Embed Size (px)
Citation preview

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
Research Article
Connexin 43 and ERK Regulate Tension-Induced Signal Transduction in Human
Periodontal Ligament Fibroblasts†
Shengnan Li1,2
, Huajing Zhang1, Shuna Li
2,Yanqi Yang
3, Bo Huo
2*, Ding Zhang
1*
1Department of Stomatology, Peking Union Medical College Hospital, Chinese Academy of
Medical Sciences, Beijing 100730, P.R. China
2Department of Mechanics, School of Aerospace Engineering, Beijing Institute of Technology,
Beijing 100081, P.R. China
3Faculty of Dentistry, University of Hong Kong, Hong Kong SAR, China
*Corresponding authors: Dr. Ding Zhang ([email protected]), Department of Stomatology,
Peking Union Medical College Hospital, Peking Union Medical College and Chinese
Academy of Medical Sciences; Dr. Bo Huo ([email protected]), Department of Mechanics,
School of Aerospace Engineering, Beijing Institute of Technology, No. 5 South
Zhongguancun Street, Beijing 100081, People’s Republic of China
Running title: Tension-Induced Signal Transduction in hPDLFs.
†This article has been accepted for publication and undergone full peer review but has not been
through the copyediting, typesetting, pagination and proofreading process, which may lead to
differences between this version and the Version of Record. Please cite this article as doi:
[10.1002/jor.22830]
Additional Supporting Information may be found in the online version of this article.
Received 12 September 2014; Revised 7 January 2015; Accepted 15 January 2015
Journal of Orthopaedic Research
This article is protected by copyright. All rights reserved
DOI 10.1002/jor.22830

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
Abstract
Periodontal ligament (PDL) fibroblasts play an important role in preserving periodontal
homeostasis and transmitting mechanical signals to alveolar bone. Connexin 43 (Cx43), a gap
junction protein, is essential for bone homeostasis and regulates bone remodeling. However,
the function of Cx43 in human PDL fibroblast-regulated bone remodeling has not yet been
elucidated. In this study, human PDL fibroblasts were exposed to cyclic mechanical tension
with a maximum 5% elongation for different durations. We then examined the expression of
signaling molecules related to osteogenesis and osteoclastogenesis at both the mRNA and
protein levels as well as the activity of extracellular signal-regulated kinase (ERK) in human
PDL fibroblasts after loading. We found that mechanical tension increased Cx43, which
further upregulated osteogenic (e.g. RUNX2, Osterix and OPG) and down-regulated
osteoclastogenic (e.g. RANKL) signaling molecules. Suppressing Cx43 gene (Gja1) by
siRNA inhibited the increase in osteogenesis-related molecules but enhanced RANKL
expression. Similar to Cx43, activated ERK1/2 was also enhanced by mechanical tension and
suppressed by Cx43 siRNA. Inhibition of ERK1/2 signaling using PD98059 reduced the
tension-regulated increase in osteogenesis-related molecules but enhanced that of
osteoclastogenesis-related ones. These findings suggest that cyclic tension may involve into
the osteogenic or osteoclastogenetic differentiation potential of human PDL fibroblasts via
the Cx43-ERK1/2 signaling pathway. This article is protected by copyright. All rights reserved
Key Words: periodontal ligament; mechanical stretch; Cx43; osteogenesis;
osteoclastogenesis

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
Introduction
The periodontal ligament (PDL), which resides between the tooth and the alveolar bone,
sensitively mediates the transmission of mechanical stimuli to the alveolar bone. In response
to applied orthodontic forces, PDL cells constitutively express numerous cytokines and
growth factors, such as osteopontin (OPN), alkaline phosphatase (ALP), receptor activator of
nuclear factor kappa-B ligand (RANKL), osteoprotegerin (OPG), matrix metalloproteinases,
vascular endothelial growth factor, and transforming growth factor-beta, as well as various
anti-inflammatory cytokines 1-3
. These factors are thought to transfer mechanical signals to
osteocytes, osteoblasts and osteoclasts to regulate osteogenesis and osteoclastogenesis 4,5
.
Periodontal ligament fibroblasts (PDLFs) are the most abundant cells in the PDL and play
essential roles in maintaining its integrity. Under mechanical forces, PDLFs can induce bone
remodeling through RANKL, OPG or OPN expression, and it is believed that remodeling of
alveolar bone during orthodontic tooth movement is greatly affected by these proteins 6-9
.
Gap junctions (GJs), an important intercellular communication system, are found among
osteoblasts, osteoclasts and osteocytes and are able to sense and transmit signals induced by
mechanical forces 10,11
. Connexins (Cxs) and pannexins (Panx) are families of
channel-forming proteins responsible for intracellular and intercellular signaling. Of the 21
known GJ subunits, connexin 43 (Cx43) is the most prevalent 12
, and is shown to be involved
with the osteogenic response of bone cells subjected to loading in vivo as well as cultured
osteoblasts and osteocytes 13-15
. One possible mechanism for Cx43-mediated regulation of
osteoblast gene expression involves the activation of runt-related transcription factor-2
(RUNX2) and Osterix (Sp7), two transcription factors that are thought to be master regulators
of osteogenesis 16
. RUNX2 binds to the osteoblast-specific cis-acting element 2 (OSE2),
which is found in the promoter regions and mediates the expression of many
osteoblast-specific genes (osteocalcin, collagen type I, bone sialoprotein, OPN, ALP, and

