Embed Size (px)
Citation preview

CELL BIOLOGY AND MORPHOGENESIS
Dehydrin genes and their expression in recalcitrant oak(Quercus robur) embryos
Vanda Sunderlıkova Æ Jan Salaj Æ Dieter Kopecky ÆTerezia Salaj Æ Eva Wilhem Æ Ildiko Matusıkova
Received: 23 January 2009 / Revised: 23 April 2009 / Accepted: 26 April 2009 / Published online: 24 May 2009
� Springer-Verlag 2009
Abstract In this work, three dehydrin genes, QrDhn1,
QrDhn2, QrDhn3, were isolated from recalcitrant oak
(Quercus robur). Their expression pattern was analyzed in
both zygotic and somatic embryos as well as in vegetative
tissues exposed to different kinds of abiotic stresses
including desiccation, osmotic stress, and chilling. The
QrDhn1 gene encoding for YnSKn type dehydrin was
expressed during later stages of zygotic embryo develop-
ment but in somatic embryos only when exposed to
osmotic or desiccation stress. In contrast, the other two oak
dehydrin genes encoding for putative Kn type dehydrins
were expressed only in somatic embryos (both not-treated
and osmotically stressed) and leaves of oak seedlings
exposed to desiccation. Behavior of these genes suggests
that different dehydrins are involved in processes of seed
maturation and response to altered osmotic (water status)
conditions in somatic embryos. Revealing further members
of dehydrin gene family in recalcitrant oak might con-
tribute to clarify non-orthodox seed behavior as well as
identify mechanisms contributing to desiccation tolerance
in plants.
Keywords LEA-2 genes � Seed desiccation �Somatic embryos � Zygotic embryos
Introduction
The species Quercus robur L. has desiccation-sensitive
(recalcitrant) seeds, which cannot be stored for long peri-
ods without serious loss of viability (Roberts 1973). In
contrast to orthodox species, recalcitrant seeds sustain
metabolic activity throughout ontogeny but burst the seed
tissues shortly after dispersal. The term recalcitrant is
generally applied to seeds that reveal immediate germina-
tion after shedding, low longevity, and desiccation intol-
erance (Barbedo and Bilia 1998). Although water is
essential for these seeds and its uncontrolled loss exerts
deleterious impacts on metabolism and development, some
decline in water content prior to shedding has been recor-
ded for seeds of several temperate species, e.g. Acer
pseudoplatanus and Q. robur (Finch-Savage and Blake
1994), leading to suggestion that measure of desiccation
tolerance might be acquired during development. Never-
theless, further dehydration is deleterious, indicating that at
least some of the defense mechanisms necessary for com-
plete desiccation tolerance are not entrained (Berjak and
Pammenter 2008).
Dehydrins (DHNs), the group 2 of LEA (late embryo-
genesis abundant) proteins, are one of the most extensively
studied putative dehydration protective molecules. Under
water deficit, these proteins often comprise a significant
portion (1–10%) of the soluble proteins induced (Close
1996). In mature embryos and in drought-stressed cereal
seedlings, for example, dehydrins alone typically accu-
mulate to levels [1–2% of the total soluble protein
extractable in low-salt, aqueous buffers (Ceccardi et al.
Communicated by A. Feher.
V. Sunderlıkova � J. Salaj � T. Salaj � I. Matusıkova (&)
Institute of Plant Genetics and Biotechnology,
Slovak Academy of Sciences, Akademicka 2,
P.O. Box 39A, 950 07 Nitra 1, Slovak Republic
e-mail: [email protected]
V. Sunderlıkova � J. Salaj � D. Kopecky � E. Wilhem �I. Matusıkova
Department of Health and Environment/Bioresources,
Austrian Research Centers GmbH-ARC,
2444 Seibersdorf, Austria
123
Plant Cell Rep (2009) 28:1011–1021
DOI 10.1007/s00299-009-0710-6

1994). According to Kermode (1997), dehydrins are part of
the developmental program of orthodox seeds, but they
have also been detected in the recalcitrant seeds of Acer
saccharinum, Aesculus hippocastanum, Araucaria angust-
ifolia, Camellia sinensis, Castanea sativa, Poncirus trifo-
liate, all of which are of temperate origin (Farrant et al.
1996). In contrast, in some other recalcitrant species they
accumulated only when individuals have developed in a
temperate climate, after exposure to additional water loss
or in response to an increase in abscisic acid accumulation
(Farrant et al. 1996). In various tropical wetland recalci-
trant species (e.g. Avicenia marina), no dehydrins have
been detected yet (Farrant et al. 1996; Kermode 1997). It
has been speculated, that the desiccation sensitivity of
the recalcitrant species is at least partially conditioned
by insufficient accumulation of dehydrins (Vertucci and
Farrant 1995; Panza et al. 2007).
In somatic embryos, desiccation has been associated
with improved germination and plantlet regeneration
(Attree et al. 1995; Bomal and Tremblay 1999), long-term
storage (Shiota et al. 1999; Bomal and Tremblay 2000) or
desiccation tolerance (Senaratna et al. 1990; Bomal et al.
2002). However, there are only few reports on activity of
dehydrins in somatic embryos (Bomal et al. 2002, Ko et al.
2006).
The distinctive feature of DHNs is a well conserved,
lysine-rich stretches of 15 amino acids, called the K
motifs that are predicted to form amphiphatic alfa-helixes
(Dure 1993; Close 1996, 1997). Other DHN domains
are the S- and Y-segments and less-conserved domains
rich in polar amino acids (U-segments). The S-segment
[(LHRSGS4-10(E/D)3] that usually precedes the K-seg-
ments, is a tract of serine residues. The consensus Y-seg-
ment (T/VDEYGNP), when present, is located in the
N-terminus (Close 1996; Campbell and Close 1997).
Besides those highly conserved domain, dehydrins share
the general tendency to be free of tryptophan and cysteine
residues, and they are rich in glycine residues (Dure 1993).
Permutations in the number and arrangement of conserved
domains assign the DHNs in higher plants to five sub-
classes YnSKn, SKn, Kn, YnKn, and KnS (Campbell and
Close 1997). Though their action is not fully understood,
they probably provide a cohesive water layer to a number
of macromolecules, preventing denaturing and coagulation
under extreme dehydration (Campbell and Close 1997) or
rescuing hydrolytic enzyme function under dry conditions
(Rinne et al. 1999).
In an effort a deeper insight into seed physiology of oak
seeds and identify genes possibly involved in their
responses to altered water status, this work was focused on
dehydrin genes from Q. robur seeds. Previously, the evi-
dence for the presence and expression of dehydrin(s) in oak
embryos has indirectly been shown by using heterologous
probes in RNA gel blot hybridisations (Finch-Savage et al.
1994; Sunderlıkova and Wilhelm 2002). Here, three
members of dehydrin family were isolated and some
aspects of their expression during zygotic and somatic
embryo development as well as upon different stresses
were studied.
Materials and methods
Plant material
Pedunculate oak (Q. robur L.) acorns were harvested in
weekly intervals from the beginning of July until the end
of September. Late cotyledonary stage zygotic embryos
(ZEs) 15–17 weeks post anthesis (WPA) were excised.
