Embed Size (px)
Citation preview

Gene 466 (2010) 26–35
Contents lists available at ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r.com/ locate /gene
Cloning, characterization and subcellular localization of a Trypanosoma cruziargonaute protein defining a new subfamily distinctive of trypanosomatids
Maria R. Garcia Silva a,1, Juan P. Tosar a,1, Magali Frugier b, Sergio Pantano c, Braulio Bonilla a, Luis Esteban d,Esteban Serra d, Carlos Rovira e, Carlos Robello f,g, Alfonso Cayota a,h,⁎a Functional Genomics Unit, Institut Pasteur de Montevideo, Mataojo 2020 CP11400 Montevideo, Uruguayb Architecture et Réactivité de l'ARN, Université de Strasbourg, CNRS, IBMC, 15 rue René Descartes, 67084 Strasbourg Cedex, Francec Molecular Simulations Unit, Institut Pasteur de Montevideo, Mataojo 2020 CP11400 Montevideo, Uruguayd Instituto de Biología Molecular y Celular de Rosario, FCByF, UNR. Suipacha 531 - S2002LRK - Rosario, Argentinae Department of Oncology, Lund University, BMC C13, 221 84 Lund, Swedenf Molecular Biology Unit, Institut Pasteur de Montevideo, Mataojo 2020 CP11400 Montevideo, Uruguayg Department of Biochemistry, Faculty of Medicine, Gral Flores 2125, CP11800, Montevideo, Uruguayh Department of Medicine, Faculty of Medicine, Av Italia s/n CP11800, Montevideo, Uruguay
Abbreviations: sRNA, small RNAs; siRNA, small interfpiRNA, Piwi-associated RNA; WAGO, worm specific ainterference; RISC, RNA-induced silencing complex; PIWtestis; PAZ, Piwi/Argonaute/Zwille; ORF, open readingtide/oligosaccharide binding folds; P-bodies, Processingbrucei AGO protein; LmPIWI, Leishmania major PIWI protPIWI protein; pAGO, prokaryote Argonaute protein; LIT⁎ Corresponding author. Functional Genomics Unit, In
Mataojo 2020 CP11400 Montevideo, Uruguay. Tel.: +5985.
E-mail addresses: [email protected] (M.R. [email protected] (J.P. Tosar), [email protected]@pasteur.edu.uy (S. Pantano), bonilla@[email protected] (L. Esteban), [email protected]@med.lu.se (C. Rovira), [email protected]@pasteur.edu.uy (A. Cayota).
1 These authors contributed equally to this work.
0378-1119/$ – see front matter © 2010 Elsevier B.V. Adoi:10.1016/j.gene.2010.06.012
a b s t r a c t
a r t i c l e i n f oArticle history:Accepted 29 June 2010Available online 16 July 2010
Received by S. Goldenberg
Keywords:Piwi-like proteinSmall RNAsProtozoan parasitesArgonaute phylogeny
Over the last years an expanding family of small non-coding RNAs (sRNA) has been identified in eukaryoticgenomes which behave as sequence-specific triggers for mRNA degradation, translation repression,heterochromatin formation and genome stability. To achieve their effectors functions, sRNAs associate withmembers of the Argonaute protein family. Argonaute proteins are segregated into three paralogous groups:the AGO-like subfamily, the PIWI-like subfamily, and theWAGO subfamily (for Worm specific AGO). Detailedphylogenetic analysis of the small RNA-related machinery components revealed that they can be traced backto the common ancestor of eukaryotes. However, this machinery seems to be lost or excessively simplified insome unicellular organisms such as Saccharomyces cerevisiae, Trypanosoma cruzi, Leishmania major andPlasmodium falciparum which are unable to utilize dsRNA to trigger degradation of target RNAs. We reportedhere a unique ORF encoding for an AGO/PIWI protein in T. cruzi which was expressed in all stages of its lifecycle at the transcript as well as the protein level. Database search for remote homologues, revealed thepresence of a divergent PAZ domain adjacent to the well supported PIWI domain. Our results stronglysuggested that this unique AGO/PIWI protein from T. cruzi is a canonical Argonaute in terms of its domainarchitecture. We propose to reclassify all Argonaute members from trypanosomatids as a distinctivephylogenetic group representing a new subfamily of Argonaute proteins and propose the generic designationof AGO/PIWI-tryp to identify them. Inside the Trypanosomatid-specific node, AGO/PIWI-tryps were clearlysegregated into two paralog groups designated as AGO-tryp and PIWI-tryp according to the presence orabsence of a functional link with RNAi-related phenomena, respectively.
ering RNA; miRNA, microRNA;rgonaute proteins; RNAi, RNAI, P-element induced wimpyframe; OB-folds, oligonucleo-bodies; TbAGO, Trypanosoma
ein; TcPIWI, Trypanosoma cruzi, Liver infusion tryptose.stitut Pasteur de Montevideo,8 522 09 10; fax: +598 522 41
ia Silva),trasbg.fr (M. Frugier),.edu.uy (B. Bonilla),ar (E. Serra),u.uy (C. Robello),
ll rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Over last decade, an expanding family of small noncoding RNAs(including miRNAs, siRNAs and piRNAs) has been identified ineukaryotic genomes which behave as sequence-specific triggers formRNA degradation, translation repression, heterochromatin forma-tion and genome stability (Ghildiyal and Zamore, 2009). Each smallRNA classes recruit RNA processing enzymes and proteins that areresponsible for either their maturation or their subsequent assemblyinto effectors complexes. These ribonucleoprotein complexes (RNP)contain at their center an AGO/PIWI protein familymember as a primecomponent (Hock and Meister, 2008; Hutvagner and Simard, 2008).
The Argonaute protein family consists of highly conserved basicproteins of approximately 100 kDa harboring characteristic structuraldomains or amino acid motifs including a central PAZ domain which

27M.R. Garcia Silva et al. / Gene 466 (2010) 26–35
binds small RNAs through 3', a C-terminal PIWI domain with a RNase-H-like fold (Song et al., 2004; Tolia and Joshua-Tor, 2007) and a MIDdomain located between the PAZ and the PIWI domain (Parker et al.,2005; Yuan et al., 2005) which anchors the 5' phosphate end of smallRNAs onto Argonaute proteins.
Argonaute proteins were previously segregated into three paraloggroups including the AGO-like subfamily (similar to Arabidopsisthaliana AGO1), the PIWI-like subfamily (closely related to Drosophilamelanogaster PIWI protein) and the WAGO subfamily (Worm specificArgonautes) reported in Caenorhabditis elegans (Carmell et al., 2002;Yigit et al., 2006; Hutvagner and Simard, 2008). Expression studies inhigher eukaryotes revealed that the AGO subfamily of proteins areubiquitously expressed, whereas expression of the PIWI subfamilymembers seems to be restricted to germline stem cells (Meister et al.,2004). Detailed phylogenetic analysis of the RNAi-related machinerycomponents AGO/PIWI and Dicer revealed that they can be tracedback to the common ancestor of eukaryotes which encoded at leastthe AGO and the PIWI paralogues (Cerutti and Casas-Mollano, 2006).However, this machinery seems to be lost or excessively simplified insome unicellular organisms such as Saccharomyces cerevisiae, Trypa-nosoma cruzi, Leishmania major and Plasmodium falciparum which areunable to utilize dsRNA to trigger degradation of target RNA(Robinson and Beverley, 2003; DaRocha et al., 2004; Ullu et al.,2004) suggesting that this machinery could have been lost indepen-dently several times during eukaryotic evolution.
