Embed Size (px)
Citation preview

R E V I E W A R T I C L E
Cyanobacterial hydrogenases:diversity, regulationandapplicationsPaula Tamagnini1,2, Elsa Leitao1, Paulo Oliveira3, Daniela Ferreira1,2, Filipe Pinto1, David James Harris4,5,Thorsten Heidorn3 & Peter Lindblad3
1IBMC – Instituto de Biologia Molecular e Celular, Universidade do Porto, Porto, Portugal; 2Departmento de Botanica, Faculdade de Ciencias,
Universidade do Porto, Porto, Portugal; 3Department of Photochemistry and Molecular Science, The Angstrom Laboratories, Uppsala University,
Uppsala, Sweden; 4CIBIO/UP, Centro de Investigacao em Biodiversidade e Recursos Geneticos, Universidade do Porto, Vairao, Portugal; and5Departamento de Zoologia e Antropologia, Faculdade de Ciencias da Universidade do Porto, Porto, Portugal
Correspondence: Paula Tamagnini, IBMC –
Instituto de Biologia Molecular e Celular, Rua
do Campo Alegre, 823. 4150-180 Porto,
Portugal. Tel.: 1351 2260 74900; fax: 1351
2260 99157; e-mail: [email protected]
Received 5 January 2007; revised 12 July 2007;
accepted 9 August 2007.
First published online October 2007.
DOI:10.1111/j.1574-6976.2007.00085.x
Editor: Annick Wilmotte
Keywords
cyanobacteria; hydrogenase; hup ; hox ; hyp ;
transcriptional regulator.
Abstract
Cyanobacteria may possess two distinct nickel-iron (NiFe)-hydrogenases: an
uptake enzyme found in N2-fixing strains, and a bidirectional one present in both
non-N2-fixing and N2-fixing strains. The uptake hydrogenase (encoded by hupSL)
catalyzes the consumption of the H2 produced during N2 fixation, while the
bidirectional enzyme (hoxEFUYH) probably plays a role in fermentation and/or
acts as an electron valve during photosynthesis. hupSL constitute a transcriptional
unit, and are essentially transcribed under N2-fixing conditions. The bidirectional
hydrogenase consists of a hydrogenase and a diaphorase part, and the correspond-
ing five hox genes are not always clustered or cotranscribed. The biosynthesis/
maturation of NiFe-hydrogenases is highly complex, requiring several core
proteins. In cyanobacteria, the genes that are thought to affect hydrogenases
pleiotropically (hyp), as well as the genes presumably encoding the hydrogenase-
specific endopeptidases (hupW and hoxW) have been identified and characterized.
Furthermore, NtcA and LexA have been implicated in the transcriptional regula-
tion of the uptake and the bidirectional enzyme respectively. Recently, the
phylogenetic origin of cyanobacterial and algal hydrogenases was analyzed, and it
was proposed that the current distribution in cyanobacteria reflects a differential
loss of genes according to their ecological needs or constraints. In addition, the
possibilities and challenges of cyanobacterial-based H2 production are addressed.
Introduction
Cyanobacteria, one of the largest and most important
groups of bacteria on Earth, are able to perform oxygenic
photosynthesis using water as an electron donor and may be
found in almost any ecological niche from fresh to salt water,
terrestrial and extreme environments (Whitton & Potts,
2000). The knowledge on such a diverse group of prokar-
yotic organisms has greatly increased since cyanobacterial
genomes became available. In 1996, the entire sequence of
Synechocystis sp. PCC 6803 was published (Kaneko et al.,
1996; Nakamura et al., 1998), and since then, many other
cyanobacterial genome projects have been completed and
released, including that of Nostoc punctiforme ATCC 29133/
PCC 73102, one of the largest microbial genomes sequenced
so far (Meeks et al., 2001; Anderson et al., 2006).
Fossil traces of cyanobacteria are claimed to have been
found from around 3.5 billion years ago (Schopf, 2000), and
ancestors of cyanobacteria most probably played a key role
in the formation of atmospheric oxygen, and are thought
to have evolved into present-day chloroplasts of algae and
green plants (Miyagishima, 2005; Mulkidjanian et al., 2006).
Cyanobacteria display a relatively wide range of morpholo-
gical diversity, including unicellular, filamentous and colo-
nial forms. Some filamentous strains form differentiated
cells specialized in nitrogen fixation – heterocysts, and
spore-like resting cells – akinetes. A number of nonhetero-
cystous strains are also able to perform N2 fixation under
certain conditions. The fact that several cyanobacteria
are able to reduce nitrogen and carbon under aerobic
conditions may be responsible for their evolutionary and
ecological success. In cyanobacteria, as in any diazotrophic
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved

bacteria, the reduction of N2 to NH3 is accompanied by the
formation of molecular hydrogen (Berman-Frank et al.,
2003). The H2 produced by the nitrogenase is rapidly
consumed by an uptake hydrogenase, an enzyme that has
been found in almost all the N2-fixing cyanobacteria exam-
ined so far, with one reported exception – Synechococcus sp.
BG 043511 (Ludwig et al., 2006). Additionally, these strains
may contain a bidirectional hydrogenase, an enzyme that is
generally present in the non nitrogen-fixing cyanobacteria
(Tamagnini et al., 2002, 2005), but absent in Gloeobacter
violaceus PCC 7421, a cyanobacterium that possesses a
number of unique characteristics such as the absence of
thylakoids (Nakamura et al., 2003; Ludwig et al., 2006). The
distribution of genes related to hydrogenases among repre-
sentative cyanobacterial strains is displayed in Table 1. Both
cyanobacterial hydrogenases are NiFe enzymes, which are
the most common hydrogenases found in bacteria and
Archaea. The core enzyme consists of an ab heterodimer
with the large/a subunit hosting the bimetallic active site,
and the small/b-subunit containing the FeS clusters, which
function as electron transfer domains between the electron
acceptors/donors and the catalytic center of the enzyme
(Fig. 1). In general, the NiFe hydrogenases are divided into
four groups, with the cyanobacterial uptake hydrogenases
clustering together with the cytoplasmic H2 sensors of group
2, and the bidirectional enzymes belonging to group 3
comprising the bidirectional heteromultimeric cytoplasmic
hydrogenases (for reviews on this subject, see Vignais et al.,
2001; Vignais & Colbeau, 2004).
In the present review, recent advances on cyanobacterial
hydrogenases, have been summarized focusing on achieve-
ments on the diversity and molecular regulation of both the
uptake and the bidirectional enzyme.
Photobiological production of H2 by microorganisms is
of great public interest because it promises a renewable
energy carrier from nature’s most plentiful resources: solar
energy and water. Cyanobacteria and green algae are the
only organisms known so far that are capable of both
oxygenic photosynthesis and hydrogen production. In a
separate section, the possibilities and challenges in cyano-
bacterial-based hydrogen production are outlined.
Uptake hydrogenase
The cyanobacterial uptake hydrogenase, found exclusively in
N2-fixing strains and encoded by the hup – hydrogen uptake
– genes, is at least a heterodimeric enzyme with a large
subunit of about 60 kDa containing the active site (HupL)
and a small subunit of c. 35 kDa playing a role in electron
transfer (HupS) (Fig. 1). Because the physiological and
biochemical data point to a membrane-bound enzyme
(Houchins & Burris, 1981b; Houchins, 1984; Lindblad &
Sellstedt, 1990; Rai et al., 1992), and the hydropathy profiles
of the HupL and the HupS proteins do not indicate any
transmembrane domains (Tamagnini et al., 2005), the
existence of a polypeptide that anchors the HupSL hetero-
dimer to the membrane seems likely. In fact, analysis of the
available genomes revealed the presence of ORFs whose
products could potentially fulfill this anchoring role (Lind-
berg, 2003). However, to date no definitive proof was
obtained, and the existence of both a soluble and a mem-
brane-bound form of the enzyme cannot be excluded (see
for e.g. Houchins & Burris, 1981b).
Immunolocalization studies, using antibodies produced
against hydrogenases from other bacteria, showed that the
hydrogenase antigens are present in both the vegetative cells
and heterocysts of N. punctiforme, and several symbiotic
Nostoc strains (Lindblad & Sellstedt, 1990; Rai et al., 1992;
Tamagnini et al., 1995). However, these studies do not
clarify whether the enzyme is in its active form in both
cell types. In Anabaena/Nostoc sp. PCC 7120, the uptake
hydrogenase activity was essentially associated with the
particulate fraction of the heterocysts (Houchins & Burris,
1981b); however, one must bear in mind that in this strain
the hupL gene undergoes a rearrangement, allowing its
expression in the heterocysts only, and that this process does
not occur in N. punctiforme (Oxelfelt et al., 1998). Moreover,
the presence/levels of the cyanobacterial uptake hydrogenase
are certainly dependent on the growth conditions. In
heterocystous cyanobacteria grown in air and without
combined nitrogen, the uptake hydrogenase activity is
mainly confined to heterocysts, where it is protected from
oxygen inactivation; however, the exact location of the
enzyme in cyanobacteria should be further investigated in
both heterocystous and nonheterocystous strains.
A strong correlation between the nitrogen-fixation pro-
cess and the uptake hydrogenase activity has been demon-
strated for cyanobacteria (Lambert & Smith, 1981;
Houchins, 1984; Wolk et al., 1994; Oxelfelt et al., 1995;
Schutz et al., 2004), and this indicates that the main
physiological function of the uptake hydrogenase is to
reutilize and regain the H2/electrons produced by the H2
evolution through the nitrogenase. This recycling has been
suggested to have at least three beneficial functions to the
organism: (1) it provides ATP via the oxyhydrogen reaction,
minimizing the loss of energy; (2) it removes the oxygen
from nitrogenase, thereby protecting it from inactivation;
and (3) it supplies reducing equivalents (electrons) to
various cell functions (Bothe et al., 1977, 1991; Howarth &
Codd, 1985; Weisshaar & Boger, 1985; Smith, 1990).
Physical organization of hup genes and thecorresponding proteins
The physical arrangement of the structural genes encoding
the uptake hydrogenase is very similar in all the
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
693Cyanobacterial hydrogenases

Tab
le1.
Dis
trib
ution
of
gen
esre
late
dto
hyd
rogen
ases
inre
pre
senta
tive
cyan
obac
terial
stra
ins
Org
anis
ms
Bid
irec
tional
hyd
rogen
ase
Upta
ke
hyd
rogen
ase
hupL
reco
mbin
ase
Bid
irec
tional
spec
ific
endopep
tidas
e
Upta
kesp
ecifi
c
endopep
tidas
e
Oth
er
mat
ura
tion
gen
es
Gen
Ban
k
acce
ssio
n
num
ber
/
Ref
eren
ces
hoxF
UY
HhoxE
hupSL
Xis
C�
hoxW
hupW
hyp
FCD
EAB
Unic
ellu
lar
non-N
2-fi
G.vi
ola
ceus
PCC
7421
��
��
��
NC
_005125
Nak
amura
etal
.
(2003)
Synec
hocy
stis
sp.
PCC
6803
1 Appel
&Sc
hulz
(1996)
1�
1�
1 Scat
tere
d
NC
_000911
Kan
eko
etal
.
(1996)
Unic
ellu
lar
N2-fi
C.w
atso
nii
WH
8501
��
1�
11 Sc
atte
red
NZ_
AD
V00000000
Fila
men
tous
nonhet
erocy
stousL.
maj
usc
ula
CC
AP
1446/4
1N
D1
ND
11 O
per
on
Leitao
etal
.(2
005,
2006)
N2-fi
T.er
ythra
eum
IMS
101
��
1�
11
NC
_008312
Fila
men
tous
het
erocy
stous
N2-fi
A.va
riab
ilis
ATC
C29413
1 Schm
itz
etal
.
(1995)
11 H
appe
etal
.(2
000)
�1
11
NC
_007413
Nost
oc
sp.
PCC
7120
11
1 Car
rasc
oet
al.(1
995)
11
11 G
ubili
&Bort
hak
ur
(1996,1998)
NC
_003272
Kan
eko
etal
.(2
001)
N.punct
iform
e
PCC
73102
��
1 Oxe
lfel
tet
al.(1
998)
��
11 O
per
on
Han
sele
tal
.(2
001)
NZ_
AA
AY
00000000
� Rea
rrag
emen
tocc
urr
ing
during
the
diffe
rentiat
ion
of
ave
get
ativ
ece
llin
toa
het
erocy
st.
ND
,not
det
erm
ined
.
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
694 P. Tamagnini et al.

cyanobacteria studied so far: hupS and hupL are contiguous,
with the gene encoding the smaller subunit located up-
stream from the gene encoding the larger one (Carrasco
et al., 1995; Oxelfelt et al., 1998; Happe et al., 2000; Lindberg
et al., 2000; Oliveira et al., 2004; Leitao et al., 2005) (Fig. 2).
Transcriptional start sites have been identified upstream of
the hupS start codon (Happe et al., 2000; Lindberg et al.,
2000; Oliveira et al., 2004; Leitao et al., 2005), and a putative
transcriptional terminator, located immediately down-
stream of hupL, has been found in N. punctiforme (Lindberg
et al., 2000). In agreement, reverse transcriptase (RT)-PCR
experiments, and the sizes of transcripts determined by
Northern blot, indicate that hupSL constitute a transcrip-
tional unit in Anabaena variabilis ATCC 29413, N. puncti-
forme and Lyngbya majuscula CCAP 1446/4 (Happe et al.,
2000; Lindberg et al., 2000; Leitao et al., 2005). In the
unicellular Gloeothece sp. ATCC 27152 and in the filamen-
tous Trichodesmium erythraeum IMS 101 hupW – the gene
encoding for the putative uptake hydrogenase-specific endo-
peptidase – is the ORF located immediately downstream of
hupL, and was shown to be cotranscribed with hupSL in
Gloeothece sp. ATCC 27152 (Oliveira et al., 2004). In other
strains, the position of hupW related to the hupSL varies
considerably, and in the strains examined they are tran-
scribed independently (Wunschiers et al., 2003) (Fig. 2).
Analysis of the predicted proteins encoded by the hupSL
operon demonstrated that whereas HupS has the same
number of amino acid residues in all the cyanobacteria
investigated [320 amino acids (aa)], HupL generally has
531 aa with the exception of the filamentous nonheterocys-
tous L. aestuarii CCY 9616 (six extra), L. majuscula (six extra),
and T. erythraeum (three extra). To date, the physiological
significance (if any) of these extra residues is still unknown.
In the NiFe hydrogenases, the large subunit harbors the
active center that is deeply buried inside the protein, close to
the large interface between the two subunits, and the small
subunit contains the FeS clusters that conduct electrons
between the active center and the physiological electron
acceptor/donor (Vignais et al., 2001; Vignais & Colbeau,
2004). In concordance, the cyanobacterial HupL sequences
contain the four conserved cysteine residues that are in-
volved in the coordination of the bimetallic NiFe center of
the active site, and HupS contains eight cysteine residues
clearly corresponding to those involved in the formation of
the FeS clusters, and a ninth cysteine slightly shifted
compared with other bacteria (Tamagnini et al., 2002). In
addition, HupL contains the C-terminal region that is
presumably cleaved off, by a specific endopeptidase, as the
last step of the maturation of the large subunit. In contrast
with other organisms, HupS lacks both the twin-arginine
HupLHupL
NifKNifK NifDNifD
NifDNifD
NifHNifH
NifHNifH NifKNifK
Dinitrogenasereductase Dinitrogenase
Uptake hydrogenaseUptake hydrogenase
NitrogenaseNitrogenase
NH
N2+ H+
HH22
2H++ 2e−
2H++ 2e−
e−
HupSHupS
HupCHupC??
Hydrogenase
HoxHHoxH
HoxYHoxY HoxFHoxF
HoxEHoxE
Diaphorase
BiBi--directional hydrogenasedirectional hydrogenase
HH22 HoxUHoxU
NAD+
NADH
Hox(EFUYH)2
HupLHupL
NifKNifK NifDNifD
NifDNifD
NifHNifH
NifHNifH NifKNifK
NifKNifK NifDNifD
NifDNifD
NifHNifH
NifHNifH NifKNifK
β α
α β
Uptake hydrogenaseUptake hydrogenase
NitrogenaseNitrogenase
NH
HH22
HupSHupS
HupCHupC??
HupCHupC??
HoxHHoxH
HoxYHoxY HoxFHoxF
HoxEHoxE
BiBi--directional hydrogenasedirectional hydrogenase
HH22 HoxUHoxU
Hox(EFUYH)2
HoxHHoxH
HoxYHoxY HoxFHoxF
HoxEHoxE
BiBi--directional hydrogenasedirectional hydrogenase
HH22 HoxUHoxU
Hox(EFUYH)2
Fig. 1. Enzymes directly involved in hydrogen metabolism in cyanobacteria. While the uptake hydrogenase is present in most of the nitrogen-fixing
strains tested (with only one exception reported so far; see text and Table 1), the bidirectional enzyme seems to be present in non-N2-fixing and N2-fixing
strains but is not a universal enzyme. The existence of a third subunit (HupC) anchoring the uptake hydrogenase to the membrane is yet to be
confirmed, and the molecular weight of the native bidirectional hydrogenase indicates a dimeric assembly of the enzyme complex Hox(EFUYH)2.
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
695Cyanobacterial hydrogenases

1000 bpGloeothece sp. ATCC 27152
S L
hup
W
Trichodesmium erythraeum IMS101
S L
hup
W
Tery0800Tery0790Tery0799Tery0801
FDE CAB
hyp
~ 589 kb
Nostoc punctiforme ATCC 29133 / PCC 73102
FDEB A C
hyp
S L
hup
W
NpR0370NpR0363NpR0364
NpR0366
NpF0371
NpR0365NpF0372
NpF0373
NpR0367
Lyngbya majuscula CCAP 1446/4
FDEB A C
hyp
S L
hup
W
ORF1 ORF3ORF2
ORF4 ORF5 ORF6 ORF7ORF8
ORF9ORF10
ORF11
Nostoc sp. PCC 7120
FDE
alr0691
SB
alr0692 all0675
A
asr0689
C
asr0690
~ 880 kb
alr0693
L
hyp hup
W
hup
asr0697
(a)
Fig. 2. Organization of the loci containing the genes encoding (a) the uptake hydrogenase (hup) and (b) the bidirectional hydrogenase (hox) in selected
cyanobacterial strains (black ORFs). The accessory genes (hyp, hupW and hoxW), encoding proteins involved in the maturation of the hydrogenases are
also depicted, as gray ORFs, as well as some additional ORFs (identified, when available, with the corresponding ORF-number in respective annotated
genomes, and shown as white ORFs). Gloeothece sp. ATCC 27152 (Oliveira et al., 2004 – GenBank accession no. AY260103), Trichodesmium
erythraeum IMS101 (http://genome.jgi-psf.org/finished_microbes/trier/trier.home.html), Lyngbya majuscula CCAP 1446/4 (Leitao et al., 2005 –
GenBank accession no. AF368526), Nostoc punctiforme ATCC 29133/PCC 73102 (http://genome.jgi-psf.org/draft_microbes/nospu/nospu.home.html),
Nostoc sp. PCC 7120 (Kaneko et al., 2001), Synechocystis sp. PCC 6803 (Kaneko et al., 1996), Synechococcus elongatus PCC 7942 (http://genome.
jgi-psf.org/finished_microbes/synel/synel.home.html), Arthrospira platensis FACHB341 (Zhang et al., 2005a, b – GenBank accession nos. DQ309870
and AY345594) and Anabaena variabilis ATCC 29413 (http://genome.jgi-psf.org/finished_microbes/anava/anava.home.html).
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
696 P. Tamagnini et al.

