Embed Size (px)
Citation preview

www.elsevier.com/locate/pain
Pain 130 (2007) 137–143
Descending analgesia – When the spine echoes what the brain expects
Philippe Goffaux a, William John Redmond a, Pierre Rainville b, Serge Marchand a,*
a Universite de Sherbrooke, Faculte de Medecine, Sherbrooke, Que., Canada J1H 5N4b Universite de Montreal, Faculte de Medecine Dentaire, Montreal, Que., Canada H3C 3J7
Received 21 June 2006; received in revised form 12 November 2006; accepted 21 November 2006
Abstract
Changes in pain produced by psychological factors (e.g., placebo analgesia) are thought to result from the activity of specificcortical regions. However, subcortical nuclei, including the periaqueductal gray and the rostroventral medulla, also show selectiveactivation when subjects expect pain relief. These brainstem regions send inhibitory projections to the spine and produce diffuseanalgesic responses. Regrettably the precise contribution of spinal mechanisms in predicting the strength of placebo analgesia isunknown. Here, we show that expectations regarding pain radically change the strength of spinal nociceptive responses in humans.We found that contrary to expectations of analgesia, expectations of hyperalgesia completely blocked the analgesic effects ofdescending inhibition on spinal nociceptive reflexes. Somatosensory-evoked brain potentials and pain ratings further confirmedchanges in spino-thalamo-cortical responses consistent with expectations and with changes in the spinal response. These findingsprovide direct evidence that the modulation of pain by expectations is mediated by endogenous pain modulatory systems affectingnociceptive signal processing at the earliest stage of the central nervous system. Expectation effects, therefore, depend as much aboutwhat takes place in the spine as they do about what takes place in the brain. Furthermore, complete suppression of the analgesicresponse normally produced by descending inhibition suggests that anti-analgesic expectations can block the efficacy of pharmaco-logically valid treatments which has important implications for clinical practice.� 2006 International Association for the Study of Pain. Published by Elsevier B.V. All rights reserved.
Keywords: Expectations; Withdrawal reflex; RIII; Spine; SEP; Brain; DNIC
1. Introduction
A number of studies have shown that pain relief islargely dependent upon what we expect regarding theeffect of a given treatment (Montgomery and Kirsch,1997; Price et al., 1999; Charron et al., 2006). Neuro-imaging studies that have focused on expectancy-medi-ated analgesia reveal that the dorsolateral, orbitofrontaland medial prefrontal cortex may be involved in trigger-ing this form of analgesia (Ploghaus et al., 1999; Wageret al., 2004; Rainville and Duncan, 2006). Similar studiesare also beginning to show that prefrontal and midbrainregions are co-activated during expectations of analgesia
0304-3959/$32.00 � 2006 International Association for the Study of Pain. P
doi:10.1016/j.pain.2006.11.011
* Corresponding author. Tel.: +1 819 346 1110x15889; fax: +1 819564 5424.
E-mail address: [email protected] (S. Marchand).
(Petrovic et al., 2002; Wager et al., 2004). This finding isimportant because several midbrain nuclei are involvedin descending inhibitory control. The activation ofdescending inhibitory influences by expectations of anal-gesia may thereby reduce the intensity of pain signals atthe earliest stages of nociceptive processing in the spinalcord. The putative effects of mesencephalic descendingpathways have never been tested in a paradigm thatmanipulates expectation. Our goal in the present studywas to investigate the effects of expectation on thestrength of descending inhibition and spinal painprocessing.
Endogenous pain-modulating systems includedescending inhibitory projections that are coordinatedin the rostroventral medulla and make up part of thespinal-bulbo-spinal diffuse noxious inhibitory controlpathway or DNIC (Le Bars et al., 1979; Fields and
ublished by Elsevier B.V. All rights reserved.

138 P. Goffaux et al. / Pain 130 (2007) 137–143
Basbaum, 1999; Millan, 2002). DNIC activity is typical-ly triggered using a counterirritation technique (applica-tion of a second painful stimulus). Early studies showedthat it was possible to quantify the effects of DNIC byrecording changes in electrophysiological activity andsubjective pain ratings (Willer et al., 1989, 1990). In thisway, electromyographic (EMG) recordings were used toshow that counterirritation procedures reduce theamplitude of a spinal nociceptive flexion reflex (RIII)elicited at the bicep femoris by a painful transcutaneouselectrical stimulation of the sural nerve (Willer et al.,1990; Dowman, 2002; Sandrini et al., 2005). Counterir-ritation procedures also reduce the amplitude ofsomatosensory-evoked brain potentials (SEPs) triggeredby painful sural nerve stimulations (Dowman, 2002).
