Embed Size (px)
Citation preview

at SciVerse ScienceDirect
Biochimie 94 (2012) 2687e2698
Contents lists available
Biochimie
journal homepage: www.elsevier .com/locate/biochi
Research paper
Chronic gastric ulceration causes matrix metalloproteinases-9 and -3augmentation: Alleviation by melatonin
Krishnendu Ganguly 1, Snehasikta Swarnakar*
Department of Physiology, Drug Development Diagnostics and Biotechnology Division, CSIR-Indian Institute of Chemical Biology, 4, Raja S.C. Mullick Road, Jadavpur, Kolkata 700032,India
a r t i c l e i n f o
Article history:Received 16 March 2012Accepted 7 August 2012Available online 13 August 2012
Keywords:IndomethacinChronic ulcerCytokineMatrix metalloproteinaseMelatoninReactive oxygen speciesSignalingNF-kBAP-1
Abbreviations: AP, activator protein; b.w., bodymatrix; ERK, extracellular-regulated kinase; i.p., intJNK, c-Jun N-terminal kinase; MMPs, matrix metalsteroidal anti-inflammatory drugs; NF-kB, nuclear faoxide radical; �OH, hydroxyl radical; H2O2, hydrogen pspecies; RT-PCR, reverse transcriptase-PCR; Tmetalloproteinase.* Corresponding author. Tel.: þ91 33 2473 3491x15
E-mail address: [email protected] (S. Swa1 Present address: Laboratory of Molecular Neurob
teura 3, 02-093 Warsaw, Poland.
0300-9084/$ e see front matter � 2012 Elsevier Mashttp://dx.doi.org/10.1016/j.biochi.2012.08.004
a b s t r a c t
Matrix metalloproteinases (MMPs) are a family of zinc-dependent enzymes capable of degradation ofextracellular matrix (ECM) and key player in various inflammatory diseases. We investigated the regu-lation of MMPs in chronic gastric ulceration in mice. We generated chronic gastric ulcers in mice byindomethacin and examined the activity and expression of MMP-9 and -3 in stomach. Melatonin (N-acetyl-5-methoxytryptamine) treatment has also been applied to mice to characterize the changes inexpression and activities of MMPs in gastric tissues. We observed significant upregulation of MMP-9and -3 expressions and activities in stomach with increasing doses and duration of indomethacin thatcorroborated with increased activity of activator protein (AP)-1. Substantial damage in gastric epitheliallayer was found during chronic ulceration. Melatonin suppressed MMP-9 and -3 expressions andactivities during prevention and healing of chronic gastric ulcers. It also suppressed protein oxidation,lipid peroxidation and antioxidant enzymes. Additionally, expression of tumor necrosis factor (TNF)-a,interleukin (IL)-1b and IL-8 was significantly high in ulcerated stomachs while melatonin treatmentblocked them to control level. We found elevated phosphorylation of extracellular-regulated kinase(ERK)1/2 and c-Jun N-terminal kinase (JNK) during chronic gastric ulceration, which were significantlyreversed by melatonin. Moreover, expression of NF-kB, c-fos and c-jun were inhibited by melatoninresulting down regulation of MMP-9 and -3 expressions. In summary, oxidative stress is preceded bychronic inflammation that enhances the expression of MMP-9 and -3, while melatonin arrests both ofthem via reduction of AP-1 activity during protection of ulcer.
� 2012 Elsevier Masson SAS. All rights reserved.
1. Introduction
Gastric inflammation is more prevalent than gastric ulcer whichpredisposes to both ulceration and in more advanced stages toadenocarcinoma [1]. Apart from the infection with Helicobacterpylori, indiscriminate use of nonsteroidal anti-inflammatory drugs(NSAIDs) are major risk factors for gastritis and gastric ulcers
weight; ECM, extracellularraperitoneal; IL, interleukin;loproteinases; NSAIDs, non-ctor kappa beta; O2
��, super-eroxide; ROS, reactive oxygenIMP, tissue inhibitor of
9; fax: þ91 33 2473 5197.rnakar).iology, Nencki Institute, Pas-
son SAS. All rights reserved.
because both types of damages are accompanied by tissueinflammation and secretion of cytokines, including interleukin (IL)-1b and tumor necrosis factor (TNF)-a [2e4]. Indomethacin causesinjury in the gastroduodenal tract through expression of interleu-kins (IL)-1, -6, and -8 and infiltration of polymorphonuclearleukocyte and in turn apoptosis in lesion area of gastric mucosa[2,5,6]. Indomethacin also activates the infiltrated inflammatorycells in lesion areas that generates reactive oxygen species (ROS)i.e., superoxide (O2�), which is rapidly converted to hydrogenperoxide (H2O2) by superoxide dismutase (SOD) or hydroxyl radical(�OH) by Fenton reaction and the superoxide driven HabereWeissreaction [7e9]. This ROS can induce lipid peroxidation, proteinoxidation and glutathione depletion and, thereby cause tissueinjury in gastric mucosa [10,11]. Melatonin, (N-acetyl-5 methoxy-tryptamine), a hormone produced from the pineal gland, retina andenterochromaffin cells of gastrointestinal tract that maintainscircadian rhythmicity, seasonal reproduction, sleep and gastroin-testinal homeostasis [15,16]. It also acts as an antioxidant and thusprotects against various types of gastric ulceration through

K. Ganguly, S. Swarnakar / Biochimie 94 (2012) 2687e26982688
stimulating glutathione reductase activity, scavenging the hydroxylradical (�OH), reducing the activity of myeloperoxidase (MPO) andlipid peroxidation [17,18].
Matrix metalloproteinases (MMPs), a group of zinc-dependentenzyme remodel healthy connective tissue by specifically digestingcollagen and other matrix molecules [19,20]. In chronic inflam-matory conditions, such as rheumatoid arthritis, osteoarthritis andin tumor invasion, the extracellular secretion ofMMPs substantiallycontributes to stromal tissue destruction [19,21]. Among the MMPfamily there are two unique members i.e. gelatinases (MMP-2 and-9) and stromilysin-1 (MMP-3) collectively can cleave type I, IV, V,VII, and XI collagens, elastin, fibronectin and laminin [19]. Manyfactors including cytokines, growth factors and extracellular matrixproteins have been reported to promote expressions of MMP-9and -3, however the intracellular signaling pathway in chronicgastric ulceration induced by NSAIDs remains to be characterized.
Recent evidences suggest that chronic inflammation leads toincreased production of certain forms of ROS especially H2O2,remain for longer durations in certain compartments of cell andlow concentration of H2O2 may act as second messengers forvarious physiological signaling [12e14] through the activation oftranscription factors such as nuclear factor-kappa B (NF-kB) andactivator protein-1 (AP-1) [22,23]. The increased ROS generationcan activate mitogen-activated protein kinase (MAPK)s, includingextracellular-regulated kinase (ERK)1/2 and c-Jun N-terminalkinase (JNK) [23]. JNK phosphorylates Jun transcription factorfamily members c-Jun and JunD, whereas ERK-1/2 phosphorylatesJun transcription factor family members (c-Jun) and Fos familymembers (c-Fos). Upon dimerization, JuneJun or JuneFos trans-locate into the nucleus and augment several MMP-gene as well asinflammatory gene transcription [23]. However, the regulation andmechanism of action of MMPs at transcription and proteinexpression levels are poorly understood during chronic gastriculceration and healing process.
In the present study, we demonstrate that chronic gastriculceration by indomethacin caused inflammation in a dose andtime dependent manner which led to increased ROS productionfollowed by MMP-9 and -3 mRNA and protein upregulation. Theincreased production of cytokines i.e., TNF-a, IL-1b and IL-8 affectedthe signaling of MMP-9 and -3 by NF-kB and AP-1 in ulcer milieu.Melatonin, herein blocks the inflammation by suppressing cyto-kines expression during protection and healing of gastric ulcer.Additionally, it reduced oxidative stress by reducing SOD-2 andcatalase expressions thereby reducing MMP-9 and -3 activities andexpressions. Melatonin also reduced the phosphorylation of ERK1/2and JNK and, suppressed MMPs by lowering AP-1 (c-fos and c-jun)expression during prevention of chronic ulceration. In conclusion,melatonin offers MMP-9 and -3 gene suppression in addition to itsanti-inflammatory and antioxidant property while arrestingchronic gastric ulcer.
2. Materials and method
2.1. Materials
Gelatin from porcine skin, indomethacin, melatonin, Triton X-100 (TX), protease inhibitors cocktail, gelatin fused with 4% beadedagarose, 2,4,dinitrophenyl hydrazine, guanidine HCl, tetraguaiacol,benzenesulfinic acid and 5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazonium solution were obtained from Sigma (St.Louis, MO, USA). Bradford reagent was purchased from Bio-Rad(Hercules, CA). Trichloroacetic acid (TCA) and Tris base acetate(TBA) were obtained from SRL, India. Monoclonal anti-MMP-2antibody, polyclonal anti-TNFa, anti-IL-1b, anti-IL-8, anti-MMP-9,anti-pERK1/2(Tyr-204), anti-p-JNK (Thr-183, Tyr-185), anti-NF-kB,
anti-cFos, anti-p-c-Jun (Ser-73) anti-Histone (H)4 and anti-GAPDHantibodies were purchased from Santa Cruz (CA, USA). TRIzolreagent, Superscript II Reverse Transcriptase and oligo (dT)15primer were purchased from Invitrogen (CA, USA).
