Embed Size (px)
Citation preview

Comparing excitatory backward and forward
conditioning
Raymond C. Chang, Steven Stout, and Ralph R. Miller
State University of New York at Binghamton, New York, USA
Three Pavlovian lick suppression studies with rats were conducted to compare the role of the
conditioning context in excitatory backward and forward conditioning. The experiments
explored the possibility that excitatory backward conditioning, but not forward conditioning, is
mediated by the context. That is, in excitatory backward conditioning, the conditioning context
may function as an excitatory mediator, which supports second-order conditioning of the target
cue. This possibility contrasts with traditional accounts, which suggests that common processes
underlie excitatory backward and forward conditioning. Experiment 1 found that conditioned
responding following backward conditioning was attenuated as a result of posttraining extinction
of the training context, but the same manipulation elevated responding after forward condition-
ing. Experiments 2 and 3 found that posttraining and pretraining associative inflation of the
context (presenting unsignalled USs) increased conditioned responding to the target of a back-
ward conditioning procedure but either had no effect or reduced responding to the target of a
forward conditioning procedure. Thus, excitatory backward and forward conditioning appear to
differ in their dependence on the status of the conditioning context.
In a backward conditioning procedure, the unconditioned stimulus (US) is soon followed by a
conditioned stimulus (CS). Even though the occurrence of backward conditioning has been
frequently reported (e.g., Barlow, 1956; Cole & Miller, 1999; Doslatek & Krasa, 1972, 1973;
Heth, 1976; Keith-Lucas & Guttman, 1975; Moscovitch & LoLordo, 1968; Plotkin & Oakley,
1975; Romaniuk & Williams, 2000; Siegel & Domjam, 1971, 1974; Spooner & Kellogg, 1947;
Williams & Hurlburt, 2000; Williams & Overmier, 1990), we know little about the mecha-
nisms that determine conditioned responding in such procedures. Indeed, most contempo-
rary learning models eschew any detailed consideration of backward conditioning (e.g.,
Mackintosh, 1975; Pearce & Hall, 1980; Rescorla & Wagner, 1972; but see Wagner, 1981).
Correspondence should be addressed to Ralph R. Miller, Department of Psychology, SUNY-Binghamton,
Binghamton, NY 13902-6000, USA. Email: [email protected]
Support for this research was provided by NIMH Grant 33881. The authors would like to thank Jeffrey
Amundson, Francisco Arcediano, Martha Escobar, and Daniel Wheeler for commenting on an earlier version of the
paper. We also thank Doreen Yirenchi for assistance in running the experiments. The experiments were conducted in
partial fulfillment of the requirements for Masters degree at SUNY Binghamton for the first author (RCC). The first
author thanks his committee members, Peter Gerhardstein, Ralph Miller, and Lisa Savage, for their encouragements
and supports in completing this project.
2004 The Experimental Psychology Society
http://www.tandf.co.uk/journals/pp/02724995.html DOI:10.1080/02724990344000015
THE QUARTERLY JOURNAL OF EXPERIMENTAL PSYCHOLOGY, 2004, 57B (1), 1–23
Q1357—QJ02B/19 JG/WTG. 23 Pages, 3 Tables, 3 Figures . READ AS KEYED

Thus, a backward-trained CS that is capable of evoking excitatory conditioned responding is
theoretically problematic (Spetch, Wilkie, & Pinel, 1981).
Even though the underlying mechanisms of backward conditioning have not been agreed
upon, some variables that affect stimulus control by a backward-trained CS have been identi-
fied (e.g., Chang, Blaisdell, & Miller, 2002; Heth, 1976). In a study of human finger with-
drawal, Spooner and Kellogg (1947) found that backward pairings of a CS and US resulted in
the development of conditioned responding to the CS. A critical determinant for the degree of
responding was the number of US–CS pairings. With repeated pairings of the US and CS,
after the first few trials conditioned responding declined in a pattern resembling experimental
extinction following excitatory forward pairings. Later studies suggested that backward
US–CS pairings tend to endow a CS with inhibitory rather than excitatory properties. For
instance, using a Sidman avoidance task with dogs as subjects, Moscovitch and LoLordo
(1968) found that a backward-trained CS suppressed avoidance behaviour more (indicative of
inhibitory potential) than did a CS that was randomly paired with a US. Subsequent studies
confirmed that a backward-trained CS could act as a conditioned inhibitor (e.g., Plotkin &
Oakley, 1975; Siegel & Domjan, 1971; reviewed by LoLordo & Fairless, 1985). However,
these studies did not manipulate trial number, as had Spooner and Kellogg (1947). Heth
(1976) directly examined the effect of trial number in backward conditioning on excitation and
inhibition. Using a bar-press suppression preparation with rats, he found a shift from excita-
tion with 10 backward pairings to inhibition with 160 backward pairings. Other reports fur-
ther confirmed that the properties of a backward-trained CS shifts from excitatory to
inhibitory over trials. These include studies that found excitatory backward conditioning with
a single trial (Ayres, Haddad, & Albert, 1987; Keith-Lucas & Guttman, 1975) and increasing
inhibition with increasing numbers of trials (e.g., Siegel & Domjan, 1974). Our own laboratory
has replicated Heth’s basic relationship (Cole & Miller, 1999). In a conditioned suppression of
drinking preparation with rats, we found that 4 backward pairings endowed a backward-
trained CS with excitatory properties, whereas 96 pairings were sufficient to make the back-
ward-trained CS a conditioned inhibitor.
Another seemingly crucial variable for behavioural control by a backward-trained CS is the
associative status of the conditioning context. In backward conditioning, the onset of a US is
always preceded by the context alone. The lack of a discrete predictor of the US should
encourage the contextual cues that are present during training to become strongly associated
with the US (e.g., LoLordo & Fairless, 1985). Moreover, the context alone overlaps with and
follows each CS presentation, thereby allowing the CS and the context to become strongly
associated. Thus, it is reasonable to suspect that the training context may have the potential to
mediate excitatory behavioural control by the CS by serving as a representational bridge
between it and the US. There is some empirical support for this view. For example, Marlin
(1983) demonstrated that a conditioning context that is paired with a US can support
Pavlovian conditioning of a CS later paired with that context. Marlin (1986) found that sub-
sequent devaluation of that context, but not an equivalent excitatory context, reduced
responding to the CS. Moreover, Droungas and LoLordo (1994) found that a CS explicitly
unpaired with the US within the same conditioning session could become excitatory. They
also demonstrated that this excitation depended on the associative status of the context, as
posttraining extinction of the conditioning context eliminated the excitation.
2 CHANG, STOUT, MILLER

In fact, the trial dependency of backward conditioning and learning about the conditioning
context may be intimately linked. Few backward pairings may not allow good encoding of
the temporal localization of the US within the overall [global] training context, thereby
precluding a discrimination between the pre-US context and post-CS context and making a
backward CS excitatory. In contrast, many pairings may encourage more precise timing of
USs. Provided that some relatively large minimum intertrial (CS–US) interval is scheduled,
then subjects should learn that the [local] context immediately following each CS is devoid of
the US. Once a subject learns that the CS is reliably followed by a “safety period”, the CS
should become a conditioned inhibitor (see Moscovitch & LoLordo, 1968). Interpreted in this
way, backward conditioning is effectively a discrimination problem of the form: A → US,
AX–, in which A represents the training context. Early in training, the CS becomes excitatory
because of its association with the excitatory context (i.e., there is second-order conditioning
to the CS), and later in training the CS becomes a conditioned inhibitor as the organism learns
that it signals a US-free period within the otherwise excitatory context. This progression from
second-order excitation to inhibition with extended training is mirrored in training of the
same form but with a discrete mediating cue in place of the context (Yin, Barnet, & Miller,
1994).
Further consideration suggests that the encoded information underlying an excitatory
backward-trained CS is qualitatively different from that of a forward-trained CS. In forward
conditioning, a CS presumably directly signals that a US will follow CS onset (as opposed to
the backward conditioning procedure in which the CS does not signal that the US will shortly
occur). Thus in forward conditioning the CS is in a position to compete with the context for
predictive status. Consistent with this consideration, prior studies have observed an inverse
relationship between the excitatory status of a forward CS and the context in which it has been
conditioned. For instance, Grahame, Barnet, and Miller (1992) observed enhancement of
excitatory responding to a forward-trained CS after degraded contingency training (i.e.,
unsignalled USs interspersed among the CS–US pairings) when the training context was
massively extinguished after training. This finding suggested that the training context formed
an association with the US, which, in turn, competed with the target CS–US association at
the time of testing. Seemingly, in forward conditioning the context serves as a competing
stimulus—or comparator stimulus, in the terminology of Miller and Matzel’s (1988) account
of this phenomenon—with respect to the CS. Thus, in conventional forward conditioning the
context might be said to negatively mediate the excitatory potential of the CS, whereas
the second-order conditioning hypothesis asserts that excitatory backward conditioning is
positively mediated by the context.
In contrast to the second-order conditioning hypothesis of excitatory backward con-
ditioning is the view that excitatory backward conditioning is the result of first-order
conditioning between a memory trace of the just-presented US and the CS. Recent experi-
ments by Romaniuk and Williams (2000) were consistent with this position. They suggested
that a backward-trained CS might be decomposed into at least two distinct proximal stimuli,
stimulus onset and stimulus termination, and that each proximal stimulus would condition
separately. They assumed, like Wagner (1981), that at onset a US is activated into an excit-
atory state and then, starting at termination, decays into a refractory state. Wagner’s theory
states that a CS that is paired with the US representation when it is in the excitatory state
MEDIATION BY THE CONTEXT 3

