Embed Size (px)
Citation preview

C. J. O’Shea, M. B. Lynch, T. Sweeney, D. A. Gahan, J. J. Callan and J. V. O’Doherty
composition, and manure odor and ammonia emissions from finishing pigstractoat-based diet on nutrient digestibility, nitrogen utilization, distal gastrointestinal
Comparison of a wheat-based diet supplemented with purified ²-glucans, with an
doi: 10.2527/jas.2010-2882 originally published online Oct 15, 2010; 2011.89:438-447. J Anim Sci
http://jas.fass.org/cgi/content/full/89/2/438the World Wide Web at:
The online version of this article, along with updated information and services, is located on
www.asas.org
at Swets Blackwell Inc on January 25, 2011. jas.fass.orgDownloaded from

ABSTRACT: It was hypothesized that supple-mentation of an oat-extracted mixed-linkage (1→3), (1→4)-β-d-glucan (β-glucans) to a wheat-based diet may beneficially mitigate manure odor and ammonia emissions associated with intensive pig production, without depressing nutrient digestibility as has been observed with oat-based diets. An experiment was conducted to investigate the effect of dietary β-glucan source and the inclusion of an enzyme composite con-taining β-glucanase on energy and nutrient digestibili-ty, N utilization, distal gastrointestinal tract (GIT) fer-mentation, and manure emissions from finisher boars. Twenty-eight boars (BW = 74.2 ± 3.6 kg) were assigned to 1 of 4 dietary treatments (n = 7/treatment): 1) an oat-based diet (oat), 2) an oat diet + enzyme compos-ite (OE), 3) a wheat-based diet + purified β-glucans (WG), and 4) a wheat-based diet + purified β-glucans
+ enzyme composite. The wheat-based diets containing purified β-glucans were formulated to contain concen-trations of total β-glucans comparable with the oat-based diet. Consumption of the WG diet resulted in a greater digestibility of GE (P = 0.001) and an increase in the urine:feces N excretion ratio (P = 0.049) com-pared with the oat diet. In the distal GIT, pigs offered the WG diet contained fewer bifidobacteria (P = 0.027) and lactobacilli (P = 0.050) compared with the oat diet. Pigs offered the WG diet had increased manure odor emissions compared with the oat diet (P = 0.023). In conclusion, although supplementing wheat-based di-ets with extracted oat-β-glucan did not reduce nutrient digestibility, there was a negligible effect in beneficially influencing manure emissions from pigs when compared with a conventional oat formulation.
Key words: ammonia, beta-glucan, intestinal fermentation, oats, odor, pig
©2011 American Society of Animal Science. All rights reserved. J. Anim. Sci. 2011. 89:438–447 doi:10.2527/jas.2010-2882
INTRODUCTION
Gaseous emissions from intensive pig production units reduce air quality, inducing behavioral aberrations in local human communities (Schiffman et al., 1995). These pollutants partly reflect the inefficient utilization of dietary CP by the growing pig, resulting in consider-able urinary excretion of N (Carpenter et al., 2004) and the fermentation of undigested protein to malodorous end products (Le et al., 2005). Such processes subse-quently contribute to manure odor and ammonia emis-sions (Mackie et al., 1998; O’Shea et al., 2009).
The production of odorous metabolites may be re-duced through stimulation of carbohydrate fermentation in the distal gastrointestinal tract (GIT), with benefi-cial moderation of odor and ammonia emissions (Garry et al., 2007). Such decreases in gaseous emissions have been attributed to providing a dietary source of mixed-linkage (1→3), (1→4)-β-d-glucan (β-glucans), which may induce a more saccharolytic microbiota (O’Connell et al., 2005; Garry et al., 2007). Previous studies have indicated that oats may present the optimal source of dietary β-glucans for modulating the composition of the distal GIT and thus improve manure emissions from finishing pigs (O’Shea et al., 2010b).
However, the integration of fermentable carbohydrate with the intention of ameliorating manure emissions may be accompanied by impaired nutrient utilization (Lynch et al., 2007; O’Shea et al., 2009). Therefore, it is desirable to impart the beneficial properties of β-glucans to a conventional wheat-based formulation. To validate the role of β-glucans in both extracted and intact form,
Comparison of a wheat-based diet supplemented with purified β-glucans, with an oat-based diet on nutrient digestibility,
nitrogen utilization, distal gastrointestinal tract composition, and manure odor and ammonia emissions from finishing pigs1
C. J. O’Shea, M. B. Lynch, T. Sweeney, D. A. Gahan, J. J. Callan, and J. V. O’Doherty2
School of Agriculture, Food Science and Veterinary Medicine, University College Dublin, Ireland
1 Funding for this research was provided under the National De-velopment Plan, through the Commercialisation Fund administered by Enterprise Ireland (Dublin, Ireland).
2 Corresponding author: [email protected] February 3, 2010.Accepted October 6, 2010.
438
at Swets Blackwell Inc on January 25, 2011. jas.fass.orgDownloaded from

supplementation with an enzyme composite containing β-glucanase may elucidate whether induced intestinal variables are a response to β-glucans or an additional fermentable component in oats.
It was hypothesized that supplementation of a wheat-based diet with a purified β-glucan source may posi-tively influence manure odor and ammonia emissions from finishing pigs comparable with an oat-based diet. An experiment was conducted to investigate the effect of dietary β-glucan source and the inclusion of an en-zyme composite containing β-glucanase on nutrient di-gestibility, N utilization, distal GIT fermentation, and manure emissions from finisher boars.
MATERIALS AND METHODS
All procedures described in this experiment were con-ducted under experimental license from the Irish De-partment of Health in accordance with the Cruelty to Animals Act 1876 and the 1994 European Communi-ties (Amendments of the Cruelty to Animals Act 1876) Regulations.
Experimental Design and Dietary Treatments
The experiment was devised as a complete random-ized block design comprising 4 dietary treatments as fol-lows: 1) oat-based diet (oat), 2) oat diet + enzyme com-posite (OE), 3) wheat-based diet + purified β-glucan (WG), and 4) wheat-based diet + purified β-glucan + enzyme composite (WGE). All experimental diets were formulated to be isocalorific and isonitrogenous, containing similar concentrations of calculated NE (9.3 MJ/kg) and standardized ileal digestible Lys (8.6 g/kg; Sauvant et al., 2004) and offered in meal form. All other ileal digestible AA requirements were met relative to Lys (NRC, 1998). Wheat-based dietary treatments were supplemented with an oat-derived β-glucan source (65%; Cambridge Commodities Ltd., Cambridgeshire, UK) comparable with concentrations of total β-glucans typically observed in oat diets. The enzyme supplement was derived from Penicillium funiculosum (IMI SD 101) and contained endo-1,3 (4)-β-glucanase (EC 3.2.1.6) and endo-1,4-β-xylanase (EC 3.2.1.8; DSM, Belfast, Northern Ireland). The dietary composition and analy-sis are presented in Table 1.
