seminoma part of a mixed tu-
mor, as the cells grow very well in culture; as far as we are aware, no one has yet succeeded in establishing a seminoma cell line. TIB4DA cultures c and d were both fibroblast-like, as were cultures f, g, and j, with a modal number of 64, from the same tumor.
Karyotype of Four Test icular Teratomas 95
A
lo T T 3 2 G A
m
10
i: I lOr
50 .55 60
T T 3 4 DA
C
65 90 95 ~00 t05 t10 t~5 ~20
i _ _ ! _ _ ! _ _ 3
B ~
1:f_
,o
5
c f
Figure 2
f,
II m
50 55 60 65 90 95 ~00 105 t~0 1~5 f20
TT36ER
e I 50 55 60 65 90 95 t00 105 lf0 1t5 120
Chromosome number per metophose Histogram showing the chromosome number per metaphase in different parallel cultures from tumor
samples. A) two from TT32GA; B) five from TT34DA; and C) two from ~T36ER.
96 J .M. Parrington et al.
TT16MU
A B
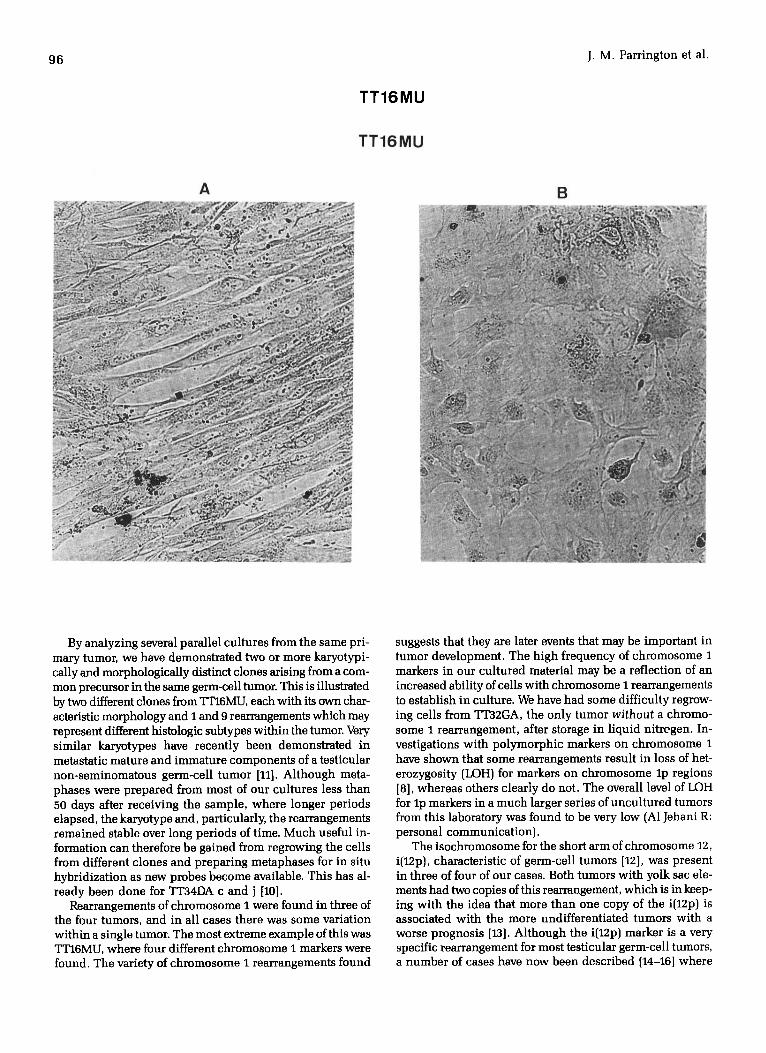
Figure 3 Morphologically distinct cell monolayers from two clones of TTI6MU differing in chromosomes 1 and 9 rearrangements. A) Ne(fb): + lp- and t(9;20); B) Tue(ep): +i(lq) and t(9;10).
By analyzing several parallel cultures from the same pri- mary tumor, we have demonstrated two or more karyotypi- cally and morphologically distinct clones arising from a com- mon precursor in the same germ-cell tumor. This is illustrated by two different clones from"ffl"16MU, each with its own char- acteristic morphology and I and 9 rearrangements which may represent different histologic subtypes within the tumor. Very similar karyotypes have recently been demonstrated in metastatic mature and immature components of a testicular non-seminomatous germ-cell tumor [11]. Although meta- phases were prepared from most of our cultures less than 50 days after receiving the sample, where longer periods elapsed, the karyotype and, particularly, the rearrangements remained stable over long periods of time. Much useful in- formation can therefore be gained from regrowing the cells from different clones and preparing metaphases for in situ hybridization as new probes become available. This has al- ready been done for TT34DA c and j [10].