becomes a conditioned excitor, whereas a CS that is paired with the US representation when it
is in the refractory state becomes a conditioned inhibitor. In Romaniuk and Williams’s view
(2000), in a backward conditioning preparation, the onset elements of the CS are presented
when the memory traces of the US are still largely in an excitatory state, thus supporting
excitatory conditioning. The termination elements of the CS occur when the US has largely
entered the refractory state, and thus support inhibitory conditioning. This inhibitory con-
ditioning should lag behind the excitatory conditioning because US traces in the refractory
state are assumed to be less salient than those in the excitatory state. Consistent with this
interpretation, they found that the onset of a backward-trained CS was a conditioned excitor,
despite the entire CS passing the summation test for inhibition. Thus, they concluded that
backward conditioning has the same associative structure as forward conditioning, but that a
backward-trained CS is actually two stimuli, CS onset and CS termination, the former
becoming excitatory through first-order conditioning early in training, the latter becoming
inhibitory with more training.
The present experiments directly contrast the first-order conditioning hypothesis and the
second-order conditioning accounts of excitatory backward conditioning by examining the
effect of various manipulations of the training context on forward-trained and backward-
trained excitatory CSs in the same experiments with all parameters and procedures identical
except for the order of the CS and US during training. According to the second-order con-
ditioning hypothesis, because the conditioning context serves as a mediating stimulus for a
backward-trained CS, either posttraining or pretraining manipulation of the associative status
of the context should alter responding to the backward-trained CS. Consistent with this view,
recent unpublished experiments in our laboratory (Chang et al., 2002) found that massive
posttraining extinction of the conditioning context attenuated the excitatory potential of a
backward-trained CS (excitatory as a result of a few pairings with the US). The second-order
conditioning hypothesis of backward conditioning does not speak to forward conditioning,
but other research has often found that there is an inverse relationship between the response
potential of a forward-trained CS and its conditioning context (e.g., Grahame et al., 1992; Yin
et al., 1994; see also the comparator hypothesis of Miller & Matzel, 1988). Thus, we expected
that the same manipulation would have the opposite effect on a backward-trained CS relative
to that which we previously observed with a forward-trained CS. In contrast, the view that
backward conditioning is first-order conditioning of memory traces of the US and the physical
CS suggests that manipulation of the conditioning context before or after training should have
an identical effect on backward- and forward-trained CSs because backward conditioning and
forward conditioning presumably arise from identical associative processes. (Another, albeit
less likely, explanation of backward conditioning in terms of first-order conditioning between
the CS and US is that an association is formed between the CS and the next US. This view also
does not anticipate posttraining extinction of the context to decrease responding.) Therefore,
the basic issue addressed by the current series of experiments was whether the conditioning
context has different mediating functions in excitatory backward conditioning relative to
(excitatory) forward conditioning. Importantly, these experiments did not address why the
mediating function of the context may depend on the temporal arrangement of the CS and
US, only whether it does so.
In Experiment 1, the conditioning context was associatively deflated (i.e., extinguished)
after conditioning. If the context functions as a positive mediator for an excitatory backward-
4 CHANG, STOUT, MILLER

trained CS and as a negative mediator for a forward-trained CS, massive posttraining context
extinction should attenuate excitatory responding to the backward-trained CS, whereas the
same manipulation should increase excitatory responding to the forward-trained CS. In
subsequent experiments, the conditioning context was associatively inflated either after
conditioning (Experiment 2) or prior to conditioning (Experiment 3). The second-order
conditioning hypothesis predicts that both posttraining and pretraining associative context
inflation should increment the excitatory potential of a backward CS due to a positive medi-
ating role of the conditioning context. In contrast, the same associative inflation of the context
should degrade the excitatory potential of a forward CS due to the negative mediating role of
the conditioning context.
EXPERIMENT 1Posttraining context deflation
Experiment 1 had two aims. The primary aim was to compare the consequences of post-
training associative deflation (i.e., extinction treatment) of the conditioning context for
behavioural control by backward-trained and forward-trained CSs. As stated, across-
experiment comparisons suggest fundamentally different outcomes for context deflation
following each type of conditioning, as opposed to the view of Williams and colleagues, who
have proposed that forward and backward conditioning share essentially the same associative
structure and thus should respond equally to the same postconditioning treatment. The
second aim of Experiment 1 was to illuminate the theoretical basis of excitatory backward
conditioning and test the previously discussed assumption of the context-mediation hypo-
thesis that an excitatory backward-trained CS receives its excitation through contextual
mediation.
In this experiment, either the training context or an irrelevant context was extinguished
following backward or forward conditioning (see Table 1). Two forward conditions were
included. The first was a perfect analogue to the backward conditioning groups, except for the
reversed temporal order of the CS and US during training. However, stronger excitatory
responding to the CS in the forward-trained groups than in the backward-trained groups was
anticipated. If this were to occur, it would create a difference in baseline responding to the excit-
atory CSs, which would confound assessment of the orthogonal manipulation—that
is, posttraining extinction of the conditioning context. Hence, a second forward-training condi-
tion was included. This condition used as a US a footshock with its intensity reduced to a low
level such that forward training with it was expected to yield stimulus control approximately
equal to that of the backward CS paired with the “full”-intensity US. Obviously, this forward
condition confounded US intensity, whereas the first forward condition confounded baseline
behaviour. However, if the two conditions responded equivalently to the posttraining context
deflation manipulation, it would suggest that neither potential confound was of consequence.
Method
Subjects
The subjects were 36 experimentally naive male (192–260 g) and 36 female (190–245 g) Sprague-
Dawley-descended rats bred in our colony. They were individually housed in wire-mesh cages in a
MEDIATION BY THE CONTEXT 5

vivarium maintained on a 16-hr light/8-hr dark cycle. Training was conducted approximately midway
through the light phase of the cycle. A progressive water-deprivation schedule was imposed over the
week prior to the beginning of the experiment, until water availability was limited to 20 mins per day. All
the animals were handled three times per week for 30 s from time of weaning to the initiation of the study.
Subjects were randomly assigned to one of six groups (n = 12) counterbalanced for sex: Bk–Ext,
Bk–NoExt, Fd–Ext, Fd–NoExt, LS–Fd-Ext, and LS–Fd-NoExt, with Bk = backward-trained, Fd =
forward-trained, LS-Fd = forward-trained with low footshock intensity, Ext = posttraining extinction
of the context, and NoExt = no extinction of the context.
Apparatus
Three types of enclosures were used. A total of 12 of each of two types (enclosures R and V) served as
Contexts I and II, counterbalanced for physical identity within groups. Enclosure R was a clear Plexiglas
chamber in the shape of a rectangular box 22.75 × 8.25 × 13.0 cm (l × w × h), with a floor constructed of
0.48-cm-diameter rods 1.5 cm centre-to-centre, connected by NE-2 neon bulbs, which allowed
constant-current footshock to be delivered by means of a high-voltage AC circuit in series with a 1.0-MΩresistor. Each copy of enclosure R had its own environmental isolation chest. Enclosure R was dimly illu-
minated by a 2-Watt (nominal at 120 VAC) bulb driven at 60 VAC mounted on an inside wall of the envi-
ronmental isolation chest approximately 30 cm from the animal enclosure. Background noise, primarily
from a ventilation fan, was 75 dB (C-scale).
Enclosure V was a 25.5-cm-long box in the shape of a vertical truncated V (28 cm high, 21 cm wide at
the top, 5.25 cm wide at the bottom). Each copy of enclosure V had its own environmental isolation chest.
The floor and sides were constructed of sheet metal. The ceiling was clear Plexiglas. The floor consisted
of two parallel metal plates each 2 cm wide, with a 1.25-cm gap between them. The two metal plates were
connected by an NE-2 neon bulb, and, as was the case with enclosure R, footshock could be delivered
through the floor by means of a high-voltage AC circuit in series with a 1.0-MΩ resistor. Enclosure V was
dimly illuminated by a 7-watt (nominal at 120 VAC) bulb driven at 60 VAC mounted on an inside wall of
the environmental isolation chest approximately 30 cm from the animal enclosure, with the light
entering the animal enclosure being primarily that reflected from the roof of the environmental chest.
Due to differences in the opaqueness of the walls of the enclosures, this level of illumination roughly
matched that of enclosure R. Background noise, primarily from a ventilation fan, was 78 dB (C-Scale).
A third context (III) was used for testing in order to avoid the confounds of testing in contexts (a) to
which subjects had previously had differing amounts of exposure due to the extinction phase of treat-
ment, and (b) which might have been more excitatory in the backward condition than in the forward
condition because the CS could better compete with the context in the forward condition. Context III
was a different instance of chamber R, with significant alterations, including the addition of a lick tube, a
Plexiglass floor plate, an odour cue, and the absence of the house light. The odour cue was produced by
two drops of methyl salicylate (a mint odour [98% by weight]) on the top surface of a wooden cube, which
was then placed inside the sound-attenuating environmental isolation chest and outside the animal
chamber. Context III was equipped with a water-filled lick tube, which extended about 1 cm into a cylin-
drical niche (axis perpendicular to a chamber end-wall) 4.5 cm in diameter, left–right centred with its
bottom 1.75 cm above the floor of the apparatus, and 5.0 cm deep. There was a photobeam detector 1 cm
in front of the lick tube, which sensed when a subject had its snout inserted in the niche.
Context I was used in Phases 1 (training) and 2A. Context II was only used in Phase 2B. Context III
was only used for the acclimation, reacclimation, and testing phases. A 45-Ω speaker mounted on the
interior of each environmental chest could deliver a click train (6/s) approximately 8 dB (C-Scale) above
the ambient background, which served as CS X. The stimulus duration in training was 30 s. Two
different intensities of footshock US were used. The high-intensity footshock (US) was a 1.0-mA, 1-s
footshock, whereas the low-intensity footshock (US) was a 0.65-mA, 1-s footshock (see Table 1).
6 CHANG, STOUT, MILLER