If, expectations affect descending inhibitory activity,then differences in expectation should change the analge-sic potential of counterirritation. To explore this possi-bility, we activated the DNIC system with a standardcounterirritation procedure (immersion of the arm incold water). In a first group, subjects expected this pro-cedure to provoke analgesic effects while in a secondgroup subjects expected this procedure to provokehyperalgesic (pain-enhancing) effects. We hypothesizedthat if expectations modulate DNIC activity, then thisshould be manifest as a change in the amplitude of thespinal RIII-reflex, the SEP and the pain evoked by elec-trical stimulation of the sural nerve. If, however, expec-tancy effects are purely a cortical epiphenomenon thatfails to interact with descending inhibitory systems, thenonly cortical responses (SEPs and subjective ratings)should change as a function of expectation. In this casecounterirritation should trigger a full strength DNICresponse regardless of what is expected. As a conse-quence, spinal reflexes should decrease in amplitude inspite of the analgesia or hyperalgesia that may beexpected or subjectively experienced.
2. Methods
2.1. Subjects
Twenty healthy volunteers were randomly assigned to oneof the two experimental groups. Ten were assigned to the anal-gesia expectancy group and 10 were assigned to the hyperalge-sia expectancy group. Participants in the analgesia group were22.9 (±2.0) years of age and participants in the hyperalgesiagroup were 21.9 (±3.1) years of age. For each group, an equalnumber of men and women were tested. Participants wereexcluded if they were under any medication. All participantssigned a consent form approved by the local Ethics Committee.
2.2. Neurophysiological measures
Withdrawal reflex activity was evoked by stimulating theleft sural nerve over its retromalleolar path. Stimulationsconsisted of a volley of five electrical pulses (each 1 ms long)
administered at a rate of 240 Hz using a constant currentstimulator. A stimulation volley lasted only 21 ms. Stimula-tions produced a withdrawal reflex that was measured viaan electromyographic (EMG) recording of the knee flexormuscle. Stimulations were set at 15% above the thresholdvalue required to provoke the reflex. Threshold values weredetermined using an iterative staircase method. EMG activitywas considered reflexive when its amplitude exceeded baselineactivity levels by at least 1.5 standard deviations. Withdrawalreflex activity was quantified by calculating the integral ofthe rectified EMG signal between 90 and 150 ms post-stimulation.
Cortical activity was also recorded and time-locked to theonset of all sural nerve stimulations. This allowed us to obtainevent-related waveforms. Waveforms were obtained at the ver-tex (Cz) using a monopolar montage with a right ear-reference.Using a Powerlab system, our signal was sampled at 200 Hzand filtered online within a 0.5 to 30 Hz bandpass. Electrocu-lographic (EOG) activity was recorded bipolarly from elec-trodes placed at the outer canthi of both eyes (horizontalEOG) and above and below the left eye (vertical EOG). Trialscontaminated with EOG artifacts (±60 lV) were eliminatedfrom the analyses. Cortical activity was baseline correctedbetween 0 and 100 ms prior to stimulation onset and averagedtogether. From the mean waveforms specific epochs were iso-lated in order to capture both early (P45, N100) and late(N150, P260) somatosensory components. The P45 deviationis known to reflect primary somatosensory cortex activityand is insensitive to stimulus intensity or magnitude ratings.N100, N150 and P260 deviations, on the other hand, increasewith increasing pain intensity. The N100 component has a cen-tro-temporal maximum and is characterized by sources locatednear the secondary somatosensory cortex whereas the N150component is most negative near the vertex and includes gen-erators located in the insular and anterior cingulate cortices.The P260 is also maximal at vertex and is typically associatedwith sources located near the caudal sector of the supracallosalACC. A number of different localization studies support thesefindings (Tarkka and Treede, 1993; Valeriani et al., 1996; Lenz,1998) and confirm that the neuroelectric activity of specificbrain regions can be reliably measured from surface scalppotentials.
