Embed Size (px)
Citation preview

lsevier.com/locate/yclim
Clinical Immunology 11
Commensal microbiota alter the abundance and TCR responsiveness
of splenic naıve CD4+ T lymphocytes
Tiffany Huang a, Bo Wei b, Peter Velazquez b,1, James Borneman c, Jonathan Braun a,b,*
a Molecular Biology Institute, David Geffen School of Medicine at UCLA, University of California,
Los Angeles, 10833 Le Conte Avenue, CHS 13-222, Los Angeles, CA 90095, USAb Department of Pathology and Laboratory Medicine, David Geffen School of Medicine at UCLA, University of California,
Los Angeles, 10833 Le Conte Avenue, CHS 13-222, Los Angeles, CA 90095, USAc Department of Plant Pathology, University of California, Riverside, CA 92521, USA
Received 12 November 2004; accepted with revision 27 September 2005
Available online 14 November 2005
Abstract
The epidemiologic risk of certain systemic immunologic diseases is affected by commensal or environmental microbiota, but the cellular basis
of the ‘‘hygiene hypothesis’’ is poorly understood. In this study, we demonstrate that composition of the commensal microbiota affects the
functional state of the peripheral naıve (CD62LhiCD44lo) T lymphocyte populations. Restricted flora (RF) mice (stably colonized with excess
nonpathogenic Clostridium sp., and changes in other bacterial and fungal taxa) were distinguished after the neonatal period by a progressive
deficiency in absolute numbers of naıve CD4+ and CD8+ T lymphocytes. SPF and RF mice had comparable levels of memory CD4+ and CD8+ T
cells. This phenotype was attributable to the altered levels of certain commensals and their products, since germ-free mice had normal absolute
numbers of splenic CD4+ and CD8+ T cells and their respective naıve and memory subsets. The naıve CD4+ T cell subset was functionally
distinguished in RF mice versus SPF mice by TCR hyperresponsiveness, pro-inflammatory cytokine production, and increased activation-induced
cell death. Biochemically, these traits were associated with higher basal phosphorylation of the TCR signaling proteins ZAP-70, Lck, and LAT.
These findings indicate that enteric microbial products, through unknown cellular circuitry, influence steps in CD4 T cell differentiation
moderating basal TCR signaling and immune responsiveness.
D 2005 Elsevier Inc. All rights reserved.
Keywords: T cell receptors; Bacteria; Fungi; Commensals; Signaling; T lymphocytes; Mucosal immunology; Hygiene hypothesis
Introduction
Mucosal lymphocytes require the colonization of commensal
bacteria to develop and function appropriately [1]. The reduced
size of gut-associated mucosal tissues and Peyer’s patch, and
reduced cell numbers of lamina propria or intraepithelial T cells
are the characteristics of germ-free rodents which lack
commensal microflora [2–6]. Restoration of the normal
microflora reverses the germ-free mucosal phenotype, further
1521-6616/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.clim.2005.09.012
Abbreviations: AICD, activation-induced cell death; RF, restricted flora;
SPF, specific pathogen free.
* Corresponding author. Department of Pathology and Laboratory Medicine,
CHS 13-222, UCLA School of Medicine, Los Angeles, CA 90095-1732, USA.
Fax: +1 310 825 5674.
E-mail address: [email protected] (J. Braun).1 Present address: Skirball Institute of Biomolecular Medicine, New York
University School of Medicine, 540 First Ave., New York, NY 10016, USA.
suggesting the requirement of commensal bacteria for mucosal
lymphocytes development. Germ-free mice also have decreased
and delayed immune responses after mucosal antigen challenge,
indicating that the presence of commensal bacteria in the
gastrointestinal tract helps to prime mucosal lymphocytes for
quicker immune responses to antigen exposure [7–11]. The
induction and/or maintenance of mucosal tolerance requires the
presence of bacteria in the intestine [12–15]. The loss of
commensal bacteria augments disease in certain models of
autoimmunity, including diabetes onset in NOD mice, collagen-
induced arthritis, and myelin basic protein-induced experimen-
tal autoimmune encephalomyelitis [16–20].
Attenuation of TCR activation is an important homeostatic
feature of mucosal CD4+ Tcells. However, the process directing
this mode of Tcell development and the biochemical basis of the
modified signaling pathway itself are poorly understood. Most
mucosal T lymphocytes have an activated or memory phenotype
7 (2005) 221 – 230
www.e
T. Huang et al. / Clinical Immunology 117 (2005) 221–230222
with CD45RBlo, CD62Llow, CD44hi, and CD69hi surface
markers. This state in part may reflect ongoing innate or
adaptive stimulation of the mucosal immune system by enteric
bacterial products. However, immune ignorance or tolerance
autologous enteric flora by these mucosal lymphocytes has also
been reported [21,22]. Mucosal T cells produce both pro- and
anti-inflammatory cytokines such as IFN-g and IL-10, indicat-
ing that homeostasis is maintained through an active immuno-
regulatory process.
In this study, we tested whether there is an effect of enteric
microbial colonization on the composition of the systemic T
cell population. Germ-free mice were comparable to SPF mice
in their abundance of the major splenic T cell subsets. In
contrast, RF mice (bearing an altered enteric microbiota) had a
selective numerical deficit in naıve CD4+ and CD8+ T cells,
accompanied by functional hyperresponsiveness and Th1
polarization. Collectively, these observations demonstrate a
role for commensal enteric bacteria in shaping the systemic T
cell population.
Materials and methods
Mice
Specific pathogen free (SPF) C57BL/6 mice were obtained
from Jackson Laboratories (Bar Harbor, ME) and monitored
for a panel pathogenic microorganisms based on serologic
and microbiologic screening by the UCLA Division of
Laboratory Animal Medicine. A restricted flora (RF) colony
was established 12 years ago in a separate facility maintained
by the UCLA Department of Radiation Oncology, by
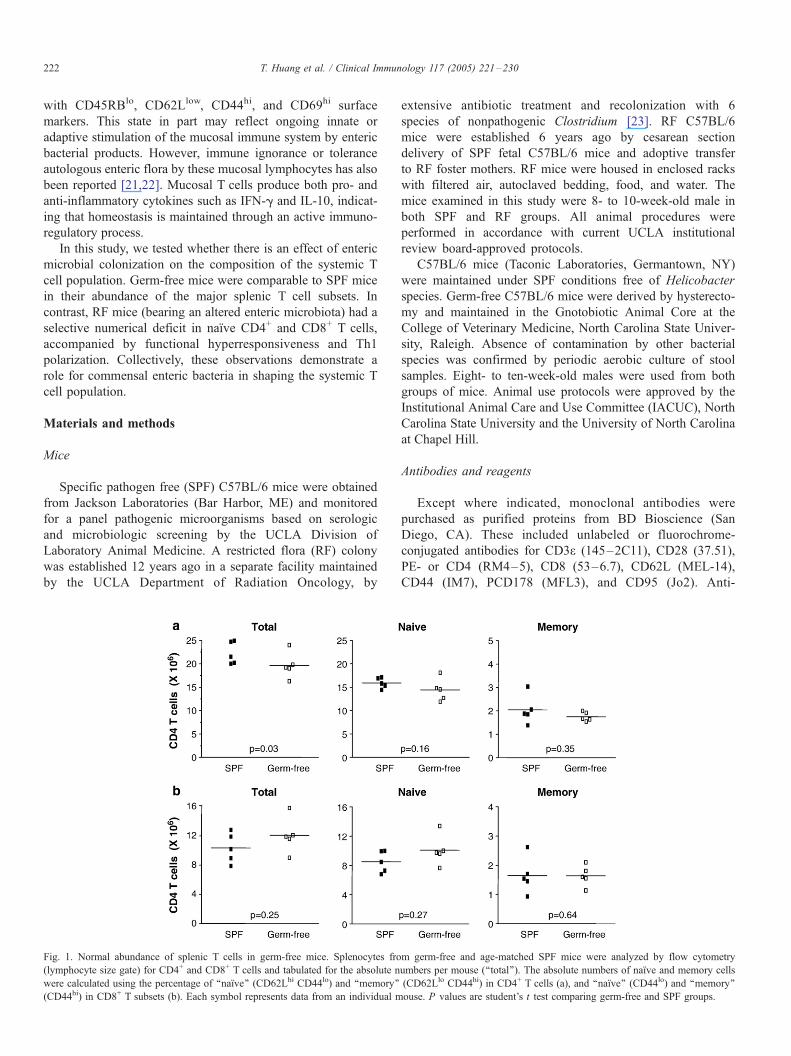
Fig. 1. Normal abundance of splenic T cells in germ-free mice. Splenocytes fro
(lymphocyte size gate) for CD4+ and CD8+ T cells and tabulated for the absolute n
were calculated using the percentage of ‘‘naıve’’ (CD62Lhi CD44lo) and ‘‘memory’’
(CD44hi) in CD8+ T subsets (b). Each symbol represents data from an individual m
extensive antibiotic treatment and recolonization with 6
species of nonpathogenic Clostridium [23]. RF C57BL/6
mice were established 6 years ago by cesarean section
delivery of SPF fetal C57BL/6 mice and adoptive transfer
to RF foster mothers. RF mice were housed in enclosed racks
with filtered air, autoclaved bedding, food, and water. The
mice examined in this study were 8- to 10-week-old male in
both SPF and RF groups. All animal procedures were
performed in accordance with current UCLA institutional
review board-approved protocols.
