Embed Size (px)
Citation preview

JOURNAL OF CELLULAR PHYSIOLOGY 156:443452 (1993)
Comparative Analysis of Mitogenic and Morphogenic Effects of HGF and EGF on Rat
and Human Hepatocytes Maintained in Collagen Gels
GEORGE K. MICHALOPOULOS,* WILLIAM BOWEN, ANDREAS K. NUSSLER, MICHAEL J. BECICH, AND TAMARA A. H O W A R D
Departments of Pathology (C.K.M., W. B., M.1. B., T.A.H.) and Surgery (A.K.N.), University of Pittsburgh, Pittsburgh, Pennsy/vania 1526 1
Hepatocytes maintained in collagen gels remain differentiated for prolonged peri- ods of time compared to cells maintained on conventional cultures. Previous studies with other culture systems in which chemical supplements or substratum modifications enhanced hepatocyte differentiation showed that in all of these systems hepatocytes do not respond to mitogens. In this study it is shown that hepatocytes maintained between two layers of collagen gels respond to mitogens HGF (also known as scatter factor (HGF/SF)) and epidermal growth factor (EGF). Cell density did not affect the responsiveness to mitogens as in conventional cultures. In addition both mitogens (HGF more pronounced) induce characteristic morphogenic changes in which hepatocytes form processes and join in formation of cords. Hepatocytes respond to mitogens for up to 6 days in culture at which point they become refractory to fu r the r mitogenic stimulation. This occurs despite electron microscopic evidence that these cells are fully viable when they become refractory to mitogenesis. The refractory state is not modified by substitution of one growth factor for the other or by addition of growth factors at different times. Hepatocytes in the refractory state become again responsive to mitogens when the collagen gels are dispersed by collagenase and the cells are replated on conven- tional substrates. o 1993 WiIey-Liss, Inc.
Primary cultures of hepatocytes maintained on plas- tic or collagen-coated rigid plates respond to complete mitogens with DNA synthesis limited to a few rounds of DNA replication (Vintermyr and Doskeland, 1987; Michalopoulos et al., 1982). These systems have been used as bioassays to identify the complete mitogens and comitogens for hepatocytes (Michalopoulos and Zarne- gar, 1992; Michalopoulos, 1990). Other studies have shown that modification of the substrate of culture has profound effects on hepatocyte gene expression and cell proliferation (DiPersio et al., 1991; Jefferson et al., 1984). Hepatocytes maintained on high laminin gels derived from the EHS sarcoma express differentiation- related genes for prolonged periods of time (Bissell et al., 1987). The same is true for hepatocytes maintained in 2% dimethyl sulfoxide (DMSO) (Isom et al., 1985; Kost and Michalopoulos, 1991) or 3 mM phenobarbital (Chan et al., 1990). In the latter three systems, while hepatocyte differentiation and liver-specific gene ex- pression is prolonged, hepatocytes cease responding to mitogens.
Previous studies have shown that collagen gels com- posed primarily of type I collagen also enhance hepato- cyte differentiation and longevity (Michalopoulos and Pitot, 1975). Improved performance was achieved when hepatocytes were maintained in between two layers of 0 1993 WILEY-LISS, INC.
collagen gels (Dunn et al., 1992). In this study we ex- plored the response to the complete mitogens HGF and epidermal growth factor (EGF) of hepatocytes main- tained between two layers of collagen gels. These stud- ies were also extended to the observation of the morpho- logic changes induced by these mitogens on hepatocytes in this system. HGF has been identified as a potent motogenic and morphogenic factor for other cell types in collagen gels, including endothelial cells (Bussolino et al., 19921, the kidney-derived MDCK cell line (Ue- hara and Kitamura, 1992), and keratinocytes (Matsu- mot0 et al., 1991). In this study, we extended these findings to hepatocytes and compared the morphogenic effects of HGF and EGF. Comparisons were also made between rat and human hepatocytes.
MATERIALS AND METHODS Materials
Male Fischer 344 rats from Charles River were used for all the experiments involving hepatocyte isolation. EGF was obtained from Collaborative Research
Received January 20,1993; accepted March 23,1993. *To whom reprint requestskorrespondence should be addressed.

444 MICHALOPOULOS ET AL.
(Waltham, Massachusetts). 3H Thymidine was ob- tained from ICN Radiochemicals, Irvine, California. Collagenase for hepatocyte isolation was obtained from Boehringer, Mannheim, Germany. Vitrogen (from Cel- trix Labs., Palo Alto, California) was used for the con- struction of the collagen gels. General reagents were obtained from Sigma (St. Louis, MO). EGF was pur- chased from Collaborative Research. HGF was kindly donated by Genentech.
Isolation and culture Isolation and culture of rat hepatocytes were per-
formed by an adaptation of the calcium two-step colla- genase perfusion technique as previously described from our laboratory (Kost and Michalopoulos, 1991). For cell culture, minimal essential medium (MEM) with Earle's salts and with nonessential amino acids was supplemented with 1 mM pyruvate, gentamicin 1 pg/ml, and 10' M insulin. Growth factors were added as required for the experiments. Hepatocytes were plated on a single layer of collagen gel (see below) and left to attach for 2 hours. Twelve well cluster plates (3.8 cm2 per plate) from Corning were used. Unless otherwise specified, 100,000 hepatocytes were inoculated per plate for all the experiments described, except for the data shown in Figure 5 in which cell density was varied as described. Subsequently, another layer of collagen gel was overlayed on top of the attached hepatocytes. The medium was changed at 2 hours after cells were plated and every 48 hours thereafter. Thymidine, growth factors, etc. were added at the time of medium change as required.
