Embed Size (px)
Citation preview

Nanomedicine: Nanotechnology, B
Diagnostic
Developing scanning probe–based nanodevices—
stepping out of the laboratory into the clinic
Martin Stolz, PhD, Ueli Aebi, PhD4 Daniel Stoffler, PhDM.E. Muller Institute for Structural Biology, Biozentrum, University of Basel, Switzerland
Received 25 April 2006; accepted 19 July 2006
www.nanomedjournal.com
Abstract This report focuses on nanotools based on the scanning force microscope (SFM) for imaging,
1549-9634/$ – see fro
doi:10.1016/j.nano.20
No conflict of inte
4 Corresponding
University of Basel, B
E-mail address: u
measuring, and manipulating biological matter at the sub-micron scale. Because pathophysiological
processes often occur at the (sub-) cellular scale, the SFM has opened the exciting possibility to spot
diseases at a stage before they become symptomatic and cause functional impairments in the affected
part of the body. Such presymptomatic detection will be key to developing effective therapies to slow
or halt disease progression.
D 2007 Elsevier Inc. All rights reserved.
Key words: Multifunctional SFM probes; Articular cartilage; Osteoarthritis; Coronary arteries; Arteriosclerosis; Molecular
motors; AFM; Clinical diagnostics; Nanotechnology; Tissue engineering; Actin; Asymmetric unit membrane;
Nuclear pore complex; Nanomedicine; IT SFM; Advanced health care
Introduction
Living matter is composed of a large number of
biological bnanomachinesQ, so it seems hardly surprising
that many diseases have their onset at the nanometer scale.
In fact, the idea of using bnanotoolsQ for detecting diseases
and repairing the human body was first put forward in the
late 1950s by the renowned American physicist and Nobel
laureate Richard Feynman during his famous speech,
bThere’s Plenty of Room at the BottomQ [1]. One such
nanotool is the scanning force microscope (SFM), a
variation of the scanning tunneling microscope for which
Gerd Binnig and Heinrich Rohrer were awarded the Physics
Nobel Prize in 1986 [2,3]. The SFM uses a fine tip at the
end of a cantilever, which scans over a specimen surface and
thereby maps its elevations and depressions with great
accuracy. The SFM provides not only the beyesQ for imaging
biological matter at scales from nanometers up to milli-
meters, but also the bfingersQ for measuring and manipulat-
ing biological matter at the level of single molecules,
nt matter D 2007 Elsevier Inc. All rights reserved.
07.01.001
rest was reported by the authors of this paper.
author. M.E. Mqller Institute for Structural Biology,
asel, Switzerland.
[email protected] (U. Aebi).
organelles, up to entire cells and tissues. For example, by
varying the force by which the tip of the SFM presses onto
the specimen surface while monitoring its deformation at a
given point—called indentation-type (IT) SFM—the stiff-
ness of the specimen may be determined at this position.
Moreover, the SFM tip may be bfunctionalizedQ to serve as asensor for a broad range of molecular targets. It is expected
that in the near future novel biochemical and biophysical
tests will emerge that, in turn, will greatly improve the
sensitivity and speed of medical diagnostics. Evidently, the
prospects of this unique nanosensor and nanoactuator in
biomedical research and clinical applications are only
limited by our imagination.
Imaging: Visualizing dynamic processes at the molecular
scale, from bsnapshotsQ to movies
Transmission electron cryomicroscopy (cryo-EM) [4-6]
is a powerful method for studying the architecture and
assembly of macromolecules and their interactions with
substrates in a close-to-native environment [7]. However,
major drawbacks regarding applications in biology are the
high investment for a state-of-the-art microscope and the
ability of this technique to provide only bsnapshotsQ of
iology, and Medicine 3 (2007) 53–62

Fig 1. SFM imaging of the mammalian urothelial plaques and plaque particles. A, SFM imaging provides absolute height measurements of the asymmetric unit
membrane (AUM) on stable immobilized urothelial plaques at a mica surface in a close-to-native buffer environment: The overall height of the entire urothelial
plaque was measured to ~12.5 nm; the height of the lipid membrane is ~5.5 nm (A, substrate level, i.e., mica surface; step size between A and B; B, lipid
bilayer (~5.5 nm) plus cytoplasmic face (~0.5 nm); C, overall height 12.5 F 0.7 nm); the leaflet above the lipid bilayer at the luminal side raised by ~6.5 nm
(step size between B and C) above the luminal membrane surface and the donut-shaped protrusions of the cytoplasmic side is ~0.5 nm above the lipid
membrane. (B–E) Surface topography of the 16-nm particle by SFM. B, Low-magnification view of the luminal face of several interconnected urothelial
plaques. C, High-magnification view of the luminal surface of an urothelial plaque. D, Low-magnification view of the cytoplasmic face of several urothelial
plaques. E, High-magnification view of the cytoplasmic face of several urothelial plaques. F, The averaged images of the luminal (upper left half in orange)
and cytoplasmic (lower right half in blue) surfaces of urothelial plaques. (A–E, Scale bars = 100 nm; F, center-to-center distance = 16 nm).
M. Stolz et al. / Nanomedicine: Nanotechnology, Biology, and Medicine 3 (2007) 53–6254
the biological specimen (being immobilized in a thick layer
of vitrified ice). Also, cryo-EM does not allow measuring of
the functional properties of tissue.
Over the past decade, the SFM has opened completely
new vistas for analyzing the surface topography of biological
matter in its aqueous environment and at a resolution
comparable to that achieved by EM [8-18]. The most recent
generation of high-speed SFMs now permits the recording
of dynamic biological processes at close-to-video speed
[19-23]. A dynamic process may involve lateral [24-26] or
rotational motions [27,28], as well as bspatially stationaryQprocesses, those that do not exhibit any lateral or rotational
movement such as, for example, the opening and closing of
gates and channels [29-32].