Oak zygotic embryos after 18 WPA were considered
mature. Embryos were frozen in liquid nitrogen and stored
at -80�C prior to analysis. Acorns collected after being
shed from trees on the grounds were germinated in sand in
the greenhouse. One-month-old seedlings were kept under
normal greenhouse conditions and used as a control group.
For cold treatment, seedlings were transferred to 4�C for
24 or 48 h, respectively. Desiccation stress was performed
by maintaining plants on dry filter paper for 7 or 24 h
under growth conditions in the greenhouse. The leaves
wounded by mechanical trimming with scissors were
collected after 12 or 24 h. Total RNA was extracted
(Chang et al. 1993) from the first two leaves or stems
(*5 mm under shoot apex) collected prior to and fol-
lowing each treatment.
The oak embryogenic culture line 6QR5 was maintained
by means of repetitive embryogenesis on P24 medium
(Teasdale 1992) supplemented with 3% (w/v) sucrose,
0.89 lM BA and solidified with 0.8% (w/v) agar as
described earlier (Endemann and Wilhelm 1999). Matura-
tion was induced by culturing translucent somatic embryos
(SEs) (size B 5 mm) on P24 medium supplemented with
6% (w/v) sorbitol for 5 weeks. To promote plant conver-
sion, SEs were subjected to partial desiccation treatment by
placing them into Petri dishes in the dark until they lost
approximately 25–30% of their moisture (Sunderlıkova and
Wilhelm 2002). SEs derived from each treatment were
frozen in liquid nitrogen and stored at -80�C until used.
ABA treatment
Cotyledonary somatic embryos (C3 mm) derived from the
proliferation medium were manually separated and cul-
tured in liquid P24 medium containing 50 or 100 lM ABA
for 2 days. Zygotic embryos were treated with 100 lM
ABA for 2 days as described previously (Sunderlıkova and
Wilhelm 2002).
1012 Plant Cell Rep (2009) 28:1011–1021
123

Isolation of dehydrin gene sequences
To isolate dehydrin genes (fragments) from Q. robur leaf
genomic DNA (Bekesiova et al. 1999), degenerate PCR
primers were designed positioned in the Y segment (V/T)
DEYGNP and the K segment MDKIKEKL (Close 1997)
[forward: 50-AC(C/T) GA(C/T) GA(A/G) TA(C/T) GG(A/
C/T) AAC C-30 and reverse: 50-AGC TTC TCC TT(A/G)
ATC TT(G/C) TCC-30, respectively]. The amplification
profile was: 12 min at 95�C, 40 cycles of 94�C/30 s, 47�C/
45 s, 72�C/1 min, and final extension at 72�C for 10 min.
The amplicons obtained were cloned into pCR 4-TOPO
vector (Invitrogen). The fragments were sequenced from
both sides with universal M13 primers using the BigDyeTM
Terminator Cycle Sequencing Ready Reaction Kit (PE
Applied Biosystem) in the ABI 3100 Genetic Analyzer
(Applied Biosystems). Database searches were carried out
using the BLAST program (Altschul et al. 1997) and
sequence comparison using CLUSTAL W software
(Thompson et al. 1994).
The fragments encoding for putative dehydrin genes
served as a basis for isolation of the corresponding full
length cDNAs. The first-strand cDNA synthesis was per-
formed with 2 lg of RNA sample from ABA-treated ZE
using random primers and SuperScript II reverse trans-
criptase (Life Technologies). The second strand was
amplified in PCR using degenerate primers for dehydrin
genes.
Screening for dehydrins in the local oak EST depository
The isolated oak dehydrin sequences were blasted locally
against the PICME (Platform for Integrated Clone
Management, Austrian Research Centers GmbH (ARC),
Seibersdorf, Austria) depository of oak ESTs comprising
44.178 clones from Q. robur and Q. petraea (cDNA, SSH,
and DiffSSH). The clones revealing sequence homology to
known dehydrin genes from other plant species (at signif-
icance e \ 10-10) were selected, put into the EST2Uni
pipeline (Forment et al. 2008), cleaned from vectors and
clustered in order to remove redundant sequences. For this
purpose, EST2Uni uses a two-step procedure, doing a
pairwise BLAST of all sequences to identify preliminary
clusters in a first step, and refining these clusters in a more
precise alignment step using CAP3. The resulting singleton
and contig sequences were used to predict protein trans-
lations using Translate tool (http://www.expasy.ch/tools/
dna.html) at the ExPASy Proteomic Server of the Swiss
Institute of Bioinformatics (SIB). Alignments of the entire
sequences as well as the guide tree were done using
CLUSTAL W software. Phylogenetic tree was drawn using
the TreeView 1.6.6 software (Page 1996).
RNA gel blot analyses
Aliquots (10 lg) of total RNA samples were separated on a
1.2% (w/v) agarose-formaldehyde gel and transferred to
the nylon membrane (Hybond N?, Amersham) by capillary
blotting according to the manufacturer’s instructions. The
membranes were fixed by UV-cross-linking followed by
baking at 80�C for 2 h. The RNA blot was stained with
methylene blue (Sambrook et al. 1989) to check the
amount of RNA loaded.
The blots were probed with cDNA fragments that were
radioactively labelled with [a-32P]-dCTP using the Ran-
dom Oligonucleotide Priming Kit (Stratagene). Hybrid-
ization and washing were carried out at 65�C in the manner
reported earlier (Sunderlıkova and Wilhelm 2002). The
membranes were exposed to Kodak X-Omat films
(Rochester, NY, USA) using an intensifying screen at
-80�C overnight.
In situ hybridization (ISH)
Plant tissues were fixed in 4% (w/v) paraformaldehyde and
0.25% (v/v) glutaraldehyde in PBS (phosphate buffer sal-
ine), embedded in Paraplast Plus (Sigma), sectioned and
mounted on poly-L-lysine coated slides. The RNA probes
were prepared by cloning the dehydrin PCR fragments into
pCR 4-TOPO vector (Invitrogen). The plasmids carrying
the QrDhn1 and QrDhn2 were linearised with Pst I and
Not I and in vitro transcribed using T7 and T3 RNA
polymerases in the presence of digoxigenin-UTP (Roche).
The in situ hybridization protocol followed the procedure
described earlier (Sunderlıkova et al. 2009). Sections were
observed with optical Axiovert 200 M microscope (Carl
Zeiss, Gottingen) and photographed by AxioCam system
(Carl Zeiss, Gottingen).
