Embed Size (px)
Citation preview

1
DEVELOPMENT OF A SUITABLE SEMEN
EXTENDER FOR THE CRYOPRESERVATION
OF NILI RAVI BUFFALO BULL (Bubalus
bubalis) SEMEN
By
FAZAL WADOOD
2007-VA-557
A thesis submitted in the partial fulfillment of the requirement for the
degree
Of
DOCTOR OF PHILOSOPHY
In
THERIOGENOLOGY
DEPARTMENT OF THERIOGENOLOGY
UNIVERSITY OF VETERINARY AND ANIMAL
SCIENCES LAHORE-PAKISTAN
2015

2
The Controller of Examinations,
University of Veterinary and Animal Sciences,
Lahore.
We the members of the Supervisory Committee, certify that the contents
and form of thesis submitted by Mr. Fazal Wadood have been found satisfactory and
recommend that it should be processed for the evaluation by the External Examiner for
the award of the degree.
SUPERVISORY COMMITTEE
CHAIRMAN ____________________________________
PROF. DR. MUHAMMAD ALEEM
CO-SUPERVISOR ____________________________________
DR. MUHAMMAD YOUNAS
MEMBER ____________________________________
PROF. DR. NASIM AHMAD
MEMBER ____________________________________
PROF. DR. IJAZ AHMAD

3
IN THE NAME OF
ALLAH,
THE COMPASSIONATE,
THE MERCIFUL

4
ACKNOWLEDGEMENTS
I situate my sincere and humble thanks before Almighty Allah, who created the
universe and bestowed the mankind with knowledge and wisdom to search for its secrets.
I wish to express sincere gratitude to my hardworking, responsive and
praiseworthy supervisor Prof. Dr. Muhammad Aleem, Department of Theriogenology,
University of Veterinary and Animal Sciences, Lahore for his affectionate supervision,
demonstrative guidance, sympathetic behavior and great help not only in
accomplishment of present study but also in every aspect of my life.
I am also grateful to the members of my supervisory committee, Dr. Muhammad
Younas, Deputy Director, Semen Production Unit, Qadirabad, Sahiwal, Prof. Dr. Nasim
Ahmad, Dean, Faculty of Veterinary Sciences, University of Veterinary and Animal
Sciences, Lahore and Prof. Dr. Ijaz Ahmad, Dean, Faculty of Bio Sciences, University of
Veterinary and Animal Sciences, Lahore for their skillful guidance and critical insight.
Special thanks are extended to Dr. Muhammad Shahbaz Yousuf, Dr. Dawar
Hameed,Dr. Amjad Riaz, Dr. Arshad Javed and Dr. Sajid Iqbal and their team members,
all my friends especially members of the PhD club, colleagues and Lab. staff including
Mr. Abdul Razzaq and Hafiz Muhammad Adnan for their wishes and cordial cooperation
during the studies. Lastly, I wish to thank my family especially my father, mother, wife, in
laws, brothers and sisters who always pray for my success. They taught, love and
supported me to achieve higher goals in life. Their concern in me can never be fully
returned but will always be remembered.
This pioneering study on cryopreservation of buffalo bull semen would have not
been possible without the financial assistance of the Higher Education Commission of
Pakistan, the monetary support provided by this funding agency is gratefully
acknowledged.
Fazal Wadood

5
TABLE OF CONTENTS
Sr. No. Title Page No.
1 Acknowledgements I
2 List of Tables Iv
3 List of Figures V
CHAPTERS I. INTRODUCTION 1
Objectives 4
II. REVIEW OF LITERATURE 5
Semen cryopreservation 5
Extenders for semen cryopreservation 6
Osmotic pressure and semen 7
Reactive oxygen species (ROS) 8
Ros production 9
Oxidative stress 9
Antioxidants in semen 10
Butylated hydroxy toluene (BHT)
L-cystein
Fertility rate with cryopreserved semen
Factors affecting fertility rate
Fertility rate with different extenders
Fertility rate with antioxidants supplemented extenders
11
11
12
12
12
13
III. MATERIALS AND METHODS 14
Experimental design
Experimental stations
Management of experimental animal
EXPERIMENT-I
14
14
15
15
Extenders evaluated
Schematic diagram of experiment- i
Preparation of stock solutions
15
16
17
Tris citric acid extender (TCAE) stock solution 17
Skim milk extender (SME) stock solution 17
Coconut water extender (CWE) stock solution
Maintenance of working solutions
Storage of working solutions
18
19
20
Extender’s preparation
Semen collection and evaluation
20
21
Spermatozoa motility 21
Spermatozoa concentration 21
Semen dilution 22
The 3-(4, 5-dimethylthiazol-2-yl) -2, 5-
diphenyltetrazolium bromide (MTT) reduction assay
22
Straws filling 23
Cryopreservation 23
Post thaw evaluation 23

6
Spermatozoa motility 23
Spermatozoa viability 24
Spermatozoa acrosomal integrity 24
Spermatozoa plasma membrane integrity 25
Spermatozoa DNA damage 25
Thiobarbituric acid assay (lipid peroxidation) 26
Experiment-II 28
Stock’s solution preparation and storage 28
Preparation of antioxidants stock solutions
Semen collection and initial evaluation
Extender’s preparation
Semen extension and processing
Post thaw evaluation
28
28
29
29
30
Experiment-III 30
Semen doses 30
Trial doses 30
Control doses 30
Inseminations 31
Pregnancy rate 32
Statistical analysis 32
IV. RESULTS 33
Experiment-I 33
Post-thawed semen characteristics 33
Spermatozoa motility 33
Spermatozoa viability 34
Spermatozoa acrosomal integrity 35
Spermatozoa plasma membrane integrity 37
Spermatozoa DNA Damage 38
Lipid peroxidation 40
Experiment-II 41
Post-thawed semen characteristics 42
Spermatozoa motility 42
Spermatozoa viability 43
Spermatozoa acrosomal integrity 43
Spermatozoa plasma membrane integrity 44
Spermatozoa DNA damage 45
Lipid peroxidation 47
Experiment-III 48
V DISCUSSION 52
VI SUMMARY 65
VII LITERATURE CITED 67

7
LIST OF TABLES
Table No. Title Page No.
1. Suitable osmotic pressure for semen extenders in different
species 8
2. Mean seminal parameters of fresh ejaculates and pooled
buffalo semen (n=10) 33
3. Mean post thawed motility (%) of buffalo spermatozoa at
different osmotic pressures of various extenders (n=5) 34
4. Mean post thawed viability (%) of buffalo spermatozoa at
different osmotic pressures of various extenders (n=5) 35
5.
Mean post thawed normal apical ridge (%) of buffalo
spermatozoa at different osmotic pressures of various extenders
(n=5)
36
6.
Mean post thawed plasma membrane integrity (%) of buffalo
spermatozoa at different osmotic pressures of various extenders
(n=5)
38
7. Mean post thawed damaged DNA (%) of buffalo spermatozoa
at different osmotic pressures of various extenders (n=5) 40
8. Mean post thawed oxidative status (nm) of buffalo spermatozoa
at different osmotic pressures of various extenders (n=5) 41
9. Antioxidant’s effect on mean post thawed motility (%) of
buffalo spermatozoa (n=5) 42
10. Antioxidant’s effect on mean post thawed viability (%) of
buffalo spermatozoa (n=5) 43
11. Antioxidant’s effect on mean post thawed normal apical ridge
(%) of buffalo spermatozoa (n=5) 44
12. Antioxidant’s effect on mean post thawed plasma membrane
integrity (%) of buffalo spermatozoa (n=5) 45
13. Antioxidant’s effect on mean post thawed damaged DNA (%)
of buffalo spermatozoa (n=5) 46
14. Antioxidant’s effect on mean post thawed oxidative status (nm)
of buffalo spermatozoa (n=5) 48
15. Effect of milk production on pregnancy (%) of trial and control
doses 50
16. Effect of buffalo parity on pregnancy (%) of trial and control
doses 50
17. Effect of post partum interval on pregnancy (%) of trial and control
doses 51

8
LIST OF FIGURES
Figure No. Title Page No.
1. Standard curve of the stock solution (TMP) 27
2. Comparative effect of trial and control doses on pregnancy rate 49

9
Chapter I
INTRODUCTION
Total world buffalo population is 177.247 million and 97% buffaloes are found in
Asia (Presicce, 2007). Pakistan has 31.7 million buffalo population which is a major
proportion to world and Asian population of this species (Anonymous, 2010-2011).
Buffalo contributes 68% of total milk produced in the country (Bilal et al., 2006).
With the increase in human population and their demand for animal products
there is a need to increase per animal milk production. Artificial insemination (A.I.) with
cryopreserved semen is the most viable biotechnology to increase milk production of
buffalo population as we can disseminate elite bull genome to next progeny.
The basic purpose of semen extender was to extend the volume of semen and to
maintain the semen fertility during cryopreservation. Buffalo spermatozoa are more
susceptible to damage than cattle spermatozoa during freezing (Raizada et al., 1990). It is
therefore, essential to develop a suitable extender for buffalo semen for viable conception
rates.
Up to 50% spermatozoa livability is dropped during semen cryopreservation
(Watson, 2000). Following three factors contributes in damage during cryopreservation
(1) osmotic stress (Watson, 2000) (2) oxidative stress (Agarwal et al., 2003) and (3) cold-
shock (Watson, 2000). Consequently, the objective of spermatozoa-freezing is to prevent
lethal intracellular ice crystal formation to reduce membrane damage due to osmotic
shock during and after cryopreservation (Amirat-Briand et al., 2004).
During cryopreservation, the spermatozoa is exposed to sudden changes in
osmotic pressure which leads to variation in solute concentration and results in formation
1

10
of intra and extra cellular ice crystals that causes irreversible damage to its integrity
(Jeyendran et al., 1984; Hammerstedt et al., 1990). This spermatozoa damage due to
osmotic changes during cryopreservation can be minimized by developing an iso-osmotic
semen extender (Blackshah and Emman, 1951). Semen extenders for buffalo semen
cryopreservation ignore the osmolality requirements of buffalo whole semen (~269
mOsm/L; Khan and Ijaz, 2008). Preservation of buffalo semen in such an extender may
expose spermatozoa to osmotic stress (Mazur, 1980). Thus, along with other semen
quality parameters, osmotic pressure of semen diluents play key role during semen
cryopreservation.
Oxidants (reactive oxygen species) level is another factor that can affect semen
quality (Thuwanut, 2007). Lower reactive oxygen species (ROS) concentrations is
advantageous for spermatozoa acrosome reaction and capacitation in female reproductive
tract (de Lamirande and Gagnon, 1993). Studies revealed that oxidative stress can cause
apoptosis, reduced spermatozoa motility, DNA and spermatozoa membrane damage and
spermatozoa protein denaturation in human, bull, ram and stallion respectively (Aitken,
1999; Nair et al., 2006 and Filho, 2009).
It is observed that addition of antioxidants to semen reduces oxidative stress and
improves post thaw semen quality. Cryopreservation reduces the natural antioxidants
(enzymatic and non enzymatic) in buffalo semen because these antioxidants oxidizes
themselves and neutralize the produced oxidants, the remaining ones antioxidants are so
much diluted by the semen extender that they cannot protect the spermatozoa from
oxidative stress. Moreover, buffalo have higher lipid oxidation rate due to higher
membrane contents of poly unsaturated fatty acids and reduced activity and quantity of

11
naturally present antioxidants. Therefore, external antioxidants addition in semen is
necessary during cryopreservation to protect the spermatozoa integrity (Andrabi et al.,
2008a).
Antioxidant, L-Cysteine (one form of cysteine) is a amino acid (propionic acid),
included in thiols group of antioxidants (Van Zandwijk, 1995) and is a precursor of
glutathione (GSH) which play role in hydrogen peroxide (H2O2) oxidation (Meister,
1994). That is why; L- cysteine can prevent DNA damage during cryopreservation. L-
cysteine addition to semen extender prevents apoptosis, spermatozoa motility loss, lipid
peroxidation during cryopreservation (Erkkila et al., 1998; Bilodeau et al., 2001).
Butylated hydroxytoluene (BHT), a phenolic organic compound that is a synthetic
analogue of vitamin E. BHT significantly decreases the membrane permeability changes
during cryopreservation by acting as membrane protectant (Khalifa et al., 2008).
Low conception rate in buffaloes with cryopreserved semen may be due to
acrosomal damage (Akhtar and Chaudry, 1989), reduced spermatozoa motility (Tuli et al.
1981; Budworth et al., 1988) and alterations in spermatozoa membrane integrity (Rasul et
al., 2001) during the process of cryopreservation. So, the need exists to minimize all
these damages during cryopreservation by developing a suitable semen extender for
buffalo semen cryopreservation. By keeping in view all the above mentioned issues, this
study was designed with following objectives.