C57BL/6 mice (Taconic Laboratories, Germantown, NY)
were maintained under SPF conditions free of Helicobacter
species. Germ-free C57BL/6 mice were derived by hysterecto-
my and maintained in the Gnotobiotic Animal Core at the
College of Veterinary Medicine, North Carolina State Univer-
sity, Raleigh. Absence of contamination by other bacterial
species was confirmed by periodic aerobic culture of stool
samples. Eight- to ten-week-old males were used from both
groups of mice. Animal use protocols were approved by the
Institutional Animal Care and Use Committee (IACUC), North
Carolina State University and the University of North Carolina
at Chapel Hill.
Antibodies and reagents
Except where indicated, monoclonal antibodies were
purchased as purified proteins from BD Bioscience (San
Diego, CA). These included unlabeled or fluorochrome-
conjugated antibodies for CD3( (145–2C11), CD28 (37.51),
PE- or CD4 (RM4–5), CD8 (53–6.7), CD62L (MEL-14),
CD44 (IM7), PCD178 (MFL3), and CD95 (Jo2). Anti-
m germ-free and age-matched SPF mice were analyzed by flow cytometry
umbers per mouse (‘‘total’’). The absolute numbers of naıve and memory cells
(CD62Llo CD44hi) in CD4+ T cells (a), and ‘‘naıve’’ (CD44lo) and ‘‘memory’’
ouse. P values are student’s t test comparing germ-free and SPF groups.
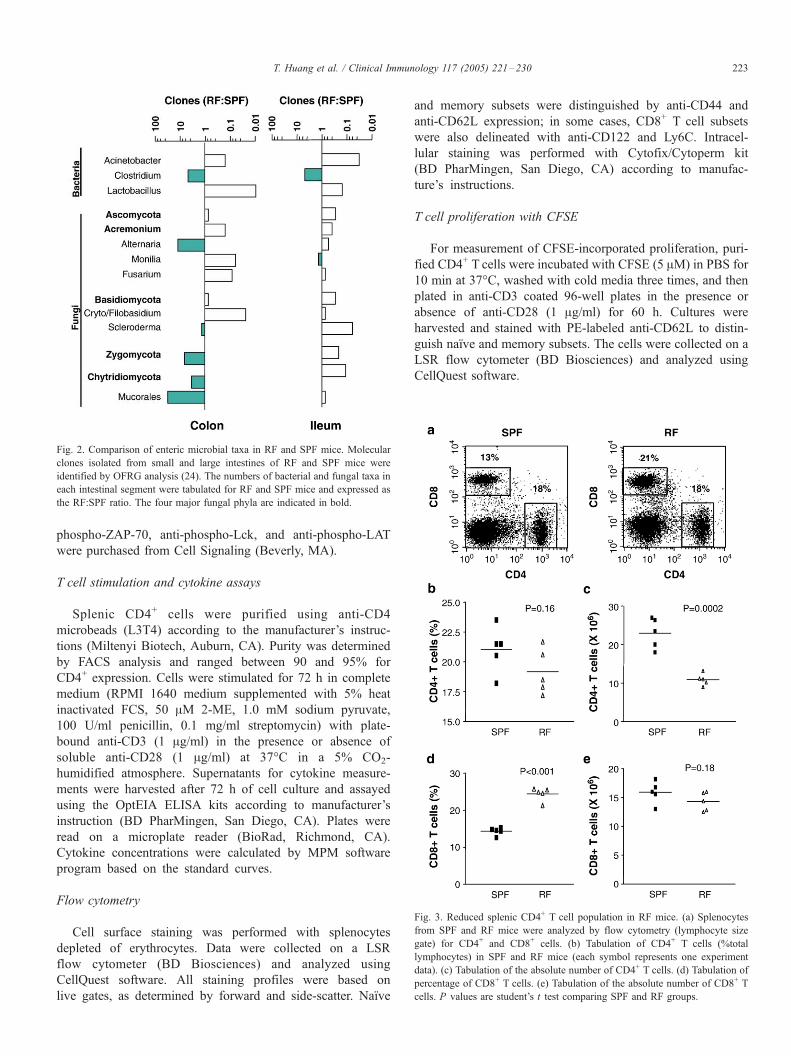
Fig. 3. Reduced splenic CD4+ T cell population in RF mice. (a) Splenocytes
from SPF and RF mice were analyzed by flow cytometry (lymphocyte size
gate) for CD4+ and CD8+ cells. (b) Tabulation of CD4+ T cells (%total
lymphocytes) in SPF and RF mice (each symbol represents one experiment
data). (c) Tabulation of the absolute number of CD4+ T cells. (d) Tabulation of
percentage of CD8+ T cells. (e) Tabulation of the absolute number of CD8+ T
cells. P values are student’s t test comparing SPF and RF groups.
Fig. 2. Comparison of enteric microbial taxa in RF and SPF mice. Molecular
clones isolated from small and large intestines of RF and SPF mice were
identified by OFRG analysis (24). The numbers of bacterial and fungal taxa in
each intestinal segment were tabulated for RF and SPF mice and expressed as
the RF:SPF ratio. The four major fungal phyla are indicated in bold.
T. Huang et al. / Clinical Immunology 117 (2005) 221–230 223
phospho-ZAP-70, anti-phospho-Lck, and anti-phospho-LAT
were purchased from Cell Signaling (Beverly, MA).
T cell stimulation and cytokine assays
Splenic CD4+ cells were purified using anti-CD4
microbeads (L3T4) according to the manufacturer’s instruc-
tions (Miltenyi Biotech, Auburn, CA). Purity was determined
by FACS analysis and ranged between 90 and 95% for
CD4+ expression. Cells were stimulated for 72 h in complete
medium (RPMI 1640 medium supplemented with 5% heat
inactivated FCS, 50 AM 2-ME, 1.0 mM sodium pyruvate,
100 U/ml penicillin, 0.1 mg/ml streptomycin) with plate-
bound anti-CD3 (1 Ag/ml) in the presence or absence of
soluble anti-CD28 (1 Ag/ml) at 37-C in a 5% CO2-
humidified atmosphere. Supernatants for cytokine measure-
ments were harvested after 72 h of cell culture and assayed
using the OptEIA ELISA kits according to manufacturer’s
instruction (BD PharMingen, San Diego, CA). Plates were
read on a microplate reader (BioRad, Richmond, CA).
Cytokine concentrations were calculated by MPM software
program based on the standard curves.
Flow cytometry
Cell surface staining was performed with splenocytes
depleted of erythrocytes. Data were collected on a LSR
flow cytometer (BD Biosciences) and analyzed using
CellQuest software. All staining profiles were based on
live gates, as determined by forward and side-scatter. Naıve
and memory subsets were distinguished by anti-CD44 and
anti-CD62L expression; in some cases, CD8+ T cell subsets
were also delineated with anti-CD122 and Ly6C. Intracel-
lular staining was performed with Cytofix/Cytoperm kit
(BD PharMingen, San Diego, CA) according to manufac-
ture’s instructions.
T cell proliferation with CFSE
For measurement of CFSE-incorporated proliferation, puri-
fied CD4+ T cells were incubated with CFSE (5 AM) in PBS for
10 min at 37-C, washed with cold media three times, and then
plated in anti-CD3 coated 96-well plates in the presence or
absence of anti-CD28 (1 Ag/ml) for 60 h. Cultures were
harvested and stained with PE-labeled anti-CD62L to distin-
guish naıve and memory subsets. The cells were collected on a
LSR flow cytometer (BD Biosciences) and analyzed using
CellQuest software.
