Embed Size (px)
Citation preview

ORIGINAL PAPER
Consilience of genetics and archaeobotany in the entangledhistory of rice
Dorian Q. Fuller & Yo-Ichiro Sato & Cristina Castillo & Ling Qin & Alison R. Weisskopf &Eleanor J. Kingwell-Banham & Jixiang Song & Sung-Mo Ahn & Jacob van Etten
Received: 30 March 2010 /Accepted: 18 May 2010# Springer-Verlag 2010
Abstract Major leaps forward in understanding rice bothin genetics and archaeology have taken place in the pastdecade or so—with the publication of full draft genomes forindica and japonica rice, on the one hand, and with thespread of systematic flotation and increased recovery ofarchaeological spikelet bases and other rice remains onearly sites in China, India and Southeast Asia. This paperwill sketch a framework that coherently integrates theevidence from these burgeoning fields. This frameworkimplies a reticulate framework in the phylogeny of earlycultivated rice, with multiple starts of cultivation (two isperhaps not enough) but with the key consolidations ofadaptations that must have been spread through hybrid-isation and therefore long-distance cultural contacts.
Archaeobotanical evidence allows us to document thegradual evolutionary process of domestication through ricespikelet bases and grain size change. Separate trends ingrain size change can be identified in India and China. Theearliest centre of rice domestication was in the Yangtzebasin of China, but a largely separate trajectory into ricecultivation can be traced in the Ganges plains of India.Intriguingly, contact-induced hybridisation is indicated for theearly development of indica in northern India, ca. 2000 BC.An updated synthesis of the interwoven patterns of thespread of various rice varieties throughout Asia and toMadagascar can be suggested in which rice reached most ofits historical range of important cultivation by the Iron Age.
Keywords Archaeology .Oryza sativa . Domestication .
Dispersal . Neolithic
The distribution of wild rice and genetic diversityin domesticated rices
Rice is a highly diversified crop, being grown from theequator to over 40° N, from sea-level ca. 2,700 m in partsof the Himalayas and in a wide ecological range ofcultivation systems. Although there is much less ecologicalvariation found within its wild progenitor complex (Oryzarufipogon and Oryza nivara), these are nonetheless distrib-uted over a wide geographical range and a spectrum ofecological niches from permanent to seasonal wetlands. Asthe origins of cultivation must have developed in placeswhere hunter–gatherers were utilising wild populations, thedistribution of the wild progenitor, in the past whencultivation began, is a key element in identifying theorigins of rice. There are three lines of approach to inferringthis without archaeological evidence, including (1) the
Electronic supplementary material The online version of this article(doi:10.1007/s12520-010-0035-y) contains supplementary material,which is available to authorized users.
D. Q. Fuller (*) : C. Castillo :A. R. Weisskopf :E. J. Kingwell-Banham : J. SongInstitute of Archaeology, University College London,London, UKe-mail: [email protected]
Y.-I. SatoResearch Institute for Humanity and Nature,Kyoto, Japan
L. QinSchool of Archaeology and Museology, Peking University,Beijing, China
S.-M. AhnWonkwang University,Iksan, South Korea
J. van EttenIE University,Segovia/Madrid, Spain
Archaeol Anthropol SciDOI 10.1007/s12520-010-0035-y

modern geographical and ecological distribution of wildrices, (2) an assessment of the likely effect of past climaticchanges on this distribution, and (3) phylogenetic relation-ships between various modern wild populations anddomesticated populations. There are complicating factors,however. Apparent genetic relationships between wild andcultivated populations may be affected by recent introgres-sion and hybridisation across lineages, as well as by deepershared histories. Thus, a careful comparison of comple-mentary and contrasting genetic datasets is necessary. Asfor geography, the modern distribution of wild rice mayhave been impacted not just by climatic change but alsohuman land-use as suitable habitats for wild rices weredestroyed or modified, for example by becoming cultivatedlandscapes. This can be accounted for to some extent byreference to ancient Chinese historical sources whichindicate the presence of wild rices about 1,000 years agoin regions where rice does not occur wild today.
The accepted wild progenitor complex for rice is that ofO. rufipogon sensu stricto, predominantly perennial, andthe annual O. nivara (Vaughan et al. 2005, 2008a, b), whichare distributed in South and Southeast Asia and parts ofChina (Fig. 1). An understanding of the past distribution ofwild rice must also take into account probable reports fromTang and Song Dynasty sources (eighth to tenth centuryAD; Ho 1977; You 1987). We follow You (1987), inseparating those which are from plausible primary habitats,such as lakes as opposed to those “wild” rices that sound
more like weedy types (“Oryza spontanea”) or late tillerscollected from cultivated areas in times of famine.
During periods of drier climate, such as Pleistocene glacialperiods, we expect that these wild populations would havebeen more restricted into wetter tropical refugia, in parts ofEastern India, Yunnan and mainland Southeast Asia (e.g.Fig. 2 P). This is consistent with the genetic evidence that theancestral haplotypes of O. rufipogon are found in SoutheastAsia and India (Londo et al. 2006). The most recentgeographic expansion and radiation of wild rice populationswould have started ca. 15000 BP (16500 cal. BC) as post-glacial climatic conditions set in. In addition to the changesin temperature and rainfall regimes, it should be noted thatthis period also saw a dramatic increase in atmospheric CO2
levels from 18 Pa (atmospheric partial pressure) to 27 Pa, by11000 BP, which is expected to have had a major impact onthe photosynthetic productivity and biomass production ofplants, but especially C3 plants like rice (Sage 1995;Cunniffe et al. 2008). We expect this period to havewitnessed a major expansion in the geographic range of ricetowards higher latitudes, especially in East Asia where anorthwards expansion was not blocked by the high Hima-layas. We might also expect some adaptive changes in ricemorphology in response to new climatic and atmosphericconditions and will return below to how this might confoundthe recognition of some domestication traits.
A massive geographical range expansion of wild Oryzaspp. took place between the dry conditions of the glacial
Fig. 1 Map of modern and historical distribution of rice’s wild progenitors, O. rufipogon and O. nivara
Archaeol Anthropol Sci

maximum, the warm and wet conditions of the TerminalPleistocene (Fig. 2 P) and early Holocene (Fig. 2 H). Dryseasons of 7 months or more are likely to precludecultivation of traditional late ripening rices and wild rices.As indicated by Chinese historical sources, the majoradoption of early-ripening rice (which can be grown in 4 to5 months), generally more tolerant of water shortage, tookplace from the eleventh century AD onwards (Ho 1956). Forthis reason, the climatic pattern devised by Huke (1982), tiedto unchanging topography, seems a suitable corrective to thereconstructed vegetation zones of pollen indicators (Adamsand Faure 1997; also, Shi et al. 1993; Ren and Beug 2002).This indicates that wild rice probably grew throughout theYangtze basin and eastern China as far north as Shandong,and thus, archaeobotanical finds of rice, without cleardomestication traits or indicators of cultivation, could relateto wild gathering. The potential of inferring cultivationthrough archaeological finds of rice beyond the range of thewild progenitors is thus quite limited, although finds fromnorthwest Pakistan from 2500 to 2000 BC qualify (Costantini1987), as does Early Yangshao rice from Nanjiaokou,northern Henan (Qin and Fuller 2009) and Late Yangshaorice from Xishanping, Gansu (Li et al. 2007).
Rice is divided into two main sub-species, indica andjaponica (e.g. Oka 1988; Vaughan et al. 2008b), although
numerous other varieties and sub-varieties are recognised.Recent genetic classifications (e.g. Garris et al. 2005;Caicedo et al. 2007; McNally et al. 2009) suggest thatthere are five analytically useful groups which can berecognised: indica, aus, temperate japonica, tropicaljaponica (including javanica) and fragrant, although mostdiscussions of origins have focussed on whether the indica(including aus) and japonica groups derived from a singleor multiple domestications. There is a now a wide range ofgenetic studies, using markers from the chloroplast genome,neutral variation in the nuclear genome and variation withinfunctional alleles that have been used to examine this issue.Chloroplast DNA has suggested two distinct lineages inboth wild and domesticated rice since a 69-bp deletion wasreported in the early 1990s (Chen et al. 1993). Nakamura etal. (1998) discovered an additional cpDNA polymorphismin a single sequence repeat (SSR) in the PS-ID region.Differences with respect to the number of C (cytosine) andA (adenine) bases arrayed in succession enable not onlydifferentiation of indica (8C8A) and japonica (6C7A) butalso identify variation amongst japonica (some are 7C6A)and differentiation of wild rice species (several additionalvariants; see also Ishikawa et al. 2002; Takahashi et al.2008). In addition, chloroplast genome types (both in PS-ID and 57 K sequence genome types) typical of indica are
Fig. 2 Map contrasting the expected Pleistocene (P) glacial refugiumdistribution of wild rice, based on tropical vegetation zones of Adamsand Faure (1997) and phytolith data from southern Poyang Lake basin(Zhao and Piperno 2000) and the Holocene (H) maximum riceexpansion associated with conditions of the Early and MiddleHolocene, based on agroclimatic zones of Huke (1982) adjusted for
early Holocene conditions but taking into account topographicconstraints like the Qinling Mountains, as well as vegetationreconstructions of Adams and Faure (1997) and Yu et al. (2000),with adjustments in India for more precise vegetation zones (cf.Asouti and Fuller 2008)
Archaeol Anthropol Sci