Animals and Management
Twenty-eight finishing boars [Landrace boars × (Large White × Landrace) sows] were blocked based on BW and assigned to 1 of 4 dietary treatments (n = 7). Pigs were electronically tagged and individually of-fered ad libitum access to feed and water by means of a computerized single-space feeding system (Varley et al., 2010) and remained on their respective experimental diets until slaughter. At the end of a 14-d dietary adap-tion period, 16 boars (n = 4) of a uniform BW (74.2 ± 3.6 kg) were selected and transferred to individual me-
tabolism crates. The pigs were allocated an additional 5 d to adapt to the metabolism crates before collections commenced. The collection period was subdivided into 3 parts to facilitate studies on manure ammonia emis-sions (d 1 to 2), manure odor emissions (d 3 to 5), and apparent total tract nutrient digestibility and N bal-ance (d 6 to 10). The pigs were offered ad libitum ac-cess to feed. The quantity of feed consumed and refused was noted. Fresh water was provided with meals in a 1:1 ratio, and fresh water was offered ad libitum be-tween meals. The metabolism crates were located in a temperature-controlled room maintained at 22 ± 1.5°C. After the collection period, the pigs were reunited with their penmates until slaughter.
Ammonia Emissions Study
Four separate collections of total feces and urine were carried out at 12-h intervals during collection d 1 to 2, and samples were stored in sealed containers at 4°C. Af-ter the final collection, the urine and feces were pooled and macerated together according to the original ex-cretion ratio. Maceration was performed to simulate the conditions of a typical manure container, which would be more dynamic due to constant disturbance through addition of fresh excreta. The respective ma-nure sample (2 kg) was placed in a ventilated container in a temperature-controlled room maintained at 20°C. Ammonia emissions from the manure were measured over 240 h in a laboratory scale set-up as described by O’Shea et al. (2009). Briefly, ammonia emitted from manure was collected in 3 impingers through the use of a pump with a ventilation rate calibrated at 2.5 L/min. The first 2 impingers contained 1 mol/L of nitric acid, and the third impinger contained distilled water. The first impinger was sampled at 48, 96, 144, 192, and 240 h, and the second impinger was sampled at 96 h. Samples were collected from all 3 impingers at 240 h. The concentration of ammonia-N contained in each impinger was determined by the microdiffusion tech-nique of Conway (1957). Ammonia emissions (mg) from each manure sample were compared between different dietary treatments using the quantity volatilized from 0 to 240 h per kg of sample. A 2-kg duplicate sample was retained in a separate ventilated container for each 2-kg manure sample used in the ammonia emission study. From the duplicate sample, an aliquot (50 g) was re-tained at 0 and 240 h and frozen at −20°C for subse-quent VFA determination.
Air Sample Collection and Odor Concentration Measurement
Collections of total urine and feces production were carried out at 12-h intervals during collection d 3 to 4 and stored together in an open container below the me-tabolism crate until sampling on d 5. Air samples were used to measure odor concentration (European odor units/m3). Air sampling was performed as described
Purified and intact dietary β-glucans in pigs 439
at Swets Blackwell Inc on January 25, 2011. jas.fass.orgDownloaded from

by Lynch et al. (2008). Samples were contained in 20-L Nalophan bags and analyzed for odor concentration (Ecoma T07 dynamic olfactometer, Ecoma, Honig-see, Germany) as described previously by Hayes et al. (2004).
Apparent Total Tract Energy and Nutrient Digestibility and N Balance Studies
Daily urine production was collected separately be-neath the metabolism crate in a plastic container and recorded. To avoid N volatilization, 20 mL (25%) of sulphuric acid was added to the container daily. As a further preventative, a weak solution (2%) of sulphuric acid was applied to the connecting funnel twice daily. A 50-mL aliquot of urine and 100-g aliquot of feces were retained daily and frozen (−20°C) for subsequent determination of N.
Microbiology Study
Immediately postslaughter, digesta samples (approx-imately 10 ± 1 g) were aseptically recovered from the distal colon of each animal (n = 7) in sterile condi-tions. Digesta samples were stored in sterile containers
(Sarstedt Ltd., Wexford, Ireland), placed in insulated containers with dry ice, and transported to the labora-tory within 2 h. A 1.0 g of sample was removed from each digesta sample, serially diluted (1:10) in 9.0-mL aliquots of maximum recovery diluents (Oxoid Ltd., Hampshire, UK) and spread (0.1 mL aliquots) onto selective agars. Populations of lactobacilli, bifidobac-teria, and Escherichia coli were selectively isolated and enumerated as described by Pierce et al. (2005). Lac-tobacilli and bifidobacteria were selected as bacterial indicators of saccharolytic fermentation (Garry et al., 2007; O’Shea et al., 2010a), whereas E. coli were cho-sen based on their potential association with gastroin-testinal disequilibrium and protein fermentation (Heo et al., 2009). Typical colonies of each bacterium were counted, log-transformed, and are presented per gram of digesta.
pH Measurements
The pH of digesta samples from the distal colon were determined immediately postslaughter. Samples (n = 7) were placed in universal containers, and pH measurements were made using a pH meter (Mettler Toledo MP 220, Mettler-Toledo Inc., Columbus, OH),
Table 1. Composition and analysis of experimental diets (as-fed basis)
Item
Oats Wheat + β-glucans
No enzyme With enzyme No enzyme With enzyme
Ingredient, g/kg Oats 645 645 0 0 Wheat 0 0 681 681 Soybean meal 270 270 270 270 Soy oil 60 60 0 0 Dicalcium phosphate 7.5 7.5 7.5 7.5 Salt 5 5 5.0 5.0 Limestone flour 10 10 10.0 10.0 Minerals and vitamins1 2.5 2.5 2.5 2.5 Enzyme composite2 0 0.05 0 0.05 β-Glucans3 0 0 24 24Analyzed composition, g/kg DM 902.4 908.8 889.2 888.5 CP 183.9 185.0 185.7 186.3 Ash 51.7 52.6 48.6 45.0 NDF 162.7 150.5 93.5 104.0 GE, MJ/kg 17.8 17.7 15.5 15.8 Xylose 26.06 28.02 25.78 26.62 Hemicellulose 65.4 64.1 57.9 59.6 Cellulose 65.7 59.9 22.5 26.5 Lignin 15.8 13.3 6.5 8.9 Total β-glucans 20.5 19.5 22.9 22.5 Soluble β-glucans 11.9 11.5 18.7 18.3 Insoluble β-glucans 8.6 8.0 4.2 4.2 β-Glucanase activity4 30 1,800 40 1,760 β-Xylanase activity5 35 1,350 30 1,300
1Premix provided per kilogram of complete diet: 3 mg of retinol, 0.05 mg of cholecalciferol, 40 mg of α-tocopherol, 25 mg of copper as copper II sulfate, 100 mg of zinc as zinc oxide, 0.3 mg of selenium as sodium selenite, 25 mg of manganese as manganese oxide, and 0.2 mg of iodine as calcium iodate on a calcium sulfate/calcium carbonate carrier.
2DSM, Belfast, Northern Ireland.3Cambridge Commodities Ltd., Cambridgeshire, UK.4β-Glucanase activity is defined as the units of glucose produced per gram of diet.5β-Xylanase activity is defined as the units of xylose produced per gram of diet.
O’Shea et al.440
at Swets Blackwell Inc on January 25, 2011. jas.fass.orgDownloaded from

which was calibrated using certified pH 4 and 7 buf-fer solutions (Sigma-Aldrich Ireland Limited, Wicklow, Ireland). Distilled water was added to all distal colon samples in a 1:1 ratio to facilitate pH measurement. The pH measurements of manure samples at 0 and 240 h were determined similarly.