Results
Isolation of dehydrin sequences
Using degenerate primers, several fragments were obtained
on genomic DNA from oak tissue. Three of these revealed
significant sequence homology with dehydrins from other
plant sources. Full length cDNAs for QrDhn1 (Gene Bank
accession under the number AY607705) and QrDhn3
(AY607707) were obtained by 50and 30rapid amplification
of cDNA ends, for QrDhn2 (AY607706) we failed to
isolate the 50end. The putative dehydrin genes belong to
different groups (Fig. 1). The C-terminus of QrDhn1 gene
contains two well-conserved Lys-rich domains (K-seg-
ments) with the consensus motif EKKGIMDKIKEKLPG
(Fig. 1). It has been proposed that this motif could be
Plant Cell Rep (2009) 28:1011–1021 1013
123

implicated in hydrophobic interactions with partially
denatured proteins or membranes. In addition, the con-
served motif V/TDEYGNP (Y segment) is located at the
amino terminus of the QrDhn1. This motif has significant
amino acid sequence relatedness to a portion of the
nucleotide binding site of plant and bacterial chaperones
(Brini et al. 2007). Finally, the gene has the conserved
region with the track of serine residues (S segment)
involved in phosphorylation events, possibly related to the
binding of nuclear localization signal peptides and there-
fore to nuclear transport (Heyen et al. 2002). The presence
or absence of these motifs assigns the three oak dehydrin
genes to the types YK3S (QrDhn1), K3 (QrDhn3) and K4
(QrDhn2) (based on partial sequence only).
The genes encoding for dehydrins in oak form a
redundant family. Because of the presence of several
highly homologous motifs in the isolated gene sequences,
the mixture of QrDhn1-3 gene fragments was labeled and
used to probe the DNA gel blot. At least six dehydrin
family members could be identified in Q. robur genomic
DNA (Fig. 2a). The local search for oak dehydrin homo-
logues in the PICME database revealed four of them, while
an additional six putative Kn-type dehydrin genes (frag-
ments) were detected among Q. petraea clones (Table 1).
The relationship among the oak dehydrin sequences iden-
tified is depicted in the phylogenetic tree diagram (Fig. 2b).
The QrDhn2 and QrDhn3 are apparently closely related
with the Dhn3 dehydrin genes from Q. petraea but show a
more distant (evolutionary) relationship to the QrDhn1.
Expression of dehydrins in oak embryos
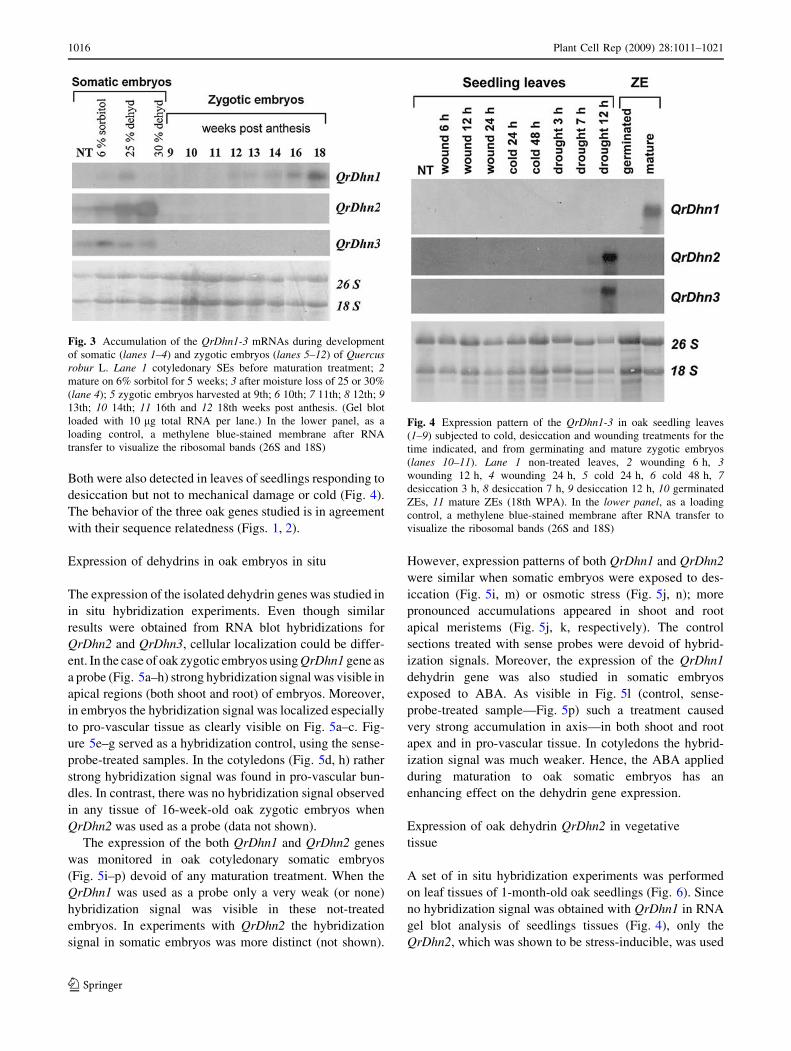
RNA gel blot analyses revealed that the QrDhn1 is expressed
in zygotic but not in untreated somatic embryos (Fig. 3) or
seedling leaves (Fig. 4). This gene is activated during later
stages of zygotic embryogenesis and responds to
Fig. 1 Multiple alignment of the predicted amino acid sequence of
the QrDhn 1-3 (GenBank accession numbers AY607705–AY607707,
respectively) with the deduced protein sequences of dehydrins from
Glycine max (Gm_AAB71225), Quercus robur (Qr_CAM98306,
Qr_CAM98307), Fagus sylvatica (Fs_CAE54590). Amino acids
which are conserved in all seven sequences aligned are marked by
asterisks. Chemically similar amino acids are marked by dots or
double dots. Dashes indicate where a sequence has been expanded to
allow optimal sequence alignment. Letters in the light shaded boxindicate the Y segment, in the dark-shaded and framed box the S
segment, and those in the mid-dark shaded box represent the K
segment. The sequences were aligned using Clustal W program
1014 Plant Cell Rep (2009) 28:1011–1021
123

dehydration as well as higher osmotic conditions (6% sor-
bitol) in somatic embryos. In contrast, the transcripts of the
two other dehydrins, QrDhn2 and QrDhn3, were detectable
in somatic but not zygotic embryos. The increased expres-
sion of QrDhn2–3 in treated somatic embryos show their
response to altered water and osmotic conditions (Fig. 3).
Table 1 Dehydrins identified in the clone collection of the PICME database
Name Singleton/contig Accession Source Length after cleaning
QrDhn1 AY607705_s AY607705 Q. robur 836
QrDhn2 AY607706_s AY607706 Q. robur 852
QrDhn3 CL2Contig1 AY607707,
MWG_721_C07a
Q. robur 739,
242
DN949901 CL1Contig1 DN949901 Q. robur 452
Qp_Dhn1 CR627795_s CR627795 Q. petraea 244
Qp_Dhn2 CL1Contig2 CR627714, CR627734,
CR627767
Q. petraea 251, 261,
259
Qp_Dhn3 CL1Contig3 CR627715, CR627736,
CR627753, CR627789,
CR627796, CR627905,
CR627909
Q. petraea 213, 247,
236, 183,
261, 268,
246
Qp_Dhn4 CL1Contig4 CR627784, CR627969,
CR627977, CR627997
Q. petraea 194, 299,
312, 290
Qp_Dhn5 CL1Contig5 CR627788,
LG0AAC25YI09RM1a,
LG0AAC31YA18RM1a
Q. petraea 309,
646,
600
Qp_Dhn6 LG0AAC14YN18RM1_s LG0AAC14YN18RM1a Q. petraea 669
Qp_AM711636 AM711636_s AM711636 Q. petraea 620
Qp_AM711635 AM711635_s AM711635 Q. petraea 656
a The clones present in the PICME depository only
Fig. 2 DNA gel blot analysis with the mixture of QrDhn1-3 cDNA
fragments (a). Genomic DNA from oak leaves was digested with BglII (lane 1), and HindIII (lane 2), separated on 0.8% agarose gel, and
blotted to a nylon membrane. The mixture of QrDhn1-3 cDNA
fragments was radioactively labeled and used as a probe in DNA gel
blot hybridisation under stringent conditions (658C). The kDNA
digested with HindIII was used as a molecular mass marker.
b Phylogram showing the sequence relatedness of the QrDhn1-3and DN949901 from Quercus robur with other dehydrins from
Quercus petraea (all GeneBank accessions viewed in the Table 1).