not represented in modern wild rice samples, suggestingthat the immediate maternal ancestor of indica has eithernot been sampled or has gone extinct (Kawakami et al.2007; Takahashi et al. 2008). More recently, a comparisonof whole chloroplast genomes for single representativejaponica and indica varieties showed a high degree ofdivergence with an age estimate for their common ancestorat 86,000–200,000 years (Tang et al. 2004).
A wide range of markers in the nuclear genome showsimilar patterns. Whole genome comparisons of represen-tative accessions for the nuclear genome suggest a similarlydeep divergence on the orders of 100,000 s of years (Vitteet al. 2004; Ma and Bennetzen 2004), whilst various genesor gene systems also point to differences such as pSINEs(Ohtsubo et al. 2004), SSR and STS datasets (Garris et al.2005; Caicedo et al. 2007), SAM and vTPASE sequencevariation (Londo et al. 2006) and ∼3,000 single nucleotidepolymorphisms assayed across 20 representative landraces(McNally et al. 2009). Whilst all of these studies agree onthe deep divergence of indica and japonica, they alsosuggest that there are some indicators of possible multiplederivations in both indica and japonica. These studies alsoshow hybridization between these lineages and mostrecently McNally et al. (2009) have provided evidence forquite frequent hybridisation between the major cultivarlineages. Much of this hybridisation may be historicallyrecent, for example taking place in China over the past800–900 years after early-ripening indica rices wereintroduced (cf. Ho 1956), or earlier around (1,500–2,500 years ago) in India and Persia when early historictrade connections were well established—the earliestarchaeological rice in the Near East is Parthian (first
century AD; Miller 1981), whilst imported rice is knownfrom Berenike on the red sea coat of Egypt (ca. 50 BC;Cappers 2006). Nevertheless, some hybridisation betweenearly japonica and indica lineages may have taken placenear the start of domestication, even when one lineage wasunder primitive pre-domestication cultivation (Fig. 3).Indeed current evidence for morphological domesticationtraits and what is known of associated genes implies asmuch (Sang and Ge 2007; Fuller and Qin 2009).
The domestication traits of rice: archaeobotanyand genetic lineages
Domestication refers to the key morphological (andgenetic) adaptations that characterise crops adapted tocultivation, traits that have often been characterised asan adaptive ‘syndrome’ of domestication (e.g. Harlan etal. 1973; Hammer 1984; Fuller 2007; Purugganan andFuller 2009; Fuller and Allaby 2010). These domesticationtraits evolved in a more or less gradual and piecemealprocess in response to the selection pressures created bythe human activities of cultivation. In other words,tracking the origins of rice requires us to try to identifythe behaviour of cultivation, then the appearance and riseto dominance of the domestication traits and finally thespread of reliance on rice agriculture. New appreciation ofthe drawn-out process of the evolution of domesticationtraits raises the likelihood that traits were selectedseparately in different regions and then brought togetherby processes of hybridisation/introgression (see Allaby2010).
Fig. 3 A simplified diagram ofthe main evolutionary pathwaysto domesticated Asian rice, withselected key mutationsindicated, differentiatingwild-type dominant form withall capital letters anddomesticated type recessivealleles with lowercase letters.Mutations: sh4 non-shattering,Prog1 erect growth, rc whitegrain pericarp, sw5 wider grains,qsh1 further non-shattering, wxywaxy/glutinous rice (lowamylase), WW wild genepool,dd domesticated genepool, cult.cultivated, X major hybridisationevent; crossed arrows indicatecontinued gene flow(introgression)
Archaeol Anthropol Sci
As with other cereals, non-shattering is a keydomestication trait which is likely to be deleterious inwild habitats. Geneticists identified six possible quanti-tative trait loci that appeared to be involved in this trait,with four that recur across studies (Xiong et al. 1999;Cai and Morishima 2002; Li et al. 2006a; Lin et al. 2007).Two of these have subsequently received much attentionprompting sequencing research. These are the recessivemutation sh4, which is a SNP in a gene on chromosome 4shared widely across japonica, fragrant, aus and indicarice (Li et al. 2006b, 2007) and qsh1 which is found onlyin some temperate japonica rices (Konishi et al. 2006) andappears to be derived from a haplotype found in some O.rufipogon. Most commentators have assumed that thewidespread sh4 was the major mutation involved in initialrice domestication (e.g. Sang and Ge 2007; Vaughan et al.2008a; Fuller et al. 2009). Fuller and Qin (2009) argued thatthis gene was introduced to proto-indica cultivars in Indiaafter japonica domestication in China, although recentlyZhang et al. (2009a, b) have suggested that this evolvedrapidly after domestication and then became widespread incultivated rices through gene flow (hybridisation; Zhanget al. 2009) therefore inferring that sh4 was not involvedin the initial evolution of non-shattering detected archae-ologically (e.g. at Tianluoshan). Later in a sub-group ofEast Asian rices, qsh1 became fixed during or after thebottleneck that led to differentiation of temperate japonica(Zhang et al. 2009). Unfortunately, little is known aboutmorphological differentiation of the spikelet bases of thesegenotypes when harvested and threshed by various
traditional means, so it is not yet possible to linkarchaeological remains to genotypes.
Archaeobotanists are interested in spikelet bases as clearindicators of whether individual rice specimens are ofdomesticated or wild type (e.g. Sato et al. 1991; Thompson1996, 1997; Sato 2002). There has been some confusion inthe use of this morphological indicator. The criteriaproposed by Zheng et al. (2007), in which there are twocategories, are not entirely consistent with those ofThompson (1996, 1997), and both present different ‘do-mestic’ morphotypes. Zheng et al. (2007) examined fourmodern populations, two wild and two domesticated;Thompson examined a few more modern populations.Based on a more extensive comparative study of wildmaterial than either of them (159 populations) and takinginto account the likely presence of immature spikelets,which is indicated in grain morphology as well as emptywaterlogged husks on some early Chinese sites, we nowhave a three category division (in addition to a fourthindeterminate category for poorly preserved specimens;Fuller and Qin 2008; Fuller et al. 2009). However, amongstthe domestic/non-shattering category, there is still a rangeof variation apparent which might relate to different non-shattering genetic loci (or combinations of loci) as well asto variation in the cultural practices of harvesting andthreshing tools/techniques. Further research is needed onthis topic.
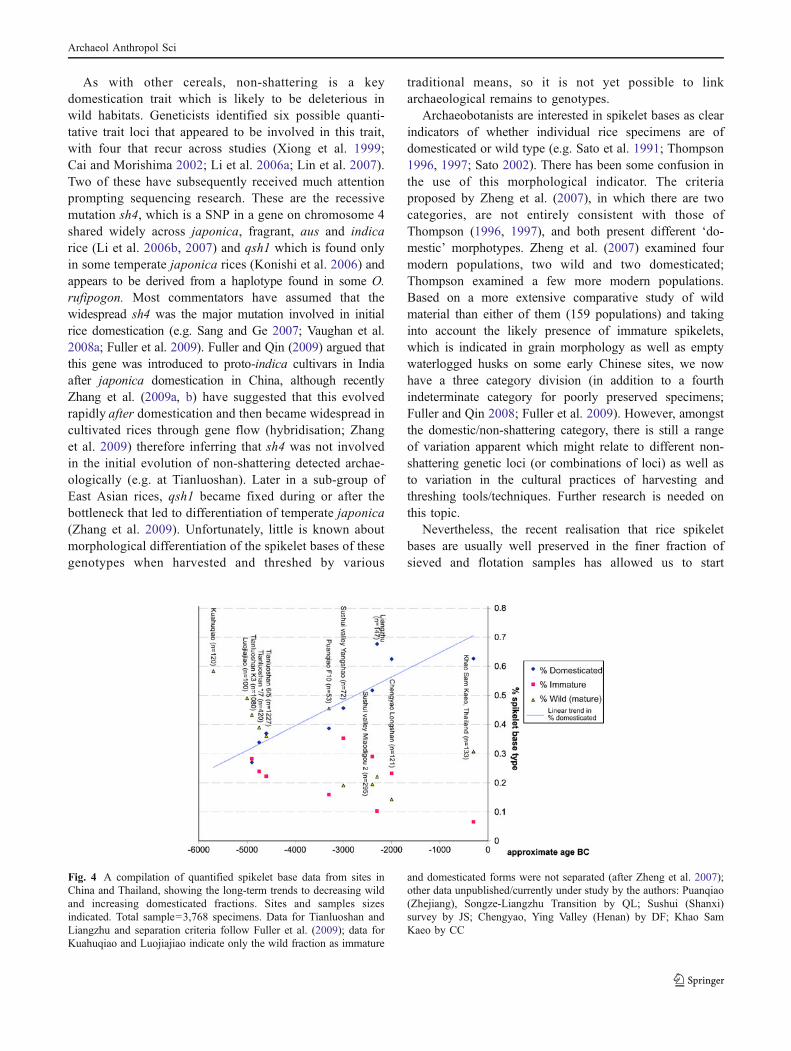
Nevertheless, the recent realisation that rice spikeletbases are usually well preserved in the finer fraction ofsieved and flotation samples has allowed us to start
Fig. 4 A compilation of quantified spikelet base data from sites inChina and Thailand, showing the long-term trends to decreasing wildand increasing domesticated fractions. Sites and samples sizesindicated. Total sample=3,768 specimens. Data for Tianluoshan andLiangzhu and separation criteria follow Fuller et al. (2009); data forKuahuqiao and Luojiajiao indicate only the wild fraction as immature
and domesticated forms were not separated (after Zheng et al. 2007);other data unpublished/currently under study by the authors: Puanqiao(Zhejiang), Songze-Liangzhu Transition by QL; Sushui (Shanxi)survey by JS; Chengyao, Ying Valley (Henan) by DF; Khao SamKaeo by CC
Archaeol Anthropol Sci