VFA Profiling of Digesta and Manure Samples
Samples of digesta from the distal colon of individual pigs (n = 7) were recovered for VFA analysis. Total and specific VFA concentrations in the digesta were deter-mined as described previously by Porter and Murray (2001) using a modified method (Pierce et al., 2006b). These short-chain fatty acids were selected due to their ready conversion into the gaseous phase from manure surfaces. Briefly, a sample of 1 g was diluted with dis-tilled water (2.5 × weight of sample) and centrifuged at 1,400 × g for 4 min at 21°C (Sorvall GLC, Block Scientific Inc., Bohemia, NY). One milliliter of the subsequent supernatant and 1 mL of internal standard (0.5 g of 3-methyl-n-valeric acid in 1 L of 0.15 mol/L of oxalic acid) were mixed with 3 mL of distilled wa-ter. After centrifugation to remove the precipitate, the sample was filtered through Whatman 0.45-µm poly-ethersulfone membrane filters (Sigma-Aldrich Ireland Limited, Wicklow, Ireland) into a chromatographic sample vial. A sample of 1 µL was injected into a gas chromatograph (model 3800, Varian, JVA Analytical, Dublin, Ireland) with a 25 m × 0.53 mm i.d. mega-bore column [coating CP-Wax 58 (FFAP) – CB (No. CP7614); Varian, Middelburg, the Netherlands]. The VFA concentrations of manure from the odor study at 0 and 240 h were similarly determined with minor ad-aptations as described previously by O’Connell et al. (2005). Isovaleric acid, isobutyric acid, and valeric acid were designated as odorous VFA because of their as-sociation with protein fermentation and disproportion-ately large contribution to the perception of malodor from pig manure.
Laboratory Analysis of Samples
The proximate analysis of diets and feces for DM (method 934.01) and ash (method 942.05) were carried out according to the AOAC (1995) procedures. The DM of the diets and feces was determined after drying for 24 h at 100°C. The ash content of diets and feces was determined after ignition of a weighed sample in a muffle furnace (Nabertherm, Bremen, Germany) at 500°C for 4 h. The GE of diets and feces samples was determined using an adiabatic bomb calorimeter (Parr Instruments, Moline, IL). The NDF, ADF, and ADL fractions of diets and feces were determined using a fiber extraction unit (Fibertec, Tecator, Hoganas, Swe-den) as described by Van Soest et al. (1991). The N concentration of diets and urine was determined (Leco FP 528, Leco Instruments, Cheshire, UK). The N con-
centration of fresh feces was analyzed by the macro-Kjeldahl technique using a digestion and distillation apparatus (Buchi Labortechnik, Flawil, Switzerland). The dietary concentrations of Lys, Thr, Trp, Met, and Cys were determined by HPLC (Varian Prostar, Santa Clara, CA) as described by Iwaki et al. (1987). The total β-glucan and xylose content of diets were deter-mined colorimetrically after using an enzymatic hydro-lysis assay (Megazyme International Ireland Ltd., Co. Wicklow, Ireland). The insoluble and soluble β-glucan contents were determined as described by McCleary and Glennie-Holmes (1985). The hemicellulose content of diets was estimated as the numerical difference be-tween NDF and ADF. The cellulose content was esti-mated as the numerical difference between ADF and ADL. The lignin content is presented as ADL. The dietary β-glucanase and β-xylanase activity were de-termined as described by Somogyi (1960). The activ-ity of β-glucanase and β-xylanase refers to the units of glucose and xylose, respectively, produced per gram of diet.
Statistical Analysis
Experimental data were analyzed as a complete ran-domized design using the GLM procedure (SAS Inst. Inc., Cary, NC). The statistical model investigated the effect of dietary treatment, and BW was included in the statistical model as a covariate. A contrast state-ment was used to compare 1) oat diet vs. WG diet, 2) oat diet vs. OE diet, and 3) WG diet vs. WGE diet. All data in the tables are presented as least squares means and pooled SEM. The individual pig served as the ex-perimental unit.
RESULTS
The chemical analysis indicated an average CP con-tent of 184.5 and 186.0 g/kg for the oat and wheat-based diets, respectively (Table 1). The average total β-glucan content was 22.7 g/kg for the wheat-based di-ets containing a source of exogenous β-glucans, whereas it was 20.0 g/kg for the oat diets.
Apparent Total Tract Digestibility and N Balance Study
There was no effect of supplementing the oat diet or the WG diet with an enzyme composite contain-ing β-glucanase and β-xylanase on energy and nutri-ent digestibility or N balance (Table 2). Pigs offered the WG diet had increased water intake (P = 0.032), digestibility of DM (P = 0.001), OM (P = 0.001), GE (P = 0.001), and NDF (P = 0.001), and decreased fe-cal excretion (P = 0.001) and DM output (P = 0.001) compared with the oat diet. Offering the WG diet in-creased urine excretion (P = 0.018) and increased the urine:feces excretion ratio (P = 0.001) compared with pigs offered the oat diet. Offering the WG diet increased
Purified and intact dietary β-glucans in pigs 441
at Swets Blackwell Inc on January 25, 2011. jas.fass.orgDownloaded from

urinary N (P = 0.038), the urine:feces N excretion ratio (P = 0.049), and decreased N retention (P = 0.013) and N absorption (N retention/intake; P = 0.022) com-pared with the oat diet. Pigs offered the WGE diet had an increased urine:feces N excretion ratio (P = 0.017) compared with the WG diet.