Distances are displayed according to a scale of ‘‘0.1’’, meaning 0.1
nucleotide substitutions per site. The actual value depends on the
branch lengths in the tree
Plant Cell Rep (2009) 28:1011–1021 1015
123
Both were also detected in leaves of seedlings responding to
desiccation but not to mechanical damage or cold (Fig. 4).
The behavior of the three oak genes studied is in agreement
with their sequence relatedness (Figs. 1, 2).
Expression of dehydrins in oak embryos in situ
The expression of the isolated dehydrin genes was studied in
in situ hybridization experiments. Even though similar
results were obtained from RNA blot hybridizations for
QrDhn2 and QrDhn3, cellular localization could be differ-
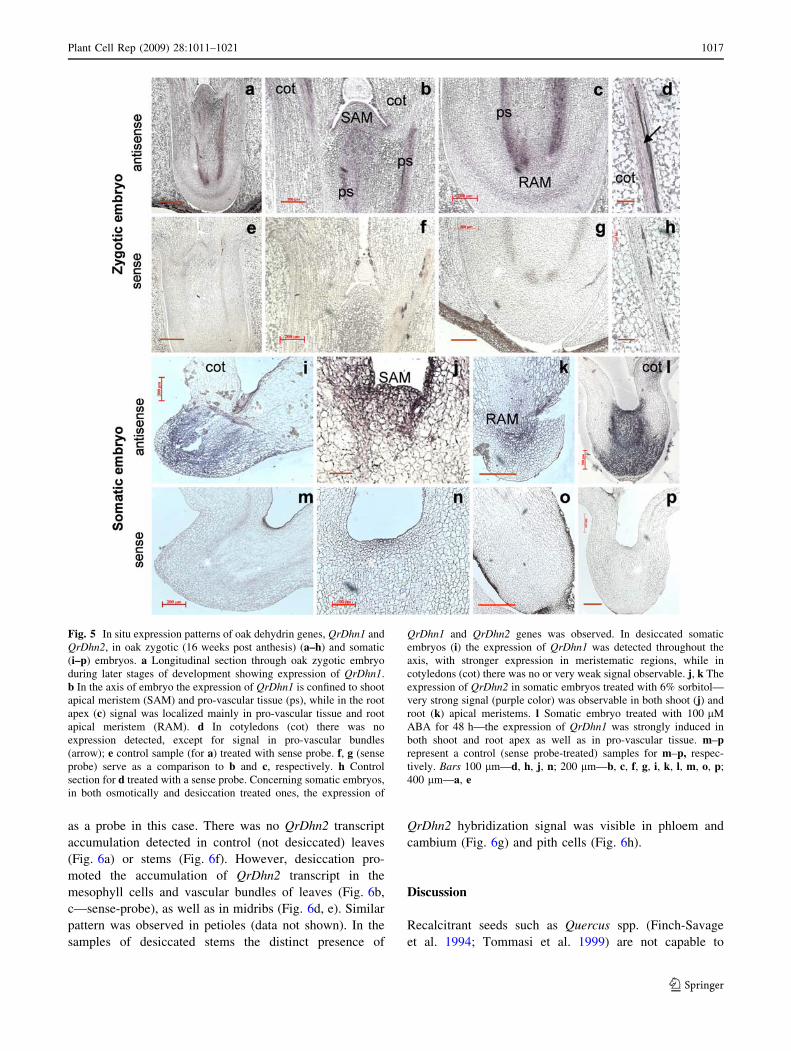
ent. In the case of oak zygotic embryos using QrDhn1 gene as
a probe (Fig. 5a–h) strong hybridization signal was visible in
apical regions (both shoot and root) of embryos. Moreover,
in embryos the hybridization signal was localized especially
to pro-vascular tissue as clearly visible on Fig. 5a–c. Fig-
ure 5e–g served as a hybridization control, using the sense-
probe-treated samples. In the cotyledons (Fig. 5d, h) rather
strong hybridization signal was found in pro-vascular bun-
dles. In contrast, there was no hybridization signal observed
in any tissue of 16-week-old oak zygotic embryos when
QrDhn2 was used as a probe (data not shown).
The expression of the both QrDhn1 and QrDhn2 genes
was monitored in oak cotyledonary somatic embryos
(Fig. 5i–p) devoid of any maturation treatment. When the
QrDhn1 was used as a probe only a very weak (or none)
hybridization signal was visible in these not-treated
embryos. In experiments with QrDhn2 the hybridization
signal in somatic embryos was more distinct (not shown).
However, expression patterns of both QrDhn1 and QrDhn2
were similar when somatic embryos were exposed to des-
iccation (Fig. 5i, m) or osmotic stress (Fig. 5j, n); more
pronounced accumulations appeared in shoot and root
apical meristems (Fig. 5j, k, respectively). The control
sections treated with sense probes were devoid of hybrid-
ization signals. Moreover, the expression of the QrDhn1
dehydrin gene was also studied in somatic embryos
exposed to ABA. As visible in Fig. 5l (control, sense-
probe-treated sample—Fig. 5p) such a treatment caused
very strong accumulation in axis—in both shoot and root
apex and in pro-vascular tissue. In cotyledons the hybrid-
ization signal was much weaker. Hence, the ABA applied
during maturation to oak somatic embryos has an
enhancing effect on the dehydrin gene expression.