Surprisingly, the genome of some RNAi negative organisms such asT. cruzi and L. major bear one putative ORF potentially coding for atleast onemember of the AGO/PIWI family although database searchesfor protein domains revealed the lack of a conserved PAZ domain(Ullu et al., 2004; Cerutti and Casas-Mollano, 2006; Hutvagner andSimard, 2008). The presence of an isolated PIWI domain conductedinitially to denominate these putative proteins as TcPIWI and LmPIWIrespectively or generically as PiwiS (for Piwi “Solo”) to distinguishthem from the canonical PAZ-PIWI containing proteins in spite oftheir very low homology with the reported PIWI-like subfamilymembers. Since there is still no evidence that they could carry outfunctions related with any kind of small RNA-mediated pathways, thebiological significance of these AGO/PIWI-like genes lacking arecognizable PAZ domain remains to be elucidated. Although a singlecopy of the PiwiS is present in genomes of all trypanosomatids, anadditional paralog gene is present in some members includingTrypanosoma brucei, Trypanosoma congolense, Trypanosoma vivaxand Leishmania braziliensis (Durand-Dubief and Bastin, 2003; Ullu etal., 2004) which were generically dubbed “AGO-like.” Interestingly, T.congolense and T. brucei were demonstrated to have functional RNAipathways although only the AGO-like member from T. brucei wasdemonstrated to participate directly in RNAi-related phenomena.(Ngo et al., 1998; Inoue et al., 2002).
Lineage-specific loss of either of the AGO/PIWI paralogs mighthave occurred during evolution as seen in plants which evolved toencode only the AGO paralogs and in animals which evolved toencode both paralogs in their genomes (Hock and Meister, 2008).
In this work we cloned and sequenced a unique ORF from T. cruziencoding for an Argonaute protein which was expressed in all stagesof its life cycle at the transcript as well as the protein level. State of theart methods for remote homology detection using profile vs. profilealignments (Soding, 2005) suggested the presence of a divergent PAZdomain adjacent to a well supported PIWI domain indicating that thisprotein could be a canonical AGO/PIWI in terms of its domainarchitecture. Based on phylogenetic tree constructions, we propose toclassify all Argonaute members from trypanosomatids as a distinctivephylogenetic group representing a new subfamily of Argonauteproteins and propose the generic designation of AGO/PIWI-tryp todistinguish them from the AGO/PIWI members from higher eukar-yotes. The two paralogs groups identifiedwithin the AGO/PIWI familywere designated either “AGO-tryp” subfamily (identifying members
with demonstrated associationwith RNAi phenomena) or “PIWI-tryp”subfamily for members present in all trypanosomatids but withoutreported relationship with RNAi-related phenomena and with adivergent PAZ domain. Thus, the unique recognizable protein from T.cruzi was designated as TcPIWI-tryp.
2. Materials and methods
2.1. T. cruzi cultures and total RNA isolation
T. cruzi epimastigotes from the Dm28c clonewere used throughoutthis work (Contreras et al., 1988). Epimastigotes were grown in liverinfusion tryptose (LIT) medium supplemented with 10% of heat-inactivated fetal calf serum at 28 ºC. Parasite cultures were started at1×106/ml and harvested in late logarithmic growth phase at a celldensity of ~50×106/ml. Metacyclic forms were obtained in axeniccultures by incubating epimastigotes for 3 h in TAU medium(Triatome Artificial Urine medium) at room temperature and furtherincubated in TAU3AAG medium (TAU medium supplemented with10 mM glucose; aspartic acid 2 mM, glutamic acid 50 mM and proline10 mM) as reported (Contreras et al., 1988). Cultures were prolongedfor 2–4 days until more than 80% of parasites were transformed. Theamastigote formswere obtained by culturingmetacyclic forms at 37 ºCfor 3–4 days under chemically defined conditions (MEM-TAU3AAGmedium) as previously reported (Contreras et al., 2002). Total RNAwas extracted from cell pellets with the Trizol reagent (Invitrogen)and processed according to the manufacturer's instructions.
2.2. Prediction of the translation initiation start (TIS) for the putativeTcPIWI-tryp gene
A classifier was trained to discriminate between coding(c) andnon-coding or intergenic sequences as previously described (Ornellaet al., 2005). We used the AdaBoost Decision Stumps classifier(Schapire and Singer, 1999) and implementations provided by theWEKA Java Machine Learning library (Witten and Frank, 2000). Theclassifier was trained with a curated dataset from T. cruzi obtainedfrom GenBank database (Release 146) using a Perl script designed adhoc. The training datasets containing raw coding and intergenicsequences were converted to a binary dataset containing 64 attributes(the frequency of trimers) plus a binary class label indicating thecoding/intergenic identity of original sequences. The 5' extreme of theTcPIWI-tryp sequencewas scanned by slidingwindows of fixed size of100 nt. Each fragment was classified, the score returned by theclassifier was collected, and finally the coordinate pairs x=nucleotideposition+(Windowsize/2) and y=score value (a measure ofprobability of being coded), were represented in a graphic (http://www.r-project.org).
2.3. Homology modeling and phylogenetic tree constructions
We used the HHpred server (http://toolkit.tuebingen.mgp.de/hhpred), which allows inferring more remotely homology relation-ships (Soding, 2005). A structural model of the PIWI domain wasconstructed with the HHpred server using Modeller 8.0 (Eswar et al.,2000). The template structure used (best hit from HHpred) was thatof the Pyrococcus furiosus Argonaute protein (PfAgo, UniProt code:Q8U3D2, PDB id 1U04) (Song et al., 2004). The model of the PIWIdomain included residues 825 to 1070 and 545 to 770 for TcPIWI-trypand PfAgo proteins respectively.
Multiple sequences alignments were performed with PRANKSTER(http://www.ebi.ac.uk/goldman-srv/prank/), which is a novel algo-rithm that treats insertions correctly and avoids over-estimation of thenumber of deletion events and correctly takes into account theevolutionary distances between sequences (Loytynoja and Goldman,2005; Loytynoja and Goldman, 2008) using aWAG substitution matrix.

28 M.R. Garcia Silva et al. / Gene 466 (2010) 26–35
For phylogenetic tree constructions, full length eukaryotic andprokaryotic AGO/PIWI proteins were aligned with PRANKSTER asdescribed above. Bayesian inference of phylogeny was performedwith the MrBayes3.1 software (http://mrbayes.csit.fsu.edu/index.php) using a WAG substitution matrix.
2.4. RT-PCR and sequencing
For cDNA synthesis, 2 μg of total RNA was reverse transcribed withSuperscript II reverse transcriptase system (Invitrogen, Karlsruhe,Germany) using oligo(dT) primers according to the manufacturer'sinstructions. The entire predicted ORF of the TcPIWI-tryp gene wasamplified by using several primer pairs spanning the availableannotated sequence for the T. cruzi CL-Brener clone (SupplementaryFig. 1). After cloning into pGEM T-easy cloning vectors (PromegaCorp., Buenos Aires, Argentina) amplicons were submitted for customsequencing.