T. Huang et al. / Clinical Immunology 117 (2005) 221–230224
Apoptosis assay
Purified CD4+ or CD8+ T cells (1 � 106 cells) were cultured
in 24-well plates coated with anti-CD3 and anti-CD28 as
described above, but also with anti-CD95. The cells were
cultured for 18 h, harvested, and stained with annexin-V and
propidium iodide (BD PharMingen). Cell death was measured
using a LSR flow cytometer as the %cells positive for annexin-
V or propidium iodide.
Results
Normal formation of systemic T cell populations in germ-free
mice
In germ-free mice lacking detectable enteric bacteria, B and T
cell populations associated with the intestinal mucosa are
dramatically altered in abundance and function. To evaluate
whether the changes in the mucosal compartment affected the
overall state of T cells in the periphery, we evaluated the
abundance of splenic CD4+ and CD8+ T cells in germ-free and
age-matched SPF mice. A minor but significant reduction was
observed in the absolute numbers of CD4+ T cells (Fig. 1a).
However, phenotypically identified naıve and memory subsets
of CD4+ T cells were within normal range (Fig. 1a), and no
differences from SPF mice were observed for CD8+ T cells and
their subsets (Fig. 1b). Analysis of these CD4+ and CD8+ T cell
subpopulations on a percent basis resulted in concordant
findings (data not shown). These results indicate that enteric
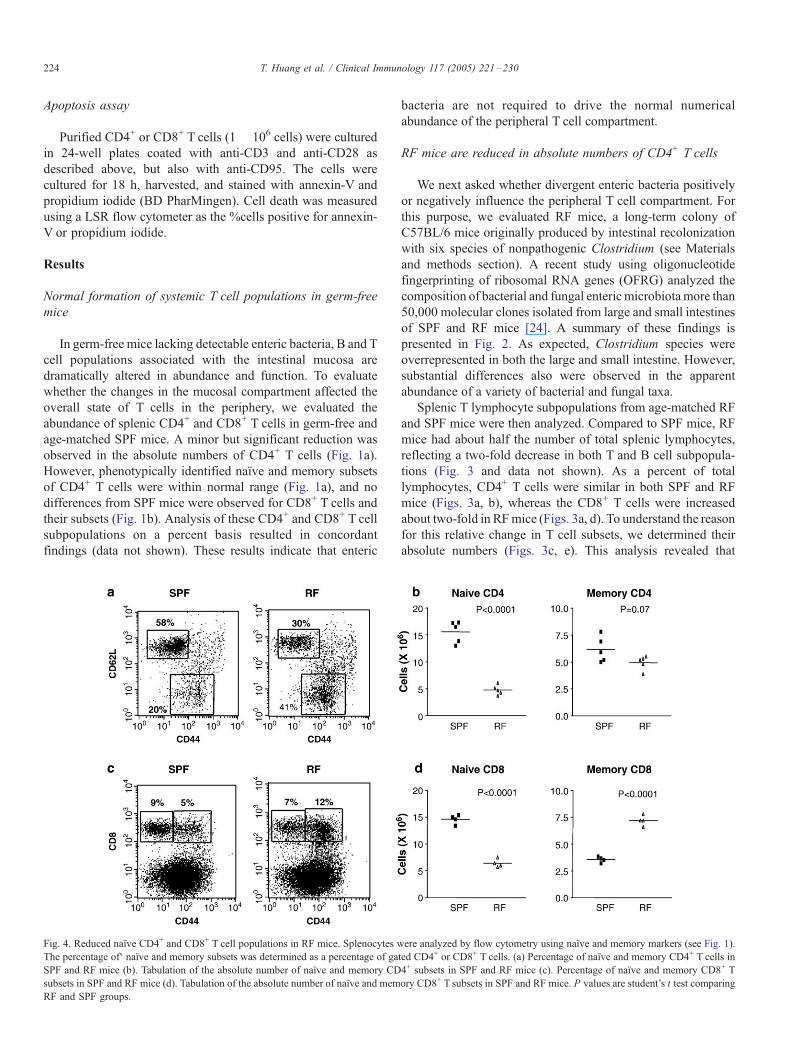
Fig. 4. Reduced naıve CD4+ and CD8+ T cell populations in RF mice. Splenocytes w
The percentage of‘ naıve and memory subsets was determined as a percentage of ga
SPF and RF mice (b). Tabulation of the absolute number of naıve and memory CD
subsets in SPF and RF mice (d). Tabulation of the absolute number of naıve and mem
RF and SPF groups.
bacteria are not required to drive the normal numerical
abundance of the peripheral T cell compartment.
RF mice are reduced in absolute numbers of CD4+ T cells
We next asked whether divergent enteric bacteria positively
or negatively influence the peripheral T cell compartment. For
this purpose, we evaluated RF mice, a long-term colony of
C57BL/6 mice originally produced by intestinal recolonization
with six species of nonpathogenic Clostridium (see Materials
and methods section). A recent study using oligonucleotide
fingerprinting of ribosomal RNA genes (OFRG) analyzed the
composition of bacterial and fungal entericmicrobiotamore than
50,000 molecular clones isolated from large and small intestines
of SPF and RF mice [24]. A summary of these findings is
presented in Fig. 2. As expected, Clostridium species were
overrepresented in both the large and small intestine. However,
substantial differences also were observed in the apparent
abundance of a variety of bacterial and fungal taxa.
Splenic T lymphocyte subpopulations from age-matched RF
and SPF mice were then analyzed. Compared to SPF mice, RF
mice had about half the number of total splenic lymphocytes,
reflecting a two-fold decrease in both T and B cell subpopula-
tions (Fig. 3 and data not shown). As a percent of total
lymphocytes, CD4+ T cells were similar in both SPF and RF
mice (Figs. 3a, b), whereas the CD8+ T cells were increased
about two-fold in RFmice (Figs. 3a, d). To understand the reason
for this relative change in T cell subsets, we determined their
absolute numbers (Figs. 3c, e). This analysis revealed that
ere analyzed by flow cytometry using naıve and memory markers (see Fig. 1).
ted CD4+ or CD8+ T cells. (a) Percentage of naıve and memory CD4+ T cells in
4+ subsets in SPF and RF mice (c). Percentage of naıve and memory CD8+ T
ory CD8+ T subsets in SPF and RF mice. P values are student’s t test comparing
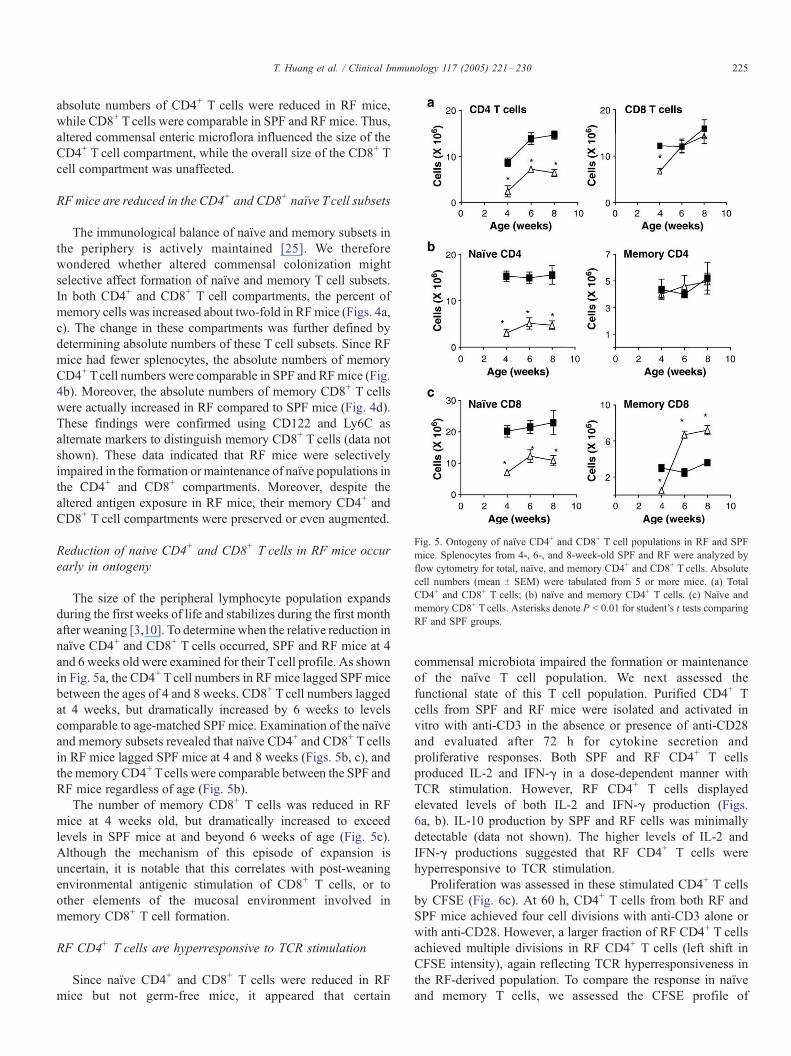
Fig. 5. Ontogeny of naıve CD4+ and CD8+ T cell populations in RF and SPF
mice. Splenocytes from 4-, 6-, and 8-week-old SPF and RF were analyzed by
flow cytometry for total, naıve, and memory CD4+ and CD8+ T cells. Absolute
cell numbers (mean T SEM) were tabulated from 5 or more mice. (a) Total
CD4+ and CD8+ T cells; (b) naıve and memory CD4+ T cells. (c) Naıve and
memory CD8+ T cells. Asterisks denote P < 0.01 for student’s t tests comparing
RF and SPF groups.