Procedure (see Table 1)
Acclimation. On Day 1, all the groups were acclimated to Context III for 30 min. During this
session, no stimulus occurred. The animals had free access to the water-filled tubes during this phase.
Conditioning (Phase 1). On Day 2, backward and forward conditioning took place in Context I
during a single 15-min session. For all the groups, the first and last 2 min of the session were empty
waiting periods (following Cole & Miller, 1999). Thus, the actual conditioning occurred between
minutes 2 and 13. All the groups received either 4 backward US → X (Bk–Ext and Bk–NoExt) or 4
forward X → US pairings (Fd–Ext, Fd–NoExt, LS–Fd–Ext, and LS–Fd–NoExt). The trial presenta-
tions occurred with short intertrial intervals (ITIs) or 180, 120, and 240 s (mean ITI = 180 s), which
would presumably encourage the context to become excitatory. Specifically, the trial presentations
began 2, 5.5, 8, and 12.5 min into the 15-min-long session. The backward (Bk) and forward (Fd) condi-
tions (groups Bk-Ext, Bk–NoExt, Fd–Ext, and Fd–NoExt) were conditioned with the 1.0-mA
footshock, whereas the low-shock-forward (LS-Fd) conditions (groups LS–Fd–Ext and
LS–Fd–NoExt) were conditioned with the 0.65-mA footshock. In the case of backward conditioning,
US termination and the CS onset were simultaneous. In the case of forward conditioning, CS termina-
tion and US onset were simultaneous.
Context extinction (Phases 2A and 2B). On Days 3–10, all the groups received exposure to Context I
alone for 150 min on each of 4 days, alternated with 4 days of 2-min daily exposure to Context II (groups
Bk–Ext, Fd–Ext, and LS–Fd–Ext), or, alternatively, 2-min context-alone exposure to Context I and
150-min context-alone exposure to Context II (groups Bk–NoExt, Fd–NoExt, and LS–Fd–NoExt). All
the groups were exposed to the contexts according to the following schedule by day: I, II, II, I, II, I, I, II.
Reacclimation. On Days 11 and 12, reacclimation took place in Context III (the test context). The
daily session for all groups was 30 min in duration.
Testing CS X. On Day 13, testing with CS X took place in Context III. The first instance of 5 cumu-
lative seconds of licking after placement of the subjects in the test context was recorded. CS X was
presented at that time. Times for the subjects to complete 5 more seconds of licking, now in the presence
MEDIATION BY THE CONTEXT 7
TABLE 1
Design summary of Experiment 1
Expected response
Group Phase 1I Phase 2AII Phase 2BII to CS XIII
Bk–Ext 4 US → X (600 min) (8 min) cr
Bk–NoExt 4 US → X (8 min) (600 min) CR
Fd–Ext 4 X → US (600 min) (8 min) CR
Fd–NoExt 4 X → US (8 min) (600 min) CR
LS–FD–Ext 4 X → US (600 min) (8 min) CR
LS–Fd–NoExt 4 X → US (8 min) (600 min) cr
Note: CS X was a click train; the boldface “US” represents a 1-mA, 1-s footshock, and the lightface “US”
represents a 0.65-mA, 1-s footshock. Number (4) in front of the trial types indicates number of trials; Bk and Fd
denote backward and forward conditioning, respectively; LS denotes low US intensity; Ext indicates long context-
alone exposure to Context I and short exposure to Context II, whereas NoExt denotes short exposure to Context I and
long exposure to Context II. Times (600 or 8) denote the duration of context-alone exposure; the subscripts (I, II, and
III) denote different contexts; cr = weak responding; CR = medium responding; and CR = strong responding (hence,
cr < CR < CR).
of the CS, were recorded as the dependent variable. Notably, with this procedure, all subjects were
drinking at CS onset. Any subject that took more than 60 s to complete the first 5 cumulative seconds of
licks (i.e., preCS scores) was excluded from all subsequent analyses because such preCS scores suggested
an unusually strong fear of the test context. Based on this a priori criterion of rejection, one subject in
group Bk–NoExt was eliminated. Any delay in continued drinking during the stimulus presentation was
assumed to be indicative of an excitatory X–US association. A 15-min ceiling was imposed on all CS
scores. All the drinking latencies (both preCS and during the CS presentation) were converted into log
(base 10) times to better approximate the normality assumption of parametric tests. An α of 0.05 was
selected as the criterion for statistical significance.
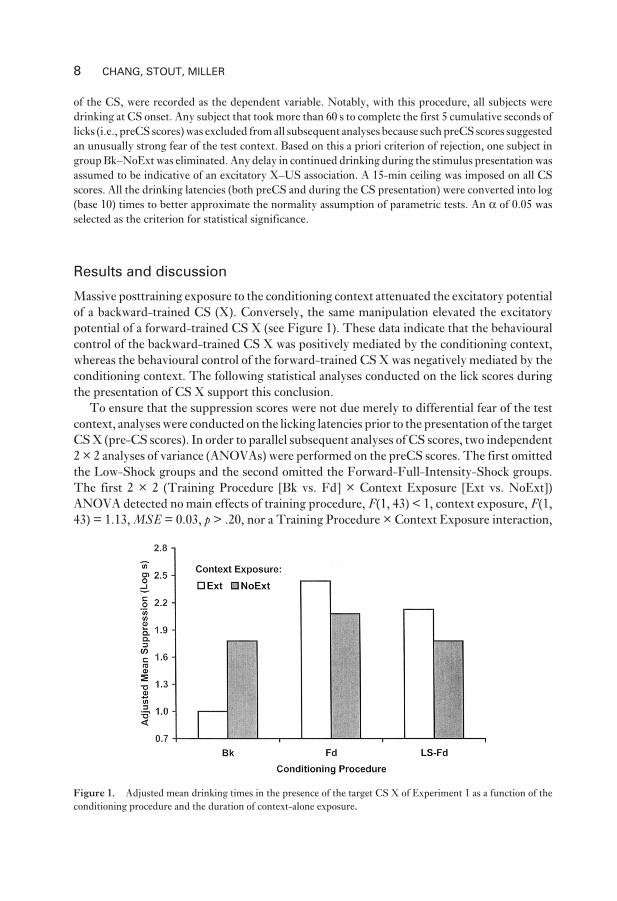
Results and discussion
Massive posttraining exposure to the conditioning context attenuated the excitatory potential
of a backward-trained CS (X). Conversely, the same manipulation elevated the excitatory
potential of a forward-trained CS X (see Figure 1). These data indicate that the behavioural
control of the backward-trained CS X was positively mediated by the conditioning context,
whereas the behavioural control of the forward-trained CS X was negatively mediated by the
conditioning context. The following statistical analyses conducted on the lick scores during
the presentation of CS X support this conclusion.
To ensure that the suppression scores were not due merely to differential fear of the test
context, analyses were conducted on the licking latencies prior to the presentation of the target
CS X (pre-CS scores). In order to parallel subsequent analyses of CS scores, two independent
2 × 2 analyses of variance (ANOVAs) were performed on the preCS scores. The first omitted
the Low-Shock groups and the second omitted the Forward-Full-Intensity-Shock groups.
The first 2 × 2 (Training Procedure [Bk vs. Fd] × Context Exposure [Ext vs. NoExt])
ANOVA detected no main effects of training procedure, F(1, 43) < 1, context exposure, F(1,
43) = 1.13, MSE = 0.03, p > .20, nor a Training Procedure × Context Exposure interaction,
8 CHANG, STOUT, MILLER
Figure 1. Adjusted mean drinking times in the presence of the target CS X of Experiment 1 as a function of the
conditioning procedure and the duration of context-alone exposure.

F(1, 43) < 1. The second 2 × 2 (Training Procedure [Bk vs. LS-Fd] × Context Exposure [Ext
vs. NoExt] ANOVA detected no main effect of context exposure, F(1, 43) = 1.32, MSE =
0.03, p > .20, nor a Training Procedure × Context Exposure interaction, F(1, 43) < 1.
However, a marginal main effect of training procedure was detected, F(1, 43) = 3.97, .05 < p <
.06, suggesting that Context III drinking baseline among the subjects prior to the stimulus
presentation was influenced by shock intensity in Context I. Because of this main effect,
analyses of covariance (ANCOVAs) with the preCS score as the covariate were used in all of
the following analyses on the suppression scores during the presence of CS X. These
ANCOVAs also presumably reduced the influence of individual differences.
The first 2 × 2 (Training Procedure [Bk vs. Fd] × Context Exposure [Ext vs. NoExt])
ANCOVA performed on the suppression scores obtained from the forward “full-intensity”
footshock groups during the presentation of CS X detected a main effect of training proce-
dure, F(1, 42) = 60.44, MSE = 0.15, p < .001, but no main effect of context exposure, F(1, 42)
= 3.29, MSE = 0.15, p = .08. Most important, a Training Procedure × Context Exposure
interaction was detected, F(1, 42) = 25.16, MSE = 0.15, p < .001. Planned contrasts using the
overall error term from the 2 × 2 ANCOVA found that group Bk–Ext suppressed less than did
group Bk–NoExt, F(1, 42) = 22.64, p < .001, indicating that the massive extinction of the
conditioning context attenuated the excitatory properties of the backward-trained CS X.
However, an opposite effect was found in the forward condition, in which group Fd–Ext
suppressed longer than did group Fd–NoExt, F(1, 42) = 5.10, p < .05, indicating that the
deflation of the conditioning context increased the excitatory properties of the forward-
trained CS X. As expected, differences in excitatory baseline between backward conditioning
and forward conditioning, although marginal, were detected, F(1, 42) = 3.80, p = .06 (groups
Bk–NoExt vs. Fd–NoExt; see Figure 1). In order to overcome this confound in baseline, the
second ANCOVA was performed with the forward-low-shock groups substituted for the
“full-intensity”-shock groups.
The first 2 × 2 (Training Procedure [Bk vs. LS–Fd] × Context Exposure [Ext vs. NoExt])
ANCOVA detected a main effect of training procedure, F(1, 42) = 37.99, MSE = 0.10, p <
.001, and a main effect of context exposure, F(1, 42) = 4.42, MSE = 0.10, p < .05. Critically, a
Training Procedure × Context Exposure interaction was also detected, F(1, 42) = 36.94, MSE
= 0.10, p < .001. Planned comparisons confirmed these observations. Group Bk–Ext again
suppressed less than did group Bk–NoExt, F(1, 42) = 32.53, p < .001. However, the opposite
outcome was observed in the low-shock–forward condition, in which group LS–Fd–Ext
suppressed longer than did group LS–FD–NoExt, F(1, 42) = 7.82, p < .01. The crucial obser-
vation of this particular 2 × 2 ANCOVA was that groups Bk–NoExt and LS–NoExt were not
statistically different in suppression, F(1, 42) < 1, which suggests that the differences in excit-
atory baseline between backward and forward conditioning were greatly attenuated by the
lower-intensity shock in the low-shock–forward groups. That is, the effects of the context
manipulations on backward- and forward-trained CSs in the present study could not have
differed due to the differences in excitatory baseline but were due to the training procedures
per se and the differential role of the conditioning context.
The present experiment showed that the context in which the backward trained CS X
was embedded functioned not as a competing cue but, rather, as an excitatory mediating cue.
This replicates previous findings from our laboratory (Chang et al., 2002) and is in accord
with a second-order conditioning interpretation of excitatory backward conditioning. In
MEDIATION BY THE CONTEXT 9