accumulating spikelet base data in recent years from sites inChina and Thailand (Fig. 4). Rechecking the fine fractionof previously sorted samples from Mahagara, India (Harveyet al. 2003; Harvey 2006) has also revealed spikelet bases.The largest assemblage yet studied is that from Tianluoshanin the Lower Yangtze region, which is now central todiscussions of the domestication process in the LowerYangtze (Fuller et al. 2009; Zhao 2010; Nakamura 2010).Unfortunately, we lack analogue population studies of theproportions of spikelet base types found in moderntraditional rice harvests, in which weedy rices and someimmatures should occur. The timing of rice harvests usuallybalances loss to incomplete grain maturation with earlierharvests (e.g. ∼30 days after harvesting) and loss due toshattering as panicles become ‘over-mature’ with laterharvests (e.g. ∼40 days after flowering; Horiuchi et al.1971). Some modern agronomic data from China wouldsuggest that levels of 5–10% immature spikelets might beexpected, affected in part by factors like water and diseasein a given season (cf. Pan 2008), but as indicated byethnographic and experimental data in Malaysia (Horiuchiet al. 1971), these are also affected by harvest time. Inaddition, we know little at present about how differingharvest and processing methods might bias spikelet baseassemblages that enter routine domestic refuse or seasonalthreshing waste.
The other domestication trait often considered inarchaeobotany is grain size (Fig. 5), although any use ofthis as an absolute indicator of domestication status onindividual specimens is problematic. It is generally true thatdomesticated grain crops have larger (often wider) seedsthan their wild ancestors, which is thought to be selected forby deeper burial in more disturbed, tilled soils (Harlan et al.1973; Heiser 1988; Fuller 2007). In rice, however, this traitis complicated by the great range of variation representedby modern cultivars and wild populations (Fig. 5a). Also,much variation in rice grain length can be linked to climaticvariation and ecological adaptations (Oka 1988; Kitano etal. 1993): More northerly temperate japonica landraces areshort-grained, whilst tropical varieties (the javanica racerices) are massively long. Complicating this trend is that oflonger-grained upland rices versus shorter-grained lowlandrices in East Asia generally (Nitsuma 1993), whilst somevarieties at high elevations in the tropical mountains (e.g.Yunnan, Nepal) may be very short-grained. The latitudinaltrend found in modern rices is indeed evident in the data ofNeolithic northern China (e.g. Liu et al. 2007; Qin andFuller 2009) and Bronze Age Korea (Ahn 2010), wheregrains are generally shorter than those from the Yangtze ofsimilar date. Seed size is affected by many factors,including environment, and is polygenically controlled(Fuller and Allaby 2010). For the most part, these genesand regulatory networks are poorly understood, but the
many QTL analyses that have been carried out have shownseed size to be associated with many genes of varyingeffect (Gupta et al. 2006). At least three loss-of-functionmutations have been identified in rice that directly influencegrain shape/size (Fan et al. 2006; Song et al. 2007;Shomura et al. 2008). In addition, mutation to the genespd6 appears to increase grain size as well as grain number(Shan et al. 2009). Two of these result in an increase in thenumber of glume cells, thereby giving the grain milk alarger cavity to fill, resulting in wider grains (Song et al.2007; Shomura et al. 2008). This shift in husk cellarchitecture is important as it offers an explanation of someof the variation documented amongst and between thedomesticated and wild rice husk phytoliths (Zhao et al.1998; Harvey 2006)—although there is so much variationand overlap amongst modern rices to make this anunreliable separator in many cases (Harvey 2006). Inaddition, this implies that ‘domestic’-type phytoliths foundin pre-agricultural Pleistocene cave sites (Zhao 1998, 2010)could be the product of shifts in climate and latitudinalvegetation belts in the post-glacial period.
In the archaeobotanical record of India and of theYangtze, one finds trends towards larger grain sizes overtime (Fig. 5b, c). However, this is also not the only trend:Grains from the Yellow River in China tend to be as smallor smaller than those in the Yangtze (Liu et al. 2007),meaning that this trait cannot be used in a straight forwardway to indicate domestication, but local trends of changemay be considered useful indicators. In India, by contrast,grain size increase appears to be dampened within theGanges core region by contrast to early grains in regionswithout wild rice. Whether this indicates a role for recurrentgene flow from wild populations or different selectionpressures in differing regional cultivation regimes (whichhas been argued for pulses: Fuller and Harvey 2006)requires further research.
Another classic domestication trait is apical dominance,reflected in reduced lateral branching or tillering that resultsin a more erect growth habit. The gene PROG1 which hasattracted particular attention in rice genetics recentlyappears to have a large effect on morphology when therecessive form is expressed (Tan et al. 2008; Jin et al.2008). The domestic-type prog1 sequence was sharedacross all 182 domestic lines tested, with a similar sequencefound in a subset of perennial O. rufipogon from SouthChina, Taiwan and Thailand (Tan et al. 2008). Whilst thissuggests that this trait is derived from variation that existedin the wild, wild bearers of the mutation do not express anerect growth habit indicating the countervailing effect ofother factor(s). Other genes that have been implicated inrice architecture include Moc1, MAX2 and MAX3, but asreviewed by Doust (2007), branching pattern is influencedalso by environmental conditions.
Archaeol Anthropol Sci

Chronological and geographical patternsin the archaeology of rice
Archaeobotanical evidence for the use of wild rice remainsquite limited, whilst the date of the start of cultivation of rice iscontroversial in both China and India. The locations ofPleistocene and Early Holocene reports are shown inFig. 6a, with site details in Table S2. A single report ofPleistocene rice from Xom Trai, Vietnam, at ca. 16000 BP, ispresumably wild (Glover and Higham 1996: 421). SomePleistocene rice finds are not directly associated with humanarchaeology (i.e. no associated artefacts) and might be
suggested to relate to wild populations, including phytolithsoff of the Shanghai coast (Lu et al. 2002) and perhaps theinfamous rice husks of the palaeosol (peat) deposit at Sororiin Korea (Lee and Woo 2006; Kim et al. 2008; however, thelatter may represent later Holocene intrusive material: seecritical comments in Ahn 2010). Interestingly, both sites dateto ca. 13000 BP uncalibrated or ca. 13000 cal. BC which isafter a period of rapid global warming and CO2 increase thatshould be expected to have extended the wild range of ricenorthwards, perhaps even further north than in the EarlyHolocene. A few finds of rice from the Late Pleistocenearchaeological occupation sites in China are presumably
Fig. 5 Grain size data.a Modern ranges based onmeasured populations in UCLcollection (same as thosereported in Fuller et al. 2007)and full range of extrememodern (reported in Vaughan etal. 2008a). b Grain width ofarchaeological site populations(mean and standard deviation),plotted against time in the LowerYangtze, with trend line throughaverages shown. c Grain widthof archaeological site populations(mean and standard deviation),plotted against time in northernIndia, with trend line throughaverages shown. Forarchaeological detailson archaeological data,see Table S1
Archaeol Anthropol Sci

from wild use, such as Yuchanyan, Xianrendong andDiaotonghuan (Lu 2006; Zhang and Hung 2008). Despitethe suggestion from phytoliths that they document a shifttowards domesticated morphology in husk cells (Zhao 1998,2010), we would suggest that this might be mimicked byrice’s natural response to rapid post-glacial climatic change.What the shift in husk phytoliths suggests is a shift in grainsize, which is just as likely the result of adaptation to climaticchanges at ca. 15000 cal. BC, the date of the phytolith shiftbased on the association with early ceramics that are nowwell dated to this time (see Kuzmin 2006; Zhang and Hung2008; Boaretto et al. 2009).
Setting aside Pleistocene finds, we are still faced withwhether Early Holocene finds of rice indicate foraging orcultivation. In the Lower Yangtze region, the earliest evidencefor rice dates between 10000 and 8000 BC, especially at thesite of Shangshan (Jiang and Liu 2006; Nakamura 2010; Zhao2010), but evidence documents neither clear cultivationbehaviour nor morphological domestication. Two spikelet
bases could be isolated, which include one unambiguouswild type and one immature type (cf. Zheng et al. 2007; butcriteria follow Fuller et al. 2009), and thus, there are noconfirmed domesticated types and a uselessly small samplesize. In the middle Yangtze, ceramics with husk temper fromthe site of Pengtoushan have direct AMS dates back to8000–7000 BC, whilst the culturally related site of Bashi-dang (7000–6000 BC) produced 1,000 s of rice grains,within an overall broad spectrum assemblage, but withoutany good evidence for domestication traits one way or theother (e.g. HPIACR 2006). It should be noted that theconventional view assumed that the earliest rice in Chinawas domesticated indica (e.g. at Hemudu, Bashidang) or‘intermediate’ between indica and japonica and evolved intojaponica over the course of the Neolithic (e.g. Zhang andWang 1998; Zhang 2002). Such a model is contrary to whatwould be expected based on current genetic knowledge (e.g.Sato 2002; Sweeney and McCouch 2007), and morphomet-ric data on leaf bulliform phytoliths indicate that these are
Fig. 6 The chronologicalspread of rice in two maps and11 ‘thrusts’. Sites plotted inbroad time horizons. Arrows andnumbers refer to the numberedthrusts referred to in the text.For details of the sites plotted,see Table S2
Archaeol Anthropol Sci