12
Objectives
The objectives of this study are:
To select best osmotic pressure for various buffalo bull semen extenders
To determine effects of various antioxidants on post thaw semen quality
To develop a best semen extender for cryopreservation of buffalo semen

13
Chapter II
REVIEW OF LITERATURE
Artificial insemination (A.I.) with cryopreserved semen is the most viable
biotechnology to increase milk production of buffalo. Buffalo spermatozoa are more
susceptible to damage than cattle during cryopreservation (Raizada et al., 1990). It is
therefore, essential to develop a suitable extender for buffalo semen cryopreservation for
viable conception rates.
2.1. Semen Cryopreservation:
Cryopreserved semen generally has impaired fertility compared to fresh semen
because of the fact that 40-50 % spermatozoa do not survive cryopreservation (Watson,
2000). Moreover, cryopreservation reduces number of motile spermatozoa and lowers the
chances of spermatozoa to reach at site of fertilization. Capacitation like changes
(increased intracellular calcium and acrosome reaction) do take place during
cryopreservation and that impairs fertility, too (Bailey et al., 2000). Cryopreservation
primarily damaged the spermatozoa plasma membrane (Yildiz et al., 2007), which
ultimately lowers fertility.
The plasma membrane is mainly composed of phospholipids, neutral lipids, and
glycolipids and its composition vary in different species (Flesch and Gadella, 2000), and
a higher cholesterol to phospholipids ratio in plasma membrane offer more resistance to
cold shock.
Buffalo spermatozoa have higher ratio of phospholipids in its membrane
compared to cattle, i.e. phosphatidyl choline content of plasma membrane is 66% of total
phospholipids in buffalo (Cheshmedjieva and Dimov, 1994) compared to plasma
5

14
membrane of cattle spermatozoa that contains only 50% phospholipids (Parks et al.,
1987). Similarly, phosphatidyl ethanolamine ratio in buffalo and cattle bull spermatozoa
plasma membrane has been reported as 23% and 10%, respectively (Parks et al., 1987;
Cheshmedjieva and Dimov, 1994). Variation in plasma membrane profile is one of the
reasons for poor freezability of buffalo semen compared to cattle (Tatham, 2000).
2.2. Extenders for Semen Cryopreservation:
Extenders for semen cryopreservation have vital role on post thawed semen
quality. Previously, extenders of different compositions i.e. sodium citrate, tris, citric acid
whey, lactose (Chinnaiya and Ganguli, 1980; Heuer, 1980; Matharoo and Singh, 1980;
Tuli et al., 1981), skim milk (Kakar and Anand., 1981), zwitterion buffers (TCA, Tes,
Hepes, Bes, Mes, Mops, Pipes and Tricine; Rasul et al. (2000) , Cornell university,
Illinoise variable temperature, Minnesotta university (Hashemi et al., 2007) and coconut
water (Vale et al., 1997) extenders having egg yolk were used for the cryopreservation of
buffalo semen.
Buffalo spermatozoa quality was higher in tris citric acid egg yolk extender
compared to Cornell university, Illinois variable and Minnesota extenders (Hashemi et al.
2007). Likewise, Rasul et al. (2000) found tris citric acid egg yolk extender a suitable
extender to improve post-thaw semen quality compared to citrate, HEPEST and TEST
extenders. Buffalo semen quality was comparable for tris citric acid egg yolk and bioxcell
(contains soy lecithin instead of egg yolk) extenders (Akhtar et al., 2010).
In another comparison, buffalo spermatozoa quality parameters in liquid stored
semen were higher in bioxcell, milk and tris citric acid extenders compared to citrate
(Akhtar et al., 2011). Ari et al. (2011) compared both pure and processed milk of cow

15
and goat milk as extender and found goat milk based extender as an effective semen
dilution media for ram semen cryopreservation.
Viveiros et al. (2008) efficiently used reconstituted powdered coconut with
methyl glycol for fish semen cryopreservation. Lower agouti spermatozoa quality was
noted in pasteurized and unpasteurized coconut water compared to sterilized skim milk
based extenders (Mollineau et al., 2011).
2.3. Osmotic Pressure and Semen:
During cryopreservation osmotic pressure changes that causes osmotic stress
which may leads to spermatozoa damage (Jeyendran et al., 1984; Hammerstedt et al.,
1990; Watson, 2000). The spermatozoa undergo sustained swelling or shrinkage when
exposed to changes in osmotic pressure (Mazur, 1984; Hammerstedt et al., 1990; Du et
al., 1994; Gilmore et al., 1996). Phenomenon of cell swelling or shrinkage is regulated by
simple diffusion and ionic pumps (Darnell et al., 1986; Padan and Schuldiner, 1993;
Lingrel and Kuntzweiler, 1994).
There are controversial reports about osmotic pressure of buffalo semen. Ibrahim
et al. (1985) documented an osmotic pressure of 293.33±3.39 mOsm/kg, whereas, Khan
and Ijaz (2008) reported 268.8 1.± 1.17 mOsm/kg. The highest metabolic activity of
spermatozoa was noted when semen was extended in an iso-osmotic extender. Most of
the buffalo semen extenders ignore the osmolality requirements of the specie (Khan and
Ijaz, 2008). Preservation of buffalo semen in such extenders may expose spermatozoa to
osmotic stress, which not only affects the structure and function of spermatozoa
mitochondria, nucleus, flagella, cell signaling and plasma membrane but also leads to
spermatozoa death (Mazur, 1980; Latif et al., 2005; Meyers, 2005).
16
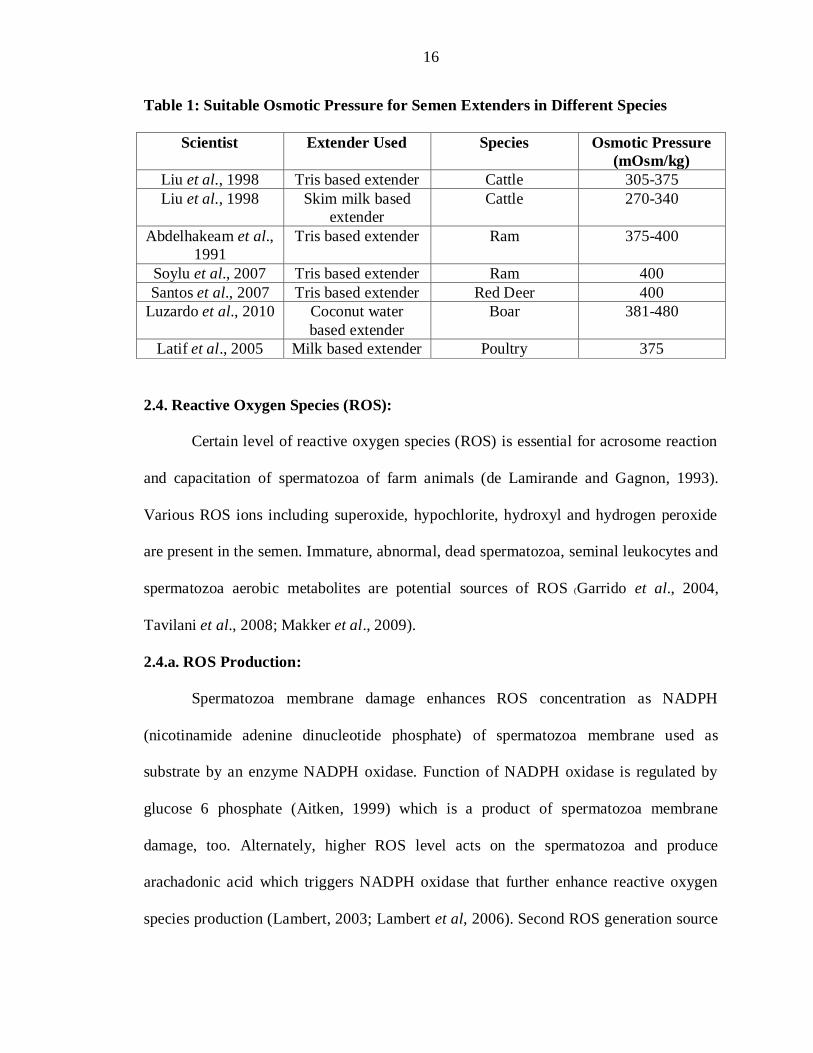
Table 1: Suitable Osmotic Pressure for Semen Extenders in Different Species
Scientist Extender Used Species Osmotic Pressure
(mOsm/kg)
Liu et al., 1998 Tris based extender Cattle 305-375
Liu et al., 1998 Skim milk based
extender
Cattle 270-340
Abdelhakeam et al.,
1991
Tris based extender Ram 375-400
Soylu et al., 2007 Tris based extender Ram 400
Santos et al., 2007 Tris based extender Red Deer 400
Luzardo et al., 2010 Coconut water
based extender
Boar 381-480
Latif et al., 2005 Milk based extender Poultry 375
2.4. Reactive Oxygen Species (ROS):
Certain level of reactive oxygen species (ROS) is essential for acrosome reaction
and capacitation of spermatozoa of farm animals (de Lamirande and Gagnon, 1993).
Various ROS ions including superoxide, hypochlorite, hydroxyl and hydrogen peroxide
are present in the semen. Immature, abnormal, dead spermatozoa, seminal leukocytes and
spermatozoa aerobic metabolites are potential sources of ROS (Garrido et al., 2004,
Tavilani et al., 2008; Makker et al., 2009).
2.4.a. ROS Production:
Spermatozoa membrane damage enhances ROS concentration as NADPH
(nicotinamide adenine dinucleotide phosphate) of spermatozoa membrane used as
substrate by an enzyme NADPH oxidase. Function of NADPH oxidase is regulated by
glucose 6 phosphate (Aitken, 1999) which is a product of spermatozoa membrane
damage, too. Alternately, higher ROS level acts on the spermatozoa and produce
arachadonic acid which triggers NADPH oxidase that further enhance reactive oxygen
species production (Lambert, 2003; Lambert et al, 2006). Second ROS generation source

17
is the spermatozoa mitochondria which are regulated by the NADPH oxidoreductase and
that is regulated by glucose 6 phosphate dehydrogenase (Makker et al., 2009).
2.4.b. Oxidative Stress:
Oxidative stress is one of the factors that can adversely affect the semen quality
(Thuwanut, 2007). Oxidative stress damage lipids, sugars, nucleic acids and proteins of
spermatozoa (Chatterjee and Gangon, 2001) and is responsible for spermatozoa
apoptosis, changes in plasma membrane integrity and decrease spermatozoa motility in
human, stallion, bull and ram, respectively (Aitken, 1999; Baumber et al., 2000; Nair et
al., 2006; Peris et al., 2007). Higher content of poly unsaturated fatty acids (PUFA) in
spermatozoa plasma membrane (compared to somatic cell membrane) increases the
membrane fluidity of membrane (Sikka, 2004). Oxidation of plasma membrane decrease
membrane fluidity. This loss of membrane fluidity impairs the function of membrane
ATPases which ultimately cause spermatozoa damage as ATPases regulate the entry of
nutrients/ions into the spermatozoa (Aitken and Clarkson, 1987; Ernster, 1993).
Secondly, oxidative stress causes lipid peroxidation of protein’s thiols of spermatozoa
plasma membrane and makes the spematozoa susceptible to macrophages attack (Alvarez
and Storey, 1989). Thirdly, oxidation process causes DNA modifications, chromosomal
rearrangements and breakage of single and double stranded DNA (Kemal et al., 2000).
Oxidative stress also influences adenosine triphosphate production by affecting the
tyrosine phosphorylation cycle (Sikka, 2004). Oxidative stress adversely affects both
spermatozoa function and fertilizing potential of the semen.
2.5. Antioxidants in Semen:

18
Buffalo semen contains both natural enzymatic (glutathion peroxidase, glutathion
reductase, superoxide dismutase and catalase etc.) and non enzymatic (cysteine, Vitamin
C, E and glutathion etc.) antioxidants. These antioxidants protect the spermatozoa from
damages during cryopreservation (Ansari et al., 2011a) by scavenging the reactive
oxygen species (Sikka, 2004).
Buffalo semen is more susceptible to oxidative stress as it has higher membrane
contents of poly unsaturated fatty acid (Chatterjee and Gagnon, 2001). Moreover, semen
extension and cryopreservation reduce the natural antioxidants in semen to such an extent
that they cannot protect the spermatozoa from oxidative stress. Therefore, external
antioxidants addition in semen is necessary during cryopreservation to protect the
spermatozoa integrity (Andrabi et al., 2008a).
Different antioxidants i.e. cysteine, butylated hydroxy toluene (BHT), trehalose,
taurine, tocopherol, ascorbic acid, glutathione etc. (Andrabi et al., 2008a; Ijaz et al.,
2009; Ansari et al., 2010; Reddy et al., 2010; Ansari et al., 2011a; Beheshti et al., 2011)
have been used for buffalo semen cryopreservation. In present study, BHT and L-cysteine
were also added to study their effect on post thawed buffalo semen quality.
2.5.f. Butylated Hydroxy Toluene (BHT):
Significantly improved buffalo semen quality was reported in extended semen
having BHT (1.5 mM) inclusion at room and refrigerated temperature (Pankaj et al.,
2009). Similarly, buffalo post thawed semen quality parameters were significantly higher
at BHT 1.5 mM concentration (Munir, 2011). Ijaz et al. (2009) reported significant
improvement in post thawed semen quality at addition of 2.0 mM concentration of BHT.
2.5.g. L-Cysteine:

19
In the literature, variable levels of Cysteine are reported to improve post thaw
semen quality. In one study, significantly higher buffalo post thawed semen quality was
observed in extender having 1.0mM cysteine (Ansari et al., 2011a). In another
experiment, significantly higher post thawed buffalo spermatozoa quality parameters
were noted in extender having 7.5 mM cysteine (Beheshti et al., 2011). Significantly
positive effect of cysteine (5mM) on post-thawed buffalo semen quality was observed by
El-Sheshtawy et al. (2008).
2.6. Fertility Rate with Cryopreserved Semen:
Conventional parameters used for evaluation of semen have limited application
because they only help to assess the structural integrity of the spermatozoa, but fertility
rate is the most appropriate means to evaluate the quality of post thaw buffalo semen
(Vale, 1997). Reported conception rate after insemination with frozen thawed semen in
buffalo is about 33% (Chohan et al., 1992; Bhosrekar et al., 2001) which is lower than
cattle.
2.6. Factors Affecting Fertility Rate:
Water buffalo is known as seasonal breeder, and is not much sexually active in
summer as compared to winter (Tahir et al., 1981). Second factor is nutrition that is vital
issue upsetting reproductive performance and fertility in ruminants (Zarazaga et al.,
2005; Anzar et al., 2003). Fertility in buffalo may also be affected due to weak
behavioural signs, inexperienced inseminators and silent ovulation (Vale, 1997; Qureshi,
1998). Whereas, low fertility rates could also be related to wrong side uterine

20
insemination due to smaller size of uterus (Zicarelli et al., 1997b). Fertility rate is also
affected by: the presence of vasectomised bull (Zicarelli et al., 1997a) as estrus detection
got improved, environmental factors, size of the semen straw (Haranath et al., 1990) and
type of semen extender used (Dhami and Kodagali, 1990).
2.6.a. Fertility Rate with Different Extenders:
Pregnancy rate more than 50% in buffaloes with cryopreserved semen would be
considered good (Vale et al., 1997). Pregnancy rates of cryopreserved semen in tris,
citrate and lactose based extenders were 42.7%, 39.8% and 37.5%, respectively (Dhami
and Kodagali, 1990). Similar fertility rates of bioxcell and tris citric acid extenders (44%
vs. 47%) were observed, respectively by Akhtar et al. (2010). Whereas, first service
conception rate (45.85%) of frozen semen diluted in tris based extender was noted in
buffaloes by Singh et al. (1980). In another similar experiment, Heuer et al. (1987) noted
no significant difference regarding pregnancy rates of lactose, skim milk and tris
extenders in buffaloes.
2.9.b. Fertility Rate with Antioxidants Supplemented Extenders:
Fertility was significantly improved in does when inseminated with tris extender
having 5 mM BHT as compared to extender with 0.0 mM BHT, respectively (Khalifa et
al., 2008). Whereas, no BHT inclusion outcome on oocyte cleavage rate was observed,
but blastocyst development was significantly enhanced from spermatozoa having 0.4 mM
BHT inclusion (Roca et al., 2004).
The difference regarding fertility rate for glutathione, cysteine and control group
in cows was not significant (Tuncer et al., 2010). Similarly, non-return rates was not
significant for taurine, cysteine & control groups (Sariozkan et al., 2009).