Human hepatocytes Human hepatocytes were isolated by an adaptation
of the collagenase perfusion technique as described pre- viously from our laboratory (Strom et al., 1982). Cells were cultured as described above for the rat hepatocytes.
Collagen gels Vitrogen solution from Celtrix was used as the source
of collagen. This solution is derived from acid extrac- tion of bovine achilles tendon and consists primarily of type I collagen. Collagen gel solution was prepared as previously described (Michalopoulos and Pitot, 1975) by mixing on ice ten parts ofvitrogen, 0.55 parts of 0.34 M NaOH, and 1 part of 1OX concentrated Waymouth medium. The freshly prepared solution was kept on ice and was poured into plastic plate wells kept at room temperature. The solution became, gel within 10-30 minutes after pouring into the wells. Gels formed faster at higher (up to 37°C) temperatures. The underlay and overlay gel volume each was .25 ml/well.
DNA synthesis DNA synthesis was measured by uptake of tritiated
thymidine into trichloroacetic acid (TCA) precipitable material as previously used in our laboratory (Michalo- poulos et al., 1982; Kost and Michalopoulos, 1991). Col- lagen gels were digested with 2 mg of collagenase/ml of MEM. Incubation was done for 30 minutes at 37°C. The digested gels were treated with NaOH followed by TCA
in order to precipitate DNA, RNA, and proteins as pre- viously described (Kost and Michalopoulos, 1991).
Replating of hepatocytes from day 6 collagen gels
Media overlaying collagen gels were removed and 1 ml of collagenase solution (1 mg/ml) in MEM supple- mented as above and with 5% serum was added to the wells. Collagenase solution was sterilized by filtration through 0.22 m filters. The wells were incubated at 37°C in 5% C02-95% air until collagen gels were dis- solved. This usually required 30-40 minutes of incuba- tion. After digestion, the total contents of each well (dissolved gels, media, collagenase, etc.) were diluted 1:3 with ice cold MEN supplemented as above and with 5% serum. The suspension was centrifuged at 170g for 10 minutes. The supernatant was removed and the cell pellet was resuspended with 10 cc of MEM as supple- mented before. This process was repeated three times to remove any residual collagenase. The final resuspen- sion was done in medium at the required volume for plating the cells. The isolated cells were inoculated into collagen-coated plates and allowed 30 minutes to reat- tach. After 30 minutes, the media were changed to se- rum free and growth factors to stimulate DNA synthe- sis were added at that time.
Transmission electron microscopy Samples for transmission electron microscopy (TEM)
were fixed on the culture plates for 1-1.5 hours in 0.1 M cacodylate buffer (pH 7.4) containing 2.5% glutaralde- hyde and 2% paraformaldehyde. The plates were then rinsed two times with 0.1 M cacodylate buffer (pH 7.4) and two times with 0.1 M cacodylate buffer containing 5% sucrose (pH 7.4). They were held in the sucrose buffer for 1-7 days, rinsed twice with 0.1 M cacodylate buffer (pH 7.4), and then postfixed for 1 hour in 1% OsO, in 0.1 M cacodylate buffer. The plates were then rinsed again in buffer and the fixed and processed colla- gen gels were cut in strips with a razor blade. The strips were transferred to glass specimen vials, dehydrated through a graded series of ethanol (25100%) and two propylene oxide changes, and infiltrated with Epon- Araldite resin. Several changes of resin were made over 2 days, as the collagen gels tended to hold the propylene oxide. The collagen strips were flat embedded and cured overnight a t 60C".
RESULTS Morphologic effects of HGF and EGF on hepatocytes maintained in collagen gels
Phase contrast microscopy. Distinct morphologic changes were induced on hepatocytes in cultures sup- plemented with HGF or EGF. In the control cultures (gels with no addition of growth factors), the cells main- tain a rounded contour. Hepatocytes gradually make contact and aggregate to form short cords by days 3 and 4 (Fig. 1A-C). Hepatocytes exposed to HGF developed many sharp and long processes a t day 2 in culture. By day 3, these processes were making contact and hepato- cytes began to form cord-like structures. These cord- like structures became more prominent by day 4 (Fig.

HGF AND EGF EFFECTS ON HEPATOCYTES IN COLLAGEN GELS 445
Fig. 1. A-C: Hepatocytes from control cultures at days 1 , 2 , 3 , corre- spondingly. Hepatocytes maintain a rounded contour and form cords and plates by day 3. X 100. D-F Hepatocytes from cultures exposed to HGF at days 1,2,3, correspondingly. Cells form long pointed processes
by 24 hours. These processes gradually make contact as the cells aggregate to make larger cord shaped structures at day 2. These struc- tures coalesce even further and become stabilized by day 3. Cells still maintain processes in the free surfaces even beyond day 3.
1D-F). Hepatocytes in the cords maintained their pro- cesses for several days after cord formation if they were not directly adjoined by other hepatocytes. Hepatocytes exposed to EGF also developed processes by day 2 but
fewer than hepatocytes exposed to HGF. The cords in EGF supplemented cultures also tended to form by day 4. The above changes were observed in both rat and human hepatocytes.