consistently of the japonica type (or its wild ancestor; e.g.Sato et al. 1991; Fujiwara 1993; Zheng et al. 2003).
The situation is similar in India, where well-dated earlyfinds come only from Lahuradewa in the middle Gangesregion, which has ceramics and charred rice remains datingback to 6000–8000 BC (Tewari et al. 2008). Although thelatest report claims to have ‘domesticated’ rice on the basisof grain L/W/Th ratios and spikelet bases, the reportedmeasurements and illustrated spikelet bases actually con-tradict these claims, with the grain ratios, and absolutemeasurements clearly overlapping with O. nivara and O.rufipogon and one population with Oryza officinalis (fordetails, see Fuller 2009). Archaeological grain metrics forthe region suggest that a marked increase in rice grainbreadth occurred after 2000 BC continuing through the firstmillennium BC (Fig. 5c). Rice bulliforms from the nearbyLahuradewa Lake pollen core have been suggested toinclude domesticated types, although the original method-ological study for the criteria used—the number of fan-edgechips (Lu et al. 2002)—would imply domesticated ricesalready introduced into the Lower Yangtze by 13000 BP,which is so at odds with the archaeobotanical evidence(from habitation sites, spikelet bases etc.) that this criterionmust be regarded with suspicion. Rather, as alreadysuggested, the Lower Yangtze sea core phytoliths suggestthe colonisation of the region by wild rice with post-glacialwarming, and this thus undermines fan-edge bulliformarchitecture as a domestication trait.
Based on unambiguous morphological domesticationtraits, we can recognise the presence of domesticated ricefrom 6000 cal. BC in the Lower Yangtze and from ca.2000 cal. BC in the Ganges. In the Lower Yangtze,Kuahuqiao (6000–5400 BC) is reported to have a majorityof mature wild-type spikelet bases (Zheng et al. 2007),whilst insufficient criteria make it unclear how the restdivide between immature and domesticated types. AtTianluoshan, it is clearer that a large minority of domesti-cated types in the earliest samples (ca. 4900 BC) increaseover the subsequent three centuries to outnumber maturewild types (39:31%), whilst immature types also decrease(Fuller et al. 2009). By the time of the proto-urbancivilisation of Liangzhu, this ratio had tipped to 68:25 infavour of domesticates. It is probable that 20–25% wildtypes can be derived from weedy forms of rice that havepersisted alongside domesticates. Indeed, evidence fromLate Yangshao Shanxi (Sushui valley survey) and the YingValley (Chengyao) show similar proportions of wild typesand similarly reduced proportions of immature types (5–10%). In the Ganges, data are only just starting to beaccumulated but so far the few dozen spikelet basesrecovered from Mahagara are almost entirely domesticated,as are those from early historic sites in the South like Terrand Paithan. In the latter cases, this may indicate that the
cultivation regime (dry cropping during the monsoon?) wasone where weedy rices are less of a problem. By contrast inIron Age southern Thailand, represented by Khao SamKaeo, associated with an upland dry-rice weed flora, a highproportion of wild-type spikelet bases suggest a highpresence of weedy types and probable continuing intro-gression. Nevertheless, domesticated morphotypes werepresent in Neolithic Thailand, by ca. 2000 BC, where 27spikelet bases recovered from coprolites at Khok PhanomDi are said to be dominated by domesticated types(Thompson 1996).
Elsewhere, our record of early rice is limited merely todata on presence and sometimes quantitative importance incomparison to other crops. Nevertheless, we can sketch thebroad patterns of the dispersal of rice cultivation. Dispersalscan be divided into 11 distinct thrusts, which are reviewedbelow. These thrusts are numbered below and on the arrowsin Figs. 6a, b. The evidential basis of these 11 regional/chronological units is summarised through a list of sites inthe on-line supplementary material (Table S2).
The spread of rice in 11 thrusts
1. The northern thrust and early temperate japonica
Rice is found in the middle Yellow River, separated by theQinling Mountains from the wild rices of the Yangtzewatershed, from about 6,000 years ago, and much moreextensively by 5,000 years ago. For Yangshao or Dawenkousites, 16 have apparently reliable reports of rice, whilst for theLongshan period (2500–1900 BC, including Miaodigou 2),this tally doubles to 33 (Fig. 6a). Recent systematic samplingby flotation during excavations and regional surveys hasbeen extensive and accounts for the majority of thisevidence, including numerous direct AMS dates on rice orother crop grains. Typically, rice is a minor componentalongside millets. Setaria italica tends to dominate thearchaeobotanical evidence whereas second place is takenby Panicum miliaceum or rice. This is true of the earliestfinds from Nanjiaokou from Early Yangshao cultural levels(from 4500 BC, with direct AMS on rice of ca. 3800 BC;Qin and Fuller 2009). In the subsequent Middle Yangshaoperiod (3800–3000 BC), Nanjiaokou and several other siteshave produced rice, and it is fairly widespread by 3000 BC.Geoarchaeological and phytolith evidence from the YiluoValley suggest that rice agriculture in this region was carriedout in wetland fields and was facilitated by a period of activeriver alluviation and hillslope erosion, which created suitablelow-lying environments for irrigated rice (Rosen 2008).Available grain metrics indicate notably short-grained formsof rice, consistent with a temperate japonica lineage. After apeak of rice farming (by number of sites) in the Longshan
Archaeol Anthropol Sci

period, rice finds decrease into the Bronze Age with the riseof the Shang state, which may relate to the climaticaridification after 2200–2000 BC. Those varieties whichpersisted must have been adapted to more northerlyconditions. This reduction in rice land should also be seenin relation to the introduction of bread wheat and in someareas barley, at about this time from Central Asia (Li et al.2007; Lee et al. 2007; Fuller and Zhang 2007). After thisperiod, wheat and rice took their place alongside the milletsand soybean as the five grains of classical Chinesecivilisation (Chang 1980).
2. An earlier Jomon and Chulmun upland japonica?
There is a hypothesis that rice cultivation was introducedto Japan already during the late Jomon period prior to2000 BC and certainly by 1000 BC (e.g. Sato 1996;D'Andrea 1999). Evidence for this remains limited andmainly comes from phytoliths from Jomon site sedimentsand ceramic fabrics with rice inclusions from Kyushu (e.g.Fujio 2004; Takahashi 2009; Toyama 2002: Table 2).Whilst the presence of rice in pre-Yayoi cultural contextsseems clear, for sites in Kyushu (e.g. Uenoihara, Ooishi,Fukuda, Minami Mizote), some commentators put the dateof these finds only marginally earlier than 1000 BC (e.g.Keally 2004). The earliest direct AMS dates on grains comefrom terminal Jomon Kazahari far to the north in Aomoriprefecture, with dates of ca. 900–700 BC. The presence ofrice in this era may also be indicated in South Korea,although data are limited and ambiguous (Ahn 2010). Thedatabase for South Korea is quite extensive (56 sites) bycomparison to its land area, but the vast majority ofevidence dates to after 1000 BC, and all earlier finds arerather controversial. Unambiguous rice finds date to after2000 BC—based on the earliest direct rice AMS date atOun 1, although the wide error margin could be consistentwith 1500–1450 BC (Crawford and Lee 2003). This earlyrice may have been grown in dry field systems, whilst thelater Mumun is generally associated with paddy fieldagriculture (Ahn 2010), like the Yayoi in Japan (Imamura1996; Kuraku 2002; Takahashi 2009).
This Jomon rice has been interpreted as a dry field crop,differentiated from the Yayoi paddy field rice, because theJomon sites were located in upland areas. Whether thisimplies photo-period insensitive, early-ripening varieties isnot clear. It has been suggested that these rices would haverepresented an early and primitive form of tropical japonicacultivars, distinct from the temperate japonica paddy riceintroduced in the Yayoi period (Sato 1996; cf. Takamiya2001: 216). Ancient DNA analyses of Yayoi and later riceremains suggest a mixture of temperate and tropicaljaponica types which would be consistent with anintroduced Yayoi temperate paddy rice over a substratumof earlier tropical japonica (Sato 1996; Tanaka et al. 2010).
If dates for rice in Japan prove to be truly earlier than thosein Korea, then diffusion directly from the Lower Yangtze/Shandong coast area would seem to be indicated. However,uncertainties over Chulmun rice reports in Korea (see Ahn2010) mean that there may have been an earlier, dry-cropped, form of rice cultivation that spread from theShandong peninsula to Korea and Japan earlier than themore obvious paddy field diffusion wave of the Mumunand Yayoi period. Archaeological evidence for upland ricein China at this period is so far limited, although the weedflora of Liangchenzheng in Shandong (2500–1800 BC)could be consistent with rain-fed cultivation (Crawford etal. 2005). In addition to systematic archaeobotany anddirect dating in Korea and Japan from this era, further workaimed at understanding the ecology of rice, e.g. throughweed flora, is needed in China.
3. To Taiwan and beyond: a southerly quasi-temperatejaponica
Rice appears in Taiwan along with foxtail millet in the earlythird millennium BC (Tsang 2005), presumably with thespread of agriculture from mainland China. Hard data arelimited, with five Neolithic sites having reported rice andnone of them with a full, systematic archaeobotanical study.The interesting co-occurrence of rice and millet together withcultural practices, like front tooth removal, and apparentlyshared early vocabulary may suggest a derivation from thesouthern Shandong coast/northern Jiangsu (Sagart 2008)rather than the Lower Yangtze area (postulated by Bellwood1997, 2005), as millet and rice are unknown together inLower Yangtze archaeobotany. Although quantitative dataare unavailable, Tsang’s (2005) photograph suggests a short-grained rice morphotype, like early grains from Shandongand the Yellow River basin.
From Taiwan, rice is expected to have spread furthersouth into island Southeast Asia, although hard evidence isscarce (Paz 2003; Barton and Paz 2007), with one Neolithicsite in the Philippines, three in Borneo, two in Bali and onein Sulawesi. Half of these are based on husk impressions inpottery only. Though some of these may be as early as2000 BC, there are few morphological details available.This may indicate that the Austronesian-associated ceramicNeolithic of the islands although linked with some riceagriculture in its early stages was mainly driven by otherforms of subsistence (Bellwood 1997).
4. Proto-indica origins and spread
Although rice use is established by around 7500 BC atthe site of Lahuradewa (Tewari et al. 2006, 2008; Saxena etal. 2006), whether this rice was cultivated or gathered wildremains controversial (see reviews in Fuller 2006; Fullerand Qin 2009). Later evidence from around 2500–2000 BCis stronger which suggests that rice was an established
Archaeol Anthropol Sci