signal peptide at the N-terminal, and the hydrophobic motif
at the C-terminal proposed to be involved in translocation
and anchorage to the membrane, respectively. As mentioned
previously, these general features of the cyanobacterial
hydrogenases cluster them together with the soluble H2-
sensing enzymes (Vignais et al., 2001; Vignais & Colbeau,
2004). However, the construction of hup� mutants proved
that the cyanobacterial uptake hydrogenase is indeed a
Synechocystis sp. PCC 6803
hox
E F U Y H WF DE A2B2C
hyp
B1A1
sll1222 ssl2420
sll1225
~ 192 kb ~743 kb~ 236 kb~ 445 kb~ 79 kb~ 150 kb~ 47 kb~ 477 kb
hyp
1000 bp
Synechococcus elongatus PCC 7942
hox
EDEF
hyphox
U HY W
hyp
AB F Cbk 685 ~bk 433 ~ ~ 172 kb
Arthrospira platensis FACHB 341
unknown
hox
E F U Y H
Anabaena variabilis ATCC 29413
F D E BAC
hyp
Ava4652
~ 52 kb
hox
E F U Y H W
Ava4655
Ava4656
Ava4658
Ava4660
Ava4662 Ava4664
Ava4663
Ava4605
Nostoc sp. PCC 7120
~ 8.8 kb
hox
E F U Y H W~ 65 kb
F D E BAC
hyp
9670lla7670lla5670rla3670rla0570rla
all0768asl0749
asr0697
(b)
Fig. 2. Continued
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
697Cyanobacterial hydrogenases

physiological functional enzyme rather than a regulatory
one (Happe et al., 2000; Lindberg et al., 2002; Lindblad et al.,
2002; Masukawa et al., 2002).
hupL rearrangement in heterocystous strains
Programmed DNA rearrangements have been described in
eukaryotes and prokaryotes but are relatively uncommon
events. In cyanobacteria, developmentally regulated DNA
rearrangements have been reported to occur in heterocys-
tous strains (for a review, see Golden, 1997). Generally, the
ORF is interrupted in the vegetative cells by a 10–60-kb
DNA element, which is excised during the differentiation of
a photosynthetic vegetative cell into a N2-fixing heterocyst,
restoring the structure of the gene/operon and allowing its
expression in heterocysts only.
The rearrangement within hupL (large subunit of the
uptake hydrogenase) was first described for Nostoc sp. PCC
7120 (Carrasco et al., 1995). In the vegetative cells of this
cyanobacterium, hupL is interrupted by a 9.5-kb element
that is excised late during the heterocyst differentiation
process by a site-specific recombination between the 16-bp
direct repeats that flank the element (Fig. 3). The hupL
element contains, in one of its borders, the gene that encodes
the recombinase necessary for the excision – xisC (Carrasco
et al., 1995, 1998, 2005). Site-directed mutagenesis revealed
that the XisC protein has a functional similarity to the phage
integrase family of recombinases. Recently, it has been
unequivocally demonstrated that the inactivation of xisC
blocks the hupL rearrangement and that XisC alone is
sufficient to catalyze the hupL element site-specific recom-
bination in Nostoc sp. PCC 7120 (Carrasco et al., 2005). It
was also shown that the xisC-mutant forms heterocysts
without any obvious developmental defects and that the
mutant grown under N2-fixing conditions (BG110) was not
only defective for hydrogen uptake activity but evolves
H2 (Lindblad et al., 2002; Carrasco et al., 2005). Moreover,
Lindblad et al. (2002) showed that, in a competitive growth
environment with increased light intensity, the wild-type
strain has an advantage over the xisC-mutant, probably
because these specific conditions induced higher rates of
H2 evolution that only the wild type has the capacity of
reutilizing through the oxyhydrogen reaction. These find-
ings support the hypothesis that the uptake hydrogenase
plays a role in minimizing the loss of energy caused by the
nitrogenase-dependent H2 formation.
Despite the hupL element being absent from the two other
heterocystous strains for which genome sequences are avail-
able, A. variabilis and N. punctiforme (see also Oxelfelt et al.,
1998; Happe et al., 2000), DNA hybridization studies
showed that sequences similar to xisC were present in about
half of the heterocystous strains tested (Tamagnini et al.,
2000). These authors also showed that the presence of the
bidirectional hydrogenase is not ubiquitous among hetero-
cystous cyanobacteria, although they could not establish a
correlation between the presence/absence of the bidirec-
tional enzyme and hupL rearrangement.
hupSL intergenic region
The regions between hupS and hupL in cyanobacteria are
longer than in other microorganisms, differ considerably in
Vegetative cell Heterocyst
hupS hupL
9.5 kb ?
enzyme
?
9.5 kb elementcontaining xisC
5´ 3´hupLhupS
Uptake hydrogenase enzyme
hupS hupL
9.5 kb ?
Uptake hydrogenaseenzyme
?
hupS hupLhupS hupLhupS hupL
5´ 3´hupLhupS
Fig. 3. Schematic representation of the hupL rearrangement occurring in Nostoc sp. PCC 7120 and other heterocystous cyanobacteria (adapted from
Carrasco et al., 2005). In the vegetative cells, hupL is interrupted by a DNA element that is excised late during the heterocyst differentiation process by a
site-specific recombination. Subsequently, the structure of the hupL gene is restored, allowing its expression in the heterocysts only. The destiny of the
9.5-kb excised element is currently unknown. In aerobically grown filaments of Nostoc sp. PCC 7120, most of the uptake hydrogenase activity is
recovered in the membrane fraction of heterocysts (Houchins & Burris, 1981b). The question marks represent events that have not been elucidated so
far: the fate of the excised DNA element, and the attachment of the uptake hydrogenase to a cell membrane.
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
698 P. Tamagnini et al.
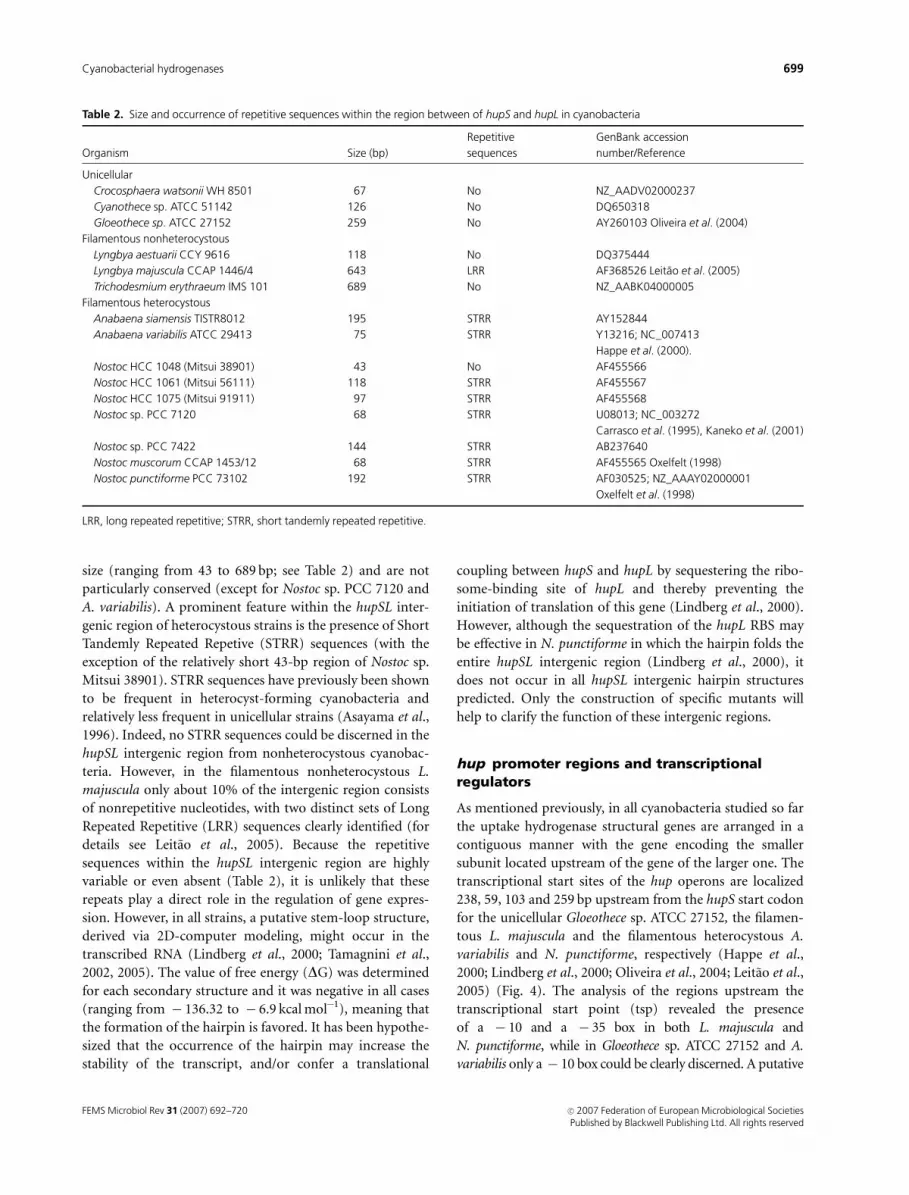
size (ranging from 43 to 689 bp; see Table 2) and are not
particularly conserved (except for Nostoc sp. PCC 7120 and
A. variabilis). A prominent feature within the hupSL inter-
genic region of heterocystous strains is the presence of Short
Tandemly Repeated Repetive (STRR) sequences (with the
exception of the relatively short 43-bp region of Nostoc sp.
Mitsui 38901). STRR sequences have previously been shown
to be frequent in heterocyst-forming cyanobacteria and
relatively less frequent in unicellular strains (Asayama et al.,
1996). Indeed, no STRR sequences could be discerned in the
hupSL intergenic region from nonheterocystous cyanobac-
teria. However, in the filamentous nonheterocystous L.
majuscula only about 10% of the intergenic region consists
of nonrepetitive nucleotides, with two distinct sets of Long
Repeated Repetitive (LRR) sequences clearly identified (for
details see Leitao et al., 2005). Because the repetitive
sequences within the hupSL intergenic region are highly
variable or even absent (Table 2), it is unlikely that these
repeats play a direct role in the regulation of gene expres-
sion. However, in all strains, a putative stem-loop structure,
derived via 2D-computer modeling, might occur in the
transcribed RNA (Lindberg et al., 2000; Tamagnini et al.,
2002, 2005). The value of free energy (DG) was determined
for each secondary structure and it was negative in all cases
(ranging from � 136.32 to � 6.9 kcal mol�1), meaning that
the formation of the hairpin is favored. It has been hypothe-
sized that the occurrence of the hairpin may increase the
stability of the transcript, and/or confer a translational
coupling between hupS and hupL by sequestering the ribo-
some-binding site of hupL and thereby preventing the
initiation of translation of this gene (Lindberg et al., 2000).
However, although the sequestration of the hupL RBS may
be effective in N. punctiforme in which the hairpin folds the
entire hupSL intergenic region (Lindberg et al., 2000), it
does not occur in all hupSL intergenic hairpin structures
predicted. Only the construction of specific mutants will
help to clarify the function of these intergenic regions.
hup promoter regions and transcriptionalregulators
As mentioned previously, in all cyanobacteria studied so far
the uptake hydrogenase structural genes are arranged in a
contiguous manner with the gene encoding the smaller
subunit located upstream of the gene of the larger one. The
transcriptional start sites of the hup operons are localized
238, 59, 103 and 259 bp upstream from the hupS start codon
for the unicellular Gloeothece sp. ATCC 27152, the filamen-
tous L. majuscula and the filamentous heterocystous A.
variabilis and N. punctiforme, respectively (Happe et al.,
2000; Lindberg et al., 2000; Oliveira et al., 2004; Leitao et al.,
2005) (Fig. 4). The analysis of the regions upstream the
transcriptional start point (tsp) revealed the presence
of a � 10 and a � 35 box in both L. majuscula and
N. punctiforme, while in Gloeothece sp. ATCC 27152 and A.
variabilis only a � 10 box could be clearly discerned. A putative
Table 2. Size and occurrence of repetitive sequences within the region between of hupS and hupL in cyanobacteria
Organism Size (bp)
Repetitive
sequences
GenBank accession
number/Reference
Unicellular
Crocosphaera watsonii WH 8501 67 No NZ_AADV02000237
Cyanothece sp. ATCC 51142 126 No DQ650318
Gloeothece sp. ATCC 27152 259 No AY260103 Oliveira et al. (2004)
Filamentous nonheterocystous
Lyngbya aestuarii CCY 9616 118 No DQ375444
Lyngbya majuscula CCAP 1446/4 643 LRR AF368526 Leitao et al. (2005)
Trichodesmium erythraeum IMS 101 689 No NZ_AABK04000005
Filamentous heterocystous
Anabaena siamensis TISTR8012 195 STRR AY152844
Anabaena variabilis ATCC 29413 75 STRR Y13216; NC_007413
Happe et al. (2000).
Nostoc HCC 1048 (Mitsui 38901) 43 No AF455566
Nostoc HCC 1061 (Mitsui 56111) 118 STRR AF455567
Nostoc HCC 1075 (Mitsui 91911) 97 STRR AF455568
Nostoc sp. PCC 7120 68 STRR U08013; NC_003272
Carrasco et al. (1995), Kaneko et al. (2001)
Nostoc sp. PCC 7422 144 STRR AB237640
Nostoc muscorum CCAP 1453/12 68 STRR AF455565 Oxelfelt (1998)
Nostoc punctiforme PCC 73102 192 STRR AF030525; NZ_AAAY02000001
Oxelfelt et al. (1998)
LRR, long repeated repetitive; STRR, short tandemly repeated repetitive.
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
699Cyanobacterial hydrogenases

binding site for NtcA (a protein that operates global nitro-
gen control in cyanobacteria) could be found in Gloeothece
sp. ATCC 27152, L. majuscula and N. punctiforme, although
its relative position to the tsp varied depending on the
strain. Moreover, in L. majuscula and N. punctiforme a
possible binding site for the integration host factor (IHF) –
WATCAAN4TTR (Craig & Nash, 1984; Goodrich et al.,
1990; Goodman et al., 1999) – could be recognized in the
region between the NtcA motif and the tsp (Fig. 4). It has
been postulated that the possible binding of the IHF to the
promoter could bend the DNA (Friedman, 1988), and
consequently allow the contact of the NtcA with the RNA
polymerase complex, activating the hupSL transcription. In
the unicellular Gloeothece sp. ATCC 27152, the potential
NtcA-binding site is centered at � 41.5 bp with respect to
the tsp in place of the � 35 box, like in the canonical NtcA-
activated promoters with the consensus sequence signature
GTAN8TAC (Herrero et al., 2001), a structure similar to that
of class II bacterial promoters activated by catabolite acti-
vator protein (CAP). In L. majuscula and N. punctiforme, the
NtcA-binding sites were found to be centered at positions
� 233.5 and � 258.5, respectively, resembling class I CAP-
dependent promoters (Busby & Ebright, 1999; Herrero
et al., 2001, 2004). These data indicate that the type of the
NtcA-activated promoter (class I vs II) is not correlated to
the strategies used by heterocystous and nonheterocystous
cyanobacteria to separate N2 fixation and photosynthesis. In
the filamentous heterocystous A. variabilis, half of a se-
quence motif identical to the consensus Fnr-binding se-
quence was identified 144-bp upstream of the tsp (Happe
et al., 2000) (Fig. 4). Fnr is a regulator of a fumarate nitrate
reductase, which has been found to be involved in the
regulation of the hyp operon in Escherichia coli (Lutz et al.,
1991), and it is responsible for the induction of several
operons in E. coli grown under anaerobic conditions (Spiro
& Guest, 1990). In A. variabilis, although there is no
rearrangement of the hupL gene, hupSL are expressed in
heterocysts only. These differentiated cells have very low
intracellular O2 pressures which led Happe et al. (2000) to
suggest that the hupSL operon in A. variabilis could be
regulated in a manner similar to that of the anaerobically
induced operons in E. coli.
hyp
NtcA2 NtcA1
hox
Synechocystis sp. PCC 6803
LexA2 LexA1LexA2 LexA1
100 bp
Lyngbya majuscula CCAP 1446/4
hypFNtcA2 NtcA1
LexA
+1
17 bp
+1
ORF
Nostoc punctiforme PCC 73102
21 bp
hoxE
+1
−35 −10−35 −10
−35 −10
−35 −10
−10
−10
−10
168 bp
hup
ATCC 29413
hupS
+1
+1
Fnr103 bp
Gloeothece sp. ATCC 27152
hupS
+1+1
NtcA238 bp
Lyngbya majuscula CCAP 1446/4
hupSNtcA IHF
+1
59 bp
hupSNtcA IHF
Nosctoc punctiforme PCC 73102
259 bp
Anabaena variabilis
hupS
Fig. 4. Promoter regions upstream of hupS, hoxE and hypF in cyanobacteria. The following regions are highlighted: putative NtcA-, IHF-, Fnr- and LexA-
binding sites, the � 10 and � 35 boxes and the transcriptional start points (11). The following ORFs are not to scale. In Nostoc punctiforme, the ORF
represented here is immediately upstream of hypF and in the same direction. Analysis of the available genomes revealed the presence of homologues of
this ORF, in the same position and direction, in other filamentous cyanobacteria, and the encoded proteins can be assigned to COG0583 that includes
transcriptional regulators from the LysR family (Leitao et al., 2006). In Synechocystis sp. PCC 6803 hox promoter region, the two putative pairs of LexA-
binding motifs were identified by two different groups (Gutekunst et al., 2005; Oliveira & Lindblad, 2005).
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
700 P. Tamagnini et al.