Expression of oak dehydrin QrDhn2 in vegetative
tissue
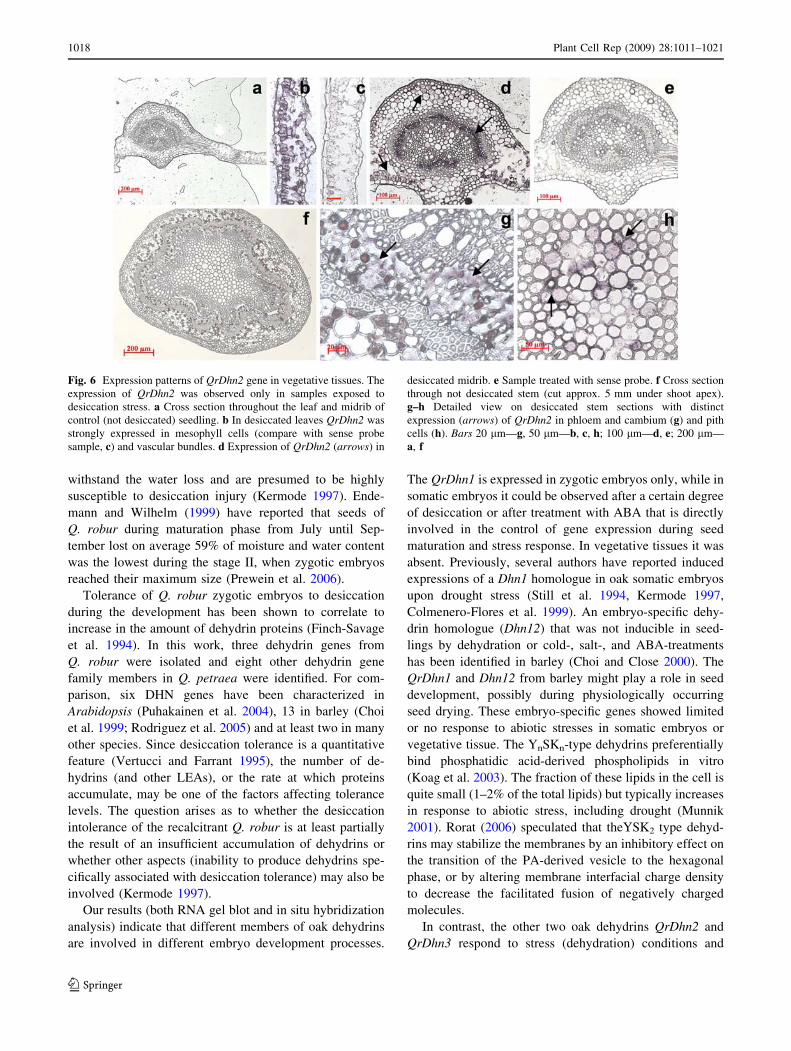
A set of in situ hybridization experiments was performed
on leaf tissues of 1-month-old oak seedlings (Fig. 6). Since
no hybridization signal was obtained with QrDhn1 in RNA
gel blot analysis of seedlings tissues (Fig. 4), only the
QrDhn2, which was shown to be stress-inducible, was used
Fig. 3 Accumulation of the QrDhn1-3 mRNAs during development
of somatic (lanes 1–4) and zygotic embryos (lanes 5–12) of Quercusrobur L. Lane 1 cotyledonary SEs before maturation treatment; 2mature on 6% sorbitol for 5 weeks; 3 after moisture loss of 25 or 30%
(lane 4); 5 zygotic embryos harvested at 9th; 6 10th; 7 11th; 8 12th; 913th; 10 14th; 11 16th and 12 18th weeks post anthesis. (Gel blot
loaded with 10 lg total RNA per lane.) In the lower panel, as a
loading control, a methylene blue-stained membrane after RNA
transfer to visualize the ribosomal bands (26S and 18S)
Fig. 4 Expression pattern of the QrDhn1-3 in oak seedling leaves
(1–9) subjected to cold, desiccation and wounding treatments for the
time indicated, and from germinating and mature zygotic embryos
(lanes 10–11). Lane 1 non-treated leaves, 2 wounding 6 h, 3wounding 12 h, 4 wounding 24 h, 5 cold 24 h, 6 cold 48 h, 7desiccation 3 h, 8 desiccation 7 h, 9 desiccation 12 h, 10 germinated
ZEs, 11 mature ZEs (18th WPA). In the lower panel, as a loading
control, a methylene blue-stained membrane after RNA transfer to
visualize the ribosomal bands (26S and 18S)
1016 Plant Cell Rep (2009) 28:1011–1021
123
as a probe in this case. There was no QrDhn2 transcript
accumulation detected in control (not desiccated) leaves
(Fig. 6a) or stems (Fig. 6f). However, desiccation pro-
moted the accumulation of QrDhn2 transcript in the
mesophyll cells and vascular bundles of leaves (Fig. 6b,
c—sense-probe), as well as in midribs (Fig. 6d, e). Similar
pattern was observed in petioles (data not shown). In the
samples of desiccated stems the distinct presence of
QrDhn2 hybridization signal was visible in phloem and
cambium (Fig. 6g) and pith cells (Fig. 6h).
Discussion
Recalcitrant seeds such as Quercus spp. (Finch-Savage
et al. 1994; Tommasi et al. 1999) are not capable to
Fig. 5 In situ expression patterns of oak dehydrin genes, QrDhn1 and
QrDhn2, in oak zygotic (16 weeks post anthesis) (a–h) and somatic
(i–p) embryos. a Longitudinal section through oak zygotic embryo
during later stages of development showing expression of QrDhn1.
b In the axis of embryo the expression of QrDhn1 is confined to shoot
apical meristem (SAM) and pro-vascular tissue (ps), while in the root
apex (c) signal was localized mainly in pro-vascular tissue and root
apical meristem (RAM). d In cotyledons (cot) there was no
expression detected, except for signal in pro-vascular bundles
(arrow); e control sample (for a) treated with sense probe. f, g (sense
probe) serve as a comparison to b and c, respectively. h Control
section for d treated with a sense probe. Concerning somatic embryos,
in both osmotically and desiccation treated ones, the expression of
QrDhn1 and QrDhn2 genes was observed. In desiccated somatic
embryos (i) the expression of QrDhn1 was detected throughout the
axis, with stronger expression in meristematic regions, while in
cotyledons (cot) there was no or very weak signal observable. j, k The
expression of QrDhn2 in somatic embryos treated with 6% sorbitol—
very strong signal (purple color) was observable in both shoot (j) and
root (k) apical meristems. l Somatic embryo treated with 100 lM
ABA for 48 h—the expression of QrDhn1 was strongly induced in
both shoot and root apex as well as in pro-vascular tissue. m–prepresent a control (sense probe-treated) samples for m–p, respec-
tively. Bars 100 lm—d, h, j, n; 200 lm—b, c, f, g, i, k, l, m, o, p;
400 lm—a, e
Plant Cell Rep (2009) 28:1011–1021 1017
123
withstand the water loss and are presumed to be highly
susceptible to desiccation injury (Kermode 1997). Ende-
mann and Wilhelm (1999) have reported that seeds of
Q. robur during maturation phase from July until Sep-
tember lost on average 59% of moisture and water content
was the lowest during the stage II, when zygotic embryos
reached their maximum size (Prewein et al. 2006).
Tolerance of Q. robur zygotic embryos to desiccation
during the development has been shown to correlate to
increase in the amount of dehydrin proteins (Finch-Savage
et al. 1994). In this work, three dehydrin genes from
Q. robur were isolated and eight other dehydrin gene
family members in Q. petraea were identified. For com-
parison, six DHN genes have been characterized in
Arabidopsis (Puhakainen et al. 2004), 13 in barley (Choi
et al. 1999; Rodriguez et al. 2005) and at least two in many
other species. Since desiccation tolerance is a quantitative
feature (Vertucci and Farrant 1995), the number of de-
hydrins (and other LEAs), or the rate at which proteins
accumulate, may be one of the factors affecting tolerance
levels. The question arises as to whether the desiccation
intolerance of the recalcitrant Q. robur is at least partially
the result of an insufficient accumulation of dehydrins or
whether other aspects (inability to produce dehydrins spe-
cifically associated with desiccation tolerance) may also be
involved (Kermode 1997).