Colonic Composition
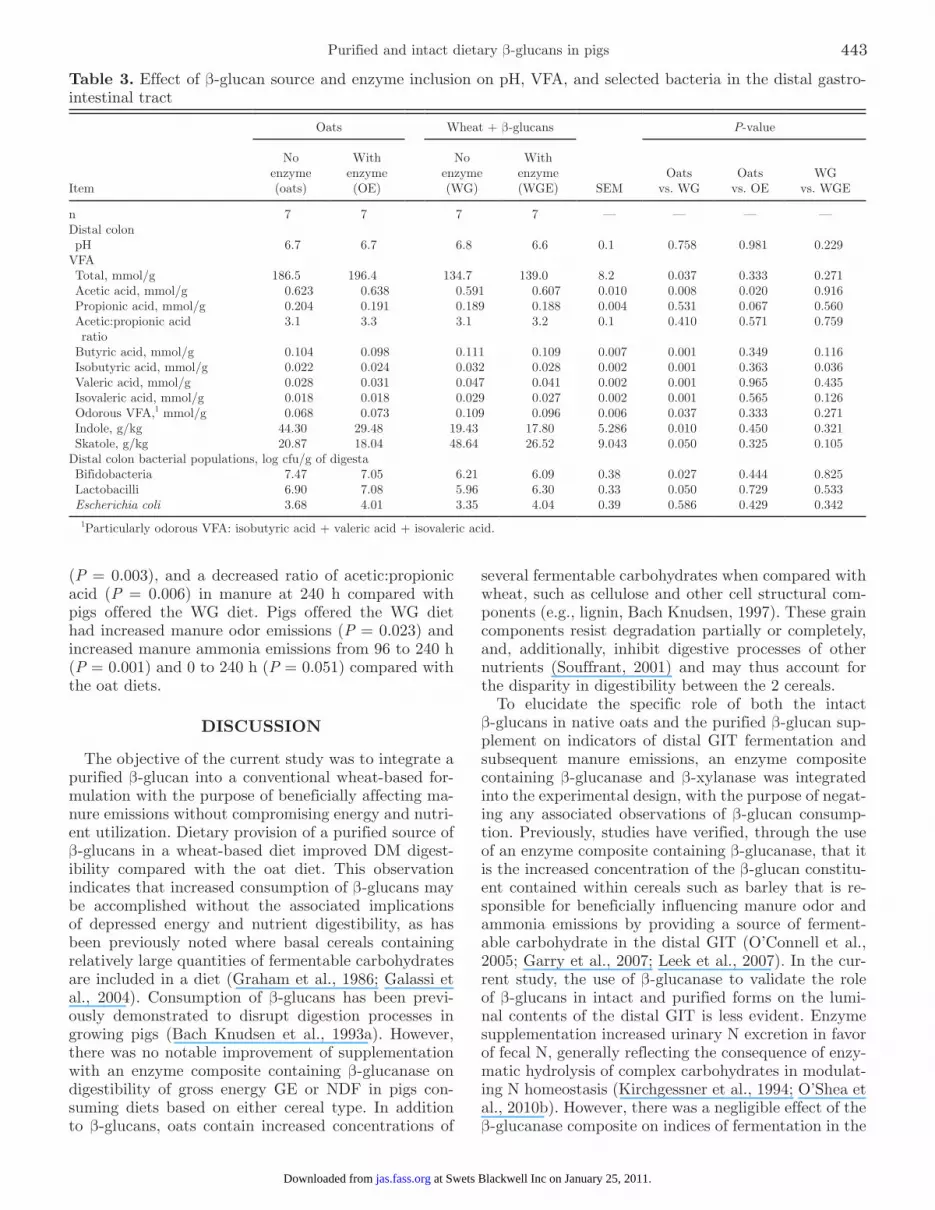
In the colon, pigs offered the WG diet had decreased numbers of bifidobacteria (P = 0.027) and decreased numbers of lactobacilli (P = 0.050) compared with pigs offered the oat diet (Table 3). In the colon, pigs offered the WG diet had decreased concentrations of total VFA (P = 0.037) and molar proportion of acetic acid (P = 0.008) and an increased molar proportion of butyric acid (P = 0.001), isobutyric acid (P = 0.001), valeric acid (P = 0.001), isovaleric acid (P = 0.001), and odor-ous VFA (P = 0.037) compared with offering the oat diets. In the colon, pigs offered the OE diet had an increased molar proportion of acetic acid (P = 0.020) compared with pigs offered the oat diet. In the colon, pigs offered the WGE diet had a decreased molar pro-portion of isobutyric acid (P = 0.036) when compared with the WG diet. In the colon, pigs offered the WG diet had decreased concentrations of indole (P = 0.010)
and increased concentrations of skatole (P = 0.050) compared with the oat diet.
Manure Composition and Emissions
At 0 h, manure from pigs offered the WG diet con-tained increased concentrations of total VFA (P = 0.009), acetic acid (P = 0.005), butyric acid (P = 0.035), valeric acid (P = 0.013), isovaleric acid (P = 0.039), and overall odorous VFA (P = 0.021), and an increased acetic:propionic acid ratio (P = 0.001) com-pared with pigs offered the oat diets (Table 4). Offering the WGE diet resulted in decreased concentrations of total VFA (P = 0.009) and acetic acid (P = 0.005), and a decreased acetic:propionic acid ratio (P = 0.001) in manure compared with the WG diet.
At 240 h, manure from pigs offered the WG diet contained increased concentrations of total VFA (P = 0.013), acetic acid (P = 0.013), propionic acid (P = 0.050), isobutyric acid (P = 0.029), valeric acid (P = 0.006), isovaleric acid (P = 0.007), and overall odorous VFA (P = 0.013), and an increased acetic:propionic acid ratio (P = 0.001) compared with pigs offered the oat diet. Pigs offered the WGE diet had decreased con-centrations of total VFA (P = 0.017), acetic acid (P = 0.018), propionic acid (P = 0.008), and isovaleric acid
Table 2. Effect of dietary β-glucan source and exogenous enzyme supplementation on apparent energy and nutri-ent digestibility
Item
OatsWheat
+ β-glucans
SEM
P-value
No enzyme (oats)
With enzyme (OE)
No enzyme (WG)
With enzyme (WGE)
Oats vs. WG
Oats vs. OE
WG vs. WGE
n 4 4 4 4 — — — —Intake variable DMI, kg/d 2.0 1.9 1.9 1.9 0.0 0.277 0.496 0.959 Water intake, kg/d 5.3 5.3 7.0 5.8 0.5 0.032 0.984 0.120Digestibility coefficient DM 0.792 0.786 0.905 0.893 0.007 0.001 0.544 0.259 OM 0.806 0.798 0.922 0.913 0.006 0.001 0.432 0.372 GE 0.802 0.799 0.900 0.894 0.007 0.001 0.729 0.505 NDF 0.280 0.296 0.585 0.601 0.044 0.001 0.873 0.697 N 0.871 0.888 0.889 0.914 0.009 0.185 0.180 0.077 Ash 0.583 0.602 0.626 0.536 0.029 0.345 0.645 0.055Excretory variable Urine, kg/d 2.1 2.7 4.2 3.0 0.5 0.018 0.433 0.145 Feces, kg/d 1.3 1.3 0.6 0.6 0.1 0.001 0.877 0.751 Urine:feces 1.5 2.5 5.9 5.1 0.5 0.001 0.231 0.329 Total manure output, g/d 3.4 3.9 4.8 3.6 0.5 0.091 0.460 0.150 Fecal DM, % 31.7 33.7 29.2 34.0 1.5 0.289 0.362 0.051 DM output, kg/d 0.406 0.412 0.169 0.201 0.015 0.001 0.762 0.167N utilization N intake, g/d 63.6 62.3 63.1 63.2 1.3 0.811 0.485 0.990 Urinary N, g/d 24.1 25.9 30.6 33.0 1.8 0.038 0.499 0.373 Fecal N, g/d 8.1 7.0 7.1 5.5 0.6 0.252 0.201 0.084 Urinary:fecal N 3.0 3.9 4.4 6.1 0.4 0.049 0.163 0.017 Total N excretion, g/d 32.2 32.9 37.6 38.5 1.8 0.071 0.790 0.747 N retention, g/d 31.4 29.4 25.5 24.7 1.3 0.013 0.310 0.666 N retention/intake 0.500 0.475 0.410 0.391 0.023 0.022 0.437 0.580
O’Shea et al.442
at Swets Blackwell Inc on January 25, 2011. jas.fass.orgDownloaded from
(P = 0.003), and a decreased ratio of acetic:propionic acid (P = 0.006) in manure at 240 h compared with pigs offered the WG diet. Pigs offered the WG diet had increased manure odor emissions (P = 0.023) and increased manure ammonia emissions from 96 to 240 h (P = 0.001) and 0 to 240 h (P = 0.051) compared with the oat diets.
