Embed Size (px)
Citation preview

Chemosphere 63 (2006) 175–181
www.elsevier.com/locate/chemosphere
Degradation of CL-20 by white-rot fungi
Diane Fournier, Fanny Monteil-Rivera, Annamaria Halasz,Manish Bhatt, Jalal Hawari *
Biotechnology Research Institute, National Research Council of Canada, Environmental Chemistry Group,
6100 Royalmount Avenue, Montreal, Que., Canada H4P 2R2
Received 22 March 2005; received in revised form 16 June 2005; accepted 23 June 2005Available online 19 August 2005
Abstract
In previous studies, we found that the emerging energetic chemical, CL-20 (C6H6N12O12, 2,4,6,8,10,12-hexanitro-2,4,6,8,10,12-hexaazaisowurtzitane), can be degraded following its initial denitration using both aerobic and anaerobicbacteria. The C and N mass balances were not determined due to the absence of labeled starting compounds. The pres-ent study describes the degradation of the emerging contaminant by Phanerochaete chrysosporium using ring-labeled[15N]-CL-20 and [14C]-CL-20. Ligninolytic cultures degraded CL-20 with the release of nitrous oxide (N2O) in amountscorresponding to 45% of the nitrogen content of CL-20. When ring-labeled [15N]-CL-20 was used, both 14N14NO and15N14NO were observed, likely produced from –NO2 and N–NO2, respectively. The incubation of uniformly labeled[14C]-CL-20 with fungi led to the production of 14CO2 (>80%). Another ligninolytic fungus, Irpex lacteus, was also ableto degrade CL-20, but as for P. chrysosporium, no early intermediates were observed. When CL-20 was incubated withmanganese peroxidase (MnP), we detected an intermediate with a [M�H]� mass ion at 345 Da (or 351 and 349 Dawhen using ring-labeled and nitro-labeled [15N]-CL-20, respectively) matching a molecular formula of C6H6N10O8.The intermediate was thus tentatively identified as a doubly denitrated CL-20 product. The concomitant release ofnitrite ions (NO�2 ) with CL-20 degradation by MnP also supported the occurrence of an initial denitration prior tocleavage and decomposition.� 2005 Elsevier Ltd. All rights reserved.
Keywords: CL-20; Degradation; Nitramine; Phanerochaete chrysosporium; Fungi
1. Introduction
The environmental fate and impact of the emergingenergetic chemical CL-20, C6H6N12O12 (compound 1,Fig. 1) are not well defined. Preliminary toxicologicalstudies indicated that CL-20 is not acutely toxic to the
0045-6535/$ - see front matter � 2005 Elsevier Ltd. All rights reservdoi:10.1016/j.chemosphere.2005.06.052
* Corresponding author. Tel.: +1 514 496 6267; fax: +1 514496 6265.
E-mail address: [email protected] (J. Hawari).
marine bacteria Vibrio fisheri, freshwater green algaeSelenastrum capricornutum, terrestrial higher plants(perennial ryegrass and alfalfa), and indigenous soilmicroorganisms (Gong et al., 2004). However, the poly-cyclic nitramine has significant lethal and sublethaleffects on the earthworm Eisenia andrei (Robidouxet al., 2004). At lower doses, the toxicity of the nitraminewas more important in a sandy soil than in an organicrich soil (Robidoux et al., 2004), a phenomenon ex-plained by the favored sorption of CL-20 to soil organicfraction (Balakrishnan et al., 2004a).
ed.

N
N
N
NN
N
NO2
NO2NO2
NO2
2a Doubly denitrated CL-20
Abiotic secondary decomposition
Fe0 (Balakrishnan et al. 2004b)
Nitroreductase (Bhushan et al. 2004b)
1 CL-20NO2
O2N
O2N
O2NNO2NN
N
NN
N
NO2
CHOCHO + N2O + HCOO- + NH3
NO2-
NO2-
N
N
N
NN
N
NO2
NO2
NO2NO2
2b Isomer of compound 2a
+
Fig. 1. Structure of CL-20 (1) and CL-20 denitration products (2a and 2b) observed during treatment with Fe0 (Balakrishnan et al.,2004b) or with nitroreductase (Bhushan et al., 2004b).