MICHALOPOULOS ET AL. 446
TEM. Hepatocytes in control cultures maintained normal organelle and membrane structure until at least day 6 (Figs. 2A,B, 3A,B). In contrast, in HGF supplemented cultures the hepatocyte morphology showed distinct changes (Figs. 2C7D7 3C,D). Similar changes were also observed in the EGF supplemented rat hepatocyte cultures, but to a lesser extent (Fig. 2E,F). The untreated hepatocytes in culture formed canalieular complexes both within themselves and be- tween adjacent cells (Figs. 2A,B7 3A,B). The treated cells exhibited fewer and less extensive canaliculi, or none at all (Figs. 2C-F, 3C,D). The formation of junc- tional complexes between the treated cells was also decreased as compared with the controls. Treated cells, however, exhibited more coated pit per vesicle activity than that observed in untreated cells. We also observed increases in the amount per density of cytoplasmic fila- ments in the treated cells; some HGF-treated cells had very dense filament accumulations (see Fig. 2D). The amount of glycogen stored in the treated cells was less than that in untreated cells after the same residence in culture. The reduced glycogen stores observed in the treated cells were segregated into distinct areas of the cytoplasm; a subpopulation of the HGF-treated cells lacked glycogen. The formation of cytoplasmic pro- cesses was increased in the cells exposed to E G F more so in those treated with HGF. The typical round, even nuclear profile of hepatocytes was modified following exposure to either EGF or HGF. Nuclei in EGF-treated cells were somewhat ovoid, but relatively regular. The nuclei of HGF-treated cells varied in nuclear shape from the typical round shape to elongate with smooth or irregular edges.
The changes observed in the human hepatocytes treated with HGF were not as reproducible from one preparation to the next as those seen in the rat hepato- cytes. Overall, however, the same morphologic modifi- cations are present.
All cultures regardless of the presence of mitogens were completely viable and in a state of good morpho- logic preservation at day 6. This was the time at which these cultures became unresponsive to mitogens (see below). The evidence for the viability of the cultures is shown by the degree of structural preservation and or- ganization at day 6. We also examined HGF-treated and untreated cells after 9 days in culture, at which time the cells were still morphologically intact.
Induction of DNA synthesis Addition of HGF or EGF to the medium resulted in
DNA synthesis which lasted up until day 6 of cultures. A dose-response relationship for HGF, EGF, and DNA synthesis is shown in Figure 4. The doses of HGF and EGF required for stimulation of maximal DNA synthe- sis are higher than those usually employed for studies on rigid substrates. The effect of density of hepatocytes on DNA synthesis was also studied (Fig. 5). In contrast to previously described effects with hepatocytes on col- lagen-coated plates (Michalopoulos et al., 19821, no sig- nificant inhibition of DNA synthesis was seen even in cultures with up to 50,000 cells/cm2. A slight decrease was seen in cultures with densities of 100,000/cm2. A density of 25,000/cmZ was used for the subsequent stud- ies. The responsiveness of hepatocytes at different
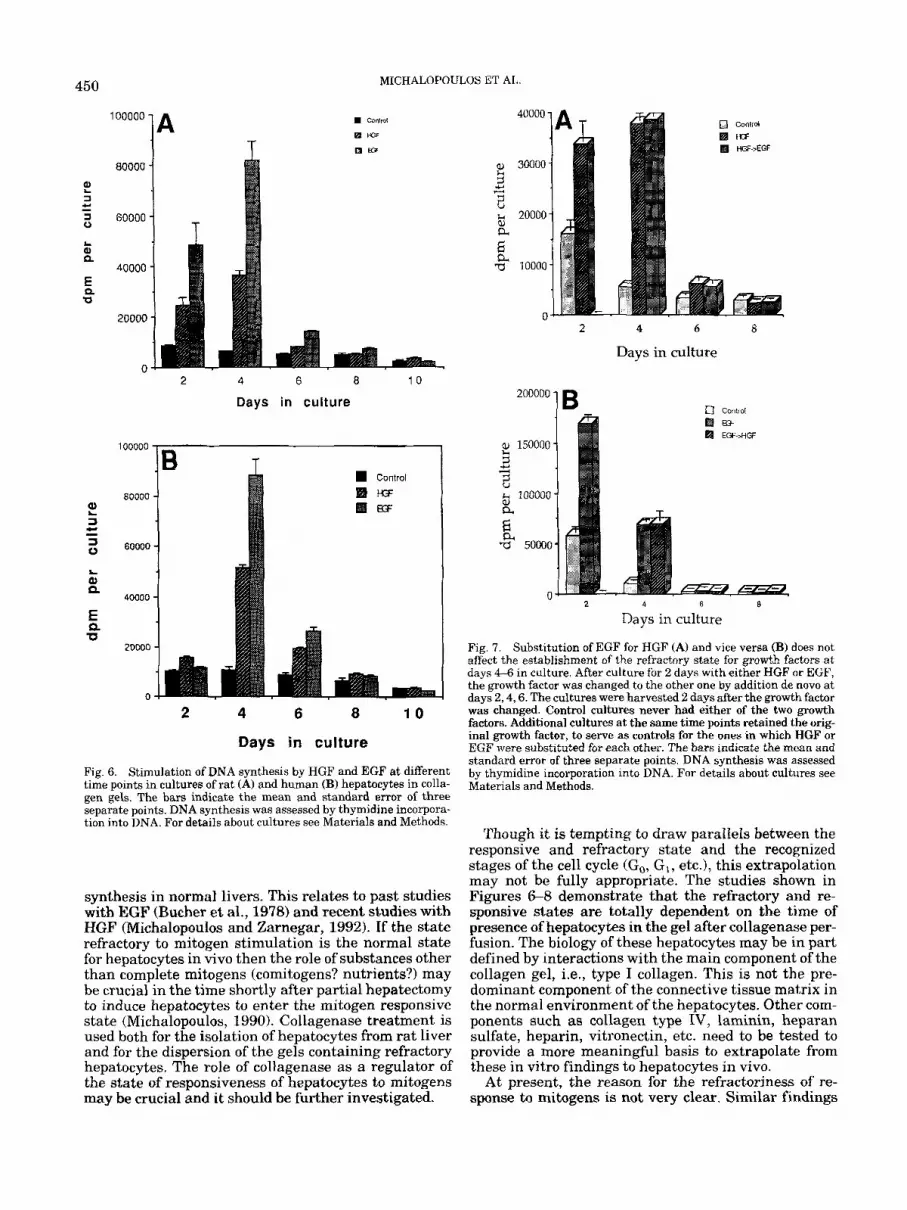
times after initiation of cultures to mitogens was also examined. The results are shown in Figure 6. Contin- ual exposure of rat hepatocytes (Fig. 6A) to either HGF (50 ng/rnl) or EGF (25 ng/ml) stimulated DNA synthe- sis lasting until day 4. No significant responses were seen after day 6, despite the excellent preservation of hepatocytes as seen with electron microscopy (see above). Similar results were obtained with human he- patocytes (Fig. 6B).