P45 activity was examined in the 45–55 ms post-stimulusinterval whereas N100 activity was examined in the90–120 ms interval. N150 activity was examined in the135–150 ms interval and P260 activity was examined inthe 280–350 ms interval. These intervals were selected becausethey correspond to a series of consecutive time points duringwhich the scalp topographic pattern is stable and during whichdistinct source configurations are present (Dowman, 1994a,b;Dowman and Darcey, 1994). Although the epoch of ourP260 component spans a timeframe that includes the fallingarm of the P260, we will continue to refer to this componentas a P260 potential. It is important to point out that peak-scor-ing of the P260 is not advised when using electrical stimula-tions, since, during this time (and slightly earlier), there is aspatial and temporal overlap of the scalp electric potentialsgenerated by both innocuous and pain-related sources(Dowman and Schell, 1999). In contrast, the 280–350 ms inter-val of this late potential has been shown to reflect pain-relatedactivity more specifically (Dowman, 1994a,b).
Ch
ang
e in
Pai
n In
ten
sity
(%
rel
ativ
e to
bas
elin
e)
-100
-50
0
50
ExpectedExperienced
Per
ceiv
ed P
ain
Inte
nsi
ty
(% r
elat
ive
to b
asel
ine)
Expected Pain Intensity (% relative to baseline)
Hyperalgesia
Analgesia
200
100
00 100
a
b
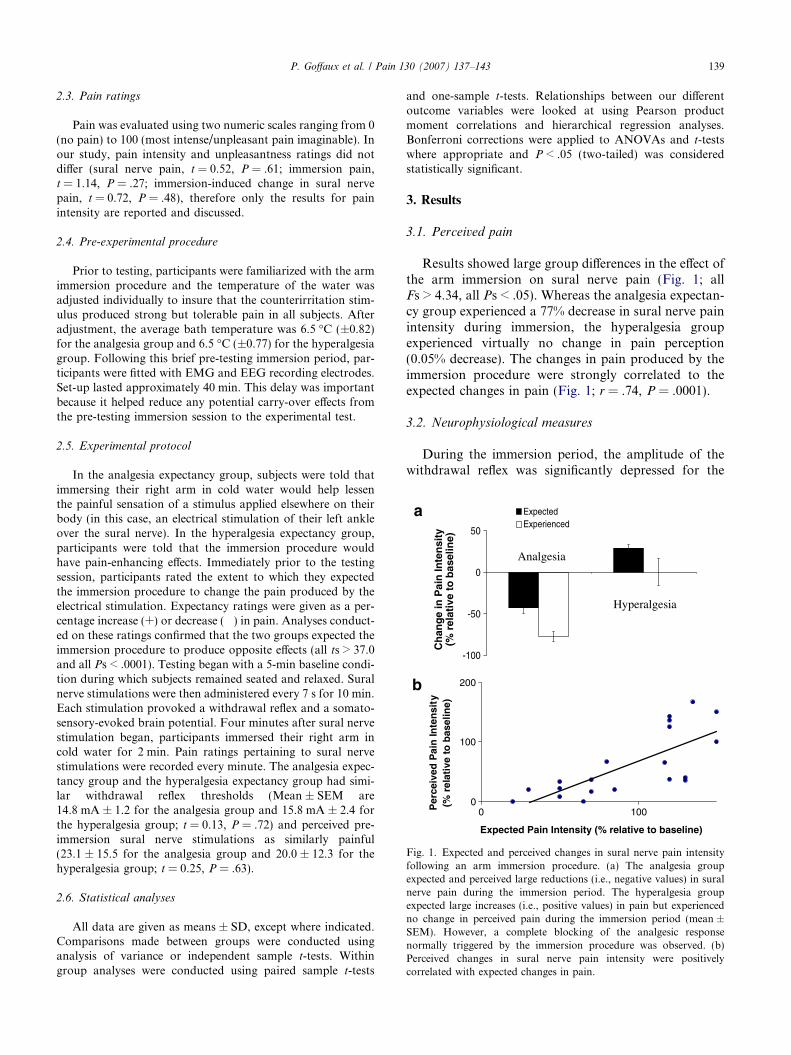
ig. 1. Expected and perceived changes in sural nerve pain intensityollowing an arm immersion procedure. (a) The analgesia groupxpected and perceived large reductions (i.e., negative values) in suralerve pain during the immersion period. The hyperalgesia groupxpected large increases (i.e., positive values) in pain but experiencedo change in perceived pain during the immersion period (mean ±EM). However, a complete blocking of the analgesic responseormally triggered by the immersion procedure was observed. (b)erceived changes in sural nerve pain intensity were positivelyorrelated with expected changes in pain.