176 D. Fournier et al. / Chemosphere 63 (2006) 175–181
While two aerobic RDX-degrading strains (Rhodo-
coccus sp. DN22 and Rhodococcus rhodochrous 11Y)failed to degrade CL-20 (unpublished data), several aero-bic and anaerobic microorganisms degraded the ener-getic compound (Trott et al., 2003; Bhushan et al.,2003, 2004a; Karakaya et al., 2004). CL-20 was foundto transform via an N-denitration mechanism, as con-firmed by the detection of nitrite ions (NO�2 Þ and puta-tive doubly denitrated CL-20 intermediates (compound2a and its isomer 2b, Fig. 1) during treatment with nitro-reductase (Bhushan et al., 2004b). The proposed deni-trated intermediates have also been observed whenreacting CL-20 with zero valent iron (Balakrishnanet al., 2004b). In the biotic like in the abiotic processes,compound 2a and its isomer 2b degraded into the end-products nitrous oxide (N2O), formate (HCOO�),ammonia (NH3), and glyoxal (CHOCHO) (Fig. 1).
Despite the important progresses that have beenachieved in the understanding of CL-20 degradationpathways, the fate of carbons from the emerging chem-ical is not thoroughly known. In the present study, weassessed the ability of the ligninolytic white-rot fungi,Phanerochaete chrysosporium and Irpex lacteus, to de-grade CL-20. For the first time, the carbon mass balancewas determined using the radiotracer [14C]-CL-20.
P. chrysosporium and I. lacteus are lignin degraders thatsynthesize nonspecific enzymes such as lignin peroxidase(LiP), manganese peroxidase (MnP), and other criticalmetabolic components such as oxidases, reductases,hydrogen peroxide, veratryl alcohol, oxalate, and qui-nones (Kersten et al., 1985; Stahl and Aust, 1995).Due to the complex nature of fungal metabolic chemis-try, we used MnP to get insight into the degradationmechanism.
2. Material and methods
2.1. Chemicals
2,4,6,8,10,12-Hexanitro-2,4,6,8,10,12-hexa-azaisowurtzitane (e-CL-20), ring-labeled [15N]-CL-20,nitro-labeled 15N-[CL-20], and uniformly labeled [14C]-CL-20 (chemical purity > 97%, specific activity of0.75 mCi/g) were provided by A.T.K. Thiokol Propul-sion (Brigham City, UT). [1,2-14C]-glyoxal (chemicalpurity > 99%, specific activity of 56 mCi/mmol) wasfrom American Radio-labeled Chemicals, Inc. (St.Louis, MO). All other chemicals used in this study wereof reagent grade.

D. Fournier et al. / Chemosphere 63 (2006) 175–181 177
2.2. Bio-transformation of CL-20 in liquid culture
by P. chrysosporium and I. lacteus
P. chrysosporium strain ATCC 24725 and I. lacteus
(obtained from Dr. V. Sasek, Institute of Microbiology,Prague, Czech Republic) were maintained on YPDplates, and were cultivated in a modified Kirk�s nitro-gen-limited medium (pH 4.5) as previously described(Fournier et al., 2004). Kirk�s modified medium containsa carbon source (10 g/l glucose) and a limited source ofnitrogen (1.2 mM ammonium tartrate), which are en-ough to promote the initial development of fungal myce-lia and induce the production of ligninolytic enzymes (themedium is referred below as the N-limited mineral med-ium). Bio-transformation experiments were performed inthe N-limited mineral medium (10 ml in 125 ml serumbottle) inoculated with 2 · 106 fungal spores. CL-20(stock solution of approximately 15000 mg l�1, preparedin acetone) was added to the spore suspension. Heat-killed fungal spores were incubated with CL-20 as con-trols for possible abiotic degradation. The bottles weresealed with Teflon coated serum septa and aluminumcaps. The cultures were incubated statically in the darkat 37 ± 2 �C (for P. chrysosporium) and 20 ± 2 �C forI. lacteus, and were aerated every 3–4 days.
2.3. Mineralization of CL-20 and glyoxal with fungi
Mineralization experiments were performed by add-ing the radiotracer [14C]-CL-20 (0.048 lCi, 162.6 nmol)or [14C]-glyoxal (0.288 lCi, 3100 nmol) to P. chrysospo-
rium spore suspensions. Formation of 14CO2 was moni-tored as described previously (Fournier et al., 2004).Once again, the microcosms were aerated every 3–4days.
2.4. Bio-transformation of CL-20 with MnP
MnP from a high potential fermentation strain ofNemalotoma frowardii was purchased from Jena Biosci-ence GmbH (Jena, Germany). The MnP reactions wereperformed in 6 ml vials in a total of 2 ml: sodium malon-ate (50 mM, pH 4.5), MnCl2 (1 mM), H2O2 (0.33 mM),MnP (3 U) and CL-20 (38 mg l�1 using the above-de-scribed stock solution). CL-20 was present at a concentra-tion superior to its aqueous solubility (Monteil-Riveraet al., 2004) in an attempt to generate sufficient amountsof reaction intermediates to allow detection. The vialswere sealed with Teflon coated serum septa and alumi-num caps. Reactions were carried out at 37 ± 2 �C underagitation at 150 rpm.