To further explore the reasons leading to cessation of responsiveness to mitogens, the growth factors em- ployed were changed from HGF to EGF or the reverse, at different times (days 2, 4) in culture. As shown in Figure 7, regardless of the type of mitogen substitution or the time of substitution, hepatocytes in collagen gels ceased to respond to mitogens by day 6.
In all experiments described in this study, mitogens were added shortly after hepatocytes were isolated and placed in culture. From this design, it was not clear whether the time of entry into the state in which hepa- tocytes become refractory to mitogens was dependent on the time of the addition of mitogens or the time of isolation of hepatocytes. If the initiation of the refrac- tory state was dependent on the time of addition of mitogens and independent of the time of hepatocyte isolation, then hepatocytes would become refractory to the mitogen at a given point after addition of the mito- gen, independent of the time of initiation of the cul- tures. To resolve this dilemma, HGF was added for the first time in cultures at 2,4,6, and 8 days. Each of these culture groups was followed by two 48 hour thymidine pulses. Samples of cultures were harvested at 2 and 4 days after addition of the mitogen. The results are shown in Figure 8. Regardless of the day in culture in which the mitogen was added for the first time, hepato- cytes responded the same way as if mitogen had been added immediately after initiation of the culture. A maximal response was seen a t days 2-4, with no fur- ther response to mitogen after culture day 6. These results show that cessation of hepatocyte proliferation is timed not in relation to the first exposure to the mitogen but in relation to the time after initiation of the culture.
The refractory state of hepatocytes to mitogens did not appear to be irreversible, however. Collagen gels from control cultures a t day 6 were digested with colla- genase. Hepatocytes were isolated and replated on col- lagen-coated plates. The replated hepatocytes were ex- posed to mitogens HGF and EGF. Cultures from the same preparations of hepatocytes were also kept in in- tact collagen gels.
As shown in Figure 9, replated hepatocytes re- sponded immediately to HGF or EGF whereas hepato- cytes from identical cultures maintained in collagen gels remained unresponsive to the same mitogens.
DISCUSSION The present paper describes the responses to mito-
gens HGF and EGF of hepatocytes maintained in colla- gen gels. The findings reveal new information about the effects of these mitogens on hepatocytes in a system where morphogenic and motogenic effects can be ob- served. They also reveal new information, however, rel- evant to control of hepatocyte growth.

HGF AND EGF EFFECTS ON HEPATOCYTES IN COLLAGEN GELS 447
Fig. 2. Cultured rat hepatocytes. A: Untreated, 9 days in culture. The cells possess the standard features of hepatocytes (circular nu- cleus, fairly homogeneous cytoplasm, glycogen throughout the cyto- plasm, etc.). The large canalicular complex and junctional complexes are evident. Bar = 2.0 pm. B: Untreated, 4 days in culture. A higher magnification of a n area adjacent to a canalicular complex. Bar = 0.5 pm. C: HGF-treated, 6 days in culture. Glycogen stores are reduced. The cells do not form the canalicular complexes seen in the untreated cells, although there may be interdigitations. Junctional complexes are reduced both in number and extent. Bar = 2.0 wm. D. HGF-
treated, 4 days in culture. Cytoplasmic filaments and microtubules are more pronounced than in untreated cells. Bar = 0.5 pm. E: EGF- treated, 6 days in culture. Canalicular and junctional complexes are reduced as compared to those observed in the untreated cultures. Bar = 2.0 pm. F EGF-treated, 4 days in culture. Cytoplasmic fila- ments are more pronounced than in controls, but are not as developed as are those in HGF-treated cultures. C, canaliculus; F, cytoplasmic filaments; G1, glycogen; JC, junctional complex; MT, microtubule; ar- rowhead, interdigitations. Bar = 0.2 pm.