The possible interaction between NtcA and the hupSL(W)
promoter regions in cyanobacteria was assessed by perform-
ing band shift assays. These experiments indicate a specific
binding of NtcA to DNA sequences upstream of hupS in the
three cyanobacterial strains tested (Gloeothece sp. ATCC
27152, L. majuscula and N. punctiforme), suggesting, indeed,
the involvement of NtcA in the transcription regulation of
the uptake hydrogenase gene cluster (Lindberg, 2003;
Oliveira et al., 2004; Leitao et al., 2005). The fact that the
transcription of the uptake hydrogenase structural genes
is under the control of the transcriptional regulator that
operates global nitrogen control in cyanobacteria reinforces
the correlation observed between the activity of the
uptake hydrogenase and N2 fixation, already demonstrated
in several filamentous heterocystous cyanobacteria (Hou-
chins, 1984; Wolk et al., 1994; Oxelfelt et al., 1995; Troshina
et al., 1996).
Transcription and expression patterns ofhup genes
The first transcriptional data on cyanobacterial uptake
hydrogenases arose from RT-PCR experiments on Nostoc
sp. PCC 7120, revealing that hupL is expressed only after a
photosynthetic vegetative cell differentiates into a N2-fixing
heterocyst (see above details about the DNA rearrangement
occurring within this strain, Carrasco et al., 1995, 2005).
Subsequent studies with other filamentous heterocystous
strains have shown that hupSL is a transcriptional unit
(Happe et al., 2000; Lindberg et al., 2000), present in cells
grown under N2-fixing conditions (Axelsson et al., 1999;
Happe et al., 2000; Hansel et al., 2001). Non-N2-fixing
cultures of Nostoc muscorum, a strain without the hupL
rearrangement, exhibit no in vivo H2-uptake activity (Ax-
elsson et al., 1999). However, the transfer of N. muscorum
cells from non-N2-fixing (ammonia) to N2-fixing condi-
tions induced the appearance of a transcript (after c. 24 h),
and the relative amounts of transcript increased in parallel
with the H2-uptake activity (Axelsson et al., 1999). A similar
pattern of transcription was observed for A. variabilis
and N. punctiforme, two other strains with noninterrupted
hupL genes (Happe et al., 2000; Hansel et al., 2001). These
authors demonstrated that hupSL transcripts were
missing in A. variabilis and in N. punctiforme cells grown
with ammonia (and in A. variabilis cells grown with
nitrate), but were present in both organisms grown under
N2-fixing conditions.
While the heterocyst provides a microaerobic environ-
ment protecting the oxygen-sensitive nitrogenases and up-
take hydrogenases from the atmospheric and intracellulary
generated oxygen, the nonheterocystous cyanobacteria de-
veloped different approaches. The temporal separation
between photosynthesis (light) and nitrogen-fixation/
hydrogen uptake (dark) seems to be the most common
strategy adopted by the later cyanobacteria (Bergman et al.,
1997; Bohme, 1998; Berman-Frank et al., 2003). In fact, in
the nonheterocystous Gloeothece sp. ATCC 27152 (unicellu-
lar) and L. majuscula (filamentous), grown under nitrogen-
fixing conditions and 12 h light/12 h dark cycles, there is an
evident light/dark regulation with the highest levels of
hupSL(W) transcripts detected during the light phase or in
the transition between the light and dark phase, respectively
(Oliveira et al., 2004; Leitao et al., 2005). It has also been
demonstrated that both organisms exhibit higher hydrogen-
uptake activities during the dark period (in agreement with
the nitrogen fixation rates; see Reade et al., 1999; Lundgren
et al., 2003). In L. majuscula, the increase of the HupL
protein levels coincides with the increase of hydrogenase
uptake activity during the dark phase. In the beginning of
the light phase, no hupSL transcription is detectable, and the
levels of both polypeptides and H2 uptake activity begin to
decline (Leitao et al., 2005). These results suggest that in
L. majuscula, a protein turnover occurs, with degradation
taking place during the light period and de novo synthesis
taking place during the dark phase. The time difference
between the hupSL transcription and the hydrogen uptake
activity, both in Gloeothece sp. ATCC 27152 and L. majuscu-
la, might be due to the complexity of the maturation process
of the uptake hydrogenase. Thus, it is possible that the
translation occurs as soon as the transcript is available, while
the enzyme becomes active only after the maturation
process is completed. The temporal separation between
the photosynthesis and nitrogen fixation/hydrogen uptake
activity may also influence the time lag between transcrip-
tion and activity.
In the presence of combined nitrogen, hupSLW transcrip-
tion is totally repressed in Gloeothece sp. ATCC 27152, while
in L. majuscula the levels of hupSL transcription and
expression are significantly reduced but it is possible to
discern a pattern similar to the one observed in cells grown
under N2-fixing conditions (Oliveira et al., 2004; Leitao
et al., 2005, Ferreira et al., 2007). The results obtained for
L. majuscula under non-N2-fixing conditions could be ex-
plained by the mode of growth of this cyanobacterium, in
which the inner cells are probably not in the same conditions
notably in terms of access to the combined nitrogen.
Besides the source of nitrogen, other factors were proven
to influence the transcription/expression of the cyanobac-
terial uptake hydrogenases. Similar to any NiFe hydroge-
nase, the activity of the cyanobacterial uptake enzyme was
shown to be dependent on nickel availability, and the
addition of external nickel to the growth medium (up to a
certain concentration) increased the uptake hydrogenase
activity in several strains (Xiankong et al., 1984; Daday
et al., 1985; Kumar & Polasa, 1991; Oxelfelt et al., 1995;
Axelsson & Lindblad, 2002). Furthermore, the addition of
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
701Cyanobacterial hydrogenases

exogenous hydrogen was shown to induce hupSL transcrip-
tion and hydrogen uptake activity in N. muscorum and N.
punctiforme (Oxelfelt et al., 1995; Axelsson & Lindblad,
2002), as well as hydrogen uptake activity in Nostoc sp.
PCC 7120 (Houchins & Burris, 1981b). Both cyanobacterial
hydrogenases are affected by the oxygen partial pressure.
Nostoc muscorum and N. punctiforme cultures transferred
from aerobic to anaerobic conditions showed an increase in
both the transcription of hupL and hydrogen uptake activity
(Axelsson & Lindblad, 2002). Similarly, the uptake hydro-
genase activity could be elicited by removing oxygen from
the sparging gas of a culture of Nostoc sp. PCC 7120
(Houchins & Burris, 1981b). The addition of organic carbon
to the culture medium can also influence the hydrogen
uptake activity. Cells of N. punctiforme grown either photo-
or chemoheterotrophically reach both higher nitrogenase
and hydrogen uptake activities than photoautotrophically
grown cells (Oxelfelt et al., 1995). However, the effect of
carbon substrates on the cyanobacterial uptake hydrogenase
activity is difficult to assess, and apparently contradictory
results are reported in the literature (Houchins, 1984;
Kumar et al., 1986; Chen et al., 1989; Margheri et al., 1991).
Bidirectional hydrogenase
The soluble or loosely membrane associated cyanobacterial
bidirectional hydrogenase might be present in both N2- and
non-N2-fixing strains (Tamagnini et al., 2000, 2002). Initi-
ally, the bidirectional hydrogenase was thought to be com-
posed of four subunits (encoded by the hox – hydrogen
oxidation – genes), in which HoxFU constitute the diaphor-
ase part, and HoxYH constitute the hydrogenase part
(Schmitz et al., 1995; Appel & Schulz, 1996; Boison et al.,
1996, 1998; Sheremetieva et al., 2002). However, because
HoxE was shown to copurify with the active bidirectional
enzyme, the cyanobacterial bidirectional hydrogenase is
considered to be a heteropentameric enzyme encoded by
hoxEFUYH, HoxE belonging to the diaphorase part
(Schmitz et al., 2002). Bidirectional hydrogenases with more
than four subunits have also been identified in other
bacteria, such as the photosynthetic purple sulfur bacteria
Thiocapsa roseopersicina and Allochromatium vinosum which
contain heteropentameric cyanobacterial-type bidirectional
hydrogenases (Rakhely et al., 2004; Long et al., 2007), and
Ralstonia eutropha, which possess two HoxI subunits besides
HoxFUYH (Burgdorf et al., 2005). In recent years, the
number of reports showing the presence of a functional
and active bidirectional hydrogenase in cyanobacteria has
increased significantly, ranging from unicellular strains
(Gloeocapsa alpicola CALU 743 – Sheremetieva et al.,
2002; Troshina et al., 2002) to filamentous nonheterocystous
(L. majuscula – Schutz et al., 2004; Leitao et al., 2005;
Arthrospira and Spirulina spp. – Zhang et al., 2005a, b), and
filamentous heterocystous strains (Nostoc spp. – Tamagnini
et al., 2000; Schutz et al., 2004). Furthermore, the increasing
number of cyanobacterial sequenced genomes is contribut-
ing toward a better understanding of both the distribution
and the diversity of this enzyme.
The physiological function of the bidirectional hydroge-
nase in cyanobacteria is not totally clear. It has been
suggested that the enzyme acts as an electron valve during
photosynthesis in Synechocystis sp. PCC 6803. This is based
on the fact that hoxH� mutants are impaired in the oxida-
tion of PSI, have higher fluorescence of PSII and have
different transcript levels of the photosynthetic genes psbA,
psaA and petB when compared with the wild type (Appel
et al., 2000). The enzyme has also been proposed to play a
role in fermentation functioning as a mediator in the release
of excess reducing power under anaerobic conditions (Stal &
Moezelaar, 1997; Troshina et al., 2002). Furthermore, it has
been suggested previously that the bidirectional hydroge-
nase could be part of the respiratory complex I (Appel &
Schulz, 1996; Schmitz & Bothe, 1996), because only 11
subunits out of 14 conserved subunits of the prokaryotic
complex I have been identified in cyanobacteria. Some of
the subunits of the bidirectional hydrogenase indeed show
sequence similarities with the missing subunits of the
respiratory complex I (Schmitz et al., 1995). However, the
bidirectional hydrogenase has been demonstrated to be
absent from several cyanobacterial strains (Tamagnini et al.,
1997, 2000; Schutz et al., 2004; Ludwig et al., 2006). More-
over, N. punctiforme, a strain naturally lacking the bidirec-
tional hydrogenase (Tamagnini et al., 1997), has rates of
respiration comparable to cyanobacteria containing the
bidirectional hydrogenase (Boison et al., 1999). In addition,
mutants of hoxU in Synechococcus sp. PCC 6301 (former
Anacystis nidulans) (Boison et al., 1998) and hoxEF in
Synechocystis sp. PCC 6803 (Howitt & Vermaas, 1999)
showed nonimpaired respiratory O2 uptake while being
affected in H2 evolution. Furthermore, inactivation of hoxH
in Synechocystis sp. PCC 6803 and Nostoc sp. PCC 7120
resulted only in a small decrease in the growth rate com-
pared with the respective wild types (Appel et al.,
2000; Masukawa et al., 2002). Taking into account all the
data, it seems that in general the bidirectional hydrogenase
does not play an essential role for cell survival in the strains
where it is present.
Attempting to shed some light on the physiological
function of the bidirectional hydrogenase, Cournac et al.
(2004) demonstrated that the bidirectional hydrogenase in
Synechocystis sp. PCC 6803 is insensitive to light, reversibly
inactivated by O2 and can be quickly reactivated by NADH
or NADPH. This work also reported H2 evolution by cells
incubated anaerobically in the dark, after an adaptation
period. This dark H2 evolution was enhanced by exogen-
ously added glucose and resulted from the oxidation of
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
702 P. Tamagnini et al.

NAD(P)H produced by fermentation reactions. Upon illu-
mination, a short (o 30 s) burst of H2 output was observed,
followed by rapid H2 uptake, and a concomitant decrease in
CO2 concentration in the cyanobacterial cell suspension,
which were both linked to photosynthetic electron transport
in the thylakoids (Cournac et al., 2004). Moreover, in this
experimental setup, in anoxia (or microaerobiosis) and in
the presence of H2, H2 uptake was of the same magnitude as
photosynthetic activity and could therefore contribute sig-
nificantly to CO2 fixation. Therefore, although the bidirec-
tional hydrogenase in Synechocystis sp. PCC 6803 is
constitutively expressed in the presence of O2 (Appel et al.,
2000), it probably plays a role mainly under anaerobic or
microaerobic conditions, and at the onset of light before the
enzyme is inactivated by photosynthetic O2. In the ndhB
mutant M55, which is defective in the type I NADPH-
dehydrogenase complex (NDH-1) and produces only low
amounts of O2 in the light, H2 uptake was negligible during
dark-to-light transitions, allowing several minutes of con-
tinuous H2 production. It was further shown that two
pathways of electron supply for H2 production operate in
M55, namely photolysis of water at the level of photosystem
II and carbohydrate-mediated reduction of the plastoqui-
none pool. When comparing the features of the Synechocys-
tis sp. PCC 6803 hydrogenase with those of the homologous
NAD1-dependent hydrogenase of R. eutropha, despite se-
quence homologies between the two enzymes, their char-
acteristics are not identical, which might indicate that this
enzyme might have slightly different functions in different
organisms (Cournac et al., 2004).
If the function of the bidirectional hydrogenase is still open
to debate, its subcellular localization is not less controversial.
The bidirectional hydrogenase can be found in both the
heterocysts and the vegetative cells (Hallenbeck & Benemann,
1978; Houchins & Burris, 1981a), and in Nostoc sp. PCC 7120
appears in the soluble fraction after cell disruption, and
consequently has been considered to be a soluble enzyme
(Houchins & Burris, 1981b). Nevertheless, investigations in
other cyanobacteria suggest a weak association of the bidirec-
tional hydrogenase with cell membranes: in A. variabilis and
Synechocystis sp. PCC 6803, an association with the thylakoid
membrane was proposed (Serebriakova et al., 1994; Appel
et al., 2000), while in Synechococcus sp. PCC 6301 immuno-
logical data implied an association with the cytoplasmic
membrane (Kentemich et al., 1989, 1991).
Physical organization of hox genes and thecorresponding proteins
In cyanobacteria, the structural genes encoding the bidirec-
tional hydrogenase are organized in a dissimilar way (see
Fig. 2). In some strains (e.g. Synechocystis sp. PCC 6803 and
A. variabilis), the hox genes are localized in one cluster,
although interspersed with different ORFs at diverse posi-
tions. In other cases, the hox genes are found in two different
clusters separated by several kilobase (c. 333 and 8.8 kb in
Synechococcus sp. PCC 6301 and Nostoc sp. PCC 7120,
respectively). Despite this fact, the similarities at the de-
duced amino acid level of their homologous hydrogenase
proteins range between 55% and 81%.
The bidirectional hydrogenase has been purified from
several cyanobacterial strains: A. cylindrica (Hallenbeck &
Benemann, 1978), Spirulina maxima (Llama et al., 1979),
Microcystis aeruginosa (Asada et al., 1987), Synechococcus sp.
PCC 6301 (Schmitz et al., 1995, 2002) and Synechocystis sp.
PCC 6803 (Schmitz et al., 2002), but the data collected by
Schmitz et al. (2002) finally helped to clarify the picture of
the subunit composition and molecular mass of the cyano-
bacterial bidirectional hydrogenase. Thus, it is widely ac-
cepted that the bidirectional hydrogenase is composed of
five subunits, HoxE, HoxF, HoxU, HoxY and HoxH, with
apparent molecular weights of c. 20, 61, 28, 24 and 49 kDa,
respectively. The molecular weight of the native protein
(375 kDa) indicates a dimeric assembly of the enzyme
complex, Hox(EFUYH)2 (Schmitz et al., 2002).
Similar to the uptake hydrogenase, the large subunit of the
hydrogenase dimer (HoxH) harbors the active metal center
containing nickel and iron. The two metal atoms are held in
close proximity by two disulfide bridges provided by two
cysteine residues of the protein. The iron has two cyanide ions
and one carbon monoxide as ligands, whereas the nickel ion
is coordinated by two additional cysteines (Volbeda et al.,
1995). The small subunit of the hydrogenase dimer (HoxY),
and the different components of the diaphorase part of the
bidirectional hydrogenase (HoxF and HoxU) also contain
several conserved cysteine residues putatively involved in the
coordination of FeS clusters (Schmitz et al., 2002; for a review,
see Tamagnini et al., 2002). In addition, in the middle region
of HoxF, typical glycine-rich binding sites for NAD1
(GxGxxGxxxG) and flavin mononucleotide (GxGxxxxGx10
GxxG) can be identified (Schmitz et al., 1995). HoxE may be
involved as a bridging subunit in membrane attachment.
Moreover, a functional role in electron transport directed to
membrane components, as demonstrated experimentally for
the Hox-hydrogenase of Thiocapsa roseopersicina (Rakhely
et al., 2004), could be considered because sequence motifs for
binding of an additional FeS cluster are present in this gene
(Schmitz et al., 2002).
hox promoter regions and transcriptionalregulators
The information about the transcription and regulation of
the hox genes is limited in cyanobacteria, but the under-
standing of these mechanisms is now emerging. Recent
studies showed that the hox genes in Synechocystis
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
703Cyanobacterial hydrogenases