2.5. Analytical techniques
After sampling the headspace for N2O analysis(Fournier et al., 2004), the liquid phase of fungal cul-
tures or enzymatic reaction were sacrificed by addingone equivalent volume of acidified acetonitrile (0.25 mlof concentrated H2SO4 per liter of acetonitrile) to onevolume of reaction mixture. Whole fungal cultures weresubmitted to sonication at 20 �C for 18 h. In all cases,the resulting solution was then filtered through a0.45 lm membrane (Millipore PTFE), the first 3 mlbeing discarded. CL-20 was analyzed by HPLC con-nected to a photodiode array (PDA), as described previ-ously (Monteil-Rivera et al., 2004). Analyses of nitrate(NO�3 ), formate (HCOO�), glycolate (HOCH2COO�),ammonia (NH3), and glyoxal (HCOHCO) were also per-formed as previously described (Balakrishnan et al.,2004b). Intermediate products of CL-20 were analyzedusing a Bruker Esquire 3000 plus ion trap spectrometer(Bruker-Daltonics, Boston, MA, USA) equipped withan Agilent 1100 HPLC system (Agilent, Waldbronn,Germany). The atmospheric electrospray source wasoperated in negative mode using nitrogen as a dryinggas (15.0 psi), at a flow rate of 5 l/min and a temperatureof 150 �C. The capillary voltage was set at 4000 V withan end plate offset of �500 V. The scanning mass tocharge range was 40–550 m/z. Details on the LC-MSset up can be found in Hawari et al. (2004). Nitrite(NO�2 ) was analyzed using the photometric US EPAMethod 354.1 (US EPA, 1979).
3. Results and discussion
3.1. Bio-transformation of CL-20 by fungi
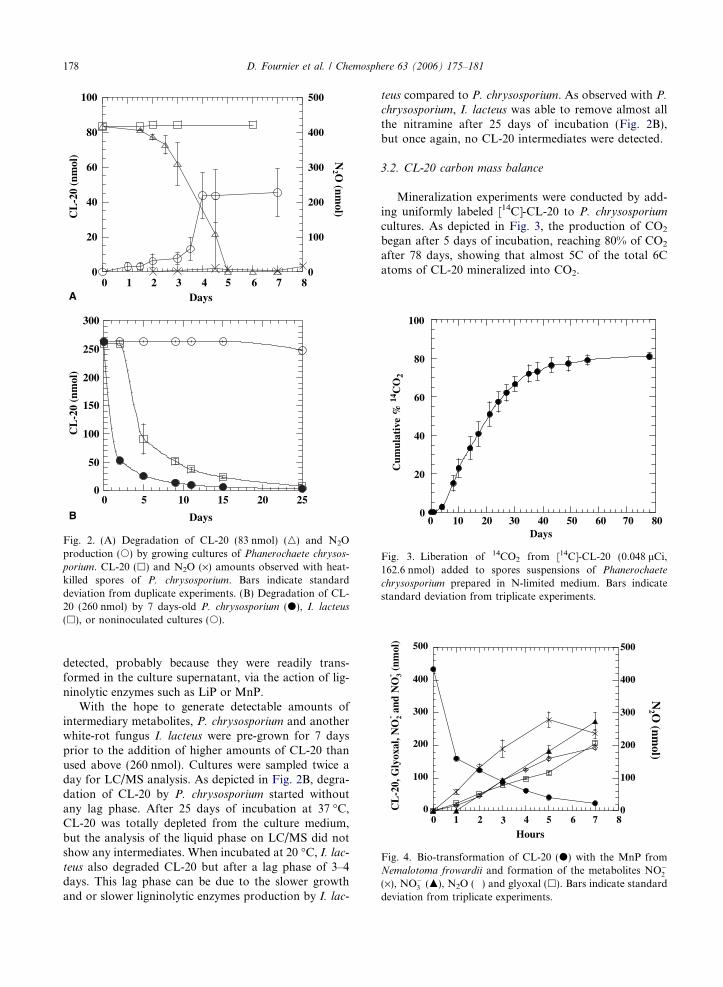
Degradation of CL-20 added to spores suspension ofP. chrysosporium was monitored for up to 8 days. Fig. 2Ashows the disappearance of CL-20 in the presence ofgrowing or heat-killed fungal cultures. In heat-killed con-trols the initial amount of CL-20 (85 nmol) remained sta-ble during all the incubation period. The degradationwith active P. chrysosporium started after a lag phaseof 2 days (Fig. 2A). After 5 days of incubation, CL-20was totally depleted from the culture medium. The for-mation of N2O started after 3 days of incubation(Fig. 2A). On the seventh day, the ratio of N2O overCL-20 degraded indicated that 45% of the nitrogen con-tent of CL-20 was transformed into nitrous oxide. Whenring-labeled [15N]-CL-20 was used in degradation experi-ments, the GC/MS analysis of N2O showed the presenceof both 14N14NO and 15N14NO, likely produced fromperipheral nitro (–NO2) and the nitramine (N–NO2)groups, respectively. No nitrite ions (NO�2 Þ were de-tected, but when P. chrysosporium was cultivated withNaNO2 as the sole nitrogen source, the growth of themycelia was observed with the concomitant productionof N2O. It should be noted that no growth and no N2Oproduction were observed in the absence of either CL-20 or NaNO2. No CL-20 intermediary metabolites were