2.5. Polyclonal antibody production
Immunizations were performed with a His-tagged recombinantpeptide fragment of 13.5 kDa specific for TcPIWI-tryp (from residues287 to 392) which was obtained by using the pQE30 vector (QIAGEN)in competent cells derived from the M15 strain of Escherichia coli.Immunizations were performed in New Zealand rabbits with the Ni2+
affinity chromatography-purified peptide. Rabbits were bled 15 daysafter the fourth booster according to serum anti-TcPIWI-tryp titrationas assessed by ELISA against the specific recombinant peptide. Rabbitpolyclonal antiserum was enriched in immunoglobulin fraction byammonium sulfate fractionation.
2.6. Western blot and immunofluorescence assays
For immunofluorescence, harvested parasites were washed andfixed in 4% of buffered paraformaldehyde for 10 min at roomtemperature, washed twice with PBS and further incubated in25 mM NH4Cl for 10 min. Parasites were allowed to adhere topolylysine-coated microscope slides and permeabilized with 0.2%Triton X-100 in PBS for 5 min. Slides were then incubated with therabbit anti-TcPIWI-tryp polyclonal antibody in 0.2% BSA Tween 0.1%PBS (1/500) for 2 h at room temperature. After extensive washing theslides were further incubated with a TRITC-conjugated swine anti-rabbit secondary antibody (DAKO) for 1 h at room temperature. Afterwashing, slides were incubated with 4,6-diamidino-2-phenylindole(DAPI) at 1 mg.ml−1 for 5 min at room temperature. Slides weremounted with 5 μl of Fluor Save Reagent (DAKO). Analyses wereperformed in an Olympus IX 81 microscope coupled to a HamamatsuOrca-ER camera (Diagnostic Instruments).
For western blots total proteins were extracted from cell pelletswith lysis buffer (10 mMNaH2 PO4, 1% SDS, 1% β-mercapthoetanol andurea 5M). 30 μg of total protein extracts were separated in 8% SDS-PAGE and after electrophoretic transfer, nitrocellulose membraneswere blocked in PBS plus 5% BSA and Tween 0.1%. Membranes wereincubated with anti-TcPIWI-tryp antibodies for 2 h at 1:2000 in PBS-Tween 0.5% and BSA 1 mg/ml at room temperature. After extensivewashing membranes were incubated with a horseradish peroxidase-conjugated secondary antibody and developed using ECL reagents(Amersham Biosciences).
2.7. Northern blots
For northern blot, 20 micrograms of total RNA was separated bydenaturing 1% formaldehyde-agarose electrophoresis and transferredto nylonmembranes (Amersham Inc., USA). A DNA probe labeledwithα-32P-dCTP was prepared with a 732 bp amplicon generated by PCRspanning the Piwi domain of the Dm28c (2473 to 3204 bp). The
hybridization was performed in hybridization buffer (1 mM EDTA, 0.5M, Na2HPO4, pH 7.2, 5% SDS, 0,5%BSA) overnight at 42 °C. Themembranes were washed once with SSC 2× for 30 min, and once withSSC 1× for 15 min. and autoradiography was performed using KodakX-AR films.
3. Results
3.1. Translation initiation site (TIS) prediction and analysis of theTcPIWI-tryp complete transcript from the Trypanosome cruzi strainDm28c
In silico analysis of the complete genome reported for the referenceT. cruzi clone CL-Brener revealed a putative AGO/PIWI-like gene witha predicted open reading frame of 3315 bp encoding a putativeprotein of 1104 amino acids with a MW of 122.5 kDa and a pI of 9.72(http://tritrypdb.org; accession number Tc00.1047053511367.240and NCBI Reference Sequence: XP_819430). With these data wedesigned a cloning protocol to clone and sequence the correspondingtranscript from the T. cruzi strain Dm28c. The complete nucleotidesequence for the Dm28c AGO/PIWI-like mRNA (herein TcPIWI-tryp)was 3.616 bp in length including an ORF of 3213 bp, a 5'UTR region of32 bp and a 3'UTR of 371 bp (Supplementary Fig. 2A, GeneBankaccession number: HM126462). The 5'UTR was identified by using acombination of several reverse primers associated to a forward primercorresponding to the spliced leader which confirmed the transcript asthe actual mRNA for the respective TcPIWI-tryp gene. The start codonfor the Dm28c strain was identified by bioinformatic prediction andlocalized 78 bp downstream of that predicted for the putative CL-Brener transcript (Supplementary Fig. 2C). As expected, in silicotranslation of the sequenced TcPIWI-tryp transcript yields a 1070amino acids long protein with a predicted molecular weight of~118.2 kDa and a theoretical pI of 9.36 (Supplementary Fig. 2B).
The search of TcPIWI-tryp protein for conserved domains (NCBIConserved Domain Database and Pfam) clearly revealed a C-terminalPIWI domain (Fig. 1) from amino acids 696 to 1049 with an E-value of2e10−13. As previously reported for others trypanosomatids norecognizable PAZ domains were detected (Ullu et al., 2004).Additionally, our search clearly revealed an OB-fold domain forTcPIWI-tryp from amino acids (positions 355 to 413) with an E-valueof 8×10−4. OB-folds are known subdomains of the PAZ domainwhichare compact structural motifs frequently used for ssDNA or ssRNArecognition and binding (Song et al., 2003; Theobald et al., 2003).
The TcPIWI-tryp transcript also codes for a glutamine rich region(polyQ) also found at the N-terminal end of AGO1 and AGO2 proteinsdescribed in A. thaliana and D. melanogaster respectively (Supple-mentary Fig. 2B). Nevertheless, when compared to the CL-Brenerstrain, in the Dm28c strain this polyQ box showed a 24 bp deletionwhich shortened it from 12 glutamines intercalated with twohistidines to only 6 contiguous glutamines. Two allelic variants atpositions 956 and 1059 and a total of 54 nucleotide substitutionsresulting in a total of 25 changes in amino acids and 29 synonymoussubstitutions were identified compared to the reference CL-Brenerstrain.
Multiple sequence alignments with the founding members ofAGO/PIWI proteins (AtAGO1 and DmPIWI proteins from A. thalianaand D. melanogaster respectively) revealed that TcPIWI-tryp share avery low similarity with these proteins (9% and 10% respectively).
In agreement with the sequenced transcript, northern blot assaysconfirmed the presence of a unique transcript for TcPIWI-tryp ofabout 3.6 kb (Fig. 2A) which included the spliced leader and therespective 5' and 3' UTR regions.