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
collagenase-3) 17
. Osterix (Osx), a zinc-finger-containing transcription factor of the sp family,
is expressed in osteoblasts of all endochondral and membranous bones, and is required for the
differentiation of preosteoblasts into mature osteoblasts 18-20
.
Previous research has identified Cx proteins (Cx32, Cx40, Cx43, Cx45) in PDLFs 21
,
and cyclic stretch has been shown to influence the membrane Cx43 protein expression in
cultured human PDL cells 22
. However, whether Cx43 modulates signal transduction related
to osteogenesis in PDLFs under mechanical stimulation is unknown. In addition, some
studies have identified a role for extracellular signal-regulated kinase (ERK) in influencing
osteogenesis in mechanical force-exposed periodontal fibroblasts 6,8
. Given that GJs regulate
signal transduction through the ERK pathway 23
, we hypothesized that mechanical tension
may regulate osteogenesis and the expression of Cx43 in PDLFs, and that Cx43 is required
for the osteogenic response of PDLFs to mechanical tension through ERK signaling.
Materials and Methods
Cell culture
Human PDLFs (hPDLFs; ScienCell, Carlsbad, CA) were maintained in Dulbecco's modified
Eagle's medium (Hyclone, Thermo Scientific, Wilmington, DE) containing 10% fetal bovine
serum, 100 μg/mL penicillin, and 100 μg/mL streptomycin (all from Invitrogen, Carlsbad,
CA) at 37°C in a humidified atmosphere of 5% CO2. Cells were passaged at 80–90%
confluence, and generally cells from passages 4 to 6 were used for experiments.
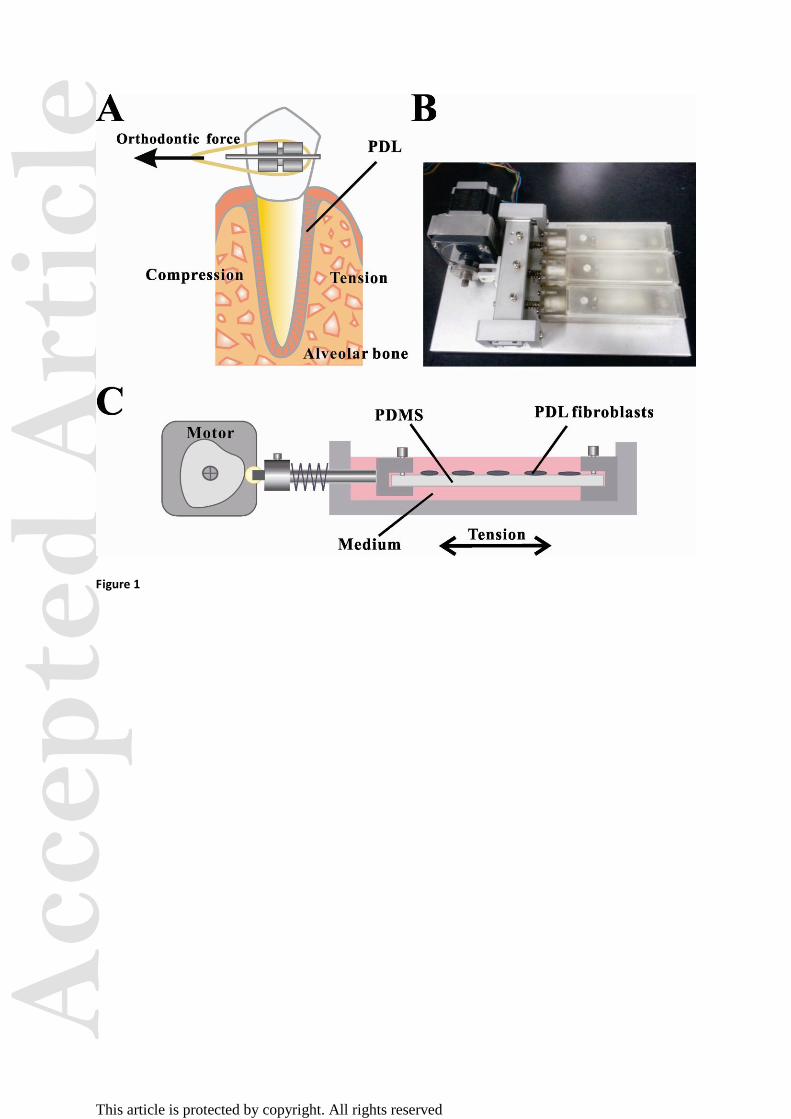
Cyclic mechanical tension
A custom-made tensile device, comprising elastic membranes, culture chambers and a motor,
was adopted to stretch cells adhering to an elastic membrane in a uniaxial manner (Fig. 1).
Elastic membranes were made using poly dimethyl-siloxane (PDMS) gel (Sylgard 184, Dow
Corning, Midland, MI). Two components mixed at a ratio of 1:10, and the mixture was

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
poured onto plates in suitable dimensions for the tensile device and left to stand for 12 h. The
membranes were then cured at 65°C for 2 h and at room temperature (~25°C) for 12 h. For
seeding cells, the resultant 3-mm-thick membranes were treated by oxygenized plasma.
Fibroblasts were grown on the membranes until 75–85% confluence and the cultures were
then mounted to the tensile device. Cells were cyclically stretched (3 min/cycle) with a
maximal deformation of 5% for 0, 0.5, 1, 2, 4, 8, 12 and 24 h, respectively. In the control
experiments, PDLFs were cultured on the PDMS membranes in the absence of mechanical
tension.
Real-time quantitative polymerase chain reaction (qPCR)
Total RNA was obtained using Trizol (Invitrogen), and reverse transcribed using a Revert Aid
First-strand cDNA Synthesis Kit (TransGen Biotech, Beijing, China) according to the
manufacturer’s instructions. Real-time quantitative PCR was carried out with the SYBR
Premix EXTaq (TaKaRa Biotechnology, Dalian, China) and Bio-Rad 5700 (Applied
Biosystems, Foster City, CA). Standard polymerase chain reaction were performed at 95°C
for 30 s followed by 40 cycles at 95°C for 5 s, then 60°C for 30 s. The experiment was
repeated three times. Relative mRNA levels per gene were normalized to the GAPDH
housekeeping gene. Oligonucleotide sequences for the genes of interest are listed in Table 1.
Western blotting
After stretch loading, cells were harvested, centrifuged, washed with PBS, and lysed for 30
min on ice in Cytobuster Lysis Buffer (Sigma-Aldrich, St Louis, MO). The whole cell
extracts from adherent cells were centrifuged at 14,000 rpm for 30 min at 4°C. Proteins were
quantified using the bicinchoninic acid (BCA) protein quantification kit (Pierce
Biotechnology, Rockford, IL). Samples were then separated using sodium dodecyl sulfate
polyacrylamide gel electrophoresis on 12% gels and blotted onto nitrocellulose membranes.
Membranes were blocked for 1 h at room temperature with 5% nonfat milk or bovine serum