Imaging the asymmetric unit membrane
The distinct nanostructure of the urothelial plaque, also
termed basymmetric unit membraneQ (AUM), offers an ideal
system for membrane visualization using the SFM. Accord-
ingly, the apical surface of the mammalian bladder
urothelium is covered almost entirely by rigid-looking,
concave membrane plaques consisting of 16-nm protein
particles that naturally form two-dimensional crystalline
arrays (Figure 1, A, B, and D) [33-36]. These plaques play
important roles in urothelial function and diseases, including
(1) the permeability barrier function to protect the underly-
ing tissues from the potentially harmful components of the
urine; (2) the reversible adjustment of the apical urothelial
surface area during the urination cycle; and (3) the receptor
function for the uropathogenic type 1-piliated bacteria,
which cause the majority of urinary tract infections, one of
the most common bacterial infections [37,38].
Urothelial plaques are composed of distinct 16-nm
diameter particles (Figure 1, C). Figure 1, F depicts the
averaged SFM images of the luminal and cytoplasmic
surfaces of urothelial plaques. In their native environment,
the urothelial plaque particles are hexagonally packed and,
based on their bfolded-ribbonQ topology [33], are predes-
tined to cooperatively rearrange during bladder distention.
Possibly the plaque particles undergo conformational
changes induced by effector molecules contained in the
urine. Experiments are now needed to track the formation of
plaques by time-lapse SFM and to eventually investigate
different conformations of individual plaque particles [36].
Imaging actin dynamics and assembly
Actin’s oligomerization, polymerization, and polymor-
phism have been investigated over the past decade at atomic
detail by combining x-ray and EM data. Figure 2, A depicts
negatively stained actin filaments recorded by transmission
electron microscopy (TEM). Actin exhibits a myriad of
diverse functions, most of which ultimately depend on its
intrinsic ability to rapidly assemble and disassemble
filamentous structures [39-41]. Actin controls the mechan-
ical stability and dynamics of the cytoskeleton and thereby
the overall shape, motility, and plasticity of cells. Actin is
also directly involved in muscle contraction where actin
filaments (F-actin) are serving as bmolecular tracksQ for
myosin motors to step along.
The bulk of structural information portrays actin in one
specific functional state. Hence, imaging F-actin under
native conditions by SFM, such as shown in Figure 2, B, is
useful not only to compare the native structure of the
filament with its atomic model, but also to eventually
directly monitor structural changes induced by chemical or
mechanical effectors [20,21,42]. For example, the new
generation of high-speed SFMs promises to directly
monitor muscle contraction at work. Here, the SFM tip

Fig 2. Imaging of actin filaments by transmission electron microscopy (TEM) and by SFM. A, Negative-stain TEM image of actin filaments. B, Native actin
filaments grown on the positively charged surface of a lipid film and transferred onto a hydrophobic support suitable for SFM and kept in buffer solution. SFM
imaging done in tapping mode of native actin filaments revealed filament dimensions (36-nm crossover repeat of the long-pitch helix crossover strand) similar
to those obtained by x-ray diffraction and electron microscopy. The width represents ~40 to 60 nm. (Scale bars: A, 100 nm, 20 nm (inset); B, 100 nm.
M. Stolz et al. / Nanomedicine: Nanotechnology, Biology, and Medicine 3 (2007) 53–62 55
would be employed to track myosin heads in their different
conformational states during their cyclic interaction with
F-actin filaments.
Consequently, our knowledge about actin filaments has
evolved from a rigid bbeads-on-a-stringQ model to that of a
complex, highly dynamic protein polymer. To this end, the
SFM is the sole microscope providing us with high-
resolution images revealing the dynamics of actin in its
various functional states. Thus, time-lapse SFM should
further our present understanding of actin filament structure,
polymerization, and dynamics, as well as the interaction of
actin with myosin motors during muscle contraction and
cell locomotion.
Imaging nuclear pore complex architecture and dynamics
Nuclear pore complexes (NPCs; Figure 3, A) are large
macromolecular assemblies embedded in the double mem-
brane of the nuclear envelope (NE). They are the major
gateways mediating transport of ions, small molecules,
proteins, RNAs, and ribonucleoprotein particles in and out
of the nucleus in interphase cells (see Fahrenkrog and Aebi
[43]). Delineating the three-dimensional (3D) structure and
molecular architecture of the NPC is a prerequisite to
mechanistically understand its role in mediating nucleocy-
toplasmic transport. Over the past several years significant
advances have been made in the use of electron microscopy
techniques to elucidate the 3D structure and architectural
design of the NPC [44]. These studies indicated that NPCs
are rather dynamic supramolecular assemblies exhibiting a
high degree of structural plasticity, where significant
structural changes can be induced by cellular signals such
as calcium or nucleotides [44].
Whereas EM provides only snapshots of distinct func-
tional states, time-lapse SFM allows the imaging of the
same specimen in various stages of activity. Hence, many
laboratories began imaging NPCs by SFM (listed in [29]).
However, in most cases the specimen was chemically fixed,
exposed to detergents, or dehydrated and rehydrated at some
stage during its preparation for SFM. Thus, it remained
unclear whether the responses of NPCs to effector mole-
cules depicted in some of these studies did indeed represent
bona fide structural changes or distinct functional states
in response to adding effectors or ligands, or whether
they were merely due to specimen preparation effects,
for example, differential extraction of labile or weakly
bound NPC constituents or plastic deformations caused by
surface tension.
To overcome these limitations, it was important to
develop an isolation-preparation protocol for Xenopus
oocyte NEs that avoids exposure to detergents, chemical
fixation, or dehydration-rehydration steps. Based on such a
protocol [29], we established SFM imaging conditions
allowing the repeated scanning of these native NE samples
in physiological buffer environment, so that one and the
same NPC can be watched in response to chemical effectors.