Western blot assays for TcPIWI-tryp showed a ~120 kDa immu-noreactive band which was closed to the predicted molecular massbased on the sequenced transcript and ORF estimation (Fig. 2B). Asecond immunoreactive band with lower relative molecular weight
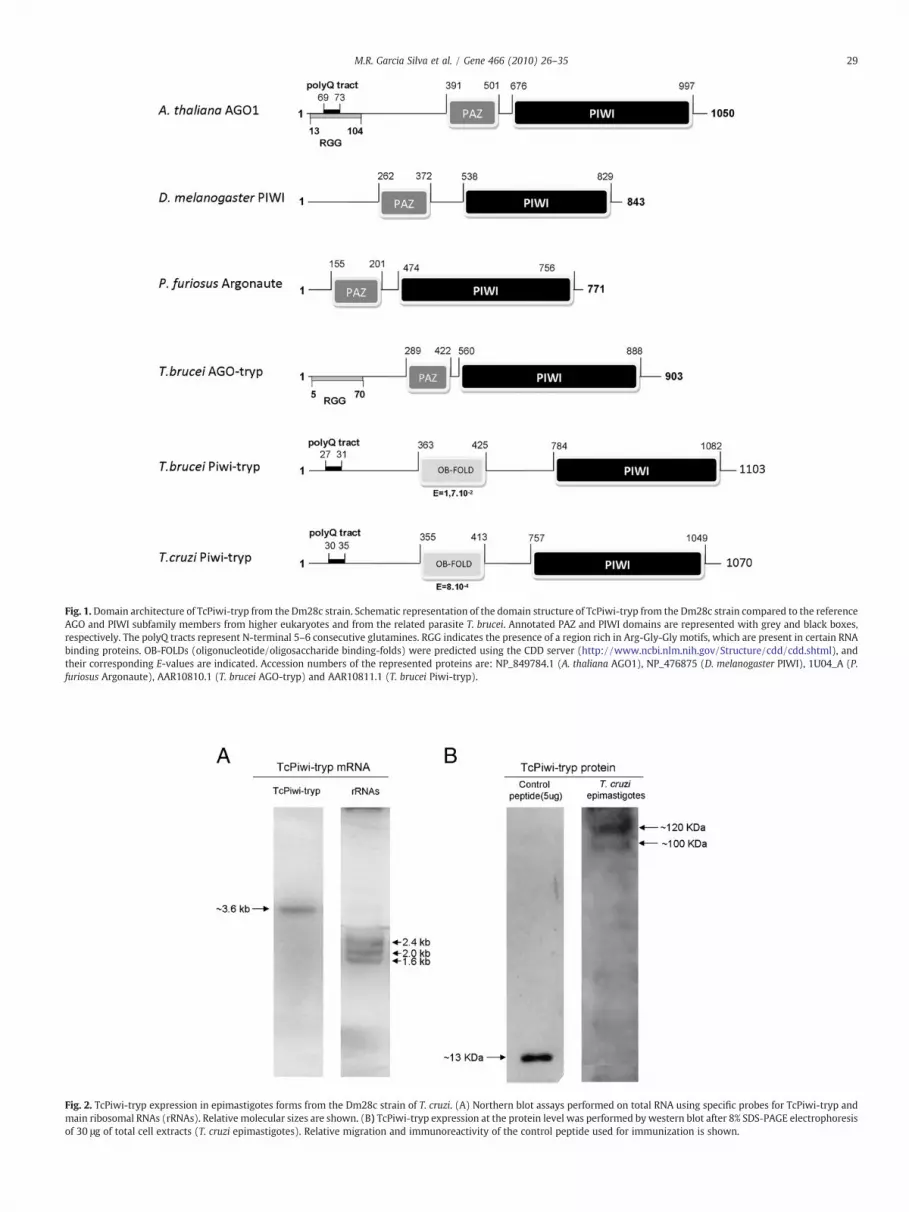
Fig. 1.Domain architecture of TcPiwi-tryp from the Dm28c strain. Schematic representation of the domain structure of TcPiwi-tryp from the Dm28c strain compared to the referenceAGO and PIWI subfamily members from higher eukaryotes and from the related parasite T. brucei. Annotated PAZ and PIWI domains are represented with grey and black boxes,respectively. The polyQ tracts represent N-terminal 5–6 consecutive glutamines. RGG indicates the presence of a region rich in Arg-Gly-Gly motifs, which are present in certain RNAbinding proteins. OB-FOLDs (oligonucleotide/oligosaccharide binding-folds) were predicted using the CDD server (http://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml), andtheir corresponding E-values are indicated. Accession numbers of the represented proteins are: NP_849784.1 (A. thaliana AGO1), NP_476875 (D. melanogaster PIWI), 1U04_A (P.furiosus Argonaute), AAR10810.1 (T. brucei AGO-tryp) and AAR10811.1 (T. brucei Piwi-tryp).
Fig. 2. TcPiwi-tryp expression in epimastigotes forms from the Dm28c strain of T. cruzi. (A) Northern blot assays performed on total RNA using specific probes for TcPiwi-tryp andmain ribosomal RNAs (rRNAs). Relative molecular sizes are shown. (B) TcPiwi-tryp expression at the protein level was performed by western blot after 8% SDS-PAGE electrophoresisof 30 μg of total cell extracts (T. cruzi epimastigotes). Relative migration and immunoreactivity of the control peptide used for immunization is shown.
29M.R. Garcia Silva et al. / Gene 466 (2010) 26–35

30 M.R. Garcia Silva et al. / Gene 466 (2010) 26–35
was observed which could correspond to a non-specific reaction or toa partial proteolysis of the mature peptide. These results confirm thepresence of a protein in T. cruzi that positively reacts with an antibodydesigned against the TcPIWI-tryp protein.
3.2. Remote homology prediction reveals a TcPIWI-tryp with a completeArgonaute architecture
Owing to the characteristic very low sequence conservation of PAZdomains we sought to use more sensitive methods for databasesearching of remote homologues using the HHpred prediction tool.This search revealed several PAZ-containing homologues with knownstructures within the region from amino acids 500 to 1071 on TcPIWI-tryp (Fig. 3A). The best scoring hits corresponded to Argonauteproteins from several archaebacteria and eubacteria for which thestructure was reported. Excluding the Archeoglobus fulgidus Argo-naute, which lacks of a canonical PAZ domain, these proteins feature acomplete Argonaute architecture comprising PAZ, MID and PIWIdomains, suggesting that an analogous functional domain organiza-
Fig. 3. Structural insights into TcPIWI-tryp protein. (A) Selected hits of remote homology desequence of TcPiwi-tryp. Structural domains are colored in orange for OB folds, in red for PAZin the corresponding segments followed by HHpred scores enclosed in parenthesis. (B)representative eukaryotic AGO/PIWI and AGO/PIWI-tryp proteins. Gapped residues betweewere colored in red. Residues which were present in most members of the AGO-like subfadefining residues were colored in blue. The AGO/PIWI-tryp-defining residues were coloredindicated by asterisks. (C) Structural model of the PAZ/MID/PIWI domains from TcPIWI-trypThe inset shows the spatial close vicinity of the putative catalytic triad conformed by residuDynamics Software (VMD, http://www.ks.uiuc.edu/Research/vmd). The different domains
tion is present in TcPIWI-tryp protein. Additionally, high structuralhomology in the region from amino acids 500 to 610 was found withPAZ domains from human and fly Argonautes (PDB: 1r4k, 1si2 and1vyn).