sp. PCC 6803 are transcribed as a single operon (Gutekunst
et al., 2005; Oliveira & Lindblad, 2005; Antal et al., 2006)
with the transcription start point located 168-bp upstream
of the hoxE start codon (Gutekunst et al., 2005; Oliveira &
Lindblad, 2005).
Up to now, only one regulator – LexA – has been proven
to bind and regulate the transcription of the hox genes in
cyanobacteria. Two independent studies (Gutekunst et al.,
2005; Oliveira & Lindblad, 2005) demonstrated an interac-
tion between LexA and the promoter region of the bidirec-
tional hydrogenase in Synechocystis sp. PCC 6803. However,
two distinct regions were analyzed and both were demon-
strated to be targets for this interaction. Oliveira & Lindblad
(2005) showed that LexA binds to a region located between
the nucleotides � 198 and � 338 bp, respective to transla-
tional start point, while Gutekunst et al. (2005) found that
LexA interacts further upstream on the hox promoter, at the
positions � 592 to � 690 bp, in relation to the hoxE ATG
codon (see Fig. 4). Furthermore, a LexA-depleted mutant
showed a reduced hydrogenase activity compared with the
wild-type, suggesting that LexA works as a transcription
activator of the hox genes in Synechocystis sp. PCC 6803
(Gutekunst et al., 2005). Synechocystis sp. PCC 6803 LexA
has been detected in different proteomic studies (Wang
et al., 2000; Gan et al., 2005; Srivastava et al., 2005; Fulda
et al., 2006; Kurian et al., 2006; Slabas et al., 2006), and its
transcript has also been identified in microarray experi-
ments (Hihara et al., 2001; Kamei et al., 2001; Li et al., 2004;
Singh et al., 2004; Tu et al., 2004; Shapiguzov et al., 2005).
Interestingly, in some proteomic studies, LexA has been
identified in association with thylakoid membrane fractions
(Wang et al., 2000; Srivastava et al., 2005), which represents
an unexpected location for a transcription regulator.
Based on the observations that the bidirectional hydro-
genase activity is directly affected by the redox status of the
cell, either in photosynthesis or in fermentation, and that
the regulation of the hox gene expression can be operated by
LexA, hypothesis was recently put forward on the direct
involvement of the transcription regulator LexA as a med-
iator of the redox-responsive regulation of the hox gene
expression in Synechocystis sp. PCC 6803 (Antal et al., 2006).
Interestingly, the expression of the cyanobacterial DEAD-
box RNA helicase, crhR, which is regulated in response to
conditions that elicit reduction of the photosynthetic elec-
tron transport chain, was recently shown as being directly
controlled by LexA in Synechocystis sp. PCC 6803 (Patter-
son-Fortin et al., 2006). Transcript analysis indicated that
lexA and crhR are divergently expressed, with the respective
transcripts accumulating differently under conditions,
which, respectively, oxidize and reduce the electron trans-
port chain, suggesting that LexA works as a repressor of the
crhR transcription (Patterson-Fortin et al., 2006). Although
these results are in agreement with the initial hypothesis, the
signal transduction pathways directly or indirectly involved
in the regulation of LexA, and consequently its downstream
targets, definitely require further investigation.
Transcription and expression patterns of hoxgenes
The number of studies focusing on the transcription and
regulation of the hox genes in cyanobacteria is scarce.
Nevertheless, transcripts of the bidirectional hydrogenase
have been shown to be present in NH41-grown filaments,
and in both vegetative cells and heterocysts under nitrogen-
fixing conditions in A. variabilis (Boison et al., 2000). In
addition, hoxFUYH were shown to be transcribed as a single
unit together with other two ORFs with unknown function.
However, it should be kept in mind that these experiments
were performed using RT-PCR and do not exclude addi-
tional promoters within the operon (Boison et al., 2000). On
the other hand, in the unicellular Synechococcus sp. PCC
6301 and Synechococcus sp. PCC 7942 the hox genes are
located apart and give rise to two different transcripts
(Boison et al., 2000; Schmitz et al., 2001). While hoxEF are
cotranscribed in both strains, the second transcript is
constituted by hoxUYH together with hoxW, hypA and hypB
in Synechococcus sp. PCC 6301 (Boison et al., 2000), and by
hoxUYHW only in Synechococcus sp. PCC 7942 (Schmitz
et al., 2001). For the last strain, using real-time PCR and
reporter gene constructs, it was suggested that a second
promoter might be present between hoxH and hoxW
(Schmitz et al., 2001). Furthermore, it was demonstrated
that the hox genes have a circadian clock expression
(Schmitz et al., 2001), a fact that has also been demonstrated
for hoxE in Synechocystis sp. PCC 6803 (Kucho et al., 2005).
Very few studies focusing on the regulation of hox genes
transcription have been performed in cyanobacteria. Analy-
sis of the transcription of hoxY and hoxH in G. alpicola,
under combined nitrogen-limiting growth conditions, de-
monstrated an increase in the enzyme activity, but no
regulation at the transcript level (Sheremetieva et al., 2002).
In contrast, Northern blot analyses of the hox genes expres-
sion in Synechocystis sp. PCC 6803 under combined nitro-
gen-limiting growth conditions demonstrated an increase in
transcription (Antal et al., 2006), followed by an increase in
enzyme activity (T.K. Antal, P. Oliveira & P. Lindblad,
unpublished data). A similar increase in the hox genes
transcription has also been observed with microarray in
Synechocystis sp. PCC 6803 cells undergoing nitrogen starva-
tion for 4 h (Osanai et al., 2006 – supplementary material).
Furthermore, a transfer to a low level of oxygen in
A. variabilis induced both the enzyme activity as well as the
relative amount of hoxH (Sheremetieva et al., 2002). It has
long been demonstrated that microaerobic/anaerobic con-
ditions influence hox transcription and bidirectional
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
704 P. Tamagnini et al.

hydrogenase activity in heterocystous cyanobacteria (Hou-
chins & Burris, 1981a; Houchins, 1984; Serebryakova et al.,
1994; Schmitz & Bothe, 1996; Axelsson & Lindblad, 2002;
Sheremetieva et al., 2002). The bidirectional hydrogenase in
Nostoc sp. PCC 7120 is active in both vegetative cells and in
heterocysts in aerobically grown filaments, with heterocysts
having several fold more activity than vegetative cells. When
the filaments were transferred to anaerobic conditions, the
activity of the bidirectional hydrogenase increased by about
two orders of magnitude with approximately the same
activity levels in both types of cells (Houchins & Burris,
1981a). Similar results have been observed in A. variabilis
(Serebryakova et al., 1994). In contrast to the filamentous
cyanobacteria, the activity of the bidirectional hydrogenase
in the unicellular G. alpicola is not directly dependent on
oxygen (Troshina et al., 2002). Higher activity is observed
under nitrogen starvation and low light, and it was sug-
gested that the bidirectional hydrogenase could act as an
alternative electron donor to PSI after inactivation of PSII
due to nitrogen starvation. Under dark anoxic conditions,
the unicellular cyanobacterium G. alpicola produces H2
catalyzed by the bidirectional hydrogenase (Troshina et al.,
2002). In addition, the unicellular strain Chroococcidiopsis
thermalis CALU 758 contains a bidirectional hydrogenase
with some catalytic properties more related to an uptake
hydrogenase, i.e. not inducible under anaerobic conditions
or under nitrate-starving conditions (Serebryakova et al.,
2000).
Because the bidirectional hydrogenase in cyanobacteria is
a metal-dependent enzyme, containing nickel and iron in
its active center and FeS clusters involved in electron
transfer, the availability of these elements in the growing
medium has been a subject of research. Axelsson & Lindblad
(2002) showed that in the heterocystous N. muscorum
CCAP 1453/12, the addition of external nickel to the
growing medium increased the mRNA abundance of hoxH
(monitored by RT-PCR). Making use of reporter gene
constructs, Gutekunst et al. (2006) were able to show that
the transcription of the bidirectional hydrogenase genes in
Synechocystis sp. PCC 6803 increased with lower concentra-
tions of iron, the signal being 10 times higher in cells grown
with 0.22mM iron compared with nonstarved cells. In the
same work, measurements of the hydrogenase activity
revealed a reduction of the enzyme activity alongside the
decrease in the iron concentration. The increase in tran-
scription of the hox genes, when the cells undergo iron
starvation, might be a feedback mechanism to compensate
for the lack of functionally active enzyme (Gutekunst et al.,
2006). The availability of sulfur in the growth medium has
also been shown to influence the bidirectional hydrogenase
activity in Synechocystis sp. PCC 6803 and G. alpicola (Antal
& Lindblad, 2005). Both strains showed an enhanced (more
than fourfold) H2 production capacity during fermentation
via hydrogenase, when grown under sulfur starvation con-
ditions.
Although the understanding of the regulation and the
physiological role of the bidirectional hydrogenase is becom-
ing clearer, intriguing recent results on the hydrogenase
activity from two substrains of Synechocystis sp. PCC 6803
have shown that they do not have comparable values
(Gutekunst et al., 2006). The authors suggested that these
phenotypic differences in the hydrogenase activity might be
due to divergences in their metabolism. In fact, maintenance
of these strains in culture collections, or under various
laboratory conditions, may have led to spontaneous muta-
tions and unintended selective pressures, resulting in the
observable variations in each subculture (Ikeuchi & Tabata,
2001). Therefore, special care must be taken when interpret-
ing results coming from different laboratories and different
cyanobacterial strains, even from the same strain, but
cultured in different laboratories.
Maturation of cyanobacterialhydrogenases
The biosynthesis/maturation of NiFe-hydrogenases is a
highly complex process requiring at least seven core proteins
for the incorporation of the metal ions and CO and CN
ligands in to the active center, the orientation of the FeS
clusters within the small subunit and the cleavage of the C-
terminus as the final step in the maturation of the large
subunit (for a recent review on this subject, see Bock et al.,
2006, and also Casalot & Rousset, 2001; Blokesch et al., 2002;
Mulrooney & Hausinger, 2003; Kuchar & Hausinger 2004;
Vignais & Colbeau, 2004; Theodoratou et al., 2005). The
genes encoding the proteins involved in the maturation of
hydrogenases were firstly characterized for E. coli, and while
most of the Hyp proteins affect hydrogenases pleiotropically,
the large subunit of each hydrogenase is proteolytically
processed by a specific endopeptidase (Lutz et al., 1991;
Jacobi et al., 1992; Menon et al., 1994; Rossmann et al., 1995;
Theodoratou et al., 2005; Bock et al., 2006). Homologues of
the hyp genes are present in all organisms capable of forming
NiFe hydrogenases. Although little is known about the
biosynthesis/maturation of the cyanobacterial hydrogenases,
several genes presumably involved in this process have been
identified clustered or scattered throughout the genomes of
several cyanobacterial strains (Boison et al., 1996; Gubili &
Borthakur, 1996, 1998; Kaneko et al., 1996; Sakamoto et al.,
1998; Hansel et al., 2001; Tamagnini et al., 2002; Wunschiers
et al., 2003; Hoffmann et al., 2006; Leitao et al., 2006). The
presence of a single copy of most of the hyp genes (hypFC-
DEAB) in the genome of cyanobacteria, regardless of
possessing only the uptake hydrogenase (e.g. N. puncti-
forme), the bidirectional hydrogenase (e.g. Synechocystis sp.
PCC 6803) or both enzymes (e.g. Nostoc sp. PCC 7120)
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
705Cyanobacterial hydrogenases

suggests that they might be responsible for the maturation
of both hydrogenases. In contrast, the genes encoding for the
putative hydrogenase C-terminal endopeptidases – hupW
and hoxW – were identified and seem to be specific for the
cyanobacterial uptake and the bidirectional hydrogenase,
respectively, resembling the situation in other organisms
(Wunschiers et al., 2003; Oliveira et al., 2004; Leitao et al.,
2006).
Physical organization of hyp genes and thecorresponding proteins
The hyp genes in cyanobacteria are frequently clustered and
in the vicinity of the structural genes of one of the hydro-
genases (Fig. 2), with a well-known exception – the uni-
cellular non-N2-fixing Synechocystis sp. PCC 6803 – in which
the hypABCDEF genes are scattered throughout the genome.
Still, in this organism the homologs hypA2 and hypB2 are
clustered (Kaneko et al., 1996), but these two do not seem to
play a key role in the maturation of the bidirectional
hydrogenase (Hoffmann et al., 2006). In three Synechococ-
cus, closely related strains (Synechococcus elongatus PCC
6301, Synechococcus elongatus PCC 7942 and Synechococcus
sp. PCC 7002) hypABFC are together and downstream of
hox genes, while hypD and hypE are apart in the two first
organisms (Boison et al., 1996). In the heterocystous strains,
N. punctiforme, Nostoc sp. PCC 7120 and A. variabilis and in
the N2-fixing but nonheterocystous L. majuscula, the hyp
genes are located in a cluster with all genes orientd in the
same direction, and relatively close to the uptake hydro-
genase structural genes, although in the opposite direction
(Gubili & Borthakur, 1998; Hansel et al., 2001; Leitao et al.,
2006). However, this organization does not constitute a
pattern for N2-fixing strains, because it contrasts with the
organization observed for other nonheterocystous strains,
such as the filamentous T. erythraeum, in which hyp genes
are located much further upstream of hupSL (ca. 589 kb),
and the unicellular Crocosphaera watsonii WH 8501, in
which the genes are scattered over the genome resembling
the non-N2-fixing Synechocystis sp. PCC 6803. When the
genes are grouped, the order varies in non-N2-fixing com-
pared with N2-fixing strains being hypABFC and hypFC-
DEAB, respectively. In the former case, ORFs interspersed
with the hyp genes can be found in several organisms.
The putative cyanobacterial Hyp proteins possess con-
served motifs and may fulfill functions similar to the
corresponding proteins in other organisms (Tamagnini
et al., 2002; Vignais & Colbeau, 2004; Hoffmann et al.,
2006; Leitao et al., 2006). It is believed that from the two
metal ions present in the active center of a NiFe hydro-
genase, Fe is the first to be incorporated into the enzyme.
HypF and HypE are the proteins involved in the synthesis of
the CN, and maybe the CO, ligands of iron (Paschos et al.,
2001, 2002; Bock et al., 2006). HypF accepts carbamoyl
phosphate (CP) as a substrate, catalyzes a CP-dependent
hydrolysis of ATP into AMP and inorganic phosphate (PPi)
and forms an adenylated CP derivative. The carbamoyl
group of CP is transferred to the cysteine at the C-terminus
of HypE (Paschos et al., 2002; Reissmann et al., 2003). It was
demonstrated in vitro that the CN group from HypE-
thiocyanate can be transferred to the complex HypC plus
HypD (Blokesch et al., 2004a). Because the transfer of
the ligands to the iron requires the input of two electrons
(Blokesch & Bock, 2002), HypD is proposed to be the one
involved in this process, given that among all the maturation
proteins it is the only one with a redox-active cofactor
(Blokesch et al., 2004a; Roseboom et al., 2005). On the other
hand, HypC is a small chaperone-like protein that was
shown to form a complex with HypD (Blokesch & Bock,
2002) and to interact with the large subunit of the hydro-
genase (Magalon & Bock 2000a; Casalot & Rousset 2001).
Probably, the liganding of the iron takes place at the
HypC–HypD complex (Blokesch et al., 2002, 2004a), and
the interaction between HypC and the precursor of the large
subunit leads to the liberation of HypD (Blokesch et al.,
2004a; Blokesch & Bock, 2006). Subsequently, the liganded
Fe is transferred to the precursor of the hydrogenase large
subunit (Blokesch & Bock, 2006), and HypC remains
attached to the large subunit, maintaining it in an open
conformation, allowing the insertion of nickel. This step
requires the presence of HypA and HypB (Jacobi et al., 1992;
Olson et al., 2001). HypA is a zinc-containing protein that
binds nickel (Mehta et al., 2003; Blokesch et al., 2004b), and
HypB is a GTPase that probably plays a dual function: nickel
storage and nickel insertation (Maier et al., 1993, 1995). It is
thought that HypA functions as a nickel chaperone and that
HypB acts as a regulator, controlling the donation of the
metal to the apoprotein or the release of the nickel-free
chaperone (Blokesch et al., 2004b). After both metals have
been coordinated to the precursor of the large subunit, the
C–terminal extension is accessible and can be removed by
the specific endopeptidase. The cleavage can only occur after
HypC dissociation from the precursor of the large subunit
that already contains Ni and Fe(CO)(CN�)2 centers (Maga-
lon & Bock, 2000a, b), because the endopeptidase uses Ni
as a recognition motif. Following the cleavage of the
C-terminal tail from the large hydrogenase subunit, the
mature large subunit can be assembled, with the mature
small subunit forming the functional enzyme (Magalon &
Bock, 2000a). Maturation of the small subunit should occur
in parallel, and independently from the large subunit
maturation. The knowledge about this process is still scarce,
although recent studies highlighted at least four gene
products (encoded within the hup cluster, and downstream
of uptake hydrogenase structural genes) that are required for
the maturation of the small subunit of the NiFe
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
706 P. Tamagnini et al.

hydrogenases of Rhizobium leguminosarum bv. viciae (Man-
yani et al., 2005; Bock et al., 2006). In cyanobacteria, several
additional ORFs are commonly present near hyp or hup
genes (Leitao et al., 2006). The consistent location of these
ORFs might indicate that their proteins may have a role in
the uptake hydrogenase maturation process and/or its
regulation, notably regarding the small subunit.
hyp promoter regions and transcriptionalregulators
As mentioned above, the hyp genes can be found clustered or
scattered throughout the genome of cyanobacteria (Fig. 2).
Analysis of the hyp cluster promoter region of N. puncti-
forme revealed the presence of � 10 and � 35 elements, and
putative binding sites for NtcA (Hansel et al., 2001; Fig. 4).
Similarly, in the corresponding region of L. majuscula the
presence of a � 10 box, and two putative NtcA-binding sites
could be identified. In this organism, a clear � 35 box is not
present, but it should be taken into account that its sequence
is highly variable. Furthermore, a putative LexA-binding site
was also found in L. majuscula (Leitao et al., 2006; Fig. 4).
The transcriptional regulators NtcA and LexA were shown
to bind to the promoter regions of the hup and the hox
genes, suggesting their involvement in the regulation of the
uptake and bidirectional hydrogenase, respectively (see
above, Lindberg, 2003; Oliveira et al., 2004; Gutekunst
et al., 2005; Leitao et al., 2005; Oliveira & Lindblad, 2005).
The presence of putative binding sites for both transcrip-
tional factors NtcA and LexA within the hyp operon
promoter region, and preliminary results from electro-
phoretic mobility shift assays (Ferreira et al., 2007) suggest
the involvement of these proteins in the transcriptional
regulation of hyp genes in L. majuscula, a cyanobacterium
containing both hydrogenases. These data reinforce the
hypothesis that the Hyp proteins might be implicated in
the maturation/regulation of both hydrogenases, and raise
the hypothesis that the transcription of hyp genes in
cyanobacteria containing both hydrogenases could be under
the control of different transcriptional regulators, e.g. NtcA
and LexA.
Transcription and expression patterns of hypgenes
In the heterocystous N. punctiforme, the hup and hyp genes
are transcribed under N2-fixing but not under non-N2-
fixing growth conditions (Hansel et al., 2001). One should
bear in mind that N. punctiforme contains only one hydro-
genase (the uptake enzyme), and that in this organism both
the transcription of hupL and the H2 uptake activity are
repressed when combined nitrogen is present in the growth
medium (Oxelfelt et al., 1995; Hansel et al., 2001).
In the unicellular non-N2-fixing Synechocystis sp. PCC
6803, a cyanobacterium harboring only the bidirectional
hydrogenase, deletion and insertion mutants of hypA1, B1,
C, D, E and F showed no hydrogenase activity. Moreover, the
complementation of each of the above hyp- inactivated
genes restored the bidirectional hydrogenase activity to the
wild-type level in the respective mutants (Hoffmann et al.,
2006). In contrast, the deletion of the homologues hypA2
and hypB2 had no effect on the bidirectional hydrogenase
activity even though they are transcribed in the wild type,
demonstrating that the products of these genes are not
actively involved in the maturation process of the bidirec-
tional hydrogenase (Hoffmann et al., 2006).
Hydrogenase-specific endopeptidases geneshupW and hoxW, and corresponding proteins
The last step in the processing of the large subunit of NiFe-
hydrogenases is the cleavage of a C-terminal peptide, which,
most likely, allows a structural reorganization of the mole-
cule and the consequent assembly of the holoenzyme. After
both metals have been inserted into the apoprotein precur-
sor of the large subunit, the C-terminal extension is acces-
sible and can be removed by the specific endopeptidase
(Theodoratou et al., 2005; Bock et al., 2006). This process
triggers a conformational switch in which the free thiol of
the most C-terminally located cysteine residue closes the
bridge between the two metals resulting in the formation of
the complete heterobinuclear center (Maier & Bock, 1996;
Magalon & Bock, 2000a; Theodoratou et al., 2005; Bock
et al., 2006). The peptidase cleaves the hydrogenase large
subunit precursor after a histidine or an arginine residue at
the C-terminal consensus motif DPCxxCxx(H/R), liberating
a short polypeptide that varies considerably both in length
and sequence among different organisms (Wunschiers et al.,
2003). It has been postulated that the endopeptidase recog-
nizes its substrate, the nickel-containing hydrogenase pre-
cursor, at least in part via the metal that is coordinated by
three thiolates, and binds to the exposed C-terminal domain
(Theodoratou et al., 2000a, b, 2005 and Fig. 5). In addition,
the endopeptidase interacts with a structural domain to
which both the mature part of the large subunit and the C-
terminal extension contribute. Therefore, it is believed that
the recognition of the hydrogenase by the endopeptidase
does not depend on the cleavage site consensus sequence but
is mediated by the overall three-dimensional hydrogenase
and peptidase protein structures (Theodoratou et al.,
2000a, b). After the proteolytic cleavage, the mature large
hydrogenase subunit assembles with the small subunit and
eventually the enzyme becomes active.
In cyanobacteria hydrogenase large subunits, the C-
terminal consensus motif [DPCxxCxx(H/R)] was found in
all the deduced amino acid sequences; however, in the
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
707Cyanobacterial hydrogenases