DISCUSSION
The objective of the current study was to integrate a purified β-glucan into a conventional wheat-based for-mulation with the purpose of beneficially affecting ma-nure emissions without compromising energy and nutri-ent utilization. Dietary provision of a purified source of β-glucans in a wheat-based diet improved DM digest-ibility compared with the oat diet. This observation indicates that increased consumption of β-glucans may be accomplished without the associated implications of depressed energy and nutrient digestibility, as has been previously noted where basal cereals containing relatively large quantities of fermentable carbohydrates are included in a diet (Graham et al., 1986; Galassi et al., 2004). Consumption of β-glucans has been previ-ously demonstrated to disrupt digestion processes in growing pigs (Bach Knudsen et al., 1993a). However, there was no notable improvement of supplementation with an enzyme composite containing β-glucanase on digestibility of gross energy GE or NDF in pigs con-suming diets based on either cereal type. In addition to β-glucans, oats contain increased concentrations of
several fermentable carbohydrates when compared with wheat, such as cellulose and other cell structural com-ponents (e.g., lignin, Bach Knudsen, 1997). These grain components resist degradation partially or completely, and, additionally, inhibit digestive processes of other nutrients (Souffrant, 2001) and may thus account for the disparity in digestibility between the 2 cereals.
To elucidate the specific role of both the intact β-glucans in native oats and the purified β-glucan sup-plement on indicators of distal GIT fermentation and subsequent manure emissions, an enzyme composite containing β-glucanase and β-xylanase was integrated into the experimental design, with the purpose of negat-ing any associated observations of β-glucan consump-tion. Previously, studies have verified, through the use of an enzyme composite containing β-glucanase, that it is the increased concentration of the β-glucan constitu-ent contained within cereals such as barley that is re-sponsible for beneficially influencing manure odor and ammonia emissions by providing a source of ferment-able carbohydrate in the distal GIT (O’Connell et al., 2005; Garry et al., 2007; Leek et al., 2007). In the cur-rent study, the use of β-glucanase to validate the role of β-glucans in intact and purified forms on the lumi-nal contents of the distal GIT is less evident. Enzyme supplementation increased urinary N excretion in favor of fecal N, generally reflecting the consequence of enzy-matic hydrolysis of complex carbohydrates in modulat-ing N homeostasis (Kirchgessner et al., 1994; O’Shea et al., 2010b). However, there was a negligible effect of the β-glucanase composite on indices of fermentation in the
Table 3. Effect of β-glucan source and enzyme inclusion on pH, VFA, and selected bacteria in the distal gastro-intestinal tract
Item
Oats Wheat + β-glucans
SEM
P-value
No enzyme (oats)
With enzyme (OE)
No enzyme (WG)
With enzyme (WGE)
Oats vs. WG
Oats vs. OE
WG vs. WGE
n 7 7 7 7 — — — —Distal colon pH 6.7 6.7 6.8 6.6 0.1 0.758 0.981 0.229VFA Total, mmol/g 186.5 196.4 134.7 139.0 8.2 0.037 0.333 0.271 Acetic acid, mmol/g 0.623 0.638 0.591 0.607 0.010 0.008 0.020 0.916 Propionic acid, mmol/g 0.204 0.191 0.189 0.188 0.004 0.531 0.067 0.560 Acetic:propionic acid ratio
3.1 3.3 3.1 3.2 0.1 0.410 0.571 0.759
Butyric acid, mmol/g 0.104 0.098 0.111 0.109 0.007 0.001 0.349 0.116 Isobutyric acid, mmol/g 0.022 0.024 0.032 0.028 0.002 0.001 0.363 0.036 Valeric acid, mmol/g 0.028 0.031 0.047 0.041 0.002 0.001 0.965 0.435 Isovaleric acid, mmol/g 0.018 0.018 0.029 0.027 0.002 0.001 0.565 0.126 Odorous VFA,1 mmol/g 0.068 0.073 0.109 0.096 0.006 0.037 0.333 0.271 Indole, g/kg 44.30 29.48 19.43 17.80 5.286 0.010 0.450 0.321 Skatole, g/kg 20.87 18.04 48.64 26.52 9.043 0.050 0.325 0.105Distal colon bacterial populations, log cfu/g of digesta Bifidobacteria 7.47 7.05 6.21 6.09 0.38 0.027 0.444 0.825 Lactobacilli 6.90 7.08 5.96 6.30 0.33 0.050 0.729 0.533 Escherichia coli 3.68 4.01 3.35 4.04 0.39 0.586 0.429 0.342
1Particularly odorous VFA: isobutyric acid + valeric acid + isovaleric acid.
Purified and intact dietary β-glucans in pigs 443
at Swets Blackwell Inc on January 25, 2011. jas.fass.orgDownloaded from

distal GIT. This likely reflects the mechanistic location of β-glucanase, which may be more proximal along the GIT (Bedford, 2000). As such, any influence on bac-terial metabolite profiles because of partitioning of N from fecal to urinary excretion is likely to be distorted by the conveying of sources of endogenous N, such as desquamated intestinal cells and pancreatic secretions into the distal GIT (Mariscal-Landín et al., 1995).
To facilitate investigations on the effect of dietary treatments on the distal GIT microbiota, bifidobacteria and lactobacilli were suggested as bacterial indicators of normal distal GIT function (O’Doherty et al., 2005; Pierce et al., 2006a; Kiarie et al., 2007) and also because of their association with reduced manure odor (Garry et al., 2007; O’Shea et al., 2010a). The enumeration of E. coli was performed because of its potential asso-ciation with gut disequilibria (Heo et al., 2009), prote-olytic fermentation, and greater manure odor emissions (Zhu, 2000; O’Shea et al., 2010a).