T. Huang et al. / Clinical Immunology 117 (2005) 221–230 225
absolute numbers of CD4+ T cells were reduced in RF mice,
while CD8+ Tcells were comparable in SPF and RF mice. Thus,
altered commensal enteric microflora influenced the size of the
CD4+ T cell compartment, while the overall size of the CD8+ T
cell compartment was unaffected.
RF mice are reduced in the CD4+ and CD8+ naıve Tcell subsets
The immunological balance of naıve and memory subsets in
the periphery is actively maintained [25]. We therefore
wondered whether altered commensal colonization might
selective affect formation of naıve and memory T cell subsets.
In both CD4+ and CD8+ T cell compartments, the percent of
memory cells was increased about two-fold in RFmice (Figs. 4a,
c). The change in these compartments was further defined by
determining absolute numbers of these T cell subsets. Since RF
mice had fewer splenocytes, the absolute numbers of memory
CD4+ Tcell numbers were comparable in SPF and RFmice (Fig.
4b). Moreover, the absolute numbers of memory CD8+ T cells
were actually increased in RF compared to SPF mice (Fig. 4d).
These findings were confirmed using CD122 and Ly6C as
alternate markers to distinguish memory CD8+ T cells (data not
shown). These data indicated that RF mice were selectively
impaired in the formation or maintenance of naıve populations in
the CD4+ and CD8+ compartments. Moreover, despite the
altered antigen exposure in RF mice, their memory CD4+ and
CD8+ T cell compartments were preserved or even augmented.
Reduction of naive CD4+ and CD8+ T cells in RF mice occur
early in ontogeny
The size of the peripheral lymphocyte population expands
during the first weeks of life and stabilizes during the first month
after weaning [3,10]. To determine when the relative reduction in
naıve CD4+ and CD8+ T cells occurred, SPF and RF mice at 4
and 6 weeks old were examined for their Tcell profile. As shown
in Fig. 5a, the CD4+ Tcell numbers in RF mice lagged SPF mice
between the ages of 4 and 8 weeks. CD8+ T cell numbers lagged
at 4 weeks, but dramatically increased by 6 weeks to levels
comparable to age-matched SPF mice. Examination of the naıve
and memory subsets revealed that naıve CD4+ and CD8+ T cells
in RF mice lagged SPF mice at 4 and 8 weeks (Figs. 5b, c), and
the memory CD4+ Tcells were comparable between the SPF and
RF mice regardless of age (Fig. 5b).
The number of memory CD8+ T cells was reduced in RF
mice at 4 weeks old, but dramatically increased to exceed
levels in SPF mice at and beyond 6 weeks of age (Fig. 5c).
Although the mechanism of this episode of expansion is
uncertain, it is notable that this correlates with post-weaning
environmental antigenic stimulation of CD8+ T cells, or to
other elements of the mucosal environment involved in
memory CD8+ T cell formation.
RF CD4+ T cells are hyperresponsive to TCR stimulation
Since naıve CD4+ and CD8+ T cells were reduced in RF
mice but not germ-free mice, it appeared that certain
commensal microbiota impaired the formation or maintenance
of the naıve T cell population. We next assessed the
functional state of this T cell population. Purified CD4+ T
cells from SPF and RF mice were isolated and activated in
vitro with anti-CD3 in the absence or presence of anti-CD28
and evaluated after 72 h for cytokine secretion and
proliferative responses. Both SPF and RF CD4+ T cells
produced IL-2 and IFN-g in a dose-dependent manner with
TCR stimulation. However, RF CD4+ T cells displayed
elevated levels of both IL-2 and IFN-g production (Figs.
6a, b). IL-10 production by SPF and RF cells was minimally
detectable (data not shown). The higher levels of IL-2 and
IFN-g productions suggested that RF CD4+ T cells were
hyperresponsive to TCR stimulation.
Proliferation was assessed in these stimulated CD4+ T cells
by CFSE (Fig. 6c). At 60 h, CD4+ T cells from both RF and
SPF mice achieved four cell divisions with anti-CD3 alone or
with anti-CD28. However, a larger fraction of RF CD4+ T cells
achieved multiple divisions in RF CD4+ T cells (left shift in
CFSE intensity), again reflecting TCR hyperresponsiveness in
the RF-derived population. To compare the response in naıve
and memory T cells, we assessed the CFSE profile of
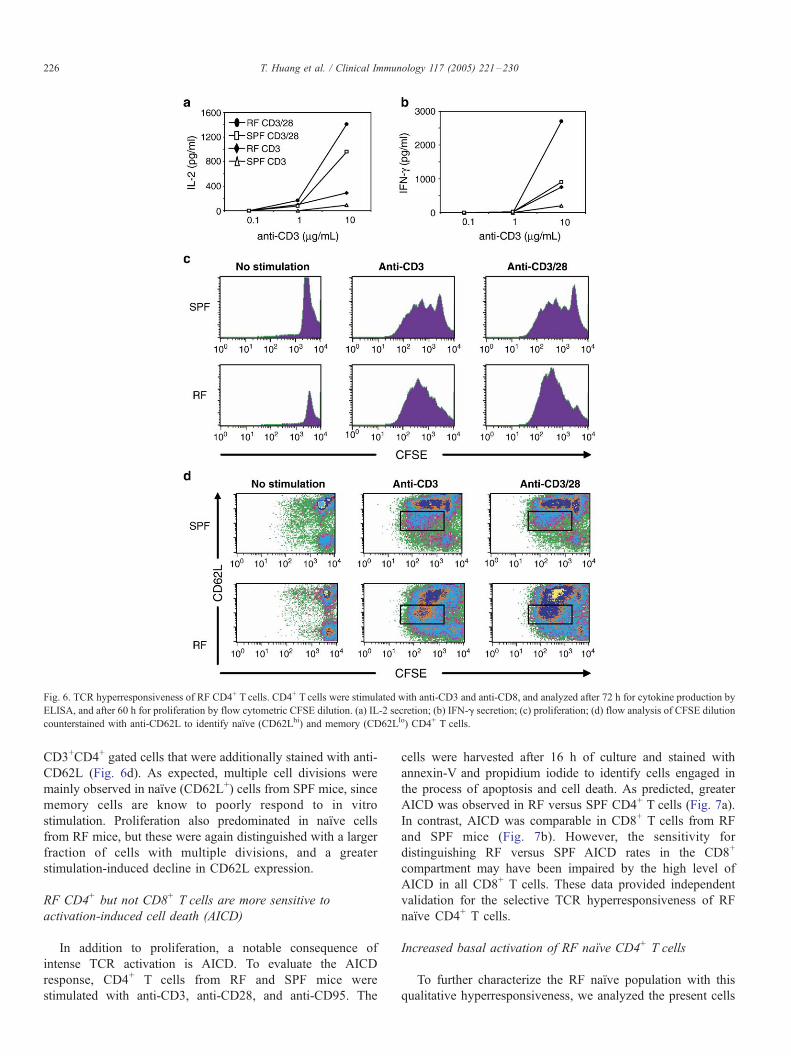
Fig. 6. TCR hyperresponsiveness of RF CD4+ T cells. CD4+ T cells were stimulated with anti-CD3 and anti-CD8, and analyzed after 72 h for cytokine production by
ELISA, and after 60 h for proliferation by flow cytometric CFSE dilution. (a) IL-2 secretion; (b) IFN-g secretion; (c) proliferation; (d) flow analysis of CFSE dilution
counterstained with anti-CD62L to identify naıve (CD62Lhi) and memory (CD62Llo) CD4+ T cells.