As depicted in Figure 3, B and C, the NE appears highly
asymmetric with regard to the cytoplasmic and nuclear
surface topography of its NPCs, similar to metal-shadowed
specimens imaged by EM [45]. In areas devoid of NPCs,
bremnantsQ of the nuclear lamina meshwork [46] could
readily be depicted (Figure 3, C). With this preparation
protocol at hand, we performed time-lapse SFM experiments
to monitor the nuclear surface topography of completely
unfixed NEs. These experiments revealed the repeated
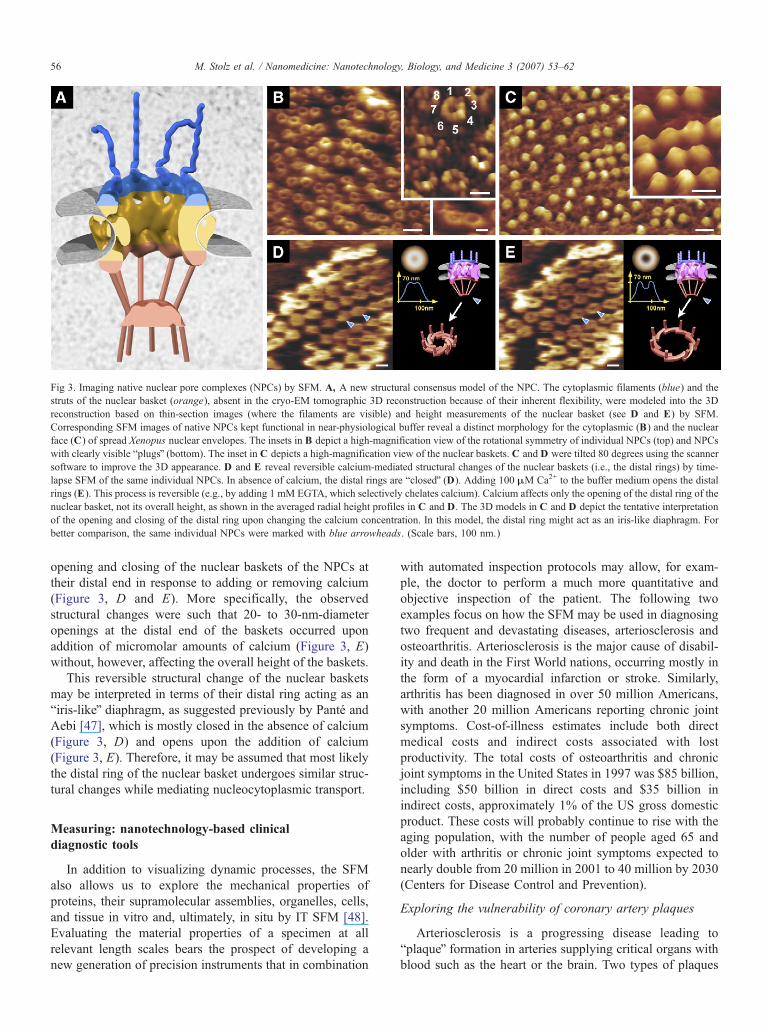
Fig 3. Imaging native nuclear pore complexes (NPCs) by SFM. A, A new structural consensus model of the NPC. The cytoplasmic filaments (blue) and the
struts of the nuclear basket (orange), absent in the cryo-EM tomographic 3D reconstruction because of their inherent flexibility, were modeled into the 3D
reconstruction based on thin-section images (where the filaments are visible) and height measurements of the nuclear basket (see D and E) by SFM.
Corresponding SFM images of native NPCs kept functional in near-physiological buffer reveal a distinct morphology for the cytoplasmic (B) and the nuclear
face (C) of spread Xenopus nuclear envelopes. The insets in B depict a high-magnification view of the rotational symmetry of individual NPCs (top) and NPCs
with clearly visible bplugsQ (bottom). The inset in C depicts a high-magnification view of the nuclear baskets. C and D were tilted 80 degrees using the scanner
software to improve the 3D appearance. D and E reveal reversible calcium-mediated structural changes of the nuclear baskets (i.e., the distal rings) by time-
lapse SFM of the same individual NPCs. In absence of calcium, the distal rings are bclosedQ (D). Adding 100 MM Ca2+ to the buffer medium opens the distal
rings (E). This process is reversible (e.g., by adding 1 mM EGTA, which selectively chelates calcium). Calcium affects only the opening of the distal ring of the
nuclear basket, not its overall height, as shown in the averaged radial height profiles in C and D. The 3D models in C and D depict the tentative interpretation
of the opening and closing of the distal ring upon changing the calcium concentration. In this model, the distal ring might act as an iris-like diaphragm. For
better comparison, the same individual NPCs were marked with blue arrowheads. (Scale bars, 100 nm.)
M. Stolz et al. / Nanomedicine: Nanotechnology, Biology, and Medicine 3 (2007) 53–6256
opening and closing of the nuclear baskets of the NPCs at
their distal end in response to adding or removing calcium
(Figure 3, D and E). More specifically, the observed
structural changes were such that 20- to 30-nm-diameter
openings at the distal end of the baskets occurred upon
addition of micromolar amounts of calcium (Figure 3, E)
without, however, affecting the overall height of the baskets.
This reversible structural change of the nuclear baskets
may be interpreted in terms of their distal ring acting as an
biris-likeQ diaphragm, as suggested previously by Pante and
Aebi [47], which is mostly closed in the absence of calcium
(Figure 3, D) and opens upon the addition of calcium
(Figure 3, E). Therefore, it may be assumed that most likely
the distal ring of the nuclear basket undergoes similar struc-
tural changes while mediating nucleocytoplasmic transport.
Measuring: nanotechnology-based clinical
diagnostic tools
In addition to visualizing dynamic processes, the SFM
also allows us to explore the mechanical properties of
proteins, their supramolecular assemblies, organelles, cells,
and tissue in vitro and, ultimately, in situ by IT SFM [48].
Evaluating the material properties of a specimen at all
relevant length scales bears the prospect of developing a
new generation of precision instruments that in combination
with automated inspection protocols may allow, for exam-
ple, the doctor to perform a much more quantitative and
objective inspection of the patient. The following two
examples focus on how the SFM may be used in diagnosing
two frequent and devastating diseases, arteriosclerosis and
osteoarthritis. Arteriosclerosis is the major cause of disabil-
ity and death in the First World nations, occurring mostly in
the form of a myocardial infarction or stroke. Similarly,
arthritis has been diagnosed in over 50 million Americans,
with another 20 million Americans reporting chronic joint
symptoms. Cost-of-illness estimates include both direct
medical costs and indirect costs associated with lost
productivity. The total costs of osteoarthritis and chronic
joint symptoms in the United States in 1997 was $85 billion,
including $50 billion in direct costs and $35 billion in
indirect costs, approximately 1% of the US gross domestic
product. These costs will probably continue to rise with the
aging population, with the number of people aged 65 and
older with arthritis or chronic joint symptoms expected to
nearly double from 20 million in 2001 to 40 million by 2030
(Centers for Disease Control and Prevention).