contrast, the context in which the forward-trained CS X was embedded functioned not as a
positive mediating cue but as a competing cue, which replicates the results of Grahame et al.
(1992). In other words, the conditioning context served different functions (negative
[competing] vs. positive mediation) for the forward- and backward-trained CSs respec-
tively. Importantly, unlike prior studies, these observations were made within a single study
that exposed both forward- and backward-trained subjects to an otherwise common set of
procedures.
Several points are worth noting. First, the present findings suggest that the context in
backward conditioning serves a qualitatively different associative function than it does in
forward conditioning, as indicated by the opposite effect that the posttraining context
extinction had on the backward-trained CS and the forward-trained CS. Second, the
present experiment is also one of relatively few instances that provide clear evidence
regarding the competing role of context conditioning on simple excitatory forward condi-
tioning, as indicated by the increase of excitation to the forward-trained CS X after context
extinction. Third, at least in the present preparation, a backward-trained CS that was paired
with the “full-intensity” footshock had a similar excitatory potential to a forward-trained
CS that was paired with a low-intensity footshock. This suggests that a forward-trained CS
paired with a “full-intensity” footshock has a higher excitatory magnitude than does an
otherwise equivalent backward-trained CS, which was paired with the same footshock
during training. This difference in excitatory magnitude potentially reflects excitatory
backward conditioning resulting from a form of second-order conditioning that is mediated
by the context, in contrast with forward conditioning, which results in a direct first-order
association between the CS and the US (albeit one that in select circumstances is down-
modulated by the training context). The present experiment provides evidence against the
first-order conditioning interpretation of excitatory backward conditioning (Romaniuk &
Williams, 2000; Williams & Hurlburt, 2000). According to this account, excitatory back-
ward conditioning and forward conditioning result from identical associative structures
based on simultaneous representation of the CS and US in the excitatory memory state of
Wagner’s SOP model. In this experiment, extinction of the conditioning context attenuated
the excitatory properties of the excitatory backward-trained CS X, whereas this manipula-
tion elevated the excitatory properties of the forward-trained CS X. In the framework of the
SOP model, as well as many other classic conditioning models, posttraining extinction of the
conditioning context should have no effect in either case because it posits changes in associa-
tion between two cues to occur only if at least one of the cues is physically presented.
Notably, posttraining extinction of a (positive) mediator in second-order conditioning does
not always attenuate (or increase) stimulus control (e.g., Rescorla, 1979; Rizley & Rescorla,
1972), but reports explicitly addressing the issue are limited in number and reports in which
it does (e.g., Rashotte, Griffin, & Sisk, 1977; Rescorla, 1982; Robertson, Krane, & Garrud,
1984; Speers, Gillan, & Rescorla, 1980) outnumber those in which it does not. Rescorla
(1979) has suggested US modality as the key determinant of whether posttraining extinction
of the mediating cue will decrease second-order conditioned responding, but this assertion
was based on very limited evidence. In addition to constituting a test of the second-order
conditioning hypothesis of excitatory backward conditioning, the present results can be
taken as further evidence that posttraining extinction of the first-order mediating stimulus
can reduce responding in fear-conditioning preparations. The following studies further
10 CHANG, STOUT, MILLER
investigated other implications of the second-order conditioning and first-order condi-
tioning accounts of excitatory backward conditioning.
EXPERIMENT 2Posttraining context inflation
In Experiment 2, posttraining inflation of the conditioning context (i.e., unsignalled footshock
USs) was implemented (see Table 2). The expectation was that, if the training context posi-
tively mediates responding to a backward-trained CS, posttraining inflation of the context
should augment responding to the backward-trained CS. Based on previous findings (e.g.,
Grahame et al., 1992), an opposite result was expected for a forward-trained CS, where
inflating the conditioning context should cause the context to become highly excitatory,
which, in turn, should reduce responding to the forward-trained CS (this claim has been
observed with other procedures, such as interspersing extra USs among CS–US pairings
[Durlach, 1983; Rescorla, 1968]). In contrast, the first-order conditioning account of excit-
atory backward conditioning predicts that posttraining inflation of the training context should
have had a common effect on both the backward-trained and the forward-trained CSs (the
precise effect depending on the conditioning model).
One potential problem with Experiment 2 is that degradation of behavioural control by a
forward-trained CS through posttraining presentation of the US in the training context has
proven difficult to obtain (e.g., Miller, Hallam, & Grahame, 1990). This difficulty has been
found to be a consequence of a forward-trained CS having acquired biological significance
(i.e., control of behaviour) prior to inflation of the conditioning context. In such cases, CSs
appear to be partly protected from posttraining changes in responding (e.g., Denniston,
Miller, & Matute, 1996; Oberling, Bristol, Matute, & Miller, 2000). In an effort to overcome
the protective effect of biological significance in the present experiment, a large number of
posttraining unsignalled USs were presented in the training context over four short training
MEDIATION BY THE CONTEXT 11
TABLE 2
Design summary of Experiment 2
Expected response
Group Phase 1I Phase 2AII Phase 2BII to CS XIII
Bk–Same 8 US → X 24 US (—) CRr
Bk–Diff 8 US → X (—) 24 US cr
Fd–Same 8 X → US 24 US (—) CR
Fd–Diff 8 X → US (—) 24 US CR
LS–Fd–Same 8 X → US 24 US (—) cr
LS–Fd–Diff 8 X → US (—) 24 US CR
Note: CS X = a click train; the boldface “US” = 1-mA, 0.5-s footshock, and the lightface “US” represents a 0.65-
mA, 0.5-s footshock. (—) denotes context-alone exposure. Numbers (8 and 24) in front of the cues (“X”, “US”, and
“US”) indicate number of trials. “Bk” and “Fd” denote backward and forward conditioning, respectively; “LS”
denotes low US intensity; “Same” and “Diff” indicate the location of unsignalled footshock presentations relative to
Phase 1. The subscripts (I, II, and III) denote different contexts; cr = weak responding; CR = medium responding;
and CR = strong responding (hence, cr < CR < CR).

sessions.1
However, it was still possible that the CS’s biological significance would interfere
with our observing an effect of posttraining associative inflation of the context. However, our
major goal was to contrast backward-trained CSs with forward-trained CSs. Although the
context inflation treatment might not induce a reduction of responding to the forward-trained
CS, it still might positively mediate responding to the backward-trained CS due to the associa-
tive chaining of backward conditioning (i.e., increasing the associative status of the context
might increase the response to the backward-trained CS).
Because detection of an effect of associative inflation of the training context was the central
goal of the present experiment, there was a concern that such an effect could be masked by a
ceiling effect in the backward condition. In Experiment 1, 4 backward pairings had been
administered during training, and they proved sufficient to make the backward-trained CS
into a moderately effective conditioned excitor. If the backward-trained CS acquires strong
excitatory properties, any effects of posttraining inflation of the conditioning context could be
obscured by a ceiling effect. In order to circumvent this problem, in the backward condition
the number of training trials was increased from the 4 in Experiment 1 to 8 in Experiment 2.
Eight backward pairings were chosen because prior data collected under similar circum-
stances by Cole and Miller (1999) suggested that 4 backward pairings made a backward-
trained CS into a conditioned excitor and 16 pairings made the backward-trained CS into a
behaviourally silent CS with neither an excitatory nor inhibitory potential. Because the func-
tion relating a backward-trained CS’s excitatory potential to the number of training trials is
largely exponential (Spooner & Kellogg, 1947), 8 trials were expected to yield low to interme-
diate excitatory responding. The greater degree of training should reduce the excitation
obtained in the backward condition relative to that of Experiment 1. With the exception of this
change, which was implemented in an additional day of Phase 1 training, the intertrial inter-
vals and session duration in this phase were identical to Experiment 1.
Method
Subjects and apparatus
The subjects were 36 experimentally naive male (260–460 g) and 36 female (175–350 g) Sprague-
Dawley descended rats bred in our colony. They were housed and maintained as described in Experi-
ment 1. The subjects were randomly assigned to one of six groups (n = 12) counterbalanced for sex:
Bk–Same, Bk–Diff, Fd–Same, Fd–Diff, LS–Fd–Same, and LS–Fd–Diff, with Bk = backward-trained,
Fd = forward trained, and LS–Fd = forward-trained with low intensity shock, Same = posttraining
12 CHANG, STOUT, MILLER
1Prior to Experiment 2, a pilot study was conducted to illuminate the effect of trial density on the context inflation
treatment. When the trial density was extremely high (mean intertrial interval of 20 s between shocks, session dura-
tion 150 s), the behavioural results matched the predictions of the second-order conditioning hypothesis. However,
there were problems with these pilot data. First, the mean suppression scores among the groups were very high, ap-
proaching the 15-min ceiling for the test trials. Though the direction of differences was suggestive, the unusually high
suppression could have been the result of generalized fear of the entire situation that the subjects acquired during the
high-density unsignalled footshock exposures in Phase 2. Second, the data were positively skewed, even after a log
transformation, such that use of parametric tests for the data analysis was questionable. To partially remedy these
problems, in Experiment 2 the intertrial intervals in Phase 2 were increased from 20 s to 60 s, the duration of the US
was changed from 1 s to 0.5 s, CS intensity was reduced from 8 dB to 6 dB, and the number of Phase 1 trials was
increased from 4 to 8.