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
albumin (BSA) in Tris-buffered solution with Tween-20 (TBST) and then incubated with the
following primary antibodies diluted in 5% nonfat milk or BSA overnight at 4°C with
shaking: rabbit anti-Cx43, anti-RANKL, anti-OPG, anti-Osterix, anti-RUNX2 (Abcam,
Cambridge, MA), p-ERK, and ERK (Cell signaling technology). After three washes with
TBST, the blots were incubated with horseradish peroxidase-conjugated affinity-purified goat
anti-rabbit IgG (Abcam; 1:5,000) for 2 h at room temperature, incubated with enhanced
chemiluminescence and exposed to X-ray film for visualization (Eastman-Kodak, China).
The integrated density value of each band was assessed using Image-Pro Plus (Media
Cybernetics Inc., Rockville, MD). Protein levels were normalized to β-actin.
Cx43 siRNA experiments
Cells were plated on PDMS membranes for 24 h and transiently transfected with siRNA
oligonucleotides using siRNA Transfection Reagent (GenePharma Corporation, Shanghai,
China) using these sequences: sense, 5′-GCCGCAAUUACAACAAGCATT-3′; antisense,
5′-UGCUUGUUGUAAUUGCGGCTT-3′. Cyclic tension was applied 24 h after transfection
and sustained for different durations. A vacant RNA carrier was used as the scrambled group
and cells not subjected to tension as the control group. All experiments were repeated thrice.
Statistical analysis
All data are presented as the mean ± standard deviation (SD). Statistical differences among
the mean values of different groups were determined with one-way analysis of variance
(ANOVA) as well as Bonferroni post-hoc test using SPSSv.10.0 (Chicago, IL), and was
considered significant when p<0.05.
Results
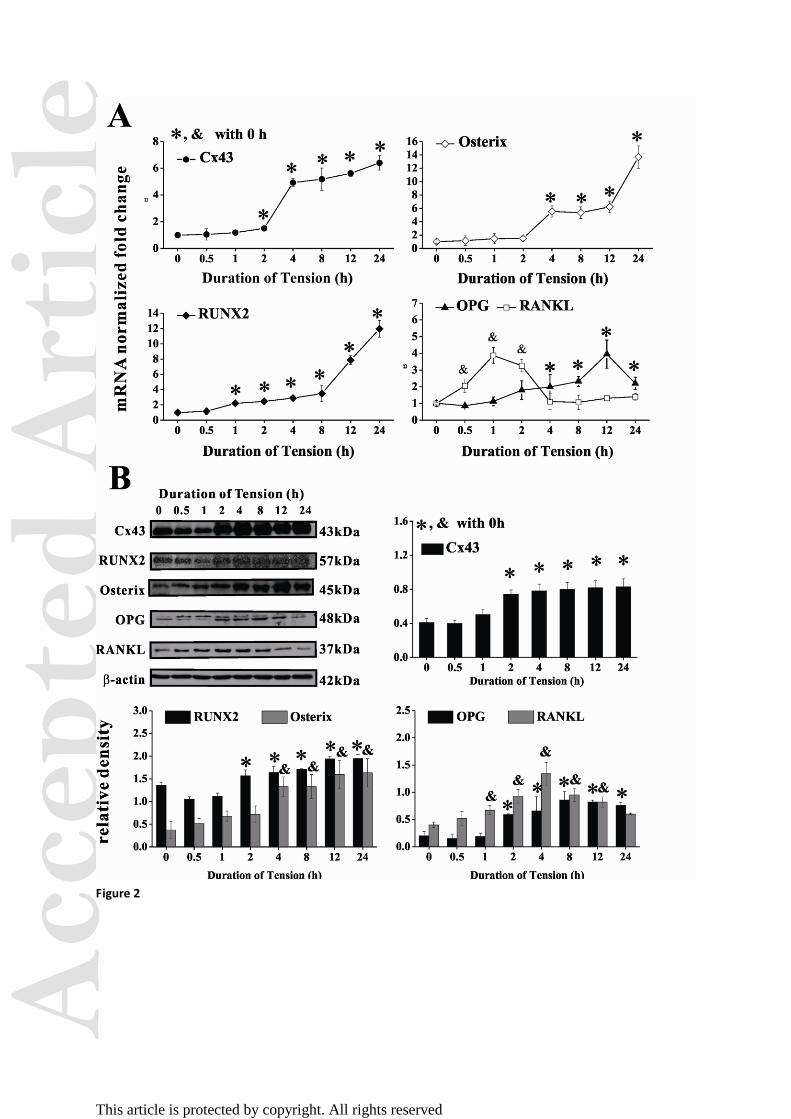
Cyclic mechanical tension modulates signaling molecules
To determine if Cx43 expression is regulated by mechanical stretching in PDLFs, we used