P. Goffaux et al. / Pain 130 (2007) 137–143 139
2.3. Pain ratings
Pain was evaluated using two numeric scales ranging from 0(no pain) to 100 (most intense/unpleasant pain imaginable). Inour study, pain intensity and unpleasantness ratings did notdiffer (sural nerve pain, t = 0.52, P = .61; immersion pain,t = 1.14, P = .27; immersion-induced change in sural nervepain, t = 0.72, P = .48), therefore only the results for painintensity are reported and discussed.
2.4. Pre-experimental procedure
Prior to testing, participants were familiarized with the armimmersion procedure and the temperature of the water wasadjusted individually to insure that the counterirritation stim-ulus produced strong but tolerable pain in all subjects. Afteradjustment, the average bath temperature was 6.5 �C (±0.82)for the analgesia group and 6.5 �C (±0.77) for the hyperalgesiagroup. Following this brief pre-testing immersion period, par-ticipants were fitted with EMG and EEG recording electrodes.Set-up lasted approximately 40 min. This delay was importantbecause it helped reduce any potential carry-over effects fromthe pre-testing immersion session to the experimental test.
2.5. Experimental protocol
In the analgesia expectancy group, subjects were told thatimmersing their right arm in cold water would help lessenthe painful sensation of a stimulus applied elsewhere on theirbody (in this case, an electrical stimulation of their left ankleover the sural nerve). In the hyperalgesia expectancy group,participants were told that the immersion procedure wouldhave pain-enhancing effects. Immediately prior to the testingsession, participants rated the extent to which they expectedthe immersion procedure to change the pain produced by theelectrical stimulation. Expectancy ratings were given as a per-centage increase (+) or decrease (�) in pain. Analyses conduct-ed on these ratings confirmed that the two groups expected theimmersion procedure to produce opposite effects (all ts > 37.0and all Ps < .0001). Testing began with a 5-min baseline condi-tion during which subjects remained seated and relaxed. Suralnerve stimulations were then administered every 7 s for 10 min.Each stimulation provoked a withdrawal reflex and a somato-sensory-evoked brain potential. Four minutes after sural nervestimulation began, participants immersed their right arm incold water for 2 min. Pain ratings pertaining to sural nervestimulations were recorded every minute. The analgesia expec-tancy group and the hyperalgesia expectancy group had simi-lar withdrawal reflex thresholds (Mean ± SEM are14.8 mA ± 1.2 for the analgesia group and 15.8 mA ± 2.4 forthe hyperalgesia group; t = 0.13, P = .72) and perceived pre-immersion sural nerve stimulations as similarly painful(23.1 ± 15.5 for the analgesia group and 20.0 ± 12.3 for thehyperalgesia group; t = 0.25, P = .63).
2.6. Statistical analyses
All data are given as means ± SD, except where indicated.Comparisons made between groups were conducted usinganalysis of variance or independent sample t-tests. Withingroup analyses were conducted using paired sample t-tests
and one-sample t-tests. Relationships between our differentoutcome variables were looked at using Pearson productmoment correlations and hierarchical regression analyses.Bonferroni corrections were applied to ANOVAs and t-testswhere appropriate and P < .05 (two-tailed) was consideredstatistically significant.
3. Results
3.1. Perceived pain
Results showed large group differences in the effect ofthe arm immersion on sural nerve pain (Fig. 1; allFs > 4.34, all Ps < .05). Whereas the analgesia expectan-cy group experienced a 77% decrease in sural nerve painintensity during immersion, the hyperalgesia groupexperienced virtually no change in pain perception(0.05% decrease). The changes in pain produced by theimmersion procedure were strongly correlated to theexpected changes in pain (Fig. 1; r = .74, P = .0001).