Exploring the vulnerability of coronary artery plaques
Arteriosclerosis is a progressing disease leading to
bplaqueQ formation in arteries supplying critical organs with
blood such as the heart or the brain. Two types of plaques

Fig 4. Imaging and measuring native coronary artery tissue. A, SFM imaging reveals the intact endothelial cell layer lining the lumen of a porcine artery in
physiological buffer. Individual cells can clearly be delineated, as can their segregating cell junctions. B, The surface endothelial layer of a wild-type mouse
coronary artery was mechanically removed by surgical tools for inspection in physiological buffer. The SFM image reveals the subsurface layer that is
supporting the endothelium. The inset documents the 67-nm axial repeat of the collagen fibrils. C, SFM imaging of rat-2 cells as imaged under culture
conditions. Different cells and their cytoskeletal actin network are visible. (Scale bars: A, 5 Mm; B, 2 Mm, inset 500 nm; C, 25 lm).
M. Stolz et al. / Nanomedicine: Nanotechnology, Biology, and Medicine 3 (2007) 53–62 57
exist: fibrous, stable plaques that are rarely life-threatening
[49-53], and vulnerable, lipid-rich plaques prone to me-
chanical rupture thus leading to myocardial infarction and
stroke. Conventionally, discrimination of the two types of
plaques is performed histologically during an autopsy [54].
In our attempt to investigate plaques at different stages of
their development by SFM, we recently established two new
preparation protocols that not only take into account the
fragility of the soft coronary arteries by maintaining its bona
fide structure in a nondestructive sample treatment, but also
permit the investigation of cultured cell lines under well-
defined culture conditions [55]. As a first result, we
succeeded in improving the imaging resolution of those
soft biological specimens as documented in Figure 4, A to C.
Figure 4, A and B, depict SFM images of native coronary
artery endothelium in physiological buffer taken from a pig
or a wild-type mouse, respectively. For imaging the
underlying vascular extracellular matrix that consists mainly
of collagen fibrils, we mechanically removed the endothelial
cell layer (Figure 4, B). In addition to the preparation of
pieces of native coronary arteries, cultured vascular endo-
thelial cells may be studied as an in vitro model system. As
a first step toward this latter goal, Figure 4, C displays an
example of imaging living rat fibroblasts by SFM. The cells
were imaged under optimal culture conditions, that is, by
continuously exchanging the culture medium at 378C to
assure viability of the cells in the SFM over extended
periods of time. Such in vitro model systems may eventually
be used for testing drugs and pharmaceutical agents
targeted, for example, at cytoskeletal elements to affect
their 3D structure and spatial organization, and hence their
mechanical properties.
The ability to inspect the bona fide structure and assess
the local biomechanical properties of soft biological tissue is
paramount, because the investigator can thereby avoid
artifacts (e.g., due to freezing and fixation). The SFM
allows imaging, measuring, and manipulating native coro-
nary artery tissue in its physiological context. Moreover,
there is great potential for directly observing changes in
individual vascular endothelial cells in response to mechan-
ical or pharmacological stimulation by time-lapse SFM. In
the context of studying atherosclerosis, plaques might not
only be localized but also differentiated in terms of their
vulnerability by measuring their local, scale-dependent
biomechanical properties, thereby depicting differences in
tissue elasticity and plasticity by an experimental approach
being similar to that reported previously [48,55].
Inspection of articular cartilage
Osteoarthritis is a painful and disabling joint disease that
affects millions of people by progressively degrading the
articular cartilage. Osteoarthritis poses a dilemma: it usually
begins by attacking different joints long before middle age,
but it cannot be diagnosed until it becomes symptomatic
decades later, at which point the structure and biomechan-
ical properties of the affected cartilage are usually irrevers-
ibly altered. Currently available devices for measuring the
biomechanical properties of cartilage typically work at the
millimeter scale or above [48,56-60] and hence are not able
to assess the cellular and molecular features of cartilage (i.e.,
the scale at which biomechanical processes occur and
pathological lesions typically start). In contrast, the SFM
can readily image cartilage morphology and measure its
biomechanical properties at the micrometer scale and below.
Thus, the SFM opens the exciting possibility to detect
pathological features of articular cartilage long before they
become symptomatic and cause a functional impairment of
the affected joint. Detection of OA at a presymptomatic
stage might be key to the development of effective therapies
to slow or stop its progression.
Recently, we used ex vivo IT SFM on articular cartilage
biopsies to measure the mechanical stiffness, which slowly

Fig 5. Imaging and measuring native articular cartilage. Surface topography of (A) normal articular cartilage, (B) osteoarthritic articular cartilage, and (C) the
corresponding elasticity measurements performed at the nanometer scale with a sharp pyramidal tip. A, The 67-nm axial repeat distance of individual collagen
fibrils was clearly resolved by SFM imaging. B, In contrast to the normal cartilage that exhibited a random orientation of the collagen fibril network, in the
diseased cartilage the collagen fibrils coalesced on top of each other and exhibited a preferred orientation. This orientation might follow the directed movement
within the joint more easily once the GAGs become digested in the course of the disease progression. C, Comparison of the two slopes of the corresponding
force displacement curves indicated stiffening of the osteoarthritic articular relative to the normal cartilage. (A and B, Scale bars = 1 lm).
M. Stolz et al. / Nanomedicine: Nanotechnology, Biology, and Medicine 3 (2007) 53–6258
but definitely changes during progression of osteoarthritis.
Interestingly, when measured at the nanometer scale—but
not at the micrometer or millimeter scale—such stiffness
changes can be depicted long before clinical symptoms of
osteoarthritis (e.g., pain) arise. As illustrated in Figure 5, A,
normal articular cartilage exhibits a stochastic 3D orienta-
tion of the collagen fibrils, revealing a characteristic 67-nm
axial repeat. In contrast, osteoarthritic cartilage exhibits a
preferred orientation and a tendency of bundling of its
collagen fibrils, as indicated by the arrows in Figure 5, B.