2.2. Animals used
BALB/c mice (25e30 g) bred in-house and maintained with freeaccess to food andwaterwere used for all experiments. Experimentsweredesigned tominimize animal sufferingand touse theminimumnumber associated with valid statistical evaluation. Animal experi-ments were carried out following the guidelines of animal ethicscommittee of the Institute. Animals of both control andexperimentalgroups were kept separately under standard controlled conditionand were fasted for 24 h with free access to water.
2.3. Indomethacin-induced chronic gastric ulceration
The chronic gastric ulcer model was standardized using indo-methacin according to the study of Yamagiwa et al. [10]. In brief,different doses of indomethacin (2, 5, 10 and 20 mg/kg b.w.) wereadministered on BALB/c mice for consecutive 3 days to study theseverity of chronic ulceration. For time dependent experiment themice were killed after 0, 1, 2 and 3 days of indomethacin posttreatment. The 10 mg/kg b.w. indomethacin was considered as aneffective dose for generation of the chronic ulcer for all experi-ments. The mice were sacrificed at different days (0, 1, 2, 3, 6 and 9days) for auto and melatonin mediated healing studies. The controlgroup received vehicle orally for consecutive 3 days for both doseand time dependent studies.
2.4. Preventive and healing studies of indomethacin-induced gastriculcer by melatonin
To study the dose dependent gastroprotective effect, differentdoses (10, 20 and 40 mg/kg b.w.) of melatonin were dissolved indistilled water with a minimal amount of ethanol (10e15%) andwere administered intraperitoneally (i.p.) to different groups ofmice prior to oral administration of indomethacin (10 mg/kg b.w.)for 5 consecutive days and were sacrificed on the following day. Totest the time-dependent effect of melatonin in indomethacin-induced chronic gastric ulceration, melatonin at a dose of 40 mg/kg b.w. was given i.p. 30 min. prior to indomethacin (10mg/kg b.w./day) treatment for consecutive 5 days. The rest of the timedependent study were conducted by administration of vehicle forulcerated groups and only melatonin plus vehicle for melatoninpretreated groups and, were monitored up to 9 days (n ¼ 8). Tostudy the healing effect of melatonin, one group served as autohealing where indomethacin (10 mg/kg b.w.) were administeredorally for consecutive 3 days and the study were continued furtherup to 12 days with vehicle treatment. The other groups served asmelatonin-mediated healing where melatonin (40 mg/kg b.w.) waspretreated i.p. along with vehicle treatment for everyday after 3consecutive days of indomethacin administration and was moni-tored up to 12 days. The mice were sacrificed at different intervalsfor both preventive as well as healing studies (n ¼ 12).
2.5. Measurement of protein carbonyl
Protein oxidation was measured as carbonyl content in the lowspeed supernatant of the fundic stomach homogenate [24,25]. Thefundic stomach from control, ulcerated, and melatonin-pretreatedindomethacin-treated mice were homogenized in 50 mM sodiumphosphate buffer, pH 7.4, in a Potter-Elvehjem glass homogenizerfor 2 min to get 20% homogenate. After centrifugation at 600� g for

K. Ganguly, S. Swarnakar / Biochimie 94 (2012) 2687e2698 2689
10 min, the proteins from 1.0 ml of the supernatant were precipi-tated with 10% trichloroacetic acid and allowed to react with 0.5 mlof 10 mM 2, 4, dinitrophenyl hydrazine for 1 h. After precipitationwith 20% trichloroacetic acid, the protein was washed thrice witha mixture of ethanol-ethyl acetate (1:1), dissolved in 1.0 ml ofa solution containing 6 M guanidine HCl in 20 mM potassiumphosphate adjusted to pH 2.3 with trifluroacetic acid followed bycentrifugation. The supernatant was read for carbonyl content at362 nm (e ¼ 22,000 M�1 cm�1).
2.6. Measurements of lipid peroxidation
The cytosolic and microsomal fractions from the gastric tissuehomogenate were used for measurement of lipid peroxide contentas thiobarbituric acid reactive species (TBARS) [25,26]. Briefly, 1 mlof the cytosolic fraction and 1 ml of microsomal fraction wereallowed to react with 2 ml of TCA-TBA-HCl (15% TCA, 0.375% TBA,0.25 N HCl) reagent, heated in a boiling water-bath for 15 min,cooled and centrifuged. The absorbance of the supernatant wasmeasured for nano moles of TBARS at 535 nm (e ¼ 1.5 � 105/M�1/cm�1).
2.7. Gelatin and casein zymography
For assay of MMP-9 and -2 activities, tissue extracts were elec-trophoresed in 8% SDS-polyacrylamide gel containing 1 mg/mlgelatin (Sigma), under non-reducing conditions. The gels werewashed twice in 2.5% Triton-X-100 (Sigma) and then incubated incalcium assay buffer (40 mM TriseHCl, pH 7.4, 0.2 M NaCl, 10 mMCaCl2) for 18 h at 37 �C. For assay of MMP-3 activity, tissue extractswere electrophoresed in casein (1 mg/ml) gel. Gels were stainedwith 0.1% coomassie blue followed by destaining (24e26). Thezones of gelatinolytic or caseinolytic activity came as negativestaining. Standard MMP-9 and -3 enzymes were purchased fromChemicon, Hampshire, UK. Quantification of zymographic bandswas done using densitometry linked to proper software (LabImage).
2.8. Preparation of nuclear extracts, Western blotting andelectrophoretic mobility shift assay (EMSA)
Control and chronic gastric ulcerated tissues were minced,hand-homogenized and centrifuged at 1000� g for 5 min at 4 �C.After washing with ice cold PBS, Mouse gastric tissues fromdifferent groups was minced in ice cold PBS (10 mM phosphatebuffer pH 7.4, 150 mM NaCl), homogenized and centrifuged at1000� g for 5 min at 4 �C. After washing once with ice cold PBS, cellpellets were suspended in 200 ml low salt buffer (10 mMHEPES pH-7.9, 1.5 mM MgCl2 and 10 mM KCl), incubated for 10 min on ice,followed by addition of 20 ml of 10% NP-40 solution and vigorousmixing for 30 s. Nuclei were collected by centrifugation andresuspending in 50 ml high salt buffer (20 mM HEPES pH-7.9,420 mM NaCl, 1.5 mM MgCl2, 0.2 mM EDTA, 25% glycerol). Bothbuffers were supplemented with protease inhibitors and 0.5 mMDTT before use. Nuclei were incubated for 15 min on ice, vortexedperiodically. Nuclear extracts were obtained by centrifugation at12,500� g for 10 min at 4 �C and stored at �80 �C. For preparationof cytosolic extracts the same procedure was followed except thatlow salt buffer was not removed by centrifugation and high saltbuffer was added in it [22]. Protein concentration was determinedby Bradford method.
2.8.1. Western blottingTissue extracts (cytosolic and nuclear) were mixed with 1�
reducing Lammeli sample loading buffer (0.0625 M TriseHCl (pH
6.8), 2% (w/v) SDS, 10% (v/v) glycerol, 0.05% BME, 0.001% (w/v)bromophenol blue) and were resolved by 8e10% SDS-polyacrylamide gel. The resolved proteins were then transferredto PDVF membranes (membrane was previously immersed inmethanol for 10e15 s followed by washing with distilled H2O for3e5 min) using Mini Trans-Blot apparatus (BioRad) at 250 mA for2.5 h. After transfer the membranes were blocked for 2 h at roomtemperaturewith 3% BSA in TBST (20mM TriseHCl, pH 7.4,150mMNaCl, and 0.02% Tween 20), followed by overnight incubation withdifferent antibodies (dilution range: 1:200 or 1:500 in TBST con-taining 3% BSA) at 4 �C under shaking conditions. The blots werewashed 4 times (15 min per wash) with 5e10 ml of TBST followedby incubation with alkaline phosphatase-conjugated secondaryantibody solution (dilution range: 1:2000 in TBST containing 3%BSA) for 2 h at room temperature under shaking conditions. Theblots were againwashed 4 times (15min per wash) with 5e10ml ofTBST followed by a last washing in 5e10 ml TBS for 10e15 minunder shaking condition at room temperature. After washing ofsecondary antibody, the blots were immersed in 1e2 ml of BCIP/NBT (5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazo-nium) substrate solution (Sigma) and kept in dark under shakingconditions for 15e20 min with intermittent observations. Once theband appears the reaction should immediately be stoppedby addition of distilled H2O. The blots shown in this study arerepresentative replicates selected from at least three experiments[24e26].