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
quantitative RT-PCR (qRT-PCR) to analyze Cx43 expression in stretched conditions over
time. Cx43 gene (Gja1) expression increased concomitant with longer periods of tension,
with a 5-fold increase in Gja1 after 4 h of stretching as compared with the static control
group (Fig. 2A). RUNX2 and Osterix mRNA levels in PDLFs were also increased in a
time-dependent manner following mechanical tension, indicating the Cx43 may be involved
in osteoblast differentiation. As for signaling molecules related to osteoclastogenesis,
RANKL mRNA increased immediately within the first hour (3.87-fold) followed by a trend
toward a down-regulation in expression. In contrast, OPG mRNA expression was upregulated
as compared with the static control groups; there was a slight down-regulation at 24 h.
Western blotting showed similar results (Fig. 2B). RUNX2 and Osterix proteins increased
along with tension duration, whereas RANKL increased until 4 h—slower than its mRNA
level—and then decreased. OPG also followed a down-regulation trend after peaking in
expression. These results indicate that a maximum of 5% cyclic tension upregulates Cx43
expression and promotes the expression of osteogenic genes but inhibits genes associated
with osteoclastogenesis.
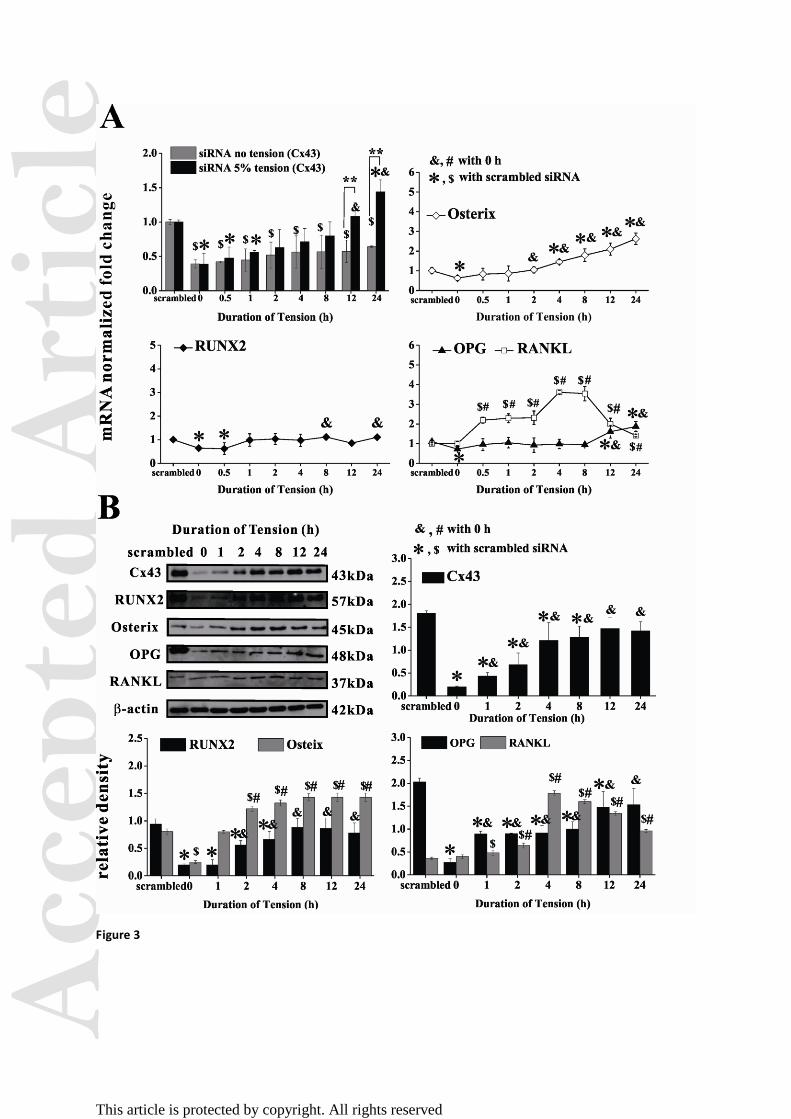
Cx43 silencing influences tension-regulated signaling
To further examine the role of Cx43 in hPDLFs under mechanical tension, we silenced Gja1
expression using siRNA. The scrambled siRNA data at each time point of tensile stimulation
showed similar trends of gene expression to those without any chemical treatments (Fig.
S1-S6). Within the first hour of tension, siRNA effectively suppressed Gja1 in stretched and
unstretched cells as compared with scrambled siRNA (Fig. 3A). After 12 h of tension, Gja1
expression in stretched fibroblasts almost recovered to that of the control group without
silencing Cx43, whereas by 24 h, Cx43 mRNA level had increased by 1.4-fold; this recovery
was not apparent in the unstimulated group. Western blotting results showed that siRNA also

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
significantly depressed Cx43 protein with or without tension, and the protein was recovered
to original levels by 24 h. Both RUNX2 and OPG mRNA were suppressed initially by siRNA
but fluctuated around baseline levels after exposure to stretching. By 8 h of stretching, OPG
expression had slightly increased (1.6-fold) as compared with the scrambled control. Osterix
mRNA was also suppressed initially following silencing of Cx43 and then upregulated in a
time-dependent manner under tension. On the contrary, RANKL mRNA was increased with
Cx43 silencing, reached a maximum expression at 4 h (3.6-fold); after this, RANKL mRNA
expression gradually decreased along with Cx43 recovery. These results were confirmed by
western blotting (Fig. 3B).
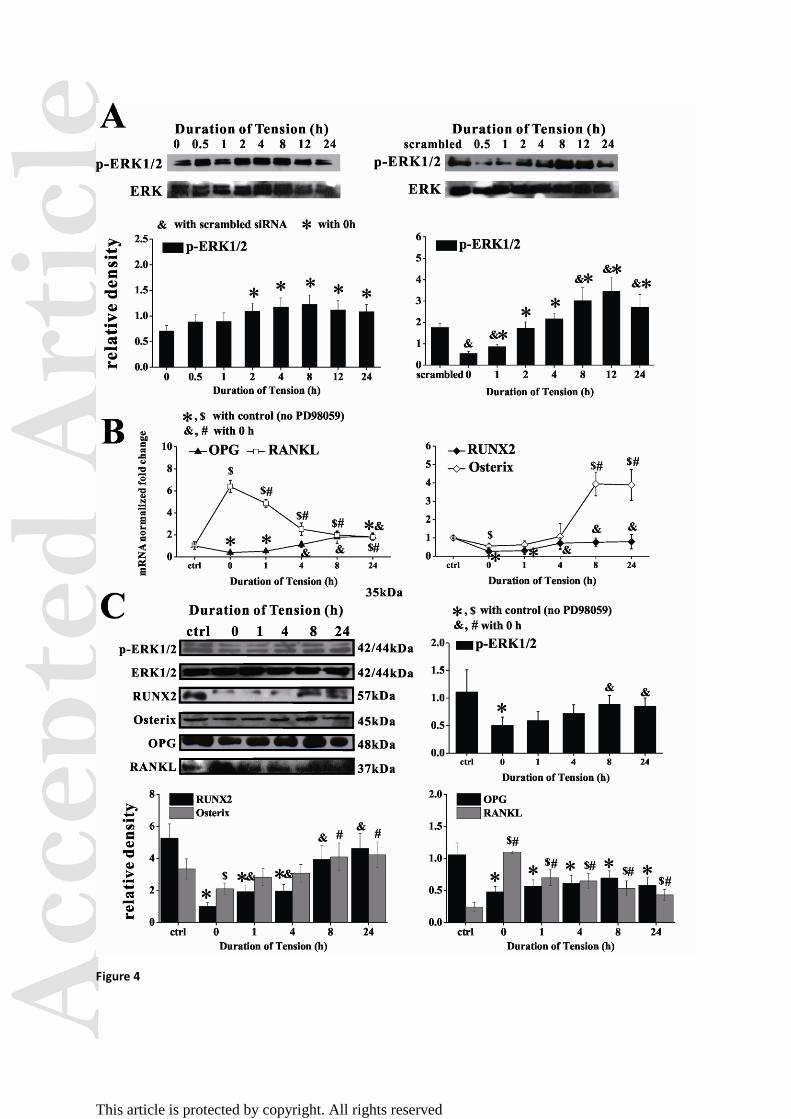
Stretching restored the signaling variation induced by blocking ERK1/2
ERK1/2 activation is required for mechanotransduction in cells 6,8
, and ERK1/2 is activated
upstream of osteoblast differentiation-related genes mediated by Cx43 24
. Thus, we next
examined ERK phosphorylation (p-ERK) as a possible signaling pathway regulating the
expression of osteogenic genes in stretched hPDLFs. We found that tension-induced ERK1/2
activation increased in a time-dependent manner, with significant increases observed after 2 h
of tension (p < 0.05; Fig. 4A). This increase could be suppressed by Cx43 siRNA (p < 0.05),
indicating that Cx43 lies upstream of ERK1/2 activation. When hPDLFs were pretreated with
10 μM PD98059 (Selleck Chemicals, Houston, TX), an ERK1/2 inhibitor, the mRNA and
protein levels of Osterix, RUNX2 and OPG were significantly reduced but those of RANKL
were enhanced (Fig. 4B and C). It should be noted DMSO, the solvent of PD98059, did not
interfere with the inhibitor (see Figure S8). Tension was able to reactivate ERK1/2 after its
phosphorylation was inhibited by Cx43-specific siRNA (Fig. 4A) or PD98059 (Fig. 4C),
whereas Osterix, RUNX2 and OPG expression increased slowly as compared with the static
control group. RANKL expression, by comparison, transiently increased but then decreased