Our results (both RNA gel blot and in situ hybridization
analysis) indicate that different members of oak dehydrins
are involved in different embryo development processes.
The QrDhn1 is expressed in zygotic embryos only, while in
somatic embryos it could be observed after a certain degree
of desiccation or after treatment with ABA that is directly
involved in the control of gene expression during seed
maturation and stress response. In vegetative tissues it was
absent. Previously, several authors have reported induced
expressions of a Dhn1 homologue in oak somatic embryos
upon drought stress (Still et al. 1994, Kermode 1997,
Colmenero-Flores et al. 1999). An embryo-specific dehy-
drin homologue (Dhn12) that was not inducible in seed-
lings by dehydration or cold-, salt-, and ABA-treatments
has been identified in barley (Choi and Close 2000). The
QrDhn1 and Dhn12 from barley might play a role in seed
development, possibly during physiologically occurring
seed drying. These embryo-specific genes showed limited
or no response to abiotic stresses in somatic embryos or
vegetative tissue. The YnSKn-type dehydrins preferentially
bind phosphatidic acid-derived phospholipids in vitro
(Koag et al. 2003). The fraction of these lipids in the cell is
quite small (1–2% of the total lipids) but typically increases
in response to abiotic stress, including drought (Munnik
2001). Rorat (2006) speculated that theYSK2 type dehyd-
rins may stabilize the membranes by an inhibitory effect on
the transition of the PA-derived vesicle to the hexagonal
phase, or by altering membrane interfacial charge density
to decrease the facilitated fusion of negatively charged
molecules.
In contrast, the other two oak dehydrins QrDhn2 and
QrDhn3 respond to stress (dehydration) conditions and
Fig. 6 Expression patterns of QrDhn2 gene in vegetative tissues. The
expression of QrDhn2 was observed only in samples exposed to
desiccation stress. a Cross section throughout the leaf and midrib of
control (not desiccated) seedling. b In desiccated leaves QrDhn2 was
strongly expressed in mesophyll cells (compare with sense probe
sample, c) and vascular bundles. d Expression of QrDhn2 (arrows) in
desiccated midrib. e Sample treated with sense probe. f Cross section
through not desiccated stem (cut approx. 5 mm under shoot apex).
g–h Detailed view on desiccated stem sections with distinct
expression (arrows) of QrDhn2 in phloem and cambium (g) and pith
cells (h). Bars 20 lm—g, 50 lm—b, c, h; 100 lm—d, e; 200 lm—
a, f
1018 Plant Cell Rep (2009) 28:1011–1021
123

appear exclusively in somatic but not in zygotic embryos of
developmental stages studied (9–18 weeks post anthesis).
The oak somatic embryos that were subjected to the
progressive dehydration treatment expressed gradually
increasing accumulations of these dehydrin transcripts that
correlates with increasing conversion rates of embryos. It
has also been found that somatic embryos that lost 55%
of their initial fresh weight were still able to recover
(Sunderlıkova 2002). The positive effect of partial drying
on SE germination has already been observed in many
species including oak (Sunderlıkova and Wilhelm 2002).
This suggests that accumulation of dehydrins during slow
drying might enhance desiccation tolerance and the via-
bility of oak somatic embryos. Moreover, the QrDhn2–3
dehydrins accumulated also in vegetative tissue under
desiccation stress. However, unlike other dehydrins
described previously (Close 1996), they do not respond to
other stimuli, for instance neither cold nor wounding. By
contrast to the YnSKn DHNs, there is no direct in vitro
evidence for the function of Kn-type dehydrins. In non-
stressed plants, these dehydrins are mainly localized on the
vascular tissue of transporting organs (roots, stems), the
vascular tissues of leaves, the roots tips and the apical parts
of the roots (Rorat et al. 2006). Their accumulation in
response to low temperature has been previously demon-
strated and shown to correlate with capacity of the plants to
develop freezing tolerance (Rorat 2006).
In our in situ hybridization experiments, dehydrin’s
transcripts were mostly localized in shoot and root apical
meristems and pro-vascular strands. These data are in
accordance with the findings of Carjuzaa et al. (2008) and
Kalemba et al. (2009) who detected the presence of
dehydrins in all axis and cotyledon tissues. However, the
cotyledon tissues of Q. robur embryos (both zygotic and
somatic) failed in the presence of dehydrin transcripts—
except for pro-vascular tissue. Buitink et al. (2003) have
reported that cotyledons were not essential for re-estab-
lishment of desiccation tolerance in germinating Medicago
truncatula seeds. Such localization of dehydrins in oak
embryos indicated their participation in vascular develop-
ment as one of the ontogenetic processes in the apical
meristems before germination of embryo. These processes
are quite likely water demanding, e.g. for rehydration of
meristems, cell expansion and metabolic pathway recovery
(Yakovlev et al. 2008), and therefore necessary for active
growth of both zygotic and somatic oak embryos.
According to Rorat (2006), plants subjected to abiotic
stress such as drought, low temperature or any other kind of
stress leading to cellular dehydratation, show not only an
increase in the content of dehydrins already present in the
tissue but often also the synthesis of new ones. Several in
situ localization studies (mostly immunolocalization ones)
have previously found dehydrins to be localized primarily
in the cytoplasm and nucleus (Ashgar et al. 1994, Brini
et al. 2007, Carjuzaa et al. 2008), in the endoplasmic
reticulum (Carjuzaa et al. 2008), protein bodies (Rinne
et al. 1999), vacuoles (Heyen et al. 2002), and other cell
compartments. On other hand, in the mature seeds of
Euterpe edulis the dehydrins have been found in the
cytoplasm and associated with chromatin, but not with
organelles or membranes (Panza et al. 2007). Nevertheless,
there is still a rather small number of reports available
dealing with in situ localization of dehydrins in plant tis-
sues (Swensson 2001, Rorat 2006, Panza et al. 2007,
Carjuzaa et al. 2008). As has been stated by Carjuzaa et al.
(2008), it is probably due to technical difficulties of pro-
cessing seed tissues that are full of reserve material and are
in a dehydrated state.
In addition to the Dhn1-3 genes isolated in this study,
the dehydrin gene family in Q. robur comprises at least
three other members, while in Q. petraea eight putative Kn
type dehydrins were identified in a local sequence depos-
itory. In addition, our analysis did not encompass the SKn
type dehydrins (if any exiting in oak). The biological sig-
nificance of the multigene families is hypothesized to be
the ability to independently and flexibly regulate the pro-
duction of different gene products in the same or different
cells (Dixon et al. 2002). Alternatively, multigene families
could ensure rapid and massive accumulation of certain
gene products at the time and position of urgent need, e.g.
during infection, abiotic stresses, etc. (Dixon et al. 2002). It
must be anticipated, however, that our analysis was based
solely on sequence similarity that could be less than would
result in physical detection on mid- to high-stringency
DNA gel blot analyses. Therefore, these analyses should
not be taken to imply proven function since some of the
genes identified and annotated as dehydrins still may in fact
encode related enzymes with different functions. Never-
theless, sequence- (EST) based approaches can reveal
potential new gene family members that can be further
tested for molecular function and a possible role in certain
physiological processes.