2.8.2. EMSAThe NF-kB specific sence: 50-AGTTGAGGGGACTTTCCCAGGC-30 &
antisence: 50-GCCTGGGAAAGTCCCCTCAACT-30 and AP-1 specificsence: 50-CTAGTGATGAGTCAGCCGGATC-30 and antisence: 50-GATCCGGCTGAGTCATCACTAG) oliginucleotides were used forEMSA. Single-stranded oligonucleotides were labeled with [g�32P]ATP (Amersham, Freiburg, Germany) in a buffer solution containingT4-polynucleotide kinase (MBI Fermentas, St Leon-Rot, Germany)and incubated at 37 �C for 45 min and then further incubated at65 �C for 10 min in a thermal cycler. To the solution 180 ml of doubledistilled water was added (200ml) followed by addition of 500 ml ofabsolute alcohol (2.5�) and 20 ml of 10 M ammonium acetate andmixed well. The total solution was then kept at �80 �C. Afterovernight incubation the tubes were centrifuged at 14,000� g for22 min at 4 �C and the supernatant was discarded. The pellet waswashed with 200 ml of 70% ethanol gently and centrifuged at14,000� g for 12 min at 4 �C. The supernatant was discarded andthe pellet was air dried for around 1.5 h, dissolved in 20 ml ofdeionized water and kept overnight at 4 �C. A twofold molar excessof unlabeled complementary oligonucleotides carried out ina thermal cycler with a following reaction condition (95 �C for10 min, 45 �C for 2 h and 25 �C for 30 min). The reaction was thenkept at room tempetature over night and directly used in EMSAassay. Binding reactions were performed for 45 min on ice in 20 mlbuffer (1 mM MgCl2; 0.5 mM EDTA; 0.5 mM DTT; 50 mM NaCl;10 mM TriseHCl, pH 7.5; 4% glycerol) containing 5 ng nuclearextract protein, 1 mg poly (dI:dC) and 10,000 cpm labeled oligo-nucleotide. Free [g�32P] ATP labeled oligonucleotides were also runas negative control. Reaction was stopped using gel-loading dye(Bromophenol blue in 20% glycerol). Binding complexes wereresolved by electrophoresis in vertical nondenaturing 7% poly-acrylamide gels, using 0.3� TBE as running buffer. Gels were driedand radioactive signals were detected by autoradiography films.
2.9. Reverse transcriptase-polymerase chain reaction (RT-PCR)
Total cellular RNA was extracted with TRIzol reagent accordingto the protocol provided by the manufacturer and quantified by

K. Ganguly, S. Swarnakar / Biochimie 94 (2012) 2687e26982690
measuring the absorbance at 260 nm. Complementary DNA wassynthesized using 1 mg of total RNA from each sample in a 20 mlreaction buffer using Superscript II Reverse Transcriptase with anoligo (dT)15 primer. Complementary DNA was amplified againstforward and reverse primers of MMP-9 (50-GACCCGAAGCGGA-CATTGTCAT-30 and 50-GGAATGATCTAAGCCCAGTGCAT-30; 755 bp),MMP-3 (50-GGATTGTGAATTATACACCGGAT-30 and 50-GGATAACCTGCTAGCTCCTCGT-30; 769 bp), MMP-2 (50-ATGGCTTCCTCTGGTGCT-30 and 50-TCGTAGTGGTTGTGGTTGC-30; 385 bp), and GAPDH(50TGGGGTGATGCTGGTGCTGAG-30 and 50-GGTTTCTCCAGGCGG-CATGTC-30; 497 bp) respectively. The reactions were subjected todenaturation (94 �C for 30 s), annealing (54 �Ce59.5 �C for 30 s),and extension (72 �C for 60 s) for 35 cycles [25,26]. The PCRproducts were fractionated on 2% agarose gels and visualized byethidium bromide staining.
2.10. Histology and immunofluorescence microscopy
For light microscopic investigations, the stomach of control andchronic gastric ulceration were dissected and sectioned into 2e3 mm2 pieces. The tissue samples were fixed in 10% formalin for48 h dehydrated in ascending alcohol series and embedded inparaffinwax. Approximately 5 mm thick serial sections were stainedwith hematoxylin and eosin (H&E) for general morphology. Theimages were assessed under an Olympus microscope (1�70). Forimmunofluorescence study antigen retrieval was performed bytrypsin (0.05% Trypsin, 0.1% CaCl2) followed by blocking with 5%BSAwith TBS (20 mM TriseHCl pH 7.4 containing 150 mMNaCl) for2 h. Then it was incubated overnight at 4 �C in primary antibodysolution (1:200 dilutions in TBS with 1% BSA) in a humid chamber.The sections were washed five times with TBST and then incubatedwith fluorescence isothiocyanate-conjugated secondary antibody(1:400 dilutions in TBS with 1% BSA) for 2 h at room temperatureand washed with TBST. Images were captured at 20X10, 40X10 and100X10 under an Olympus microscope (1�70) using Camediasoftware (E-20P 5.0 Megapixel) and processed using Adobe Pho-toshop version 7.0. Images were captured at 20� using Camediasoftware (E-20P 5.0 Megapixel) and processed using Adobe Pho-toshop version 6.0 [24,25].
2.11. Statistical analysis
The histographic representations were fitted using Sigma plot.Data were presented as the mean � SEM. Significance was calcu-lated using the Student’s Newman’s Keuels test by Graph padinstat-3 software (Germany) [25,26].
3. Results
3.1. Dose dependent effect of indomethacin on MMP-9 and -3activities during chronic gastric ulceration
Chronic gastric ulcers were developed in mice by oral adminis-trationof differentdoses of indomethacin (2, 5,10 and20mg/kgb.w.)for consecutive 3 days and were sacrificed on following day. Themacroscopic observation of gastric damagewas evident from lesionsappearing in stomach (data not shown). In order to test the changeson secreted MMP-9 and -3 activities, PBS extracts of stomach fromdifferent group of mice under treatment of varying doses of indo-methacin were subjected to gelatin and casein zymography respec-tively. Results shown in Fig. 1A and B indicated that gradual increasein both pro- and active forms of secreted MMP-9 (w108 kDa and97 kDa), and -3 (w56 kDa and 45 kDa) activities respectively withincrease indomethacin dose. The activity of secreted proMMP-9wasincreased byw1.2, 5.4,11.5 and 9.5 folds and, activeMMP-9 byw1.2,
32.5, 60.8 and 120 folds respectively following administration of 2, 5,10 and 20 mg/kg b.w. doses of indomethacin (Fig. 1C). We foundsimilar increase in secreted MMP-3 activity with increasing doses ofindomethacin. Fig. 1D shows that the activity of secreted proMMP-3was increased by w3.6, 6.4, 6.8 and 8.5 folds and active MMP-3 byw2.6, 58.8,104.5 and100.6 folds respectively at 2, 5,10 and20mg/kgb.w. indomethacin. A dose of 10 mg/kg b.w. indomethacin wasselected for the future experiments.
MMP-9 and -3 expressions in ulcerated and control stomachswere evaluated by Western blotting using anti-MMP-9 and -3antibodies (Fig. 1E). Fig. 1G shows that 2, 5, 10 and 20 mg/kg b.w.indomethacin doses caused an induction of w1.5, 2.2, 8.9 and 25folds of MMP-9 and w5, 12.5, 12.8 and �23 folds of MMP-3respectively. Surprisingly, MMP-3 expression was more than 5fold higher even at 2 mg/kg b.w. of indomethacin compared to thatof control. Western blot for GAPDHwas performed to confirm equalprotein loading in the above blots for each respective doses ofindomethacin (Fig. 1E). To analyze the dose dependent effect ofindomethacin on gene transcription of MMPs, equal amount ofcDNA from control and ulcerated tissues were subjected to RT-PCR.Fig. 1F and H show that 2, 5, 10 and 20 mg/kg b.w. of indomethacincaused an increment in gene expressions by w2.5, 3, 3.3 and 7.9folds of MMP-9 and w1.5, 2.0, 2.5 and 3.2 folds of MMP-3 respec-tively. The transcription of GAPDH gene in different tissues wasused for comparison of MMP gene transcription (Fig. 1H).
3.2. Time dependent effect of indomethacin on MMP-9 and -3activities and their regulation by transcription factor during chronicgastric ulceration
Results shown in Fig. 2A and B indicate that indomethacincaused a time dependent gradual increase in secreted MMP-9 and-3 activities. Herein, during chronic gastric ulceration the activitiesof secreted proMMP-9 was increased gradually and reachedmaximally by w12 folds. However, the ratio of pro- and activeMMP-9 was increased by w15 folds during day 3 of indomethacinadministration (Fig. 2C). Interestingly, we found a gradual increasein secreted pro-MMP-3 activity by w12 fold as that of MMP-9profile with respect of time during indomethacin treatment(Fig. 2A and B). The densitometry of zymographic bands in Fig. 2Dshow that the ratio of pro- and activeMMP-3were increased byw2and 5 folds respectively.
EMSA assay reveals that the binding of activated NF-kB complexwith NF-kB specific oligonucleotide sequences were increased withtime during chronic gastric ulceration (Fig. 2E). We also confirmedthe binding of the activated AP-1 transactivating complex with AP-1 specific oligonucleotide sequences using nuclear extract fromgastric tissues. We found that during ulceration the binding of theAP-1 specific nuclear proteins were gradually increased timedependently which was induced maximally at day 3 of chroniculceration (Fig. 2F).