inflation in the conditioning context, and Diff = posttraining inflation in the nontraining context. The
apparatus was identical to that used in Experiment 1. However, the parameters of the stimuli used were
changed for the reasons described in the Footnote 1. The intensity of the click train (X) was reduced from
8 dB to 6 dB. Furthermore, the duration of the footshock US was changed from 1 s to 0.5 s.
Procedure (see Table 2)
Acclimation. On Day 1, all the groups were acclimated to Context III for 30 min, as in Experiment 1,
Backward and forward conditioning (Phase 1). On Days 2 and 3, the conditioning pairings took place
in Context I. The session was 15 min in duration. The first and last 2 minutes of the sessions were devoid
of nominal stimulus events, as in Experiment 1. Over the 2 days, all the groups either received 8 back-
ward pairings (US → X; groups Bk–Same and Bk–Diff) or 8 forward pairings (X → US; groups
Fd–Same, Fd–Diff, LS–Fd–Same; and LS–Fd–Diff) with a mean intertrial interval of 180 s. Animals in
the backward condition (groups Bk–Same and Bk–Diff) and forward condition (groups Fd–Same and
Fd–Diff) received 8 backward (US → CS) pairings or 8 forward (CS → US) pairings with the “full-
intensity” footshock US of 1.0 mA. Animals in the low-shock–forward condition (groups LS–Fd–Same
and LS–Fd–Diff) received 8 forward pairings with the low-intensity footshock US of 0.65 mA. The
rationale for the low-shock-intensity condition was the same as in Experiment 1.
Posttraining context inflation (Phases 2A and 2B). Groups Bk–Same, Fd–Same, and LS–Fd–Same
received 6-daily unsignalled US presentations in Context I (the conditioning context) on Days 4, 7, 9,
and 10 (Phase 2A). They also experienced context-only exposure in Context II (the non-training
context) on Days 5, 6, 8, and 11 (Phase 2B). Groups Bk–Diff, Fd–Diff, and LS–Fd–Diff received the
same treatment, except the unsignalled US presentations were administered in Context II during Phase
2B, and they received exposure to Context I alone during Phase 2A. To prevent subjects learning about
“safe” (no-shock) periods in this phase, the intershock intervals (US–US) were highly variable (3–120 s)
with a mean of 60 s, and the session duration was 7 min, 20 s. The individual intershock intervals were
120, 15, 90, 3, 75, 30, and 105 s. Four intershock schedules were used. These intervals were arranged
pseudo-randomly. The restrictions were that no interval values appeared more than ever at the
beginning or the end of a session, and the 3-s ITIs never appeared at the beginning of or the end of a
session. A short session duration was expected to minimize extinction of Context I between USs.
Reacclimation and testing X. On Days 12, 13, and 14, reacclimation took place in Context III. An
extra reacclimation day was added relative to Experiment 1 because of the large number of footshock USs
that the subjects received in Phase 2. The session duration for all groups was 30 min. On Day 15, testing
with CS X took place in Context III. The testing procedures and subject rejection criterion were
identical to Experiment 1. Based on excessive preCS scores (> 60 s), two subjects from group Bk–Diff
were excluded.
Results and discussion
The present experiment found that animals that experienced backward pairings and
posttraining unsignalled USs in the training context suppressed drinking more than did
otherwise equivalent animals that experienced posttraining USs outside the training context.
This suggests that the conditioning context positively mediated the excitatory potential of the
backward-training CS X. However, no associative inflation effect was found in the forward
conditions (see Figure 2). The latter finding was not surprising based on the published data, in
which little or no decrement in response potential resulted from inflation of the training
MEDIATION BY THE CONTEXT 13
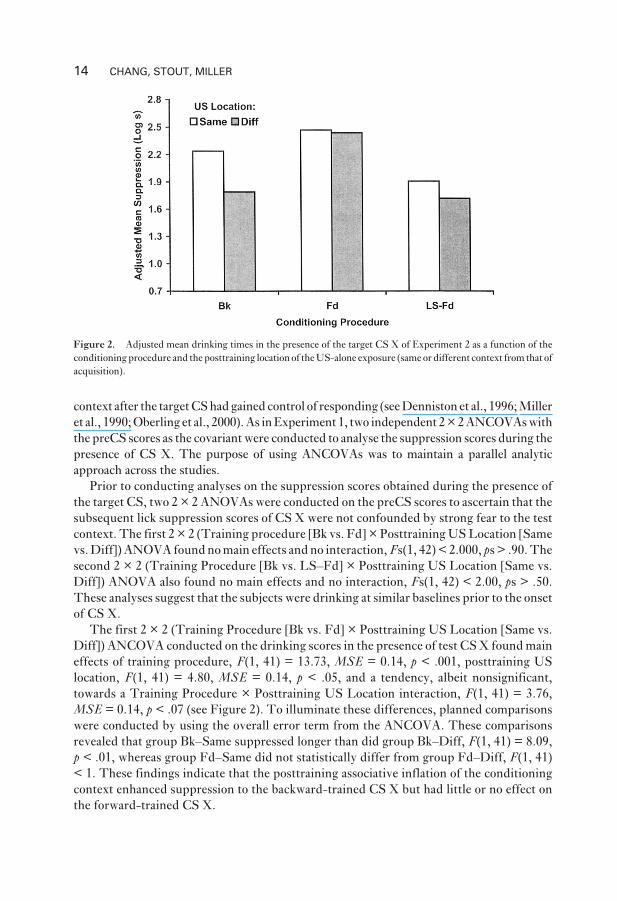
context after the target CS had gained control of responding (see Denniston et al., 1996; Miller
et al., 1990; Oberling et al., 2000). As in Experiment 1, two independent 2 × 2 ANCOVAs with
the preCS scores as the covariant were conducted to analyse the suppression scores during the
presence of CS X. The purpose of using ANCOVAs was to maintain a parallel analytic
approach across the studies.
Prior to conducting analyses on the suppression scores obtained during the presence of
the target CS, two 2 × 2 ANOVAs were conducted on the preCS scores to ascertain that the
subsequent lick suppression scores of CS X were not confounded by strong fear to the test
context. The first 2 × 2 (Training procedure [Bk vs. Fd] × Posttraining US Location [Same
vs. Diff]) ANOVA found no main effects and no interaction, Fs(1, 42) < 2.000, ps > .90. The
second 2 × 2 (Training Procedure [Bk vs. LS–Fd] × Posttraining US Location [Same vs.
Diff]) ANOVA also found no main effects and no interaction, Fs(1, 42) < 2.00, ps > .50.
These analyses suggest that the subjects were drinking at similar baselines prior to the onset
of CS X.
The first 2 × 2 (Training Procedure [Bk vs. Fd] × Posttraining US Location [Same vs.
Diff]) ANCOVA conducted on the drinking scores in the presence of test CS X found main
effects of training procedure, F(1, 41) = 13.73, MSE = 0.14, p < .001, posttraining US
location, F(1, 41) = 4.80, MSE = 0.14, p < .05, and a tendency, albeit nonsignificant,
towards a Training Procedure × Posttraining US Location interaction, F(1, 41) = 3.76,
MSE = 0.14, p < .07 (see Figure 2). To illuminate these differences, planned comparisons
were conducted by using the overall error term from the ANCOVA. These comparisons
revealed that group Bk–Same suppressed longer than did group Bk–Diff, F(1, 41) = 8.09,
p < .01, whereas group Fd–Same did not statistically differ from group Fd–Diff, F(1, 41)
< 1. These findings indicate that the posttraining associative inflation of the conditioning
context enhanced suppression to the backward-trained CS X but had little or no effect on
the forward-trained CS X.
14 CHANG, STOUT, MILLER
Figure 2. Adjusted mean drinking times in the presence of the target CS X of Experiment 2 as a function of the
conditioning procedure and the posttraining location of the US-alone exposure (same or different context from that of
acquisition).