0
20
40
60
80
100
0
100
200
300
400
500
CL
-20
(nm
ol)
CL
-20
(nm
ol)
N2 O
(nmol)
Days
Days
0
50
100
150
200
250
300
0
0 1 2 3 4 5 6 7 8
5 10 15 20 25
A
B
Fig. 2. (A) Degradation of CL-20 (83 nmol) (n) and N2Oproduction (s) by growing cultures of Phanerochaete chrysos-
porium. CL-20 (h) and N2O (·) amounts observed with heat-killed spores of P. chrysosporium. Bars indicate standarddeviation from duplicate experiments. (B) Degradation of CL-20 (260 nmol) by 7 days-old P. chrysosporium (d), I. lacteus
(h), or noninoculated cultures (s).
0 10 20 30 40 50 60 70 800
20
40
60
80
100
Days
Cum
ulat
ive
% 14
CO
2
Fig. 3. Liberation of 14CO2 from [14C]-CL-20 (0.048 lCi,162.6 nmol) added to spores suspensions of Phanerochaete
chrysosporium prepared in N-limited medium. Bars indicatestandard deviation from triplicate experiments.
0
100
200
300
400
500
0 1 2 3 4 5 6 7 80
100
200
300
400
500
N2 O
(nmol)
Hours
--
CL
-20,
Gly
oxal
, NO
2 an
d N
O3
(nm
ol)
Fig. 4. Bio-transformation of CL-20 (d) with the MnP fromNemalotoma frowardii and formation of the metabolites NO�2(·), NO�3 (m), N2O (�) and glyoxal (h). Bars indicate standarddeviation from triplicate experiments.
178 D. Fournier et al. / Chemosphere 63 (2006) 175–181
detected, probably because they were readily trans-formed in the culture supernatant, via the action of lig-ninolytic enzymes such as LiP or MnP.
With the hope to generate detectable amounts ofintermediary metabolites, P. chrysosporium and anotherwhite-rot fungus I. lacteus were pre-grown for 7 daysprior to the addition of higher amounts of CL-20 thanused above (260 nmol). Cultures were sampled twice aday for LC/MS analysis. As depicted in Fig. 2B, degra-dation of CL-20 by P. chrysosporium started withoutany lag phase. After 25 days of incubation at 37 �C,CL-20 was totally depleted from the culture medium,but the analysis of the liquid phase on LC/MS did notshow any intermediates. When incubated at 20 �C, I. lac-
teus also degraded CL-20 but after a lag phase of 3–4days. This lag phase can be due to the slower growthand or slower ligninolytic enzymes production by I. lac-
teus compared to P. chrysosporium. As observed with P.
chrysosporium, I. lacteus was able to remove almost allthe nitramine after 25 days of incubation (Fig. 2B),but once again, no CL-20 intermediates were detected.
3.2. CL-20 carbon mass balance
Mineralization experiments were conducted by add-ing uniformly labeled [14C]-CL-20 to P. chrysosporium
cultures. As depicted in Fig. 3, the production of CO2
began after 5 days of incubation, reaching 80% of CO2
after 78 days, showing that almost 5C of the total 6Catoms of CL-20 mineralized into CO2.