3.3. Localizations of MMP-9 and MMP-3 in gastric tissues duringindomethacin-induced chronic gastric ulceration
The evaluation of hemorrhagic scars in tissues of indomethacin-treated and vehicle treated groups were made to H&E stainedhistological sections. The microscopic observation of control tissuereveals that the structure of gastric mucosa had an intact epitheliallayer and glandular cells with continuous gastric pits (Fig. 3A). Incontrast, the indomethacin-treated chronic gastric ulceratedtissues were found to have disrupted epithelial layer and glandularcells with discontinuous gastric pits (Fig. 3B). To decipher thecellular localization and functional aspects of MMP-9 and -3 in ECMremodeling during chronic gastric ulceration, tissue section of mice

Fig. 1. Changes in expressions and activities of MMP-9 and -3 with severity of chronic gastric ulceration. Chronic gastric ulceration was induced by oral administration of differentindomethacin doses (0, 2, 5, 10 and 20 mg/kg b.w.) to different groups of BALB/c mice for three consecutive days followed by sacrificing day after. In order to monitor the secretedMMP-9 and -3 activities, PBS extracts of stomach from different treatment were subjected to zymography. (A) Gelatin and (B) Casein zymography for detection of secreted MMP-9and -3 activities respectively using equal amount of PBS extracts (80 mg protein) as described in “Materials and Methods”. Graphical representation of indomethacin doses versusarbitrary activities of (C) pro- & active MMP-9 and (D) pro- & active MMP-3 in stomach. In order to test the severity dependent effect on secreted MMPs expression, equal amount(80 mg protein) of PBS extract from indomethacin-treated tissues were subjected to Western blot and probed with (E) polyclonal anti-MMP-9, antiMMP-3, anti-GAPDH antibody.Total RNA were extracted from control and different doses of indomethacin treated groups and were used for RT-PCR for the detection of (F) MMP-9, MMP-3 and GAPDH mRNAexpression. Graphical representation of doses of indomethacin versus fold expression at (G) protein and (H) mRNA levels of MMP-9 and -3. Activities were measured by ImageQUANT designed densitomety values from the above zymograms or blot and three other representative zymograms or blots from independent experiments. Results are reported asthe means � SEM where #, p < 0.01; *, p < 0.001 and ns ¼ non significant versus control, using Student’s Newman’s Keuels test to compare means.
K. Ganguly, S. Swarnakar / Biochimie 94 (2012) 2687e2698 2691
from day 3 ulceration were subjected to immunofluorescencemicroscopy with MMP-9 and -3 specific antibodies. It is evidentfrom Fig. 3(EeH) that bothMMP-9 and -3were localized tomucosalcells in control animals. However, the synthesis of both MMP-9 and-3 were increased during day 3 of chronic ulceration especially inmucosal cells Fig. 3(EeH). Herein, we found that during day 3 ofchronic ulceration indomethacin caused severe mucosal disruptiontherefore the increased MMP-9 and -3 were released in the laminapropria of gastric ECM.
3.4. Preventive effect of melatonin against ulceration andregulation of MMP-9 and -3 activities
We have used 10, 20 and 40 mg/kg b.w. of melatonin to findits effect on MMP-9 and -3 during prevention of gastric
ulceration. Equal amount of PBS extracts (80 mg) from respectivesamples were subjected to gelatin and casein zymography toassess MMP-9, and -3 activities. Fig. 4A and B show that mela-tonin was significantly effective in prevention of chronic gastriculcers that are associated with the attenuation of MMP-9, -3activities. Ulceration caused an increased activity of proMMP-9,and -3 by w22, and 115 folds respectively while melatonindose dependently down regulated the activities of MMPs tocontrol levels during prevention (Fig. 4C and D). The highest doseof melatonin (40 mg kg/b.w.) offered gastroprotection by atten-uating w90e95% of secreted MMP-9 and -3 activities (Fig. 4Cand D).
To test whether melatonin offers gastroprotection in chronicgastric ulceration, melatonin (40 mg/kg b.w.) was administered30 min prior to indomethacin treatment (10 mg/kg b.w./day) for 5

Fig. 2. Time dependent regulation of MMP-9 and -3 activities by AP-1 and NF-kB during chronic gastric ulceration. The experiment was performed as described in “Materials andMethod.” In order to test the time dependent effect on secreted MMP-9 and -3 activities, during chronic gastric ulceration, PBS extracts from different groups were subjected forgelatin and casein zymogram respectively. Equal amounts (80 mg) of PBS extracts from different groups of indomethacin-treated chronic gastric ulcerated tissues were subjected to(A) gelatin and (B) casein zymography respectively. Histographic representation of duration versus arbitrary activities of gelatinases (pro- and active MMP-9 and -2) in gelatin (C)and casein (D) zymograms. Activities were measured by using LabImage densitometry program from the above zymogram and three other representative zymograms fromindependent experiments in each case. Results are reported as the mean � SEM where *, p < 0.001 and ns ¼ non significant versus control using Student’s Newman’s Keuels test tocompare means. The nuclear extracts were isolated from different stomach samples and incubated with specific oligonucleotide sequences of NF-kB and AP-1 separately and wereassayed through electrophoretic mobility shift assay (EMSA). (E) and (F) showing the NF-kB and the AP-1 activity respectively by EMSA assay.
K. Ganguly, S. Swarnakar / Biochimie 94 (2012) 2687e26982692
consecutive days. Herein, the time dependent study was conductedby administration of vehicle for indomethacin treated groups andonly melatonin for melatonin pretreated groups and the experi-ment was continued up to 5 days. Equal amounts of PBS (80 mg)extracts from both indomethacin-treated and melatonin-pretreated tissues were subjected to gelatin and casein zymog-raphy respectively. Fig. 5A and C depict that with time indometh-acin caused a gradual induction in MMP-9 and -3 (both pro- andactive) activities. The densitometry analysis of zymographic bandsin Fig. 5B and D show w(6e9) and (1.6e1.9) folds induction inproMMP-9 and -3 activities and w(41e126) and (1.6e1.7) foldsinduction in active MMP-9 and -3 activities respectively with thetime of indomethacin treatment. The interesting fact was that theactivities of MMP-3were induced significantly in day 1whileMMP-9 induction which was increased maximally at day 9 of chroniculceration. Herein, melatonin showed significant gastroprotectionby gradual inhibition of total MMP-9 and -3 activities in a timedependent manner.
We performed RT-PCR analysis with indomethacin-treatedand melatonin pretreated gastric tissue samples to study tran-scription of MMP-9 and -3 during prevention of chronic ulcer-ation. Fig. 5E and G show that the expression of MMP-9 and -3mRNA were enhanced w(2e6) and (2e10) folds respectivelyduring the chronic ulceration. Herein, melatonin gradually sup-pressed the transcription of MMP-9 and -3 to control levelsduring prevention. Using real time-PCR, a direct quantitativemeasurement of MMP-9 and -3 transcriptions in ulcerated andmelatonin pretreated tissues were measured (Fig. 5F and H).The relative expression were enhanced w(2.5e12) and (4.5e15)folds respectively for MMP-9 and -3 during ulceration.Fig. 5F and H show that during prevention melatonin graduallysuppressed the transcriptions of both MMP-9 and -3 andthe value of the products were reached almost up to day 1 ofchronic ulceration. GAPDH gene was used as an internal controlagainst which MMP-9 signal was normalized (CtMMP-9�CtGAPDH ¼ DCt).