21
Chapter III
MATERIALS AND METHODS
Experimental Design:
In experiment-I, semen extender with optimal osmotic pressure for buffalo bull
semen out of tris citric acid, skim milk and coconut water extenders was selected by
evaluating post thawed spermatozoa quality parameters. In experiment -II, best extender
chose from experiment-I with optimal osmotic pressure was improved by the addition of
antioxidants (Butylated Hydroxy Toluene (BHT) and L-cysteine). In experiment-III,
fertility trial comparison was carried out under field conditions by using best extender
with best antioxidant concentration (50 inseminations) of experiment-II (Trial group) and
50 inseminations (Control group) of semen extender presently used in Punjab, Pakistan
for buffalo semen cryopreservation by Semen Production Unit, Qadirabad, Pakistan.
3.1. Experimental Stations:
Buffalo bull semen was collected, processed and cryopreserved at Semen
Production Unit (SPU), Qadirabad, Pakistan. Post-thaw semen quality analysis was
carried out in post graduate laboratory, department of physiology, University of
Veterinary and Animal Sciences, Lahore. Fertility trial was conducted at artificial
insemination (A.I.) Centers; district Bahawal Nagar, Punjab, Pakistan.

22
3.2. Management of Experimental Animal:
Four healthy buffalo bulls (Nili-Ravi) of known fertility and comparable age (<
10 years) were used in the study. These bulls were housed individually in the North-
South directionally situated pens having sufficient cross ventilation and protection against
heat during summer and double open space than the covered space for sun bath in winter.
These breeding bulls were fed good quality seasonal fodder (Barseam) at 10 % of the
body weight. In addition, 2-3 kg concentrate (commercially prepared Wanda) was offered
daily. Drinking water was provided ad libitum. Each bull showered twice daily. Physical
exercise was practiced three times a week. Vaccination against Hemorrhagic Septicemia
(H.S), Foot and Mouth Disease (FMD) and Black Quarter was done routinely once a
year. Preventive measures against internal and external parasites was undertaken twice a
year or whenever felt necessary. Semen was collected using artificial vagina maintained
at 42˚C (Andrabi et al., 2007). One ejaculate was collected at weekly intervals for a
period of 5 weeks during winter months (November - January) for the first two trials of
this research. Semen samples with more than 70% motile spermatozoa were selected for
further processing.
EXPERIMENT-I:
(SELECTION OF SUITABLE EXTENDER WITH OPTIMAL OSMOTIC
PRESSURE)
3.3. Extenders Evaluated:
Following three different types of extenders were used in this study.
1. TRIS citric acid yolk glycerol extender (TCAE)
2. Skim milk yolk extender (SME)

23
3. Coconut water citrate yolk extender (CWE)
3.3.a. Schematic Diagram of Experiment- I:
Semen Collection
↓
Pooled Semen
(Equal volumes)
↓
Dilution either in TCAE, SME, CWE each with osmotic
pressures (260, 270, 280, 290 & 300 mOsm/kg)
↓
Semen Cooling & Cryopreservation
↓
Thawing for 30 sec (37oC)
↓
Post Thaw Evaluation
(% motility, %viability, %NAR, %HOST, %Damaged DNA & LPO)
↓
Selection of Single Extender with best osmotic pressure
LPO (lipid peroxidation)
NAR (normal apical ridge)
HOST (hypo osmotic swelling test)
3.4. Preparation of Stock Solutions:
Composition for these solutions is as under,

24
3.4.a. Tris Citric Acid Extender (TCAE) Stock Solution:
TCAE stock solution (500 ml) with following composition was prepared
Ingredient Quantity
Tris 16.78 gm
Citric Acid 9.32 gm
Fructose 7.53 gm
Above mentioned composition was derived after modification to the
formula of Liete et al. (2010).
Chemicals for TCAE stock solution were procured with following sources, Tris
(Hydroxymethyl) aminomethane, (BDH Laboratory supplies, England), citric acid
monohydrate (Merck, Germany) and D (-) Fructose (BDH Laboratory supplies, England).
3.4.b. Skim Milk Extender (SME) Stock solution:
SME stock solution (500 ml) with following composition was prepared
Ingredients Quantity
Skim Milk 55 gm
Above mentioned composition was derived after modification to the
formula of Kommisrud et al. (1996).
For TCAE and SME, after addition of above mentioned quantities of ingredients
in to a graduated cylinder, 80 ml double distilled water at 37 ˚C was added. Now,
these solutions were thoroughly mixed with the help of vortex mixer.
Now more double distilled water at 37 ˚C was added to these extender’s solutions
and made the final volume 500 ml.

25
After preparation SME stock solution was heated at 95 ˚C for 10 minutes. For
SME, skim milk (Skimmillac, Millac Foods (Pvt.), Ltd. Pakistan) was procured.
3.4.c. Coconut Water Extender (CWE) Stock Solution:
Coconut water extender (CWE) stock solution (500 ml) with following
composition was prepared
Ingredient Quantity
Coconut Water 250 ml
Tri sodium Citrate
Dihydrate 7.50 gm
Double distilled water 250 ml
Above mentioned composition was derived after modification to the
formula of Cardoso et al. (2003).
For CWE stock solution, fresh coconut were obtained from market (After
breaking coconut, water was filtered two times by using filter paper) and Tri-sodium
citrate dihydrate from (Merck, 64271 Darmstadt, Germany).
All the stock solutions were confirmed to have osmotic pressures > 300 mOsm/kg
by using cryoscopic osmometer (Gonotec 030, cryoscopic osmometer, Berlin,
Germany).
3.5. Maintenance of Working Solutions:
All three types of stock solutions were used to achieve osmotic pressures of 260,
270,280,290 and 300 mOsm/kg. This stock solution of each extender then further
subdivided into six parts. Then five subdivided parts of stock solutions were used to
prepare desired osmotic pressure levels for each extender and sixth subdivided part of

26
stock solution was spared for further use. Double distilled water was used for lowering
osmotic pressure of stock solution. To uplift osmotic pressure prepared stock solution
(spared subdivided part) of higher osmotic pressure was used. Three repeated
observations of each osmotic pressure level were taken to verify the accuracy of the
desired level. pH of the desired osmotic pressure solution was maintained at ~7.0. pH of
this solution was adjusted to ~7.0 by using pH meter (Neo Met, Istek, Korea). For pH
adjustment 1N Hcl (9 ml of 37% hydrochloric acid was taken in a glass cylinder and
mixed with bi distilled water until final volume of 100 ml was achieved, it was shaken
well to make uniform solution) and N/10 NaOH (prepared by dissolving 0.4 g of NaOH
in bi distilled water to achieve 100 ml volume. It was well mixed with vortex mixer to
make a uniform solution. Its pH was 13.22) was used. After pH adjustment again the
osmometer was used to estimate osmotic pressures. Mean of three observations was taken
as single point.

27
3.5.a. Storage Of Working Solutions:
After preparation 10 ml aliquots of working solutions of specific osmotic pressure
of each extender were stored in capped graduated test tubes and freeze at -40 ˚C in
biomedical freezer (MDF-U5411, Sanyo, Japan) until use. One day before of semen
collection these working solutions were shifted to SPU, Qadir Abad.
3.6. Extender’s Preparation:
One night before the day of semen collection, stored working solutions were
warmed in water bath maintained at 37 ˚C. Then these working solutions were
pasteurized at 65 ˚C for half an hour in water bath. After that the working solutions were
cooled down to 37 ˚C (Munir, 2011). One aliquots of specific osmotic pressure of each
extender was used to prepare extender by the addition of following ingredients. Hundred
mili litter of extender was prepared with following composition,
Ingredient Quantity
Working solution 73 ml
Egg Yolk 20 ml
Glycerol 7 ml
Streptomycin 1.0 mg/ml
Penicillin 1000 i.u./ml
Above mentioned chemicals were procured with following sources, glycerol
(Scharlab S.L. Spain), streptomycin (China National Medicine & Health Products
Chongqing, China) and penicillin (Sinochem, China).
3.7. Semen Collection and Evaluation:

28
All hygienic measures were made before semen collection from bulls. Semen was
collected at early morning by expert technicians. Before collecting the ejaculates false
mounts were given to the bulls. After taking the ejaculates from the bulls, semen was
immediately shifted to the water bath maintained at 37 ˚C. Preliminary semen assessment
e.g. color, motility and volume etc. were carried out.
3.7.a. Spermatozoa Motility:
Semen motility was assessed by using the phase contrast microscope with digital
stage warmer maintained at 37 ˚C (40X; Olympus attached with closed circuit television).
Suitable ejaculates (>70% motility) were further evaluated for spermatozoa concentration
by digital photometer (Bovine photometer n˚ 1119, IMV, France).
3.7.b. Spermatozoa Concentration:
3660ul, normal saline (0.9% Nacl, Merck) was taken in a cuvette and 40 ul
undiluted semen was added in it and then spermatozoa concentration was determined by
digital photometer. In this experiment, equal volume of each ejaculate from four bulls
was pooled to get handsome semen volume and to remove bull to bull variation. Pooled
semen concentration and dilution ratio @ 40 million motile spermatozoa/ ml were also
calculated.
3.7.c. Semen Dilution:
Calculated semen quantity was added to the graduated tubes of respective osmotic
pressure that were containing specific amount of each semen extender. After dilution
semen was immediately shifted from water bath to the cold cabinet. Diluted semen
cooled from 37 ˚C to 4 ˚C in cold cabinet in two hours. Then equilibration time of 4 hours
were given to the diluted semen at 4˚C (Rasul et al., 2001).

29
Pooled semen quality was also assessed by using standard parameters (motility,
eosin and nigrosin staining, hypo osmotic swelling test, normal acrosomal ridge and MTT
assay). Procedure of all these tests (except MTT assay) is described under in post thaw
semen evaluation. Procedure of MTT assay was described under.
3.7.c.1. MTT Reduction Assay:
This test was used at undiluted semen to detect metabolically active spermatozoa.
Procedure of test was as under.
MTT reduction assay was used to assess viability of metabolically active
spermatozoa in each concentration of the extenders as mentioned by Iqbal et al. (2010).
The pooled semen was diluted using a phosphate buffer solution (PBS) solution to attain
40×106 spermatozoa/ml. Micro-plate with 96-wells was used. This test was performed by
mixing 100 μl of semen sample and 10 μl of MTT stock solution (50 mg/10 ml of MTT
in PBS) in five wells of micro plate. MTT reduction rate was recorded immediately by
using a spectrophotometer (MS2 Reader, Titertec Plus-MS2 Reader, ICN Biomedical,
Basingstoke, UK) at a wavelength of 550 nm. Later on, this plate was incubated for an
hour at 37°C and again noted its reading. The MTT reduction rate of each sample was
computed by the average difference of pre incubated and post incubated readings.
3.7.d. Straws Filling:
Semen straws (0.5, IMV, France) with three different colors were specified for
each extender. Each color straws were further subdivided into five equal quantities. Each
quantity then further specified for specific level of osmotic pressure of extender. Then
these straws were labeled by using the straw printing machine (Domino- A200 Pin Point,
IMV Technologies, France). After that equilibrated semen stored at 4˚C was filled in

30
straws by using suction pump to achieve a spermatozoa concentration of 20×106 motile
spermatozoa/ straw, then sealed by using the poly venile powder (PVP).
3.7.e. Cryopreservation:
These straws were cooled from 4˚C to -15˚C @ 3˚C/min then from -15˚C to -80˚C
@ 10˚C/min. Now, these straws were placed 4 cm above the liquid nitrogen for 10
minutes by placing straws horizontally in straw grill. Then these straws were dipped into
liquid nitrogen. Now, these straws were shifted to University of Veterinary and Animal
Sciences, Lahore, Pakistan, where post thaw evaluation of these straws were carried out.
3.8. Post Thaw Evaluation:
Following tests were used to determine post thaw spermatozoa characteristics,
3.8.a.Spermatozoa Motility:
After removing from liquid nitrogen container semen straws were thawed for
thirty seconds in water bath maintained at 37 ˚C. A drop of semen was placed on pre
warmed glass slide placed on stage warmer maintained at 37˚C. Straws of each extender
were assessed for spermatozoa motility under phase contrast microscope at 20x. The
mean of three motility observations was taken as a single data point. Three straws of
same level for each collection were observed.
3.8.b. Spermatozoa Viability:
A small drop of thawed semen of each concentration of extenders was mixed with
a drop of Eosin and Nigrosin stain. This stain was prepared by dissolving 5% nigrosin
(w/v); 10172, Merck, Germany) and 1% eosin B (w/v); (1343, Merck, Germany) in to 3%
solution of (TSCD). After preparing and drying, smear was examined under phase
contrast microscope, for unstained heads of the spermatozoa was considered live and

31
stains or partially stained heads as dead. Two hundred spermatozoa were counted to
determine live and dead spermatozoa percentage (Khan and Ijaz, 2008). The mean of
three observations was considered a single data point.
3.8.c. Spermatozoa Acrosomal Integrity:
I used method adopted by Rasul et al. (2000) for this test, 500 µl of each thaw
semen sample of each extender was mixed in 50 µl of 1% formaldehyde citrate ( prepared
by mixing 99 ml of 2.9 % (w/v) sodium citrate and 1 ml of 37% formaldehyde). To
examine normal apical ridge two hundred spermatozoa were assessed under a phase-
contrast microscope. Abnormalities like absent, ruffled and swollen acrosome was noted.
The mean of three observations was considered a single data point.