The preferential and ready utilization of oat β-glucan by saccharolytic genera such as lactobacilli and bifi-
dobacteria has been previously demonstrated in vitro (Jonsson and Hemmingsson, 1991), whereas the sub-stitution of basal cereals containing increased β-glucan concentrations for wheat-based diets have similarly promoted these bacterial genera in vivo (O’Connell et al., 2005). In the current study, however, consumption of the wheat-based diets supplemented with purified oat β-glucans resulted in reduced numbers of bifidobac-teria and lactobacilli in the distal GIT compared with the oat diet, signifying a considerable shift in the pro-portions of these typically saccharolytic genera relative to E. coli. This demonstrates that the purified β-glucan source was ineffective in increasing the numbers of lac-tobacilli and bifidobacteria in the distal GIT to quanti-ties comparable with the oat diet. Putative explanations for this may be 2-fold. First, although it has previously been suggested that solubility may not be an exclu-sive determinant of the fate of ingested β-glucan along the GIT (O’Shea et al., 2010b), the extraction process involved in disrupting purified β-glucans from the par-ent cell wall has consequences for solubility (Izydorczyk
Table 4. Effect of β-glucan source and enzyme inclusion on manure composition and ammonia and odor emis-sions
Item
Oats Wheat + β-glucans
SEM
P-value
No enzyme (oats)
With enzyme (OE)
No enzyme (WG)
With enzyme (WGE)
Oats vs. WG
Oats vs. OE
WG vs. WGE
n 4 4 4 4 — — — —Manure (0 h) pH 7.5 8.0 8.2 8.2 0.3 0.076 0.152 0.837 VFA in manure (DM) Total VFA, mmol/g 558.1 436.9 1,188.8 553.1 143.1 0.009 0.561 0.009 Acetic acid, mmol/g 350.9 307.4 886.4 355.7 110.4 0.005 0.785 0.005 Propionic acid, mmol/g 106.3 68.8 138.7 98.0 18.9 0.248 0.186 0.153 Acetic:propionic acid ratio 3.4 4.5 6.3 3.6 0.6 0.001 0.474 0.001 Butyric acid, mmol/g 54.3 26.5 78.5 46.0 10.9 0.035 0.272 0.056 Isobutyric acid, mmol/g 12.5 8.1 21.7 13.6 2.7 0.143 0.096 0.057 Valeric acid, mmol/g 21.2 17.0 39.1 25.9 4.4 0.013 0.514 0.054 Isovaleric acid, mmol/g 12.8 9.1 24.4 13.9 3.5 0.039 0.467 0.057 Odorous VFA,1 mmol/g 46.6 34.2 85.2 53.4 10.3 0.021 0.414 0.050Manure (240 h) pH 8.9 8.8 9.0 9.0 0.1 0.399 0.498 0.864 VFA in manure (DM) Total VFA, mmol/g 943.2 925.3 2,442.7 1,022.2 364.3 0.013 0.973 0.017 Acetic acid, mmol/g 732.1 774.9 2,061.6 808.7 322.3 0.013 0.927 0.018 Propionic acid, mmol/g 90.8 68.5 165.9 56.7 24.3 0.050 0.530 0.008 Acetic:propionic acid ratio 7.8 11.4 13.4 12.2 1.6 0.001 0.416 0.006 Butyric acid, mmol/g 56.2 34.0 79.0 51.8 10.3 0.144 0.156 0.087 Isobutyric acid, mmol/g 18.4 14.7 37.3 28.8 5.4 0.029 0.636 0.285 Valeric acid, mmol/g 36.8 27.8 86.8 68.0 10.5 0.006 0.554 0.228 Isovaleric acid, mmol/g 9.0 5.4 16.0 8.2 1.4 0.007 0.091 0.003 Odorous VFA,1 mmol/g 64.2 47.8 136.2 104.9 17.4 0.013 0.519 0.229Ammonia, mg/g of N intake 0 to 96 h 32.8 32.9 27.4 29.6 5.8 0.527 0.982 0.797 96 to 240 h 38.8 42.4 60.0 57.6 2.9 0.001 0.404 0.579 0 to 240 h 71.6 75.4 87.4 87.2 5.1 0.051 0.609 0.975Odor (72 h) Concentration,2 OuE/mL 2,366 1,993 5,212 3,461 765 0.023 0.737 0.134
1Particularly odorous VFA: isobutyric acid + valeric acid + isovaleric acid.2OuE = European odor units.
O’Shea et al.444
at Swets Blackwell Inc on January 25, 2011. jas.fass.orgDownloaded from

and MacGregor, 2000). This is evident in the current study, where the oat diets returned a greater insoluble proportion of β-glucan compared with the β-glucan-supplemented wheat-based diet. The effect of extrac-tion on solubility may influence the capacity of puri-fied β-glucans to persist more distally along the GIT, possibly being degraded and absorbed prececally when compared with intact forms (Johansen et al., 1993). Despite this, Bach Knudsen et al. (1993b) demonstrat-ed that, although subject to a greater rate of degra-dation prececally compared with intact forms, purified oat β-glucans are available for fermentation in the dis-tal portion of the GIT. Second, an additional cell wall constituent or a cell wall component complexed with β-glucan in oats (Izydorczyk and MacGregor, 2000), which is not presented in the β-glucan-supplemented wheat-based diet, may be fermented readily in the dis-tal GIT and thus influence luminal composition (Bach Knudsen et al., 1993b).
Indole and skatole are indolic compounds and bacte-rial metabolites of tryptophan fermentation (Le et al., 2005; Blachier et al., 2007). Williams and Evans (1981) designated both skatole and indole as significant con-tributors to the malodorous smell associated with pig manure, whereas skatole in particular has been char-acterized as possessing a fecal or nauseating quality, detectable at minute concentrations by the human nose (Zahn et al., 2001; Le et al., 2005). There was a distinct difference in the production of skatole and indole in the distal GIT in the current study, with consumption of the oat diets producing greater concentrations of in-dole in the colon, whereas by contrast, consumption of the wheat-based diet resulted in greater concentrations of skatole. The diet-dependent response provoking the divergent pathways of tryptophan degradation is note-worthy. Previously, Jensen et al. (1995) have observed a decrease in gastrointestinal skatole in association with greater concentrations of dietary fermentable carbohy-drates, whereas Xu et al. (2002) described increased concentrations of indole at the expense of skatole in pigs offered a source of fermentable carbohydrate. These latter authors ascribed the transfer in the production of these tryptophan metabolites to promotion of carbohy-drate fermentation and saccharolytic bacterial genera such as bifidobacteria, which the findings of the current study support and indicate greater proteolytic activity in the distal GIT of pigs offered the purified β-glucan.
Consumption of the wheat-based diet supplemented with oat β-glucans resulted in a compositional shift in the contents of the distal colon compared with pigs of-fered the oat-based diets. The relative proportions of VFA concentrations were modified, such that consump-tion of the wheat-based diet containing β-glucans re-sulted in greater production of the branch-chained fatty acids, isovaleric and isobutyric acid, and also valeric acid. These fermentation end products may indicate proteolytic activity in the distal GIT because they are specific by-products of AA deamination (Mortensen et
al., 1988; Marounek et al., 2002). Furthermore, these metabolites are useful markers of potential odor devel-opment because they possess malodorous characteristics detectable at minute concentrations by human olfactory senses (Le et al., 2005). Previously, studies have found an association between typically saccharolytic bacterial genera and decreased concentrations of VFA end prod-ucts of AA deamination (O’Shea et al., 2010a,b). The greater numbers of lactobacilli and bifidobacteria and reduced proportions of odorous VFA in the distal colon of pigs offered whole-oat based diets compared with the β-glucan-supplemented wheat-based diets may offer fur-ther support for this relationship. The observation that a greater proportion of AA-derived VFA was present in the distal colon of pigs offered β-glucan-supplemented wheat-based diets contributes further evidence to the absence of functional activity of purified β-glucans at the distal portion of the colon.
The disposition toward greater odorous VFA pro-duction in the distal GIT of pigs offered the β-glucan-supplemented wheat-based diet was correspondingly reflected in the manure at 0 and 240 h compared with consumption of the oat-based diets. Our group has previously noted that patterns of VFA production ob-served in the distal GIT may persist in excreted ma-nure and thus provide an indication on the role of distal GIT fermentation in development of subsequent ma-nure odor emissions (O’Shea et al., 2010a,b). Otto et al. (2003) have similarly reported associations between these odorous compounds in manure and subsequent odor development. The current study would appear to confirm these relationships; the β-glucan-supplemented wheat-based diet emitted more than 50% more odor compared with manure from pigs offered the oat diets. Manure ammonia emissions were attenuated from 0 to 240 h in response to the consumption of the oats com-pared with the wheat-based diet supplemented with β-glucans. Supplementation with a β-glucanase com-posite induced a negligible effect on subsequent manure ammonia emissions, indicating that dietary β-glucans either in a purified form or as presented intact in oats were not entirely responsible for mediating the produc-tion of this gaseous compound. The role of dietary fer-mentable carbohydrate in influencing manure ammonia may be attributed to other polysaccharides, and not specifically to β-glucans (Kerr et al., 2006).