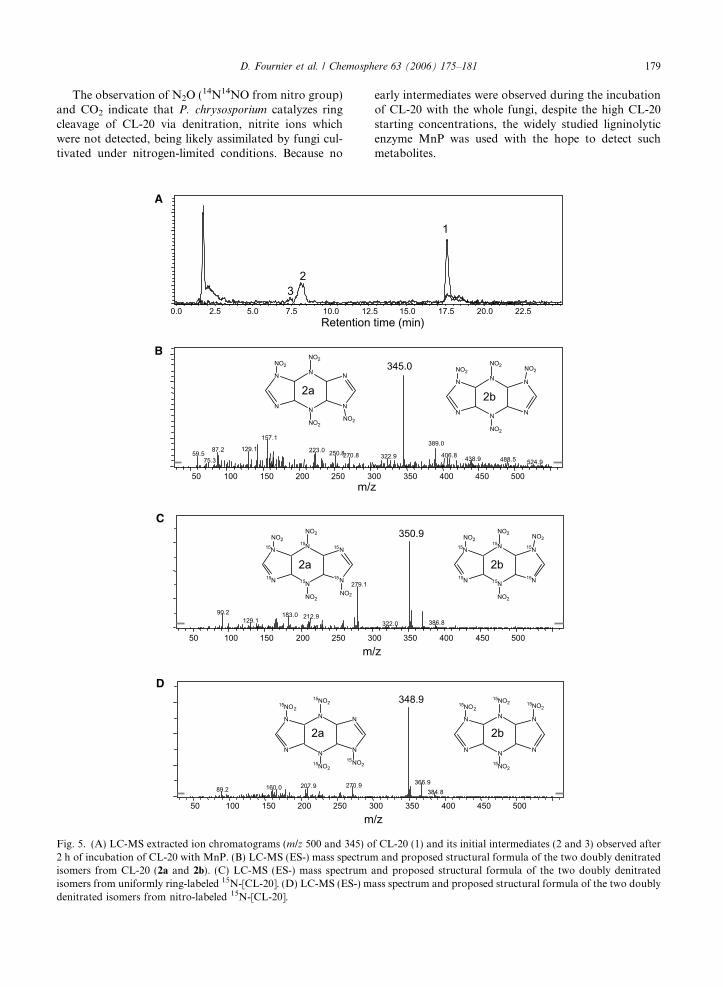
D. Fournier et al. / Chemosphere 63 (2006) 175–181 179
The observation of N2O (14N14NO from nitro group)and CO2 indicate that P. chrysosporium catalyzes ringcleavage of CL-20 via denitration, nitrite ions whichwere not detected, being likely assimilated by fungi cul-tivated under nitrogen-limited conditions. Because no
50 100 150 200 250 3
m
Retention
A
B
23
59.575.3
87.2 129.1
157.1
223.0 250.8270.8
0.0 2.5 5.0 7.5 10.0 12.
N
N
N
NN
N
NO2NO2
NO2NO2
15N
15N
15N
15N15N
15N
NO2NO2
NO2NO2
50 100 150 200 250 3m/z
C
2a
2a
N
N
N
NN
N
15NO2
15NO2
15NO2
15NO2
2a
90.2129.1
183.0 212.9
279.1
89.2 160.0 207.9 270.9
50 100 150 200 250 3
m
D
Fig. 5. (A) LC-MS extracted ion chromatograms (m/z 500 and 345) o2 h of incubation of CL-20 with MnP. (B) LC-MS (ES-) mass spectrumisomers from CL-20 (2a and 2b). (C) LC-MS (ES-) mass spectrumisomers from uniformly ring-labeled 15N-[CL-20]. (D) LC-MS (ES-) mdenitrated isomers from nitro-labeled 15N-[CL-20].
early intermediates were observed during the incubationof CL-20 with the whole fungi, despite the high CL-20starting concentrations, the widely studied ligninolyticenzyme MnP was used with the hope to detect suchmetabolites.