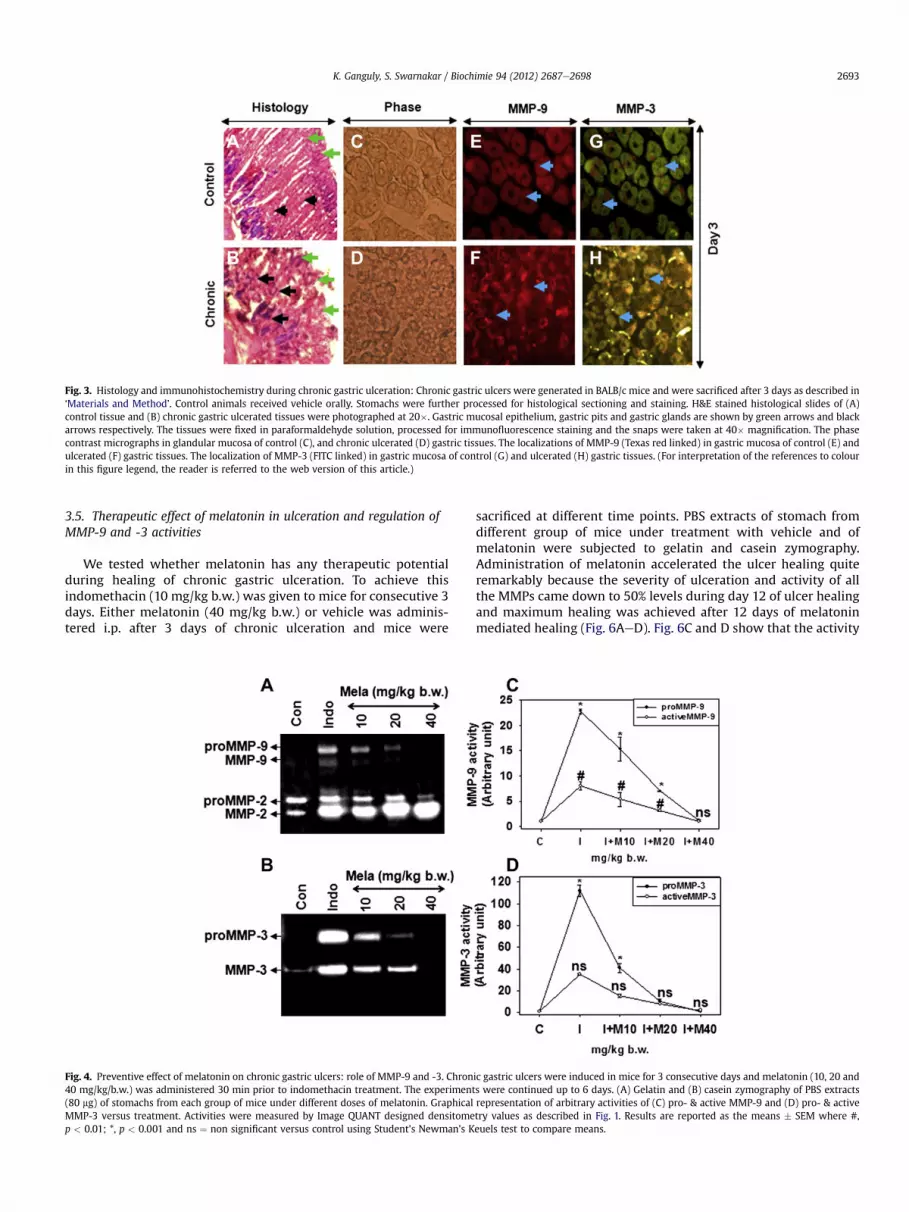
Fig. 3. Histology and immunohistochemistry during chronic gastric ulceration: Chronic gastric ulcers were generated in BALB/c mice and were sacrificed after 3 days as described in‘Materials and Method’. Control animals received vehicle orally. Stomachs were further processed for histological sectioning and staining. H&E stained histological slides of (A)control tissue and (B) chronic gastric ulcerated tissues were photographed at 20�. Gastric mucosal epithelium, gastric pits and gastric glands are shown by green arrows and blackarrows respectively. The tissues were fixed in paraformaldehyde solution, processed for immunofluorescence staining and the snaps were taken at 40� magnification. The phasecontrast micrographs in glandular mucosa of control (C), and chronic ulcerated (D) gastric tissues. The localizations of MMP-9 (Texas red linked) in gastric mucosa of control (E) andulcerated (F) gastric tissues. The localization of MMP-3 (FITC linked) in gastric mucosa of control (G) and ulcerated (H) gastric tissues. (For interpretation of the references to colourin this figure legend, the reader is referred to the web version of this article.)
K. Ganguly, S. Swarnakar / Biochimie 94 (2012) 2687e2698 2693
3.5. Therapeutic effect of melatonin in ulceration and regulation ofMMP-9 and -3 activities
We tested whether melatonin has any therapeutic potentialduring healing of chronic gastric ulceration. To achieve thisindomethacin (10 mg/kg b.w.) was given to mice for consecutive 3days. Either melatonin (40 mg/kg b.w.) or vehicle was adminis-tered i.p. after 3 days of chronic ulceration and mice were
Fig. 4. Preventive effect of melatonin on chronic gastric ulcers: role of MMP-9 and -3. Chron40 mg/kg/b.w.) was administered 30 min prior to indomethacin treatment. The experiment(80 mg) of stomachs from each group of mice under different doses of melatonin. GraphicalMMP-3 versus treatment. Activities were measured by Image QUANT designed densitomep < 0.01; *, p < 0.001 and ns ¼ non significant versus control using Student’s Newman’s K
sacrificed at different time points. PBS extracts of stomach fromdifferent group of mice under treatment with vehicle and ofmelatonin were subjected to gelatin and casein zymography.Administration of melatonin accelerated the ulcer healing quiteremarkably because the severity of ulceration and activity of allthe MMPs came down to 50% levels during day 12 of ulcer healingand maximum healing was achieved after 12 days of melatoninmediated healing (Fig. 6AeD). Fig. 6C and D show that the activity
ic gastric ulcers were induced in mice for 3 consecutive days and melatonin (10, 20 ands were continued up to 6 days. (A) Gelatin and (B) casein zymography of PBS extractsrepresentation of arbitrary activities of (C) pro- & active MMP-9 and (D) pro- & activetry values as described in Fig. 1. Results are reported as the means � SEM where #,euels test to compare means.

Fig. 5. Effect of melatonin on activity and mRNA expression of MMP-9, and -3 during prevention of chronic gastric ulcer: Experiment was performed as described in Fig. 1. (A)Gelatin and (C) casein zymography of PBS extracts (80 mg) of stomachs from each group of mice under different treatment. Graphical representation of arbitrary activities of (B) pro-& active MMP-9 and (D) pro- & active MMP-3 versus treatment. Total RNA was extracted from different groups of mice as described in ‘Materials and Method’ and was used for RT-PCR for the detection of MMP-9 (E), MMP-3 (G) and GAPDH mRNA expression. The densitomety values of fold increments in individual bands of MMP-9 and -3 as compared tocontrol were given under respective agarose gel images. An equal concentration (0.6 mg) of complementary cDNAwas used for real time-PCR analysis with specific primers of MMP-9, -3 and GAPDH mRNA. Histogram of relative expression of (F) GAPDH and MMP-9 mRNA and (H) GAPDH and MMP-3 mRNA specific products of different tissues where DRn valueswere plotted against cycle number. Results are reported as the means � SEM where *, p < 0.001 and ns ¼ non significant using Student’s Newman’s Keuels test to compare means.The data of fold expressions of different molecules of indomethacin-treated samples were compared with control samples and those for melatonin-treated samples withindomethacin-treated samples respectively.
K. Ganguly, S. Swarnakar / Biochimie 94 (2012) 2687e26982694
of proMMP-9 were increased by w(10e80) folds, and proMMP-3by w(40e120) folds respectively during auto healing. Herein,melatonin gradually suppressed the activities of MMP-9 and -3which were reached to control levels during healing of gastriculceration.
To study the therapeutic effect of melatonin on transcriptionand translation of MMPs (MMP-9 and -3) during healing of chronicgastric ulceration, we performed the western blot and RT-PCRanalysis with indomethacin treated and melatonin post-treatedindomethacin treated tissue samples. Fig. 6E and G showthat indomethacin caused a gradual induction of w4e14 folds ofMMP-9 and w6e10 folds of MMP-3 proteins respectively. Herein,melatonin significantly suppressed the increased expressions ofMMP-9, -3 during healing of chronic ulceration. The RT-PCR anal-ysis in Fig. 5F and H show that MMP-9 and -3 transcription wereenhanced w(2e6), (4e6) folds respectively during the chroniculceration phase. Herein, melatonin suppressed their transcriptionduring ulcer prevention.
3.6. Melatonin protects chronic gastric ulceration by blocking ROSand inhibiting inflammation guided NF-kB and AP-1 signalingpathway
We asked whether melatonin has any role to prevent the gastriculceration by inhibiting ROS production. Table 1 show that proteincarbonyl content in gastric tissues was increased by w3 fold,whereas lipid peroxidation in cytosolic and microsomal fractionswere increased byw10 and 13.5 folds respectively during day 1 andalmost remain unchanged till day 3e9 of ulcerogen application.Melatonin showed effective protection of protein carbonyl forma-tion by inhibiting w40% during day 1 andw68% during day 6e9 ofchronic ulceration (Table 1). Similarly it inhibited both cytosolicand microsomal lipid peroxidation by w87% and w66% respec-tively during day 1 and then byw93% andw 87% during day 6e9 ofchronic ulceration (Table 1). As several antioxidant enzymes areresponsible for conversion of the ROS, we herein looked at proteinlevel of two major antioxidant enzymes i.e. super oxide dismutase

Fig. 6. Effect of melatonin on changes in activity, mRNA and protein expressions of MMP-9 and -3 during prevention of chronic gastric ulcers. Chronic gastric ulcers were induced inmice for three consecutive days and melatonin (40 mg/kg/b.w.) was administered 30 min prior to indomethacin that were continued up to 9 days. (A) Gelatin and (B) caseinzymography of PBS extracts (80 mg protein) of stomachs from each group of mice as described in Fig. 1A. Graphical representation of activity of (C) pro- & active MMP-9 and (D) pro-& active MMP-3 in PBS extracts versus days of treatment. In order to test the MMPs expression, PBS extracts of stomach samples were subjected for Western blot analysis. Equalamount protein of PBS extract from stomach tissues were subjected to Western blot and probed with (E) polyclonal anti-MMP-9, antiMMP-3, anti-GAPDH antibody. Total RNA fromgastric tissues were extracted from different stomach tissues of mice and were used for RT-PCR for the detection of (F) MMP-9, MMP-3 and GAPDH mRNA expression. Histogram ofrelative expression of (G) MMP-9 and -3 proteins and (H) MMP-9 and -3 mRNAs are plotted against days of ulceration and healing. Band intensities were measured by Image QUANTdesigned densitometry values as described in Fig. 1. Results are reported as the means � SEM where *, p < 0.001 and ns ¼ non significant versus control using Student’s Newman’sKeuels test to compare means.