T. Huang et al. / Clinical Immunology 117 (2005) 221–230226
CD3+CD4+ gated cells that were additionally stained with anti-
CD62L (Fig. 6d). As expected, multiple cell divisions were
mainly observed in naıve (CD62L+) cells from SPF mice, since
memory cells are know to poorly respond to in vitro
stimulation. Proliferation also predominated in naıve cells
from RF mice, but these were again distinguished with a larger
fraction of cells with multiple divisions, and a greater
stimulation-induced decline in CD62L expression.
RF CD4+ but not CD8+ T cells are more sensitive to
activation-induced cell death (AICD)
In addition to proliferation, a notable consequence of
intense TCR activation is AICD. To evaluate the AICD
response, CD4+ T cells from RF and SPF mice were
stimulated with anti-CD3, anti-CD28, and anti-CD95. The
cells were harvested after 16 h of culture and stained with
annexin-V and propidium iodide to identify cells engaged in
the process of apoptosis and cell death. As predicted, greater
AICD was observed in RF versus SPF CD4+ T cells (Fig. 7a).
In contrast, AICD was comparable in CD8+ T cells from RF
and SPF mice (Fig. 7b). However, the sensitivity for
distinguishing RF versus SPF AICD rates in the CD8+
compartment may have been impaired by the high level of
AICD in all CD8+ T cells. These data provided independent
validation for the selective TCR hyperresponsiveness of RF
naıve CD4+ T cells.
Increased basal activation of RF naıve CD4+ T cells
To further characterize the RF naıve population with this
qualitative hyperresponsiveness, we analyzed the present cells
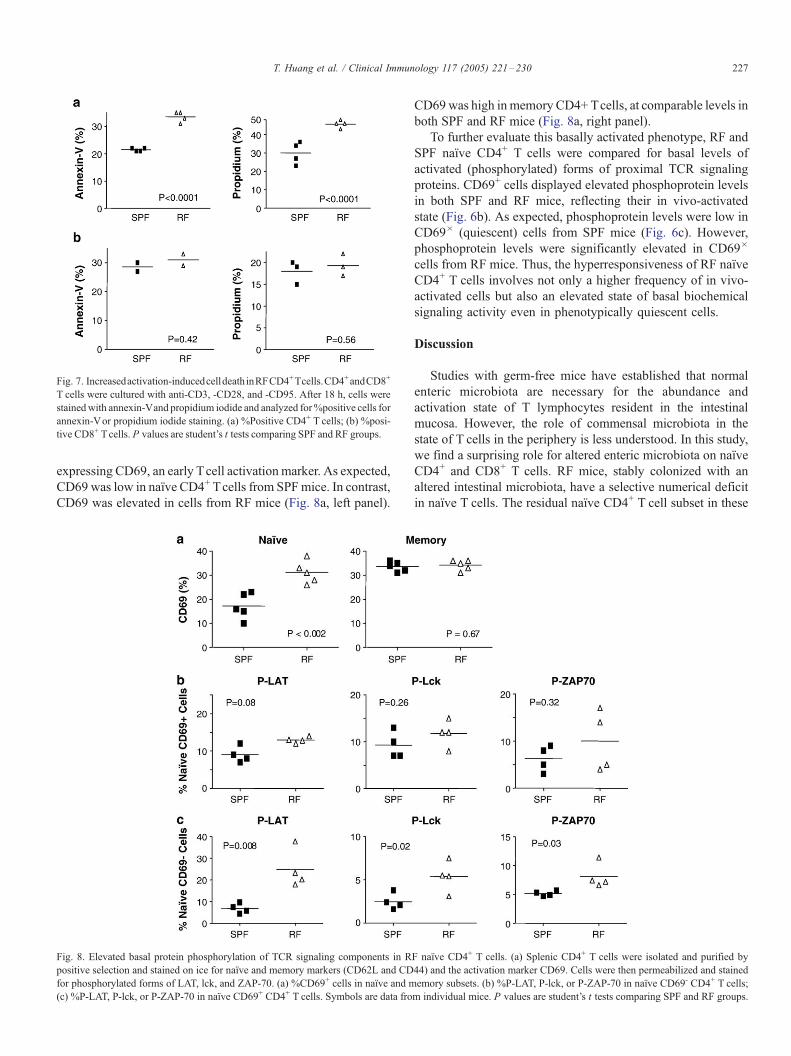
Fig. 7. Increasedactivation-inducedcelldeathinRFCD4+Tcells.CD4+andCD8+
T cells were cultured with anti-CD3, -CD28, and -CD95. After 18 h, cells were
stainedwith annexin-Vand propidium iodide and analyzed for%positive cells for
annexin-Vor propidium iodide staining. (a) %Positive CD4+ T cells; (b) %posi-
tive CD8+ Tcells. P values are student’s t tests comparing SPF and RF groups.
T. Huang et al. / Clinical Immunology 117 (2005) 221–230 227
expressing CD69, an early Tcell activation marker. As expected,
CD69 was low in naıve CD4+ Tcells from SPFmice. In contrast,
CD69 was elevated in cells from RF mice (Fig. 8a, left panel).
Fig. 8. Elevated basal protein phosphorylation of TCR signaling components in R
positive selection and stained on ice for naıve and memory markers (CD62L and CD
for phosphorylated forms of LAT, lck, and ZAP-70. (a) %CD69+ cells in naıve and m
(c) %P-LAT, P-lck, or P-ZAP-70 in naıve CD69+ CD4+ T cells. Symbols are data fro
CD69was high inmemory CD4+ Tcells, at comparable levels in
both SPF and RF mice (Fig. 8a, right panel).
To further evaluate this basally activated phenotype, RF and
SPF naıve CD4+ T cells were compared for basal levels of
activated (phosphorylated) forms of proximal TCR signaling
proteins. CD69+ cells displayed elevated phosphoprotein levels
in both SPF and RF mice, reflecting their in vivo-activated
state (Fig. 6b). As expected, phosphoprotein levels were low in
CD69� (quiescent) cells from SPF mice (Fig. 6c). However,
phosphoprotein levels were significantly elevated in CD69�
cells from RF mice. Thus, the hyperresponsiveness of RF naıve
CD4+ T cells involves not only a higher frequency of in vivo-
activated cells but also an elevated state of basal biochemical
signaling activity even in phenotypically quiescent cells.
Discussion
Studies with germ-free mice have established that normal
enteric microbiota are necessary for the abundance and
activation state of T lymphocytes resident in the intestinal
mucosa. However, the role of commensal microbiota in the
state of T cells in the periphery is less understood. In this study,
we find a surprising role for altered enteric microbiota on naıve
CD4+ and CD8+ T cells. RF mice, stably colonized with an
altered intestinal microbiota, have a selective numerical deficit
in naıve T cells. The residual naıve CD4+ T cell subset in these
F naıve CD4+ T cells. (a) Splenic CD4+ T cells were isolated and purified by
44) and the activation marker CD69. Cells were then permeabilized and stained
emory subsets. (b) %P-LAT, P-lck, or P-ZAP-70 in naıve CD69- CD4+ T cells;
m individual mice. P values are student’s t tests comparing SPF and RF groups.

T. Huang et al. / Clinical Immunology 117 (2005) 221–230228
mice are hyperresponsive, AICD-prone, and basally activated
in vivo. In this discussion, we consider the possible cellular
mechanisms for this aberrant T cell phenotype.
What properties of the commensal microbiota might
contribute to the abnormal peripheral T cell phenotype of RF
mice? RF mice differ from SPF mice in their composition of
commensal enteric bacterial and fungal taxa [24]. Microbial
composition accounts for the RF phenotype, since RF mice
conventionalized to SPF housing conditions at the time of birth
developed a normal, SPF-like T cell compartment (T. Huang
and B. Wei, unpublished data). It should be noted that
differences in microbial composition probably extend to other
commensal compartments (such as the skin or respiratory
system), so the biologically pertinent microorganisms may
reside in any of these anatomic sites.