00 350 400 450 500
/z
time (min)
1
322.9
345.0
389.0
406.8 438.9 488.5 524.9
5 15.0 17.5 20.0 22.5
N
N
N
NN
N
NO2NO2
NO2
NO2
00 350 400 450 500
15N
15N
15N
15N15N
15N
NO2NO2
NO2
NO2
2b
2b
N
N
N
NN
N
15NO2
15NO2
15NO2
15NO2
2b
322.0
350.9
386.8
348.9
366.9384.8
00 350 400 450 500
/z
f CL-20 (1) and its initial intermediates (2 and 3) observed afterand proposed structural formula of the two doubly denitrated
and proposed structural formula of the two doubly denitratedass spectrum and proposed structural formula of the two doubly
0 10 20 30 40 50 60 70 800
20
40
60
80
100
Days
Cum
ulat
ive
% 14
CO
2
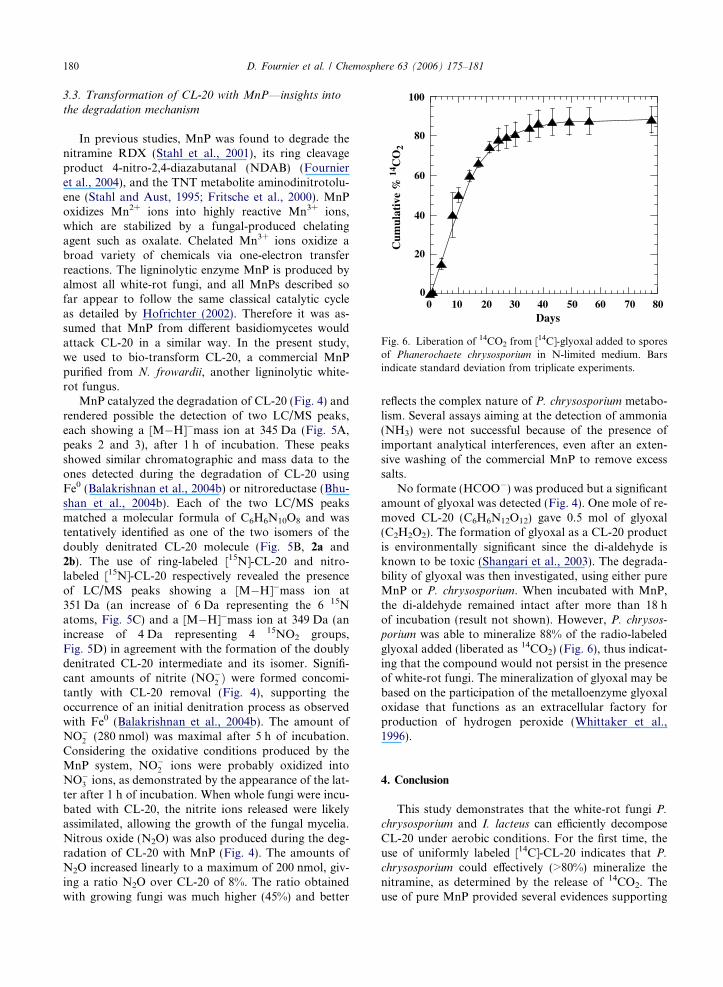
Fig. 6. Liberation of 14CO2 from [14C]-glyoxal added to sporesof Phanerochaete chrysosporium in N-limited medium. Barsindicate standard deviation from triplicate experiments.
180 D. Fournier et al. / Chemosphere 63 (2006) 175–181
3.3. Transformation of CL-20 with MnP—insights into
the degradation mechanism
In previous studies, MnP was found to degrade thenitramine RDX (Stahl et al., 2001), its ring cleavageproduct 4-nitro-2,4-diazabutanal (NDAB) (Fournieret al., 2004), and the TNT metabolite aminodinitrotolu-ene (Stahl and Aust, 1995; Fritsche et al., 2000). MnPoxidizes Mn2+ ions into highly reactive Mn3+ ions,which are stabilized by a fungal-produced chelatingagent such as oxalate. Chelated Mn3+ ions oxidize abroad variety of chemicals via one-electron transferreactions. The ligninolytic enzyme MnP is produced byalmost all white-rot fungi, and all MnPs described sofar appear to follow the same classical catalytic cycleas detailed by Hofrichter (2002). Therefore it was as-sumed that MnP from different basidiomycetes wouldattack CL-20 in a similar way. In the present study,we used to bio-transform CL-20, a commercial MnPpurified from N. frowardii, another ligninolytic white-rot fungus.