32
3.8.d. Spermatozoa Plasma Membrane Integrity (PMI):
Hypo-osmotic swelling test (HOST) according to Adeel et al. (2009) was used to
examine PMI of each concentration of extenders. HOST Solution ( 0.735 g of (TSCD)
and 1.351 g D (-) fructose were dissolved in bi-distilled water and made final volume 100
ml) osmotic pressure was measured and decreased to 75mOsm/kg by adding drop by
drop of bi-distilled water. Then I mixed 500 µl of HOS solution with 50 µl of each
thawed semen sample and incubated at 37˚C for 45 min. After this 0.5 µl of sample from
the above incubated solution was examined under a phase-contrast microscope. Two
hundred spermatozoa were counted for the percentage of spermatozoa showing coiled
tail, an indicative of intact plasma membrane was recorded. The mean of three
observations were taken as a single data point.
3.8.e. Spermatozoa DNA Damage:
Initially DNA damage of spermatozoa was validated by using acridine orange
(AO) staining technique. This protocol allowed us to differentiate between spermatozoa
having double strand (intact) DNA or single strand (denatured) DNA. A drop of diluted
semen sample was placed on a clean glass slide and smear was made and air dried. Then,
this smear was dipped in Carnoy,s solution (three parts methanol + 1 part glacial acetic
acid) and slide was placed undisturbed for 2 hours, and after this smear was again air
dried. Smear was stained with freshly prepared AO stain (0.19mg/ml) (Scharlau, Chemie
S.A, Spain) for 5 minutes in the dark (AO stain was prepared by mixing 20 ml solution of
1% AO with 80 ml of 0.1 M citric acid solution and 5.0 ml of 0.2 M Na2 HPO4.7H2O.The
pH of the solution was adjusted to 2.5). After staining, distilled water was used to wash
the slides, dried it and observed at 100x under a fluorescent microscope (Labomed, Lx

33
400, U.S.A.). A total of 200 spermatozoa were assessed by the same examiner (Chohan et
al., 2006).
When monomeric acridine orange bind to double stranded DNA, it gave green
color and acridine orange gave red or yellow color when bind to a single strand DNA
under fluorescent microscope.
3.8.f. Thiobarbituric Acid Assay (Lipid Peroxidation):
Thiobarbituric acid assay was used to judge Malondialdehyde, a stable lipid
peroxidation product according to the method of Ohkawa et al. (1979). Thawing of
semen straws from each concentration of the extenders was done for 30 seconds at 37˚C.
In a sterilized test tube, 100 μl of semen sample was mixed with 200 µl of sodium
dodecyl sulphate (8.1 %), then added 1.5 ml solution of 20% acetic acid (pH = 3.5, pH of
this solution was adjusted by adding drop by drop of 10 molar solution of NaOH). Now, i
added 1.5 ml of aqueous solution of 0.8% thiobarbituric acid (TBA) followed by addition
of distilled water to build an ultimate volume of 4 ml. This solution was heated at 95˚C
for 60 minutes followed by cooling. Then 5 ml of solution B was added (15 parts of n-
butanol and 1 part pyridine) and shaken vigorously followed by centrifugation at 4000
rpm for 10 min. centrifugation resulted in the formation of an organic layer at the top of
the test tube and inorganic layer at the bottom. The top layer was taken and absorbance
was measured by 532 nm. A standard solution of TMP (tetramethoxypropane) in nano
mole (5-50 nm) was prepared. The results of spermatozoa thiobarbituric acid reactive
substance (TBARS) were expressed in nm MDA (malondialdehyde).
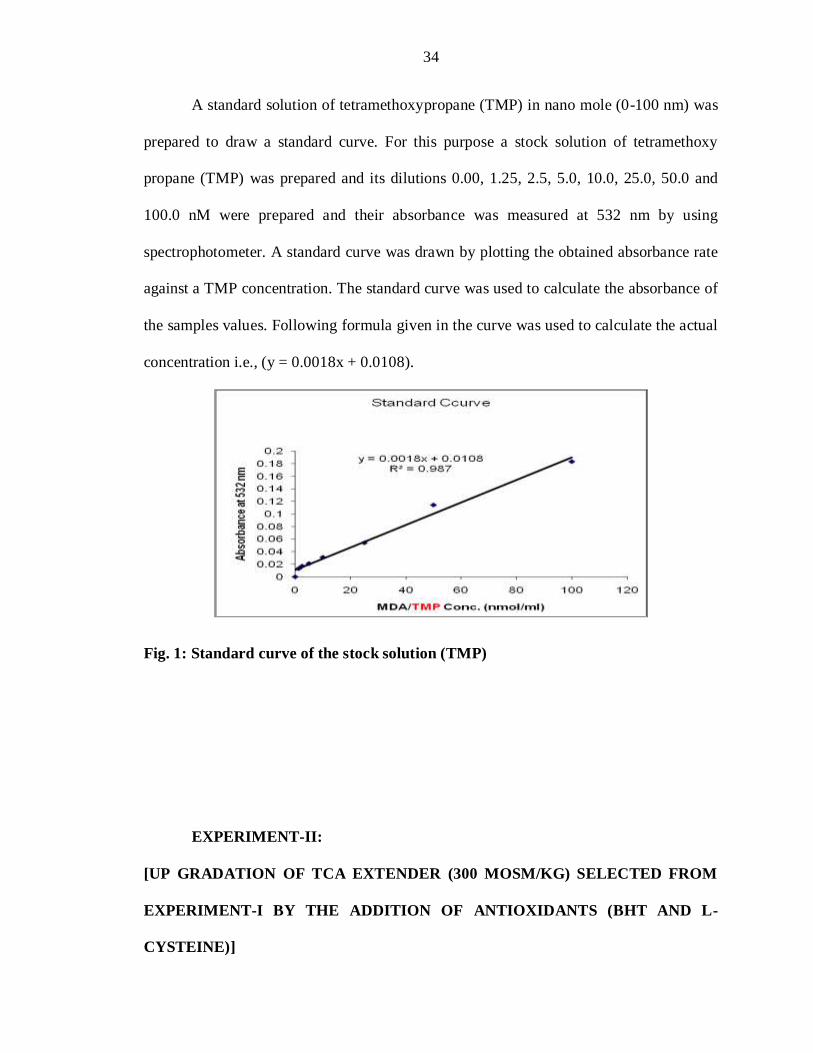
3.8.f.1. Standard curve of tetramethoxypropane (TMP):
34
A standard solution of tetramethoxypropane (TMP) in nano mole (0-100 nm) was
prepared to draw a standard curve. For this purpose a stock solution of tetramethoxy
propane (TMP) was prepared and its dilutions 0.00, 1.25, 2.5, 5.0, 10.0, 25.0, 50.0 and
100.0 nM were prepared and their absorbance was measured at 532 nm by using
spectrophotometer. A standard curve was drawn by plotting the obtained absorbance rate
against a TMP concentration. The standard curve was used to calculate the absorbance of
the samples values. Following formula given in the curve was used to calculate the actual
concentration i.e., (y = 0.0018x + 0.0108).
Fig. 1: Standard curve of the stock solution (TMP)
EXPERIMENT-II:
[UP GRADATION OF TCA EXTENDER (300 MOSM/KG) SELECTED FROM
EXPERIMENT-I BY THE ADDITION OF ANTIOXIDANTS (BHT AND L-
CYSTEINE)]

35
In experiment-I TCA extender at 300 mOsm/kg was evaluated best for post thaw
parameters. This extender was further up-graded by the addition of antioxidants in
current experiment.
3.9. Stock’s Solution Preparation and Storage:
Stock solution of TCAE at 300 mOsm/kg osmotic pressure was prepared (same
procedure as in experiment-I).
3.9.a. Preparation of Antioxidants Stock Solutions:
Due to BHT lipid solubility, it was dissolved in dimethyl sulph oxide (DMSO;
Su0151 Scharlau, Chemicals, Spain). A BHT stock solution of 500 mM was prepared by
mixing 2.2035 gram of BHT with DMSO and made the final volume 20 ml. L-cysteine
stock solution of 500 mM was prepared by mixing 1.2116 gram with double distilled
water and make the final volume 20 ml. BHT and L-cysteine solutions were made 48
hours before use and stored at -40 ˚C.
3.10. Semen Collection and Initial Evaluation:
Same as previously mentioned.
3.11. Extender’s Preparation:
Extender’s preparation was same as in experiment-I. Following concentrations of
antioxidants were achieved in TCA extender.
Butylated hydroxyl toluene (B1378, Sigma Aldrich, USA) Concentrations, 1.75, 2
and 2.25 mM were achieved in TCA extender.
L-cysteine (C7352, Sigma Aldrich, USA), 2, 5 and 8 mM concentrations were
also maintained in TCA extender.

36
As, dimethyl sulph oxide (DMSO) was also reported to have some antioxidant
properties (Donoghue and Donoghue, 1997), so, the DMSO volume equal to its volume
present in 2.25 mM BHT concentration was also run along with the BHT other
concentrations as DMSO group. Furthermore, along with these antioxidants
concentrations, a sample of TCA extender having no antioxidant was also run as negative
control.
3.12. Semen Extension and Processing:
Pooled semen from four bulls was mixed with TCAE having each antioxidant ’s
different concentrations and with negative control and DMSO group to achieve
spermatozoa concentration of 20×106 motile spermatozoa/ straw. Semen straws were
filled with diluted semen having above mentioned various antioxidants and then sealed
and cryopreserved in liquid nitrogen according to the procedure mentioned above. Single
best level of most suitable antioxidant was determined after post thaw evaluation.

37
3.13. Post thaw Evaluation:
Previously mentioned semen characteristics and assays of experiment-I were used
to assess post thaw semen quality in this experiment.
EXPERIMENT-III:
(FERTILITY RATE COMPARISON OF TRIAL AND CONTROL GROUPS)
In this experiment fertility rate comparison was done under field conditions by
using best extender with best antioxidant concentration (50 inseminations) of experiment-
II (Trial group) and 50 inseminations (Control group) of traditionally used semen
extender for buffalo semen cryopreservation by Semen Production Unit (SPU),
Qadirabad presently used for artificial insemination in Punjab, Pakistan.
3.14. Semen Doses:
3.14.a. Trial Doses:
Fifty buffaloes in heat were inseminated with the semen straws having TCAE
(osmotic pressure 300 mOsm/Kg and pH ~7.0), prepared with same procedure as in
experiment-I, supplemented with 2mM BHT (prepared and added in the same way as in
experiment-II) as concluded in experiment-II.
3.14.b. Control Doses:
Semen doses for Control group were prepared by using the conventionally used
semen extender of SPU, Qadirabad. At Semen Production Unit this extender was
prepared by dissolving 24.2 g of Tris (Hydroxymethyl) aminomethane (Research
Organics Inc, Ohio, U.S.A.), 13.4 g of citric acid monohydrate (Riedel-de Haën,
Germany) and 10 g of D (-) Fructose (Riedel-de Haën, Germany) and added distilled
water to made final volume 730 ml. Osmotic pressure of this stock solution was 312

38
mOsm/kg and pH ~6.8. This stock solution was pasteurized at 65 ˚C for half an hour and
then cooled down to 38 ˚C. Then egg yolk 200 ml, Glycerol (Riedel-de Haën, Germany)
70 ml, antibiotics 1 g streptopencillin (Sinobiotic, Shanxi Shuguang Pharmaceutical Co,
China) were added. This extender was mixed at 37 ˚C while stirring with magnetic stirrer
for 20 minutes. Then this extender was stored in a refrigerator overnight and warmed at
37 ˚C before use.
Trial and control semen doses were prepared under uniform conditions, following
same procedure of experiment-I. Both trial and control doses were prepared by the same
person. Single ejaculate of the same bull was used to prepare the control and trial doses at
the same time and uniform conditions.
3.15. Inseminations:
Buffaloes in heat were inseminated after thawing these doses at 37˚C for 30
seconds. Inseminations were made 12-24 hours after observed natural estrus. Animals
with repeat breeding history and having pathological problems of reproductive tract were
not inseminated. All inseminations were made by the expert artificial insemination
technicians working in district Bahawal Nagar, under the Livestock and Dairy
Development Department, Punjab, Pakistan. All the inseminations were made in low
breeding season (end of March to end of June). Effort was made that a single technician
should had equal number of control and trial inseminations. Technicians were also not
aware about the composition and efficiency of straws concluded on the base of previous
two experiments. Buffaloes were mainly reared by the small farmer, average holding of
buffaloes was from 2- 4. All the buffaloes were fed same fodder for the season (alfalfa
with wheat straw + concentrate), amount of concentrate was according to the milk

39
production. However, body score of buffaloes was variable. Owner name with cell
number, residential address and Animal’s feeding, milk production, post partum interval,
parity and age were recorded. Insemination date and inseminator name record was
maintained during this whole trial.
3.16. Pregnancy Rate:
Pregnancy rate was decided on the basis of rectal Palpation of all the inseminated
animals at day 60±10 post insemination.
3.17. Statistical Analysis:
Statistical analysis was conducted with the Statistical Package for Social Science
(SPSS for Windows version 12, SPSS Inc., Chicago, IL, USA). Data was presented as
mean ± S.E. The Kolmogorov Smirnov test was employed to test the normal distribution
of the data. The data was analyzed using analysis of variance. The group differences were
compared by the Duncan’s Multiple Range Test (Duncan, 1955). Pregnancy rates were
analyzed by using chi square. Difference in results was considered significant at P < 0.05.

40
Chapter IV
RESULTS
Semen volume, concentration and motility of each of the ten collections of
experiment-I and II from four bulls were noted and equal volume of ejaculates from each
bull was pooled to determine spermatozoa viability, normal apical ridge (NAR), plasma
membrane integrity (PMI) and MTT reduction rate (Table 2).
Table 2: Mean Seminal Parameters of Fresh Ejaculates and Pooled Buffalo Semen
(n=10)
Ejaculates Pooled semen
Volume
(ml)
Concentration
(million/ml)
Motility
(%)
Viability
(%) NAR (%) PMI (%)
MTT
reduction
rate
2.84±0.14 1226.43±71.48 77.13±0.71 90.05±0.71 94.67±0.30 86.23±0.34 0.290±0.06 n= no of collections Values are mean ± S.E.
EXPERIMENT-I:
Post-thaw Semen Characteristics:
Spermatozoa Motility:
At both 300 and 280 mOsm/kg TCAE spermatozoa motility was significantly
higher (P<0.05) compared to 260 mOsm/kg, however, spermatozoa motility at 270 and
290 mOsm/kg was similar when compared to all other osmotic pressures. In Skim Milk
Extender (SME), spermatozoa motility at 290 and 300 mOsm/kg was significantly higher
(P<0.05) (38.67±4.94 and 39.00±2.85%) compared to 260 mOsm/kg but there was no
difference in motility among other levels of osmotic pressures (Table 3).
41
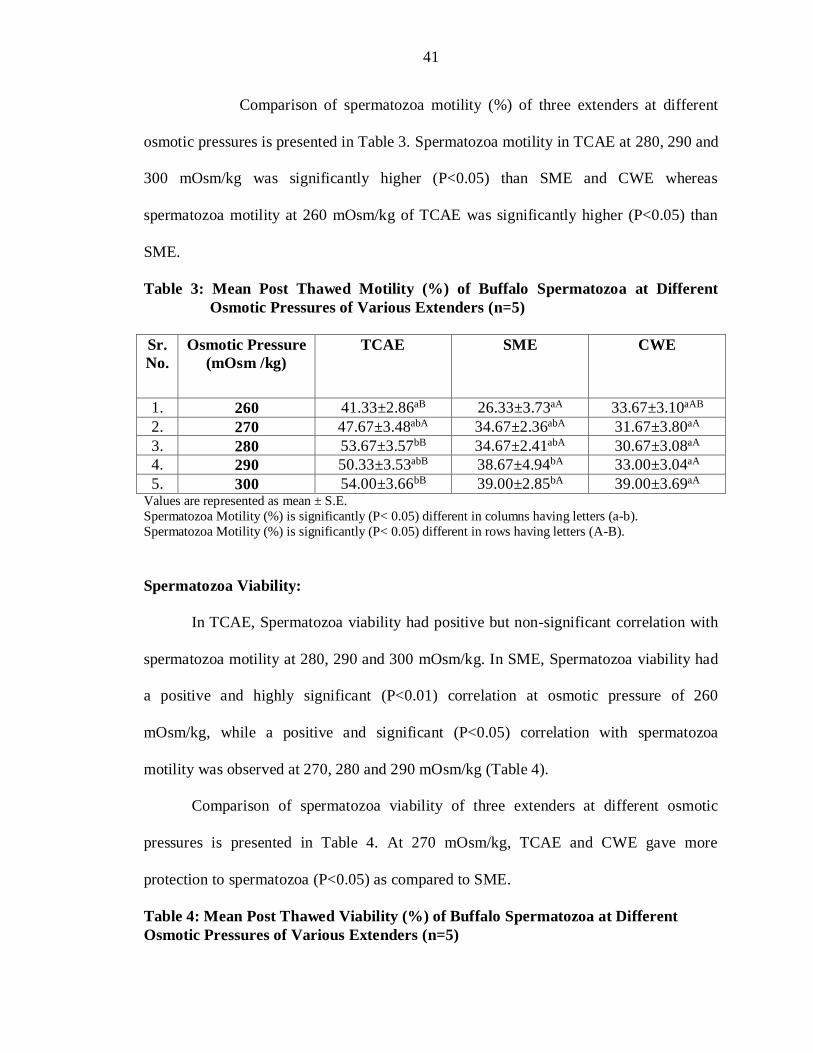
Comparison of spermatozoa motility (%) of three extenders at different
osmotic pressures is presented in Table 3. Spermatozoa motility in TCAE at 280, 290 and
300 mOsm/kg was significantly higher (P<0.05) than SME and CWE whereas
spermatozoa motility at 260 mOsm/kg of TCAE was significantly higher (P<0.05) than
SME.
Table 3: Mean Post Thawed Motility (%) of Buffalo Spermatozoa at Different
Osmotic Pressures of Various Extenders (n=5)
Sr.
No.
Osmotic Pressure
(mOsm /kg)
TCAE
SME
CWE
1. 260 41.33±2.86aB 26.33±3.73aA 33.67±3.10aAB
2. 270 47.67±3.48abA 34.67±2.36abA 31.67±3.80aA
3. 280 53.67±3.57bB 34.67±2.41abA 30.67±3.08aA
4. 290 50.33±3.53abB 38.67±4.94bA 33.00±3.04aA
5. 300 54.00±3.66bB 39.00±2.85bA 39.00±3.69aA Values are represented as mean ± S.E.
Spermatozoa Motility (%) is significantly (P< 0.05) different in columns having letters (a-b).
Spermatozoa Motility (%) is significantly (P< 0.05) different in rows having letters (A-B).
Spermatozoa Viability:
In TCAE, Spermatozoa viability had positive but non-significant correlation with
spermatozoa motility at 280, 290 and 300 mOsm/kg. In SME, Spermatozoa viability had
a positive and highly significant (P<0.01) correlation at osmotic pressure of 260
mOsm/kg, while a positive and significant (P<0.05) correlation with spermatozoa
motility was observed at 270, 280 and 290 mOsm/kg (Table 4).
Comparison of spermatozoa viability of three extenders at different osmotic
pressures is presented in Table 4. At 270 mOsm/kg, TCAE and CWE gave more
protection to spermatozoa (P<0.05) as compared to SME.
Table 4: Mean Post Thawed Viability (%) of Buffalo Spermatozoa at Different
Osmotic Pressures of Various Extenders (n=5)