The second 2 × 2 (Training Procedure [Bk vs. LS–Fd] × Posttraining US Location [Same
vs. Duff]) ANCOVA conducted on the suppression scores of target CS X found a main effect
of posttraining US location, F(1, 41) = 5.59, MSE = 0.22, p < .05, but no main effect of
training procedure, F(1, 41) = 2.01, MSE = 0.22, p > .10, and no Training Procedure ×
Posttraining US Location interaction, F(1, 41) < 1. The lack of Training Procedure ×
Posttraining US Location interaction was seemingly due to an uncommonly high MSE (0.22)
and a slight drop in suppression scores in group LS–Fd–Diff relative to group LS–Fd–Same.
However, this difference was not statistically significant, F(1, 41) = 1.08, p = .30, suggesting
that this apparent decrease was a result of statistical variability. Similarly to the first 2 × 2
ANCOVA, group Bk–Same suppressed longer than did group Bk–Diff, F(1, 41) = 5.09, p <
.05. This indicates that the inflation manipulation had a positive mediating effect on the back-
ward-trained CS X but little or no effect on the forward-trained CS X. Consistent with this
conclusion, no difference was detected between groups LS–Fd–Same and LS–Fd–Diff,
F(1, 41) = 1.08, p < .30. Notably, group Bk–Diff did not differ from group LS–Fd–Diff,
F(1, 41) < 1, indicating that our effort to obtain similar baselines was successful. The observed
differential effect of posttraining unsignalled US in the conditioning context as a function of
backward as opposed to forward conditioning trials is consistent with the second-order condi-
tioning account of backward conditioning but not with the first-order conditioning account of
excitatory backward conditioning.
One obvious concern with the present study is that, partly due to the null effects in the
forward conditions, the predicted Training Procedure × Posttraining US Location interac-
tions were short of statistical significance. Nevertheless, positive mediation was obtained in
backward conditioning, but little or no mediation effect was found in forward conditioning.
Given this concern, Experiment 3 examined the effects of a related manipulation on backward
and forward conditioning—namely the effect of inflating the conditioning context prior to
target training (i.e., US preexposure).
EXPERIMENT 3Pretraining context inflation
Experiments 1 and 2 provided support for the second-order conditioning hypothesis, but
not for the first-order conditioning account of excitatory backwards conditioning.
Specifically, the posttraining inflation treatment in Experiment 2 increased the excitatory
potential of the backward-trained CS X but had little or no effect on the forward-trained CS
X. However, the absence of an effect on forward conditioning might have reflected insensi-
tivity of forward conditioning to the inflation treatment due either to inappropriate parame-
ters or to protection derived from the biological significance of the CS. Experiment 3 was
conducted with the desire to obtain a clear increase or decrease in stimulus control in
forward conditioning as a result of associative inflation of the training context (Matzel,
Brown, & Miller, 1987; Randich, 1981). Toward this end, the inflation treatment used in
Experiment 3 was administered prior to conditioning. Thus, the subjects received
pretraining US-alone exposure in Phase 1 and conditioning pairings (backward or forward)
in Phase 2 (see Table 3). Pretraining inflation of the context has been shown to disrupt excit-
atory forward conditioning (e.g., Randich & LoLordo, 1979), and this experiment assessed
the influence of this manipulation on excitatory backward conditioning.
MEDIATION BY THE CONTEXT 15
Method
Subjects and apparatus
The subjects were 72 male (260–360 g) and 72 female (191–257 g) Sprague–Dawley-descended
experimentally naïve rats bred in our colony. They were housed and maintained as described in Experi-
ments 1 and 2. The subjects were randomly assigned to one of six groups, counterbalanced for sex (n =
24); Bk–Same, Bk–Diff, Fd–Same, Fd–Diff, LS-Fd–Same, and LS–Fd–Diff, with Bk = backward-
trained, Fd = forward trained, and LS–Fd = forward-trained with low shock intensity, Same = US
preexposure in Content I, and Diff = US preexposure in Context II. The apparatus and stimuli were
identical to those used in Experiments 1 and 2.
Procedure (see Table 3)
Acclimation. On Day 1, all the groups were acclimated to Context III for 30 min, as in Experiments
1 and 2.
US preexposure (Phases 1A and 1B) and conditioning (Phase 2). On Days 2, 5, 7 and 8 groups
Bk–Same, Fd–Same, and LS–Fd–Same received 6 daily unsignalled footshock USs in Context I, and on
Days 3, 4, 6, and 9 they experienced context-only exposure in Context II (i.e., the nontraining context).
Groups Bk–Diff, Fd–Diff, and LS–Fd–Diff received the same treatment, except that they received the
unsignalled USs in Context II on Days 2, 5, 7 and 8 and context-alone exposure in Context I on Days 3, 4,
6, and 9. The intershock intervals (US–US) were identical to those used in Phase 2 of Experiment 2. On
Days 10 and 11, backward conditioning or forward conditioning took place in Context I, as in Phase 1 of
Experiment 2.
Reacclimation and testing CS X. On Days 12, 13, and 14, reacclimation took place in Context III.
Session duration on each day for all groups was 30 min. On Day 15, testing with CS X was conducted in
Context III. The testing procedure and the subject exclusion criterion were identical to those described
in Experiment 1. Based on the subject exclusion criterion, two animals from group Fd–Diff were
excluded from the overall analysis.
16 CHANG, STOUT, MILLER
TABLE 3
Design summary of Experiment 3
Expected response
Group Phase 1AI Phase 1BII Phase 2I to CS XIII
Bk–Same 24 US (—) 8 US → X CR
Bk–Diff (—) 24 US 8 US → X cr
Fd–Same 24 US (—) 8 X → US CR
Fd–Diff (—) 24 US 8 X → US CR
LS–FD–Same 24 US (—) 8 X → US cr
LS–Fd–Diff (—) 24 US 8 X → US CR
Note: CS X = a click train; the boldface “US” = 1-mA, 0.5-s footshock, and the lightface “US” = a 0.65-mA, 0.5-s
footshock. “(—)” denotes context-alone exposure. Number (8 and 24) in front of the cues (“X”, “US”, and “US”)
indicate number of trials. “Bk” and “Fd” denote backward and forward conditioning, respectively; “LS” denotes low
US intensity; “Same” and “Diff” indicate whether the preexposure to the footshock US occurred in or out of the
training context, respectively. The subscripts (I, II, and III) denote different contexts; cr = weak responding; CR =
medium responding; and CR = strong responding (hence, cr < CR < CR).
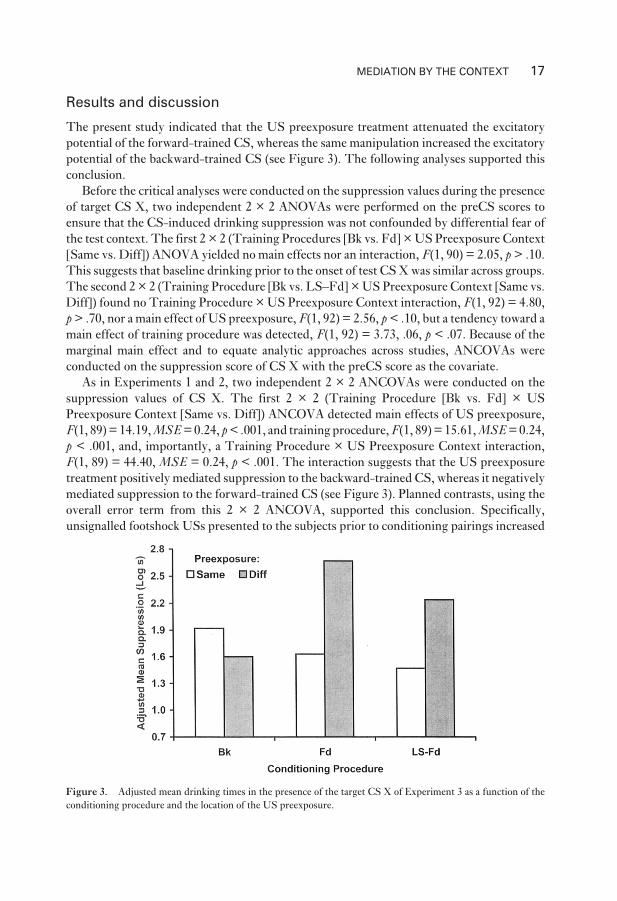
Results and discussion
The present study indicated that the US preexposure treatment attenuated the excitatory
potential of the forward-trained CS, whereas the same manipulation increased the excitatory
potential of the backward-trained CS (see Figure 3). The following analyses supported this
conclusion.
Before the critical analyses were conducted on the suppression values during the presence
of target CS X, two independent 2 × 2 ANOVAs were performed on the preCS scores to
ensure that the CS-induced drinking suppression was not confounded by differential fear of
the test context. The first 2 × 2 (Training Procedures [Bk vs. Fd] × US Preexposure Context
[Same vs. Diff]) ANOVA yielded no main effects nor an interaction, F(1, 90) = 2.05, p > .10.
This suggests that baseline drinking prior to the onset of test CS X was similar across groups.
The second 2 × 2 (Training Procedure [Bk vs. LS–Fd] × US Preexposure Context [Same vs.
Diff]) found no Training Procedure × US Preexposure Context interaction, F(1, 92) = 4.80,
p > .70, nor a main effect of US preexposure, F(1, 92) = 2.56, p < .10, but a tendency toward a
main effect of training procedure was detected, F(1, 92) = 3.73, .06, p < .07. Because of the
marginal main effect and to equate analytic approaches across studies, ANCOVAs were
conducted on the suppression score of CS X with the preCS score as the covariate.
As in Experiments 1 and 2, two independent 2 × 2 ANCOVAs were conducted on the
suppression values of CS X. The first 2 × 2 (Training Procedure [Bk vs. Fd] × US
Preexposure Context [Same vs. Diff]) ANCOVA detected main effects of US preexposure,
F(1, 89) = 14.19, MSE = 0.24, p < .001, and training procedure, F(1, 89) = 15.61, MSE = 0.24,
p < .001, and, importantly, a Training Procedure × US Preexposure Context interaction,
F(1, 89) = 44.40, MSE = 0.24, p < .001. The interaction suggests that the US preexposure
treatment positively mediated suppression to the backward-trained CS, whereas it negatively
mediated suppression to the forward-trained CS (see Figure 3). Planned contrasts, using the
overall error term from this 2 × 2 ANCOVA, supported this conclusion. Specifically,
unsignalled footshock USs presented to the subjects prior to conditioning pairings increased
MEDIATION BY THE CONTEXT 17
Figure 3. Adjusted mean drinking times in the presence of the target CS X of Experiment 3 as a function of the
conditioning procedure and the location of the US preexposure.