3.3. PIWI-tryp proteins, but not AGO-tryp proteins, seem to have a DDHcatalytic triad
It was previously reported that the “slicer” catalytic activity ofmany Argonaute proteins depend on two Asp and one His residuespresent in a specific region of the PIWI domain. (Rivas et al., 2005).Visual inspection of the multiple sequence alignments (Fig. 3B) alsodenotes the existence of a conserved DDH triad in the PIWI-trypsubfamily of proteins. The structural model of the PIWI domain ofTcPIWI-tryp constructed using PfAGO as a template indicates that thethree aligned DDH residues are located in the “slicer site” of the PIWIdomain, and spatially close to each other (Fig. 3C). The distancesbetween the carboxylates from D841 and D908 and the imidazol ringof H1039 are 0.4 nm and 0.5 nm, respectively. Other residues that
tection as obtained from the HHpred server. Structural domains are aligned against the, in green for Mid and in pink for Piwi domains. The PDB code of each protein is indicatedMultiple sequence alignment (Prankster graphical aligner) of PIWI domains from
n alignment blocks are enclosed in parenthesis. Eukaryotic AGO/PIWI-defining residuesmily (AGO-defining) but not in the PIWI-like subfamily were colored in green. PIWI-in black. Residues corresponding to the DDH catalytic triad were colored in yellow andusingModeller and the Argonaute protein form P. furiosus as template (PDB code:1u04).es Asp841, 908 and His1039. 3D-images were generated by using the Visual Molecularare identified following the same color codes used in (A).

31M.R. Garcia Silva et al. / Gene 466 (2010) 26–35
could putatively form this catalytic triad, namely, D898, D901 andD910, are located more than 1 nm far away from the “active” site. Thisprovides further support for the idea that the catalytic triad–if itactually exists–is formed by D841, D908 and H1039. Although theslicer catalytic motif was demonstrated moderately degenerated inthe third residue (D-D-E/D/H/K) (Tolia and Joshua-Tor, 2007) it isnoteworthy that a catalytic triad could not be identified in the AGO-tryp subfamily of proteins which include both the LbAGO-tryp from L.braziliensis and the RNAi active TbAGO-tryp, from T. brucei (Fig. 3B).
3.4. AGO/PIWI proteins from trypanosomatids share a commonevolutionary origin and do not belong to the previously defined AGO-and PIWI-like subfamilies
To study the evolutionary relatedness of Trypanosomatid AGO/PIWI with previously established groups regardless the presence of anannotated PAZ domain, we performed a phylogenetic analysis usingthe whole sequence from representative eukaryotic and prokaryoticArgonaute proteins. As previously reported (Cerutti and Casas-Mollano, 2006; Murphy et al., 2008), phylogenetic analysis showedthat most eukaryotic Argonautes grouped either into the AGO-likesubfamily or into the PIWI-like subfamily of Argonaute proteins(Fig. 4). The C. elegans group 3 Argonautes (WAGOs) included in thisanalysis clustered together with the AGO subfamily but formed amonophyletic group as previously reported (Yigit et al., 2006).Surprisingly, AGO/PIWI proteins from Trypanosomatids do notgrouped with any of previously delimited AGO/PIWI subfamilies.Instead, they form a well supported monophyletic group whichremains as an out-group to any other eukaryotic sequence (Fig. 4). Inaddition, this trypanosomatid-specific group is subdivided in twosubgroups, one of which comprises the PAZ-PIWI domain-containingand RNAi functionally active TbAGO-tryp and its predicted orthologs,and the other which comprises the predicted orthologs of TbPIWI-tryp (including TcPIWI-tryp), which seem to bare a very divergentform of the PAZ domain.
In agreement with these data, the analysis of residues defining theArgonaute members and their respective subfamilies based on thecriterion proposed by Parker et al. (2004), showed that residuesdefining the AGO-like and the PIWI-like subfamilies (colored in greenand blue, respectively, in Fig. 3B) were shared only partially, makingdifficult to assign AGO/PIWI-tryp proteins to either of the canonicalAGO-like and PIWI-like subfamilies.
3.5. TcPIWI-tryp protein is expressed at all stages in the life cycle of T.cruzi and localizes in the cytoplasm
Since trypanosomes display different life cycle stages, each with aspecific pattern of protein expression, we used immunofluorescenceassays to verify TcPIWI-tryp expression and subcellular localization(Fig. 5). TcPIWI-tryp was found expressed and recruited to thecytoplasm in all differentiating forms of the parasite. The cytoplasmicdistribution of TcPIWI-tryp observed in epimastigotes resembled thedistribution previously reported by TbAGO-tryp in T. brucei (Durand-Dubief and Bastin, 2003).
4. Discussion
In this work we have cloned and sequenced the complete ORFencoding a putative AGO/PIWI protein from Trypanosoma cruzidesignated here as TcPIWI-tryp. Similarly to most eukaryotic AGO/PIWI proteins the translated protein was highly basic (pI of 9.36) witha predictedmolecular weight of ~118 kDa (Cerutti et al., 2000; Song etal., 2004; Tolia and Joshua-Tor, 2007). The AGO/PIWI proteins arecharacterized by the presence of three main structural featuresdefined by the presence of the PAZ, MID and PIWI domains (Cerutti etal., 2000; Hutvagner and Simard, 2008). However, the sequence
analysis of the TcPIWI-tryp revealed only a well conserved PIWIdomain and an OB-fold in the N-terminal region. We speculated onthe possibility that TcPIWI-tryp could have really lost its PAZ domainor retained a divergent PAZ which could not be detected byconventional prediction methods based on sequence to sequencealignments. Surprisingly, the search for remote structurally relatedhomologues performed with HHpred, strongly suggested the pres-ence of a deviant PAZ domain on TcPIWI-tryp. Thus, TcPIWI-trypcomprises a full Argonaute architecture suggesting that the assump-tion that T. cruzi has lost all its RNAi-related silencing machineryincluding their Argonaute genes should be reconsidered.
For both prokaryotic and lower eukaryote Argonautes, primarysequence comparisons also failed to reveal conserved PAZ domains.For instance, although the PIWI domain could be easily detected in P.furiosus Argonaute by BLAST searches, it has been recently reportedthat a PAZ domain could not be detected until its structure was solvedrevealing a close structural similarity with the PAZ domain fromhuman Argonaute AGO1 (Song et al., 2004). It is highly likely that inthese organisms Argonaute proteins have poorly conserved PAZdomains which could not be detected by using primary sequencescomparisons (Ullu et al., 2004; Makarova et al., 2009). Thus theexpression of a PAZ-MID-PIWI domain containing protein in T. cruzi,not related to RNAi functional pathways, could be relevant since it wasthought that this parasitic protozoon had lost all its Argonaute genestogether with the entire RNAi machinery (Cerutti and Casas-Mollano,2006; Shabalina and Koonin, 2008).
While T. cruzi and L. major may have lost their AGO-tryp genestogether with the entire RNAi machinery, all Trypanosomatids ana-lyzed retained a member of the PIWI-tryp subfamily. Thus, PIWI-trypproteins may possibly play essential regulatory roles in these protists.