uptake hydrogenase large subunits (HupL) the neutral pro-
line (P) at position 2 of the cutting site motif is exchanged
for an uncharged polar serine (S). The sequence of the
cutting site motif is totally conserved for each of the
cyanobacterial hydrogenases large subunits: HoxH (bidirec-
tional hydrogenase) – DPCLSCSTH; HupL (uptake hydro-
genase) – DSCLVCTVH (see Wunschiers et al., 2003 and
Fig. 5). The putative cleaved C-terminal polypeptide varies
in length (25–32 aa residues) and sequence (10–96% simi-
larity) for HoxH, while for HupL the polypeptide always has
the same length (16 aa residues) and is highly conserved for
all the deduced sequences [AHDAKTG(E/K)ELARFRT(A/
N/S)].
In cyanobacteria, the genes encoding for the putative
hydrogenase-specific C-terminal endopeptidases were iden-
tified and named hupW and hoxW for the gene encoding the
enzyme processing the uptake and the bidirectional hydro-
genase, respectively (Kaneko et al., 1995, 2001; Boison et al.,
2000; Schmitz et al., 2001; Wunschiers et al., 2003; Oliveira
et al., 2004; Leitao et al., 2005).
The position of hupW and hoxW in the cyanobacterial
chromosome is rather variable; however, in several cases
hupW is in the vicinity and in the same direction of hupSL
(uptake hydrogenase structural genes). In the nonheterocys-
tous Gloeothece sp. ATCC 27152 and T. erythraeum, hupW is
even the ORF located immediately downstream of hupL, and
was shown to be cotranscribed with hupSL in Gloeothece
sp. ATCC 27152 (Oliveira et al., 2004). In contrast, in the
heterocystous strains A. variabilis, Nostoc sp. PCC 7120 and
N. punctiforme, hupW is not part of any known hydrogenase
cluster (Fig. 2), and it was shown to be transcribed under
N2- and non-N2-fixing conditions in the last two strains
(Wunschiers et al., 2003). These authors postulated that the
transcription of hupW under conditions in which the
transcripts of the uptake hydrogenase structural genes could
not be detected (presence of ammonia) could imply that
hupW is constitutively expressed. Taking into account all the
available data, it is not yet possible to establish whether the
expression of hupW is or is not constitutive or whether this
depends on the strain/existence of cell differentiation.
Similar to what happens for hupW, analysis of the
available cyanobacterial genomic sequences revealed that
the position and orientation of hoxW in the chromosome is
also variable but, in most of the cases, hoxW is downstream
of hoxH, one of the bidirectional hydrogenase structural
genes (Fig. 2). RT-PCR experiments indicate that in the
unicellular non-N2-fixing Synechococcus sp. PCC 6301,
hoxW is part of a polycistronic message containing hoxUYH-
WhypAB (Boison et al., 2000), while in Synechococcus sp.
PCC 7942 it was demonstrated that although hoxW consti-
tute a unit together with hoxUYH, it is mainly expressed by
its own promoter (Schmitz et al., 2001). In the heterocystous
Nostoc sp. PCC 7120, similar to hupW, hoxW is transcribed
under both N2- and non-N2-fixing conditions (Wunschiers
et al., 2003). Although some data indicate that endo-
peptidases transcripts are present when the corresponding
hydrogenase large subunit transcript is absent, and it has
been proposed that their expression is independently
regulated from the expression of both the hydrogenase
structural and the other accessory genes in cyanobacteria
(Wunschiers et al., 2003), it is premature to make any
general conclusion.
To date, two different hydrogenase specific-endopepti-
dases have been purified and studied, namely HycI and
HybD from E. coli (Rossmann et al., 1995; Fritsche et al.,
1999). Both are monomeric proteins of a molecular mass of
Endo-peptidase
C - S
DPCxx
Cxx
(H/R
)
S
HoxH DPCLSCSTH 25-32 a.a.
HupL D
Endo-
CO CN CN
C - S
DPCxx
Cxx
(H/R
)
COOH
Large Subunit
C
CO CN CN
C S - C
S - C
SmallSubunit
Large Subunit
S
DSCLVCTVH 16 a.a.
- S
- S
Fig. 5. Schematic representation of the putative final step of the maturation process of the Ni–Fe hydrogenases large subunit: cleavage of a small
peptide by a specific endopeptidase, followed by a conformational change that encloses the bimetallic center. This structural reorganization of the large
subunit will allow the consequent assembly of the holoenzyme. In the large subunits of cyanobacterial hydrogenases – HoxH (bidirectional hydrogenase)
and HupL (uptake hydrogenase) – the C-terminal consensus motif DPCxxCxx(H/R) was found in all the deduced sequences, but in HupL the proline is
exchanged by a serine (see box). The putative cleaved polypeptide varies in length and sequence for HoxH, while for HupL is always has the same length
and is highly conserved.
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
708 P. Tamagnini et al.

c. 17 kDa, and they are devoid of metal or other cofactors.
Alignments of the amino acid sequences showed that
hydrogenase- specific C-terminal endopeptidases share low
sequence similarity, with only a few positions fully con-
served (Theodoratou et al., 2005). As a general feature, they
have three highly conserved amino acid residues (Glu, Asp
and His) that, most likely, have the function of interacting
with the nickel in the hydrogenase large subunit precursor
(Theodoratou et al., 2000a). The alignment of the putative
cyanobacterial endopeptidases with the corresponding pro-
teins from E. coli clearly shows that although the amino acid
sequence identity is low, they are indeed structurally related
(67–77% structural identity) (Wunschiers et al., 2003).
Phylogenetic analysis
Recently, the phylogenetic origin of cyanobacterial and algal
hydrogenases was analyzed (Ludwig et al., 2006), leading to
the conclusion that Chloroflexus is probably the closest
ancestor of cyanobacteria. In all cyanobacterial genomes
sequenced to date, and in the genome of Chloroflexus, the
two hydrogenase operons – hup and hox – are widely
separated on the chromosome, rendering simultaneous gene
transfer unlikely. The authors claim that the current dis-
tribution of the hydrogenases in cyanobacterial strains
probably reflects a differential loss of the genes from their
last common ancestor, and that the two sets of genes,
encoding the uptake and bidirectional, were either kept in
the genome or lost differentially in the different strains
according to their ecological needs or constraints. Although
the phylogenetic analysis of Ludwig et al. (2006) clearly
demonstrated the monophyly of cyanobacteria, and their
relationship with other photosynthetic bacteria, relation-
ships within the cyanobacteria were poorly resolved using
HupL sequences. This is probably related to the difficulties
of aligning the cyanobacteria with the other highly divergent
lineages, and acerbated by the low number of sequences
available and long branches leading to terminal nodes. The
high variability of the sequences also means that more
distant bacterial outgroups cannot be unambiguously
aligned. However, analysis solely within the cyanobacteria
for both HupS and HupL is less complex, because analysis of
the predicted proteins demonstrates that in HupS the
number of residues in all known cyanobacteria is constant
(320 aa), while HupL generally has 531 aa, with the excep-
tion of the filamentous nonheterocystous strains L. majus-
cula and L. aestuarii with six extra (one insertion of 5 aa and
another of one), and T. erythraeum with three extra, coin-
ciding with the position of the five inserts in Lyngbya spp.
Owing to this relatively conserved identity, alignment of the
amino acids for phylogenetic analysis was facile.
Amino acid sequences were analyzed under the criterion
of maximum parsimony, with gaps treated separately as
either missing data or as a fifth state. Support for nodes was
estimated by bootstrapping with 10 000 replicates.
Both analyses gave widely congruent estimates of phylo-
geny (Fig. 6). The three heterocystous strains form a clade
with 100% support, separated from the nonheterocystous
strains by between 16% and 21% divergence. Within the
nonheterocystous strains, two pairs of taxa – Cyanothece
with Crocosphaera and the two Lyngbya species are well
supported. Other relationships are poorly supported.
Although the analysis with gaps treated as missing data
suggests that the filamentous taxa are not a clade, analysis
with gaps treated as a fifth character supported a relation-
ship between T. erythraeum and Lyngbya spp., although with
weak support (51%). Thus, exact relationships within this
group cannot be ascertained by these sequences, although a
sister-taxa relationship between T. erythraeum and L. ma-
juscula is strongly supported through analysis of the Hyp
sequences. These results are not in conflict with those
suggested by Ludwig et al. (2006), in which the only well-
supported node within cyanobacteria is that of the three
heterocystous strains. Evidence for the position of the
ancestral root within the cyanobacteria is weak, although T.
erythraeum may be sister taxa to the remaining sampled
cyanobacteria (Ludwig et al., 2006).
Phylogenetic analysis of cyanobacterial hydrogenases ac-
cessory proteins (Hyp A,B,C,D,E and F) and the bidirec-
tional hydrogenases structural proteins (Hox) is
complicated by the higher level of variation between species,
and in particular greater length variation that leads to
uncertain alignment for many positions. Further, not all of
the amino acid sequences of hydrogenase accessory proteins
are available for all the species analyzed for the uptake
hydrogenase structural genes. However, unweighted parsi-
mony analyses indicate that supported estimates of relation-
ships recovered for these proteins do not conflict with the
estimate of phylogeny shown in Fig. 6 (analyses not shown).
Genetic engineering/cyanobacterial H2
production
Cyanobacteria can be used for the production of molecular
hydrogen (H2), a possible future energy carrier, which has
been the subject of several recent reviews (Levin et al., 2004;
Dutta et al., 2005; Kruse et al., 2005; Prince & Kheshgi, 2005;
Sakurai & Masukawa, 2007). As the main advantages,
cyanobacteria can use sunlight as an energy source, water
as an electron source and air as a carbon (CO2) and a
nitrogen (N2) source. Therefore, no complicated or expen-
sive media are needed for the cultivation of cyanobacteria,
and the overall theoretical energy conversion efficiency
(from solar energy sun to H2) may be the highest possible.
In cyanobacteria, two natural pathways for H2 produc-
tion can be used.
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
709Cyanobacterial hydrogenases

H2 production as a by-product during nitrogenfixation by nitrogenases
In N2-fixing strains, H2 is produced as a by-product by the
nitrogenase enzymatic complex. As this reaction needs the
input of ATP (at least two ATP per electron), the overall
energy efficiency for hydrogen production is rather low. The
turnover of the nitrogenase enzyme is not very high
(o 10 s�1), and the H2 produced is efficiently taken up
by an uptake hydrogenase. The overall oxygen sensitive
N2-fixation process is occurring in an anaerobic environ-
ment achieved using a number of different strategies includ-
ing spatial or/and temporal separation of N2 fixation and
oxygenic photosynthesis and increased respiration.
Cyanobacterial nitrogenases contain molybdenum (Mo),
vanadium (V) or iron (Fe) in the active site, with different
genes and gene products making up the different nitro-
genases (Eady, 1996; Zhao et al., 2006). With sufficient
amounts of molybdenum available, the active site harbors
molybdenum and iron. Under molybdenum-deprived con-
ditions, the conventional molybdenum-nitrogenase is re-
placed by an alternative vanadium-nitrogenase, and if
vanadium is also limited, some N2-fixing microorganisms
are able to synthesize an alternative iron-nitrogenase.
Depending on the type of nitrogenase (molybdenum,
vanadium or iron), different amounts of electrons are
allocated for N2 fixation or H2 production. The general
equation for the nitrogenase-catalyzed reaction is as follows
(Rees et al., 2005):
N2 þ ð2nþ 6Þe� þ ð2nþ 6ÞHþ þ pð2nþ 6ÞATP! 2NH3 þ nH2 þ pð2nþ 6ÞADPþ pð2nþ 6ÞPi
It has been reported that n is 1 for the molybdenum-
containing enzymes, 3 for the vanadium-nitrogenases and
7.5 for the iron-only nitrogenases, respectively. As a conse-
quence, the alternative nitrogenases, although still very little
is known in cyanobacteria, may be better H2 producers
compared with the more conventional molybdenum-nitro-
genases.
H2 production by the bidirectional hydrogenase
The cyanobacterial bidirectional hydrogenase may, under
anaerobic conditions, produce and evolve significant
Fig. 6. Unrooted single most parsimonious tree recovered from an MP analysis with gaps treated as missing data of combined small (HupS) and large
(HupL) subunit amino acid sequences of cyanobacterial uptake hydrogenases. 210 characters were parsimony informative, and a single tree of 552
steps was recovered (CI = 0.83, RI = 0.76). NJ recovered an identical topology. Treating gaps as a fifth state altered the topology as indicated in the text.
Values beside nodes indicate bootstrap support for MP/NJ. 100� indicates 100% support in both analyses.
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
710 P. Tamagnini et al.

amounts of H2. Because this reaction is not dependent on
ATP, it is energetically more efficient and favorable for H2
production, with a much higher turnover (1 million turn-
overs per second) compared with the nitrogenase-based H2
production. At the same time, the enzyme is not specifically
located in an oxygen-protected environment, and the reac-
tion turns into the opposite direction (H2 uptake) above a
certain H2 partial pressure. Therefore, a continuous and very
effective removal of both O2 and H2 from the cells and the
culture is necessary to lower the overall energy conversion
efficiency significantly. Furthermore, an accumulation of
ATP could inhibit the electron flow, because it is produced
during the linear or cyclic electron flow around PSI, but is
not used by the electron acceptor hydrogenase.
Besides the specific challenges for H2 production con-
nected to the H2-evolving enzymes, there are additional
unsolved issues for photoautotrophical H2 production in
general. These are related to the low quantum efficiency, due
to the naturally large antenna systems of the photosystems,
and to electron consuming pathways directly competing
with e.g. nitrogenases and hydrogenases.
In summary, to achieve a sustainable, renewable cyano-
bacterial-based H2 production, the following challenges
have to be addressed:
(1) efficient H2 uptake by the cells,
(2) low energy efficiency and turnover of the nitrogenase
and/or the hydrogenase,
(3) limiting amounts of active H2-evolving enzymes,
(4) high oxygen sensitivity of the nitrogenase and/or the
hydrogenase,
(5) electron flow inhibition by accumulation of ATP in a
hydrogenase-driven system,
(6) low quantum efficiency due to too large antennas in
both Photosystem II (PSII) and PSI and
(7) electron-consuming pathways competing with an effi-
cient electron transfer to the H2 enzymes.
In recent years, there have been attempts to overcome
these barriers and problems, mainly by targeted genetic
engineering of cyanobacterial strains:
(1) Efficient H2 uptake by the cells: Cyanobacteria have
evolved an effective mechanism to recycle the H2 evolved
during nitrogen fixation: an uptake hydrogenase that oxi-
dizes the H2 evolved, and transfers electrons to e.g. the
respiratory-chain. As this reaction significantly lowers the
H2 production efficiency of a nitrogenase-based system,
targeted mutants with reduced or deficient uptake hydro-
genase activity have been produced. This was first achieved
by chemical mutagenesis (Kumar & Kumar, 1991; Mikheeva
et al., 1995), and later, since the molecular biology tools for
genetic engineering were established, by targeted knock-out
of structural or accessory genes of the uptake hydrogenase.
Uptake hydrogenase-deficient mutants of A. variabilis
(Happe et al., 2000), N. punctiforme (Lindberg et al., 2002,
2004), Nostoc sp. PCC 7120 (Lindblad et al., 2002; Masuka-
wa et al., 2002; Carrasco et al., 2005) and Nostoc sp. PCC
7422 (Yoshino et al., 2007) have been shown to be signifi-
cantly better H2 producers compared with the respective wild
types. In general, the H2 produced by a nitrogenase in the
wild type will be quickly reoxidized by the uptake hydro-
genase, whereas in an uptake hydrogenase-deficient mutant
the H2 produced will leave the cells. One should bear in mind
that all these strains, with the exception of N. punctiforme,
also possess a bidirectional hydrogenase. However, only for
Nostoc sp. PCC 7120 (Masukawa et al., 2002) the effect of a
hox-defective mutant (DhoxH) has been investigated. A
Nostoc sp. PCC 7120 mutant deficient in both hydrogenases
(DhupL/DhoxH) showed the same increase in H2 evolution as
the uptake hydrogenase-deficient mutant (DhupL), whereas
the bidirectional hydrogenase-deficient mutant (DhoxH) pro-
duced less H2 compared with the wild type.
In gas exchange experiments with an uptake hydrogenase-
deficient mutant of Nostoc punctiforme (Lindberg et al.,
2004), the amount of H2 produced per molecule of N2 fixed
varied with the light conditions. The ratio of H2 produced/
N2 fixed under low light and high light was 1.4 and 6.1,
respectively. This showed that, under the specific conditions,
the energy flow through the nitrogenase may be directed
towards the H2 production rather than the N2 fixation.
(2) Low energy efficiency and turnover of the nitrogenase
and/or the hydrogenase: H2-evolving enzymes with the high-
est reported turnover are the Fe-hydrogenases (Houchins,
1984; Adams, 1990). These enzymes are irreversibly inacti-
vated by oxygen, and are present in e.g. fermentative bacteria
(e.g. Clostridium) and green algae (e.g. Chlamydomonas) but
not in cyanobacteria. An elegant strategy for the creation of
an efficient H2 producer, which will not be inhibited by the
surrounding oxygen, would be the expression of a highly
active Fe-hydrogenase in the heterocysts of filamentous
cyanobacteria unable to reoxidase any H2 (i.e. an uptake
hydrogenase-deficient strain). The heterologous expression
of different iron-hydrogenases in various organisms such as
Synechococcus (Asada et al., 2000), E. coli (Posewitz et al.,
2004; King et al., 2006) and Clostridium (Girbal et al., 2005)
has already been achieved. Recently, the accessory genes
necessary for the maturation of iron-hydrogenases into
active enzymes were identified (Posewitz et al., 2004; Bock
et al., 2006; King et al., 2006). Therefore, the heterologous
expression of an active iron-hydrogenase in a cyanobacterial
host, e.g. in the heterocyst of a strain for which the genome
has been sequenced, is an interesting and realistic project.
Moreover, because the iron-hydrogenases are able to use a
wide variety of primary electron donors (Vignais et al.,
2001), including ferredoxin, which is the electron donor of
the cyanobacterial nitrogenase, it may be possible to link the
introduced iron-hydrogenase to an existing electron transfer
pathway within the cyanobacterial cell.
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
711Cyanobacterial hydrogenases