448 MICHALOPOULOS ET AL.
Fig, 3. Cultured human hepatocytes, 5 days in culture. A Un- treated. The nucleus presents the standard hepatocyte profile; as do the cytoplasmic elements (profuse glycogen and mitochondria, well- developed Golgi and rough endoplasmic reticulum). Bar = 2.0 p n . B: Untreated. A large canalicular complex has formed between two adja- cent cells. Bar = 2.0 pn. C: HGF-treated. Glycogen stores are reduced and localized; the cell shape is irregular. Bar = 2.0 pm. D HGF-
treated. Increased amounts of cytoplasmic filaments are evident. There are no junctional complexes where two adjacent cells meet. Canalicular complexes are absent, although the cells do interdigitate at certain points (arrowheads). Bar = 1.0 pm. C, canaliculus; F, cyto- plasmic filaments; G, Golgi; G1, glycogen; JC, junctional complex; arrowheads, interdigitations.
The induction of long processes of hepatocytes by HGF probably relates to the motogenic and morpho- genic effects described by HGF in other systems. It is not entirely clear whether the processes formed were instrumental in the formation of the cords seen at day 4 in culture. Conceivably hepatocytes may be aligning themselves in cords after contacts established by the long processes. EGF also induced processes, though of a shorter type. Both mitogens induced formation of the cords by day 4. Hepatocytes arranged in cords in the presence of HGF continued to have short processes even after formation of the cords. The findings demonstrate that EGF in addition to HGF may induce motogenic and morphogenic effects on hepatocytes. Though HGF
is recognized as a powerful motogen and morphogen (Michalopoulos and Zarnegar, 1992; Bussolino et al., 1992; Uehara and Kitamura, 1992), the data shown here demonstrate that other growth factors can also have similar effects on suitable cell targets. The pheno- type of the cells at day 4-6 in culture also differed between control and mitogen-treated cultures.
The concentrations of mitogens used for these studies are much higher than what is reported as the optimal concentrations in plastic or collagen-coated plates. This may reflect sequestration of the growth factors by the type I collagen, requiring larger concentrations to achieve the same effective concentration of free growth factor in the medium. Alternatively, the difference may
HGF AND EGF EFFECTS ON HEF 'ATOCYTES IN COLLAGEN GELS 449
w Control
T 5 HGF20
H f f i F Y )
0 Control 6oooo 1
HGF100
E =I
=I 0
= 200000
=I 0
9)
z E 100000 P Q L 30000
Q 0 E 20000
P 0
10000 0
0
2 4
Days in culture
50000 1
8oooo 1 B E 60000
a 3 c - 0
L 40000 0 P
5 u 20000
0 2 4
Days in culture
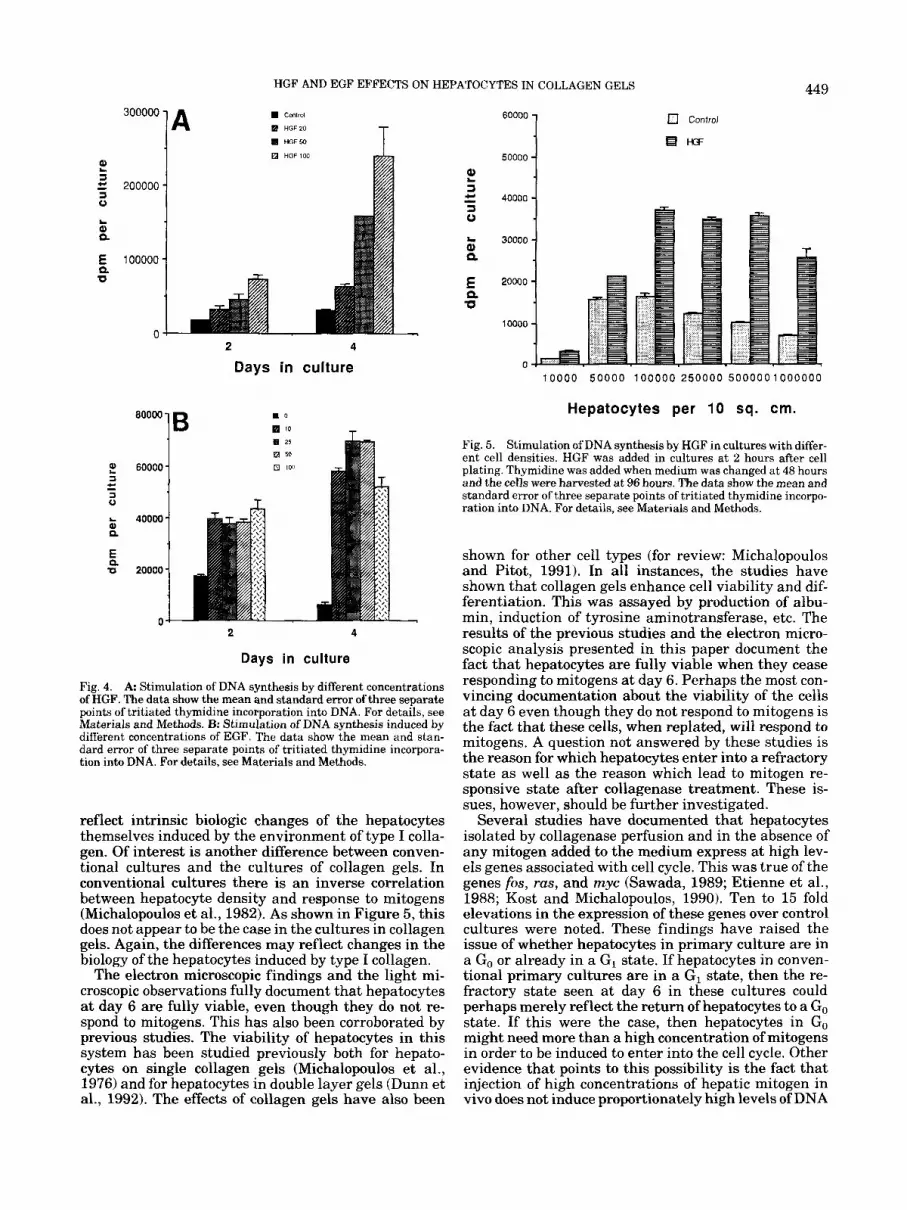
Fig. 4. A Stimulation of DNA synthesis by different concentrations of HGF. The data show the mean and standard error of three separate points of tritiated thymidine incorporation into DNA. For details, see Materials and Methods. B: Stimulation of DNA synthesis induced by different concentrations of EGF. The data show the mean and stan- dard error of three separate points of tritiated thymidine incorpora- tion into DNA. For details, see Materials and Methods.