Rearrangements of chromosome I were found in three of the four tumors, and in all cases there was some variation within a single tumor. The most extreme example of this was TT16MU, where four different chromosome I markers were found. The variety of chromosome I rearrangements found
suggests that they are later events that may be important in tumor development. The high frequency of chromosome 1 markers in our cultured material may be a reflection of an increased ability of cells with chromosome I rearrangements to establish in culture. We have had some difficulty regrow- ing cells from TT32GA, the only tumor without a chromo- some I rearrangement, after storage in liquid nitregen. In- vestigations with polymorphic markers on chromosome 1 have shown that some rearrangements result in loss of het- erozygosity (LOH) for markers on chromosome lp regions [8], whereas others clearly do not. The overall level of LOH for lp markers in a much larger series of uncultured tumors from this laboratory was found to be very low (A1Jehani R: personal communication).
The isochromosome for the short arm of chromosome 12, i(12p), characteristic of germ-cell tumors [12], was present in three of four of our cases. Both tumors with yolk sac ele- ments had two copies of this rearrangement, which is in keep- ing with the idea that more than one copy of the i(12p) is associated with the more undifferentiated tumors with a worse prognosis [13]. Although the i(12p) marker is a very specific rearrangement for most testicular germ-cell tumors, a number of cases have now been described [14-16] where

TT16MU Culture Tu b A
6
lp- 2 3
7
13 14p+
8
i i i i ¸
15
4 5
~9;2o) ;o 11
16 17 18
12q-i12p
+++ I 19 20
++ +, + +++
21 22
+i
X Y
B
TT16MU Culture Tu e
?lp+ ilq l+i
2 3 i(i i ! I
4 5
6 7 ?8q- t~;lO) 10 11 12q-i12p
13 14 15 16 17 18
i • i : i + 6
19 fragment 21 22
l!, × Y
F igu re 4 Karyotypes of the two main clones from TT16MU, showing the different rearrangements of the chromo- somes I and 9, and the same i{12p) and 12q - chromosomes found in both. (note: most cells from culture e had two normal chromosomes 4 and two normal chromosomes 20).
98 ]. M. Parrington et al.
Patient C h r . Normal Tu mou r
A TT32
GA
13
Y
13p+
B TT36 ER
13
Y
13p+
tiY;7 )
Figure 5 Partial Q-band karyotypes from A) TT32GA and B) TT36ER, normal lymphocytes and tumors, showing the pale Q-band material in both 13p + rearrangements, and the reduced Y heterochromatin in der(7)t(Y;7) from ~'36ER.
other chromosome 12 rearrangements are present instead of, or in add i t ion to, the i(12p). It is par t icular ly important to establish which other regions of the genome are associated with 12p sequences in these exceptional cases. Cultures from this type of test icular tumor are now being investigated fur- ther in our own laboratory and in Hol land (RF Suijkerbuijk) using in situ hybridizat ion. Several new breakpoints have been revealed which were not apparent with G-banding. The mixed tumor wi thout the i(12p) but wi th a rearranged chro- mosome 12, original ly thought to have been a delet ion of the
long arm (12q-) (TT34DA), has now been shown to contain only 12p sequences and no material from 12q [10]. The C-band material on the long arm of the 12 marker comes from chro- mosome 18 [9]. Material from the short arm of chromosome 12 in tuj has been inserted into the same region, Iq42, of the two different homologues of chromosome 1, which can be dis t inguished by C-band polymorphisms. The chromosome region receiving the 12p DNA could be very important for alteration of oncogenes and/or inactivation of tumor suppres- sor genes, especial ly in the i(12p)-negative cases. It is also
Table 3 Breakpoints in independen t rearrangements found in cultures from four test icular tumors
Chromosome
1 4 7 8 9 10 11 12 13 14 16 17 18 20 Y
p36 q35 q l l p21 p l l q l l q22 pl0 pl0 p36 q32 q13 q l l p l0 plO p32 pl0 p22 p l l p22 p l l ql0 qlO q12 q21 q21 q24 q42 q42
p l l q22 p13 pl0 p13 q l l q23 q12
Karyotype of Four Testicular Teratomas 99
A
TT32GA Culture
!li i¢ =l 2 3 4 5
6 ? 8 9 10 11
!!!" i12p
Ii i S i l ¢ . 6 A ~ L ti ¢ 13p+ 14 15 16 17 18
19 20 21 22 X Y
B
TT36ER Cutture e
I i i i i l i ) lp* 2 3 & 5 M
ll~=. l t l i i i | | I Z¢ 9 10 11 6 7 der7 8p- i12p
| t 13p" 14
i n , . AL~ i b 15 16 17 18
Ii • t Y t | , , a . . . ~ o
19 20 21 22 X Y o . . . . . . . . .