Interestingly, our analysis also revealed that TcPIWI-tryp harbor anOB-fold motif in the region from amino acids 355 to 413 upstreamfrom the putative PAZ motif. OB-folds (for oligonucleotide/oligosac-charide binding folds) are compact structural motifs frequently usedfor nucleic acid recognition (Theobald et al., 2003) where most familymembers share low degree of sequence similarities. OB-folds havebeen also reported as integral subdomains of PAZ motifs (Song et al.,2003). These features reinforce the idea that TcPIWI-tryp could harbora deviant PAZ domain and additionally retain an additional OB-foldable to bind ssRNA or ssDNA (Song et al., 2003; Theobald et al., 2003;Yan et al., 2003). However, these assumptions will require validationthrough specific biochemical and structural experimental approaches.Another structural aspectwas the presence of a short polyQ tract in theN-terminal region of the TcPIWI-tryp which is also present in otherPIWI-tryp proteins as well as in some higher eukaryotic ArgonautesincludingAGO2 fromD.melanogaster andAGO1 fromA. thaliana. Theseglutamine-rich domains have been implicated in protein–proteininteractions which were linked to the protein aggregation observed incertain neurodegenerative diseases (Ross and Poirier, 2004). SinceAGO2 is an essential component of protein complexes, such as theRISC, it can be speculated that control of its aggregation state isconceivably important for its function. Mammalian AGO-like proteinsare localized to GW bodies (cytoplasmic compartments analogous toyeast P-bodies)which are centers ofmRNAdegradation (Sen and Blau,2005). Of note GW182 familymembers bear poly-Q rich regionswhichwere demonstrated to be essentials for their interaction withArgonaute proteins (Eulalio et al., 2009a,b).
Multiple sequence alignments including representative membersof the three paralogous groups (AGO-like, PIWI-like and WAGO) ofArgonaute proteins (Carmell et al., 2002; Yigit et al., 2006) and severalAGO/PIWI-tryps proteins revealed that AGO/PIWI proteins fromtrypanosomatids do not share significant similarity with any of thesepreviously identified subfamilies. Additionally, the analysis of themost conserved residues of the PIWI domains of AGO/PIWI proteinsrevealed that the AGO- and PIWI-defining residues present in highereukaryotic Argonautes were shared only partially even if many

32 M.R. Garcia Silva et al. / Gene 466 (2010) 26–35
residues defining the AGO/PIWI family were conserved. Theseassumptions strongly suggested that AGO/PIWI-tryp proteins couldrepresent a distinctive subfamily different from those previouslyreported and could not be classified as belonging to either the PIWI-like, the AGO-like or the WAGO subfamilies.
In support of this assumption, recent phylogenetic tree recon-structions of Argonaute proteins (Cerutti and Casas-Mollano, 2006;Yigit et al., 2006; Shabalina and Koonin, 2008) revealed that TbAGO1(the only Argonaute member from trypanosomatids included in theanalysis) could not be reliably resolved in terms of their grouping asmember of any of these well-established subfamilies.
Thus, a classification of AGO/PIWI-tryp proteins by means ofphylogenetic criteria was not performed in the past and their assignmentas either “AGO” or “PIWI” actually leads to confusion. Interestingly, in ourphylogenetic analysis all AGO/PIWI-trypproteins formed amonophyleticgroup. Furthermore, this group is in fact an out-group to every othereukaryotic Argonaute from the AGO-like and PIWI-like subfamilies. TheBayesian posterior probability for the node connecting AGO and PIWIsubfamilies and excluding all AGO/PIWI-tryp proteins was 1 (Fig. 4), aswell as the posterior probability for the nodewhich is responsible for theAGO/PIWI-tryp monophyly. This observation implies that the lastcommon ancestor between any given AGO-like or PIWI-like protein andany given Trypanosomatid Argonaute is earlier to the gene duplicationevent which originated AGO and PIWI subfamilies. This large phyloge-netic distance explains the low degree of sequence similarity betweenTrypanosomatid Argonautes and other reference eukaryotic Argonautes.Alternatively, it can be speculated that AGO/PIWI-tryp genes could havediverged faster than the rest of eukaryotic Argonautes avoiding theirclustering into either of AGO- or PIWI-like groups.
Inside the Trypanosomatid-specific node, AGO/PIWI-tryp mem-bers were segregated into two presumable paralogous groups. Onesub-group was represented by members having clearly recognizablePAZ and PIWI domains including the AGO-tryp members from T.brucei and T. congolense which represent the only membersdemonstrated to have functionally active RNAi pathways (Ngo et al.,1998; Inoue et al., 2002; Durand-Dubief and Bastin, 2003; Tschudi etal., 2003). The AGO-tryp node includes also L. braziliensis, and T. vivaxfor which there are no published functional evidences of RNAi but byorthology are presumed to be strong candidates for a functional RNAipathway (Militello et al., 2008). Due to their possible involvement inRNAi related phenomena and by forming a distinctive monophyleticgroup we proposed the term “AGO-tryp,” in order to distinguish themfrom the canonical subfamilies of higher eukaryotic identified as AGO-like proteins.
The other subfamily of trypanosomatids was designated as PIWI-tryp (denomination referring to the presence of a PIWI domain but donot implying its classification as a member of the PIWI-like subfamilyof higher eukaryotes) and includes TcPIWI-tryp and TbPIWI-tryp fromT. cruzi and T. brucei, respectively. The genome of all trypanosomatidsretained at least one PIWI-tryp protein suggesting that this sub-groupof proteins may play a key regulatory role related or not to RNA-mediated RNA cleavage.
Since two paralogs of AGO/PIWI-tryp genes are present in thegenomes of Leishmania spp. and Trypanosoma spp., we inferred that agene duplication event should have taken place before the origin ofboth genders. This fact implies an independent loss of one of theparalogs (AGO-tryp) occurring later in the evolution of T. cruzi, L.major and L. infantum. This hypothesis, based on the parsimony
Fig. 4. Phylogenetic analysis of Trypanosomatid Argonaute proteins. Argonaute proteins fromhigher eukaryotes but in a new monophyletic group which is grouped as the “AGO/PIWI-tryAGO/PIWI-tryp: the PAZ containing homologs of TbAGO1, which is depicted as “AGO-tryp”sequence alignments), which are depicted as “PIWI-tryp” proteins. Representative prokaryotare identified by “WAGO” .The phylogenetic tree was made using a WAG substitution matrixfor each node, multiplied by a factor of 100. Sequences correspond to full length eukaryoticthe sequences used are listed as Supplementary File 1.
principle, is analogous to the accepted theory of eukaryotic Argonauteevolution (Cerutti and Casas-Mollano, 2006). Since a PAZ motif can beunambiguously identified in TbAGO-tryp (Durand-Dubief and Bastin,2003), and since AGO-tryp and PIWI-tryp proteinsmay have appearedas a consequence of a gene duplication event, it is reasonable topredict that PIWI-tryp proteins should have inherited a PAZ domain,as was strongly suggested by results described here.
The classification of Trypanosomatid Argonautes as either AGO-tryp or PIWI-tryp and globally as AGO/PIWI-tryp is consistent withphylogenetic data and emphasizes the difference between AGO/PIWI-tryp proteins and the rest of eukaryotic Argonautes.