reflect intrinsic biologic changes of the hepatocytes themselves induced by the environment of type I colla- gen. Of interest is another difference between conven- tional cultures and the cultures of collagen gels. In conventional cultures there is an inverse correlation between hepatocyte density and response to mitogens (Michalopoulos et al., 1982). As shown in Figure 5, this does not appear to be the case in the cultures in collagen gels. Again, the differences may reflect changes in the biology of the hepatocytes induced by type I collagen.
The electron microscopic findings and the light mi- croscopic observations fully document that hepatocytes a t day 6 are fully viable, even though they do not re- spond to mitogens. This has also been corroborated by previous studies. The viability of hepatocytes in this system has been studied previously both for hepato- cytes on single collagen gels (Michalopoulos et al., 1976) and for hepatocytes in double layer gels (Dunn et al., 1992). The effects of collagen gels have also been
2 3 = 40000
10000 50000 100000 250000 5000001000000
Hepatocytes per 10 sq. cm.
Fig. 5. Stimulation of DNA synthesis by HGF in cultures with differ- ent cell densities. HGF was added in cultures a t 2 hours after cell plating. Thymidine was added when medium was changed at 48 hours and the cells were harvested a t 96 hours. The data show the mean and standard error of three separate points of tritiated thymidine incorpo- ration into DNA. For details, see Materials and Methods.
shown for other cell types (for review: Michalopoulos and Pitot, 1991). In all instances, the studies have shown that collagen gels enhance cell viability and dif- ferentiation. This was assayed by production of albu- min, induction of tyrosine aminotransferase, etc. The results of the previous studies and the electron micro- scopic analysis presented in this paper document the fact that hepatocytes are fully viable when they cease responding to mitogens at day 6. Perhaps the most con- vincing documentation about the viability of the cells at day 6 even though they do not respond to mitogens is the fact that these cells, when replated, will respond to mitogens. A question not answered by these studies is the reason for which hepatocytes enter into a refractory state as well as the reason which lead to mitogen re- sponsive state after collagenase treatment. These is- sues, however, should be further investigated.
Several studies have documented that hepatocytes isolated by collagenase perfusion and in the absence of any mitogen added to the medium express at high lev- els genes associated with cell cycle. This was true of the genes fos, ras, and myc (Sawada, 1989; Etienne et al., 1988; Kost and Michalopoulos, 1990). Ten to 15 fold elevations in the expression of these genes over control cultures were noted. These findings have raised the issue of whether hepatocytes in primary culture are in a Go or already in a G, state. If hepatocytes in conven- tional primary cultures are in a GI state, then the re- fractory state seen at day 6 in these cultures could perhaps merely reflect the return of hepatocytes to a Go state. If this were the case, then hepatocytes in Go might need more than a high concentration of mitogens in order to be induced to enter into the cell cycle. Other evidence that points to this possibility is the fact that injection of high concentrations of hepatic mitogen in vivo does not induce proportionately high levels of DNA
450 MICHALOPOULOS ET AL.
looooo 1 A T 00000
60000
40000
20000
0
100000
80000
60000
40000
20000
0
2 4 6 0 1 0
Days in culture
0 Control
KF
2 4 6 8 1 0
Days in culture
Fig. 6. Stimulation of DNA synthesis by HGF and EGF at different time points in cultures of rat (A) and human (B) hepatocytes in colla- gen gels. The bars indicate the mean and standard error of three separate points. DNA synthesis was assessed by thymidine incorpora- tion into DNA. For details about cultures see Materials and Methods.
synthesis in norma1 livers. This relates to past studies with EGF (Bucher et al., 1978) and recent studies with HGF (Michalopoulos and Zarnegar, 1992). If the state refractory to mitogen stimulation is the normal state for hepatocytes in vivo then the role of substances other than complete mitogens (comitogens? nutrients?) may be crucial in the time shortly after partial hepatectomy to induce hepatocytes to enter the mitogen responsive state (Michalopoulos, 1990). Collagenase treatment is used both for the isolation of hepatocytes from rat liver and for the dispersion of the gels containing refractory hepatocytes. The role of collagenase as a regulator of the state of responsiveness of hepatocytes to mitogens may be crucial and it should be further investigated.