Because of the mechanical stress, it is conceivable that upon
degradation of the interspersed glycosaminoglycan network,
the collagen fibrils are no longer spaced apart and coalesce
on top of each other, thereby slowly but definitely aligning
themselves in a direction representing the predominant joint
movement. As documented in Figure 5, C, when assessed
by IT SFM at the nanometer scale the osteoarthritic cartilage
appears mechanically significantly stiffer than the normal
cartilage. It should be noted, however, that there are also
stages and/or conditions of osteoarthritis where the cartilage
appears mechanically softer than normal cartilage, indicat-
ing that there exist multiple osteoarthritic sequelae, all
eventually leading to complete degeneration and/or disap-
pearance of the cartilage normally covering the moving
parts of a joint. Nevertheless, this finding opens the prospect
of early diagnosis, hopefully at a stage when disease
progression can still be halted.
Manipulating: using multifunctional SFM probes
as bfingersQ for minimally invasive therapeutic
interventions
Minimally invasive therapeutic interventions are increas-
ingly being performed using surgical tools with embedded
miniature sensors providing real-time information to im-
prove surgical results, hence benefiting both the patient and
the surgeon. Based on such tools, an envisioned scenario in
a future surgical theater might be the following: the
coronary arteries are first inspected by catheter-based SFM
for identifying and differentiating potential plaques into
bstableQ versus bvulnerableQ plaques based on their biome-
chanical properties. Next, a more detailed characterization
of the state of the vulnerable plaques might be achieved by
multifunctional SFM probes that will be able to measure
heat production or pH in the near field of individual plaques,
or can detect specific plaque surface markers using
fluorescent reporter molecules by a scanning near-field
optical microscopy (SNOM)-type probe being incorporated
into the SFM tip [61,62]. Obviously, the next step will be to
develop such multifunctional SFM probes into multipurpose
SFM-type actuators that will permit us to move from
diagnostics to treatment and, consequently, toward repairing
the diseased or damaged tissue locally.
Hence, novel, nanotechnology-based surgical devices are
now needed to permit the surgeon to perform interventions
that are currently not possible, for example, diagnosing and/
or treating a disease directly at the site where it occurs. In
this context, a minimally invasive, nanoactuator-type
bnanoscalpelQ would offer unprecedented precision for
performing delicate surgery at the level of individual cells
without damaging the surrounding tissue. Once a diseased
tissue can directly be targeted by a multifunctional nano-
device, a drug or a small amount of radioactive material
could be locally administered to the diseased site, destroy-
ing, for example, cancer cells one by one. The selectivity of
nanosurgery could even be further enhanced by tagging
specific cellular components with light-sensitive markers for
specific localization or by activating photolabile chemical
reactions by light-emitting local probes [63-65].
Ultimately, such surgical nanotools will adopt and
integrate a variety of functionalities that are currently being
explored in life science applications [66]. For example,

Fig 6. The arthroscopic SFM. A, Measurement by the arthroscopic SFM (arrows) in a phantom knee. B, Prototype of an arthroscopic SFM. C, The
arthroscopic SFM is a new tool that can be inserted into a standard arthroscopic tube for evaluating the health status (e.g., the elasticity) of cartilage and also for
quality control in follow-up examinations after cartilage replacements.
M. Stolz et al. / Nanomedicine: Nanotechnology, Biology, and Medicine 3 (2007) 53–62 59
multifunctional SFM devices are being developed and
implemented that include SNOM and conductive scanning
probe microscopy (CSPM). Whereas SNOM permits local
detection of fluorescently tagged cell membrane targets at the
single-event level [67,68], CSPM probes allow detection of
currents through individual membrane channels [69]. Fur-
thermore, SFM probes have been built that can detect small,
local temperature changes [70], which are often an indicator
of a local inflammation. Next, these devices will be
customized for minimally invasive endoscopic or catheter-
based interventions, ultimately, with single-cell precision.
A more precise diagnostics and early therapeutic
intervention will provide an accurate detection of the
clinical relevant parameters even before a disease becomes
symptomatic. In addition, tissue engineering may solve the
problem of the limited amount of autologous tissue
available for transplantation. Tissue engineers are producing
bspare partsQ for replacing defective tissue in the human
body, such as bone, cartilage, or tendon. As yet, however,
no synthesized tissue can mimic the long-term mechanical
stability, durability, and biocompatibility of authentic tissue.
One major obstacle in trying to bwatch and copyQ nature
concerns the primary reliance of our perception on the
macroscopic world that is accessible to our senses. Hence,
almost everything ever created by humans is based on our
macroscopic understanding of the phenomena and actions to
which we are constantly exposed. For example, all human-
made technology follows the rule of assembling macroscale
components or by carving bulk pieces of matter into smaller
chunks. As yet, we by and large lack a rational understand-
ing of the self-assembly processes that nature uses to
generate hierarchically organized, multifunctional biological
matter from a finite set of elementary building blocks. In
fact, the elementary building blocks of biological matter
(e.g., the proteins) are nanometer-scale structures that are
endowed with a remarkable functional versatility. They are
used for creating supramolecular assemblies such as, for
example, cytoskeletal filaments, or higher order functional
units like ribosomes or the mitotic apparatus. It is these
distinct supramolecular assemblies acting in concert that, in
turn, render a tissue or organ highly versatile in terms of its
structure, plasticity, and functional adaptability.
Articular cartilage is a hierarchically organized biomate-
rial exhibiting distinct nanoscale, mesoscale, and tissue-scale
structural features. Unless being subjected to pathological
alterations, it is built to warrant lifelong performance within
the human body. Yet, cartilage is a finely tunable tissue: its
biomechanical properties, for example, may be affected by
(1) physical or chemical effectors, (2) overexpression or
knock down of some of its molecular constituents [71-73], or
(3) controlled enzymatic digestion [48,74-76]. However, for
capturing the thus induced structural and biomechanical
changes, we may need a better understanding body the old-
timer, tools that are capable to image and measure cartilage at
the sub-micrometer scale. IT SFM, for example, can measure
changes in the stiffness of articular cartilage when operated
with a sub-micrometer tip that goes undetected with
micrometer to millimeter indenters [48]. Moreover, sub-
micrometer IT SFMmay be very effective for controlling and
optimizing the quality of engineered cartilage [48].