(3) Limiting amounts of active H2-evolving enzymes:
Because the H2-evolving enzymes (nitrogenase(s) and/or
bidirectional hydrogenases) are strictly regulated on several
different levels (transcription, translation and maturation),
one possible way to enhance the production of H2 may be
the overexpression of these enzymes. For this purpose, the
genes encoding the selected enzyme/protein to be over-
expressed are placed under the control of an artificial
promoter and ribosomal-binding site (RBS) combination
on an expression vector or placed directly in the genome.
The choice of a constitutive or an inducible promoter,
together with a strong RBS, takes the enzyme biosynthesis
out of the control of the organism’s natural regulation
system, allowing a significant increase in the amount of
enzyme produced.
In heterocystous cyanobacteria grown under N2-fixing
conditions, c. 5–10% of the vegetative cells differentiate into
heterocysts. These specialized cells are compartments with
reduced O2 pressure, and thus suitable for H2 production,
either via nitrogenase or via an introduced hydrogenase.
Therefore, the increase of the heterocyst frequency should
result in a higher overall H2 production capacity by the
organism. Interestingly, it has been shown that the hetero-
cyst frequency can be increased, e.g. by overexpression of
hetR or by inactivation of patS, or hetN (Buikema &
Haselkorn, 2001; Golden & Yoon, 2003; Borthakur et al.,
2005). However, this has not been coupled to H2 production
or increased H2 production.
(4) High oxygen sensitivity of the nitrogenase and/or the
hydrogenase: One main obstacle in H2 production using
photosynthetic microorganisms is the high sensitivity of the
H2-evolving enzymes, and some attempts have been made to
introduce less oxygen sensitive hydrogenases into cyanobac-
teria. At the Craig Venter Institute (US), work is being
carried out aiming at transferring the O2-tolerant NiFe-
hydrogenase of the purple-sulfur photosynthetic bacterium
T. roseopersicina (Kovacs et al., 2005) into a Synechococcus
strain. In addition, several other putative O2-tolerant NiFe-
hydrogenases have been identified from the marine environ-
ment that could be alternative candidates to be introduced
into a cyanobacterial background (Xu et al., 2005). Also in
the US, the genes encoding the more O2-tolerant NiFe
hydrogenase of the purple nonsulfur photosynthetic bacter-
ium Rubrivivax gelatinosus CBS (Ghirardi et al., 2005), and
its accessory proteins, are being introduced into Synechocys-
tis. The in vivo half-life of this hydrogenase is 21 h in air and
6 h in air when the protein is partially purified (Ghirardi
et al., 2005). However, to the authors’ knowledge, the
heterologous expression of a more oxygen-tolerant hydro-
genase in any cyanobacterium remains to be shown.
(5) Electron flow inhibition by accumulation of ATP in a
hydrogenase-driven system: In an optimal H2 production
system, all electrons derived from water splitting in PSII
should be directed to the H2-evolving enzyme (nitrogenase
or hydrogenase) to reach maximal energy conversion effi-
ciency. In addition to the electron flow in the photosynthetic
electron transport chain, a transmembrane potential is built
up that is used for generating ATP through an ATP synthase.
In a nitrogenase-based system, the ATP is clearly needed for
N2 fixation. However, in the hydrogenase-catalyzed reaction
no ATP is consumed and the electron flow could, in a
photosynthetic microorganism, be inhibited by the accu-
mulated transmembrane potential across the thylakoid
membrane (Lee & Greenbaum, 2003). It has been observed
in the green algae Chlamydomonas reinhardtii that oxygen
serves as an electron sink, competing for electrons with the
H2-producing pathway, resulting in a ‘new oxygen sensitiv-
ity’ (Lee & Greenbaum, 2003). A possible solution may be
the introduction of a synthetic, polypeptide based on the
proton channel into the thylakoid membranes, with its
transcription controlled by a hydrogenase promoter (Lee &
Greenbaum, 2003). The proton channel may be expressed
under H2-producing (anaerobic) conditions, to dissipate the
proton gradient across the thylakoids membrane. A similar
strategy may work for cyanobacteria.
(6) Low quantum efficiency due to too large antennas in
both PSII and PSI: In nature, phototrophic organisms have
developed to handle and compete during very low light
intensities, and, therefore, large antenna systems have been
evolved. However, for biotechnological applications in
photobioreactors, where an optimal supply with light can
be engineered, only a small part of the light will be used by
the microorganisms. In addition, self-shading may become
a severe limitation causing reduced efficiency in utilizing
incoming solar energy. To overcome this problem, a reduc-
tion of the antenna size has been proposed (Melis et al.,
1998; Nakajima & Ueda, 1999; Lee et al., 2002). The
phycocyanin-deficient mutant PD1 of Synechocystis PCC
6714, generated by chemical mutagenesis, showed up to
50% higher maximal photosynthesis activity under high
light conditions compared with the wild type (Nakajima &
Ueda, 1997, 1999). Antenna-deficient green algal mutants,
created by chemical mutagenesis (Lee et al., 2002) or genetic
engineering (Polle et al., 2003; Tetali et al., 2007), also
showed remarkably greater solar conversion efficiencies and
a higher photosynthetic productivity than the respective
wild type under mass culture conditions.
(7) Electron-consuming pathways competing with an effi-
cient electron transfer to the H2 enzymes: For the production
of H2 through the action of an active nitrogenase or
hydrogenase electrons are required to combine with protons
to form H2. Because protons are abundant within the cell,
the main limitation is the available number of electrons. The
primary electron donors for the H2-producing enzymes
are ferredoxin (nitrogenase, Fe-hydrogenases) and NADH/
NADPH (NiFe-hydrogenases). However, the electrons are
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
712 P. Tamagnini et al.

mainly used by other pathways, the so-called ‘competing
pathways’, e.g. respiration and the Calvin cycle. Therefore,
one strategy for an enhanced H2 production is to direct the
electron flow towards the H2-producing enzymes and away
from any other competing pathway.
Experiments with the ndhB mutant M55 of Synechocystis
PCC 6803, which is defective in the type I NADPH-
dehydrogenase complex (NDH-1) (Cournac et al., 2004),
showed that this mutant produces only low amounts of O2
in the light, has a poor capacity to fix CO2 and evolves H2
for several minutes during dark-to-light transitions, while
the H2 uptake was negligible. The electrons used to produce
H2 were mainly coming from water splitting in PSII and
from the carbohydrate-mediated reduction of the PQ pool.
In another study, the level of reduced NADP shifted from
50% in the wild type to 100% in the NDH-1 mutant (Cooley
& Vermaas, 2001). As the cyanobacterial bidirectional hy-
drogenase evolves H2 at a relatively high level of reduced
NAD(P), the construction of mutants with blocked electron
transfer in selected key pathways, increasing the relative level
of reduced NAD(P), may be a promising strategy to increase
the H2 production capacity.
Another strategy for directing electrons toward the hy-
drogenase is to directly link the hydrogenase to PSI.
Ihara et al. (2006b) fused the membrane-bound NiFe
hydrogenase from Ralstonia eutropha H16 to the peripheral
PSI subunit PsaE of the cyanobacterium Thermosynechococ-
cus elongatus (Hyd-PsaE), and used a PsaE-free PSI
(PSI�) extract from a PsaE-deficient mutant of Synechocystis
sp. PCC 6803. The resulting hydrogenase/PSI complex
showed light-driven hydrogen production in vitro, which
was five times higher compared with a control without a
direct coupling of the hydrogenase to PSI. However, as the
activity of the hydrogenase-PsaE fusion protein was only
16% of that of the wild-type hydrogenase protein and was
totally suppressed by adding ferredoxin (Fd) and ferredox-
in-NADP1-reductase (FNR), the authors concluded that the
linker between the hydrogenase and PsaE has to be opti-
mized. In another work by the same group (Ihara et al.,
2006a), PsaE from Synechocystis sp. PCC 6803 was
chemically cross-linked with cytochrome c3 (cytc3) from
Desulfovibrio vulgaris and stoichiometrically assembled with
PsaE-free PSI to form a cytc3/PSI complex. The NADPH
production by this complex coupled with Fd and FNR
decreased to c. 10% of the original activity, whereas the H2
production by the cytc3/PSI complex coupled with hydro-
genase from Desulfovibrio vulgaris was enhanced sevenfold.
This clearly demonstrated that it is possible, in vitro, to
direct the electron-flow toward the hydrogenase by changing
the environment of the electron-donating PSI. The
next challenge will be to develop an in vivo, or even
in situ, functional system based on the assembly of different
enzymes and proteins.
As a future perspective, the development of synthetic
biology reveals new possibilities for the direct construction
of efficient H2-evolving cyanobacterial strains. Both in the
US (e.g. the Craig Venter Institute) and in Europe (e.g. the
EU/NEST project ‘BioModularH2’), the first attempts have
been initiated to use this new concept aiming at designing
reusable, standardized molecular building blocks that will
produce a photosynthetic bacterium containing engineered
chemical pathways for competitive, clean and sustainable H2
production.
Concluding remarks
The fundamental aspects of cyanobacterial hydrogenases,
and their more applied potential use as future producers of
renewable H2 from sun and water, are receiving increased
international attention. At the same time, significant progress
is being made in the understanding of the molecular regula-
tion of the genes encoding both the enzymes as well as the
accessory proteins needed for the correct assembly of an
active hydrogenase. In the last few years, the transcription
factors directly involved in the regulation of cyanobacterial
hydrogenases have been identified. Moreover, the first steps to
use isolated components from cyanobacteria and other
microorganisms in order to create a functional H2-producing
unit are being taken. With the increasing scientific commu-
nity and public interest in clean and renewable energy
sources, and consequent funding opportunities, rapid pro-
gress will be made in the fundamental understanding of the
regulation and maturation of cyanobacterial hydrogenases at
both genetic and protein levels. Unique and unexpected
results in the transcriptional regulation of cyanobacterial
hydrogenases will emerge during the coming years. Moreover,
the more applied aspects will be highlighted with progress in
generating genetically modified strains with an increased
capacity for renewable H2 from sun and water. The possibi-
lities and challenges within synthetic biology, including the
use of isolated proteins and parts, will be explored, aiming at
creating both cyanobacteria with a high potential for H2
production as well as functional in vitro systems.
Acknowledgements
This work was financially supported by FCT (POCTI/BIO/
44592/2002; SFRH/BD/4912/2001, SFRH/BD/16954/2004),
ESF (III Quadro Comunitario de Apoio), the Swedish
Research Council, the Swedish Energy Agency, the Nordic
Energy Research Program (project BioHydrogen), the EU/
NEST Projects SOLAR-H (contract # 516510) and BioMo-
dularH2 (contract # 043340).
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
713Cyanobacterial hydrogenases

References
Adams MWW (1990) The structure and mechanism of iron-
hydrogenases. Biochim Biophys Acta 1020: 115–145.
Anderson DC, Campbell EL & Meeks JC (2006) A soluble 3D LC/
MS/MS proteome of the filamentous cyanobacterium Nostoc
punctiforme. J Prot Res 5: 3096–3104.
Antal TK & Lindblad P (2005) Production of H2 by sulphur-
deprived cells of the unicellular cyanobacteria Gloeocapsa
alpicola and Synechocystis sp. PCC 6803 during dark
incubation with methane or at various extracellular pH. J Appl
Microbiol 98: 114–120.
Antal TK, Oliveira P & Lindblad P (2006) The bidirectional
hydrogenase in the cyanobacterium Synechocystis sp. strain
PCC 6803. Int J Hydrogen Energy 31: 1439–1444.
Appel J & Schulz R (1996) Sequence analysis of an operon of
NAD(P)-reducing nickel hydrogenase from the
cyanobacterium Synechocystis sp. PCC 6803 gives additional
evidence for direct coupling of the enzyme to NADP(H)-
dehydrogenase (complex I). Biochim Biophys Acta 1298:
141–147.
Appel J, Phunpruch S, Steinmuller K & Schulz R (2000) The
bidirectional hydrogenase of Synechocystis sp. PCC 6803 works
as an electron valve during photosynthesis. Arch Microbiol 173:
333–338.
Asada Y, Kawamura S & Ho K-K (1987) Hydrogenase from the
unicellular cyanobacterium, Microcystis aeruginosa.
Phytochemistry 26: 637–640.
Asada Y, Koike Y, Schnackenberg J, Miyake M, Uemura I &
Miyake J (2000) Heterologous expression of clostridial
hydrogenase in the cyanobacterium Synechococcus PCC7942.
Biochim Biophys Acta 1490: 269–278.
Asayama M, Kabasawa M, Takahashi I, Aida T & Shirai M (1996)
Highly repetitive sequences and characteristics of genomic
DNA in unicellular cyanobacterial strains. FEMS Microbiol Lett
137: 175–181.
Axelsson R & Lindblad P (2002) Transcriptional regulation of
Nostoc hydrogenases: effects of oxygen, hydrogen, and nickel.
Appl Environ Microbiol 68: 444–447.
Axelsson R, Oxelfelt F & Lindblad P (1999) Transcriptional
regulation of Nostoc uptake hydrogenase. FEMS Microbiol Lett
170: 77–81.
Bergman B, Gallon JR, Rai AN & Stal LJ (1997) N2 fixation by
non-heterocystous cyanobacteria. FEMS Microbiol Rev 19:
139–185.
Berman-Frank I, Lundgren P & Falkowski P (2003) Nitrogen
fixation and photosynthetic oxygen evolution in
cyanobacteria. Res Microbiol 154: 157–64.
Blokesch M & Bock A (2002) Maturation of [NiFe]-hydrogenases
in Escherichia coli: the HypC cycle. J Mol Biol 324: 287–96.
Blokesch M & Bock A (2006) Properties of the [NiFe]-
hydrogenase maturation protein HypD. FEBS Lett 580:
4065–4068.
Blokesch M, Paschos A, Theodoratou E, Bauer A, Hube M, Huth
S & Bock A (2002) Metal insertion into NiFe-hydrogenases.
Biochem Soc Trans 30: 674–680.
Blokesch M, Albracht SP, Matzanke BF, Drapal NM, Jacobi A &
Bock A (2004a) The complex between hydrogenase-
maturation proteins HypC and HypD is an intermediate in the
supply of cyanide to the active site iron of [NiFe]-
hydrogenases. J Mol Biol 344: 155–167.
Blokesch M, Rohrmoser M, Rode S & Bock A (2004b) HybF, a
zinc-containing protein involved in NiFe hydrogenase
maturation. J Bacteriol 186: 2603–2611.
Bock A, King PW, Blokesch M & Posewitz MC (2006) Maturation
of hydrogenases. Adv Microb Physiol 51: 1–71.
Bohme H (1998) Regulation of nitrogen fixation in heterocyst-
forming cyanobacteria. Trends Plant Sci 3: 346–351.
Boison G, Schmitz O, Mikheeva L, Shestakov S & Bothe H (1996)
Cloning, molecular analysis and insertional mutagenesis of the
bidirectional hydrogenase genes from the cyanobacterium
Anacystis nidulans. FEBS Lett 394: 153–158.
Boison G, Schmitz O, Schmitz B & Bothe H (1998) Unusual gene
arrangement of the bidirectional hydrogenase and functional
analysis of its diaphorase subunit HoxU in respiration of the
unicellular cyanobacterium Anacystis nidulans. Curr Microbiol
36: 253–258.
Boison G, Bothe H, Hansel A & Lindblad P (1999) Evidence
against a common use of the diaphorase subunits by the
bidirectional hydrogenase and by the respiratory complex I in
cyanobacteria. FEMS Microbiol Lett 174: 159–165.
Boison G, Bothe H & Schmitz O (2000) Transcriptional analysis
of hydrogenase genes in the cyanobacteria Anacystis nidulans
and Anabaena variabilis monitored by RT-PCR. Curr Microbiol
40: 315–321.
Borthakur PB, Orozco CC, Young-Robbins SS, Haselkorn R &
Callahan SM (2005) Inactivation of patS and hetN causes
lethal levels of heterocyst differentiation in the filamentous
cyanobacterium Anabaena sp. PCC 7120. Mol Microbiol 57:
111–123.
Bothe H, Tennigkeit J & Eisbrenner G (1977) The utilization of
molecular hydrogen by the blue-green alga Anabaena
cylindrica. Arch Microbiol 114: 43–49.
Bothe H, Kentemich T & Dai H (1991) Recent aspects on the
hydrogenase–nitrogenase relationship in cyanobacteria. Dev
Plant Soil Sci 48: 367–375.
Buikema WJ & Haselkorn R (2001) Expression of the Anabaena
hetR gene from a copper-regulated promoter leads to
heterocyst differentiation under repressing conditions. PNAS
98: 2729–2734.
Burgdorf T, van der Linden E, Bernhard M, Yin QY, Back JW,
Hartog AF, Muijsers AO, de Koster CG, Albracht SP &
Friedrich B (2005) The soluble NAD1-reducing [NiFe]-
hydrogenase from Ralstonia eutropha H16 consists of six
subunits and can be specifically activated by NADPH. J
Bacteriol 187: 3122–3132.
Busby S & Ebright RH (1999) Transcription activation by
catabolite activator protein (CAP). J Mol Biol 293: 199–213.
Carrasco CD, Buettner JA & Golden JW (1995) Programed DNA
rearrangment of a cyanobacterial hupL gene in heterocysts.
Proc Natl Acad Sci USA 92: 791–795.
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
714 P. Tamagnini et al.