Figure 6 Karyotypes of A) TT32GA and B) TT36ER, showing the two copies of i(12p) and the 13p + rearrangements in both, and the der(7)t(Y;7) in TT36ER.

100
TT 34 DA
A
CuLture c
1 2 3 4q÷ 5
6 7q+ 8 9 10 11q-
13 14 15 16q+ 17p+ 18
12q-
19 20 21 22 X
PO
Y
B TT34 DA CuLture f
lp+ 2 3 4q+ 5
6 7q+ 8 9 10 11q- 12q-
13 14 15 16q, 17p+ 18
19 20 21 22 X Y
Figure 7 Karyotypes of the two clones from TT34DA showing the six common rearrangements, and the lp + found only in cultures f, g and j.
A
Patient
TT16 MU
Chr. C Band s (culture)
9 b)
Rear r(:l ngeme nt
t (9 ;20 )
14 lap* ,tub)
B TT34 DA
9 t19;101 g e ~ )
.... (j)
, ,q. (j)
7 j) 9 +
,._ 11q-
~2 ~ i ! i i 7 ) 12q-
16 (c)
17 ~ ~ ~ *'(j) 1 ? p +
16q+
Figure 8 Partial C band karyotypes from A) TT16MU and B) TT34DA showing the interstitial position of the centro- meric heterochromatic (arrowed) in some of the rearranged chromosomes.

102 J .M. Parrington et al.
interesting to note the presence of a 12q- marker in an i(12p)- positive tumor (TT16MU), which has been seen in a num- ber of other NSGCTs [15]. This could be an interstitial or ter- minal deletion (12q21-12qter), which would result in the de- letion of band 12q22, the site of the more distal tumor suppressor gene postulated by Murty et al. [19]. No LOH, however, was found for any of three informative 12q probes, either proximal or distal to this region, in more than 20 3GCTs [20].
It is worth noting the 13p + rearrangement in the two tumors with yolk sac elements. In both cases there was ad- dit ion of Q-negative material; it may be possible to inves- tigate these further with in situ hybridization. Two of the three i(12p)-negative cases described by Atkin et al. [16] had rearrangements involving chromosomes 12 and 13 with prob- able breakpoints at 13qll and 13q22, and 13pll is mentioned as a non-random breakpoint by Rodriguez et al. [17] in a much larger survey of germ-cell tumors. The breakpoints involved in clonal rearrangements from all four tumors are shown in Table 3. Apart from the expected clustering of breakpoints on chromosomes 1 and 12, only 13p10 (or 13p11) is found in two different tumors, and the only other breakpoint which co-incides with the non-random breakpoints of Rodriguez et al. [17] is 7q11.
The involvement of chromosome 18 in a complicated rear- rangement with chromosome 12 in the i(12p)-negative tumor may be important. In situ hybridization with the chromo- some 18 paint probe has revealed a deletion of about half the long arm in another of our cell lines, KDK2B [TT6K1) [8]. This work will be presented at a later date after further investigation.
The translocation between the Y chromosome and what appears to be the short arm of a chromosome 7 found in ad- dition to a normal Y in TT36ER is the result of breakage in the heterochromatic region of the Y chromosome. It is not clear whether the rest of the Y heterochromatin is still pres- ent in the karyotype. Although rearrangements involving the heterochromatic region of the Y chromosome have been de- scribed in testicular tumor cell lines [18], those involving autosomes as well are not found often.
The clear-cut underrepresentat ion of chromosomes 4, 5, 13, and 18, which were never found to be trisomic in any of the four teratomas of intermediate differentiation described here, is consistent with previous findings [6], at least for chro- mosomes 5, 13, and 18, and is an indicator that tumor sup- pressor genes on these chromosomes may play a part in the progression of TGCTs. Further cytogenetic, I£)H, and link- age analysis should reveal which parts of the genome are of prime importance at different stages and lead to the iden- tification of any tumor suppressor genes that may be involved.
We are grateful to Dr. Michael Cullen for supplying tissue samples from one of the patients, to Gloria Jones, Margaret Fox, and Toni Lee for help in the preparation of the figures, and to Nalini Pillay and Naomi Tobi for typing the manuscript.
REFERENCES
1. Oliver RTD (1990): Atrophy, hormones, genes and viruses in ae- tiology of germ cell tumours. Cancer Surveys 9:263-286.
2. Forman D, Oliver RTD, Brett AR, Marsh SGE, Moses JH, Bod- mer JG, Chilvers CED, Pike MC (1992): Familial testicular can- cer: a report of the UK family register, estimation of risk and an HLA class I sib-pair analysis. Br J Cancer 65:255-262.