In summary, the expression patterns of the three oak
dehydrin studied confirm the conclusions of Carjuzaa et al.
(2008) that in orthodox seeds (1) different dehydrins can be
distributed in different tissues during plant growth and
development, (2) some dehydrins are specific to tissue or
cell types, (3) different types of dehydrins can localize to
the same tissue. In addition, dehydrins might play a more
specific role in different processes such as embryo devel-
opment or response to environmental stress (Choi and
Close 2000). The role(s) of dehydrins in development of
species with recalcitrant seeds, however, remains obscured.
Slowly dried recalcitrant seeds (unlike the orthodox ones)
generally die at water contents far above the range at which
few, if any, of the mechanisms of protection suggested for
Plant Cell Rep (2009) 28:1011–1021 1019
123

dehydrins would be operative (Berjak and Pammenter
2008). In addition, the occurrence of dehydrins in recalci-
trant seeds is equivocal, as they have been found to occur
in a range of species from different habitats, while appar-
ently being absent from others (Farrant et al. 1996).
However, their presence in some recalcitrant species,
including Quercus species (Turco et al. 2004, this work)
and wild rice, suggests that dehydrins are necessary, but
not sufficient, to achieve desiccation tolerance in seeds
(Blackman et al. 1991, Bradford and Chandler 1992).
Additional features such as water replacement, tissue dif-
ferentiation, lipid, protein, and sugar accumulation, mem-
brane phase transition and glassy state formation are also
involved in anhydrobiosis (Panza et al. 2007). The pres-
ence and activity of dehydrins in recalcitrant embryos of
particular species, however, apparently facilitates more
effective survival of (recalcitrant) seeds to lowered water
contents.
Acknowledgments This work was supported from FTSP (Austria)
and EEA Financial Mechanism ‘‘SAV-EHP-2008-02-01’’.
References
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W,
Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new
generation of protein database search programs. Nucl Acids Res
25:3389–3402
Ashgar R, Fenton RD, DeMason DA, Close TJ (1994) Nuclear and
cytoplasmic localization of maize embryo and aleurone dehy-
drin. Protoplasma 177:87–94
Attree SM, Pomeroy MK, Fowke LC (1995) Development of white
spruce (Picea glauca (Moench) Voss) somatic embryos during
culture with abscisic acid and osmoticum, and their tolerance to
drying and frozen stage. J Exp Bot 46:433–439
Barbedo CJ, Bilia DAC (1998) Evolution of research on recalcitrant
seeds. Sci Agric 55:121–125
Bekesiova I, Nap JP, Mlynarova L (1999) Isolation of high quality
DNA and RNA from leaves of the carnivorous plant Droserarotundifolia. Plant Mol Biol Rep 17:269–277
Berjak P, Pammenter NW (2008) From Avicennia to Zizania: seed
recalcitrance in perspective. Ann Bot 101:213–228
Blackman SA, Wettlaufer SH, Obendorf RL, Leopold AC (1991)
Maturation proteins associated with desiccation tolerance in
soybean. Plant Physiol 96:868–874
Bomal C, Tremblay FM (1999) Effect of desiccation to low moisture
content on germination, synchronization of root emergence, and
plantlet regeneration of black spruce somatic embryos. Plant Cell
Tiss Org Cult 56:193–200
Bomal C, Tremblay FM (2000) Dried cryopreserved somatic embryos
of two Picea species provide suitable material for direct plantlet
regeneration and germplasm storage. Ann Bot 72:177–183
Bomal C, Van Quy Le, Tremblay FM (2002) Induction of tolerance to
fast desiccation in black spruce (Picea mariana) somatic
embryos: relationship between partial water loss, sugars, and
dehydrins. Physiol Plant 115:523–530
Bradford KJ, Chandler PM (1992) Expression of ‘‘dehydrin-like’’
proteins in embryos and seedlings of Zizania palustris and Oryzasativa during dehydration. Plant Physiol 99:488–494
Brini F, Hanin M, Lumbreras V, Irar S, Pages M, Masmoudi K (2007)
Functional characterization of DHN-5, a dehydrin showing a
differential phosphorylation pattern in two Tunisian durum
wheat (Triticum durum Desf.) varieties with marked differences
in salt and drought tolerace. Plant Sci 172:20–28
Buitink J, Benoit LV, Satour P, Leprince O (2003) The re-
establishment of desiccation tolerance in germinateg radicles
of Medicago truncatula Gaertn. seeds. Seed Sci Res 13:273–286
Campbell SA, Close TJ (1997) Dehydrins: genes, proteins and
associations with phenotypic traits. New Phytol 137:61–74
Carjuzaa P, Castellion M, Distefano AJ, des Vas M, Maldonado S
(2008) Detection and subcellular localization of dehydrin-like
proteins in quinoa (Chenopodium quinoa Willd) embryos.
Protoplasma 233:149–156
Ceccardi TL, Meyer NC, Close TJ (1994) Purification of a maize
dehydrin. Protein Expr Purif 5:266–269
Chang S, Puryear J, Cairney J (1993) A simply and efficient method
for isolating RNA from pine trees. Plant Mol Biol Rep 11:113–
116
Choi DW, Close TJ (2000) A newly identified barley gene, Dhn12encoding a YSK2 DHN, is located on chromosome 6H and has
embryo-specific expression. Theor Appl Genet 100:1274–1278
Choi DW, Zhu B, Close TJ (1999) The barley (Hordeum vulgare L.)