Analysis of germ-free mice displayed a normal abundance
of the major splenic T cell subsets. This confirms that the
biologic niche required by the peripheral T cell compartment
forms independently of molecules derived from commensal
bacteria. Accordingly, the RF phenotype probably is not due to
the absence of certain microorganisms. Instead, it implies that
the increased action of unconventional commensals in RF mice
may impair formation or maintenance of the naıve T cell
compartment. While the microorganisms contributing to this
role are uncertain, candidates include microorganisms over-
represented in RF mice, including Clostridium and certain
fungi taxa.
What are the possible mechanisms that caused immunolog-
ically developmental defects in the RF mice? Various microbial
sensing systems, notably toll-like receptors (TLRs) in hemo-
poietic and epithelial cell types [26–28], play a critical role in
differentiating antigen presentation function by these cell types
[29–31]. Compared to memory T cells, naıve T cells are
particularly dependent on optimal functional state of antigen-
presenting cells [32–34]. Impairment of this differentiation
process is thus a plausible target for the RF phenotype,
exemplified by the microbial inactivation of NFkB signaling in
antigen-presenting cells [35–37]. As a second precedent, T cell
superantigens promote AICD in a process that is selective for
naıve T cells, and such molecules are produced by uncommon
enteric commensals [38,39].
Another possible cellular mechanism is the alterations of
cytokine/chemokine profiles in the RF mice, resulting in the
impaired recruitment and survival of naıve T cell subsets.
Innate microbial sensing through several receptor systems
elicits TNF-a and IL-12, which selectively differentiate naıve T
cells [29,40]. Dendritic cells produce a variety of chemokines
to recruit T cells [41–45], and molecules such as CXCL12 and
CCL21 increase the adhesion of naıve CD8 cells to the antigen
presenting cells in an antigen-independent manner [46].
Cytokines are also selective trophic for naıve versus memory
cell expansion, such as IL-2 versus IL-15 in CD8+ T cells
[33,34,47–55]. Mucosal homing and clonal expansion of
intraepithelial and mesenteric T cells are impaired in RF mice
[23,56]. This phenotype in part may be due to a deficiency in
the commensals producing innate and adaptive antigenic
stimuli required for normal mucosal immune system formation,
as observed in germ-free mice. However, the present study
suggests that the mucosal T cell deficiency also may be
compounded by the reduced abundance and aberrant functional
state of the systemic naıve T cell compartment.
A surprising feature of the naıve T cell population in RF
mice was their TCR hyperresponsiveness and elevated basal
state of activation in vivo. CD4+ T cells from mice loss of
function mutations in positive regulators such as ZAP-70 and
LAT exhibited hyporesponsiveness to TCR-mediated stimula-
tion, whereas overexpression of these molecules reduces in
hyperresponsiveness [57–59]. Conversely, deficiency of neg-
ative regulators such as c-cbl and CTLA-4 has a lymphopro-
liferative phenotype [60–63]. In RF T cells, an elevated basal
state of activation involved the entire proximal segment of
TCR signaling. This finding suggests that the RF T cell
disorder might involve abortive TCR signaling, such as
observed with partial peptide agonists, or due to aberrant T
cell expression of regulatory molecules affecting the phos-
phorylation state of the CD3 complex itself [64,65]. It is also
possible that this phenotype is augmented by some degree of
homeostatic expansion in these partially lymphopenic mice
[66,67].
In contrast to the phenotype of RF mice, increased AICD
might be expected to mainly impair formation of memory cells
and preserve naıve T cells. However, the ultimate size of the
memory pool is determined by many additional factors,
including the degree of recurrent antigenic challenge and
immunoregulatory processes; moreover, the maintenance of the
memory pool is relatively independent of TCR stimulation
[68,69]. With respect to naıve cells, premature activation and
AICD would be expected to deplete the naıve T cell pool. Thus,
a reduced antigenic threshold for activation and AICD in naıve
T cells is a plausible factor in the selective depletion of these
cells in RF mice.
The hyperresponsiveness of RF T cells reflects a discrete,
systemic T cell abnormality resulting from altered commensal
microbiota. The epidemiology of allergy, atopy, and certain
autoimmune diseases relates this immunologic susceptibility to
certain profiles of microbial colonization or antigenic exposure,
including products of helminths, bacteria, and fungi [70–75].
The cellular mechanisms underlying this ‘‘hygiene hypothesis’’
are relatively unaddressed, although particularly in the case of
helminths, model systems suggest that reduced TCR respon-
siveness is one important factor. The present study indicates that
the RF phenotype may provide an animal model for this
epidemiologic relationship and could be used to delineate the
microbial products and host traits resulting in the hyperrespon-
sive T cell trait. It also suggests a set of T cell biomarkers that
might be used to determine whether this phenotype is observed
in a subset of individuals at risk for atopy in the human
population.
Acknowledgments
Supported by NIH DK46753 (J.B.), DK69434 (J.B.),
DK43026 (CURE), AI07126 (T.H.), the Eli and Edythe L.
Broad Medical Research Foundation (J.B., J.B.), the Crohn’s

T. Huang et al. / Clinical Immunology 117 (2005) 221–230 229
and Colitis Foundation of America (B.W.), and the Jonsson
Comprehensive Cancer Center. Flow cytometry was performed
in the UCLA Jonsson Comprehensive Cancer Center for AIDS
Research Flow Cytometry Core Facility, supported by NIH
CA-16042 and AI-28697. Germ-free mice were produced
by and analyzed at the Center for Gastrointestinal Biology
and Disease, UNC-Chapel Hill School of Medicine (NIH
DK34987, R. Balfour Sartor, P.I.).
References
[1] R.D. Berg, The indigenous gastrointestinal microflora, Trends Microbiol.
4 (1996) 430–435.
[2] S. Casola, K.L. Otipoby, M. Alimzhanov, S. Humme, N. Uyttersprot, J.L.
Kutok, M.C. Carroll, K. Rajewsky, B cell receptor signal strength
determines B cell fate, Nat. Immunol. 5 (2004) 317–327.
[3] S. Fagarasan, M. Muramatsu, K. Suzuki, H. Nagaoka, H. Hiai, T. Honjo,
Critical roles of activation-induced cytidine deaminase in the homeostasis
of gut flora, Science 298 (2002) 1424–1427.
[4] T. Shikina, T. Hiroi, K. Iwatani, M.H. Jang, S. Fukuyama, M. Tamura, T.
Kubo, H. Ishikawa, H. Kiyono, IgA class switch occurs in the organized
nasopharynx- and gut-associated lymphoid tissue, but not in the diffuse
lamina propria of airways and gut, J. Immunol. 172 (2004) 6259–6264.
[5] H. Hamada, T. Hiroi, Y. Nishiyama, H. Takahashi, Y. Masunaga, S.
Hachimura, S. Kaminogawa, H. Takahashi-Iwanaga, T. Iwanaga, H.
Kiyono, H. Yamamoto, H. Ishikawa, Identification of multiple isolated
lymphoid follicles on the antimesenteric wall of the mouse small intestine,
J. Immunol. 168 (2002) 57–64.
[6] R.G. Lorenz, D.D. Chaplin, K.G. McDonald, J.S. McDonough, R.D.
Newberry, Isolated lymphoid follicle formation is inducible and
dependent upon lymphotoxin-sufficient B lymphocytes, lymphotoxin
beta receptor, and TNF receptor I function, J. Immunol. 170 (2003)
5475–5482.
[7] P.M. Bealmear, E.A. Mirand, O.A. Holtermann, Miscellaneous immune
defects in gnotobiotic and SPF mice, Prog. Clin. Biol. Res. 132C (1983)
423–432.
[8] T. Yamanaka, L. Helgeland, I.N. Farstad, H. Fukushima, T. Midtvedt, P.
Brandtzaeg, Microbial colonization drives lymphocyte accumulation and
differentiation in the follicle-associated epithelium of Peyer’s patches,
J. Immunol. 170 (2003) 816–822.
[9] L.V. Hooper, T. Midtvedt, J.I. Gordon, How host-microbial interactions
shape the nutrient environment of the mammalian intestine, Annu. Rev.
Nutr. 22 (2002) 283–307.
[10] J.J. Cebra, Influences of microbiota on intestinal immune system
development, Am. J. Clin. Nutr. 69 (1999) 1046S–1051S.