MnP catalyzed the degradation of CL-20 (Fig. 4) andrendered possible the detection of two LC/MS peaks,each showing a [M�H]�mass ion at 345 Da (Fig. 5A,peaks 2 and 3), after 1 h of incubation. These peaksshowed similar chromatographic and mass data to theones detected during the degradation of CL-20 usingFe0 (Balakrishnan et al., 2004b) or nitroreductase (Bhu-shan et al., 2004b). Each of the two LC/MS peaksmatched a molecular formula of C6H6N10O8 and wastentatively identified as one of the two isomers of thedoubly denitrated CL-20 molecule (Fig. 5B, 2a and2b). The use of ring-labeled [15N]-CL-20 and nitro-labeled [15N]-CL-20 respectively revealed the presenceof LC/MS peaks showing a [M�H]�mass ion at351 Da (an increase of 6 Da representing the 6 15Natoms, Fig. 5C) and a [M�H]�mass ion at 349 Da (anincrease of 4 Da representing 4 15NO2 groups,Fig. 5D) in agreement with the formation of the doublydenitrated CL-20 intermediate and its isomer. Signifi-cant amounts of nitrite (NO�2 Þ were formed concomi-tantly with CL-20 removal (Fig. 4), supporting theoccurrence of an initial denitration process as observedwith Fe0 (Balakrishnan et al., 2004b). The amount ofNO�2 (280 nmol) was maximal after 5 h of incubation.Considering the oxidative conditions produced by theMnP system, NO�2 ions were probably oxidized intoNO�3 ions, as demonstrated by the appearance of the lat-ter after 1 h of incubation. When whole fungi were incu-bated with CL-20, the nitrite ions released were likelyassimilated, allowing the growth of the fungal mycelia.Nitrous oxide (N2O) was also produced during the deg-radation of CL-20 with MnP (Fig. 4). The amounts ofN2O increased linearly to a maximum of 200 nmol, giv-ing a ratio N2O over CL-20 of 8%. The ratio obtainedwith growing fungi was much higher (45%) and better
reflects the complex nature of P. chrysosporium metabo-lism. Several assays aiming at the detection of ammonia(NH3) were not successful because of the presence ofimportant analytical interferences, even after an exten-sive washing of the commercial MnP to remove excesssalts.
No formate (HCOO�) was produced but a significantamount of glyoxal was detected (Fig. 4). One mole of re-moved CL-20 (C6H6N12O12) gave 0.5 mol of glyoxal(C2H2O2). The formation of glyoxal as a CL-20 productis environmentally significant since the di-aldehyde isknown to be toxic (Shangari et al., 2003). The degrada-bility of glyoxal was then investigated, using either pureMnP or P. chrysosporium. When incubated with MnP,the di-aldehyde remained intact after more than 18 hof incubation (result not shown). However, P. chrysos-
porium was able to mineralize 88% of the radio-labeledglyoxal added (liberated as 14CO2) (Fig. 6), thus indicat-ing that the compound would not persist in the presenceof white-rot fungi. The mineralization of glyoxal may bebased on the participation of the metalloenzyme glyoxaloxidase that functions as an extracellular factory forproduction of hydrogen peroxide (Whittaker et al.,1996).
4. Conclusion
This study demonstrates that the white-rot fungi P.
chrysosporium and I. lacteus can efficiently decomposeCL-20 under aerobic conditions. For the first time, theuse of uniformly labeled [14C]-CL-20 indicates that P.
chrysosporium could effectively (>80%) mineralize thenitramine, as determined by the release of 14CO2. Theuse of pure MnP provided several evidences supporting

D. Fournier et al. / Chemosphere 63 (2006) 175–181 181
the occurrence of an initial denitration step on CL-20prior to ring cleavage and decomposition. The detectionof glyoxal as a CL-20 metabolite and its mineralizationby P. chrysosporium indicate that the di-aldehyde is aCL-20 product but that it would not persist in the pres-ence of white-rot fungi. This study demonstrates thatCL-20 would not be persistent in environments that fa-vor the growth of ligninolytic fungi.
Acknowledgements
The authors wish to thank Alain Corriveau, ZoranaRadovich-Hrapovich, Dominic Manno, and ChantaleBeaulieu for their technical assistance. We also thankThiokol Propulsion (Brigham City, UT) for providingCL-20, ring-labeled and nitro-labeled [15N]-CL-20, anduniformly-labeled [14C]-CL-20. We are grateful to theU.S. Strategic Environmental Research and Develop-ment Program (SERDP CP-1256) and to Defense Re-search and Development Canada (DRDC), Valcartier,Canada for financial support.
References
Balakrishnan, V.K., Monteil-Rivera, F., Gautier, M., Hawari,J., 2004a. Sorption and stability of the polycyclic nitramineexplosive CL-20 in soil. J. Environ. Qual. 33, 1362–1368.
Balakrishnan, V.K., Monteil-Rivera, F., Halasz, A., Corbeanu,A., Hawari, J., 2004b. Decomposition of the polycyclicnitramine explosive, CL-20, by Fe0. Environ. Sci. Technol.38, 6861–6866.
Bhushan, B., Paquet, L., Spain, J.C., Hawari, J., 2003.Biotransformation of 2,4,6,8,10,12-hexanitro-2,4,6,8,10,12-hexaazaisowurtzitane (CL-20) by denitrifying Pseudomonas
sp. strain FA1. Appl. Environ. Microbiol. 69, 5216–5221.Bhushan, B., Halasz, A., Thiboutot, S., Ampleman, G.,
Hawari, J., 2004a. Chemotaxis-mediated biodegradationof cyclic nitramine explosives RDX HMX and CL-20 byClostridium sp. EDB2. Biochem. Biophys. Res. Co. 316,816–821.