In conclusion, the current study demonstrates that increased consumption of oat β-glucans via supplemen-tation to a wheat-based diet may be achieved without reducing energy and nutrient digestibility, as has been observed with oat diets. However, the application of purified β-glucans in modulating the distal GIT and subsequently influencing manure composition and gas-eous emission may be limited. Nonetheless, a definite relationship would appear to be emerging between sev-eral bacterial and metabolite markers in the distal GIT and manure odor. Although the strategy to use the β-glucan component of cereals such as oats to benefi-
Purified and intact dietary β-glucans in pigs 445
at Swets Blackwell Inc on January 25, 2011. jas.fass.orgDownloaded from

cially modulate these markers was unsuccessful, the current study may offer a template for future interven-tions to modify gastrointestinal fermentation, and thus manure composition and gaseous emissions from finish-ing pig production.
LITERATURE CITED
AOAC. 1995. Official Methods of Analysis. 16th ed. Assoc. Off. Anal. Chem., Washington, DC.
Bach Knudsen, K. E. 1997. Carbohydrate and lignin contents of plant materials used in animal feeding. Anim. Feed Sci. Tech-nol. 67:319–338.
Bach Knudsen, K. E., B. B. Jensen, and I. Hansen. 1993a. Digestion of polysaccharides and other major components in the small and large intestine of pigs fed on diets consisting of oat fractions rich in beta-d-glucan. Br. J. Nutr. 70:537–556.
Bach Knudsen, K. E., B. B. Jensen, and I. Hansen. 1993b. Oat bran but not a β-glucan-enriched oat fraction enhances butyrate pro-duction in the large intestine of pigs. J. Nutr. 123:1235–1247.
Bedford, M. R. 2000. Exogenous enzymes in monogastric nutrition—Their current value and future benefits. Anim. Feed Sci. Tech-nol. 86:1–13.
Blachier, F., F. Mariotti, J. F. Huneau, and D. Tome. 2007. Ef-fects of amino acid-derived luminal metabolites on the colonic epithelium and physiopathological consequences. Amino Acids 33:547–562.
Carpenter, D. A., F. P. O’ Mara, and J. V. O’ Doherty. 2004. The effect of dietary crude protein concentration on growth perfor-mance, carcass composition and nitrogen excretion in entire grower-finisher pigs. Ir. J. Agric. Food Res. 43:227–236.
Conway, E. J. 1957. Microdiffusion Analysis and Volumetric Error. Crosby Lockwood and Son, London, UK.
Galassi, G., G. M. Crovetto, L. Rapetti, and A. Tamburini. 2004. Energy and nitrogen balance in heavy pigs fed different fibre sources. Livest. Prod. Sci. 85:253–262.
Garry, B. P., M. Fogarty, T. P. Curran, M. J. O’Connell, and J. V. O’Doherty. 2007. The effect of cereal type and enzyme addition on pig performance, intestinal microflora, and ammonia and odour emissions. Animal 1:751–757.
Graham, H., K. Hesselman, and P. Aman. 1986. The influence of wheat bran and sugar-beet pulp on the digestibility of dietary components in a cereal-based pig diet. J. Nutr. 116:242–251.
Hayes, E. T., A. B. Leek, T. P. Curran, V. A. Dodd, O. T. Carton, V. E. Beattie, and J. V. O’Doherty. 2004. The influence of diet crude protein level on odour and ammonia emissions from fin-ishing pig houses. Bioresour. Technol. 91:309–315.
Heo, J. M., J. C. Kim, C. F. Hansen, B. P. Mullan, D. J. Hampson, and J. R. Pluske. 2009. Feeding a diet with decreased pro-tein content reduces indices of protein fermentation and the incidence of postweaning diarrhea in weaned pigs challenged with an enterotoxigenic strain of Escherichia coli. J. Anim. Sci. 87:2833–2843.
Iwaki, K., N. Nimura, Y. Hiraga, T. Kinoshita, K. Takeda, and H. Ogura. 1987. Amino acid analysis by reversed-phase high-performance liquid chromatography. J. Chromatogr. 407:273–279.
Izydorczyk, M. S., and A. W. MacGregor. 2000. Evidence of inter-molecular interactions of β-glucans and arabinoxylans. Carbo-hydr. Polym. 41:417–420.
Jensen, M. T., R. P. Cox, and B. B. Jensen. 1995. Microbial pro-duction of skatole in the hind gut of pigs given different diets and its relations to skatole deposition in backfat. Anim. Sci. 61:293–304.
Johansen, H. N., P. J. Wood, and K. E. B. Knudsen. 1993. Molecular weight changes in the (1→3)(1→4)-β-d-glucan of oats incurred by the digestive processes in the upper gastrointestinal tract of pigs. J. Agric. Food Chem. 41:2347–2352.
Jonsson, E., and S. Hemmingsson. 1991. Establishment in the pig-let gut of lactobacilli capable of degrading mixed-linked beta-glucans. J. Appl. Bacteriol. 70:512–516.
Kerr, B. J., C. J. Ziemer, S. L. Trabue, J. D. Crouse, and T. B. Par-kin. 2006. Manure composition of swine as affected by dietary protein and cellulose concentrations. J. Anim. Sci. 84:1584–1592.
Kiarie, E., C. M. Nyachoti, B. A. Slominski, and G. Blank. 2007. Growth performance, gastrointestinal microbial activity, and nutrient digestibility in early-weaned pigs fed diets containing flaxseed and carbohydrase enzyme. J. Anim. Sci. 85:2982–2993.
Kirchgessner, M., M. Kreuzer, A. Machmüller, and D. A. Roth-Maier. 1994. Evidence for a high efficiency of bacterial protein synthesis in the digestive tract of adult sows fed supplements of fibrous feedstuffs. Anim. Feed Sci. Technol. 46:293–306.
Le, P. D., A. J. A. Aarnink, N. W. M. Ogink, P. M. Becker, and M. W. A. Verstegen. 2005. Odour from animal production facili-ties: Its relationship to diet. Nutr. Res. Rev. 18:3–30.
Leek, A. B. G., J. J. Callan, P. Reilly, V. E. Beattie, and J. V. O’Doherty. 2007. Apparent component digestibility and manure ammonia emission in finishing pigs fed diets based on barley, maize or wheat prepared without or with exogenous non-starch polysaccharide enzymes. Anim. Feed Sci. Technol. 135:86–99.
Lynch, M. B., C. J. O’Shea, T. Sweeney, J. J. Callan, and J. V. O’Doherty. 2008. Effect of crude protein concentration and sug-ar-beet pulp on nutrient digestibility, nitrogen excretion, intes-tinal fermentation and manure ammonia and odour emissions from finisher pigs. Animal 2:425–434.
Lynch, M. B., T. Sweeney, J. J. Callan, and J. V. O’Doherty. 2007. Effects of increasing the intake of dietary β-glucans by exchang-ing wheat for barley on nutrient digestibility, nitrogen excretion, intestinal microflora, volatile fatty acid concentration and ma-nure ammonia emissions in finishing pigs. Animal 1:812–819.