3.2. Neurophysiological measures
During the immersion period, the amplitude of thewithdrawal reflex was significantly depressed for the
FfenenSnPc

140 P. Goffaux et al. / Pain 130 (2007) 137–143
analgesia group (Fig. 2; t = 3.85, P = .004) whereas anon-significant increase was observed for the hyperalge-sia group (t = 1.81, P = .10). The increase in RIII-relfexamplitude did not reach significance in the hyperalgesiagroup because of the greater response variabilityobserved in this group. Immersion-induced changes inreflex amplitude represented a 23% (±18.6) decreasefor the analgesia group and a 24% (±42.6) increase forthe hyperalgesia group. Importantly, immersion-in-duced changes in the spinal responses to the shock inthe analgesia group were significantly different fromthe immersion-induced changes recorded in the hyperal-gesia group (t = 3.20, P = .007). These results suggestthat expectations of pain relief can modulate the analge-sic response triggered by the counterirritation procedureand that the expectation of hyperalgesia completely
Fig. 2. Immersion-induced change in spinal and cortical activity differs depenhyperalgesia expected group show the average reflex amplitudes recorded forbaseline (i.e., the period of time prior to immersion). Group differences in refland show a reduction in reflex amplitude only for the analgesia expected groshow the average sural-nerve-evoked brain potentials recorded prior to andcomponents are shown. Our waveforms clearly demonstrate a large group dreductions are observed when analgesia is expected (b), than when hyperalg
blocked the normal analgesic response induced byDNIC.
As shown in Fig. 2, the immersion procedure alsoreduced N100, N150 and P260 amplitudes, consistentwith a DNIC-induced reduction in the spino-thalamo-cortical nociceptive response. This was observed forboth the analgesia expectancy and hyperalgesia expec-tancy groups (all Fs > 7.55, all Ps < .01). However, thesize of the P45 component did not change (F = 1.52,P = .23). Since the P45 component is the only compo-nent that is not related to nociceptive processing andsince its amplitude did not vary in our study, this sug-gests that the inhibition induced by counterirritationwas indeed pain-specific. Importantly, the reduced posi-tive amplitude of the late P260 brain potential was muchgreater for the analgesia group (�13.95 lV) than for the
ding on expectations of pain relief. (a) Analgesia expected group and (c)each sural nerve stimulation and expressed as a percentage value fromex amplitude are noticeable only during the immersion period (arrows)up. (b) Analgesia expected group and (d) hyperalgesia expected groupduring arm immersion. Early (P45, N100) and late (N150, P260) brainifference in the suppression of the late P260 component. Larger P260
esia is expected (d).

P. Goffaux et al. / Pain 130 (2007) 137–143 141
hyperalgesia group (�4.86 lV; t = 3.20, P = .005). Allother components showed immersion-induced reduc-tions that were similar in amplitude for both groups(all ts < .45, all Ps > .65).
3.3. Association between expectations, perceived pain, andneurophysiological responses
To better appreciate the relative role of cortical andspinal mechanisms in the observed modulation of pain,we examined the relationship between expectations andimmersion-induced changes in nociceptive reflex activi-ty, P260 amplitude and perceived pain intensity. Corre-lation analyses revealed that changes in withdrawalreflex activity correlated with expected (r = .55,P = .01) and perceived (r = .58, P = .008) changes inpain. Likewise, changes in the amplitude of the P260potential correlated with expected (r = .48, P = .03)and perceived (r = .43, P = .05) changes in pain. How-ever, after removing the effects of expectation, the resid-ual variance in subjective pain relief (changes inperceived pain intensity) was no longer related to chang-es in the withdrawal reflex or P260 amplitude (r = .20,P = .21 for reflex activity and r = .09, P = .60 forP260 amplitude). This confirms that expectations doindeed play a critical role in explaining the link betweenphysiological changes and subjective reports of painrelief.