42
Sr.
No.
Osmotic Pressure
(mOsm /kg)
TCAE
SME
CWE
1. 260 69.90±2.57aA 60.75±6.47aA 70.41±2.49aA
2. 270 72.22±2.36aB 62.11±4.41aA 74.17±1.99aB
3. 280 73.18±2.05aA 68.19±2.70aA 73.24±2.24aA
4. 290 72.63±1.91aA 70.59±2.96aA 72.31±2.32aA
5. 300 73.37±2.47aA 68.07±2.86aA 71.10±3.10aA Values are represented as mean ± S.E.
Means in columns are non-significantly (P> 0.05) different from one another.
Spermatozoa Live (%) is significantly (P< 0.05) different in rows having letters (A-B).
Spermatozoa Acrosomal Integrity:
In TCAE, acrosomal integrity at 300 mOsm/kg was significantly higher (P<0.05)
than other osmotic pressures. The highest acrosomal integrity (69.78±2.31%) was
recorded at 300 mOsm/kg, whereas, lowest was at 260 mOsm/kg (60.61±2.32%; Table
5). Correlation between spermatozoa acrosomal integrity and spermatozoa motility was
recorded as positive but significant (P<0.05) at 280 and 290 mOsm/kg.
In SME, acrosomal integrity at 300 mOsm/kg was higher (P<0.05) compared to
260, 280 and 290 mOsm/kg (table 5). Spermatozoa acrosomal integrity had a positive and
significant (P<0.05) correlation was recorded at 260 mOsm/kg. Positive and significant
(P<0.05) at 270 and 300 mOsm/kg, while highly significant (P<0.01) at 280 mOsm/kg. In
CWE, positive and significant (P<0.05) correlation was recorded at 260 mOsm/kg.
Correlation between spermatozoa acrosomal integrity and spermatozoa viability was
positive and significant (p<0.05) correlation was noted at 270 mOsm/kg.
Relative spermatozoa acrosomal integrity rate of three extenders at various
osmotic pressures is given in Table 5. Acrosomal integrity rate improved (P<0.05) in
CWE compared to TCAE at 290 mOsm/kg.
43
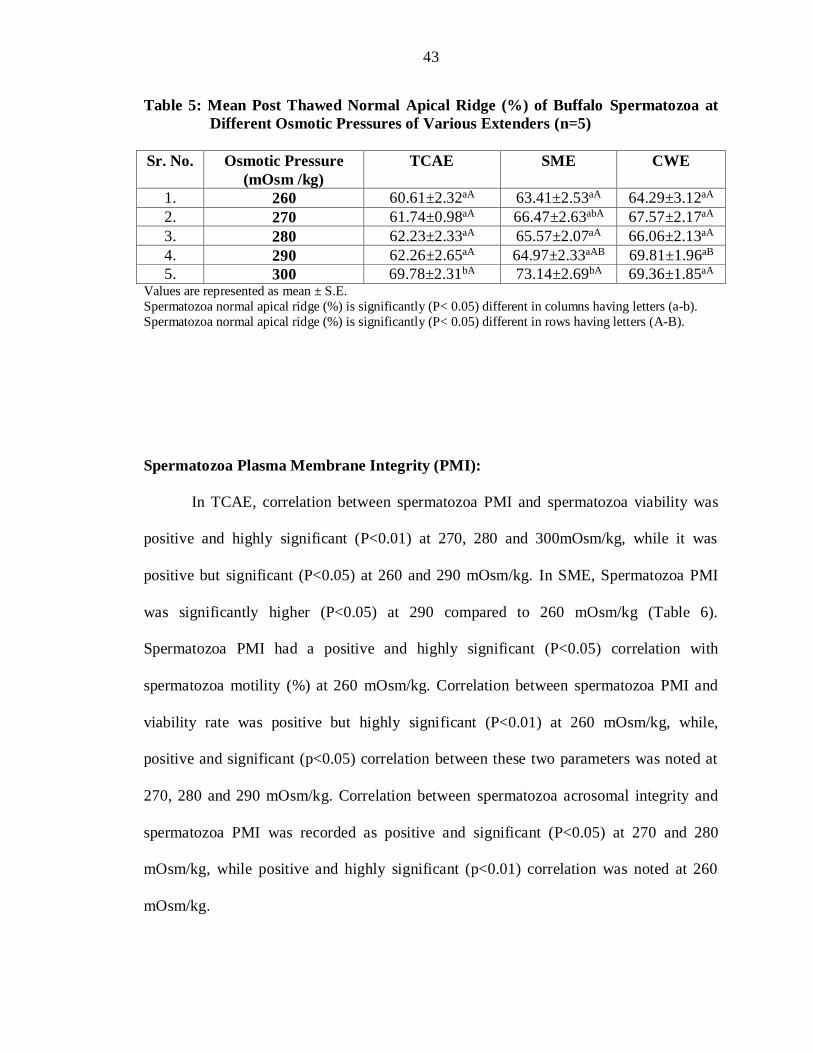
Table 5: Mean Post Thawed Normal Apical Ridge (%) of Buffalo Spermatozoa at
Different Osmotic Pressures of Various Extenders (n=5)
Sr. No. Osmotic Pressure
(mOsm /kg)
TCAE
SME
CWE
1. 260 60.61±2.32aA 63.41±2.53aA 64.29±3.12aA
2. 270 61.74±0.98aA 66.47±2.63abA 67.57±2.17aA
3. 280 62.23±2.33aA 65.57±2.07aA 66.06±2.13aA
4. 290 62.26±2.65aA 64.97±2.33aAB 69.81±1.96aB
5. 300 69.78±2.31bA 73.14±2.69bA 69.36±1.85aA Values are represented as mean ± S.E.
Spermatozoa normal apical ridge (%) is significantly (P< 0.05) different in columns having letters (a-b).
Spermatozoa normal apical ridge (%) is significantly (P< 0.05) different in rows having letters (A-B).
Spermatozoa Plasma Membrane Integrity (PMI):
In TCAE, correlation between spermatozoa PMI and spermatozoa viability was
positive and highly significant (P<0.01) at 270, 280 and 300mOsm/kg, while it was
positive but significant (P<0.05) at 260 and 290 mOsm/kg. In SME, Spermatozoa PMI
was significantly higher (P<0.05) at 290 compared to 260 mOsm/kg (Table 6).
Spermatozoa PMI had a positive and highly significant (P<0.05) correlation with
spermatozoa motility (%) at 260 mOsm/kg. Correlation between spermatozoa PMI and
viability rate was positive but highly significant (P<0.01) at 260 mOsm/kg, while,
positive and significant (p<0.05) correlation between these two parameters was noted at
270, 280 and 290 mOsm/kg. Correlation between spermatozoa acrosomal integrity and
spermatozoa PMI was recorded as positive and significant (P<0.05) at 270 and 280
mOsm/kg, while positive and highly significant (p<0.01) correlation was noted at 260
mOsm/kg.
44
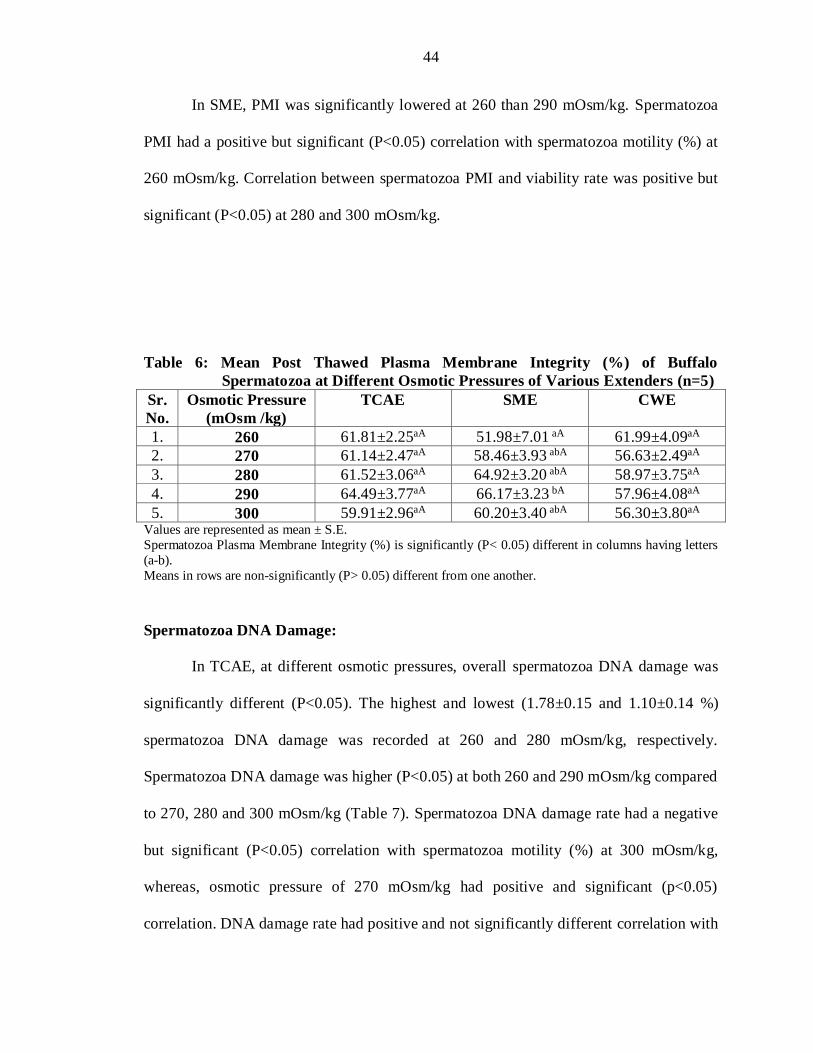
In SME, PMI was significantly lowered at 260 than 290 mOsm/kg. Spermatozoa
PMI had a positive but significant (P<0.05) correlation with spermatozoa motility (%) at
260 mOsm/kg. Correlation between spermatozoa PMI and viability rate was positive but
significant (P<0.05) at 280 and 300 mOsm/kg.
Table 6: Mean Post Thawed Plasma Membrane Integrity (%) of Buffalo
Spermatozoa at Different Osmotic Pressures of Various Extenders (n=5)
Sr.
No.
Osmotic Pressure
(mOsm /kg)
TCAE
SME
CWE
1. 260 61.81±2.25aA 51.98±7.01 aA 61.99±4.09aA
2. 270 61.14±2.47aA 58.46±3.93 abA 56.63±2.49aA
3. 280 61.52±3.06aA 64.92±3.20 abA 58.97±3.75aA
4. 290 64.49±3.77aA 66.17±3.23 bA 57.96±4.08aA
5. 300 59.91±2.96aA 60.20±3.40 abA 56.30±3.80aA Values are represented as mean ± S.E.
Spermatozoa Plasma Membrane Integrity (%) is significantly (P< 0.05) different in columns having letters
(a-b).
Means in rows are non-significantly (P> 0.05) different from one another.
Spermatozoa DNA Damage:
In TCAE, at different osmotic pressures, overall spermatozoa DNA damage was
significantly different (P<0.05). The highest and lowest (1.78±0.15 and 1.10±0.14 %)
spermatozoa DNA damage was recorded at 260 and 280 mOsm/kg, respectively.
Spermatozoa DNA damage was higher (P<0.05) at both 260 and 290 mOsm/kg compared
to 270, 280 and 300 mOsm/kg (Table 7). Spermatozoa DNA damage rate had a negative
but significant (P<0.05) correlation with spermatozoa motility (%) at 300 mOsm/kg,
whereas, osmotic pressure of 270 mOsm/kg had positive and significant (p<0.05)
correlation. DNA damage rate had positive and not significantly different correlation with