2 4 6 8
Days in culture
200000
8 150000 s + 4
2 K E
k 100000
$4 % 5000C
c
B
2 4 6 8
Days in culture
Fig. 7. Substitution of EGF for HGF (A) and vice versa (B) does not affect the establishment of the refractory state for growth factors at days 4-6 in culture. After culture for 2 days with either HGF or EGF, the growth factor was changed to the other one by addition de novo at days 2,4,6. The cultures were. harvested 2 days after the growth factor was changed. Control cultures never had either of the two growth factors. Additional cultures at the same time points retained the orig- inal growth factor, to serve as controls for the ones in which HGF or EGF were substituted for each other. The bars indicate the mean and standard error of three separate points. DNA synthesis was assessed by thymidine incorporation into DNA. For details about cultures see Materials and Methods.
Though it is tempting to draw parallels between the responsive and refractory state and the recognized stages of the cell cycle (Go, GI, etc.), this extrapolation may not be fully appropriate. The studies shown in Figures 6-8 demonstrate that the refractory and re- sponsive states are totally dependent on the time of presence of hepatocytes in the gel after collagenase per- fusion. The biology of these hepatocytes may be in part defined by interactions with the main component of the collagen gel, i.e., type I collagen. This is not the pre- dominant component of the connective tissue matrix in the normal environment of the hepatocytes. Other com- ponents such as collagen type IV, laminin, heparan sulfate, heparin, vitronectin, etc. need to be tested to provide a more meaningful basis to extrapolate from these in vitro findings to hepatocytes in vivo.
At present, the reason for the refractoriness of re- sponse to mitogens is not very clear. Similar findings

HGF AND EGF EFFECTS ON HEPATOCYTES IN COLLAGEN GELS 45 1
40000
Replated cells
3 1 T T ?! 30000
2 4 6 8 1 0
100000
n 2 4 6 8 1 0
I .:--*
200000
100000
0 -- -- 2 4 6 8 1 0
HGF C ! 200000
i 100000
2 4 6 8 1 0
Fig. 8. Responsiveness of hepatocytes in collagen gels to HGF does not depend on the time in which HGF was first added. HGF was added for the first time a t days 0 ,2 ,4 ,6 in culture. In each group of cultures the hepatocytes were harvested at 48 and 96 hours after addition of HGF. The abscissa denotes the absolute number of days in which the hepatocytes were present in culture. Day 0 is the day in which the cells were plated. The days in which HGF was added are shown by superim- posed arrows. The presence of HGF is shown by a horizontal line over the bars. The bars indicate the mean and standard error of three separate points. DNA synthesis was assessed by thymidine incorpora- tion into DNA. For details about cultures see Materials and Methods.
,Control, 81 RGF.
have been reported for hepatocytes maintained in 2% DMSO (Kost and Michalopoulos, 1991). In the latter system, hepatocytes are maintained phenotypically stable for prolonged periods of time but they cease re- sponding to mitogens immediately after exposure to 2% DMSO. Removal of DMSO in the presence of HGF or EGF stimulates bursts of DNA synthesis which stop spontaneously 4 days after DMSO removal. If DMSO is added and moved again, DNA synthesis reoccurs al- though at a reduced scale. This also occurs in hepato- cyte cultures phenotypically stabilized with 3 mM phe- nobarbital (Chan et al., 1990). Hepatocytes maintained in EHS gels also do not respond to mitogens (Bucher et al., 1990). This may be due to the mitoinhibitor trans- forming growth factor beta (TGFP), a component of the
1 c - 3 0
$ 20000
5 P
ZI 10000
0
Cells in intact gels
Fig. 9. Hepatocytes from collagen gels a t day 6 in culture were treated with collagenase, dispersed, and replated into collagen-coated dry plates as described in Materials and Methods. Addition of either HGF or EGF elicited mitogenic response. Hepatocytes from cultures derived from the same rats and maintained in gels without collage- nase dispersion were exposed to the same growth factors a t the time when the sister cultures were dispersed. The two categories are de- noted as “Replated cells” and “Cells in intact gels,” respectively. The bars indicate the mean and standard error of three separate points. DNA synthesis was assessed by thymidine incorporation into DNA. For details about cultures see Materials and Methods. a , control, m, HGF; El, EGF.
EHS gel. The refractory state may be induced by the synthesis of collagenase susceptible connective tissue proteins. In that regard, the already shown require- ment of proline for hepatocytes to enter in to DNA synthesis in response to EGF may be relevant.
The findings of this paper need to be extended into further analysis on the role of other connective tissue proteins in inducing the responsive and refractory states as well as modifying the mitogenic, motogenic, and morphogenic effects of HGF and other growth fac- tors. These studies may define the overall role of con- nective tissue proteins in the regulation of the cell cycle and define the parameters that allow cells to enter into DNA synthesis in response to mitogens.
ACKNOWLEDGMENTS This work was supported by N.I.H. grants CA43632,
CA35373, and CA30241.