4. Discussion
The present study demonstrates that expectations canmodulate the activation of endogenous pain inhibitorysystems affecting spinal and cortical nociceptive respons-es. Contrary to the analgesia group, nociceptive reflexactivity and pain perception were not reduced duringcounterirritation in subjects expecting hyperalgesia. Thisimplies that expectations of increased pain blocked theendogenous analgesic processes normally activated bythe counterirritation stimulus. This finding providesdirect support to a growing number of neuroimagingstudies showing that psychological factors (expectation,emotion, attention,. . .) affect the neuronal activity ofbrain regions involved in descending pain inhibition(Petrovic et al., 2002; Tracey et al., 2002; Valet et al.,2004; Wager et al., 2004). In particular, it confirms thatexpectation-dependent changes in pain are associatedwith genuine changes in the activation of spinal pain-modulating circuits. Complete blocking of the normalanalgesic response produced by DNIC also supportsthe idea that anti-analgesic expectations can dramatical-ly reduce the effect of active analgesic treatments. Thisfinding corroborates previous results showing that theadministration of a pharmacologically active agent hasdifferent effects depending on the information provided(Flaten et al., 1999). For example, if participants are
told that a relaxant drug (carisoprodol) acts as a stimu-lant, then the amplitude of their blink reflexes is largerthan if they are told that carisoprodol acts as a relaxant(Flaten et al., 1999). This implies that expectations caneffectively block and potentially reverse the action ofactive drugs.
One potential limitation of our study is worth noting.In order to maximize the effects of expectation on per-ceived pain, we directly manipulated expectation effects.Unfortunately, this means that we do not have a controlcondition where the baseline DNIC response is mea-sured. Although this may appear problematic, severalreasons motivated the exclusion of a ‘‘no-expectation’’condition. First, pilot testing confirmed that participantsalways carry some degree of expectation regarding theimmersion procedure. As a result a no-expectation con-dition is impossible to provide. Second, our goal was tomaximize the effects of expectation. This was bestachieved by polarizing the anticipated effects of theimmersion procedure. Notwithstanding these reasons,it is important to note that numerous human and animalstudies confirm that the classic counterirritation phe-nomenon normally produces analgesic responses (seeMillan, 2002 for a review). We can be confident, there-fore, that our expectation effects are truly modulatingan analgesic phenomenon.
Finally, we found that expectancy-related changes inspinal inhibition were accompanied only by a change inthe amplitude of the late P260 potential. This means thatvariations in spinal inhibition do not produce corre-sponding changes throughout the pain matrix of thebrain. Although our reflex responses suggest that partof the expectation effect is mediated by changes in painfiber transmission, our SEP responses suggest that partof the expectation effect is also mediated cortically, bychanges in specific brain regions. In particular, theP260 surface potential, and the functions subsumed byits cortical generator, may be important for the effectof expectation on pain perception. EEG source localiza-tion studies confirm that the P260 activity evoked byacute painful stimuli is associated with the response ofneurons located in the ACC (Garcia-Larrea et al.,2003). This part of the brain is functionally connectedto the dorsolateral prefrontal cortex (Paus et al.,2001), the same region involved in placebo analgesia(Wager et al., 2004). Given this connectivity, it is possi-ble that the ACC facilitate the integration and interpre-tation of sensory signals based on the expectationsgenerated by the dorsolateral prefrontal cortex (Paus,2001). In other words, the dorsolateral prefrontal cortexand the ACC may serve to regulate perceptions in rela-tion to changes in cognition or motivational states.Future studies conducted using multi-lead electrodemontages could confirm these interactions and facilitatethe measurement of subtle topographic changes result-ing from differences in expectation.

142 P. Goffaux et al. / Pain 130 (2007) 137–143
Recent functional imaging studies showing connec-tivity between the prefrontal cortex, the ACC and thebrainstem (Lorenz et al., 2003; Wager et al., 2004) alsosupport the proposal that expectations activate a com-plex and highly modifiable pain-processing network.These studies suggest that interactions between prefron-tal and cingulate regions of the cortex underlie subjec-tive changes in the experience of pain. These changesare thought to reflect mechanisms recruited in anattempt to align the felt internal state of the body withan anticipated outcome (Rainville, 2002). On the otherhand, the coupling between activity in the prefrontalcortex and the brainstem is thought to be directly relatedto decreases in subjective pain ratings and is assumed toreflect the engagement of descending modulatoryresponses (Lorenz et al., 2003). For the first time, wehave been able to show that expectation effects actuallychange the physiological responses associated with both
Brain stem• Mesencephalon (including the
periaqueductual gray)• Pons• Medulla (including the nucleus
raphe magnus)
Spine
- +
+/-
Prefrontal regions• rostral ACC • orbitofrontal cortex• dorsolateral prefrontal cortex
Brain evoked p
Dorsalneur
Fig. 3. Representation of the different structures and pathways suggested to p(suggestions, conditioned-learning, etc.) activate anticipatory reactions thatand cortico-subcortical pathways. Cortico-cingulate connections are thougregions), resulting in a reappraisal of the significance of pain as a functionconnections are thought to affect descending inhibitory systems located in thespinal responses and modifies afferent sensory pain transmission. Descendinline). In the present study, expectations of hyperalgesia: (1) blocked the inhactivity), (2) attenuated the inhibition of the cortical response (somatosensorcounterirritation on pain perception (subjective ratings).