45
spermatozoa viability at 280 and 290 mOsm/kg. Spermatozoa DNA damage had positive
and significant (p<0.05) correlation with spermatozoa PMI at 290 mOsm/kg.
Overall spermatozoa DNA damage was significantly different at various osmotic
pressures of SME (P<0.05). The highest (1.45±0.1%) and lowest (0.78±0.11%)
spermatozoa DNA damage was noticed at 260 and 300 mOsm/kg. Significantly increased
(P<0.05) spermatozoa DNA damage was noted at 260 and 270 mOsm/kg compared to
290 and 300 mOsm/kg. Similarly, significant (P<0.05) damage to spermatozoa DNA
noted at 280 compared to 300 mOsm/kg osmotic pressure (Table 7). Spermatozoa DNA
damage rate had a positive but significant (P<0.05) correlation with spermatozoa motility
(%) at 280 mOsm/kg.
Overall effect of different osmotic pressures on spermatozoa DNA damage was
significant (P<0.05) in case of CWE. The highest (2.00±0.088%) and the lowest
(0.80±0.13%) spermatozoa DNA damage was observed at 260 and 290 mOsm/kg,
respectively. Spermatozoa DNA damage was significantly higher (P<0.05) at 260
mOsm/kg when compared to all other levels of osmotic pressures. Increased (P<0.05)
spermatozoa DNA damage observed at 280 mOsm/kg than 290 and 300 mOsm/kg (Table
7). A positive but not significantly different correlation with PMI rate was recorded at
270 280 and 290 mOsm/kg.
Comparison of spermatozoa DNA damage of three extenders at different osmotic
pressures is presented in Table 7. Spermatozoa DNA damage at 260 mOsm/kg
significantly increased (P<0.05) in TCAE and CWE than in SME. Spermatozoa DNA
damage in TCAE at 290 mOsm/kg was also significantly higher (P<0.05) than SME and
CWE (Table 7).
46
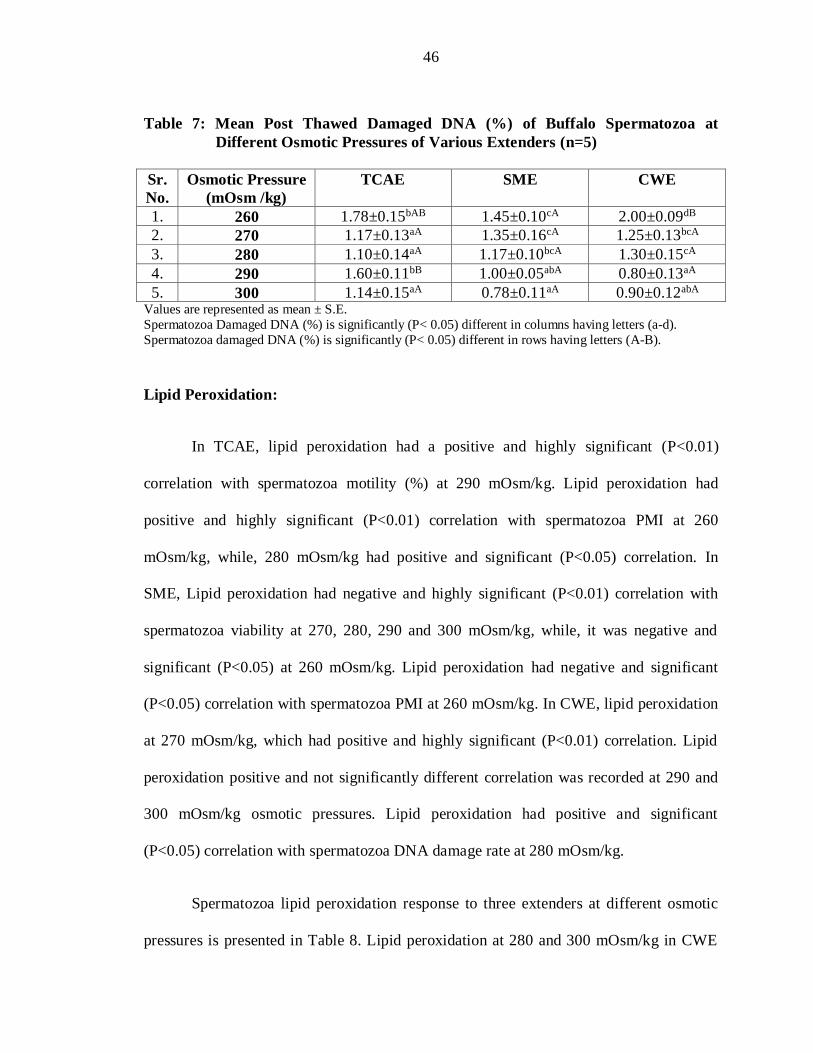
Table 7: Mean Post Thawed Damaged DNA (%) of Buffalo Spermatozoa at
Different Osmotic Pressures of Various Extenders (n=5)
Sr.
No.
Osmotic Pressure
(mOsm /kg)
TCAE
SME
CWE
1. 260 1.78±0.15bAB 1.45±0.10cA 2.00±0.09dB
2. 270 1.17±0.13aA 1.35±0.16cA 1.25±0.13bcA
3. 280 1.10±0.14aA 1.17±0.10bcA 1.30±0.15cA
4. 290 1.60±0.11bB 1.00±0.05abA 0.80±0.13aA
5. 300 1.14±0.15aA 0.78±0.11aA 0.90±0.12abA Values are represented as mean ± S.E.
Spermatozoa Damaged DNA (%) is significantly (P< 0.05) different in columns having letters (a-d).
Spermatozoa damaged DNA (%) is significantly (P< 0.05) different in rows having letters (A-B).
Lipid Peroxidation:
In TCAE, lipid peroxidation had a positive and highly significant (P<0.01)
correlation with spermatozoa motility (%) at 290 mOsm/kg. Lipid peroxidation had
positive and highly significant (P<0.01) correlation with spermatozoa PMI at 260
mOsm/kg, while, 280 mOsm/kg had positive and significant (P<0.05) correlation. In
SME, Lipid peroxidation had negative and highly significant (P<0.01) correlation with
spermatozoa viability at 270, 280, 290 and 300 mOsm/kg, while, it was negative and
significant (P<0.05) at 260 mOsm/kg. Lipid peroxidation had negative and significant
(P<0.05) correlation with spermatozoa PMI at 260 mOsm/kg. In CWE, lipid peroxidation
at 270 mOsm/kg, which had positive and highly significant (P<0.01) correlation. Lipid
peroxidation positive and not significantly different correlation was recorded at 290 and
300 mOsm/kg osmotic pressures. Lipid peroxidation had positive and significant
(P<0.05) correlation with spermatozoa DNA damage rate at 280 mOsm/kg.
Spermatozoa lipid peroxidation response to three extenders at different osmotic
pressures is presented in Table 8. Lipid peroxidation at 280 and 300 mOsm/kg in CWE