LITERATURE CITED Bissell, D.M., Arenson, D.M., Maher, J.J., and Roll, F.J. (1987) Sup-
port of cultured hepatocytes by a laminin-rich gel. Evidence for a functionally significant subendothelial matrix in normal rat liver. J. Clin. Invest., 79:801-812.
Bucher, N.L.R., Patel, U., and Gohen, S. (1978) Hormonal factors and liver growth. Adv. Enzyme Regul., 16t205-213.
Bucher, N.L., Robinson, G.S., and Farmer, S.R. (1990) Effects ofextra- cellular matrix on hepatocyte growth and gene expression: Implica- tions for hepatic regeneration and the repair of liver injury. Semin. Liver Dis., 10:11-19.
Bussolino, F., DiRenzo, M.F., Ziche, M., Bocchietto, E., Olivero, M., Naldini. L.. Gaudino, G.. Tamaenone. L.. Coffer, A,. and Comoelio, P.M. (1992) Hepatocyte growth-factor is a potent angiogenic d t o r which stimulates endothelial cell motility and growth. J. Cell. Biol., 119t629-641.
Chan, K., Kost, D.P., and Michalopoulos, G.K. (1990) Multiple sequen- tial Deriods of DNA svnthesis and auiescence in Drimarv henatocvte cu1t;res maintained“ on the DMSO-EGF onioff prot&ol. ‘J. Ckll. Physiol., 141 584-590.
DiPersio, C.M., Jackson, D.A., and Zaret, K.S. (1991) The extracellu-

452 MICHALOPOl LJLOS ET AL
lar matrix coordinately modulates liver transcription factors and hepatocyte morphology. Mol. Cell Biol., IIt4405-4414.
Dunn, J.C., Tompkins, R.G., and Yarmush, M.L. (1992) Hepatocytes in collanen sandwich Evidence for transcriptional and translational regulagon. J. Cell. Biol., 116:1043-1053.
Etienne, P.L., Baffet, G., Desvergne, B., Boisnard-Rissel, M., Glaise, D.. and Guguen-Guillouzo. C. (1988) Transient expression of c-fos and constagt expression of c-myc in freshly isolated and cultured normal adult rat hepatocytes. Oncogene Res., 3t255262.
Isom, H.C., Secott, T., Georgoff, I., Woodworth, C., and Mummaw, J. (1985) Maintenance of differentiated rat hepatocytes in primary culture. Proc. Natl. Acad. Sci. USA, 82:32523256.
Jefferson, D.M., Clayton, D.F., Darnell, J.E. J r , and Reid, L.M. (1984) Posttranscriptional modulation of gene expression in cultured rat hepatocytes. Mol. Cell Biol., 4t1929-1934.
Kost, D.P., and Michalopoulos, G.K. (1990) Effect of epidermal growth factor on the expression of protooncogenes c-myc and c-Ha-ras in short-term primary hepatocyte culture. J . Cell. Physiol., 144t122- 127.
Kost, D.P., and Michalopoulos, G.K. (1991) Effect of 2% dimethyl sulfoxide on the mitogenic properties of epidermal growth factor and hepatocyte growth factor in primary hepatocyte culture. J. Cell. Physiol., 147:27&280.
Matsumoto, K., Hashimoto, K., Yoshikawa, K., and Nakamura, T. (1991) Marked stimulation of growth and motility of human kerati- nocytes by hepatocyte growth factor. Exp. Cell. Res., 196t114-120.
Michalopoulos, G., Cianciulli, H.D., Novotny, A.R., Kligerman, A.D.,
Strom, S.C., and Jirtle, R.L. (1982) Liver regeneration studies with rat hepatocytes in primary culture. Cancer Res., 42:4673-4682.
Michalopoulos, G., and Pitot, H.C. (1975) Primary culture of paren- chymal liver cells on collagen membranes: Morphological and bio- chemical observations. Exp. Cell Res., 94t70-78.
Michalopoulos, G., and Pitot, H.C. (1991) Citation Classic, Current Contents, International Scientific Institute, 34:ll.
Michalopoulos, G., Sattler, C.A., Sattler, G.L., and Pitot, H.C. (1976) Cytochrome P-450 induction by phenobarbital and 3-methylcholan- threne in primary cultures of hepatocytes. Science, 193t907-909.
Michalopoulos, G.K. (1990) Liver regeneration: Molecular mecha- nisms of growth control. FASEB J . , 4:240-249.
Michalopoulos, G.K., and Zarnegar, R. (1992) Hepatocyte growth fac- tor. Hepatology, 15t149-155.
Sawada, N. (1989) Hepatocytes from old rats retain responsiveness of c-myc expression to EGF in primary culture but do not enter S phase. Exp. Cell Res., 181t584-588.
Strom, S.C., Jirtle, R.L., Jones, R.S., Rosenberg, M.R., and Michalo- poulos, G. (1982) Isolation, culture and transplantation of human hepatocytes. J . Natl. Cancer Inst., 68:771-775.
Uehara, Y., and Kitamura, N. (1992) Expression of a human hepato- cyte growth factorkcatter factor cDNA in MDCK epithelial cells influences cell morphology, motility, and anchorage-independent growth. J. Cell Biol., 117:889-894.
Vintermyr, O.K., and Doskeland, S.O. (1987) Cell cycle parameters of adult rat hepatocytes in a defined medium. A note on the timing of nucleolar DNA replication. J Cell. Physiol., 132~12-21.





