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
in a time-dependent manner in stretched cells following PD98059. These results strongly
suggest a relationship between Cx43 and tension-induced osteogenic signaling in hPDLFs
through ERK1/2 signaling.
Discussion
During orthodontic treatment, the PDL is indispensable for controlling tooth movement via
bone remodeling. Human PDL cells exposed to mechanical strain in vitro are thought to
express an osteogenic and osteoclastogenic transcriptional profile 1,6,25
. In the present study,
we used a tensile device to investigate the influence of mechanical tension on osteogenic and
osteoclastogenic signaling pathways, as regulated by Cx43 and ERK1/2, in hPDLFs.
Cx43 is an important modulator of bone cell response to mechanical cues, hormones or
growth factors 26,27
. Our previous research has demonstrated that Cx43 regulates
osteoclastogenesis and osteogenesis in osteocytes under fluid shear stress 28
. Recent evidence
also implicates a role for Cx43 in human PDL cell function 29
. In agreement with the previous
in vivo and in vitro studies 22,30
, our present data show that mechanical tension increases the
expression of Cx43, which might facilitate gap junctional communication in a
time-dependent manner in hPDLFs. Using siRNA, we efficiently silenced Cx43 expression,
and found that even when subjected to tension for 24 h, Cx43 protein showed a 60%
reduction in expression as compared with that observed before Cx43 silencing (Fig. 3B). This
suggests that there might be a decrease in gap junctional coupling and intercellular
communication with Cx43 silencing.
Along with the tension-induced increase in Cx43, the expression of Osterix and RUNX2,
two important regulators of osteogenesis, was upregulated. One previous study adopted the
mechanical vibration for 30 min every 24 h to stimulate hPDL stem cells, showing that 50 Hz
vibration significantly increased the mRNA and protein levels of Osterix and RUNX2,

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
respectively 31
. The increase of relative mRNA levels, 2.01-fold for Osterix and 3.59-fold for
RUNX2, coincided with our present study (Fig. 2B). Another study showed that 10%
elongation of tensile loading in 0.5 Hz frequency, greatly stronger than 5% elongation in
0.0056 Hz (3 min/cycle) used in our study, led to a slight increase 1.25-fold of Osterix at 24 h
stimulation in PDL cells 32
. When silencing Cx43, our current results revealed that both
Osterix and RUNX2 were suppressed at either mRNA or protein level. Therefore the present
study indicates that Cx43 is the critical factor to regulate the osteogenesis-related pathways in
PDLFs.
This study also showed that the cyclic tension significantly enhanced the mRNA and
protein levels of OPG, an osteoclastogenesis inhibitory factor, throughout the stretching
period. RANKL, an osteoclast differentiation factor, revealed an initial increase followed by
gradual decrease along with the increase of stimulation duration. This biphasic phenomenon
of RANKL, which may be due to the stress response of hPDLFs against the initial exposure
to tension, was also observed in our previous study on osteocytes under fluid flow 28
. Several
stydies have showed that during orthodontic treatment, the concentration of RANKL was
elevated at compression sides, and OPG expression was significantly reduced; While at
tension sides the opposite pattern was observed, supporting the pressure–tension hypothesis
33,34. One previous study has provided evidence that expression of OPG was upregulated and
RANKL mRNA expression did not change when PDL cells were loaded by intermittent
stretching with 20% elongation at 10 cycles/min similar to physiologically masticatory
condition35
. Furthermore, our present data reported that silencing Cx43 significantly inhibited
the tension-induced increase of OPG expression although the tensile loading for 12 h and 24
h can slightly restore it. RANKL, as expected, showed an opposite trend, with a strong