47
was significantly higher (P<0.05) than SME and TCAE. Lipid peroxidation at 290
mOsm/kg in CWE was also significantly higher (P<0.05) than in SME.
Table 8: Mean Post Thawed Oxidative Status (nm) of Buffalo Spermatozoa at
Different Osmotic Pressures of Various Extenders (n=5)
Sr.
No.
Osmotic Pressure
(mOsm /kg)
TCAE
SME
CWE
1. 260 37.20±7.44aA 30.73±4.32aA 56.70±6.38aB
2. 270 37.00±9.98aA 27.40±4.25aA 46.70±6.70aA
3. 280 33.70±6.34aA 31.53±5.32aA 55.10±8.11aB
4. 290 35.90±10.70aAB 28.17±4.92aA 55.70±7.61aB
5. 300 29.30±6.19aA 25.13±3.86aA 52.30±5.82aB Values are represented as mean ± S.E.
Means in columns are non-significantly (P> 0.05) different from one another.
Spermatozoa oxidative status (nm) is significantly (P< 0.05) different in rows having letters (A-B).
Using the post thawed spermatozoa quality parameters the best semen extender
(Tris citric acid extender) with optimal osmotic pressure (300 mOsm/kg) for buffalo
semen was decided to be sustained in the next experiment.
EXPERIMENT-II:
In this experiment best extender of TCAE at 300 mOsm/kg osmotic pressure was
used by adding antioxidants (BHT or L-cysteine) for optimum cryopreservation of
buffalo bull semen.
Post-thaw Semen Characteristics:
Spermatozoa Motility:
Significant higher (P<0.05) spermatozoa motility was recorded at 2.0 mM BHT
group compared to other BHT and DMSO groups. Overall mean comparison of
spermatozoa, 2mM L-cysteine had significantly higher (P<0.05) spermatozoa motility
than 1.75 mM of BHT. The highest (47.00±2.33%) and lowest (36.67±2.79%)
48
spermatozoa motility was observed with 2 mM of L-cysteine and 1.75 mM of BHT,
respectively (Table 9).
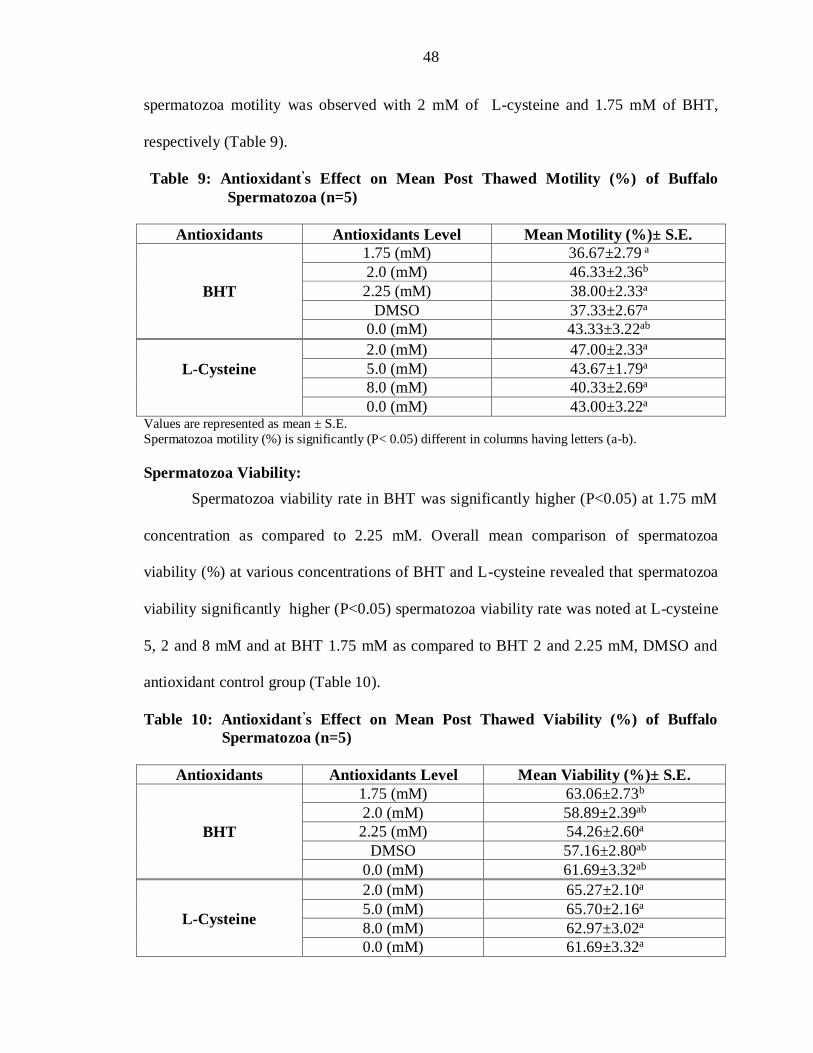
Table 9: Antioxidant’s Effect on Mean Post Thawed Motility (%) of Buffalo
Spermatozoa (n=5)
Antioxidants Antioxidants Level Mean Motility (%)± S.E.
BHT
1.75 (mM) 36.67±2.79 a
2.0 (mM) 46.33±2.36b
2.25 (mM) 38.00±2.33a
DMSO 37.33±2.67a
0.0 (mM) 43.33±3.22ab
L-Cysteine
2.0 (mM) 47.00±2.33a
5.0 (mM) 43.67±1.79a
8.0 (mM) 40.33±2.69a
0.0 (mM) 43.00±3.22a Values are represented as mean ± S.E.
Spermatozoa motility (%) is significantly (P< 0.05) different in columns having letters (a-b).
Spermatozoa Viability:
Spermatozoa viability rate in BHT was significantly higher (P<0.05) at 1.75 mM
concentration as compared to 2.25 mM. Overall mean comparison of spermatozoa
viability (%) at various concentrations of BHT and L-cysteine revealed that spermatozoa
viability significantly higher (P<0.05) spermatozoa viability rate was noted at L-cysteine
5, 2 and 8 mM and at BHT 1.75 mM as compared to BHT 2 and 2.25 mM, DMSO and
antioxidant control group (Table 10).
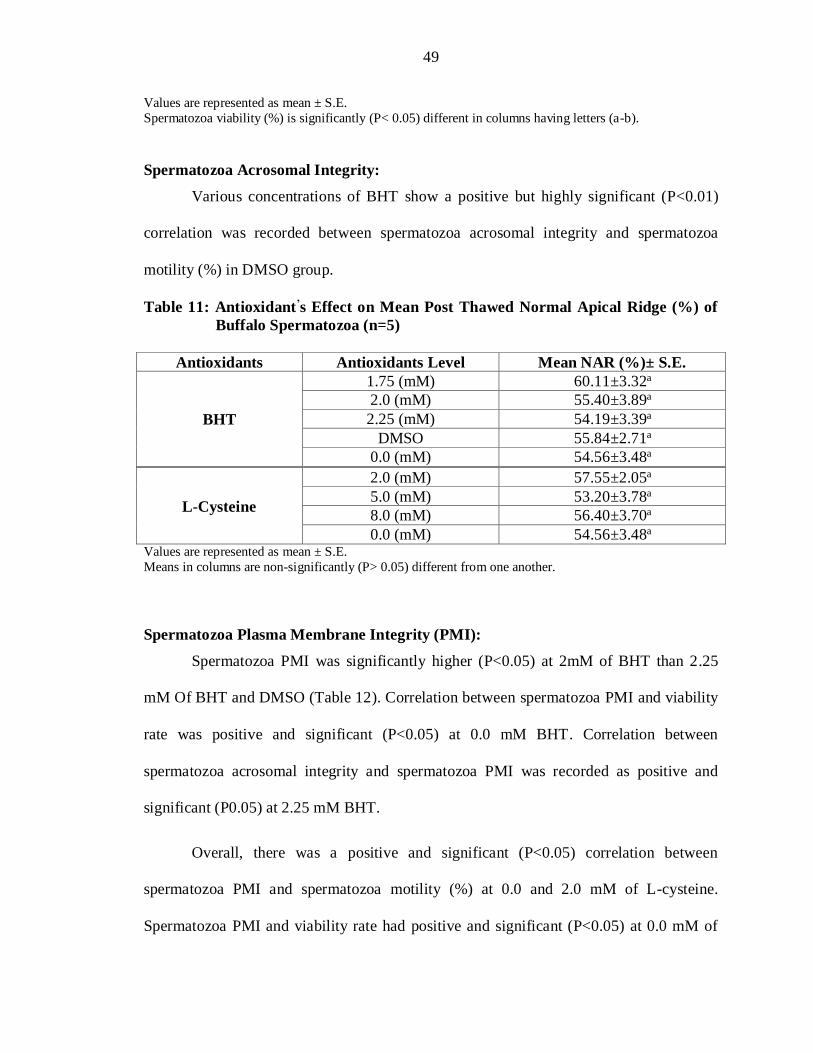
Table 10: Antioxidant’s Effect on Mean Post Thawed Viability (%) of Buffalo
Spermatozoa (n=5)
Antioxidants Antioxidants Level Mean Viability (%)± S.E.
BHT
1.75 (mM) 63.06±2.73b
2.0 (mM) 58.89±2.39ab
2.25 (mM) 54.26±2.60a
DMSO 57.16±2.80ab
0.0 (mM) 61.69±3.32ab
L-Cysteine
2.0 (mM) 65.27±2.10a
5.0 (mM) 65.70±2.16a
8.0 (mM) 62.97±3.02a
0.0 (mM) 61.69±3.32a
49
Values are represented as mean ± S.E.
Spermatozoa viability (%) is significantly (P< 0.05) different in columns having letters (a-b).
Spermatozoa Acrosomal Integrity:
Various concentrations of BHT show a positive but highly significant (P<0.01)
correlation was recorded between spermatozoa acrosomal integrity and spermatozoa
motility (%) in DMSO group.
Table 11: Antioxidant’s Effect on Mean Post Thawed Normal Apical Ridge (%) of
Buffalo Spermatozoa (n=5)
Antioxidants Antioxidants Level Mean NAR (%)± S.E.
BHT
1.75 (mM) 60.11±3.32a
2.0 (mM) 55.40±3.89a
2.25 (mM) 54.19±3.39a
DMSO 55.84±2.71a
0.0 (mM) 54.56±3.48a
L-Cysteine
2.0 (mM) 57.55±2.05a
5.0 (mM) 53.20±3.78a
8.0 (mM) 56.40±3.70a
0.0 (mM) 54.56±3.48a Values are represented as mean ± S.E.
Means in columns are non-significantly (P> 0.05) different from one another.
Spermatozoa Plasma Membrane Integrity (PMI):
Spermatozoa PMI was significantly higher (P<0.05) at 2mM of BHT than 2.25
mM Of BHT and DMSO (Table 12). Correlation between spermatozoa PMI and viability
rate was positive and significant (P<0.05) at 0.0 mM BHT. Correlation between
spermatozoa acrosomal integrity and spermatozoa PMI was recorded as positive and
significant (P0.05) at 2.25 mM BHT.
Overall, there was a positive and significant (P<0.05) correlation between
spermatozoa PMI and spermatozoa motility (%) at 0.0 and 2.0 mM of L-cysteine.
Spermatozoa PMI and viability rate had positive and significant (P<0.05) at 0.0 mM of
50
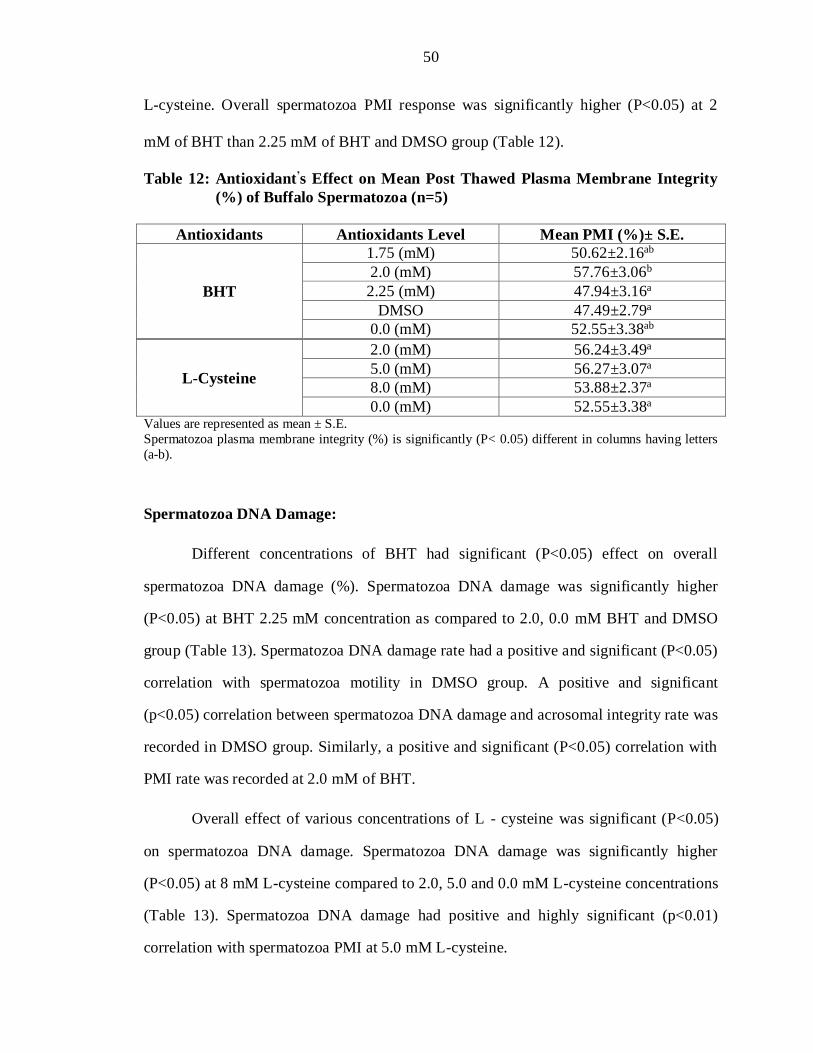
L-cysteine. Overall spermatozoa PMI response was significantly higher (P<0.05) at 2
mM of BHT than 2.25 mM of BHT and DMSO group (Table 12).
Table 12: Antioxidant’s Effect on Mean Post Thawed Plasma Membrane Integrity
(%) of Buffalo Spermatozoa (n=5)
Antioxidants Antioxidants Level Mean PMI (%)± S.E.
BHT
1.75 (mM) 50.62±2.16ab
2.0 (mM) 57.76±3.06b
2.25 (mM) 47.94±3.16a
DMSO 47.49±2.79a
0.0 (mM) 52.55±3.38ab
L-Cysteine
2.0 (mM) 56.24±3.49a
5.0 (mM) 56.27±3.07a
8.0 (mM) 53.88±2.37a
0.0 (mM) 52.55±3.38a Values are represented as mean ± S.E.
Spermatozoa plasma membrane integrity (%) is significantly (P< 0.05) different in columns having letters
(a-b).
Spermatozoa DNA Damage:
Different concentrations of BHT had significant (P<0.05) effect on overall
spermatozoa DNA damage (%). Spermatozoa DNA damage was significantly higher
(P<0.05) at BHT 2.25 mM concentration as compared to 2.0, 0.0 mM BHT and DMSO
group (Table 13). Spermatozoa DNA damage rate had a positive and significant (P<0.05)
correlation with spermatozoa motility in DMSO group. A positive and significant
(p<0.05) correlation between spermatozoa DNA damage and acrosomal integrity rate was
recorded in DMSO group. Similarly, a positive and significant (P<0.05) correlation with
PMI rate was recorded at 2.0 mM of BHT.
Overall effect of various concentrations of L - cysteine was significant (P<0.05)
on spermatozoa DNA damage. Spermatozoa DNA damage was significantly higher
(P<0.05) at 8 mM L-cysteine compared to 2.0, 5.0 and 0.0 mM L-cysteine concentrations
(Table 13). Spermatozoa DNA damage had positive and highly significant (p<0.01)
correlation with spermatozoa PMI at 5.0 mM L-cysteine.
51
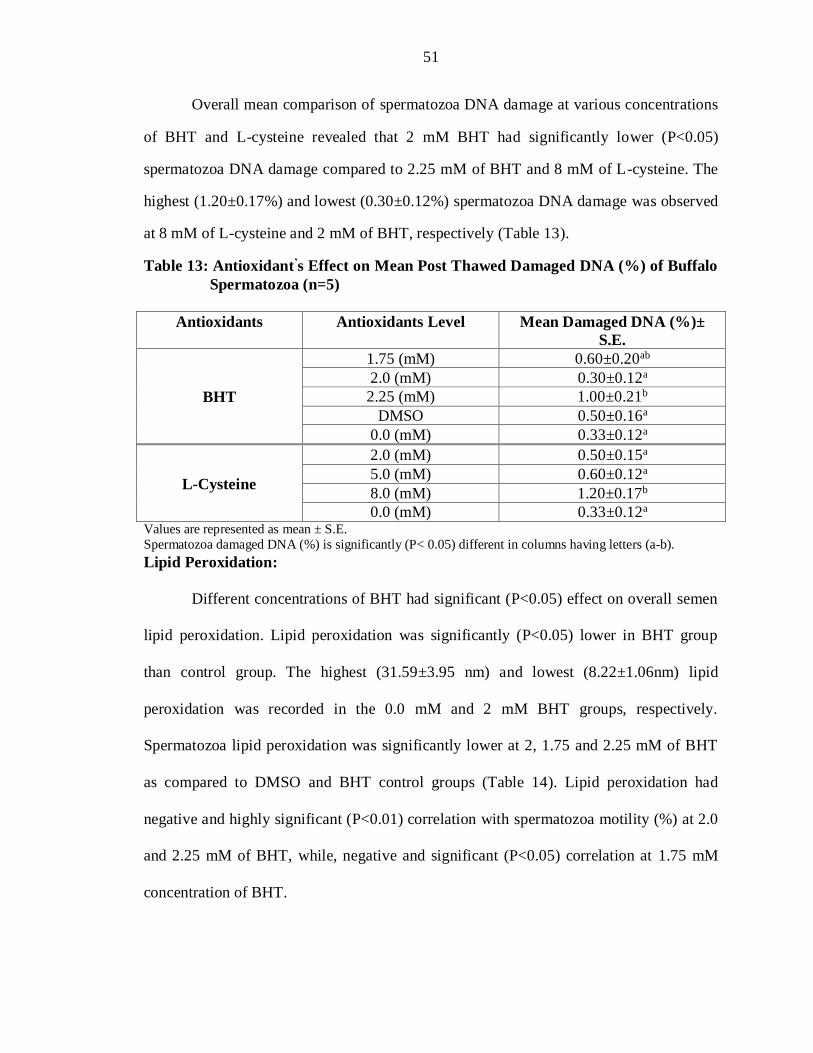
Overall mean comparison of spermatozoa DNA damage at various concentrations
of BHT and L-cysteine revealed that 2 mM BHT had significantly lower (P<0.05)
spermatozoa DNA damage compared to 2.25 mM of BHT and 8 mM of L-cysteine. The
highest (1.20±0.17%) and lowest (0.30±0.12%) spermatozoa DNA damage was observed
at 8 mM of L-cysteine and 2 mM of BHT, respectively (Table 13).
Table 13: Antioxidant’s Effect on Mean Post Thawed Damaged DNA (%) of Buffalo
Spermatozoa (n=5)
Antioxidants Antioxidants Level Mean Damaged DNA (%)±
S.E.
BHT
1.75 (mM) 0.60±0.20ab
2.0 (mM) 0.30±0.12a
2.25 (mM) 1.00±0.21b
DMSO 0.50±0.16a
0.0 (mM) 0.33±0.12a
L-Cysteine
2.0 (mM) 0.50±0.15a
5.0 (mM) 0.60±0.12a
8.0 (mM) 1.20±0.17b
0.0 (mM) 0.33±0.12a Values are represented as mean ± S.E.
Spermatozoa damaged DNA (%) is significantly (P< 0.05) different in columns having letters (a-b).
Lipid Peroxidation:
Different concentrations of BHT had significant (P<0.05) effect on overall semen
lipid peroxidation. Lipid peroxidation was significantly (P<0.05) lower in BHT group
than control group. The highest (31.59±3.95 nm) and lowest (8.22±1.06nm) lipid
peroxidation was recorded in the 0.0 mM and 2 mM BHT groups, respectively.
Spermatozoa lipid peroxidation was significantly lower at 2, 1.75 and 2.25 mM of BHT
as compared to DMSO and BHT control groups (Table 14). Lipid peroxidation had
negative and highly significant (P<0.01) correlation with spermatozoa motility (%) at 2.0
and 2.25 mM of BHT, while, negative and significant (P<0.05) correlation at 1.75 mM
concentration of BHT.
52
A positive and significant (P<0.05) correlation with acrosomal integrity rate was
recorded at 0.0 mM concentration of L-cysteine. Lipid peroxidation had negative and
highly significant (P<0.01) correlation with spermatozoa DNA damage at 0.0 mM
concentration of L-cysteine.
Overall mean comparison of spermatozoa lipid peroxidation at various
concentrations of BHT and L-cysteine was significant (P<0.05). The highest and lowest
response was recorded at 2 mM of L-cysteine and 2 mM of BHT, which was recorded as
35.17±5.23 and 8.22±1.06 nm, respectively. Spermatozoa lipid peroxidation was
significantly (P<0.05) higher at 2 mM of L-cysteine as compared to 1.75, 2.0 and 2.25
mM of BHT and DMSO groups (Table 14).
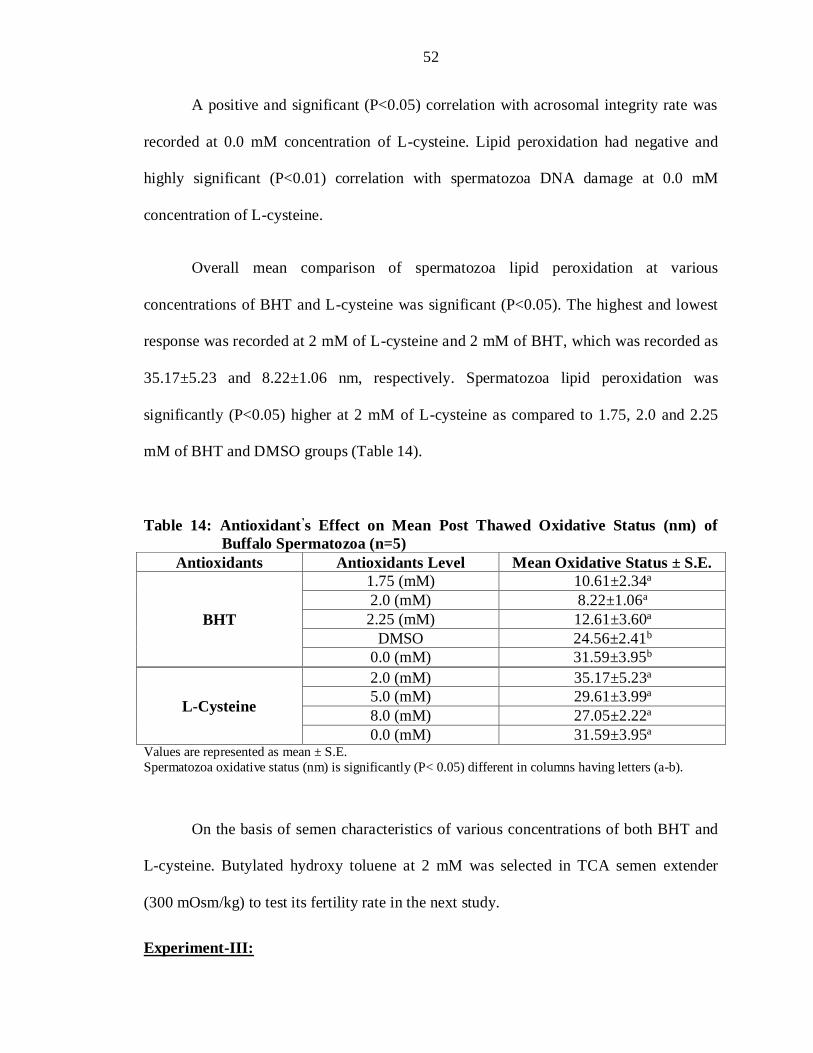
Table 14: Antioxidant’s Effect on Mean Post Thawed Oxidative Status (nm) of
Buffalo Spermatozoa (n=5)
Antioxidants Antioxidants Level Mean Oxidative Status ± S.E.
BHT
1.75 (mM) 10.61±2.34a
2.0 (mM) 8.22±1.06a
2.25 (mM) 12.61±3.60a
DMSO 24.56±2.41b
0.0 (mM) 31.59±3.95b
L-Cysteine
2.0 (mM) 35.17±5.23a
5.0 (mM) 29.61±3.99a
8.0 (mM) 27.05±2.22a
0.0 (mM) 31.59±3.95a Values are represented as mean ± S.E.
Spermatozoa oxidative status (nm) is significantly (P< 0.05) different in columns having letters (a-b).
On the basis of semen characteristics of various concentrations of both BHT and
L-cysteine. Butylated hydroxy toluene at 2 mM was selected in TCA semen extender
(300 mOsm/kg) to test its fertility rate in the next study.
Experiment-III:
53
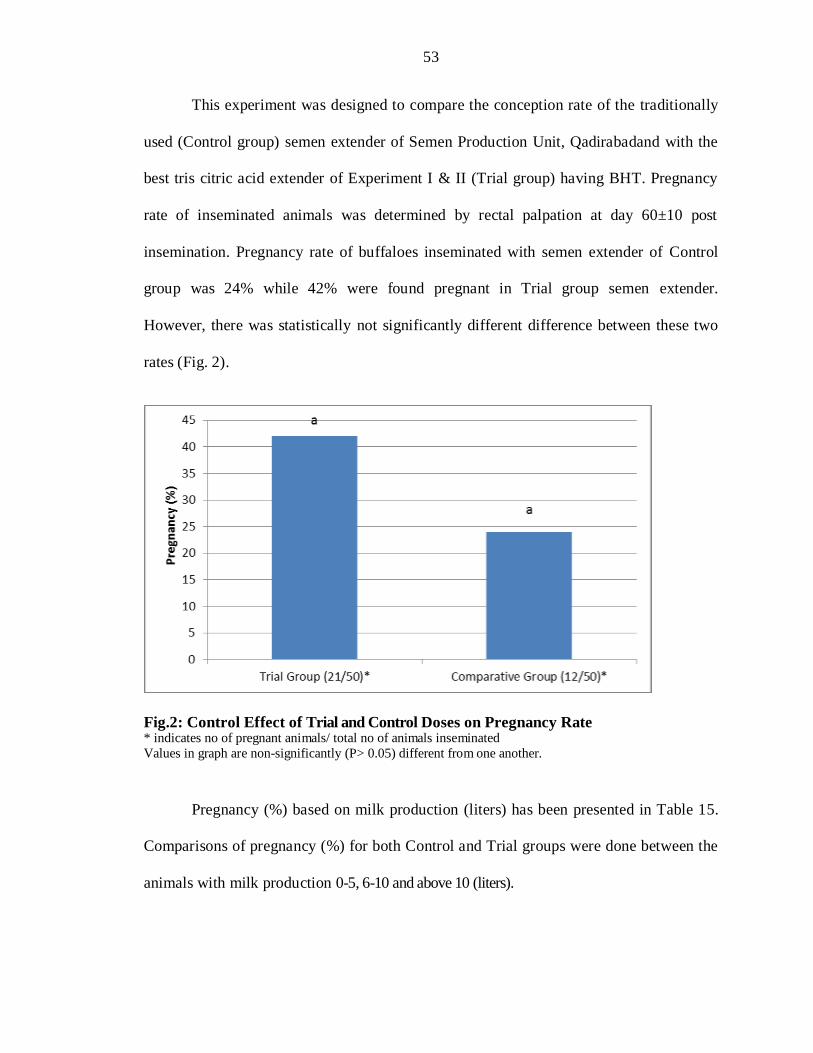
This experiment was designed to compare the conception rate of the traditionally
used (Control group) semen extender of Semen Production Unit, Qadirabadand with the
best tris citric acid extender of Experiment I & II (Trial group) having BHT. Pregnancy
rate of inseminated animals was determined by rectal palpation at day 60±10 post
insemination. Pregnancy rate of buffaloes inseminated with semen extender of Control
group was 24% while 42% were found pregnant in Trial group semen extender.
However, there was statistically not significantly different difference between these two
rates (Fig. 2).
Fig.2: Control Effect of Trial and Control Doses on Pregnancy Rate * indicates no of pregnant animals/ total no of animals inseminated
Values in graph are non-significantly (P> 0.05) different from one another.
Pregnancy (%) based on milk production (liters) has been presented in Table 15.
Comparisons of pregnancy (%) for both Control and Trial groups were done between the
animals with milk production 0-5, 6-10 and above 10 (liters).