cultivar and staple food and for some sites the only crop inthis region. On this basis, archaeologists working in Indiacontend that rice was separately brought into cultivation inthe Ganges region, although good evidence for the processof morphological domestication is unclear. Spikelet baseremains have not been reported from India, although recentwork by one of us (DF) has recovered domesticatedspikelet bases from Mahagara from between 1800 and1600 BC. If it is assumed that domestication was controlledby the sh4 non-shattering mutation, then this evidenceprovides the minimum age by which the non-shattering traitwas introduced into ancestral indica. Alternatively, wecould suggest that a different locus was involved in earlynon-shattering in indica and sh4 was introduced later butbecame universal as it was in some ways more effective.
What is clear from the evidence at present is the end of theprocess of agricultural origins in this region, as rice/othercereal/pulse-cultivating sedentary villages with domesticlivestock become widely established from 2000 BC (Fuller2006). Crop, livestock and ceramic forms point towards linksto the upper Indus/Ravi River (Harappan) region to the west.This may indicate that an intensive and expansive agricul-tural economy focussed on rice did not develop prior to theinput of other crops, including winter crops adopted from theWest. One reason for this may be that domesticated japonicarice was needed to introduce some key domestication alleles,including prog1 (erect growth), sh4 (non-shattering) and rc(white pericarp; Kovach et al. 2007; Jin et al. 2008; Tan et al.2008). As indicated in Fig. 4c, grain size increase (thicken-ing) is indicated only after 2000 BC through the Chalcolithicand Iron Age periods.
5. Tropical japonica from South China to Southeast Asia
Despite a tally of 67 sites/phases in mainland SoutheastAsia with reported rice (from Neolithic through Iron Agetimes), 66% consist only of the casual report of rice-tempered pottery, with only 17 sites in Thailand having hadsystematic sampling. Southern China, including Guangxi,Guangdong and Hong Kong, is still data poor (Lu 2009):Six sites plus two more from southern Hunan have reportsof rice, with little published in any detail (Fig. 6b). Thismeans that we can sketch some broad chronologicalpatterns (see Rispoli 2008; Zhang and Hung 2008), butonly for a few sites in Thailand can we attempt to saysomething more about the local significance of rice in thediet, or cropping ecology. Broadly speaking, in mainlandSoutheast Asia, rice appears by 2000 BC, especiallyassociated with the lower reaches of the Red River(Vietnam), the Mekong River (Cambodia) and Chao Phraya(Thailand). Interior finds up river are at present consistentlylater (perhaps mainly from 1500 BC, see e.g. Higham andHigham 2009; cf. Vincent 2003). This argues against theHigham hypothesis (e.g. Higham 2003) of a dispersal down
the spoke-like rivers from Yunnan, but rather a morecoastally focussed, or perhaps on the lower mountain slopezones, zonal dispersal from southern China (Guangdong).Recent assessment of ceramic complexes by Rispoli (2008)similarly points to links from the middle Yangtze toGuangdong and Guangxi (see also Lu 2009, Zhang andHung 2010) and onwards to Vietnam and mainlandSoutheast Asia. This revised view would fit with a processthat saw the development of true tropical japonicas of thelowlands and lower slopes, with an ecology based onrainfall or natural flooding (as suggested by White 1995).Modern genetics suggests that this trajectory saw the earlydevelopment of true glutinous (waxy; cf. Olsen andPurugganan 2002) and fragrant (badh2.1) varieties of rice(cf. Kovach et al. 2009). Also of note is the evidence thatfoxtail millet was a major crop within this trajectory datingto the Neolithic and Bronze Age in mainland SoutheastAsia (Weber et al. 2010), tracing its roots to northern China(Lee et al. 2007), via the middle Yangtze, where it isreported from Chengtoushan at ca. 4500 BC (Nasu et al.2007), via Gantouyan near the Guangxi–Vietnam borderwith both rice and foxtail millet by 3000–2000 BC (Lu2009). Economically and ecologically, rice may not havebeen any more significant than foxtail millet, but diversi-fication of its cooking qualities suggests that it hadimportant cultural or aesthetic significance.
6. Carry on up the Yangtze
Rice cultivation also presumably spread up the YangtzeRiver into the Sichuan basin and to Yunnan (Fig. 6b). Riceis known in the Chegdu plain from Jinsha during theBaodun period, 2500–1800 BC (Zhao, Zhijun, personalcommunication.) In Yunnan, rice is reported from Neolithicsites that date to the third millennium BC, such as atBaiyangcun, ca. 2100 BC (An 1999). However, in bothprovinces, archaeobotanical sampling has been rare. It isalso worth noting that, in both areas, millet(s) occur as wellas rice and the evidence from Karuo in the mountains ofQamdo, eastern Tibet puts foxtail millet there by 3000 BC(An 1999) and might indicate that foxtail millet cultivationin mountain areas was established centuries to a millenniumearlier than rice cultivation.
7. The introduction of japonica genes and the makingof indica
Recent genetic evidence has confirmed the model of Sato(1996) that indica is of hybrid origin including nuclear genesfrom domesticated japonica, although alternative scenariosmay be conceived for the initial meeting of proto-indica,either as wild populations or as a cultivar under pre-domestication cultivation (Sang and Ge 2007; Vaughan etal. 2008b; Zhang et al. 2009; Fuller and Qin 2009). Whilstdirect dispersal from the upper Yangtze via Assam is
Archaeol Anthropol Sci

proposed by some and would fit with hypotheses that deriverice cultivation from the dispersal of farmers in theAustroasiatic language family (e.g. Higham 2003; Bellwood2005), current archaeobotanical evidence for rice does notprovide support for the model. As noted above, rice use, ifnot cultivation, has a millennia long history in the Ganges,and rice, of unclear domestication status, seems to becomewidespread in the upper Ganges and upper Indus region by2500 BC. It is only at this same time horizon that ricecultivation appears to have spread to the upper Yangtze inboth Sichuan and Yunnan, leaving a large geographical gapto be covered in no time at all. In addition, historicallinguistic data from Indian Austroasiatic (Munda) languagesand archaeological evidence from eastern India point towardsan even later arrival (ca. 2000 BC), after the dispersal ofIndian pulse and western animal domesticates (Fuller 2003).
On the other hand, evidence for several cultivarsoriginating in China occurs widely in Late Harappan timesin northwest India and Pakistan (Fuller 2006: 36; Fuller andQin 2009). The earliest plausible japonica rice grains withshort and wide grains is reported from the Swat valley andBaluchistan (see Costantini 1979, 1987), from ca. 1900 BC.In addition, there is a list of other crops of Chinese originthat occur for the first time in South Asia in the northwest,including P. miliaceum (see Jarrige 1985; Fuller and Boivin2009), apricots, peaches and Cannabis. Also found are thedistinctive stone harvesting knives of a form that must havebeen adopted from China into Northern Pakistan andKashmir (Sharma 1982: Stacul 1992). It can therefore besuggested that domesticated japonica followed other cropsand tools westward and then south to India, via a precursorof the Silk Road. Once japonica rice was availablealongside local, unimproved proto-indica cultivars, hybrid-isation and back-crossing could rapidly improve crops inthe active selection regimes of farmers.
By contrast, if we set rice aside, the earliest plausibleintroductions from the northeast to the Ganges are later,including the fibre crop ramie (Boehmeria nivea), a citrusfruit (Citrus medica sensu lato) and mango, which occur allafter ca. 1400 BC (on mango and citron, see Asouti andFuller 2008: 91, 114–115; on ramie, see Saraswat et al. 1994:287).
8. The dispersal of rice through South Asia
The sub-continental record for rice (Fig. 6) indicates thediffusion of this species from the Ganges region and/orEastern India (Orissa) starting in the late third millenniumand an important presence of rice cultivation on thepeninsula only in the first millennium BC (Fuller 2002).Outside of a Ganges core, rice occurs first in parts of thegreater Indus valley and perhaps in parts of Gujarat andRajasthan, although this may have also involved introducedjaponica rice or hybrids thereof. It is not clear if any of this
early rice cultivation was based on paddy field systems orwas merely supported by seasonal floods or monsoon rains.With development of paddy-like systems in the Ganges,however, a major southwards push along the east coast toTamil Nadu occurred in Late Iron Age times (Fuller andQin 2009). In this same general period, indica rice movedprobably by overseas trade to Southeast Asia and SriLanka.
9. The spread of paddy field cultivation through Koreaand Japan
Setting aside the problematic report of Oryzoid remainsat Late Pleistocene Sorori and debates over the reliability ofChulmun finds of around 2000 BC (see Ahn 2010), there isno doubt that rice was widely cultivated in Korea by theend of the second millennium BC. Based on materialculture evidence, rice, including wetfield cultivation, isinferred to spread from the Shandong Peninsula to theLiaoning peninsula and to Korea (Norton 2007; Ahn 2010).Most early rice dates are associated with the Early Mumunperiod which starts from ca. 1200 BC. Mumun sites havealso yielded apparent paddy field systems, although dryfields are also reported, and it seems likely that both rain-fed and irrigated rice were part of the agricultural repertoireof Mumun Korea (Ahn 2010).
It is broadly accepted that the introduction of paddy fieldrice farming into Japan came from Korea at the transition tothe Yayoi period, between 1000 and 500 BC. The presence ofrice paddy agriculture in northern Kyushu, western Japanduring the early first millennium BC is well documented (e.g.Imamura 1996; Kuraku 2002; Hudson 2003; Takahashi2009). The database from Japan is extensive and cannot befully reviewed here, although Takahashi (2009) maps 29sites with early paddy field evidence. Many of these haveincluded geoarchaeology, phytolith, diatom and insectstudies (e.g. Mori 2002; Kuraku 2002).
10. Further Asian spreads and crossings in northwesternSoutheast Asia
There was necessarily further dispersal from several ofthe above areas: westwards from Indo-China, south/west-wards from Yunnan and eastwards from the Gangestowards a ‘melting-pot’ ecologically and culturally inBangladesh, Assam and western Burma. This mixing anddiversification process may account for the highly hybri-dised but often locally divergent nature of aus rices, whichrecent genetic data suggest to have higher levels of genomichybridisation than other indica rice or japonica (McNally etal. 2009). In much of modern Southeast Asia, tropicaljaponica cultivation is focussed in upland areas, whereasindica is grown in the lowlands. The origins of this patternare unclear, but it is of interest that evidence from MetalAge Khao Sam Kaeo (400–0 BC) on the Malay Peninsula
Archaeol Anthropol Sci