Table 1Effect of melatonin on prevention of protein oxidation and lipid peroxidation duringprevention of chronic gastric ulceration. Indomethacin was administered orally todifferent groups of mice at a dose of (10 mg/kg b.w./day) for generating chronicgastric ulceration and mice were sacrificed on day 1, 3, 6 and 9. No treatment meansthat animals received vehicle orally. Protein carbonyl estimation (n ¼ 18), and lipidperoxidation (n ¼ 12) from both cytosolic fractions were measured as described inthe “Materials and Method”. Results are reported as the means � SEM. *p < 0.001versus none using StudenteNewmaneKeuls test to compare means.
Treatment Day 1 Day 3 Day 6 Day 9
Protein oxidation (nmol/mg protein)Control 1.22 � 0.03 1.21 � 0.45 1.11 � 0.02 1.09 � 0.18Indomethacin 3.82 � 0.20* 3.42 � 0.03* 3.22 � 0.20* 2.12 � 0.02*þ Melatonin 2.22 � 3.31* 2.04 � 2.22* 1.82 � 2.11* 1.42 � 2.31*Lipid peroxidation (nmol/mg protein)Control 0.43 � 0.02 0.32 � 0.03 0.21 � 0.03 0.11 � 0.068Indomethacin 0.90 � 0.04* 0.70 � 0.003* 0.50 � 0.08* 0.44 � 0.003*þ Melatonin 0.40 � 0.31* 0.30 � 0.52* 0.22 � 0.01* 0.10 � 0.01*
K. Ganguly, S. Swarnakar / Biochimie 94 (2012) 2687e2698 2695
(SOD-2) and catalase using PBS extracts of gastric tissue samples(Fig. 7A). The densitometry of Western blot analysis in Fig. 7A showthat the expression of both SOD-2 and catalase were induced w2.5and 3 folds respectively during day 1 but significantly down-regulated during day 3e9 of chronic gastric ulceration. Interest-ingly, melatonin showed a biphasic regulation of these twoenzymes i.e. during day 1e3 catalase expressions decreased almostto control level and then gradually increased by w1.7 folds duringday 3e9 of ulceration while expression of SOD-2 decreased grad-ually to control level during day 3e9 of ulcer prevention (Fig. 7D).
Because chronic ulcers are accompanied by tissue inflammationand cytokine production, we tested expression of proinflammatroymolecules in ulcerated gastric tissues with or without melatonintreatment. PBS extracts of stomach from different groups of mice atdifferent days of indomethacin treatment were subjected toWestern blot analysis. Fig. 7A depicts that with increasing time

Fig. 7. Effect of melatonin on signaling molecules and transcription factors duringprevention of chronic gastric ulceration. The time dependent chronic ulceration wasperformed as described in Fig. 3. Cytosolic and nuclear extracts were prepared fromdifferent groups of tissues according to the “Materials and Method”. The cytosolic (A)and nuclear (B) extracts were subjected to Western blot and probed with polyclonalantibodies. Cytosolic extracts (80 mg protein) from different gastric tissue samples weresubjected to Western blot and probed with polyclonal anti-TNFa, anti-IL-1b, anti-IL-8,anti SOD2, anti-catalase and anti-GAPDH antibodies while nuclear extracts wereprobed with anti-NF-kB (p65), anti p-JNK, anti-pERK1/2, anti-cFos, anti-pcJun, anti-H4antibodies. Expressions were measured by Image QUANT designed densitometryvalues as described in Fig. 2B. The densitometry values of fold increments of individualbands were given under respective Western blot images.
K. Ganguly, S. Swarnakar / Biochimie 94 (2012) 2687e26982696
there was a gradual induction in TNF-a, IL-1b and IL-8 expressions.The analysis of protein bands in Fig. 6A show that secreted TNF-a,IL-1b and IL-8 expressions were increased byw(11.5e13), (2.7e9.5)and (9.4e13.5) folds respectively during day 1e3 and then gradu-ally suppressed toward control value during day 6e9 of ulceration.While melatonin suppressed w(50e60)% expressions of proin-flammatory molecules during day 3 and then gradually to controllevels during day 6e9 of ulcer prevention (Fig. 7A).
It is evident that the expressions of MMPs were primarilyincreased due to inflammation at early phases of ulceration (day 1e3) however sustained production of ROS is vital for further upre-gulation during later phases (day 6e9). As inflammation and ROSboth are responsible for modulation of several signaling moleculesand transcription factors, we addressed their association with
chronic ulceration and melatonin’s action thereon. Western blots(Fig. 7B) reveal that the expression and nuclear translocation of p-65 subunits of NF-kB transcription factor increased by w5 and 4.5folds respectively on day 1 which then decreased to control levelson day 6 onwards of ulceration. In melatonin-mediated protectionstudies, the cytosolic and nuclear expression of NF-kB wasincreased by w4.7 and 2.3 folds respectively on day 1 and thensignificantly suppressed during day 6 (Fig. 7B). Herein, we elucidatethe role of MAP kinases, especially ERK1/2 and JNK in regulatingMMPs expression. Ulceration caused a gradual induction of phos-phorylated JNK (Thr-183, Tyr-185) by w6e14 folds and phosphor-ylated ERK1/2 (Tyr-204) by 7e11.5 folds whereas melatoninsignificantly suppressed the phosphorylation (w 60e80%) of JNKand ERK1/2 proteins (Fig. 7B). As the expression of c-Fos and c-Junare dependent upon the specific binding of the ERK1/2 and JNKswith the promoter sites of c-Fos and c-Jun genes, we next analyzedthe translocation of AP-1 into the nucleus in the presence andabsence of melatonin. The densitomety analysis of Fig. 7B showsthat nuclear translocation of p-JNK were increased byw9e12 foldsand p-ERK1/2 by w5e21 folds during chronic ulceration whichwere significantly blocked by melatonin. The MMP promoterscontain AP-1 binding motif, we therefore examined the effect ofmelatonin on the expression and nuclear translocation of AP-1complex (cFos and cJun) during protection of gastric ulceration.The expressions of cFos were upregulated by w3e6 folds andphosphorylated-c-Jun (Ser-73) by w8e15 folds whereas nucleartranslocations were enhanced by w1.5e5 folds and w2e21 foldsrespectively during 3e9 days of chronic ulceration (Fig. 7B). Herein,melatonin not only suppressed the expression of cFos and pc-junbut it also attenuated the nuclear translocation by w3e50 foldsand w1.5e45 folds respectively during day 1e6 and blocked thetranslocation almost to control value on day 9. Western blot forGAPDH and histone (H)4 were performed to confirm equal proteinloading of cytosolic and nuclear extracts respectively (Fig. 7Aand B).
4. Discussion
It is well known that H. pylori induced gastric inflammation isassociated with upregulation of MMPs i.e., MMP-2, -3, -7 and -9[3,27,28]. Our previous reports describe that upregulation of MMP-9 expression and activities are associated with ethanol andindomethacin-induced acute gastric ulceration [24e26]. Moreover,human gastric ulcers are chronic in nature and occur due toexcessive tissue inflammation and aging of wounds for days wherethe expressions of MMPs are altered depending on severity [29e31]. However, the mechanisms of ulceration in context ofsignaling by keeping special emphasis on MMPs transcriptionwerenot well clarified. The major objective of this study was to under-stand the regulation of MMPs especially MMP-9 and -3 transcrip-tions in gastric tissues during chronic gastric ulceration. Herein, wealso explored the mechanism of ulcer healing after melatonintreatment and investigated its role on MMP-9 and -3 transcriptionsand associated signaling pathway in mice gastric tissue. The resultsshow that during chronic gastric ulceration the transcription,translation and activity of MMP-9 and -3 were significantlyincreased in gastric tissues with dose and duration of indomethacintreatment whereas melatonin offered gastroprotection by downregulating transcription and translation of MMP-9 and -3.
Several reports have documented that MMPs regulation occur atthe level of zymogen activation apart from transcriptional levelsand, the activatedMMPs aremainly responsible for the degradationof ECM [29,31,32]. It has also been described that during chronicinflammation most of the proMMPs are often converted into activeMMPs with a nonreversible phenomenon which lead to

K. Ganguly, S. Swarnakar / Biochimie 94 (2012) 2687e2698 2697
malignancy [29e31]. The present study indicates that indometh-acin caused a gradual increase in MMP-3, -9 activities with severityof inflammation. The immunofluorescence study suggest that thelocalization of bothMMP-9 and -3were confined tomucosal cells ofcontrol tissues but were released in ECM during chronic ulceration.Herein, our study for the first time suggest that gastric ulceration ispropagated by the induction of endogenous MMPs activation andrelease from the gastric mucosal cells that may result in ECMdegradation of gastric tissues. It is also known that serine proteaseand extracellular plasmin activates MMP-1 and MMP-3 [20,32].Herein, we speculate that the plasmin-mediated MMP-3 inductionmight be occurring in vivo for MMP-3 activation during the firstphase (day 1e3) of chronic gastric ulceration. Based on in vitrokinetic and catalytic parameters, MMP-3 appears to be the mostefficient activator of proMMP-9 and may be a natural activatorin vivo when intracellular ROS is a critical factor [33,34]. We foundthat chronic gastric ulceration is associated with excessive ROSproduction along with increased MMP-9 and -3 activations (Fig. 1,Table 1). We speculate that the availability of intracellular orextracellular ROS might be a causative factor for the activation ofMMP-9 and -3. It is evidenced from the study of Hahn-Dantonaet al. [32] that the conversion of the proMMP-9 into MMP-9occurs by MMP-3 and uPA-plasmin mediated process. In ourpresent study, we found a significant activation of MMP-9 duringthe later phases (day 6e9) of chronic ulceration suggesting that theexcess amount of MMP-3 during the onset of ulceration might beresponsible for proMMP-9 activation in later phases (day 6e9).