spinal inhibition and cortical activity. The co-occurrenceof spinal and cortical changes suggests that expectation-dependent analgesia is not a monolithic phenomenonthat depends only on descending inhibition or cognitiveappraisal. Instead, both mechanisms appear to play arole in mediating the effects of expectation. From aneuroanatomical perspective, this suggests that multiplepathways must underlie expectation effects. We proposethat expectations of pain relief provoke a cascade ofchanges resulting from the activity of cerebro-spinaland cortico-cortical connections (Fig. 3).
Our results are also important from a clinical per-spective since they demonstrate that the efficacy of validanalgesic procedures can be completely blocked by anti-analgesic expectancy. This means that a valid pain treat-ment can lose its clinical efficacy if patients do not expectpain relief. A logical extension of this would be that pre-vious experiences with ineffective treatments will induce
+
+
Nociceptive reflex activity
+/-
+ Caudal ACC
otential
horn ons
DNICLoop
AscendingSpino-cortical Pathways
• Nociceptive transmission
lay a role in mediating expectancy-related effects. Psychosocial contextsare guided by the prefrontal cortex and mediated by cortico-cingulateht to influence the activity of ACC neurons (in particular in caudal
of what is expected or contextually appropriate. Cortico-subcorticalbrainstem (i.e., DNIC system activity). A change in these systems altersg projections representing excitatory influences are also shown (dottedibitory effect of DNIC on the spinal response (nociceptive RIII-reflexy-evoked potentials), and (3) completely blocked the analgesic effect of

P. Goffaux et al. / Pain 130 (2007) 137–143 143
expectations of failure, which may interfere with the effi-cacy of future treatments (Charron et al., 2006). Critical-ly, this situation may contribute to the poor efficacy ofpain treatments in chronic pain conditions, as mostchronic pain patients are likely to have been exposedto a number of ineffective treatments over the courseof their disease. Therefore, a conservative approach toanalgesia when addressing initial complaints of painmay be ill advised given the undesirable long-term con-sequences which may develop when analgesic failure isfirst experienced.
Acknowledgements
We thank K. von Bochmann and E. Leclerc for helpin testing and data processing and N. Julien for helpfulcomments and proof-reading. This work was supportedby grants from the Louise Edwards Awards (S.M. andP.G.) and from the CIHR Placebo Net (S.M. and P.R.).
References
Charron J, Rainville P, Marchand S. Direct comparison of placeboeffects on clinical and experimental pain. Clin J Pain2006;22:204–11.
Dowman R. Pain-evoked anterior cingulate activity generating thenegative difference potential may reflect response selection pro-cesses. Psychophysiology 2002;39:369–79.
Dowman R. SEP topographies elicited by innocuous and noxious suralnerve stimulation. I. Identification of stable periods and individualdifferences. Electroencephalogr Clin Neurophysiol1994a;92:291–302.
Dowman R. SEP topographies elicited by innocuous and noxious suralnerve stimulation. II. Effects of stimulus intensity on topographicpattern and amplitude. Electroencephalogr Clin Neurophysiol1994b;92:303–15.
Dowman R, Darcey TM. SEP topographies elicited by innocuous andnoxious sural nerve stimulation. III. Dipole source localizationanalysis. Electroencephalogr Clin Neurophysiol 1994;92:373–91.
Dowman R, Schell S. The pain-related negative difference potential: adirect measure of central pain pathway activity or of interactionsbetween the innocuous somatosensory and pain pathways? ClinNeurophysiol 1999;29:423–42.
Fields HL, Basbaum AI. Central nervous system mechanisms of painmodulation. In: Wall PD, Melzack R, editors. Textbook ofPain. London: Churchill Livingstone; 1999. p. 309–29.