is suggestive of dry, upland cultivation (Castillo, unpub-lished data). By contrast, the expansion of urban politiesacross the plains of central Thailand in the early centuriesAD culminating the Dvaravati period with dense distribu-tions of moated settlements is attributed to intensive,lowland (and presumably paddy field) cultivation (Mudar1999), which we might expect to represent indica.
11. Westward dispersals: Persia and Madagascar
During the early centuries AD, rice was dispersed furtherwestward. From India, rice moved towards western Asiathrough Persia. The only datum for placing rice in WesternAsia is Parthian evidence (first century AD) from Susa inIran (Miller 1981). This northwestern dispersal presumablyincluded some of the aromatic rices, which have theirorigins mainly from tropical japonica rices (Kovach et al.2009), although some hybridisations with indica wouldhave occurred (Jain et al. 2004). Whether or not moretypical indica rices accompanied this spread is unknown.Meanwhile, from Island Southeast Asia, rice was carried byAustronesian ancestors of the Malagasy to Madagascar. Thedate of the initial settlement of Madagascar by Asianimmigrants remains controversial, with archaeologicalevidence for human presence as old as the first centuryAD, whilst linguistics arguments suggest a fifth century ADor later date (Dewar and Wright 1993). Linguisticsnormally derives this migration from Southwest Borneo.Modern rices in Madagascar include both tropical japonicaand indica types, and it is plausible that by the earlycenturies AD, both lineages were present in Southeast Asia(as indicated above for #9).
Discussion: towards a framework of ecologicaltrajectories and cultural preferences in early rice
Asian rices have had a complex history. Genetic researchand archaeobotany have the tools to provide an increasingresolution to this history. Genetics provides access toidentifying some of the key genes in the evolution ofdomesticated rice, where they are distributed and are likelyto originate, as well as the phylogenetic and reticulaterelationships of rice lineages. Hitherto, most geneticallybased models of rice origins, with one or two origins, andstraight forward outward dispersals have been too simple toaccount for the data now available: deep neutral genomedivergence, large numbers of shared selected alleles andwidespread evidence for hybridisation between lineages.Similarly, archaeologists have been too quick to pigeonholerice as domesticated or wild, indica or japonica, and havetaken less account of the evolutionary dynamics andprocesses by which domestication traits emerged at thelevel of populations in response to human activities, or how
the modern varietal groupings of indica and japonica arethemselves emergent outcomes of evolutionary processes.This review has attempted to sketch an integrated synthesis,although gaps in the evidence have had to be filled withconjecture. Nevertheless, a series of working hypotheseshave emerged which may structure further research.
References
Adams JM, Faure H (1997) Preliminary vegetation maps of the worldsince the last glacial maximum: an aid to archaeologicalunderstanding. J Archaeol Sci 24:623–647
Ahn S (2010) The emergence of rice agriculture in Korea: archae-obotanical perspectives. Archaeol Anthropol Sci 2(2).doi:10.1007/s12520-010-0029-9
Allaby R (2010) Integrating the processes in the evolutionary systemof domestication. J Exp Bot 61:935–944. doi:10.1093/jxb/erp382
An Z (1999) The origin and eastward spread or rice-planting culture ofChina. Wenwu 2:63–70, In Chinese
Asouti E, Fuller DQ (2008) Trees and woodlands of South India:archaeological perspectives. Left Coast, Walnut Creel, Retrievedfrom http://www.amazon.co.uk/Trees-Woodlands-South-India-Archaeological/dp/1598742310
Barton H, Paz V (2007) Subterranean diets in the tropical rain forestsof Sarawak, Malaysia. In: Denham T, Iriarte J, Vrydaghs L (eds)Rethinking agriculture. Archaeological and ethnoarchaeologicalperspectives. Left Coast, Walnut Creek, pp 50–77
Bellwood P (1997) Prehistory of the Indo-Malaysian Archipelago(Second rev.). University of Hawaii Press, Honolulu
Bellwood P (2005) First farmers: the origins of agricultural societies.Blackwell, Oxford
Boaretto E, Wu X, Yuan J, Bar-Yosef O, Chu V, Pan Y, Liu K, CohenD, Jiao T, Li S, Gu H, Goldberg P, Weiner S (2009) Radiocarbondating of charcoal and bone collagen associated with earlypottery at Yuchanyan Cave, Hunan Province, China. Proc NatlAcad Sci 106(24):9595–9600
Cai HW, Morishima H (2002) QTL clusters reflect characterassociations in wild and cultivated rice. Theor Appl Genet104:1217–1228
Caicedo AL, Williamson SH, Hernandez RD, Boyko A, Fiedel-AlonA, York TL, Polato NR, Olsen KM, Nielsen R, McCouch SR,Bustamante CD, Purugganan MD (2007) Genome-wide patternsof nucleotide polymorphism in domesticated rice. PLoS Genetics3(9):e163. doi:10.1371/journal.pgen.0030163
Cappers R (2006) Roman foodprints at Berenike: archaeobotanicalevidence of subsistence and trade in the Eastern Desert of Egypt.Costen Institute of Archaeology, UCLA, Los Angeles
Chang K (1980) Shang civilization. Yale University Press, NewHaven
Chen W, Nakamura I, Sato Y, Nakai H (1993) Distribution of deletiontype in cpDNA of cultivated and wild rice. Jpn J Genet 68:597–603
Costantini L (1979) Plant remains at Pirak. In: Jarrige J, Santoni M(eds) Fouilles de Pirak, vol. 1. Diffusion de Boccard, Paris, pp326–333
Costantini L (1987) Appendix B. Vegetal remains. In: Stacul G (ed)Prehistoric and protohistoric Swat, Pakistan. Instituto Italiano peril Medio ed Estremo Orientale, Rome, pp 155–165
Crawford GW, Lee G (2003) Agricultural origins in the KoreanPeninsula. Antiquity 77(295):87–95
Crawford G, Underhill A, Zhao Z, Lee G, Feinman G, Nicholas L,Luan F, Yu H, Fang H, Cai F (2005) Late Neolithic plant remains
Archaeol Anthropol Sci

from Northern China: preliminary results from Liangchengzhen,Shandong. Curr Anthropol 46(2):309–317
Cunniffe J, Osborne CP, Ripley BS, Charles M, Jones G (2008)Response of wild C4 crop progenitors to subambient CO2highlights a possible role in the origin of agriculture. GlobChang Biol 14:576–587
D'Andrea AC (1999) The dispersal of domesticated plants into north-eastern Japan. In: Gosden C, Hather JG (eds) The prehistory offood. Appetites for change. Routledge, London, pp 166–183
Dewar RE, Wright HT (1993) The culture history of Madagascar. JWorld Prehist 7(4):417–466
Doust AN (2007) Architectural evolution and its implications fordomestication in grasses. Ann Bot 100(5):941–950
Fan C, Xing Y, Mao H, Lu T, Han B, Xu C, Li X, Zhang Q (2006)GS3, a major QTL for grain length and weight and minor QTLfor grain width and thickness in rice, encodes a putativetransmembrane protein. Theor Appl Genet 112:1164–1171
Fujio S (2004) The beginning of agriculture in the Japanese islands.In: Society of Archaeological Studies (ed) Cultural diversity andthe archaeology of the 21st century. Society of ArchaeologicalStudies, Okayama, pp 62–73
Fujiwara H (1993) Research into the history of rice cultivation usingplant opal analysis. In: Pearsall DM, Piperno DR (eds) Currentresearch in phytolith analysis: applications in archaeologyand palaeoecology. MASCA Research Papers in Science andArchaeology, volume 10. University of Pennsylvania, Philadelphia,pp 160–174
Fuller DQ (2002) Fifty years of archaeobotanical studies in India:laying a solid foundation. In: Settar S, Korisettar R, Settar S,Korisettar R (eds) Indian archaeology in retrospect, volume III.Archaeology and interactive disciplines. Manohar, New Delhi, pp247–364
Fuller DQ (2003) An agricultural perspective on Dravidian historicallinguistics: archaeological crop packages, livestock and Dravid-ian crop vocabulary. In: Bellwood P, Renfrew C (eds) Examiningthe farming/language dispersal hypothesis. Mc Donald Institutefor Archaeological Research, Cambridge, pp 191–214
Fuller DQ (2006) Agricultural origins and frontiers in South Asia: aworking synthesis. J World Prehist 20(1):1–86
Fuller DQ (2007) Contrasting patterns in crop domestication anddomestication rates: recent archaeobotanical insights from theOld World. Ann Bot 100(5):903–924
Fuller DQ (2009) Indian archaeobotany watch: Lahuradewa 2008. 25June 2009 Web-log, retrieved from the internet: http://archaeobotanist.blogspot.com/2009/06/indian-archaeobotany-watch-lahuradewa.html
Fuller DQ, Allaby RG (2010) Seed dispersal and crop domestication:shattering, germination and seasonality in evolution under cultiva-tion. In: Østergaard L (ed) Fruit development and seed dispersal,annual plant reviews 38. Wiley-Blackwell, Oxford, pp 238–295
Fuller DQ, Boivin NL (2009) Crops, cattle and commensals across theIndian Ocean: current and potential archaeobiological evidence.Étud Océan Indien 42–43:13–46
Fuller DQ, Harvey EL (2006) The archaeobotany of Indian pulses:identification, processing and evidence for cultivation. EnvArchaeol 11(2):219–246. doi:10.1179/174963106x123232
Fuller D, Qin L (2008) Immature rice and its archaeobotanicalrecognition: a reply to Pan. Antiquity 82(316), on-line. Retrievedfrom http://antiquity.ac.uk/ProjGall/fuller2/index.html
Fuller D, Qin L (2009) Water management and labour in the originsand dispersal of Asian rice. World Archaeol 41(1):88–111
Fuller DQ, Zhang H (2007) A preliminary report of the surveyarchaeobotany of the upper Ying Valley (Henan Province) [inChinese and English]. In: University and Henan ProvincialInstitute of Archaeology (ed) Dengfeng wangchenggang yizhide faxian yu yanjiu (2002–2005) [Archaeological Discovery and
Research at the Wangchenggang Site in Dengfeng (2002–2005)][in Chinese]. Great Elephant, Zhengzhou, pp 916–958
Fuller DQ, Harvey E, Qin L (2007) Presumed domestication? Evidencefor wild rice cultivation and domestication in the fifth millenniumBC of the lower Yangtze region. Antiquity 81:316–331
Fuller DQ, Qin L, Zheng Y, Zhao Z, Chen X, Hosoya LA, Sun G (2009)The domestication process and domestication rate in rice: spikeletbases from the Lower Yangtze. Science 323(5921):1607–1610
Garris AJ, Tai TH, Coburn J, Kresovich S, McCouch SR (2005)Genetic structure and diversity in Oryza sativa L. Genetics169:1631–1638
Glover IC, Higham CF (1996) New evidence for early rice cultivationin South, Southeast and East Asia. In: Harris DR (ed) The originsand spread of agriculture and pastoralism in Eurasia. UCL,London, pp 413–441
Gupta PK, Rustgi S, Kumar N (2006) Genetic and molecular basis ofgrain size and grain number and its relevance to grain productivity inhigher plants. Genome 571:565–571. doi:10.1139/G06-063
Hammer K (1984) Das Domestikationssyndrome. Kulterpflanze32:11–34
Harlan JR, De Wet JM, Price EG (1973) Comparative evolution ofcereals. Evolution 27:311–325
Harvey EL (2006) Early agricultural communities in Northern andEastern India: an archaeobotanical investigation. UnpublishedPh.D. dissertation, University College London
Harvey EL, Fuller D, Pal JN, Gupta MC (2003) Early agriculture ofthe Neolithic Vindhyas (North Central India). In: Franke-Vogt U,Weisshaar H-J (eds) South Asian archaeology, Proceedings of theseventeenth international conference of the European Associationof South Asian Archaeologists (7–11 July 2003, Bonn). DeutchesArchaeologisches Institut, Achen, pp 329–334
Heiser CB (1988) Aspects of unconscious selection and the evolutionof domesticated plants. Euphytica 37:77–81
Higham C (2003) Languages and farming dispersals: Austroasiaticlanguages and rice cultivation. In: Bellwood P, Renfrew C (eds)Examining the farming/language dispersal hypothesis. Mc DonaldInstitute for Archaeological Research, Cambridge, pp 223–232
Higham C, Higham T (2009) A new chronological framework forprehistoric Southeast Asia, based on a Bayesian model from BanNon Wat. Antiquity 83(319):125–144
Ho P-T (1956) Early-ripening rice in Chinese history. Econ Hist Rev 9(2):200–218
Ho P-T (1977) The indigenous origins of Chinese agriculture. In: ReedCA (ed) Origins of agriculture. Mouton, The Hague, pp 413–484
Horiuchi T, Samy SJ, Phang CC (1971) Grain loss during handharvesting in the rice cultivation in Kedah, West Malaysia. TonanAjia Kinkyu (Southeast Asian Studies) 9(2):220–226
Hudson M (2003) Agriculture and language change in the JapaneseIslands. In: Bellwood P, Renfrew AC (eds) Examining thefarming/language dispersal hypothesis. McDonald Institute forArchaeological Research, Cambridge, pp 311–318
HPIAC [=Hunan Provincial Institute of Archaeology and CulturalRelics] (2006) Pengtoushan and Bashidang [in Chinese].Wenwu, Beijing
Huke RE (1982) Agroclimatic and dry-season maps of the South, andSoutheast, and East Asia. International Rice Research Institute,Los Banos
Imamura K (1996) Prehistoric Japan. New perspectives on insular EastAsia. UCL, London
Ishikawa R, Sato Y-I, Tang T, Nakamura I (2002) Different maternalorigins of Japanese lowland and upland rice populations. TheorAppl Genet 104:976–980
Jain S, Jain RK, McCouch SR (2004) Genetic analysis of Indianaromatic and quality rice (Oryza sativa L.) germplasm usingpanels of fluorescently-labeled microsatellite markers. TheorAppl Genet 109:965–977
Archaeol Anthropol Sci