Chronic inflammation can be a significant source of intracellularROS production, that has been implicated as one of the criticalsignalingmolecules influencingMMPs transcription [23,35,36]. Thesteady-state concentration of H2O2 is maintained by the bufferingcapacity of the mitochondrial glutathione redox system in vivo [23].However, when Sod2 transcription level is increased by pro-inflammatory cytokines, the glutathione-buffering capacity of themitochondria is disrupted [23,37e39]. We found an elevated SOD2and catalase level during the onset (day 1e3) of ulceration underthe influence of pro-inflammatory cytokine production (TNF-a, IL-1b & IL-8) and ROS generation (Fig. 7A). Herein, Melatonin blockedthe expression of both enzymes as an antioxidant and anti-inflammatory molecule. Previous reports have indicated thatMMPs are differentially regulated by ROS that are emerging as keyeffectors in signal transduction [40e46]. ROS, either directly orindirectly plays a role in enhancing the inflammation through theactivation of stress kinases (JNK, MAPK, p38) and redox sensitivetranscription factors such as NF-kB and AP-1 [23]. Although,nuclear extracts prepared from tissues are of poor quality, the EMSAassay in our present study revealed that chronic inflammation byindomethacin increased translocation of NF-kB and AP-1 specifictranscription factors and DNA binding activity with NF-kB and AP-1specific sequence in a time dependent manner. The result of ourpresent study also corroborate with in vitro studies where it wasconfirmed that any kind of ulcerogen like H. pylori or indomethacincaused NF-kB and/or AP-1 dependent DNA binding activity witha dose and time dependent manner [45,46]. We found that ulcer-ation (day 1e3) caused phosphorylation of both ERK1/2 and JNKand theywere significantly induced during the later phases (day 6e9) of ulceration. JNK phosphorylation leads to activation of the AP-1member’s c-Jun and JunB while ERK1/2 phosphorylation leads tothe activation of c-Jun, c-Fos which finally leads to MMP tran-scription [23]. We found the enhanced expression and nucleartranslocation of JNK-dependent c-Jun and ERK1/2 dependent c-Fosduring chronic ulceration that was blocked byMelatonin. As severalproinflammatory cytokines and growth factors are primarilyattributed to the induction of specific MMPs [23], we investigatedthe expression of IL-1b, IL-8 and TNF-a, which were significantly
induced during ulceration (day 1e3) and then were graduallydownregulated to basal levels at later phases (day 3e9) of ulcera-tion. As TNF-a and IL-1b have been implicated in the activation ofNF-kB [23], we investigated whether NF-kB is responsible forregulation of MMPs in the presence and absence of melatonin. Wefound that both the expressions and nuclear translocation of p-65subunit of NF-kB were increased in day 3 of ulceration whereasmelatonin significantly blocked the expressions and nucleartranslocation of NF-kB. Melatonin herein blocked TNF-a, IL-1b andIL-8 expressions and reduced the nuclear translocation of NF-kBand in turn attenuated transcription of MMP-9 and -3 duringgastroprotection.
In summary, the study showed that MMP-9 and -3 expressionsand activities were highly dependent upon severity of inflamma-tion. The other major cause might be differential redox-mediatedNF-kB and AP-1 signaling during chronic ulceration. Herein,melatonin reduced the expression of MMP-9 and -3 by blockinginflammation in one hand and redox signaling in the other handduring ulcer healing. Melatonin also offered a significant gastro-protection by preventing JNK and ERK1/2 phosphorylation alongwith reduced activation of AP-1 and henceforth suppressing MMP-9 and -3 transcriptions. Because MMPs can be both beneficial inregulating host defense and pathological inflammation [28e30], itis essential to understand the specific molecular mechanisms bywhich individual MMPs function in inflammatory processes. Redoxregulation of MMP gene expression has emerged as a novel sub-discipline in molecular biology that has promising therapeuticimplications in both pro- and anti- inflammatory processes.However, further understanding of melatonin’s action may identifytherapeutic strategies via MMP regulation in disease cure.
Acknowledgments
KG acknowledged the receipt of Senior Research Fellowshipfrom Council of Scientific and Industrial Research, New Delhi. Workis supported by NTRF grant. Authors are grateful to Prof. SiddharthaRoy, Director CSIR-IICB for overall support.
References
[1] R.M. Peek Jr., M.J. Blaser, Helicobacter pylori and gastrointestinal tract adeno-carcinomas, Natl. Rev. Cancer 2 (2002) 28e37.
[2] M.A. Garcia-Gonzalez, A. Lanas, P.H. Savelkoul, S. Santolaria, R. Benito,J.B. Crusius, A.S. Pena, Association of interleukin 1 gene family polymorphismswith duodenal ulcer disease, Clin. Exp. Immunol. 134 (2003) 525e531.
[3] M. Gooz, M. Shaker, P. Gooz, A.J. Smolka, Interleukin 1beta induces gastricepithelial cell metalloproteinase secretion and activation during Helicobacterpylori infection, Gut 52 (2003) 1250e1256.
[4] D. Basso, N. Gallo, C.F. Zambon, F. Navaglia, E. Stockreiter, F. Di Mario,M. Rugge, M. Plebani, Different effects of H. pylori water extracts on cytokines,pepsinogen C and gastrin mucosal release in patients with or withoutduodenal ulcer, J. Med. 32 (2001) 97e112.
[5] T.A. Miller, Protective effects of prostaglandins against gastric mucosaldamage: current knowledge and proposed mechanisms, Am. J. Physiol. 245(1983) G601eG623.
[6] Y. Fujii, T. Matsura, M. Kai, H. Matsui, H. Kawasaki, K. Yamada, Mitochondrialcytochrome c release and caspase-3-like protease activation duringindomethacin-induced apoptosis in rat gastric mucosal cells, Proc. Soc. Exp.Biol. Med. 224 (2000) 102e108.
[7] J.A. Imlay, Pathways of oxidative damage, Annu. Rev. Microbiol. 57 (2003)395e418.
[8] J.M. McCord, I. Fridovich, The utility of superoxide dismutase in studying freeradical reactions. II. The mechanism of the mediation of cytochromec reduction by a variety of electron carriers, J. Biol. Chem. 245 (1970) 1374e1377.
[9] B. Halliwell, J.M. Gutteridge, Formation of thiobarbituric-acid-reactivesubstance from deoxyribose in the presence of iron salts: the role of super-oxide and hydroxyl radicals, FEBS Lett. 128 (1981) 347e352.
[10] S. Yamagiwa, Y. Yoshida, R.C. Halder, A. Weerasinghe, S. Sugahara, H. Asakura,T. Abo, Mechanisms involved in enteropathy induced by administrationof nonsteroidal anti-inflammatory drugs (NSAIDS), Dig. Dis. Sci. 46 (2001)192e199.

K. Ganguly, S. Swarnakar / Biochimie 94 (2012) 2687e26982698
[11] T. Miura, S. Muraoka, Y. Fujimoto, Lipid peroxidation induced by indometh-acin with horseradish peroxidase and hydrogen peroxide: involvement ofindomethacin radicals, Biochem. Pharmacol. 63 (2002) 2069e2074.
[12] M. Ushio-Fukai, R.W. Alexander, M. Akers, K.K. Griendling, p38 Mitogen-activated protein kinase is a critical component of the redox-sensitivesignaling pathways activated by angiotensin II. Role in vascular smoothmuscle cell hypertrophy, J. Biol. Chem. 273 (1998) 15022e15029.
[13] Y.S. Bae, S.W. Kang, M.S. Seo, I.C. Baines, E. Tekle, P.B. Chock, S.G. Rhee,Epidermal growth factor (EGF)-induced generation of hydrogen peroxide.Role in EGF receptor-mediated tyrosine phosphorylation, J. Biol.Chem. 272(1997) 217e221.
[14] C. Lafon, M. Mathieu, O. Guerrin, O. Pierre, S. Vidal, A. Valette, Transforminggrowth factor beta 1-induced apoptosis in human ovarian carcinoma cells:protection by the antioxidant N-acetylcysteine and bcl-2, Cell. Growth Differ.7 (1996) 1095e1104.
[15] R.J. Reiter, D.X. Tan, J.C. Mayo, R.M. Sainz, J. Leon, D. Bandyopadhyay, Neurally-mediated and neurally-independent beneficial actions of melatonin in thegastrointestinal tract, J. Physiol. Pharmacol. 54 (2003) 113e125.
[16] K. Ganguly, P. Maity, R.J. Reiter, S. Swarnakar, Effect of melatonin on secretedand induced matrix metalloproteinase-9 and -2 activities during preventionof indomethacin-induced gastric ulcer, J. Pineal. Res. 39 (2005) 307e315.