Carrasco CD, Garcia JS & Golden JW (1998) Programmed DNA
rearrangement of a hydrogenase gene during Anabaena
heterocyst development. BioHydrogen (Zaborsky OR,
Benemann JR, Matsunaga T, Miyake J & San Pietro A, eds), pp.
203–207. Plenum Press, New York.
Carrasco CD, Holliday SD, Hansel A, Lindblad P & Golden JW
(2005) Heterocyst-specific excision of the Anabaena sp. strain
PCC 7120 hupL element requires xisC. J Bacteriol 187:
6031–6038.
Casalot L & Rousset M (2001) Maturation of the [NiFe]
hydrogenases. Trends Microbiol 9: 228–237.
Chen PC, Almon H & Boger P (1989) Physiological factors
determining hydrogenase activity in nitrogen-fixing
heterocystous cyanobacteria. Plant Physiol 89: 1035–1038.
Cooley JW & Vermaas WFJ (2001) Succinate dehydrogenase and
other respiratory pathways in thylakoid membranes of
Synechocystis sp. strain PCC 6803: capacity comparisons and
physiological function. J Bacteriol 183: 4251–4258.
Cournac L, Guedeney G, Peltier G & Vignais PM (2004)
Sustained photoevolution of molecular hydrogen in a mutant
of Synechocystis sp. strain PCC 6803 deficient in the type I
NADPH-dehydrogenase complex. J Bacteriol 186: 1737–1746.
Craig NL & Nash AH (1984) E. coli integration host factor binds
to specific sites in DNA. Cell 39: 707–716.
Daday A, Mackerras AH & Smith GD (1985) The effect of nickel
on hydrogen metabolism and nitrogen fixation in the
cyanobacterium Anabaena cylindrica. J Gen Microbiol 131:
231–238.
Dutta D, De D, Chaudhuri S & Bhattacharya SK (2005) Hydrogen
production by cyanobacteria. Microb Cell Fact 4: 36.
Eady RR (1996) Structure–function relationships of alternative
nitrogenases. Chem Rev 96: 3013–3030.
Ferreira D, Leitao E, Sjoholm J, Oliveira P, Lindblad P, Moradas-
Ferreira P & Tamagnini P (2007) Transcription and regulation
of the hydrogenase(s) accessory genes, hypFCDEAB, in the
cyanobacterium Lyngbya majuscula CCAP 1446/4. Arch
Microbiol (Epub ahead of print, PMID: 17639348).
Friedman DI (1988) Integration host factor: a protein for all
reasons. Cell 55: 545–554.
Fritsche E, Paschos A, Beisel HG, Bock A & Huber R (1999)
Crystal structure of the hydrogenase maturating
endopeptidase HYBD from Escherichia coli. J Mol Biol 288:
989–998.
Fulda S, Mikkat S, Huang F, Huckauf J, Marin K, Norling B &
Hagemann M (2006) Proteome analysis of salt stress response
in the cyanobacterium Synechocystis sp. strain PCC 6803.
Proteomics 6: 2733–2745.
Gan CS, Reardon KF & Wright PC (2005) Comparison of protein
and peptide prefractionation methods for the shotgun
proteomic analysis of Synechocystis sp. PCC 6803. Proteomics 5:
2468–2478.
Ghirardi ML, King PW, Posewitz MC, Maness PC, Fedorov A,
Kim K, Cohen J, Schulten K & Seibert M (2005) Approaches to
developing biological H2-photoproducing organisms and
processes. Biochem Soc Trans 33: 70–72.
Girbal L, von Abendroth G, Winkler M, Benton PM, Meynial-
Salles I, Croux C, Peters JW, Happe T & Soucaille P (2005)
Homologous and heterologous overexpression in Clostridium
acetobutylicum and characterization of purified clostridial and
algal Fe-only hydrogenases with high specific activities. Appl
Environ Microb 71: 2777–2781.
Golden JW (1997) Programmed DNA Rearrangements in
Cyanobacteria. Bacterial Genomes: Physical Structure
and Analysis (de Bruijn FR, Lupski JR & Weinstok GM),
pp. 162–163. Chapman & Hall, New York.
Golden JW & Yoon H-S (2003) Heterocyst development in
Anabaena. Curr Opin Microbiol 6: 557–563.
Goodman SD, Velten NG, Gao Q, Robinson S & Segall AM (1999)
In vitro selection of integration host factor binding sites. J
Bacteriol 181: 3246–3255.
Goodrich JA, Schwartz ML & McClure WR (1990) Searching for
and predicting the activity of sites for DNA binding proteins:
compilation and analysis of the binding sites for Escherichia
coli integration host factor (IHF). Nucleic Acids Res 18:
4993–5000.
Gubili J & Borthakur D (1996) The use of a PCR cloning and
screening strategy to identify lambda clones containing the
hupB gene of Anabaena sp. strain PCC 7120. J Microbiol
Methods 27: 175–182.
Gubili J & Borthakur D (1998) Organization of the hupDEAB
genes within the hydrogenase gene cluster of Anabaena sp.
strain PCC 7120. J Appl Phycol 10: 163–167.
Gutekunst K, Phunpruch S, Schwarz C, Schuchardt S, Schulz-
Friedrich R & Appel J (2005) LexA regulates the bidirectional
hydrogenase in the cyanobacterium Synechocystis sp. PCC
6803 as a transcription activator. Mol Microbiol 58: 810–823.
Gutekunst K, Hoffmann D, Lommer M, Egert M, Suzuki I,
Schulz-Friedrich R & Appel J (2006) Metal dependence and
intracellular regulation of the bidirectional NiFe-hydrogenase
in Synechocystis sp. PCC 6803. Int J Hydrogen Energ 31:
1452–1459.
Hallenbeck PC & Benemann JR (1978) Characterization and
partial purification of the reversible hydrogenase of Anabaena
cylindrica. FEBS Lett 94: 261–264.
Hansel A, Axelsson R, Lindberg P, Troshina OY, Wunschiers R &
Lindblad P (2001) Cloning and characterisation of a hyp gene
cluster in the filamentous cyanobacterium Nostoc sp. strain
PCC 73102. FEMS Microbiol Let 201: 59–64.
Happe T, Schutz K & Bohme H (2000) Transcriptional and
mutational analysis of the uptake hydrogenase of the
filamentous cyanobacterium Anabaena variabilis ATCC 29413.
J Bacteriol 182: 1624–1631.
Herrero A, Muro-Pastor AM & Flores E (2001) Nitrogen control
in cyanobacteria. J Bacteriol 183: 411–425.
Herrero A, Muro-Pastor AM, Valladares A & Flores E (2004)
Cellular differentiation and the NtcA transcription factor in
filamentous cyanobacteria. FEMS Microbiol Rev 28: 469–487.
Hihara Y, Kamei A, Kanehisa M, Kaplan A & Ikeuchi M (2001)
DNA microarray analysis of cyanobacterial gene expression
during acclimation to high light. Plant Cell 13: 793–806.
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
715Cyanobacterial hydrogenases

Hoffmann D, Gutekunst K, Klissenbauer M, Schluz-Friedrich R
& Appel J (2006) Mutagenesis of hydrogenase accessory genes
of Synechocystis sp. PCC 6803. Additional homologues of hypA
and hypB are not active in hydrogenase maturation. FEBS J
273: 4516–4527.
Houchins JP (1984) The physiology and biochemistry of
hydrogen metabolism in cyanobacteria. Biochim Biophys Acta
768: 227–255.
Houchins JP & Burris RH (1981a) Comparative characterization
of two distinct hydrogenases from Anabaena sp. strain 7120.
J Bacteriol 146: 215–221.
Houchins JP & Burris RH (1981b) Occurrence and localization of
two distinct hydrogenases in the heterocystous
cyanobacterium Anabaena sp. strain 7120. J Bacteriol 146:
209–214.
Howarth DC & Codd GA (1985) The uptake and production of
molecular hydrogen by unicellular cyanobacteria. J Gen
Microbiol 131: 1561–1569.
Howitt CA & Vermaas W (1999) Subunits of the NAD(P)-
reducing nickel-containing hydrogenase do not act as part of
the type-1 NAD(P)H-dehydrogenase in the cyanobacterium
Synechocystis sp. PCC 6803. The Phototrophic Prokaryotes
(Peschek GA, Loffelhardt W & Schmetterer G, eds), pp.
595–601. Kluwer Academic/Plenum Publishers, New York.
Ihara M, Nakamoto H, Kamachi T, Okura I & Maeda M (2006a)
Photoinduced hydrogen production by direct electron transfer
from photosystem I cross-linked with cytochrome c3 to
[NiFe]-hydrogenase. Photochem Photobiol 82: 1677–1685.
Ihara M, Nishihara H, Yoon K-S, Lenz O, Friedrich B, Nakamoto
H, Kojima K, Honma D, Kamachi T & Okura I (2006b)
Light-driven hydrogen production by a hybrid complex of a
[NiFe]-hydrogenase and the cyanobacterial photosystem I.
Photochem Photobiol 82: 676–682.
Ikeuchi M & Tabata S (2001) Synechocystis sp. PCC 6803 – a
useful tool in the study of the genetics of cyanobacteria.
Photosynth Res 70: 73–83.
Jacobi A, Rossmann R & Bock A (1992) The hyp operon gene
products are required for the maturation of catalytically active
hydrogenase isoenzymes in Escherichia coli. Arch Microbiol
158: 444–451.
Kamei A, Hihara Y, Yoshihara S, Geng X, Kanehisa M & Ikeuchi
M (2001) Functional Analysis of lexA-like gene, sll1626 in
Synechocystis sp. PCC 6803 using DNA microarray. Plant Cell
Physiol 32suppl: S95.
Kaneko T, Tanaka A, Sato S, Kotani H, Sazuka T, Miyajima N,
Sugiura M & Tabata S (1995) Sequence analysis of the genome
of the unicellular cyanobacterium Synechocystis sp. strain PCC
6803. I. Sequence features in the 1 Mb region from map
positions 64% to 92% of the genome. DNA Res 2: 153–166.
Kaneko T, Sato S, Kotani H et al. (1996) Sequence analysis of the
genome of the unicellular cyanobacterium Synechocystis sp.
strain PCC6803. II. Sequence determination of the entire
genome and assignment of potential protein-coding regions.
DNA Res 3: 185–209.
Kaneko T, Nakamura Y, Wolk CP et al. (2001) Complete genomic
sequence of the filamentous nitrogen-fixing cyanobacterium
Anabaena sp. strain PCC 7120. DNA Res 8: 205–213.
Kentemich T, Bahnweg M, Mayer F & Bothe H (1989)
Localization of the reversible hydrogenase in cyanobacteria. Z
Naturforsch 44c: 384–391.
Kentemich T, Casper M & Bothe H (1991) The reversible
hydrogenase in Anacystis nidulans is a component of the
cytoplasmic membrane. Naturwissenschaften 78: 559–560.
King PW, Posewitz MC, Ghirardi ML & Seibert M (2006)
Functional studies of [FeFe] hydrogenase maturation in an
Escherichia coli biosynthetic system. J Bacteriol 188:
2163–2172.
Kovacs KL, Kovacs AT, Maroti G, Meszaros LS, Balogh J,
Latinovics D, Fulop A, David R, Doroghazi E & Rakhely G
(2005) The hydrogenases of Thiocapsa roseopersicina. Biochem
Soc Trans 33: 61–63.
Kruse O, Rupprecht J, Mussgnug JH, Dismukes GC & Hankamer
B (2005) Photosynthesis: a blueprint for solar energy capture
and biohydrogen production technologies. Photochem
Photobiol Sci 4: 957–970.
Kuchar J & Hausinger RP (2004) Biosynthesis of metal sites.
Chem Rev 104: 509–525.
Kucho K-i, Okamoto K, Tsuchiya Y, Nomura S, Nango M,
Kanehisa M & Ishiura M (2005) Global analysis of circadian
expression in the cyanobacterium Synechocystis sp. strain PCC
6803. J Bacteriol 187: 2190–2199.
Kumar D & Kumar HD (1991) Effect of monochromatic lights on
nitrogen-fixation and hydrogen evolution in the isolated
heterocysts of Anabaena sp. strain CA. Int J Hydrogen Energy
16: 397–401.
Kumar S & Polasa H (1991) Influence of nickel and copper on
photobiological hydrogen production and uptake in
Oscillatoria subbrevis strain 111. Proc Indian Nat Sci Acad B57:
281–285.
Kumar AP, Perraju BTVV & Singh HN (1986) Carbon nutrition
and the regulation of uptake hydrogenase activity in free-living
and symbiotic Anabaena cycadeae. New Phytol 104: 115–120.
Kurian D, Jansen T & Maenpaa P (2006) Proteomic analysis of
heterotrophy in Synechocystis sp. PCC 6803. Proteomics 6:
1483–1494.
Lambert GR & Smith GD (1981) The hydrogen metabolism of
cyanobacteria (blue-green algae). Biol Rev 56: 589–660.
Lee JW & Greenbaum E (2003) A new oxygen sensitivity and its
potential application in photosynthetic H2 production. Appl
Biochem Biotechnol 105: 303–313.
Lee JW, Mets L & Greenbaum E (2002) Improvement of
photosynthetic CO2 fixation at high light intensity through
reduction of chlorophyll antenna size. Appl Biochem Biotechnol
98: 37–48.
Leitao E, Oxelfelt F, Oliveira P, Moradas-Ferreira P & Tamagnini
P (2005) Analysis of the hupSL operon of the
nonheterocystous cyanobacterium Lyngbya majuscula CCAP
1446/4: regulation of transcription and expression under a
light-dark regime. Appl Environ Microbiol 71: 4567–4576.
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
716 P. Tamagnini et al.

Leitao E, Pereira S, Bondoso J, Ferreira D, Pinto F, Moradas-
Ferreira P & Tamagnini P (2006) Genes involved in the
maturation of hydrogenase(s) in the nonheterocystous
cyanobacterium Lyngbya majuscula CCAP 1446/4. Int J
Hydrogen Energy 31: 1469–1477.
Levin DB, Lawrence P & Murry L (2004) Biohydrogen
production: prospects and limitations to practical application.
Int J Hydrogen Energy 29: 173–185.
Li H, Singh AK, McIntyre LM & Sherman LA (2004) Differential
gene expression in response to hydrogen peroxide and the
putative PerR regulon of Synechocystis sp. strain PCC 6803.
J Bacteriol 186: 3331–3345.
Lindberg P (2003) Cyanobacterial hydrogen metabolism – uptake
hydrogenase and hydrogen production by nitrogenase in
filamentous cyanobacteria. PhD Thesis, Faculty of Science
and Technology, Uppsala University, Sweden (ISBN
91-554-5708-8).
Lindberg P, Hansel A & Lindblad P (2000) hupS and hupL
constitute a transcription unit in the cyanobacterium Nostoc
sp. PCC 73102. Arch Microbiol 174: 129–133.
Lindberg P, Schutz K, Happe T & Lindblad P (2002) A hydrogen-
producing, hydrogenase-free mutant strain of Nostoc
punctiforme ATCC 29133. Int J Hydrogen Energy 27:
1291–1296.
Lindberg P, Lindblad F & Cournac L (2004) Gas exchange in the
filamentous cyanobacterium Nostoc punctiforme strain ATCC
29133 and its hydrogenase-deficient mutant strain NHM5.
Appl Environ Microbiol 70: 2137–2145.
Lindblad P & Sellstedt A (1990) Occurrence and localization of an
uptake hydrogenase in the filamentous heterocystous
cyanobacterium Nostoc PCC 73102. Protoplasma 159: 9–15.
Lindblad P, Christensson K, Lindberg P, Fedorov A, Pinto F &
Tsygankov A (2002) Photoproduction of H2 by wildtype
Anabaena PCC 7120 and a hydrogen uptake deficient mutant:
from laboratory experiments to outdoor culture. Int J
Hydrogen Energy 27: 1271–1281.
Llama MJ, Serra JL, Rao KK & Hall DO (1979) Isolation and
characterization of the hydrogenase activity from the non-
heterocystous cyanobacterium Spirulina maxima. FEBS Lett
98: 342–346.
Long M, Liu J, Chen Z, Bleijlevens B, Roseboom W & Albracht SP
(2007) Characterization of a HoxEFUYH type of [NiFe]
hydrogenase from Allochromatium vinosum and some EPR
and IR properties of the hydrogenase module. J Biol Inorg
Chem 12: 62–78.
Ludwig M, Schulz-Friedrich R & Appel J (2006) Occurrence of
hydrogenases in cyanobacteria and anoxygenic photosynthetic
bacteria: implications for the phylogenetic origin of
cyanobacterial and algal hydrogenases. J Mol Evol 63: 758–768.
Lundgren P, Bauer K, Lugomela C, Soderback E & Bergman B
(2003) Reevaluation of the nitrogen fixation behaviour in the
marine non-heterocystous cyanobacterium Lyngbya
majuscula. J Phycol 39: 310–314.
Lutz S, Jacobi A, Schlensog V, Bohm R, Sawers G & Bock A (1991)
Molecular characterization of an operon (hyp) necessary for
the activity of the three hydrogenase isoenzymes in Escherichia
coli. Mol Microbiol 5: 123–135.
Magalon A & Bock A (2000a) Analysis of the HypC–HycE
complex, a key intermediate in the assembly of the metal
center of the Escherichia coli hydrogenase 3. J Biol Chem 275:
21114–21120.
Magalon A & Bock A (2000b) Dissection of the maturation
reactions of the [NiFe] hydrogenase 3 from Escherichia coli
taking place after nickel incorporation. FEBS Lett 473:
254–258.
Maier T & Bock A (1996) Generation of active [NiFe]
hydrogenase in vitro from a nickel-free precursor form.
Biochemistry 35: 10089–10093.
Maier T, Jacobi A, Sauter M & Bock A (1993) The product of the
hypB gene, which is required for nickel incorporation into
hydrogenases, is a novel guanine nucleotide-binding protein. J
Bacteriol 175: 630–635.
Maier T, Lottspeich F & Bock A (1995) GTP hydrolysis by HypB is
essential for nickel insertion into hydrogenases of Escherichia
coli. Eur J Biochem 230: 133–138.
Manyani H, Rey L, Palacios JM, Imperial J & Ruiz-Argueso T
(2005) Gene products of the hupGHIJ operon are involved in
maturation of the iron–sulfur subunit of the [NiFe]
hydrogenase from Rhizobium leguminosarum bv. viciae. J
Bacteriol 187: 7018–7026.
Margheri MC, Tredici MR, Allotta G & Vagnoli L (1991)
Heterotrophic metabolism and regulation of uptake
hydrogenase activity in symbiotic cyanobacteria. Plant Soil
137: 139–144.
Masukawa H, Mochimaru M & Sakurai H (2002) Disruption of
the uptake hydrogenase gene, but not of the bi-directional
hydrogenase gene, leads to enhanced photobiological
hydrogen production by the nitrogen-fixing cyanobacterium
Anabaena sp. PCC 7120. Appl Microbiol Biotechnol 58:
618–624.
Meeks JC, Elhai J, Thiel T, Potts M, Larimer F, Lamerdin J, Predki
P & Atlas R (2001) An overview of the genome of Nostoc
punctiforme, a multicellular, symbiotic cyanobacterium.
Photosynth Res 70: 85–106.
Mehta N, Olson JW & Maier RJ (2003) Characterization of
Helicobacter pylori nickel metabolism accessory proteins
needed for maturation of both urease and hydrogenase. J
Bacteriol 185: 726–734.
Melis A, Neidhardt J & Benemann JR (1998) Dunaliella salina
(Chlorophyta) with small chlorophyll antenna sizes exhibit
higher photosynthetic productivities and photon use
efficiencies than normally pigmented cells. J Appl Phycol 10:
515–525.
Menon NK, Chatelus CY, Dervartanian M, Wendt JC,
Shanmugam KT, Peck HD Jr & Przybyla AE (1994) Cloning,
sequencing, and mutational analysis of the hyp operon
encoding Escherichia coli hydrogenase 2. J Bacteriol 176:
4416–4423.
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
717Cyanobacterial hydrogenases