As a next step, we are developing an arthroscopic SFM
(Figure 6) for direct, quantitative in situ inspection of articular
cartilage morphology and biomechanical properties at the
sub-micrometer scale [60,77]. In addition to serving as a

M. Stolz et al. / Nanomedicine: Nanotechnology, Biology, and Medicine 3 (2007) 53–6260
potential diagnostic nanotool, the arthroscopic SFMmay also
be used for quality control after cartilage transplantations.
Ultimately, this novel nanodevicemight permit the surgeon to
rapidly, minimally invasively, and reliably assess the health
status of articular cartilage prophylactically and, if necessary,
to prescribe therapeutic interventions that slow or even halt
disease progression at a presymptomatic stage.
Going beyond cartilage, similar nanotools may be
designed and customized to manipulate transplanted tissue
or stimulate de novo tissue formation directly in situ more
generally, and ultimately, to design and produce tissue-
engineered body parts that exhibit long-term mechanical
stability, functionality, and biocompatibility [78].
Outlook
As documented in the case of osteoarthritis, the prospects
for detecting progression of a disease in the human body well
before it becomes symptomatic is considerable, thus opening
completely new vistas for treatment of such a disease.
Because most diseases have their origin at the cellular or
even molecular level, capturing them at this stage requires
diagnostic tools that operate at the cellular or molecular scale.
Hence, the call for nanotools, such as, for example, SFM or
more generally, scanning probe microscopes (SPMs) that
permit the imaging, measuring, and manipulation of biolog-
ical matter at the nanometer to micrometer scale. Obviously, a
prerequisite for an SPM to operate is to bring it to the
presumed defect site, for example, into the knee joint by an
arthroscopic approach. More generally, SPMs will have to be
designed and customized for endoscopic or even catheter-
based direct in situ operation. This requirement, in turn,
confronts the individual appearing for a prophylactic
examination of this kind with the need for an invasive
procedure—a circumstance that may prompt him/her to
decide against it, particularly in the absence of any
symptoms. For such prophylactic interventions to eventually
being tolerated by patient, not only must they be ambulant
and minimally invasive, but they will require a change of
paradigm vis-a-vis the patient, namely to undergo an invasive
procedure without feeling sick—indeed a challenge for
nanomedicine and managed health care!
Acknowledgments
This work was supported by an NCCR program grant on
bNanoscale ScienceQ awarded by the Swiss National ScienceFoundation, the bEU Network of Excellence 3D-EMQproject LSHG -CT-2004-502828, the M.E. Mqller Founda-tion of Switzerland, and the Canton Basel-Stadt.
References
[1] Feynman RP. There’s Plenty of Room at the Bottom. A transcript of the
classic talk that Richard Feynman gave on 29 December 1959 at the
annual meeting of the American Physical Society, Available from:
http://www.zyvex.com/nanotech/feynman.html.
[2] Binnig G, Rohrer H. Scanning tunnelling microscopy. Helv Phys Acta
1982;55(6):726 -35.
[3] Binnig G, Quate CF, Gerber C. Atomic force microscope. Phys Rev
Lett 1986;56(9):930 -3.
[4] Medalia O, Weber I, Frangakis AS, Nicastro D, Gerisch G,
Baumeister W, et al. Macromolecular architecture in eukaryotic cells
visualized by cryoelectron tomography. Science 2002;298:1209-13.
[5] Steven AC, Aebi U. The next ice age: cryo-electron tomography of
intact cells. Trends Cell Biol 2003;13:107-10.
[6] Lucic V, Forster F, Baumeister W. Structural studies by electron
tomography: from cells to molecules. Annu Rev Biochem 2005;
74:833-65.
[7] Hoenger A, Aebi U. 3-D reconstructions from ice-embedded and
negatively stained biomacromolecular assemblies: a critical compar-
ison. J Struct Biol 1996;117:99 -116.
[8] Czajkowsky DM, Hotze EM, Shao Z, Tweten RK. Vertical collapse of
a cytolysin prepore moves its transmembrane (-hairpins to the
membrane. EMBO J 2004;23:3206-15.
[9] Karrasch S, Hegerl R, Hoh JH, Baumeister W, Engel A. Atomic force
microscopy produces faithful high-resolution images of protein
surfaces in an aqueous environment. Proc Natl Acad Sci U S A
1994;91:836-8.
[10] Mou J, Yang J, Shao Z. Atomic force microscopy of cholera toxin B-
oligomers bound to bilayers of biologically relevant lipids. J Mol Biol
1995;248:507 -12.
[11] Muller DJ, Sass HJ, Muller SA, Buldt G, Engel A. Surface structures
of native bacteriorhodopsin depend on the molecular packing
arrangement in the membrane. J Mol Biol 1999;285:1903 -9.
[12] Muller DJ, Schoenenberger CA, Schabert F, Engel A. Structural
changes in native membrane proteins monitored at subnanometer
resolution with the atomic force microscope: a review. J Struct Biol
1997;119:149-57.
[13] Reviakine II, Bergsma-Schutter W, Brisson A. Growth of protein 2-D
crystals on supported planar lipid bilayers imaged in situ by AFM. J
Struct Biol 1998;121:356-61.
[14] Sanner M, Stolz M, Burkhard P, Kong XP, Min G, Sun TT, et al.
Visualizing nature at work from the nano to the macro scale.
Nanobiotechnol 2005;1:7 -22.
[15] Schabert FA, Henn C, Engel A. Native Escherichia coli OmpF
porin surfaces probed by atomic force microscopy. Science 1995;
268:92-4.
[16] Scheuring S, Ringler P, Borgnia M, Stahlberg H, Muller DJ, Agre P,
et al. High-resolution AFM topographs of the Escherichia coli water
channel aquaporin Z. EMBO J 1999;18:4981-7.
[17] Fotiadis D, Liang Y, Filipek S, Saperstein DA, Engel A, Palczewski
K, et al. Atomic-force microscopy: rhodopsin dimers in native disc
membranes. Nature 2003;421:127 -8.