[17] D. Bilici, H. Suleyman, Z.N. Banoglu, A. Kiziltunc, B. Avci, A. Ciftcioglu, S. Bilici,Melatonin prevents ethanol-induced gastric mucosal damage possibly due toits antioxidant effect, Dig. Dis. Sci. 47 (2002) 856e861.
[18] P.C. Konturek, S.J. Konturek, J. Majka, M. Zembala, E.G. Hahn, Melatoninaffords protection against gastric lesions induced by ischemia-reperfusionpossibly due to its antioxidant and mucosal microcirculatory effects, Eur. J.Pharmacol. 322 (1997) 73e77.
[19] W.C. Parks, R.P. Mecham, Matrix Metalloproteinases, Academic Press, NewYork, 1998.
[20] R. Visse, H. Nagase, Matrix metalloproteinases and tissue inhibitors of met-alloproteinases: structure, function, and biochemistry, Circ. Res. 92 (2003)827e839.
[21] V.M. Kahari, U. Saarialho-Kere, Matrix metalloproteinases and their inhibitorsin tumor growth and invasion, Ann. Med. 31 (1999) 34e45.
[22] G. Liu, Y.J. Park, E. Abraham, Interleukin-1 receptor-associated kinase (IRAK)-1-mediated NF-kB activation requires cytosolic and nuclear activity, Faseb. J.14 (2008) 14.
[23] K.K. Nelson, J.A. Melendez, Mitochondrial redox control of matrix metal-loproteinases, Free Radic. Biol. Med. 37 (2004) 768e784.
[24] L.P. Singh, P. Kundu, K. Ganguly, A. Mishra, S. Swarnakar, Novel role offamotidine in downregulation of matrix metalloproteinase-9 during protec-tion of ethanol-induced acute gastric ulcer, Free Radic. Biol. Med. 43 (2007)289e299.
[25] S. Swarnakar, K. Ganguly, P. Kundu, A. Banerjee, P. Maity, A.V. Sharma, Cur-cumin regulates expression and activity of matrix metalloproteinases 9 and 2during prevention and healing of indomethacin-induced gastric ulcer, J. Biol.Chem. 280 (2005) 9409e9415.
[26] K. Ganguly, P. Kundu, A. Banerjee, R.J. Reiter, S. Swarnakar, Hydrogenperoxide-mediated downregulation of matrix metalloprotease-2 inindomethacin-induced acute gastric ulceration is blocked by melatonin andother antioxidants, Free Radic. Biol. Med. 41 (2006) 911e925.
[27] H.C. Crawford, U.S. Krishna, D.A. Israel, L.M. Matrisian, M.K. Washington,R.M. Peek Jr., Helicobacter pylori strain-selective induction of matrixmetalloproteinase-7 in vitro and within gastric mucosa, Gastroenterology 125(2003) 1125e1136.
[28] N. Mori, H. Sato, T. Hayashibara, M. Senba, R. Geleziunas, A. Wada,T. Hirayama, N. Yamamoto, Helicobacter pylori induces matrixmetalloproteinase-9 through activation of nuclear factor kappa B, Gastroen-terology 124 (2003) 983e992.
[29] Y. Otani, Y. Sakurai, K. Kameyama, N. Igarashi, T. Yokoyama, T. Kubota,K. Kumai, M. Kitajima, Matrix metalloproteinase gene expression in chronicgastric ulcer: a potential role of eosinophils in perforation, J. Clin. Gastro-enterol. 25 (1997) S101eS104.
[30] T. Yokoyama, Y. Otani, N. Kurihara, Y. Sakurai, K. Kameyama, H. Suzuki,N. Igarashi, M. Kimata, N. Wada, T. Kubota, K. Kumai, M. Kitajima, Matrixmetalloproteinase expression in cultured human gastric wall fibroblastseinteractions with Helicobacter pylori isolated from patients with ulcers,Aliment. Pharmacol. Ther. 14 (2000) 193e198.
[31] M. Menges, C.C. Chan, M. Zeitz, A. Stallmach, Higher concentration of matrix-metalloproteinase 1 (interstitial collagenase) in H. pylori-compared to NSAID-induced gastric ulcers, Z. Gastroenterol. 38 (2000) 887e891.
[32] E. Hahn-Dantona, N. Ramos-DeSimone, J. Sipley, H. Nagase, D.L. French,J.P. Quigley, Activation of proMMP-9 by a plasmin/MMP-3 cascade in a tumorcell model. Regulation by tissue inhibitors of metalloproteinases, Ann. N.Y.Acad. Sci. 878 (1999) 372e387.
[33] C.S. He, S.M. Wilhelm, A.P. Pentland, B.L. Marmer, G.A. Grant, A.Z. Eisen,G.I. Goldberg, Tissue cooperation in a proteolytic cascade activating humaninterstitial collagenase, Proc. Natl. Acad. Sci. 86 (1989) 2632e2636.
[34] Y. Ogata, J.J. Enghild, H. Nagase, Matrix metalloproteinase 3 (stromelysin)activates the precursor for the human matrix metalloproteinase 9, J. Biol.Chem. 267 (1992) 3581e3584.
[35] P. Brenneisen, K. Briviba, M. Wlaschek, J. Wenk, K. Scharffetter-Kochanek,Hydrogen peroxide (H2O2) increases the steady-state mRNA levels ofcollagenase/MMP-1 in human dermal fibroblasts, Free Radic. Biol. Med. 22(1997) 515e524.
[36] G.A. Visner, W.C. Dougall, J.M. Wilson, I.A. Burr, H.S. Nick, Regulation ofmanganese superoxide dismutase by lipopolysaccharide, interleukin-1, andtumor necrosis factor, Role in the acute inflammatory response, J. Biol. Chem.265 (1990) 2856e2864.
[37] Y. Xu, K.K. Kiningham, M.N. Devalaraja, C.C. Yeh, H. Majima, E.J. Kasarskis,D.K. St Clair, An intronic NF-kappaB element is essential for induction of thehuman manganese superoxide dismutase gene by tumor necrosis factor-alpha and interleukin-1beta, DNA Cell. Biol. 18 (1999) 709e722.
[38] A. Satomi, S. Murakami, T. Hashimoto, K. Ishida, M. Matsuki, M. Sonoda,Significance of superoxide dismutase (SOD) in human colorectal cancer tissue:correlation with malignant intensity, J. Gastroenterol. 30 (1995) 177e182.
[39] A.C. Ranganathan, K.K. Nelson, A.M. Rodriguez, K.H. Kim, G.B. Tower,J.L. Rutter, C.E. Brinckerhoff, T.T. Huang, C.J. Epstein, J.J. Jeffrey, J.A. Melendez,Manganese superoxide dismutase signals matrix metalloproteinase expres-sion via H2O2-dependent ERK1/2 activation, J. Biol. Chem. 276 (2001) 14264e14270.
[40] M.H. Pillinger, N. Marjanovic, S.Y. Kim, J.U. Scher, P. Izmirly, S. Tolani,V. Dinsell, Y.C. Lee, M.J. Blaser, S.B. Abramson, Matrix metalloproteinasesecretion by gastric epithelial cells is regulated by E prostaglandins andMAPKs, J. Biol. Chem. 280 (2005) 9973e9979.
[41] J.A. Mengshol, M.P. Vincenti, C.I. Coon, A. Barchowsky, C.E. Brinckerhoff,Interleukin-1 induction of collagenase 3 (matrix metalloproteinase 13) geneexpression in chondrocytes requires p38, c-Jun N-terminal kinase, andnuclear factor kappa B: differential regulation of collagenase 1 and collage-nase 3, Arthritis Rheum. 43 (2000) 801e811.
[42] S.H. Yang, A.D. Sharrocks, A.J. Whitmarsh, Transcriptional regulation by theMAP kinase signaling cascades, Gene 320 (2003) 3e21.
[43] J.A. Frost, T.D. Geppert, M.H. Cobb, J.R. Feramisco, A requirement for extra-cellular signal-regulated kinase (ERK) function in the activation of AP-1 by Ha-Ras, phorbol 12-myristate 13-acetate, and serum, Proc. Natl. Acad. Sci. 91(1994) 3844e3848.
[44] Z. Han, D.L. Boyle, L. Chang, B. Bennett, M. Karin, L. Yang, A.M. Manning,G.S. Firestein, c-Jun N-terminal kinase is required for metalloproteinaseexpression and joint destruction in inflammatory arthritis, J. Clin. Invest. 108(2001) 73e81.
[45] P. Kundu, R. De, I. Pal, A.K. Mukhopadhyay, D.R. Saha, S. Swarnakar, Curcuminalleviates matrix metalloproteinase-3 and -9 activities during eradication ofHelicobacter pylori infection in cultured cells andmice, PLoSOne 6 (2011) e16306.
[46] C.Y. Ock, Y.J. Lim, Y.J. Kim, J.W. Chung, K.A. Kwon, J.H. Kim, K.B. Hahm, Acidpump antagonist provoked HSP27 dephosphorylation and accentuationrescues Stomach from indomethacin-induced damages, J. Dig. Dis. 12 (2011)71e81.





