Bhushan, B., Halasz, A., Hawari, J., 2004b. Nitroreductasecatalyzed biotransformation of CL-20. Biochem. Biophys.Res. Co. 322, 271–276.
Fournier, D., Halasz, A., Spain, J., Spanggord, R.J., Bottaro,J.C., Hawari, J., 2004. Biodegradation of the hexahydro-1,3,5-trinitro-1,3,5-triazine ring cleavage product 4-nitro-2,4-diazabutanal (NDAB) by Phanerochaete chrysosporium.Appl. Environ. Microbiol. 70, 1123–1128.
Fritsche, W., Scheibner, K., Herre, A., Hofrichter, M., 2000.Fungal degradation of explosives: TNT and related nitro-aromatic compounds, book chapter. In: Spain, J.C.,
Hughes, J.B., Knackmuss, H.-J. (Eds.), Biodegradation ofNitroaromatic Compounds and Explosives. CRC Press,Boca Raton, FL, pp. 213–237.
Gong, P., Sunahara, G.I., Rocheleau, S., Dodard, S.G.,Robidoux, P.Y., Hawari, J., 2004. Preliminary ecotoxico-logical characterization of a new energetic substance, CL-20. Chemosphere 56, 653–658.
Hawari, J., Deschamps, S., Beaulieu, C., Paquet, L., Halasz, A.,2004. Photodegradation of CL-20: insights into the mech-anisms of initial reactions and environmental fate. WaterRes. 38, 4044–4064.
Hofrichter, M., 2002. Review: lignin conversion by manganeseperoxidase (MnP). Enzyme Microb. Tech. 30, 454–466.
Karakaya, P., Sidhoum, M., Christodoulatos, C., 2004. Bioki-netic study for degradation of CL-20 with Phanerochaete
chrysosporium (ATCC-24725). In: Proceedings of the WestCoast Conference on Soils, Sediments and Water, March15–18, San Diego, CA.
Kersten, P.J., Tien, M., Kalyanaraman, B., Kirk, T.K., 1985.The ligninase of Phanerochaete chrysosporium generatescation radicals from methoxybenzenes. J. Biol. Chem. 260,2609–2612.
Monteil-Rivera, F., Paquet, L., Deschamps, S., Balakrishnan,V.K., Beaulieu, C., Hawari, J., 2004. Physico-chemicalmeasurements of CL-20 for environmental applications.Comparison with RDX and HMX. J. Chromatogr. A 1025,125–132.
Robidoux, P.Y., Sunahara, G.I., Savard, K., Berthelot, Y.,Leduc, F., Dodard, S., Martel, M., Gong, P., Hawari, J.,2004. Acute and chronic toxicity of the new explosive CL-20to the earthworm (Eisenia andrei) exposed to amendednatural soils. Environ. Toxicol. Chem. 23, 1026–1034.
Shangari, N., Bruce, W.R., Poon, R., O�Brien, P.J., 2003.Toxicity of glyoxals—role of oxidative stress, metabolicdetoxification and thiamine deficiency. Biochem. Soc.Trans. 31, 1390–1393.
Stahl, J.D., Aust, S.D., 1995. Biodegradation of 2,4,6-trinitro-toluene by the white rot fungus Phanerochaete chrysospo-
rium. In: Spain, J.C. (Ed.), Biodegradation of NitroaromaticCompounds. Plenum Press, New York, pp. 117–133.
Stahl, J.D., Van Aken, B., Cameron, M.D., Aust, S.D., 2001.Hexahydro-1,3,5-trinitro-1,3,5-triazine (RDX) biodegrada-tion in liquid and solid-state matrices by Phanerochaete
chrysosporium. Bioremed. J. 5, 13–25.Trott, S., Nishino, S.F., Hawari, J., Spain, J.C., 2003. Biode-
gradation of the nitramine explosive, CL-20. Appl. Environ.Microbiol. 69, 1871–1874.
US Environmental Protection Agency, 1979. Method 354.1 forchemical analysis of water and wastes. Nitrogen-Nitrite.Environmental monitoring and support laboratory office ofresearch and development, Cincinnati, OH.
Whittaker, M.M., Kersten, P.J., Nakamura, N., Sanders-Loehr, J., Schweizer, E.S., Whittaker, J.W., 1996. Glyoxaloxidase from Phanerochaete chrysosporium is a new radical-copper oxidase. J. Biol. Chem. 271, 681–687.





