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
increase initially after Cx43 silencing followed by its down-regulation as Cx43 expression
recovered. A similar phenomenon has been shown in osteoblasts following Cx43 silencing
36,37. Thus, our current results indicate that cyclic tension may enhance intercellular gap
junctions and Cx43 gets involved in the osteoclastogenesis-related pathways in hPDLFs (Fig.
5).
We also sought to ascertain how Cx43 transmitted mechanical signals to regulate
osteogenesis in hPDLFs. Since the ERK1/2 pathway is considered as a vital, intracellular
signaling pathway in mediating bone cell responses to mechanical force 11,38
, this was deemed
a likely candidate. We found that Cx43 siRNA could inhibit ERK1/2 activation in hPDLFs,
which suggested that Cx43 may transmit cyclic tension signals to activate the ERK1/2
pathway. In addition, previous studies have demonstrated that Cx43-mediated osteoblastic
differentiation involves the transcription factors RUNX2 and Osterix (Sp7) through ERK1/2
in parallel with the PKC signaling pathway24,39,40
. Therefore, it is reasonable to assume that
the expression of osteogenic genes in hPDLFs may increase along with the activation of ERK
under cyclic tension. Our present study tested this hypothesis with an ERK selective inhibitor,
PD98059 and found that PD98059 treatment significantly suppresses the tension-upregulated
expression of Osterix, OPG and RUNX2 in hPDLFs. RANKL, on the other hand, increased
initially with PD98059 but then gradually decreased with exposure time. A similar study
found that centrifugal force regulates the expression of osteogenic genes in hPDLFs through
ERK1/2 activity8.These results indicate that ERK1/2 activation in response to
tension-induced Cx43 expression regulates osteogenesis and osteoclastogenesis in hPDLFs.
To our knowledge, this study is the first to show that mechanical tension activates the
osteogenic potential of hPDLFs through Cx43-ERK1/2 signaling. Others have demonstrated
that Cx43 is indispensable in intercellular communication and in maintaining bone
homeostasis 10,13
, and data show that bone cells respond to mechanical forces to open Cx43

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
hemichannels for the activation of calcium oscillation 41
, or to affect prostaglandin E2(PGE2)
42 and integrin α5β1
43 expression. However, controversy still exists as to the function of
Cx43 hemichannels in bone, especially in relation to other mechanical sensors 36,44
. Thus,
future work will focus on the regulatory mechanisms between Cx43 and other
mechanoreceptors or downstream signal pathways in these cells.
In summary, our study demonstrated that cyclic tension exerted on hPDLFs induced the
expression of Cx43 and its downstream signaling pathway ERK1/2, which further promoted
pathways related to osteogenesis but inhibited those related to osteoclastogenesis. This
critical connexin-ERK1/2-osteogenesis/osteoclastogenesis cascade may help to give insight
into the molecular mechanism of orthodontic loading-induced tooth movement.
Acknowledgments
This work was supported by the National Natural Science Foundation of China
(11372043 (BH) and 31070829, 31371389 (DZ)) and the Fundamental Research Funds for
the Central Universities (GZ2013015101 (BH)).
Conflict of interest statement: We declare that we have no conflicts of interest.
References:
1. Wongkhantee, S, Yongchaitrakul, T, Pavasant, P. 2008. Mechanical stress induces
osteopontin via ATP/P2Y1 in periodontal cells. J Dent Res 87: 564-568.
2. Cho, JH, Lee, SK, Lee, JW, Kim, EC. 2010. The role of heme oxygenase-1 in mechanical
stress- and lipopolysaccharide-induced osteogenic differentiation in human periodontal
ligament cells. Angle Orthod 80: 552-559.
3. Wescott, DC, Pinkerton, MN, Gaffey, BJ, et al. 2007. Osteogenic gene expression by

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
human periodontal ligament cells under cyclic tension. J Dent Res 86: 1212-1216.
4. Zhao, M, Xiao, G, Berry, JE, et al. 2002. Bone morphogenetic protein 2 induces dental
follicle cells to differentiate toward a cementoblast/osteoblast phenotype. J Bone Miner Res
17: 1441-1451.
5. Seo, BM, Miura, M, Gronthos, S, et al. 2004. Investigation of multipotent postnatal stem
cells from human periodontal ligament. Lancet 364: 149-155.
6. Kook, SH, Son, YO, Hwang, JM, et al. 2009. Mechanical force inhibits osteoclastogenic
potential of human periodontal ligament fibroblasts through OPG production and
ERK-mediated signaling. J Cell Biochem 106: 1010-1019.
7. Jacobs, C, Grimm, S, Ziebart, T, et al. 2013. Osteogenic differentiation of periodontal
fibroblasts is dependent on the strength of mechanical strain. Arch Oral Biol 58: 896-904.
8. Hong, SY, Jeon, YM, Lee, HJ, et al. 2010. Activation of RhoA and FAK induces
ERK-mediated osteopontin expression in mechanical force-subjected periodontal ligament
fibroblasts. Mol Cell Biochem 335: 263-272.
9. Diercke, K, Sen, S, Kohl, A, et al. 2011. Compression-dependent up-regulation of
ephrin-A2 in PDL fibroblasts attenuates osteogenesis. J Dent Res 90: 1108-1115.
10. Jiang, JX, Siller-Jackson, AJ, Burra, S. 2007. Roles of gap junctions and hemichannels in
bone cell functions and in signal transmission of mechanical stress. Front Biosci 12:
1450-1462.
11. Rubin, J, Rubin, C, Jacobs, CR. 2006. Molecular pathways mediating mechanical
signaling in bone. Gene 367: 1-16.
12. Yellowley, CE, Li, Z, Zhou, Z, et al. 2000. Functional gap junctions between osteocytic