Mikheeva LE, Schmitz O, Shestakov SV & Bothe H (1995)
Mutants of the cyanobacterium Anabaena variabilis altered in
hydrogenase activities. Z Naturforsch 50c: 505–510.
Miyagishima SY (2005) Origin and evolution of the chloroplast
division machinery. J Plant Res 118: 295–306.
Mulkidjanian AY, Koonin EV, Makarova KS et al. (2006) The
cyanobacterial genome core and the origin of photosynthesis.
Proc Natl Acad Sci USA 103: 13126–13131.
Mulrooney SB & Hausinger RP (2003) Nickel uptake and
utilization by microorganisms. FEMS Microbiol Rev 27:
239–261.
Nakajima Y & Ueda R (1997) Improvement of photosynthesis in
dense microalgal suspension by reduction of light harvesting
pigments. J Appl Phycol 9: 503–510.
Nakajima Y & Ueda R (1999) Improvement of microalgal
photosynthetic productivity by reducing the content of light
harvesting pigment. J Appl Phycol 11: 195–201.
Nakamura Y, Kaneko T, Hirosawa M, Miyajima N & Tabata S
(1998) CyanoBase, a www database containing the complete
nucleotide sequence of the genome of Synechocystis sp. strain
PCC6803. Nucleic Acids Res 26: 63–67.
Nakamura Y, Kaneko T, Sato S et al. (2003) Complete genome
structure of Gloeobacter violaceus PCC 7421, a cyanobacterium
that lacks thylakoids. DNA Res 10: 137–145.
Oliveira P & Lindblad P (2005) LexA, a transcription regulator
binding in the promoter region of the bidirectional
hydrogenase in the cyanobacterium Synechocystis sp. PCC
6803. FEMS Microbiol Lett 251: 59–66.
Oliveira P, Leitao E, Tamagnini P, Moradas-Ferreira P & Oxelfelt F
(2004) Characterization and transcriptional analysis of
hupSLW in Gloeothece sp. ATCC 27152: an uptake hydrogenase
from a unicellular cyanobacterium. Microbiology 150:
3647–3655.
Olson JW, Mehta NS & Maier RJ (2001) Requirement of nickel
metabolism proteins HypA and HypB for full activity of both
hydrogenase and urease in Helicobacter pylori. Mol Microbiol
39: 176–182.
Osanai T, Imamura S, Asayama M, Shirai M, Suzuki I, Murata N
& Tanaka K (2006) Nitrogen induction of sugar catabolic gene
expression in Synechocystis sp. PCC 6803. DNA Res 13:
185–195.
Oxelfelt F (1998) Hydrogenases in the cyanobacterium Nostoc sp.
strain PCC 73102. PhD Thesis, Faculty of Science and
Technology, Uppsala University, Sweden (ISBN 91-554-4290-
0).
Oxelfelt F, Tamagnini P, Salema R & Lindblad P (1995) Hydrogen
uptake in Nostoc strain PCC 73102: effects of nickel, hydrogen,
carbon and nitrogen. Plant Physiol Biochem 33: 617–623.
Oxelfelt F, Tamagnini P & Lindblad P (1998) Hydrogen uptake in
Nostoc sp. strain PCC 73102. Cloning and characterization of a
hupSL homologue. Arch Microbiol 169: 267–274.
Paschos A, Glass RS & Bock A (2001) Carbamoylphosphate
requirement for synthesis of the active center of [NiFe]-
hydrogenases. FEBS Lett 488: 9–12.
Paschos A, Bauer A, Zimmermann A, Zehelein E & Bock A (2002)
HypF, a carbamoyl phosphate-converting enzyme involved in
[NiFe] hydrogenase maturation. J Biol Chem 277:
49945–49951.
Patterson-Fortin LM, Colvin KR & Owttrim GW (2006) A LexA-
related protein regulates redox-sensitive expression of the
cyanobacterial RNA helicase, crhR. Nucl Acids Res 34:
3446–3454.
Polle JEW, Kanakagiri SD & Melis A (2003) tla1, a DNA
insertional transformant of the green alga Chlamydomonas
reinhardtii with a truncated light-harvesting chlorophyll
antenna size. Planta 217: 49–59.
Posewitz MC, King PW, Smolinski SL, Zhang LP, Seibert M &
Ghirardi ML (2004) Discovery of two novel radical S-
adenosylmethionine proteins required for the assembly of an
active [Fe] hydrogenase. J Biol Chem 279: 25711–25720.
Prince RC & Kheshgi HS (2005) The photobiological production
of hydrogen: potential efficiency and effectiveness as a
renewable fuel. Crit Rev Microbiol 31: 19–31.
Rai AN, Borthakur M, Soderback E & Bergman B (1992)
Immunogold localization of hydrogenase in the
cyanobacterial–plant symbioses Peltigera canina, Anthoceros
punctatus and Gunnera magellanica. Symbiosis 12: 131–144.
Rakhely G, Kovacs A, Maroti G, Fodor BD, Csanadi G, Latinovics
D & Kovacs KL (2004) Cyanobacterial-type,
heteropentameric, NAD1-reducing NiFe hydrogenase in the
purple sulfur photosynthetic bacterium Thiocapsa
roseopersicina. Appl Environ Microbiol 70: 722–728.
Reade JPH, Dougherty LJ, Rogers LJ & Gallon JR (1999) Synthesis
and proteolytic degradation of nitrogenase in cultures of the
unicellular cyanobacterium Gloeothece strain ATCC 27152.
Microbiology 145: 1749–1758.
Rees DC, Akif Tezcan F, Haynes CA, Walton MY, Andrade S,
Einsle O & Howard JB (2005) Structural basis of biological
nitrogen fixation. Philos Trans A Math Phys Eng Sci 363:
971–984.
Reissmann S, Hochleitner E, Wang H, Paschos A, Lottspeich F,
Glass RS & Bock A (2003) Taming of a poison: biosynthesis of
the NiFe-hydrogenase cyanide ligands. Science 299:
1067–1070.
Roseboom W, Blokesch M, Bock A & Albracht SP (2005) The
biosynthetic routes for carbon monoxide and cyanide in the
Ni–Fe active site of hydrogenases are different. FEBS Lett 579:
469–472.
Rossmann R, Maier T, Lottspeich F & Bock A (1995)
Characterisation of a protease from Escherichia coli involved in
hydrogenase maturation. Eur J Biochem 227: 545–550.
Sakamoto T, Delgaizo VB & Bryant DA (1998) Growth on urea
can trigger death and peroxidation of the cyanobacterium
Synechococcus sp. strain PCC 7002. Appl Environ Microbiol 64:
2361–2366.
Sakurai H & Masukawa H (2007) Promoting R&D in
Photobiological hydrogen production utilizing mariculture-
raised cyanobacteria. Mar Biotechnol 9: 128–145.
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
718 P. Tamagnini et al.

Schmitz O & Bothe H (1996) The diaphorase subunit HoxU of
the bidirectional hydrogenase as electron transferring protein
in cyanobacterial respiration? Naturwissenschaften 83:
525–527.
Schmitz O, Boison G, Hilscher R, Hundeshagen B, Zimmer W,
Lottspeich F & Bothe H (1995) Molecular biological analysis of
a bidirectional hydrogenase from cyanobacteria. Eur J Biochem
233: 266–276.
Schmitz O, Boison G & Bothe H (2001) Quantitative analysis of
expression of two circadian clock-controlled gene clusters
coding for the bidirectional hydrogenase in the
cyanobacterium Synechococcus sp. PCC7942. Mol Microbiol 41:
1409–1417.
Schmitz O, Boison G, Salzmann H, Bothe H, Schutz K, Wang S &
Happe T (2002) HoxE – a subunit specific for the pentameric
bidirectional hydrogenase complex (HoxEFUYH) of
cyanobacteria. Biochim Biophys Acta 1554: 66–74.
Schopf JW (2000) The fossil record: tracing the roots of the
cyanobacterial lineage. The Ecology of Cyanobacteria (Whitton
BA & Potts M, eds), pp. 13–35. Kluwer Academic Publishers,
Dordrecht, the Netherlands.
Schutz K, Happe T, Troshina O, Lindblad P, Leitao E, Oliveira P &
Tamagnini P (2004) Cyanobacterial H2 production – a
comparative analysis. Planta 218: 350–359.
Serebriakova LT, Zorin NA & Lindblad P (1994) Reversible
hydrogenase in Anabaena variabilis ATCC 29413: presence and
localization in non-N2-fixing cells. Arch Microbiol 161:
140–144.
Serebryakova LT, Sheremetieva ME & Lindblad P (2000) H2-
uptake and evolution in the unicellular cyanobacterium
Chroococcidiopsis thermalis CALU 758. Plant Physiol Biochem
38: 525–530.
Shapiguzov A, Lyukevich AA, Allakhverdiev SI, Sergeyenko TV,
Suzuki I, Murata N & Los DA (2005) Osmotic shrinkage of
cells of Synechocystis sp. PCC 6803 by water efflux via
aquaporins regulates osmostress-inducible gene expression.
Microbiology 151: 447–455.
Sheremetieva ME, Troshina OY, Serebryakova LT & Lindblad P
(2002) Identification of hox genes and analysis of their
transcription in the unicellular cyanobacterium Gloeocapsa
alpicola CALU 743 growing under nitrate-limiting conditions.
FEMS Microbiol Lett 214: 229–233.
Singh AK, Li H & Sherman LA (2004) Microarray analysis and
redox control of gene expression in the cyanobacterium
Synechocystis sp. PCC 6803. Physiol Plantarum 120: 27–35.
Slabas AR, Suzuki I, Murata N, Simon WJ & Hall JJ (2006)
Proteomic analysis of the heat shock response in Synechocystis
PCC6803 and a thermally tolerant knockout strain lacking the
histidine kinase 34 gene. Proteomics 6: 845–864.
Smith GD (1990) Hydrogen metabolism in cyanobacteria.
Phycotalk (Kumar EHD, ed), pp. 131–143. Rastogi & Co,
Meerut, India.
Spiro S & Guest JR (1990) FNR and its role in oxygen-regulated
gene expression in Escherichia coli. FEMS Microbiol Rev 6:
399–428.
Srivastava R, Pisareva T & Norling B (2005) Proteomic studies of
the thylakoid membrane of Synechocystis sp. PCC 6803.
Proteomics 5: 4905–4916.
Stal LJ & Moezelaar R (1997) Fermentation in cyanobacteria.
FEMS Microbiol Rev 21: 179–211.
Tamagnini P, Oxelfelt F, Salema R & Lindblad P (1995)
Immunological characterization of hydrogenases in the
nitrogen-fixing cyanobacterium Nostoc sp. strain PCC 73102.
Curr Microbiol 31: 102–107.
Tamagnini P, Troshina O, Oxelfelt F, Salema R & Lindblad P
(1997) Hydrogenases in Nostoc sp. strain PCC 73102, a strain
lacking a bidirectional enzyme. Appl Environ Microbiol 63:
1801–1807.
Tamagnini P, Costa J-L, Almeida L, Oliveira M-J, Salema R &
Lindblad P (2000) Diversity of cyanobacterial hydrogenases, a
molecular approach. Curr Microbiol 40: 356–361.
Tamagnini P, Axelsson R, Lindberg P, Oxelfelt F, Wunschiers R &
Lindblad P (2002) Hydrogenases and hydrogen metabolism of
cyanobacteria. Microbiol Mol Biol Rev 66: 1–20.
Tamagnini P, Leitao E & Oxelfelt F (2005) Uptake hydrogenase in
cyanobacteria: novel input from non-heterocystous strains.
Biochem Soc Trans 33: 67–69.
Tetali SD, Mitra M & Melis A (2007) Development of the light-
harvesting chlorophyll antenna in the green alga
Chlamydomonas reinhardtii is regulated by the novel Tla1
gene. Planta 225: 813–829.
Theodoratou E, Paschos A, Magalon A, Fritsche E, Huber R &
Bock A (2000a) Nickel serves as a substrate recognition motif
for the endopeptidase involved in hydrogenase maturation.
Eur J Biochem 267: 1995–1999.
Theodoratou E, Paschos A, Mintz-Weber S & Bock A (2000b)
Analysis of the cleavage site specificity of the endopeptidase
involved in the maturation of the large subunit of hydrogenase
3 from Escherichia coli. Arch Microbiol 173: 110–116.
Theodoratou E, Huber R & Bock A (2005) [NiFe]-hydrogenase
maturation endopeptidase: structure and function. Biochem
Soc Trans 33: 108–111.
Troshina OY, Serebryakova LT & Lindblad P (1996) Induction of
H2-uptake and nitrogenase activities in the cyanobacterium
Anabaena variabilis ATCC 29413: effects of hydrogen and
organic substrate. Curr Microbiol 33: 11–15.
Troshina O, Serebryakova LT, Sheremetieva ME & Lindblad P
(2002) Production of H2 by the unicellular cyanobacterium
Gloeocapsa alpicola CALU 743 during fermentation. Int J
Hydrogen Energy 27: 1283–1289.
Tu C-J, Shrager J, Burnap RL, Postier BL & Grossman AR (2004)
Consequences of a deletion in dspA on transcript
accumulation in Synechocystis sp. strain PCC 6803. J Bacteriol
186: 3889–3902.
Vignais PM & Colbeau A (2004) Molecular biology of microbial
hydrogenases. Curr Issues Mol Biol 6: 159–188.
Vignais PM, Billoud B & Meyer J (2001) Classification and
phylogeny of hydrogenases. FEMS Microbiol Rev 25: 455–501.
Volbeda A, Charon M-H, Piras C, Hatchikian EC, Frey M &
Fontecilla-Camps JC (1995) Crystal structure of the
FEMS Microbiol Rev 31 (2007) 692–720 c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
719Cyanobacterial hydrogenases

nickel–iron hydrogenase from Desulfovibrio gigas. Nature 373:
580–587.
Wang Y, Sun J & Chitnis PR (2000) Proteomic study of the
peripheral proteins from thylakoid membranes of the
cyanobacterium Synechocystis sp. PCC 6803. Electrophoresis 21:
1746–1754.
Weisshaar H & Boger P (1985) Pathways of hydrogen uptake in
the cyanobacterium Nostoc muscorum. Arch Microbiol 142:
349–353.
Whitton BA & Potts M (2000) Introduction to the cyanobacteria.
The Ecology of Cyanobacteria (Whitton BA & Potts M, eds), pp.
1–11. Kluwer Academic Publishers, Dordrecht, the
Netherlands.
Wolk CP, Ernest A & Elhai J (1994) Heterocyst metabolism and
development. The Molecular Biology of Cyanobacteria (Bryant
DA, ed), pp. 769–823. Kluwer Academic Publishers,
Dordrecht, the Netherlands.
Wunschiers R, Batur M & Lindblad P (2003) Presence and
expression of hydrogenase specific C-terminal endopeptidases
in cyanobacteria. BMC Microbiol 3: 8.
Xiankong Z, Tabita FR & Van Baalen C (1984) Nickel control of
hydrogen production and uptake in Anabaena spp. strains CA
and 1F. J Gen Microbiol 130: 1815–1818.
Xu Q, Yooseph S, Smith HO & Venter JC (2005) Development of
a novel recombinant cyanobacterial system for hydrogen
production from water. DOE Genomics: GTL Contractor-
Grantee Workshop III 63.
Yoshino F, Ikeda H, Masukawa H & Sakurai H (2007) High
photobiological hydrogen production activity of a Nostoc sp.
PCC 7422 uptake hydrogenase-deficient mutant with high
nitrogenase activity. Mar Biotechnol 9: 101–112.
Zhang X-H, Shiraiwa Y, Sui Z-H & Zhang X-C (2005a) Cloning
and characterization of hoxY gene from Arthrospira and
Spirulina and its application in phylogenetic studies. Periodical
Ocean Univ China 35: 1021–1025.
Zhang X, Zhang X, Shiraiwa Y, Mao Y, Sui Z & Liu J (2005b)
Cloning and characterization of hoxH genes from Arthrospira
and Spirulina and application in phylogenetic study. Mar
Biotechnol 7: 287–296.
Zhao Y, Bian S-M, Zhou H-N & Huang J-F (2006) Diversity of
nitrogenase systems in diazotrophs. J Int Plant Biol 48:
745–755.
FEMS Microbiol Rev 31 (2007) 692–720c� 2007 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
720 P. Tamagnini et al.








![Nickel-[iron-sulfur]-selenium-containing hydrogenases from Desulfovibrio baculatus (DSM 1743). Redox centers and catalytic properties](https://img.pdfslide.net/doc/110x75/634b40cae2b881b8bf01c78b/nickel-iron-sulfur-selenium-containing-hydrogenases-from-desulfovibrio-baculatus.jpg)




![Structural differences between the ready and unready oxidized states of [NiFe] hydrogenases](https://img.pdfslide.net/doc/110x75/63489e5f2cd4c1a3540d161c/structural-differences-between-the-ready-and-unready-oxidized-states-of-nife-hydrogenases.jpg)