[11] R.D. Berg, D.C. Savage, Immunological responses and microorganisms
indigenous to the gastrointestinal tract, Am. J. Clin. Nutr. 25 (1972)
1364–1371.
[12] V. Gaboriau-Routhiau, M.C. Moreau, Gut flora allows recovery of oral
tolerance to ovalbumin in mice after transient breakdown mediated by
cholera toxin or Escherichia coli heat-labile enterotoxin, Pediatr. Res.
39 (1996) 625–629.
[13] M.C. Moreau, V. Gaboriau-Routhiau, The absence of gut flora, the doses
of antigen ingested and aging affect the long-term peripheral tolerance
induced by ovalbumin feeding in mice, Res. Immunol. 147 (1996) 49–59.
[14] N. Sudo, S. Sawamura, K. Tanaka, Y. Aiba, C. Kubo, Y. Koga, The
requirement of intestinal bacterial flora for the development of an IgE
production system fully susceptible to oral tolerance induction,
J. Immunol. 159 (1997) 1739–1745.
[15] M.A. Breban, M.C. Moreau, C. Fournier, R. Ducluzeau, M.F. Kahn,
Influence of the bacterial flora on collagen-induced arthritis in susceptible
and resistant strains of rats, Clin. Exp. Rheumatol. 11 (1993) 61–64.
[16] A.A. Like, A.A. Rossini, D.L. Guberski, M.C. Appel, R.M.
Williams, Spontaneous diabetes mellitus: reversal and prevention in
the BB/W rat with antiserum to rat lymphocytes, Science 206 (1979)
1421–1423.
[17] A.A. Rossini, R.M. Williams, J.P. Mordes, M.C. Appel, A.A. Like,
Spontaneous diabetes in the gnotobiotic BB/W rat, Diabetes 28 (1979)
1031–1032.
[18] O. Kohashi, J. Kuwata, K. Umehara, F. Uemura, T. Takahashi, A. Ozawa,
Susceptibility to adjuvant-induced arthritis among germfree, specific-
pathogen-free, and conventional rats, Infect. Immun. 26 (1979) 791–794.
[19] O. Kohashi, Y. Kohashi, T. Takahashi, A. Ozawa, N. Shigematsu,
Reverse effect of gram-positive bacteria vs. gram-negative bacteria on
adjuvant-induced arthritis in germfree rats, Microbiol. Immunol. 29
(1985) 487–497.
[20] D. Lehmann, A. Ben, Nun, Bacterial agents protect against autoimmune
disease: I. Mice pre-exposed to Bordetella pertussis or Mycobacterium
tuberculosis are highly refractory to induction of experimental autoim-
mune encephalomyelitis, J. Autoimmun. 5 (1992) 675–690.
[21] R. Duchmann, E. May, M. Heike, P. Knolle, M. Neurath, B.K. Zum, T cell
specificity and cross reactivity towards enterobacteria, Bacteroides,
Bifidobacterium, and antigens from resident intestinal flora in humans,
Gut 44 (1999) 812–818.
[22] R. Duchmann, I. Kaiser, E. Hermann, W. Mayet, K. Ewe, Z.B. Meyer,
Tolerance exists towards resident intestinal flora but is broken in active
inflammatory bowel disease (IBD), Clin. Exp. Immunol. 102 (1995)
448–455.
[23] V. Camerini, B.C. Sydora, R. Aranda, C. Nguyen, C. MacLean, W.H.
McBride, M. Kronenberg, Generation of intestinal mucosal lymphocytes
in SCID mice reconstituted with mature, thymus-derived T cells,
J. Immunol. 160 (1998) 2608–2618.
[24] A.J. Scupham, L.L. Presley, B. Wei, E. Bent, N. Griffith, M. McPherson,
F. Zhu, O. Oluwadara, N. Rao, J. Braun, J. Borneman, Abundant and
diverse fungal microbiota in the murine intestine. Appl. Environ.
Microbiol. (in press).
[25] L. Van Parijs, A.K. Abbas, Homeostasis and self-tolerance in the immune
system: turning lymphocytes off, Science 280 (1998) 243–248.
[26] S. Akira, Toll-like receptors and innate immunity, Adv. Immunol. 78
(2001) 1–56.
[27] C. Pasare, R. Medzhitov, Toll-like receptors: balancing host resistance
with immune tolerance, Curr. Opin. Immunol. 15 (2003) 677–682.
[28] A. Poltorak, X. He, I. Smirnova, M.Y. Liu, C. van Huffel, X. Du, D.
Birdwell, E. Alejos, M. Silva, C. Galanos, M. Freudenberg, P. Ricciardi-
Castagnoli, B. Layton, B. Beutler, Defective LPS signaling in C3H/HeJ
and C57BL/10ScCr mice: mutations in Tlr4 gene, Science 282 (1998)
2085–2088.
[29] N.V. Serbina, W. Kuziel, R. Flavell, S. Akira, B. Rollins, E.G. Pamer,
Sequential MyD88-independent and -dependent activation of innate
immune responses to intracellular bacterial infection, Immunity 19
(2003) 891–901.
[30] T. Kaisho, K. Hoshino, T. Iwabe, O. Takeuchi, T. Yasui, S. Akira,
Endotoxin can induce MyD88-deficient dendritic cells to support T(h)2
cell differentiation, Int. Immunol. 14 (2002) 695–700.
[31] A.I. Chin, P.W. Dempsey, K. Bruhn, J.F. Miller, Y. Xu, G. Cheng,
Involvement of receptor-interacting protein 2 in innate and adaptive
immune responses, Nature 416 (2002) 190–194.
[32] D.F. Tough, P. Borrow, J. Sprent, Induction of bystander T cell
proliferation by viruses and type I interferon in vivo, Science 272
(1996) 1947–1950.
[33] C. Tanchot, F.A. Lemonnier, B. Perarnau, A.A. Freitas, B. Rocha,
Differential requirements for survival and proliferation of CD8 naive or
memory T cells, Science 276 (1997) 2057–2062.
[34] S. Garcia, J. DiSanto, B. Stockinger, Following the development of a CD4
T cell response in vivo: from activation to memory formation, Immunity
11 (1999) 163–171.
[35] A.S. Neish, A.T. Gewirtz, H. Zeng, A.N. Young, M.E. Hobert, V. Karmali,
A.S. Rao, J.L. Madara, Prokaryotic regulation of epithelial responses by
inhibition of IkappaB-alpha ubiquitination [see comments], Science 289
(2000) 1560–1563.
[36] T. Lawrence, M. Bebien, G.Y. Liu, V. Nizet, M. Karin, IKKalpha limits
macrophage NF-kappaB activation and contributes to the resolution of
inflammation, Nature 434 (2005) 1138–1143.
[37] D. Kelly, J.I. Campbell, T.P. King, G. Grant, E.A. Jansson, A.G. Coutts, S.

T. Huang et al. / Clinical Immunology 117 (2005) 221–230230
Pettersson, S. Conway, Commensal anaerobic gut bacteria attenuate
inflammation by regulating nuclear-cytoplasmic shuttling of PPAR-
gamma and RelA, Nat. Immunol. 5 (2004) 104–112.
[38] H. Dalwadi, B. Wei, M. Kronenberg, C.L. Sutton, J. Braun, The Crohn’s
disease-associated bacterial protein I2 is a novel enteric T cell super-
antigen, Immunity 15 (2001) 149–158.
[39] B. Wei, T. Huang, H. Dalwadi, C.L. Sutton, D. Bruckner, J. Braun,
Pseudomonas fluorescens encodes the Crohn’s disease-associated I2
sequence and T-cell superantigen, Infect. Immun. 70 (2002) 6567–6575.
[40] T. Kaisho, O. Takeuchi, T. Kawai, K. Hoshino, S. Akira, Endotoxin-
induced maturation of MyD88-deficient dendritic cells, J. Immunol. 166
(2001) 5688–5694.
[41] J.J. Campbell, K.E. Murphy, E.J. Kunkel, C.E. Brightling, D. Soler, Z.
Shen, J. Boisvert, H.B. Greenberg, M.A. Vierra, S.B. Goodman, M.C.
Genovese, A.J. Wardlaw, E.C. Butcher, L. Wu, CCR7 expression and
memory T cell diversity in humans, J. Immunol. 166 (2001) 877–884.
[42] E.C. Butcher, L.J. Picker, Lymphocyte homing and homeostasis, Science
272 (1996) 60–66.