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
and osteoblastic cells. J Bone Miner Res 15: 209-217.
13. Ziambaras, K, Lecanda, F, Steinberg, TH, Civitelli, R. 1998. Cyclic stretch enhances gap
junctional communication between osteoblastic cells. J Bone Miner Res 13: 218-228.
14. Robinson, JA, Chatterjee-Kishore, M, Yaworsky, PJ, et al. 2006. Wnt/beta-catenin
signaling is a normal physiological response to mechanical loading in bone. J Biol Chem 281:
31720-31728.
15. Tu, X, Rhee, Y, Condon, KW, et al. 2012. Sost downregulation and local Wnt signaling
are required for the osteogenic response to mechanical loading. Bone 50: 209-217.
16. Stains, JP, Lecanda, F, Screen, J, et al. 2003. Gap junctional communication modulates
gene transcription by altering the recruitment of Sp1 and Sp3 to connexin-response elements
in osteoblast promoters. J Biol Chem 278: 24377-24387.
17. Ducy, P, Zhang, R, Geoffroy, V, et al. 1997. Osf2/Cbfa1: a transcriptional activator of
osteoblast differentiation. Cell 89: 747-754.
18. Nakashima, K, Zhou, X, Kunkel, G, et al. 2002. The novel zinc finger-containing
transcription factor osterix is required for osteoblast differentiation and bone formation. Cell
108: 17-29.
19. Nishio, Y, Dong, Y, Paris, M, et al. 2006. Runx2-mediated regulation of the zinc finger
Osterix/Sp7 gene. Gene 372: 62-70.
20. Lee, JY, Lee, YM, Kim, MJ, et al. 2006. Methylation of the mouse DIx5 and Osx gene
promoters regulates cell type-specific gene expression. Mol Cells 22: 182-188.
21. Yamaoka, Y, Sawa, Y, Ebata, N, et al. 2000. Double expressions of connexin 43 and 32
in human periodontal ligament fibroblasts. Tissue Cell 32: 328-335.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
22. Xu, C, Fan, Z, Shan, W, et al. 2012. Cyclic stretch influenced expression of membrane
connexin 43 in human periodontal ligament cell. Arch Oral Biol 57: 1602-1608.
23. Buo, AM, Stains, JP. 2014. Gap junctional regulation of signal transduction in bone cells.
Febs Lett 588: 1315-1321.
24. Niger, C, Buo, AM, Hebert, C, et al. 2012. ERK acts in parallel to PKCdelta to mediate
the connexin43-dependent potentiation of Runx2 activity by FGF2 in MC3T3 osteoblasts.
Am J Physiol Cell Physiol 302: C1035-C1044.
25. Yang, YQ, Li, XT, Rabie, AB, et al. 2006. Human periodontal ligament cells express
osteoblastic phenotypes under intermittent force loading in vitro. Front Biosci 11: 776-781.
26. Plotkin, LI, Manolagas, SC, Bellido, T. 2002. Transduction of cell survival signals by
connexin-43 hemichannels. J Biol Chem 277: 8648-8657.
27. Morelli, S, Bilbao, PS, Katz, S, et al. 2011. Protein phosphatases: possible
bisphosphonate binding sites mediating stimulation of osteoblast proliferation. Arch Biochem
Biophys 507: 248-253.
28. Li, X, Liu, C, Li, P, et al. 2013. Connexin 43 is a potential regulator in fluid shear
stress-induced signal transduction in osteocytes. J Orthop Res 31: 1959-1965.
29. Kato, R, Ishihara, Y, Kawanabe, N, et al. 2013. Gap-junction-mediated communication in
human periodontal ligament cells. J Dent Res 92: 635-640.
30. Su, M, Borke, JL, Donahue, HJ, et al. 1997. Expression of connexin 43 in rat mandibular
bone and periodontal ligament (PDL) cells during experimental tooth movement. J Dent Res
76: 1357-1366.
31. Zhang, C, Li, J, Zhang, L, et al. 2012. Effects of mechanical vibration on proliferation

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
and osteogenic differentiation of human periodontal ligament stem cells. Arch Oral Biol 57:
1395-1407.
32. Li, L, Han, M, Li, S, et al. 2013. Cyclic tensile stress during physiological occlusal force
enhances osteogenic differentiation of human periodontal ligament cells via ERK1/2-Elk1
MAPK pathway. Dna Cell Biol 32: 488-497.
33. Van Schepdael, A, Vander, SJ, Geris, L. 2013. A mechanobiological model of
orthodontic tooth movement. Biomech Model Mechanobiol 12: 249-265.
34. Garlet, TP, Coelho, U, Silva, JS, Garlet, GP. 2007. Cytokine expression pattern in
compression and tension sides of the periodontal ligament during orthodontic tooth
movement in humans. Eur J Oral Sci 115: 355-362.
35. Tsuji, K, Uno, K, Zhang, GX, Tamura, M. 2004. Periodontal ligament cells under
intermittent tensile stress regulate mRNA expression of osteoprotegerin and tissue inhibitor
of matrix metalloprotease-1 and -2. J Bone Miner Metab 22: 94-103.
36. Watkins, M, Grimston, SK, Norris, JY, et al. 2011. Osteoblast connexin43 modulates
skeletal architecture by regulating both arms of bone remodeling. Mol Biol Cell 22:
1240-1251.
37. Bivi, N, Condon, KW, Allen, MR, et al. 2012. Cell autonomous requirement of connexin
43 for osteocyte survival: consequences for endocortical resorption and periosteal bone
formation. J Bone Miner Res 27: 374-389.
38. Liu, L, Shao, L, Li, B, et al. 2011. Extracellular signal-regulated kinase1/2 activated by
fluid shear stress promotes osteogenic differentiation of human bone marrow-derived
mesenchymal stem cells through novel signaling pathways. Int J Biochem Cell Biol 43:

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
1591-1601.
39. Lima, F, Niger, C, Hebert, C, Stains, JP. 2009. Connexin43 potentiates osteoblast
responsiveness to fibroblast growth factor 2 via a protein kinase C-delta/Runx2-dependent
mechanism. Mol Biol Cell 20: 2697-2708.
40. Stains, JP, Civitelli, R. 2005. Gap junctions regulate extracellular signal-regulated kinase
signaling to affect gene transcription. Mol Biol Cell 16: 64-72.
41. Jorgensen, NR, Teilmann, SC, Henriksen, Z, et al. 2003. Activation of L-type calcium
channels is required for gap junction-mediated intercellular calcium signaling in osteoblastic
cells. J Biol Chem 278: 4082-4086.
42. Cherian, PP, Siller-Jackson, AJ, Gu, S, et al. 2005. Mechanical strain opens connexin 43
hemichannels in osteocytes: a novel mechanism for the release of prostaglandin. Mol Biol
Cell 16: 3100-3106.
43. Batra, N, Burra, S, Siller-Jackson, AJ, et al. 2012. Mechanical stress-activated integrin
alpha5beta1 induces opening of connexin 43 hemichannels. Proc Natl Acad Sci U S A 109:
3359-3364.
44. Thi, MM, Islam, S, Suadicani, SO, Spray, DC. 2012. Connexin43 and pannexin1
channels in osteoblasts: who is the "hemichannel"? J Membr Biol 245: 401-409.