Jarrige J (1985) Continuity and change in the North Kachi Plain(Baluchistan, Pakistan) at the beginning of the second millenni-um B.C. In: Schotmans J, Taddei M (eds) South Asianarchaeology 1983. Instituto Universitario Orientale, Dipartimentodi Studi Asiatici, Naples, pp 35–68
Jiang L, Liu L (2006) New evidence for the origins of sedentism andrice domestication in the Lower Yangzi River, China. Antiquity80:355–361
Jin J, Huang W, Gao J, Yang J, Shi M, Zhu M, Luo D, Lin H (2008)Genetic control of rice plant architecture under domestication.Nat Genet 40:1365–1369
Kawakami S, Ebana K, Nishikawa T, Sato Y, Vaughan DA, KadowakiK (2007) Genetic variation in the chloroplast genomes suggestmultiple domestication of cultivated Asian rice (Oryza sativa L.).Genome 50:180–187
Keally CT (2004) Bad science and the distortion of history:radiocarbon dating in Japanese Archaeology. Sophia Int Rev26. Accessed on-line (30 March 2010), http://www.t-net.ne.jp/∼keally/Reports/sir2004.html
Kim J, Yang D, Nahm W, Yi S, Hong S, Yun H et al (2008) LastGlacial and Holocene fluvial wetland sedimentary stratigraphy:comparison between Soro-ri and Jangheung-ri archeologicalsites, Korea. Quatern Int 176-177:135–142, Retrieved fromhttp://dx.doi.org/10.1016/j.quaint.2007.05.013
Kitano H, Futsuhara Y, Satoh H (1993) Morphological variations inrice cultivars. In: Matsuo T, Hoshikawa K (eds) Science of therice plant. Volume One. Morphology. Food and AgriculturePolicy Research Center, Tokyo, pp 79–88
Konishi S, Izawa T, Lin SY, Ebana K, Fukuta Y, Sasaki T, Yano M(2006) An SNP caused loss of seed shattering during ricedomestication. Science 312(5778):1392–1396
Kovach MJ, Sweeney MT, McCouch SR (2007) New insights intothe history of rice domestication. Trends Genet 23:578–587
Kovach MJ, Calingacion MN, Fitzgerald MA, McCouch SR (2009)The origin and evolution of fragrance in rice (Oryza sativa L.).Proc Natl Acad Sci USA 106(34):14444–14449
Kuraku Y (2002) The origin and development of rice cultivation inJapan. In: Yasuda Y (ed) The origins of pottery and agriculture.Lustre, Delhi, pp 312–317
Kuzmin YV (2006) Chronology of the earliest pottery in East Asia:progress and pitfalls. Antiquity 80(308):362–371
Lee Y, Woo J (2006) La culture du Paleolithique dans la region deJungwon en Coree. Anthropologie 110(2):175–186, Retrievedfrom http://www.sciencedirect.com/science/article/B6X0X-4JTRTBR-2/2/94d21c38ae6e2e732a883b8498039a8f
Lee G-A, Crawford G, Liu L, Chen X (2007) Plants and people fromthe early Neolithic to the Shang periods in North China. ProcNatl Acad Sci 104(3):1087–1092
Li C, Zhou A, Sang T (2006a) Genetic analysis of the ricedomestication syndrome with the wild annual species, Oryzanivara. New Phytol 170(1):185–193
Li C, Zhou A, Sang T (2006b) Rice domestication by reducingshattering. Science 311:1936–1939
Li X, Dodson J, Zhou X, Zhang H, Masutomoto R (2007) Earlycultivated wheat and broadening of agriculture in NeolithicChina. Holocene 17:555–560
Lin Z, Griffith ME, Li X, Zhu Z, Tan L, Fu Y et al (2007) Origin ofseed shattering in rice (Oryza sativa L.). Planta 226:11–20
Liu L, Lee G, Jiang L, Zhang J (2007) Evidence for the earlybeginning (c. 9000 cal. BP) of rice domestication in China: aresponse. Holocene 17:1059–1068
Londo JP, Chiang Y, Hung K, Chiang T, Schaal BA (2006)Phylogeography of Asian wild rice, Oryza rufipogon, revealsmultiple independent domestications of cultivated rice, Oryzasativa. Proc Natl Acad Sci USA 103:9578–9583
Lu TLD (2006) The occurrence of cereal cultivation in China. AsianPerspect 45(2):129–158
Lu TLD (2009) Prehistoric coexistence: the expansion of farmingsociety from the Yangzi River to Western South China. In: IkeyaK, Ogawa H, Mitchell P (eds) Interactions between hunter–gatherers and farmers: from prehistory to present. NationalMuseum of Ethnology, Osaka, pp 47–52
Lu H, Liu Z, Wu N, Berné S, Saito Y, Liu B, Wang L (2002) Ricedomestication and climatic change: phytolith evidence from EastChina. Boreas: An International Journal of Quaternary Research31(4):378–385
Ma J, Bennetzen JL (2004) Rapid recent growth and divergence of ricenuclear genomes. Proc Natl Acad Sci USA 101:12404–12410
McNally KL, Childs KL, Bohnert R, Davidson RM, Zhao K, Ulat VJ,Zeller G, Clark RM, Hoen DR, Bureau TE, Stokowski R,Ballinger DG, Frazer KA, Cox DR, Padhukasahasram B,Bustamante CD, Weigel D, Mackill DJ, Bruskiewich RM, RatschG, Buell CR, Leung H, Leach JE (2009) Genomewide SNPvariation reveals relationships among landraces and modernvarieties of rice. Proc Natl Acad Sci USA 106(30):12273–12278
Miller NF (1981) Plant remains from Ville Royale II, Susa. Cah DAFI12:137–142
Mori Y (2002) The origin and development of rice paddy cultivationin Japan based on evidence of insect and diatom fossils. In:Yasuda Y (ed) The origins of pottery and agriculture. Lustre,Delhi, pp 273–296
Mudar KM (1999) How many Dvaravati Kingdoms? Locationalanalysis of first millennium A.D. Moated settlements in CentralThailand. J Anthropol Archaeol 18:1–28
Nakamura S (2010) The origin of rice cultivation in the lower YangtzeRegion, China. Archaeol Anthropol Sci 2 (this issue)
Nakamura I, Urairong H, Kameya N, Fukuta Y, Chitrkon S, Sato YI(1998) Six different plastid subtypes were found in the O. sativa–O. rufipogon complex. Rice Genet Newslett 15:80–82
Nasu H, Momohara A, Yasuda Y, He J (2007) The occurrence andidentification of Setaria italica (L.) P. Beauv. (foxtail millet)grains from the Chengtoushan site (ca. 5800 cal B.P.) in centralChina, with reference to the domestication centre in Asia. VegHist Archaeobot 16(6):481–494
Nitsuma Y (1993) Upland rice. In: Matsuo T, Hoshikawa K (eds)Science of the rice plant, vol. 1. Morphology. Food andAgriculture Policy Research Center, Tokyo, pp 70–76
Norton CJ (2007) Sedentism, territorial circumscription, and theincreased use of plant domesticates across Neolithic–BronzeAge Korea. Asian Perspect 46(1):133–165
Ohtsubo H, Cheng C, Ohsawa I, Tsichimoto S, Ohtsubo E (2004) Riceretroposon p-SINE1 and origin of cultivated rice. Breed Sci 54:1–11
Oka H (1988) Origins of cultivated rice. Elsevier, AmsterdamOlsen KM, Purugganan MD (2002) Molecular evidence on the origin
and evolution of glutinous rice. Genetics 162:941–950Pan Y (2008) Immature wild rice harvesting at Kuahuqiao, China?
Antiquity 82. Online: http://www.antiquity.ac.uk/ProjGall/pan/inPaz V (2003) Island Southeast Asia: spread or friction zone? In:
Bellwood P, Renfrew AC (eds) Examining the farming/languagedispersal hypothesis. McDonald Institute for ArchaeologicalResearch, Cambridge, pp 275–285
Purugganan MD, Fuller DQ (2009) The nature of selection duringplant domestication. Nature 457(7231):843–848. doi:10.1038/nature07895
Qin L, Fuller (Fu Daolian) DQ (2009) Appendix 3. The Nanjiaokousite 2007 excavated Early to Mid Yangshao plant remains. In:Henan Provincial Institute of Cultural Relics and Archaeology(ed) Nanjiaokou site in Sanmenxia. Science, Beijing, pp 427–435, In Chinese
Ren G, Beug H (2002) Mapping Holocene pollen data and vegetationof China. Quatern Sci Rev 21:1395–1422
Archaeol Anthropol Sci