the excitatory potential of the backward-trained CS, F(1, 89) = 4.10, p < .05 (Groups
Bk–Same vs. Bk–Diff), whereas the same treatment attenuated the excitatory potential of the
forward-trained CS, F(1, 89) = 53.32, p < .001 (groups Fd–Same vs. Fd–Diff). These anal-
yses confirm that the US preexposure treatment had an opposite effect on the backward-
trained and forward-trained CSs.
The second 2 × 2 (Training Procedure [Bk vs. LS–Fd] × US Preexposure Context [Same
vs. Diff]) ANCOVA yielded a pattern similar to the first ANCOVA. Specifically, the analysis
found a main effect of US preexposure, F(1, 91) = 6.59, MSE = 0.25, p < .05, and a Training
Procedure × US Preexposure Context interaction, F(1, 91) = 27.62, MSE = 0.25, p < .001,
but no main effect of training procedure, F(1, 91) = 2.02, MSE = 0.25, p > .10. Thus, the
second 2 × 2 ANCOVA also detected the positive mediation effect on the backward-trained
CS X and the negative mediation effect on the forward-trained CS X by the training context as
a result of the US preexposure treatment in Phase 1 (see Figure 3). A series of planned
contrasts obtained similar pairwise differences. Specifically, group Bk–Same suppressed
marginally more than did group Bk–Diff, F(1, 91) = 3.46, p < .06, and group LS–Fd–Same
suppressed less than did group LS–Fd–Diff, F(1, 91) = 30.50, p < .001 (LS–Fd–Same vs.
LS–Fd–Diff). The present experiment was not fully successful in controlling for the ceiling
differences, in that group LS–Fd–diff suppressed more (2.24 log s) than did group Bk-Diff
(1.60 log s). But the control was not a complete failure in that the performance of group
LS–Fd–Diff was closer to that of group Bk–Diff than was group Fd–Diff. However, the
present difference in these control groups does somewhat qualify our comparison of the
effects of context inflation in the two cases as the baselines were slightly different.
The primary objective of Experiment 3 was achieved, in that the US-preexposure treat-
ment in the present study positively mediated excitatory conditional responding to the back-
ward-trained CS, whereas the same treatment negatively mediated excitatory conditional
responding to the forward-trained CS. These findings are consistent with the view that excit-
atory backward conditioning is a form of second-order conditioning, and they challenge any
account of excitatory backward conditioning that assumes that a common mechanism under-
lies excitatory backward and forward conditioning (e.g., Romaniuk & Williams, 2000;
Williams & Hurlburt, 2000).
GENERAL DISCUSSION
The phenomenon of excitatory backward conditioning has a long history in the study of
learning because of its elusiveness (Barlow, 1956; LoLordo & Fairless, 1985) and apparent
incompatibility with intuitive and formal notions of conditioning. Our recent work sheds
some light on the nature of backward conditioning. We have shown that the conditioning
context in which a backward-trained CS is embedded mediated conditioned responding to a
backward-trained excitatory CS and proposed that excitatory backward conditioning is a form
of second-order conditioning that is mediated by the training context. Others, including
Romaniuk and Williams (2000) and Williams and Hurlburt (2000), have suggested an alterna-
tive account of excitatory backward conditioning. This account assumes that backward and
forward conditioning are a consequence of the same underlying mechanism—specifically,
simultaneous activation of the CS with lingering excitatory memory traces of the US.
18 CHANG, STOUT, MILLER

At issue in the present research was whether excitatory backward conditioning and forward
conditioning stem from the same mechanism as has been assumed historically, or whether
excitatory backward conditioning arises from a mechanism distinctly different from simple
forward conditioning, as we have suggested. Toward differentiating these two views, the
present research manipulated the associative status of the training context to see whether it
had similar consequences on forward and backward conditioned CSs or whether the conse-
quences diverged.
In Experiment 1, the associative status of the training context was devalued after a few
backward or forward pairings. In control subjects lacking the context devaluation treatment,
this type of training resulted in excitation regardless of the order of the CS and US during
training. Rats that experienced few backward pairings in Phase 1 and received extensive expo-
sure to the training context in Phase 2 suppressed less to the excitatory backward CS X at test
than did animals that did not receive the extensive context-alone exposure in Phase 2. In
contrast, animals that experienced few forward pairings in Phase 1 and the extensive exposure
to the training context in Phase 2 suppressed more to the forward excitor X than did animals
that did not receive the extensive context-alone exposure. The reduction in conditioned
responding to the backward-trained CS X after the extinction of the training context indicates
that the excitatory status of the backward-trained CS X was positively correlated with the
excitatory value of the conditioning context. The increase in conditional responding to the
forward-trained CS X after the devaluation of the training context indicates that the excitatory
status of the forward-trained CS X was negatively correlated with the excitatory status of the
conditioning context (Miller & Matzel, 1988). Because the same posttraining manipulation of
the conditioning context produced different effects on conditioned responding to the back-
ward-trained CS X and the forward-trained CS X, the suppression data are more consistent
with the second-order conditioning hypothesis than the first-order conditioning account of
excitatory backward conditioning.
In Experiment 2, the excitatory status of the conditioning context was inflated after
CS–US pairings. The data indicate that associatively inflating the training context after the
conditioned pairings increased conditional responding to the backward-trained CS X but had
little or no impact on the forward-trained excitor X. The first-order conditioning account fails
to explain the results of Experiment 2 because it assumes that the associative inflation treat-
ment of the training will have no effect, or if it had any effect, it would have a similar impact on
responding to the backward-trained and forward-trained excitors. Even though a difference in
consequence for forward- and backward-trained CSs was observed in Experiment 2, the lack
of consequence of the posttraining manipulation on the forward-trained CS might have been
due to the use of insensitive parameters or to protection against the loss of stimulus control
that is seemingly afforded to biologically significant stimuli (Denniston et al., 1996; Miller et
al., 1990; Oberling et al., 2000).
Experiment 3 examined the consequences of pretraining associative inflation of the
training context on backward- and forward-trained CSs. The US-preexposure treatment was
found to increase the conditioned responding to the backward-trained excitory CS, whereas
the same treatment decreased responding induced by subsequent forward CS–US pairings.
These observations are consistent with the view that excitatory backward conditioning results
from the context’s serving as a representational bridge to the US.
MEDIATION BY THE CONTEXT 19

Variants of Wagner’s (1981) SOP model proposed by Dickinson and Burke (1996) and by
Holland (1983) and Hall (1996) enlarge the set of conditions that presumably result in the
formation of excitatory and inhibitory associations. Their predictions are most obvious in the
case of posttraining associative deflation of the context due to Phase 2 context exposure acti-
vating representations of both the CS and US in the refractory memory state. Dickinson and
Burke’s (1996) model predicts increased responding to both the forward- and the backward-
trained CSs, whereas Holland’s (1983) and Hall’s (1996) models predict decreased excitatory
responding to both the forward- and the backward-trained CSs. Notably, neither model
anticipates the divergence in behaviour between forward- and backward-trained CSs that was
observed in Experiment 1. Applying these models to the present Experiments 2 and 3 is more
complex because the simulations depend on both capacities of and decay rates from both
the excitatory and the refractory memory states. However, across parameters, the
Dickinson–Burke (1996) model predicts that forward- and backward-trained CSs will be
altered in the same direction although to different degrees. The Holland (1983) and the Hall
(1996) models reverse the direction, of expected change in stimulus control but also predict
that the change for forward- and for backward-trained CSs will be in the same direction.
Thus, these models are inconsistent with the observed divergence observed with forward- and
backward-trained CSs. Similar problems exist for Miller and Matzel’s (1988) comparator
hypothesis, which can account for the effects of the present context manipulation on forward-
trained CSs but not on backward-trained CSs. The difficulty of these models in accounting for
the present results no doubt stems from a fundamental difficulty presented to conditioning
models by the phenomenon of second-order conditioning itself. In this preparation, a cue that
is explicitly unpaired with a first-order excitor becomes itself excitatory—exactly the condi-
tion that makes that cue a conditioned inhibitor. Contemporary models seem to have largely
ignored this conundrum and have instead chosen to consider either the second-order excita-
tion or the conditioned inhibition, while ignoring the progression from excitation to inhibition
that occurs over training trials.
The second-order conditioning hypothesis also provides an adequate account of prior
studies that involved other manipulations of the training context in backward conditioning,
most notably the experiments by Dolan, Shishimi, and Wagner (1985) and Wagner and Terry
(1975). Specifically, Wagner and Terry, using an eyelid-conditioning preparation with
rabbits, found that the excitatory properties of a backward-trained CS were attenuated if the
US with which the backward-trained CS was paired was itself predicted by a discrete cue.
Similarly, Dolan et al., using a bar-press suppression preparation with rats, observed that a
backward-trained CS passed a test for conditioned excitation only if the US with which it was
paired was unsignalled. Dolan et al. (1985) offered a viable account of this phenomenon in
terms of Wagner’s (1981) SOP model, but, as we have already discussed, this model is not able
to account for the present data. These findings are readily accounted for by the second-order
conditioning hypothesis if we assume that a discrete signal that precedes the US presentation
blocks the excitatory status of the training context. A similar point was made by Silva and
Timberlake’s (2000) experiment. Using an appetitive lever and feeder contact preparation
with rats, Silva and Timberlake found that a backward-trained CS facilitated lever contacts
during food-reinforced conditioning of a compound of the backward-trained CS and the
lever. However, they observed that a forward-trained CS blocked lever contacts during
compound conditioning. Thus, their findings are consistent with the present interpretation.
20 CHANG, STOUT, MILLER

Facilitation of lever contacts by the backward-trained CS seems to have resulted from the
backward-trained CS predicting the excitatory context, which then summated with the lever
→ US association during compound conditioning, thereby increasing the proportion of the
lever contacts. In contrast, the forward-trained CS blocked the lever contacts because the
forward-trained CS and the lever simultaneously predicted the US presentations. The
second-order conditioning hypothesis provides an account for Silva and Timberlake’s (2000)
observations similar to its account for the results of Dolan et al.’s (1985) and Wagner and
Terry’s (1975) studies.
The differences between backward and forward conditioning, as posited by the second-
order conditioning hypothesis, had been foreshadowed by Spooner and Kellogg more than
half a century ago (1947; see also Barlow, 1956). Even though Spooner and Kellogg did not
specify what mechanisms mediate the differences between the two conditioning procedures,
they presented strong evidence that the two procedures diverge in controlling behaviour with
respect to trial numbers. Using a finger-withdrawal preparation with humans as subjects,
Spooner and Kellogg found that robust excitatory responding to backward CSs occurred
within the first 20 trials of their 100 preset trials, and the strength of the conditioned response
decreased after the first 20 trials. Little or no responding was observed after 100 trials. In
contrast, responding to forward CSs continuously increased as trials persisted, until an
asymptotic level of responding was reached within the last 20 trials. More divergence of
behavioural control was observed when Spooner and Kellogg measured the response latency
of finger withdrawal and the shock intensities necessary to maintain an excitatory response. In
the backward condition, they found that the latency of finger-withdrawal responses varied
little over trials and was consistently quicker than in the forward condition, whereas in the
forward condition response latency increased consistently as training continued. With respect
to shock intensities, Spooner and Kellogg observed that backward CSs required a stronger
shock than did forward CSs to maintain consistent responding. Our results provide further
support for the thesis that “Forward and backward conditioning are fundamentally distinct
processes” (Spooner & Kellogg, 1947, p. 334).
REFERENCES
Ayres, J. J. B., Haddad, C., & Albert, M. (1987). One-trial excitatory backward conditioning as assessed by condi-
tioned suppression of licking in rats: Concurrent observation of lick suppression and defensive behaviors. Animal
Learning & Behavior, 15, 212–217.
Barlow, J. A. (1956). Secondary motivation through classical conditioning: A reconsideration of the nature of back-
ward conditioning. Psychological Review, 63, 406–408.
Barnet, R. C., & Miller, R. R. (1996). Second-order excitation mediated by a backward conditioned inhibitor. Journal
of Experimental Psychology: Animal Behavior Processes, 22, 279–296.
Chang, R. C., Blaisdell, A. P., & Miller, R. R. (2002). Exploring backward conditioning. Manuscript submitted for
publication.
Cole, R. P., & Miller, R. (1999). Conditioned excitation and conditioned inhibition acquired through backward
conditioning. Learning and Motivation, 30, 129–156.
Denniston, J. C., Miller, R. R., & Matute, H. (1996). Biological significance as a determinant of cue competition.
Psychological Science, 7, 325–331.
Dickinson, A., & Burke, J. (1996). Within-compound associations mediate the retrospective revaluation of causality.
Quarterly Journal of Experimental Psychology, 49B, 60–80.
MEDIATION BY THE CONTEXT 21