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
Table 1. Oligonucleotide sequences for the interested genes in this article
Forward Reverse
Cx43 5′- CCCTAAGAATGGTTCTGTGTATGTG -3′ 5′- GCAGGGACTTAAGGACAATCCTC -3′
OPG 5′- CAATTTGCCTGGCACCAAAG -3′ 5′- AGGTGAGGTTAGCATGTCCAATGT -3′
RANKL 5′- TGGATGCCTTGAATAATAAGCAGGA -3′ 5′- AATTTGCGGCACTTGTGGAA-3′
Osterix 5′- GCCATTCTGGGCTTGGGTA-3′ 5′- TGTGGCAGGGCCAGAGTCTA-3′
RUNX2 5′- TCCACACCATTAGGGACCATC-3′ 5′- TGCTAATGCTTCGTGTTTCCA-3′
GAPDH 5′-AAC CTG CCA AAT ATG ATG AC-3′ 5′-ATA CCA GGA AAT GAG CTT GA-3′

Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
Figure Captions
Figure 1. Effect of cyclic mechanical tension on PDL cells. (A) Schematic diagram of the
stretched PDL during orthodontic treatment. (B) Photograph of the custom-made tensile
device. (C) Side view diagram of the application of mechanical tension to PDLFs.
Figure 2. Effect of cyclic mechanical tension on signaling molecules in hPDLFs. (A)
Relative mRNA levels of Cx43, RUNX2, Osterix, RANKL and OPG by qPCR normalized to
GAPDH. (B) Western blotting and quantitative results normalized to GAPDH for Cx43,
RUNX2, Osterix, OPG and RANKL. *, &
p < 0.05 versus 0 h (n = 3).
Figure 3. Silencing Cx43 in hPDLFs under cyclic mechanical tension. (A) Relative mRNA
levels of Cx43, RUNX2, Osterix, RANKL and OPG were normalized to GAPDH (n=3). (B)
Western blots and quantitative results normalized to β-actin of Cx43, RUNX2, Osterix, OPG
and RANKL (n = 3). *, $
p < 0.05 versus scrambled siRNA groups. &, #
p < 0.05 versus 0 h.
Figure 4. Effect of ERK1/2 activation (p-ERK1/2) on cyclic mechanical tension-regulated
signaling in hPDLFs. (A) Western blots and quantitative results (normalized to β-actin) for
p-ERK1/2 and ERK1/2 (n = 3). &
p < 0.05 versus scrambled; *p < 0.05 versus 0 h. (B)
Relative mRNA levels of RUNX2, Osterix, RANKL and OPG normalized to GAPDH (n = 3).
(C) Western blots and quantitative results normalized to β-actin of p-ERK1/2, ERK1/2,
RUNX2, Osterix, OPG and RANKL (n = 3). For (B and C), *, $
p < 0.05 versus control (no
PD98059). &, #
p < 0.05 versus 0 h.
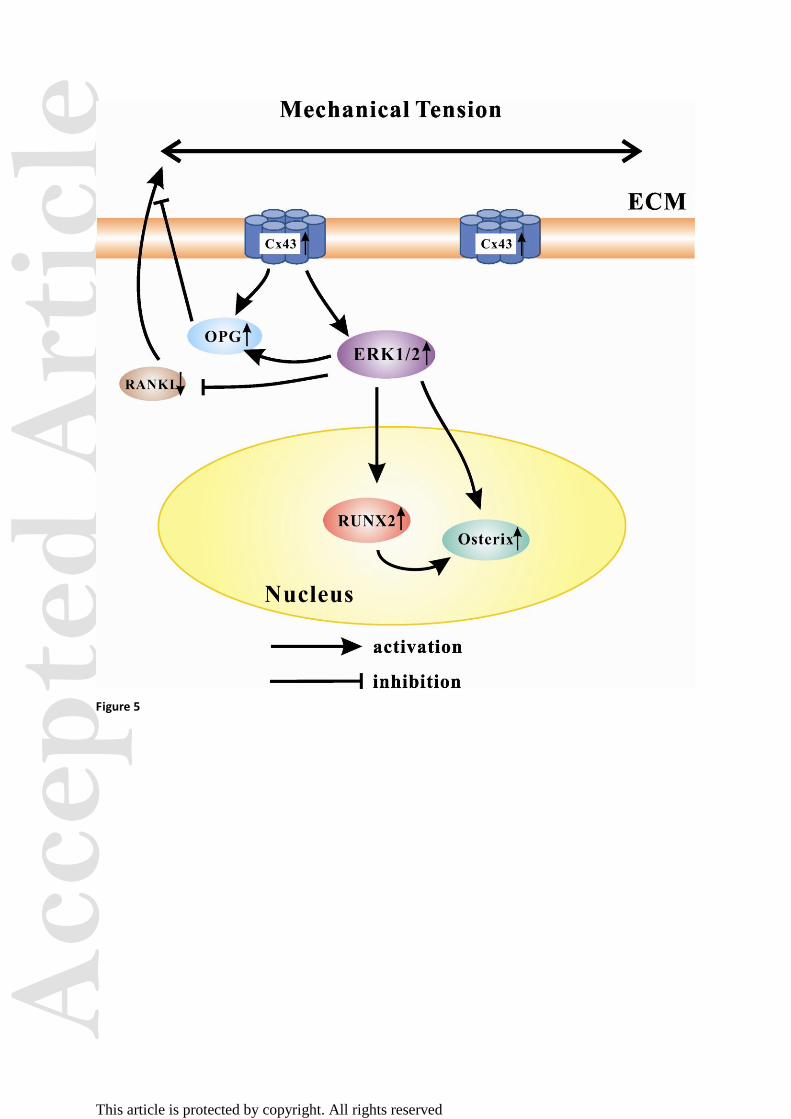
Figure 5. Schematic diagram showing the roles of Cx43 on mechanical tension-regulated
signaling of osteogenesis and osteoclastogenesis in hPDLFs.
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
Figure 1
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
Figure 2
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
Figure 3
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
Figure 4
Acc
epte
d A
rtic
le
This article is protected by copyright. All rights reserved
Figure 5





