Rispoli F (2008) The incised and impressed pottery of MainlandSoutheast Asia: following the paths of Neolithization. East West57:235–304
Rosen A (2008) The impact of environmental change and human landuse on alluvial valleys on the Loess Plateau of China during theMid-Holocene. Geomorphology 101:298–307
Sato Y, Tang SX, Yang LU, Tang LH (1991) Wild-rice seeds in theoldest rice remains. Rice Genetics Newsletter 8:76
Sagart L (2008) The expansion of Setaria farmers in East Asia: alinguistic and archaeological model. In: Sanchez-Mazas A,Blench RM, Ross M, Lin M, Pejros I (eds) Human migrationsin continental East Asia and Taiwan: matching archaeology,linguistics and genetics. Taylor & Francis, London, pp 133–157
Sage RF (1995) Was low atmospheric CO2 during the Pleistocene alimiting factor for the origin of agriculture? Glob Chang Biol1:93–106
Sang T, Ge S (2007) The puzzle of rice domestication. J Integr PlantBiol 49(6):760–768
Saraswat KS, Sharma NK, Saini DC (1994) Plant economy ay ancientNarhan (ca. 1,300 B.C.–300/400 A.D.). In: Singh P (ed)Excavations at Narhan (1984–1989). Banaras Hindu University,Varanasi, pp 255–346
Sato Y (1996) DNA-ga Kataru Inasaku Bunmaei Kingento Tenkai[The origins of rice culture inferred from DNA analysis] (inJapanese). NHK Books, Tokyo
Sato Y (2002) Origin of rice cultivation in the Yangtze River Basin.In: Yasuda Y (ed) The origins of pottery and agriculture. Lustre,New Delhi, pp 143–150
Saxena A, Prasad V, Singh IB, Chauhan MS, Hassan R (2006) On theHolocene record of phytoliths of wild and cultivated rice fromGanga Plain: evidence for rice-based agriculture. Curr Sci 90(11):1547–1552
Shan J, Zhu M, Shi M, Gao J, Lin H (2009) Fine mapping andcandidate gene analysis of spd6, responsible for small panicleand dwarfness in wild rice (Oryza rufipogon Griff.). Theor ApplGenet 119(5):827–836
Sharma AK (1982) Excavations at Gufkral, 1981. Puratattva 11:19–25Shi Y, Kong Z, Wang S, Tang L, Wang F, Yao T et al (1993) Mid-
Holocene climates and environments in China. Glob PlanetChange 7:219–233
Shomura A, Izawa T, Ebana K, Ebitani T, Kanegae H, Konishi S, YanoM(2008) Deletion in a gene associated with grain size increased yieldsduring rice domestication. Nat Genet 40:1023–1028
Song X, Huang W, Shi M, Zhu M-Z, Lin H (2007) A QTL for ricegrain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat Genet 39:623–630
Stacul G (1992) Swat, Pirak, and connected problems (mid-2ndmillennium B.C.). In: Jarrige C, Gerry JP, Meadow RH (eds)South Asian archaeology 1989. Prehistory, Madison, pp 267–270
Sweeney MT, McCouch SR (2007) The complex history of thedomestication of rice. Ann Bot 100:951–957
Takahashi R (2009) Symbiotic relations between paddy-field ricecultivators and hunter-gatherer-fishers in Japanese prehistory:archaeological considerations on the transition from the JomonAge to the Yayoi Age. In: Ikeyea K, Ogawa H, Mithcell P (eds)Interactios between hunter-gatherers and farmers: from prehistoryto present. Senri Ethnological Studies 73. National Museum ofEthnology, Osaka, pp 71–98
Takahashi H, Sato Y, Nakamura I (2008) Evolutionary analysis of twoplastid DNA sequences in cultivated and wild species of Oryza.Breed Sci 233:225–233
Takamiya H (2001) Introductory routes of rice to Japan: an examinationof the Southern Hypothesis. Asian Perspect 40(2):209–226
Tan L, Li X, Liu F, Sun X, Li C, Zhu Z et al (2008) Control of a keytransition from prostrate to erect growth in rice domestication.Nat Genet 40:1360–1364
Tanaka K, Ishikawa R, Honda T (2010) Rice archaeological remains andthe possibility of DNA archaeology: examples fromYayoi and Heianperiods of Northern Japan. Archaeol Anthropol Sci (this volume)
Tang JB, Xia HA, Cao ML, Zhang XQ, Zheng WY, Hu SN et al(2004) A comparison of rice chloroplast genomes. Plant Physiol135:412–420
Tewari R, Srivastava RK, Singh KK, Saraswat KS, Singh IB, ChauhanMS et al (2006) Second preliminary report of the excavations atLahuradewa. District Sant Kabir Nagar, U. P.: 2002–2003–2004& 2005–06. Pragdhara 16:35–68
Tewari R, Srivastava RK, Saraswat KS, Singh IB, Singh KK (2008)Early farming at Lahuradewa. Pragdhara 18:347–373
Thompson GB (1996) The excavations of Khok Phanom Di, aprehistoric site in Central Thailand. Volume IV. Subsistence andenvironment: the botanical evidence. The biological remains partIII. The Society of Antiquaries of London, London
Thompson GB (1997) Archaeobotanical indicators of rice domestica-tion—a critical evaluation of diagnostic criteria. In: Ciarla R,Rispoli F (eds) South-East Asian archaeology 1992. InstitutoItaliano per il Medio ed Estremo Orientale, Rome, pp 159–174
Toyama S (2002) The origin and spread of rice cultivation as seenfrom rice remains. In: Yasuda Y (ed) The origins of pottery andagriculture. Lustre, Delhi, pp 263–272
Tsang C (2005) Recent discoveries at a Tapenkeng culture site inTaiwan: implications for the problem of Austronesian origins. In:Sagart L, Blench R, Sanchez-Mazas A (eds) The peopling of EastAsia: putting together archaeology, linguistics and genetics.Routledge-Curzon, London, pp 63–74
Vaughan DA, Kadowaki K, Kaga A, Tomooka N (2005) On thephylogeny and biogeography of the genus Oryza. Breed Sci55:113–122
Vaughan DA, Lu B, Tomooka N (2008a) The evolving story of riceevolution. Plant Sci 174(4):394–408
Vaughan DA, Lu B, Tomooka N (2008b) Was Asian rice (Oryzasativa) domesticated more than once? Rice 1:16–24
Vincent B (2003) Rice in pottery: new evidence for early ricecultivation in Thailand. Indo-Pacific Prehistory AssociationBulletin 23:51–58
Vitte C, Ishii T, Lamy F, Brar D, Panaud O (2004) Genomicpaleontology provides evidence for two distinct origins of Asianrice (Oryza sativa L.). Mol Genet Genomics 272:504–511
Weber S, Lehman H, Barela T, Hawks S, Harriman D (2010) Rice ormillets: early farming strategies in prehistoric central Thailand.Archaeol Anthropol Sci 2 (this volume). doi:10.1007/s12520-010-0030-3
White JC (1995) Modeling the development of early rice agriculture:ethnoecological perspectives from northeast Thailand. AsianPerspect 34:37–68
Xiong LZ, Liu KD, Dai XK, Xu CG, Zhang Q (1999) Identification ofgenetic factors controlling domestication-related traits of riceusing an F2 population of a cross between Oryza sativa and O.rufipogon. Theor Appl Genet 98(2):243–251. doi:10.1007/s001220051064
You X (1987) The wild rice in Chinese ancient records(Zhongguo Gushu Zhong Jizai de Yeshengdao Tantao) [inChinese]. Gujin Nongye [Ancient and Modern Agriculture]1987(1):1–6
Yu SY, Zhu C, Song J, Qu WZ (2000) Role of climate in the rise and fallof Neolithic cultures on the Yangtze Delta. Boreas 29:157–165
Zhang W (2002) The bi-peak tubercle of rice, the character of ancientrice and the origin of cultivated rice. In: Yasuda Y (ed) Theorigins of pottery and agriculture. Lustre, Delhi, pp 205–216
Zhang C, Hung H-C (2008) The Neolithic of southern China—origin,development and dispersal. Asian Perspect 47(2):299–329
Zhang C, Hung H (2010) The emergence of agriculture in southernChina. Antiquity 84(323):11–25
Archaeol Anthropol Sci

Zhang J, Wang X (1998) Notes on the recent discovery of ancientcultivated rice at Jiahu, Henan Province: a new theoryconcerning the origin of Oryza japonica in China. Antiquity72:897–901
Zhang LB, Zhu Q, Wu ZQ, Ross-Ibarra J, Gaut B, Ge S, Sang T(2009) Selection on grain shattering genes and rates of ricedomestication. New Phytol 184:708–720
Zhao Z (1998) The Middle Yangtze region in China is one placewhere rice was domesticated: phytolith evidence from theDiaotonghuan Cave, Northern Jiangxi. Antiquity 72:885–897
Zhao Z (2010) New data and new issues for the study of origin ofrice agriculture in China. Archaeol Anthropol Sci 2 (thisvolume)
Zhao Z, Piperno D (2000) Late Pleistocene/Holocene environments inthe Middle Yangtze River Valley of the Taihu Lake in China andrice (Oryza sativa L.) domestication: the phytoliths evidence.Geoarchaeology 15:203–222
Zhao Z, Pearsall DM, Benfer RA, Piperno DR (1998) Distinguishingrice (Oryza sativa Poaceae) from wild Oryza species throughphytolith analysis II: finalised method. Econ Bot 52:134–145
Zheng Y, Dong Y, Matsui A, Udatsu E, Fuijiwara H (2003) Moleculargenetic basis of determining subspecies of ancient rice usingshape of phytoliths. J Archaeol Sci 30:1215–1221
Zheng Y, Sun G, Chen X (2007) Characteristics of the short rachillaeof rice from archaeological sites dating to 7000 years ago. ChinSci Bull 52:1654–1660
Archaeol Anthropol Sci





