Mackie, R. I., P. G. Stroot, and V. H. Varel. 1998. Biochemical identification and biological origin of key odor components in livestock waste. J. Anim. Sci. 76:1331–1342.
Mariscal-Landín, G., B. Seve, Y. Colleaux, and Y. Lebreton. 1995. Endogenous amino nitrogen collected from pigs with end-to-end ileorectal anastomosis is affected by the method of estimation and altered by dietary fiber. J. Nutr. 125:136–146.
Marounek, M., T. Adamec, V. Skivanova, and N. I. Latsik. 2002. Nitrogen and in vitro fermentation of nitrogenous substrates in caecal contents of the pig. Acta Vet. (Brno) 71:429–433.
McCleary, B. V., and M. Glennie-Holmes. 1985. Enzymatic quan-tification of (1,3)(1,4)-β-d-glucan in barley and malt. J. Inst. Brew. 91:285–295.
Mortensen, P. B., K. Holtug, and H. S. Rasmussen. 1988. Short-chain fatty acid production from mono- and disaccharides in a fecal incubation system: implications for colonic fermentation of dietary fibre in humans. J. Nutr. 118:321–325.
NRC. 1998. Nutrient Requirements of Swine. 10th rev. ed. Natl. Acad. Press, Washington, DC.
O’Connell, J. M., J. J. Callan, C. Byrne, T. Sweeney, and J. V. O’Doherty. 2005. The effect of cereal type and exogenous en-zyme supplementation in pig diets on nutrient digestibility, intestinal microflora, volatile fatty acid concentration and ma-nure ammonia emissions. Anim. Sci. 81:357–364.
O’Doherty, J. V., K. M. Pierce, and D. A. Kenny. 2005. Fermentable fibre and gut health in non- and pre-ruminants. Pages 103–128 in Recent Advances in Animal Nutrition. P. C. Garnsworthy and J. Wiseman. Nottingham University Press, Nottingham, UK.
O’Shea, C. J., B. Lynch, M. B. Lynch, J. J. Callan, and J. V. O’Doherty. 2009. Ammonia emissions and dry matter of sepa-rated pig manure fractions as affected by crude protein con-centration and sugar beet pulp inclusion of finishing pig diets. Agric. Ecosyst. Environ. 131:154–160.
O’Shea, C. J., M. B. Lynch, J. J. Callan, and J. V. O’Doherty. 2010a. Dietary supplementation with chitosan at high and low crude protein concentrations promotes Enterobacteriaceae in
O’Shea et al.446
at Swets Blackwell Inc on January 25, 2011. jas.fass.orgDownloaded from

the caecum and colon and increases manure odour emissions from finisher boars. Livest. Sci. 134:198–201.
O’Shea, C. J., T. Sweeney, M. B. Lynch, D. A. Gahan, J. J. Cal-lan, and J. V. O’Doherty. 2010b. Effect of β-glucans contained in barley- and oat-based diets and exogenous enzyme supple-mentation on gastrointestinal fermentation of finisher pigs and subsequent manure odor and ammonia emissions. J. Anim. Sci. 88:1411–1420.
Otto, E. R., M. Yokoyama, S. Hengemuehle, R. D. von Bermuth, T. van Kempen, and N. L. Trottier. 2003. Ammonia, volatile fatty acids, phenolics, and odor offensiveness in manure from growing pigs fed diets reduced in protein concentration. J. Anim. Sci. 81:1754–1763.
Pierce, K. M., T. Sweeney, P. O. Brophy, J. J. Callan, E. Fitzpat-rick, P. McCarthy, and J. V. O’Doherty. 2006a. The effect of lactose and inulin on intestinal morphology, selected microbial populations and volatile fatty acid concentrations in the gastro-intestinal tract of the weanling pig. Anim. Sci. 82:311–318.
Pierce, K. M., T. Sweeney, P. O. Brophy, J. J. Callan, P. McCarthy, and J. V. O’Doherty. 2005. Dietary manipulation post wean-ing to improve piglet performance and gastro-intestinal health. Anim. Sci. 81:347–356.
Pierce, K. M., T. Sweeney, J. J. Callan, C. Byrne, P. McCarthy, and J. V. O’Doherty. 2006b. The effect of inclusion of a high lactose supplement in finishing diets on nutrient digestibility, nitrogen excretion, volatile fatty acid concentrations and ammonia emis-sion from boars. Anim. Feed Sci. Technol. 125:45–60.
Porter, M. G., and R. S. Murray. 2001. The volatility of compo-nents of grass silage on oven drying and the inter-relationship between dry-matter content estimated by different analytical methods. Grass Forage Sci. 56:405–411.
Sauvant, D., J.-M. Perez, and G. Tran. 2004. Tables of Composition and Nutritional Value of Feed Materials. Pigs, Poultry, Cattle,
Sheep, Goats, Rabbits, Horses, Fish. 2nd ed. Wageningen Aca-demic Publishers, Wageningen, the Netherlands.
Schiffman, S. S., E. A. Sattely Miller, M. S. Suggs, and B. G. Gra-ham. 1995. The effect of environmental odors emanating from commercial swine operations on the mood of nearby residents. Brain Res. Bull. 37:369–375.
Somogyi, M. 1960. Modifications of two methods for the assay of amylase. Clin. Chem. 6:23–35.
Souffrant, W. B. 2001. Effect of dietary fibre on ileal digestibility and endogenous nitrogen losses in the pig. Anim. Feed Sci. Technol. 90:93–102.
Van Soest, P. J., J. B. Robertson, and B. A. Lewis. 1991. Meth-ods for dietary fibre, neutral detergent fiber and non starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74:3583–3597.
Varley, P. F., T. Sweeney, M. T. Ryan, and J. V. O. Doherty. 2010. The effect of phosphorus restriction during the weaner-grower phase on compensatory growth, serum osteocalcin and bone mineralization in gilts. Livest. Sci. doi:10.1016/j.livs-ci.2010.07.025.
Williams, A. G., and M. R. Evans. 1981. Storage of piggery slurry. Agric. Wastes 3:311–321.
Xu, Z.-R., C.-H. Hu, and M.-Q. Wang. 2002. Effects of fructooligo-saccharide on conversion of l-tryptophan to skatole and indole by mixed populations of pig fecal bacteria. J. Gen. Appl. Mi-crobiol. 48:83–90.
Zahn, J. A., A. A. DiSpirito, Y. S. Do, B. E. Brooks, E. E. Coo-per, and J. L. Hatfield. 2001. Correlation of human olfactory responses to airborne concentrations of malodorous volatile organic compounds emitted from swine effluent. J. Environ. Qual. 30:624–634.
Zhu, J. 2000. A review of microbiology in swine manure odor con-trol. Agric. Ecosyst. Environ. 78:93–106.
Purified and intact dietary β-glucans in pigs 447
at Swets Blackwell Inc on January 25, 2011. jas.fass.orgDownloaded from

References http://jas.fass.org/cgi/content/full/89/2/438#BIBL
This article cites 46 articles, 10 of which you can access for free at:
at Swets Blackwell Inc on January 25, 2011. jas.fass.orgDownloaded from





