Our results revealed that the PIWI domains from the TcPIWI-trypand the rest of PIWI-tryp proteins analyzed contained a DDH triadclassically found in catalytically competent Argonautes. In thisrespect, it has been demonstrated that the PIWI domain adopt anRNase H-like fold (Song et al., 2003; Parker et al., 2004; Yuan et al.,2005; Wang et al., 2008). The RNase H enzymes are Mg2+-dependentRNases that typically cleaves DNA-RNA hybrids through theirRNase-H domains performing a two-metal ion catalysis involvingthree conserved residues represented by the triad Asp-Asp-Glu (DDEmotif) which is present in prokaryotic Argonautes (Nowotny et al.,2005). Combined crystallographic and mutational analysis of eukary-otic Argonaute members determined that, although the two criticalAspartates are present, the eukaryotic counterparts use an additionalHistidine rather that the Glutamic acid in the third position (Rivas etal., 2005). It was recently reported that this catalytic triad, is the“slicer” core of the effector complex RISC (Liu et al., 2004; Tolia andJoshua-Tor, 2007). Recent reports, demonstrated that the catalytictriad is moderately degenerated in some Argonaute members andshould be re-defined as Asp-Asp-Asp/Glu/His/Lys (Rivas et al., 2005;Tolia and Joshua-Tor, 2007).
Nevertheless, experimental data demonstrated that the presenceof a catalytic motif is a prerequisite but not sufficient to assignendoribonuclease (Slicer) activity (Tolia and Joshua-Tor, 2007; Hockand Meister, 2008). Conversely, TbAGO-tryp was shown to beessential for RNAi but does not seem to harbor a DDH triad in ourmultiple sequence alignment. In this respect, there is no experimentalevidence attributing “slicer” activity for TbAGO-tryp, althoughunpredicted alternate catalytic triads could not be excluded. Asobserved for some pAGOs do not having canonical catalytic triads,specific nucleases are recruited which can act as active endonucleasesin complex with pAGOs (Makarova et al., 2009). Whatever the case,this information should be relevant for future mutational assaystempting to elucidate the biological significance of AGO/PIWI-trypproteins.
Finally, we demonstrated that TcPIWI-tryp protein is expressedthroughout the life cycle of T. cruzi andwas recruited to the cytoplasmfollowing a granular pattern. In this respect, most of eukaryoticmembers of the Ago subfamily proteins were demonstrated to localizeto the cytoplasm mainly associated to P-bodies (Hock and Meister,2008). In zebrafish, mouse and flies some members of the PIWI-likesubfamily were enriched in the cytoplasm (Brennecke et al., 2007;Houwing et al., 2007; Unhavaithaya et al., 2009).
In order to validate AGO/PIWI-tryp proteins as novel and dis-tinctive members of the Argonaute family of proteins more work isneeded to determine not only structural features but their biologicalfunction as it was demonstrated for TbAGO1which participate in RNAsilencing processes and is essential for chromosome segregation and
trypanosomatids do not group with the canonical Ago-like and Piwi-like subfamilies ofp group.” As for eukaryotic Argonautes, two paralogs subgroups are defined inside theproteins and the homologs of TbPIWI (no PAZ domain detected by algorithms based onic argonautes are depicted as “pAgos” and the worm-specific Argonautes from C. elegansand the software Mrbayes 3.1. Numbers correspond to bayesian posterior probabilitiesand prokaryotic Argonautes and were aligned with PRANKSTER. Accession numbers for

Fig. 5. Indirect immunofluorescence microscopy for TcPIWI-tryp. Representative experiments detecting TcPIWI-tryp (red fluorescence) from epimastigotes (Epi), amastigotes(Ama) and metacyclic (Met) forms of T. cruzi (Dm28c strain). Slides were counterstained with DAPI as a reference to identify nuclei and kinetoplasts. The respective differentialinterference contrast images are depicted (DIC) in order to identify the parasite cellular body and the respective flagellum origin (Scale bars=5 μm). The respective specificitycontrols performed with pre-immune sera are depicted.
34 M.R. Garcia Silva et al. / Gene 466 (2010) 26–35
mitotic spindle assembly in T. brucei (Durand-Dubief and Bastin,2003).
Acknowledgments
This work was supported partially by the “Institut Pasteur deMontevideo” and from the National Agency for Innovation and Research,Montevideo, Uruguay.
M.R.G.C. and JPT are research fellows from the “Agencia Nacionalde Investigación e Innovacion (ANII)” del Ministerio de Educacion yCultura y Presidencia de la Republica, Montevideo Uruguay.
C. Rob. and A.C. are researchers from the Sistema NacionalInvestigadores (ANII), Montevideo Uruguay
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.gene.2010.06.012.
References
Brennecke, J., Aravin, A.A., Stark, A., Dus, M., Kellis, M., Sachidanandam, R., et al., 2007.Discrete small RNA-generating loci as master regulators of transposon activity inDrosophila. Cell 128, 1089–1103.
Carmell, M.A., Xuan, Z., Zhang, M.Q., Hannon, G.J., 2002. The Argonaute family: tentaclesthat reach into RNAi, developmental control, stem cell maintenance, andtumorigenesis. Genes Dev. 16, 2733–2742.
Cerutti, H., Casas-Mollano, J.A., 2006. On the origin and functions of RNA-mediatedsilencing: from protists to man. Curr. Genet.
Cerutti, L., Mian, N., Bateman, A., 2000. Domains in gene silencing and celldifferentiation proteins: the novel PAZ domain and redefinition of the Piwidomain. Trends Biochem. Sci. 25, 481–482.
Contreras, V.T., Araujo-Jorge, T.C., Bonaldo, M.C., Thomaz, N., Barbosa, H.S., MeirellesMde, N., et al., 1988. Biological aspects of the Dm 28c clone of Trypanosoma cruziafter metacyclogenesis in chemically defined media. Mem. Inst. Oswaldo Cruz 83,123–133.
Contreras, V.T., Navarro, M.C., De Lima, A.R., Arteaga, R., Duran, F., Askue, J., et al., 2002.Production of amastigotes from metacyclic trypomastigotes of Trypanosoma cruzi.Mem. Inst. Oswaldo Cruz 97, 1213–1220.
DaRocha, W.D., Otsu, K., Teixeira, S.M., Donelson, J.E., 2004. Tests of cytoplasmic RNAinterference (RNAi) and construction of a tetracycline-inducible T7 promotersystem in Trypanosoma cruzi. Mol. Biochem. Parasitol. 133, 175–186.
Durand-Dubief, M., Bastin, P., 2003. TbAGO1, an argonaute protein required for RNAinterference, is involved in mitosis and chromosome segregation in Trypanosomabrucei. BMC Biol. 1, 2.
Eswar, N., Marti-Renom, M.A., Webb, B., Madhusudhan, M.S., Eramian, D., Shen, M., etal., 2000. Comparative protein structure modeling with MODELLER. Curr. Protoc.Bioinform. 15 5.6.1-5.6.30..
Eulalio, A., Helms, S., Fritzsch, C., Fauser, M., Izaurralde, E., 2009a. A C-terminal silencingdomain in GW182 is essential for miRNA function. RNA 15, 1067–1077.
Eulalio, A., Tritschler, F., Izaurralde, E., 2009b. The GW182 protein family in animal cells:new insights into domains required for miRNA-mediated gene silencing. RNA 15,1433–1442.
Ghildiyal, M., Zamore, P.D., 2009. Small silencing RNAs: an expanding universe. Nat.Rev. 10, 94–108.
Hock, J., Meister, G., 2008. The Argonaute protein family. Genome Biol. 9, 210.Houwing, S., Kamminga, L.M., Berezikov, E., Cronembold, D., Girard, A., van den Elst, H.,
et al., 2007. A role for Piwi and piRNAs in germ cell maintenance and transposonsilencing in Zebrafish. Cell 129, 69–82.