dehydrin multigene family: sequences, allele types, chromosome
assignments, and expression characteristics of 11 Dhn genes of
cv. Dicktoo. Theor Appl Genet 98:1234–1247
Close TJ (1996) Dehydrins: emergence of a biochemical role of
family of plant dehydration proteins. Physiol Plant 97:795–803
Close TJ (1997) Dehydrins: a commonality in the response of plants
to dehydration and low temperature. Physiol Plant 100:291–296
Colmenero-Flores JM, Moreno LP, Smith CE, Covarrubias AA
(1999) Pvlea-18, a member of a new late-embryogenesis-
abundant protein family that accumulates during water stress
and in the growing regions of well irrigated bean seedlings. Plant
Physiol 120:93–103
Dixon RA, Achnine L, Kota P, Liu Ch-J, Reddy M, Srinivasa S,
Wang L (2002) The phenylpropanoid pathway and plant
defence—a genomics perspective. Molec Plant Pathol 3:371–
390
Dure LS III (1993) The lea proteins of higher plants. In: Verma DPS
(ed) Control of Plant Gene Expression. CRC Press, Boca Raton,
pp 325–335
Endemann M, Wilhelm E (1999) Factors influencing the induction
and viability of somatic embryos of Quercus robur L. Biol Plant
42:499–504
Farrant JM, Pammenter NW, Berjak P, Farnsworth EJ, Vertucci CW
(1996) Presence of dehydrin-like proteins and levels of abscisic
acid in recalcitrant (dessication sensitive) seeds may be related to
habitat. Seed Sci Res 6:175–182
Finch-Savage WE, Blake PS (1994) Indeterminate development in
desiccation-sensitive seeds of Quercus robur L. Seed Sci Res
4:127–133
Finch-Savage WE, Pramanik SK, Bewley JD (1994) The expression
of dehydrin proteins in desiccation-sensitive (recalcitrant) seeds
of temperate trees. Planta 193:478–485
Forment J, Gilabert F, Robles A, Conejero V, Nuez F, Blanca JM
(2008) EST2uni: an open, parallel tool for automated EST
analysis and database creation, with a data mining web interface
and microarray expression data integration. BMC Bioinformatics
9:5
Heyen BJ, Alsheikh MK, Smith EA, Torvik CF, Seals DF, Randal SK
(2002) The calcium-binding activity of a vacuole-associated,
dehydrin-like protein is regulated by phosphorylation. Plant
Physiol 130:675–687
Kalemba EM, Janowiak F, Pukacka S (2009) Desiccation tolerance
acquisition in developing beech (Fagus sylvatica L.) seeds: the
1020 Plant Cell Rep (2009) 28:1011–1021
123

contribution of dehydrin-like protein. Trees doi: 10.1007/
s00468-008-0278-8
Kermode AR (1997) Approaches to elucidate the basis of desiccation-
tolerance in seeds. Seed Sci Res 7:75–95
Ko S, Tan SK, Kamada H (2006) Characterization of a dehydrin-like
phosphoprotein (ECPP44) relating to somatic embryogenesis sin
carrot. Plant Physiol Biochem 24:253a–253j
Koag M-C, Fenton RD, Wilkens S, Close TJ (2003) The binding of
maize DHN1 to lipid vesicles. Gain of structure and lipid
specificity. Plant Physiol 131:309–316
Munnik T (2001) Phosphatidic acid: an emerging plant lipid second
messenger. Trends Plant Sci 6:227–233
Page RDM (1996) TREEVIEW: an application to display phyloge-
netic trees on personal computers. CABIOS 12:357–358
Panza V, Distefano AJ, Carjuzaa P, Lainez V, del Vas M, Maldonado
S (2007) Detection of dehydrin-like proteins in embryos and
endosperm of mature Euterpe edulis seeds. Protoplasma 231:1–5
Prewein Ch, Endemann M, Reihold W, Salaj J, Sunderlıkova V,
Wilhelm E (2006) Physiological and morphological character-
istics during development of pedunculate oak (Quercus robur L.)
zygotic embryos. Trees 20:53–60
Puhakainen T, Hess MV, Makela P, Svenson J, Heino P, Palva ET
(2004) Overexpression of multiple dehydrin genes enhances
tolerance to freezing stress in Arabidopsis. Plant Mol Biol
54:743–753
Rinne PLH, Kaikuranta PLM, van der Plas LHW, van der Schoot C
(1999) Dehydrins in cold-acclimated apices of birch (Betulapubescens Ehrh.): production, localization and a potential role in
rescuing enzyme function during dehydration. Planta 209:377–
388
Roberts EH (1973) Predicting the storage life of seeds. Seed Sci
Technol 1:499–514
Rodriguez EM, Svensson JT, Malatrasi D, Choi W, Close TJ (2005)
Barley Dhn13 encodes a KS-type dehydrin with constitutive and
stress responsive expression. Theor Appl Gen 110:852–858
Rorat T (2006) Plant dehydrins—tissue location, structure and
function. Cell Mol Biol Lett 11:536–556
Rorat T, Szabala BM, Grygorowicz WJ, Wojtowicz B, Yin Z, Rey P
(2006) Expression of SK3-type dehydrin in transporting organs
is associated with cold acclimation in Solanum species. Planta
224:205–221
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: a
laboratory manual, 2nd edn. Cold Spring Harbour Laboratory
Press, Cold Spring Harbour
Senaratna T, McKersie BD, Bowley SR (1990) Artificial seeds of
alfalfa (Medicaco sativa L.). Induction of desiccation tolerance
in somatic embryos. In Vitro Cell Dev Biol-Plant 26:85–90
Shiota H, Tachibana K, Watabe K, Kamada H (1999) Successful
long-term preservation of abscisic-acid-treated and desiccated
carrot somatic embryos. Plant Cell Rep 18:749–753
Still DW, Kovach DA, Bradford KJ (1994) Development of
desiccation tolerance in rice (Oryza sativa) and wild rice
(Zizania palustris). Dehydrin expression, abscisic acid content,
and sucrose accumulation. Plant Physiol 104:431–438
Sunderlıkova V (2002) Molecular aspects of oak somatic embryo-
genesis. PhD Thesis, Nitra 2002, p 113
Sunderlıkova V, Wilhelm E (2002) High accumulation of legumin
and Lea-like mRNAs during maturation is associated with
increased conversion frequency of somatic embryos from
pedunculate oak (Quercus robur L.). Protoplasma 220:97–103
Sunderlıkova V, Salaj J, Matusıkova I, Wilhelm E (2009) Isolation
and characterization of an embryo-specific Em-like gene of
pedunculate oak (Quercus robur L.) and its temporal and spatial
expression patterns during somatic and zygotic embryo devel-
opment. Trees 23:135–144
Swensson J. (2001) Functional studies of the role of plant dehydrins
in tolerance to salinity, desiccation and low temperature.
Doctoral dissertation. Department of Plant Biology, SLU. Acta
Universitatis agriculturae Sueciae, Agraria, vol. 259, pp 47
Teasdale R (1992) Formulation of plant culture media and application
therefore. International publication No. WO 92/07460. Patent
No. Europe: 92902531.0. Forbio PTY Ltd, Queensland
Thompson JD, Higgins DG, Gibson TJ (1994) Clustal W: improving
the sensitivity of progressive multiple sequence alignment
through sequence weighting, position-specific gap penalties
and weight matrix choice. Nucl Acids Res 22:4673–4680
Tommasi F, Paciolla C, Arrigoni O (1999) The ascorbate system in
recalcitrant and orthodox seeds. Physiol Plant 105:193–198
Turco E, Close TJ, Fenton RD, Ragazzi A (2004) Synthesis of
dehydrin-like proteins in Quercus ilex L. and Quercus cerris L.
seedling subjected to water stress and infection with Phytoph-thora cinnamomi. Physiol Mol Plant Pathol 65:137–144
Vertucci CW, Farrant JM (1995) Acquisition and loss of desiccation
tolerance. In: Kigel J, Galili G (eds) Seed development and
germination. Marcel Dekker, New York, pp 237–271
Yakovlev IA, Asante DKA, Fossdal CG, Partanen J, Junttila O,
Johnsen Ø (2008) Dehydrins expression related to timing of bud
burst in Norway spruce. Planta 228:459–472
Plant Cell Rep (2009) 28:1011–1021 1021
123





