[18] Stolz M, Stoffler D, Aebi U, Goldsbury C. Monitoring biomolecular
interactions by time-lapse atomic force microscopy. J Struct Biol
2000;131:171 -80.
[19] Viani MB, Schaffer TE, Paloczi GT, Pietrasanta I, Smith BL,
Thompson JB, et al. Fast imaging and fast force spectroscopy of
single biopolymers with a new atomic force microscope designed for
small cantilevers. Rev Sci Instrum 1999;70(11):4300-3.
[20] Ando T, Kodera N, Naito Y, Kinoshita T, Furuta K, Toyoshima
YY, et al. A high-speed atomic force microscope for studying
biological macromolecules in action. Chem Phys Chem 2003;4:
1196-202.
[21] Kodera N, Kinoshita T, Ito T, Ando T. High-resolution imaging of
myosin motor in action by a high-speed atomic force microscope. Adv
Exp Med Biol 2003;538:119-27.
[22] Kindt JH, Fantner GE, Cutroni JA, Hansma PK. Rigid design of fast
scanning probe microscopes using finite element analysis. Ultra-
microscopy 2004;100:259-65.
[23] Schitter G, Stark RW, Stemmer A. Fast contact-mode atomic force
microscopy on biological specimen by model-based control. Ultra-
microscopy 2004;100:253-7.

M. Stolz et al. / Nanomedicine: Nanotechnology, Biology, and Medicine 3 (2007) 53–62 61
[24] Hoenger A, Sablin EP, Vale RD, Fletterick RJ, Milligan RA. Three-
dimensional structure of a tubulin-motor-protein complex. Nature
1995;376:271 -4.
[25] Hirokawa N, Takemura R. Molecular motors and mechanisms of
directional transport in neurons. Nat Rev Neurosci 2005;6:201-14.
[26] Yildiz A, Selvin PR. Kinesin: walking, crawling or sliding along?
Trends Cell Biol 2005;15:112-20.
[27] Wang H, Oster G. Energy transduction in the F1 motor of ATP
synthase. Nature 1998;396:279-82.
[28] Rondelez Y, Tresset G, Nakashima T, Kato-Yamada Y, Fujita H,
Takeuchi S, et al. Highly coupled ATP synthesis by F1-ATPase single
molecules. Nature 2005;433:773-7.
[29] Stoffler D, Goldie KN, Feja B, Aebi U. Calcium-mediated structural
changes of native nuclear pore complexes monitored by time-lapse
atomic force microscopy. J Mol Biol 1999;287:741-52.
[30] Yellen G. The voltage-gated potassium channels and their relatives.
Nature 2002;419:35-42.
[31] Jones SW. Calcium channels: unanswered questions. J Bioenerg
Biomembr 2003;35:461 -75.
[32] Doyle DA. Structural themes in ion channels. Eur Biophys J 2004;
33:175-9.
[33] Walz T, Haner M, Wu XR, Henn C, Engel A, Sun TT, et al. Towards
the molecular architecture of the asymmetric unit membrane of the
mammalian urinary bladder epithelium: a closed twisted ribbon
structure. J Mol Biol 1995;248:887-900.
[34] Kachar B, et al. Three-dimensional analysis of the 16 nm urothelial
plaque particle: luminal surface exposure, preferential head-to-head
interaction, and hinge formation. J Mol Biol 1999;285:595-608.
[35] Oostergetel GT, Keegstra W, Brisson A. Structure of the major
membrane protein complex from urinary bladder epithelial cells by
cryo-electron crystallography. J Mol Biol 2001;314:245-52.
[36] Min G, Stolz M, Zhou G, Liang F, Sebbel P, Stoffler D, et al.
Localization of uroplakin Ia, the urothelial receptor for bacterial
adhesion FimH, on the six inner domains of the 16 nm urothelial
plaque particle. J Mol Biol 2002;317:697 -706.
[37] Sun TT, Liang FX, Wu XR. Uroplakins as markers of urothelial
differentiation. Adv Exp Med Biol 1999;462:7 -18.
[38] Hooton TM, Stamm WE. Diagnosis and treatment of uncompli-
cated urinary tract infection. Infect Dis Clin North Am 1997;11:
551-81.
[39] Steinmetz MO, Goldie KN, Aebi U. A correlative analysis of actin
filament assembly, structure, and dynamics. J Cell Biol 1997;138:
559-74.
[40] Steinmetz MO, Stoffler D, Hoenger A, Bremer A, Aebi U. Actin: from
cell biology to atomic detail. J Struct Biol 1997;119:295-320.
[41] Schoenenberger CA, Steinmetz MO, Stoffler D, Mandinova A, Aebi
U. Structure, assembly, and dynamics of actin filaments in situ and in
vitro. Microsc Res Tech 1999;47:38-50.
[42] Ando T, Kodera N, Takai E, Maruyama D, Saito K, Toda A, et al. A
high-speed atomic force microscope for studying biological macro-
molecules. Proc Natl Acad Sci U S A 2001;98:12468-72.
[43] Fahrenkrog B, Aebi U. The nuclear pore complex: nucleocytoplasmic
transport and beyond. Nat Rev Mol Cell Biol 2003;4:757-66.
[44] Stoffler D, et al. Cryo-electron tomography provides novel insights
into nuclear pore architecture: implications for nucleocytoplasmic
transport. J Mol Biol 2003;328:119 -30.
[45] Jarnik M, Aebi U. Toward a more complete 3-D structure of the
nuclear pore complex. J Struct Biol 1991;107:291-308.
[46] Aebi U, Cohn J, Buhle L, Gerace L. The nuclear lamina is a
meshwork of intermediate-type filaments. Nature 1986;323:560-4.
[47] Pante N, Aebi U. Toward the molecular dissection of protein import
into nuclei. Curr Opin Cell Biol 1996;8:397 -406.
[48] Stolz M, et al. Dynamic elastic modulus of porcine articular cartilage
determined at two different levels of tissue organization by indentation-
type atomic force microscopy. Biophys J 2004;86:3269-83.