3. Knudson AG (1986): Genetics of human cancer. Ann Rev Genet 20:231-251.
4. Howich A (1986): Germ cell tumour chemotherapy. Br J Cancer 59:156-159.
5. JCrgensen N, M~ller J, Giwercman A, Skakkebaek NE (1990): Clinical and biological significance of carcinoma in situ of the testis. Cancer Surveys 9:287-302.
6. De Jong B, Oosterhuis JW, Castedo S, Vos A, Meerman G (1990): Pathogenesis of adult testicular germ cell tumours. Cancer Ge- net Cytogenet 48:143-167.
7. Vogelstein B, Fearon ER, Hamilton SR, Kern SE, Preisinger AC, Leppert M, Nakamura Y, White R, Smits A, Bos JL (1988): Genetic alterations during colorectal turnout development. N Engl J Med 319: 525-532.
8. Parrington JM, West LF, Povey S (1987): Loss of heterozygosity in hypotriploid cell cultures from testicular tumours. Hum Ge- net 77:269-276.
9. Parrington JM, A1-Jehani R, Wells D, Fox M (1992): Character- ization of marker chromosomes in testicular tumours by band- ing, RFLP analysis and fluorescence in situ hybridization (ab- stract). Cancer Genet Cytogenet 63:149.
10. Suijkerbuijk RF, Sinke RJ, Meloni AM, Parrington JM, Van Echten J, De Jong B, Oosterhuis JW, Sandberg AA, Geurts van Kessel A (1993): Overrepresentation of chromosome 12p sequences and karyotype evolution in i(12)p-negative testicular germ cell tumours revealed by fluorescence in situ hybridisation. Can- cer Goner Cytogenet (in press).
11. De Graaff WE, De Jong B, Oosterhuis JW, Echten-Arends JV, Wiersema-Buist J, Schraffordt Koops H, Sleijfer DT (1991): Cytogenetic analysis of the mature and immature teratoma com- ponents of a metastatic testicular non-seminomatous germ cell tumour. Cancer Genet Cytogenet 57:59-68.
12. Suijkerbuijk RF, Van de Veen AY, Van Echten J, Buys C, De Jong B, Oosterhuis JW, Warburton DA, Cassiman JJ, Schonk D, Geurts van Kessel A (1991): Demonstration of the genuine Iso-12p charac- ter of the standard marker chromosome of testicular germ cell tumours and identification of further chromosome 12 aberra- tions by competitive in situ hybridization. Am J Hum Genet 48:269-273.
13. Bosl GJ, Dmitrovsky E, Reuter VE, Samaniego F, Rodriguez E, Geller NL, Chaganti RSK (1989): Isochromosome of chromosome 12: clinically useful marker for male germ cell turnouts. J Natl Cancer Inst 81:1874-1878.
14. Castedo S, De Jong B, Oosterhuis JW, Seruca R, Idenburg VJ, Buist J, Sleijfer DT (1988): i(12p)-negative testicular germ cell tumors. A different group? Cancer Genet Cytogenet 35:171-178.
15. Samaniego F, Rodriguez E, Houldsworth J, Murty VS, Ladanyi M, Lele KP, Chen O~ Dimitrovsky E, Geller N, Reuter V. Jhanwar SC, Bosl GJ, Chaganti RSK (1990): Cytogenetic and molecular analysis of human male germ cell tumours: chromosome 12 ab- normalities and gene amplification. Genes Chrom Cancer 1:289-300.
16. Atkin NB, Fox M, Baker MC, Jackson Z (1993): Chromosome 12- containing markers, including two dicentrics, in three i(12p)- negative testicular germ cell tumours. Genes Chrom Cancer 6:218-221.
17. Rodriguez E, Mathew S, Router VE, Ilson DH, Bosl GJ, Chaganti RSK (1992): Cytogenetic analysis of 124 prospectively ascertained male germ cell tumours. Cancer Res 52:2285-2291.
18. Arnemann J, Gradl G, Casper J, Schmoll H-J, Schmidtke J, Fonatsch C (1989): Characterization of rearranged Y chromo- somes in human testicular tumour cell lines. Cancer Genet Cytogenet 37:141-151.
19. Murty VS, Houldsworth J, Baldwin S, Router VE, Hunziker W, Besmer P, Bosl GJ, Chaganti RSK (1992): Allelic deletions in the long arm of chromosome 12 identify sites of candidate tumour suppressor genes in male germ cell tumours. Proc Natl Acad Sci USA 89:11006-11010.
20. A1-Jehani R, Povey S, Delhanty J, Parrington J (1991): Loss ofhet- erozygous DNA markers in testicular turnouts (abstract). Can- cer Goner Cytogenet 52:278.





