Hutvagner, G., Simard, M.J., 2008. Argonaute proteins: key players in RNA silencing. Nat.Rev. Mol. Cell Biol. 9, 22–32.

35M.R. Garcia Silva et al. / Gene 466 (2010) 26–35
Inoue, N., Otsu, K., Ferraro, D.M., Donelson, J.E., 2002. Tetracycline-regulated RNAinterference in Trypanosoma congolense. Mol. Biochem. Parasitol. 120, 309–313.
Liu, J., Carmell, M.A., Rivas, F.V., Marsden, C.G., Thomson, J.M., Song, J.J., et al., 2004.Argonaute2 is the catalytic engine of mammalian RNAi. Science 305, 1437–1441.
Loytynoja, A., Goldman, N., 2005. An algorithm for progressive multiple alignment ofsequences with insertions. Proc. Natl Acad. Sci. USA 102, 10557–10562.
Loytynoja, A., Goldman, N., 2008. Phylogeny-aware gap placement prevents errors insequence alignment and evolutionary analysis. Science 320, 1632–1635.
Makarova, K.S., Wolf, Y.I., van der Oost, J., Koonin, E.V., 2009. Prokaryotic homologs ofArgonaute proteins are predicted to function as key components of a novel systemof defense against mobile genetic elements. Biol. Direct 4, 29.
Meister, G., Landthaler, M., Patkaniowska, A., Dorsett, Y., Teng, G., Tuschl, T., 2004.Human Argonaute2 mediates RNA cleavage targeted by miRNAs and siRNAs. Mol.Cell 15, 185–197.
Militello, K.T., Refour, P., Comeaux, C.A., Duraisingh, M.T., 2008. Antisense RNA andRNAi in protozoan parasites: working hard or hardly working? Mol. Biochem.Parasitol. 157, 117–126.
Murphy, D., Dancis, B., Brown, J.R., 2008. The evolution of core proteins involved inmicroRNA biogenesis. BMC Evol. Biol. 8, 92.
Ngo, H., Tschudi, C., Gull, K., Ullu, E., 1998. Double-stranded RNA induces mRNAdegradation in Trypanosoma brucei. Proc. Natl Acad. Sci. USA 95, 14687–14692.
Nowotny, M., Gaidamakov, S.A., Crouch, R.J., Yang, W., 2005. Crystal structures of RNaseH bound to an RNA/DNA hybrid: substrate specificity and metal-dependentcatalysis. Cell 121, 1005–1016.
Ornella, L., Esteban, L., Serra, E., Tapia, E., 2005. Classification approach for detectingcoding and non coding regions in T. cruzi genomic sequences: Actas de la AcademiaNacional de Ciencias, Argentina XIV, pp. 125–133.
Parker, J.S., Roe, S.M., Barford, D., 2004. Crystal structure of a PIWI protein suggestsmechanisms for siRNA recognition and slicer activity. EMBO J. 23, 4727–4737.
Parker, J.S., Roe, S.M., Barford, D., 2005. Structural insights into mRNA recognition froma PIWI domain-siRNA guide complex. Nature 434, 663–666.
Rivas, F.V., Tolia, N.H., Song, J.J., Aragon, J.P., Liu, J., Hannon, G.J., et al., 2005. PurifiedArgonaute2 and an siRNA form recombinant human RISC. Nat. Struct. Mol. Biol. 12,340–349.
Robinson, K.A., Beverley, S.M., 2003. Improvements in transfection efficiency and testsof RNA interference (RNAi) approaches in the protozoan parasite Leishmania. Mol.Biochem. Parasitol. 128, 217–228.
Ross, C.A., Poirier, M.A., 2004. Protein aggregation and neurodegenerative disease. Nat.Med. 10 (Suppl), S10–S17.
Schapire, R.E., Singer, Y., 1999. Improved boosting algorithms using confidence-ratedpredictions. Mach. Learn. 37, 297–336.
Sen, G.L., Blau, H.M., 2005. Argonaute 2/RISC resides in sites of mammalian mRNAdecay known as cytoplasmic bodies. Nat. Cell Biol. 7, 633–636.
Shabalina, S.A., Koonin, E.V., 2008. Origins and evolution of eukaryotic RNAinterference. Trends Ecol. Evol. 23, 578–587.
Soding, J., 2005. Protein homology detection by HMM–HMM comparison. Bioinformatics21, 951–960.
Song, J.J., Liu, J., Tolia, N.H., Schneiderman, J., Smith, S.K., Martienssen, R.A., et al., 2003.The crystal structure of the Argonaute2 PAZ domain reveals an RNA binding motifin RNAi effector complexes. Nat. Struct. Biol. 10, 1026–1032.
Song, J.J., Smith, S.K., Hannon, G.J., Joshua-Tor, L., 2004. Crystal structure of Argonauteand its implications for RISC slicer activity. Science 305, 1434–1437.
Theobald, D.L., Mitton-Fry, R.M., Wuttke, D.S., 2003. Nucleic acid recognition by OB-foldproteins. Annu. Rev. Biophys. Biomol. Struct. 32, 115–133.
Tolia, N.H., Joshua-Tor, L., 2007. Slicer and the argonautes. Nat. Chem. Biol. 3, 36–43.Tschudi, C., Djikeng, A., Shi, H., Ullu, E., 2003. In vivo analysis of the RNA interference
mechanism in Trypanosoma brucei. Methods 30, 304–312.Ullu, E., Tschudi, C., Chakraborty, T., 2004. RNA interference in protozoan parasites. Cell.
Microbiol. 6, 509–519.Unhavaithaya, Y., Hao, Y., Beyret, E., Yin,H., Kuramochi-Miyagawa, S., Nakano, T., et al., 2009.
MILI, a PIWI-interacting RNA-binding protein, is required for germ line stem cell self-renewal and appears to positively regulate translation. J. Biol. Chem. 284, 6507–6519.
Wang, Y., Sheng, G., Juranek, S., Tuschl, T., Patel, D.J., 2008. Structure of the guide-strand-containing argonaute silencing complex. Nature 456, 209–213.
Witten, I.H., Frank, E., 2000. Data mining: practical machine learning tools andtechniques with java implementations. Morgan Kaufmann, San Francisco.
Yan, K.S., Yan, S., Farooq, A., Han, A., Zeng, L., Zhou, M.M., 2003. Structure and conservedRNA binding of the PAZ domain. Nature 426, 468–474.
Yigit, E., Batista, P.J., Bei, Y., Pang, K.M., Chen, C.C., Tolia, N.H., et al., 2006. Analysis of theC. elegans Argonaute family reveals that distinct Argonautes act sequentially duringRNAi. Cell 127, 747–757.
Yuan, Y.R., Pei, Y., Ma, J.B., Kuryavyi, V., Zhadina, M., Meister, G., et al., 2005. Crystalstructure of A. aeolicus argonaute, a site-specific DNA-guided endoribonuclease,provides insights into RISC-mediated mRNA cleavage. Mol. Cell 19, 405–419.






