[49] Ross R. The pathogenesis of atherosclerosis—an update. N Engl J
Med 1986;314:488-500.
[50] Naghavi M, Libby P, Falk E, Casscells SW, Litovsky S, Rumberger J,
et al. From vulnerable plaque to vulnerable patient: a call for new
definitions and risk assessment strategies: Part II. Circulation
2003;108:1772-8.
[51] Naghavi M, Libby P, Falk E, Casscells SW, Litovsky S, Rumberger J,
et al. From vulnerable plaque to vulnerable patient: a call for new
definitions and risk assessment strategies: Part I. Circulation
2003;108:1664-72.
[52] Libby P. Vascular biology of atherosclerosis: overview and state of the
art. Am J Cardiol 2003;91:3A-6A.
[53] Libby P, Theroux P. Pathophysiology of coronary artery disease.
Circulation 2005;111:3481-8.
[54] Virmani R, Kolodgie FD, Burke AP, Farb A, Schwartz SM. Lessons
from sudden coronary death: a comprehensive morphological
classification scheme for atherosclerotic lesions. Arterioscler Thromb
Vasc Biol 2000;20:1262-75.
[55] Reichlin T, Wild A, Durrenberger M, Daniels AU, Aebi U, Hunziker
PR, et al. Investigating native coronary artery endothelium in situ and
in cell culture by scanning force microscopy. J Struct Biol
2005;152:52 -63.
[56] Tkaczuk H. Human cartilage stiffness. In vivo studies. Clin Orthop
1986;206:301 -12.
[57] Lyyra T, Jurvelin J, Pitkanen P, Vaatainen U, Kiviranta I. Indentation
instrument for the measurement of cartilage stiffness under arthro-
scopic control. Med Eng Phys 1995;17:395 -9.
[58] Shepherd DE, Seedhom BB. A technique for measuring the
compressive modulus of articular cartilage under physiological
loading rates with preliminary results. Proc Inst Mech Eng [H]
1997;211:155-65.
[59] Appleyard RC, Swain MV, Khanna S, Murrell GA. The
accuracy and reliability of a novel handheld dynamic indentation
probe for analysing articular cartilage. Phys Med Biol 2001;46:
541 -50.
[60] Stolz M, Imer R, Staufer U, Aebi U. Nanotechnology on the verge of
stepping into the clinic: development of an arthroscopic AFM.
BioWorld 2003;4:2 -5.
[61] Leinhos T, Rudow O, Stopka M, Vollkopf A, Oesterschulze E.
Coaxial probes for scanning near-field microscopy. J Microsc 1999;
194:349-52.
[62] Eckert R, Freyland JM, Gersen H, Heinzelmann H, Schurmann G,
Noell W, et al. Near-field optical microscopy based on microfabricated
probes. J Microsc 2001;202:7 -11.
[63] Khodjakov A, Cole RW, Rieder CL. A synergy of technologies:
combining laser microsurgery with green fluorescent protein tagging.
Cell Motil Cytoskeleton 1997;38:311 -7.
[64] Sacconi L, Tolic-Norrelykke IM, Antolini R, Pavone FS. Combined
intracellular three-dimensional imaging and selective nanosurgery by
a nonlinear microscope. J Biomed Opt 2005;10:14002.
[65] Colombelli J, Reynaud EG, Rietdorf J, Pepperkok R, Stelzer EH. In
vivo selective cytoskeleton dynamics quantification in interphase
cells induced by pulsed ultraviolet laser nanosurgery. Traffic 2005;6:
1093-102.
[66] Miles M. Scanning probe microscopy. Probing the future. Science
1997;277:1845-7.
[67] Menger FM. Supramolecular chemistry and self-assembly. Proc Natl
Acad Sci U S A 2002;99:4818-22.
[68] Eckel R, Ros R, Decker B, Mattay J, Anselmetti D. Supramolecular
chemistry at the single-molecule level. Angew Chem Int Ed Engl
2005;44:484 -8.
[69] Frederix PL, Gullo MR, Akiyama T, Tonin A, Rooij NF, Staufer U,
et al. Assessment of insulated conductive cantilevers for biology
and electrochemistry. Nanotechnology 2005;16:997-1005.
[70] Cannaerts M, Chamirian O, Maex K, Van Haesendonck C. Mapping
nanometre-scale temperature gradients in patterned cobalt-nickel
silicide films. Nanotechnology 2002;13:149 -52.
[71] Aszodi A, Pfeifer A, Wendel M, Hiripi L, Fassler R. Mouse models
for extracellular matrix diseases. J Mol Med 1998;76:238-52.

M. Stolz et al. / Nanomedicine: Nanotechnology, Biology, and Medicine 3 (2007) 53–6262
[72] Helminen HJ, Saamanen AM, Salminen H, Hyttinen MM. Transgenic
mouse models for studying the role of cartilage macromolecules in
osteoarthritis. Rheumatology (Oxford) 2002;41:848-56.
[73] Fournier C. Where do T cells stand in rheumatoid arthritis? Joint Bone
Spine 2005;72:527-32.
[74] Korhonen RK, Laasanen MS, Toyras J, Lappalainen R, Helminen
HJ, Jurvelin JS, et al. Fibril reinforced poroelastic model predicts
specifically mechanical behavior of normal, proteoglycan depleted
and collagen degraded articular cartilage. J Biomech 2003;36:
1373-9.
[75] Basalo IM, Mauck RL, Kelly TA, Nicoll SB, Chen FH, Hung CT, et al.
Cartilage interstitial fluid load support in unconfined compression
following enzymatic digestion. J Biomech Eng 2004;126:779 -86.
[76] Saad OM, Myers RA, Castleton DL, Leary JA. Analysis of
hyaluronan content in chondroitin sulfate preparations by using
selective enzymatic digestion and electrospray ionization mass
spectrometry. Anal Biochem 2005;344:232-9.
[77] Hunziker P, Stolz M, Aebi U. Nanotechnology in medicine: moving
from the bench to the bedside. Chimia 2002;56:520-6.
[78] Langer R, Vacanti JP. Tissue engineering. Science 1993;260:920-6.





