[43] F. Sallusto, D. Lenig, R. Forster, M. Lipp, A. Lanzavecchia, Two subsets
of memory T lymphocytes with distinct homing potentials and effector
functions, Nature 401 (1999) 708–712.
[44] C. Schaniel, E. Pardali, F. Sallusto, M. Speletas, C. Ruedl, T. Shimizu,
T. Seidl, J. Andersson, F. Melchers, A.G. Rolink, P. Sideras, Activated
murine B lymphocytes and dendritic cells produce a novel CC
chemokine which acts selectively on activated T cells, J. Exp. Med.
188 (1998) 451–463.
[45] C. Sousa, A. Sher, P. Kaye, The role of dendritic cells in the induction and
regulation of immunity to microbial infection, Curr. Opin. Immunol. 11
(1999) 392–399.
[46] S.K. Bromley, D.A. Peterson, M.D. Gunn, M.L. Dustin, Cutting edge:
hierarchy of chemokine receptor and TCR signals regulating T cell
migration and proliferation, J. Immunol. 165 (2000) 15–19.
[47] J. Sprent, D.F. Tough, T cell death and memory, Science 293 (2001)
245–248.
[48] K.S. Schluns, K. Williams, A. Ma, X.X. Zheng, L. Lefrancois, Cutting
edge: requirement for IL-15 in the generation of primary and memory
antigen-specific CD8 T cells, J. Immunol. 168 (2002) 4827–4831.
[49] A.L. Marzo, V. Vezys, K. Williams, D.F. Tough, L. Lefrancois, Tissue-
level regulation of Th1 and Th2 primary and memory CD4 T cells in
response to Listeria infection, J. Immunol. 168 (2002) 4504–4510.
[50] X. Zhang, S. Sun, I. Hwang, D.F. Tough, J. Sprent, Potent and selective
stimulation of memory-phenotype CD8+ T cells in vivo by IL-15,
Immunity 8 (1998) 591–599.
[51] T.A. Waldmann, S. Dubois, Y. Tagaya, Contrasting roles of IL-2 and IL-15
in the life and death of lymphocytes: implications for immunotherapy,
Immunity 14 (2001) 105–110.
[52] P. Marrack, T. Mitchell, D. Hildeman, R. Kedl, T.K. Teague, J. Bender, W.
Rees, B.C. Schaefer, J. Kappler, Genomic-scale analysis of gene
expression in resting and activated T cells, Curr. Opin. Immunol. 12
(2000) 206–209.
[53] T.C. Becker, E.J. Wherry, D. Boone, K. Murali-Krishna, R. Antia, A. Ma,
R. Ahmed, Interleukin 15 is required for proliferative renewal of virus-
specific memory CD8 T cells, J. Exp. Med. 195 (2002) 1541–1548.
[54] J.P. Lodolce, D.L. Boone, S. Chai, R.E. Swain, T. Dassopoulos, S.
Trettin, A. Ma, IL-15 receptor maintains lymphoid homeostasis by
supporting lymphocyte homing and proliferation, Immunity 9 (1998)
669–676.
[55] J. Marks-Konczalik, S. Dubois, J.M. Losi, H. Sabzevari, N. Yamada, L.
Feigenbaum, T.A. Waldmann, Y. Tagaya, IL-2-induced activation-induced
cell death is inhibited in IL-15 transgenic mice, Proc. Natl. Acad. Sci.
U. S. A. 97 (2000) 11445–11450.
[56] V. Camerini, C. Panwala, M. Kronenberg, Regional specialization of the
mucosal immune system: intraepithelial lymphocytes of the large intestine
have a different phenotype and function than those of the small intestine,
J. Immunol. 151 (1993) 1765–1776.
[57] E.N. Neumeister, Y. Zhu, S. Richard, C. Terhorst, A.C. Chan, A.S.
Shaw, Binding of ZAP-70 to phosphorylated T-cell receptor zeta and
eta enhances its autophosphorylation and generates specific binding
sites for SH2 domain-containing proteins, Mol. Cell. Biol. 15 (1995)
3171–3178.
[58] L. Zeitlmann, T. Knorr, M. Knoll, C. Romeo, P. Sirim, W. Kolanus, T cell
activation induced by novel gain-of-function mutants of Syk and ZAP-70,
J. Biol. Chem. 273 (1998) 15445–15452.
[59] T.S. Finco, T. Kadlecek, W. Zhang, L.E. Samelson, A. Weiss, LAT is
required for TCR-mediated activation of PLCgamma1 and the Ras
pathway, Immunity 9 (1998) 617–626.
[60] K.M. Murphy, W. Ouyang, J.D. Farrar, J. Yang, S. Ranganath, H. Asnagli,
M. Afkarian, T.L. Murphy, Signaling and transcription in T helper
development, Annu. Rev. Immunol. 18 (2000) 451–494.
[61] M. Naramura, I.K. Jang, H. Kole, F. Huang, D. Haines, H. Gu, c-Cbl and
Cbl-b regulate T cell responsiveness by promoting ligand-induced TCR
down-modulation, Nat. Immunol. 3 (2002) 1192–1199.
[62] E.A. Tivol, F. Borriello, A.N. Schweitzer, W.P. Lynch, J.A. Bluestone,
A.H. Sharpe, Loss of CTLA-4 leads to massive lymphoproliferation and
fatal multiorgan tissue destruction, revealing a critical negative regulatory
role of CTLA-4, Immunity 3 (1995) 541–547.
[63] C.A. Chambers, T.J. Sullivan, J.P. Allison, Lymphoproliferation in CTLA-
4-deficient mice is mediated by costimulation-dependent activation of
CD4+ T cells, Immunity 7 (1997) 885–895.
[64] E.N. Kersh, A.S. Shaw, P.M. Allen, Fidelity of T cell activation through
multistep T cell receptor zeta phosphorylation, Science 281 (1998)
572–575.
[65] J. Sloan-Lancaster, A.S. Shaw, J.B. Rothbard, P.M. Allen, Partial T cell
signaling: altered phospho-zeta and lack of zap70 recruitment in APL-
induced T cell anergy, Cell 79 (1994) 913–922.
[66] H. Gudmundsdottir, L.A. Turka, A closer look at homeostatic proliferation
of CD4+ T cells: costimulatory requirements and role in memory
formation, J. Immunol. 167 (2001) 3699–3707.
[67] B. Seddon, G. Legname, P. Tomlinson, R. Zamoyska, Long-term survival
but impaired homeostatic proliferation of naive T cells in the absence of
p56lck, Science 290 (2000) 127–131.
[68] P.R. Burkett, R. Koka, M. Chien, D.L. Boone, A. Ma, Generation,
maintenance, and function of memory T cells, Adv. Immunol. 83 (2004)
191–231.
[69] J. Sprent, C.D. Surh, T cell memory, Annu. Rev. Immunol. 20 (2002)
551–579.
[70] G. Borkow, Q. Leng, Z. Weisman, M. Stein, N. Galai, A. Kalinkovich, Z.
Bentwich, Chronic immune activation associated with intestinal helminth
infections results in impaired signal transduction and anergy, J. Clin.
Invest. 106 (2000) 1053–1060.
[71] R.M. Maizels, M. Yazdanbakhsh, Immune regulation by helminth
parasites: cellular and molecular mechanisms, Nat. Rev., Immunol. 3
(2003) 733–744.
[72] M. Kalliomaki, P. Kirjavainen, E. Eerola, P. Kero, S. Salminen, E. Isolauri,
Distinct patterns of neonatal gut microflora in infants in whom atopy was
and was not developing, J. Allergy Clin. Immunol. 107 (2001) 129–134.
[73] M. Kalliomaki, S. Salminen, H. Arvilommi, P. Kero, P. Koskinen, E.
Isolauri, Probiotics in primary prevention of atopic disease: a randomised
placebo-controlled trial, Lancet 357 (2001) 1076–1079.
[74] G.A.W. Rook, L.R. Brunet, Microbes, immunregulation, and the gut, Gut
54 (2005) 317–320.
[75] C. Braun-Fahrlander, J. Riedler, U. Herz, W. Eder, M. Waser, L. Grize, S.
Maisch, D. Carr, F. Gerlach, A. Bufe, R.P. Lauener, D. Nowak, E. von
MutiusAllergy and Endotoxin Study Team, Environmental exposure to
endotoxin and its relation to asthma in school-age children, N. Engl. J.
Med. 347 (2002) 869–877.





