Dolan, J. C., Shishimi, A., & Wagner, A. (1985). The effects of signaling the US in backward conditioning: A shift
from excitatory to inhibitory learning. Animal Learning & Behavior, 13, 209–214.
Doslatek, C., & Krasa, H. (1972). Backward conditioning: One-session vs. ten-session experiment. Activitas Nervosa
Superior, 14, 58–59.
Doslatek, C., & Krasa, H. (1973). The role of unconditioned stimulus intensity in backward conditioning. Activitas
Nervosa Superior, 15, 239–240.
Droungas, A., & LoLordo, V. M. (1994). Evidence for simultaneous excitatory and inhibitory associations in the
explicitly unpaired procedure. Learning and Motivation, 25, 1–25.
Duriach, P. J. (1983). Effect of signaling intertrial unconditioned stimuli in autoshaping. Journal of Experimental
Psychology: Animal Behavior Processes, 9, 374–389.
Grahame, N. J., Barnet, R. C., & Miller, R. R. (1992). Pavlovian conditioning in multiple contexts: Competition
between contexts for comparator status. Animal Learning & Behavior, 20, 329–338.
Hall, G. (1996). Learning about associatively activated stimulus representations: Implications for acquired equiva-
lence and perceptual learning. Animal Learning & Behavior, 24, 233–255.
Heth, C. D. (1976). Simultaneous and backward fear conditioning as a function of number of CS–UCS pairings. Jour-
nal of Experimental Psychology: Animal Behavior Processes, 2, 117–129.
Holland, P. C. (1983). Representation-mediated overshadowing and potentiation of conditioned aversions. Journal of
Experimental Psychology: Animal Behavior Processes, 9, 1–13.
Keith-Lucas, T., & Guttman, N. (1975). Robust single-trial delayed backward conditioning. Journal of Comparative
and Physiological Psychology, 88, 468–476.
LoLordo, V. M., & Fairless, J. L. (1985). Pavlovian conditioned inhibition: The literature since 1969. In R. R. Miller
& N. E. Spear (Eds.), Information processing in animals: Conditioned inhibition (pp. 1–49). Hillsdale, NJ: Lawrence
Erlbaum Associate Inc.
Mackintosh, N. J. (1975). A theory of attention: Variations in the associability of stimuli with reinforcement. Psycho-
logical Review, 82, 276–298.
Marlin, N. A. (1983). Second-order conditioning using the context as S1. Animal Learning & Behavior, 11, 290–294.
Marlin, N. A. (1986). Devaluation of a contextual conditioned stimulus (S1) in second-order conditioning in rats.
American Journal of Psychology, 99, 471–478.
Matzel, L. D., Brown, A. M., & Miller, R. R. (1987). Associative effects of US preexposure: Modulation of condi-
tioned responding by an excitatory training context. Journal of Experimental Psychology: Animal Behavior
Processes, 13, 65–72.
Miller, R. R., Hallam, S. C., & Grahame, N. J. (1990). Inflation of comparator stimuli following CS training. Animal
Learning & Behavior, 18, 434–443.
Miller, R. R., & Matzel, L. D. (1988). The comparator hypothesis: A response rule for the expression of association.
In G. H. Bower (Ed.), The psychology of learning and motivation. Vol. 2 (pp. 51–92). San Diego: Academic Press.
Moscovitch, A., & LoLordo, V. M. (1968). Role of safety in Pavlovian backward fear conditioning procedure. Journal
of Comparative and Physiological Psychology, 66, 673–678.
Oberling, P., Bristol, A. S., Matute, H., & Miller, R. R. (2000). Biological significance attenuates overshadowing,
relative validity, and degraded contingency effects. Animal Learning & Behavior, 28, 172–186.
Pearce, J. M., & Hall, G. (1980). A model for Pavlovian learning: Variations in the effectiveness of conditioned but not
of unconditioned stimuli. Psychological Review, 87, 532–552.
Plotkin, H. C., & Oakley, D. A. (1975). Backward conditioning in the rabbit (Oryctolagus Cuniculus). Journal of
Comparative and Physiological Psychology, 88, 586–590.
Randich, A. (1981). The US preexposure phenomena in the conditioned suppression paradigm: A role for condi-
tioned situational stimuli. Learning and Motivation, 12, 321–341.
Randich, A., & LoLordo, V. M. (1979). Preconditioning exposure to the unconditioned stimulus affects the acquisi-
tion of a conditioned emotional response. Learning and Motivation, 10, 245–277.
Rashotte, M. E., Griffin, R. W., & Sisk, C. L. (1977). Second-order conditioning of the pigeons keypeck. Animal
Learning & Behavior, 5, 25–38.
Rescorla, R. A. (1968). Probability of shock in the presence and absence of CS in fear conditioning. Journal of Compar-
ative and Physiological Psychology, 66, 1–5.
Rescorla, R. A. (1979). Aspects of the reinforcer learned in second-order Pavlovian conditioning. Journal of Experi-
mental Psychology: Animal Behavior Processes, 5, 79–95.
22 CHANG, STOUT, MILLER

Rescorla, R. A. (1982). Simultaneous second-order conditioning produces S-S learning in conditioned suppression.
Journal of Experimental Psychology: Animal Behavior Processes, 8, 23–32.
Rescorla, R. A., & Wagner, A. R. (1972). A theory of Pavlovian conditioning: Variations in the effectiveness of
reinforcement and nonreinforcement. In A. H. Black & W. F. Prokasy (Eds.), Classical conditioning II: Current
research and theory (pp. 64–99). New York: Appleton-Century–Crofts.
Rizley, R. C., & Rescorla, R. A. (1972). Associations in second-order conditioning and sensory preconditioning.
Journal of Comparative and Physiological Psychology, 81, 1–11.
Robertson, D., Krane, R. V., & Garrud, P. (1984). Second-order conditioned taste aversion in the rat: Shared modal-
ity is not sufficient to promote an association between S1 and S2. Animal Learning & Behavior, 12, 316–322.
Romaniuk, C. B., & Williams, D. A. (2000). Conditioning across the duration of a backward conditioned stimulus.
Journal of Experimental Psychology: Animal Behavior Processes, 26, 454–461.
Savastano, H. I., & Miller, R. R. (1998). Time as content in Pavlovian conditioning. Behavioural Processes, 44,
147–162.
Siegel, S., & Domjan, M. (1971). Backward conditioning as an inhibitory procedure. Learning and Motivation, 2,
1–11.
Siegel, S., & Domjan, M. (1974). The inhibitory effect of backward conditioning as a function of the number of back-
ward pairings. Bulletin of the Psychonomic Society, 4, 122–124.
Silva, F. J., & Timberlake, W. (2000). A clarification of the nature of backward excitatory conditioning. Learning and
Motivation, 31, 67–80.
Speers, M. A., Gillan, D. J., & Rescorla, R. A. (1980). Within-compound associations in a variety of compound condi-
tioning procedures. Learning and Motivation, 11, 135–149.
Spetch, M. L., Wilkie, D. M., & Pinel, J. P. J. (1981). Backward conditioning: A reevaluation of the empirical
evidence. Psychological Bulletin, 89, 163–175.
Spooner, A., & Kellogg, W. N. (1947). The backward conditioning curve. American Journal of Psychology, 60,
321–334.
Wagner, A. R. (1981). SOP: A model of automatic memory processing in animal behavior. In N. E. Spear & R. R.
Miller (Eds.), Information processing in animals: Memory mechanisms (pp. 5–47). Hillsdale, NJ: Lawrence Erlbaum
Associates, Inc.
Wagner, A., & Terry, W. S. (1975). Backward conditioning to a CS following an expected vs. a surprising UCS.
Animal Learning & Behavior, 3, 370–374.
Williams, D. A., & Hurlburt, J. L. (2000). Mechanisms of second-order conditioning with a backward conditioned
stimulus. Journal of Experimental Psychology: Animal Behavior Processes, 26, 340–351.
Williams, D. A., & Overmier, J. B. (1990). Nonassociative habituation, US preexposure, and backward inhibitory
conditioning with signaled and unsignalled USs. Animal Learning & Behavior, 18, 35–43.
Yin, H., Barnet, R. C., & Miller, R. R. (1994). Trial spacing and trial distribution effects in Pavlovian conditioning:
Contributions of a comparator mechanism. Journal of Experimental Psychology: Animal Behavior Processes, 20,
123–134.
Original manuscript received 24 June 2002
Accepted revision received 20 January 2003
MEDIATION BY THE CONTEXT 23





