Flaten MA, Simonsen T, Olsen H. Drug-related information generatesplacebo and nocebo responses that modify the drug response.Psychosom Med 1999;61:250–5.
Garcia-Larrea L, Frot M, Valeriani M. Brain generators of laser-evoked potentials: from dipoles to functional significance. ClinNeurophysiol 2003;33:279–92.
Le Bars D, Dickenson AH, Besson JM. Diffuse noxious inhibitorycontrols (DNIC). I. Effects on dorsal horn convergent neurones inthe rat. Pain 1979;6:283–304.
Lenz FA, Rios M, Zirh A, Chau D, Krauss G, Lesser RP. Painfulstimuli evoke potentials recorded over the human anterior cingu-late gyrus. J Neurophysiol 1998;79:2231–4.
Lorenz J, Minoshima S, Casey KL. Keeping pain out of mind: the roleof the dorsolateral prefrontal cortex in pain modulation. Brain2003;126:1079–91.
Millan MJ. Descending control of pain. Prog Neurobiol2002;66:355–474.
Montgomery GH, Kirsch I. Classical conditioning and the placeboeffect. Pain 1997;72:107–13.
Paus T. Primate anterior cingulate cortex: where motor control, driveand cognition interface. Nature Rev Neurosci 2001;2:417–24.
Paus T, Castro-Alamancos MA, Petrides M. Cortico-cortical connec-tivity of the human mid-dorsolateral frontal cortex and itsmodulation by repetitive transcranial magnetic stimulation. Eur JNeurosci 2001;14:1405–11.
Petrovic P, Kalso E, Petersson KM, Ingvar M. Placebo and opioidanalgesia – imaging a shared neuronal network. Science2002;295:1737–40.
Ploghaus A, Tracey I, Gati JS, Clare S, Menon RS, Matthews PM,Rawlins JNP. Dissociating pain from its anticipation in the humanbrain. Science 1999;284:1979–81.
Price DD, Milling LS, Kirsch I, Duff A, Montgomery GH, Nicholls SS.An analysis of factors that contribute to the magnitude of placeboanalgesia in an experimental paradigm. Pain 1999;83:147–65.
Rainville P. Brain mechanisms of pain affect and pain modulation.Curr Opin Neurobiol 2002;12:195–204.
Rainville P, Duncan GH. Functional brain imaging of placeboanalgesia: methodological challenges and recommendations. Pain2006;121:177–80.
Sandrini G, Serrao M, Rossi P, Romaniello A, Cruccu G, Willer JC. TheLower limb flexion in humans. Prog Neurobiol 2005;77:353–95.
Tarkka IM, Treede RD. Equivalent electrical source analysis of pain-related somatosensory evoked potentials elicited by a CO2 laser. JClin Neurophysiol 1993;10:513–9.
Tracey I, Ploghaus A, Gati JS, Clare S, Smith S, Menon RS, MatthewsPM. Imaging attentional modulation of pain in the periaqueductalgrey in humans. J Neurosci 2002;22:2748–52.
Valeriani M, Rambaud L, Mauguiere F. Scalp topography and dipolarsource modelling of potentials evoked by CO2 laser stimulation ofthe hand. Electroencephalogr Clin Neurophysiol 1996;100:343–53.
Valet M, Sprenger T, Boecker H, Willoch F, Rummeny E, Conrad B,Erhard P, Tolle TR. Distraction modulates connectivity of thecingulo-frontal cortex and the midbrain during pain-an fMRIanalysis. Pain 2004;109:399–408.
Wager TD, Rilling JK, Smith EE, Sokolik A, Casey KL, Davidson RJ,Kosslyn SM, Rose RM, Cohen JD. Placebo-induced changes infMRI in the anticipation and experience of pain. Science2004;303:1162–7.
Willer JC, De Broucker T, Le Bars D. Encoding of nociceptive thermalstimuli by diffuse noxious inhibitory controls in humans. JNeurophysiol 1989;62:1028–38.
Willer JC, Le Bars D, De Broucker T. Diffuse noxious inhibitorycontrols in man: involvement of an opioidergic link. Eur JPharmacol 1990;182:347–55.





