54
Comparison of pregnancy (%) based on buffalo parity has been presented in Table
16. Pregnancy (%) for control and trial groups was done for buffaloes with pirity 0, 1-2, 3-
4 and 5 and above (Table 16).
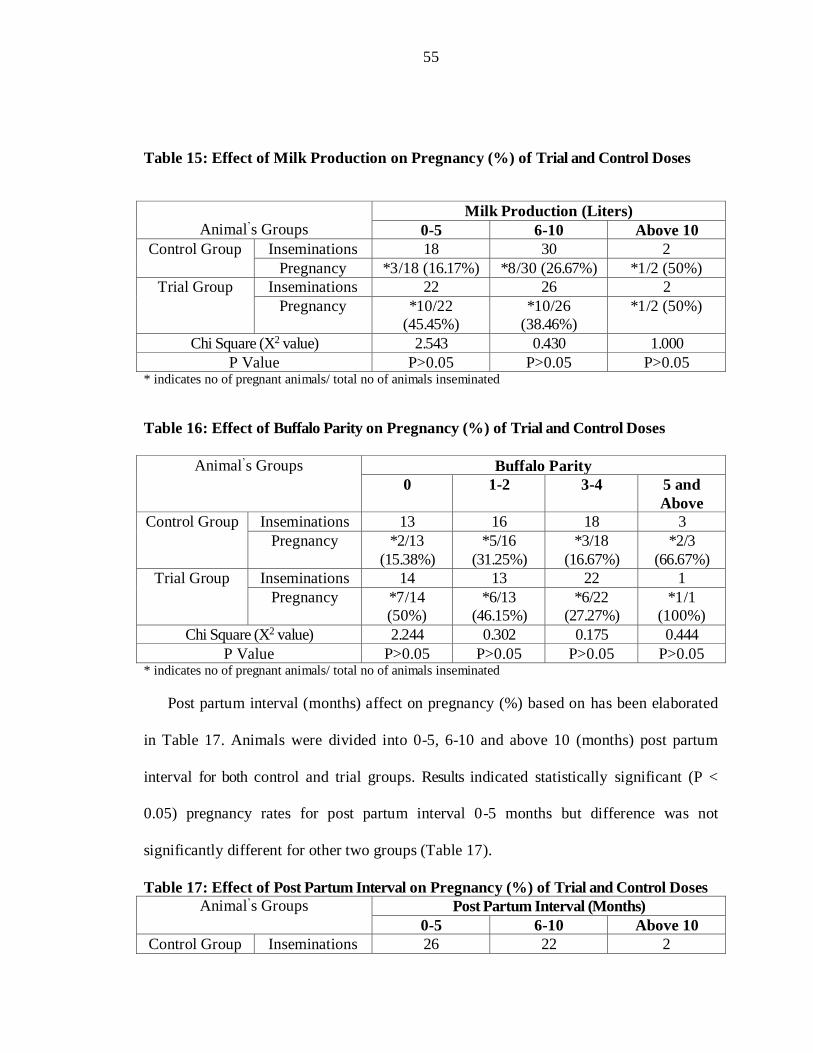
55
Table 15: Effect of Milk Production on Pregnancy (%) of Trial and Control Doses
Animal’s Groups
Milk Production (Liters)
0-5 6-10 Above 10
Control Group Inseminations 18 30 2
Pregnancy *3/18 (16.17%) *8/30 (26.67%) *1/2 (50%)
Trial Group Inseminations 22 26 2
Pregnancy *10/22
(45.45%)
*10/26
(38.46%)
*1/2 (50%)
Chi Square (X2 value) 2.543 0.430 1.000
P Value P>0.05 P>0.05 P>0.05 * indicates no of pregnant animals/ total no of animals inseminated
Table 16: Effect of Buffalo Parity on Pregnancy (%) of Trial and Control Doses
Animal’s Groups Buffalo Parity
0 1-2 3-4 5 and
Above
Control Group Inseminations 13 16 18 3
Pregnancy *2/13
(15.38%)
*5/16
(31.25%)
*3/18
(16.67%)
*2/3
(66.67%)
Trial Group Inseminations 14 13 22 1
Pregnancy *7/14
(50%)
*6/13
(46.15%)
*6/22
(27.27%)
*1/1
(100%)
Chi Square (X2 value) 2.244 0.302 0.175 0.444
P Value P>0.05 P>0.05 P>0.05 P>0.05 * indicates no of pregnant animals/ total no of animals inseminated
Post partum interval (months) affect on pregnancy (%) based on has been elaborated
in Table 17. Animals were divided into 0-5, 6-10 and above 10 (months) post partum
interval for both control and trial groups. Results indicated statistically significant (P <
0.05) pregnancy rates for post partum interval 0-5 months but difference was not
significantly different for other two groups (Table 17).
Table 17: Effect of Post Partum Interval on Pregnancy (%) of Trial and Control Doses
Animal’s Groups Post Partum Interval (Months)
0-5 6-10 Above 10
Control Group Inseminations 26 22 2
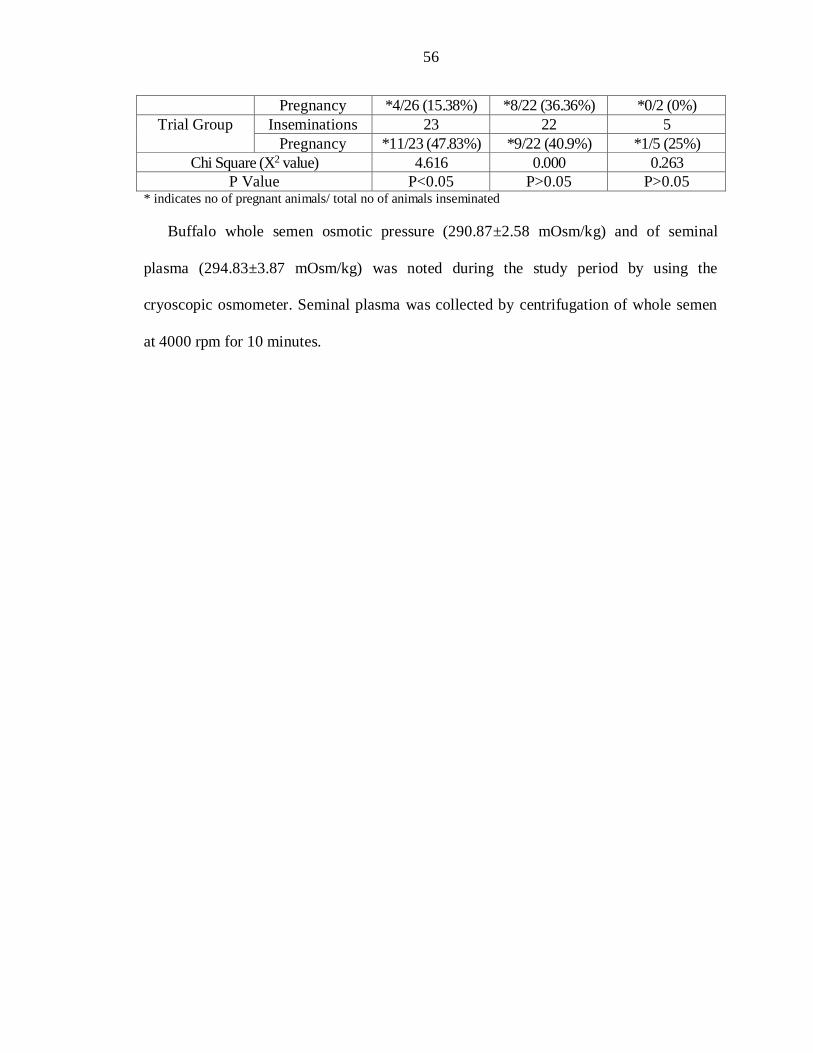
56
Pregnancy *4/26 (15.38%) *8/22 (36.36%) *0/2 (0%)
Trial Group Inseminations 23 22 5
Pregnancy *11/23 (47.83%) *9/22 (40.9%) *1/5 (25%)
Chi Square (X2 value) 4.616 0.000 0.263
P Value P<0.05 P>0.05 P>0.05 * indicates no of pregnant animals/ total no of animals inseminated
Buffalo whole semen osmotic pressure (290.87±2.58 mOsm/kg) and of seminal
plasma (294.83±3.87 mOsm/kg) was noted during the study period by using the
cryoscopic osmometer. Seminal plasma was collected by centrifugation of whole semen
at 4000 rpm for 10 minutes.

57
Chapter V
DISCUSSION
Higher plasma membrane contents of poly unsaturated fatty acids (PUFA) could
be a reason for higher oxidation rate in buffalo semen (Chatterjee and Gagnon, 2001).
Secondly, ignoring of osmolality requirement in buffalo semen extenders could be
another factor for lower conception rates after use of cryopreserved semen (~269
mOsm/L; Khan and Ijaz, 2008).
Pre Freeze Semen Analysis:
Semen volume noted in present study was comparable to that reported by Younis,
(1996). However, higher semen volume was reported by Ahmad (1984). Mean
spermatozoa concentration of the present study was comparable to the report of Adeel et
al. 2009. However, Reddy et al. (1992) reported a lower concentration of spermatozoa in
Murrah buffalo bulls. Mean spermatozoa motility of ejaculates recorded during this trial
was in agreement with Rasul et al. (2001). Comparatively higher motility of was reported
by Hashemi et al. (2007). Mean spermatozoa viability (%) in ejaculates was close to the
findings of Kanwal et al. (2000). Our result of spermatozoa viability was comparatively
higher as reported by Iqbal et al., (2010). Mean spermatozoa acrosomal integrity in
ejaculates was precisely similar as reported by Aguiar et al. (1994). Comparatively higher
acrosomal integrity was observed by Hashemi et al. (2007). Spermatozoa plasma
membrane integrity (PMI) in ejaculates was in agreement with Rasul et al. (2001). A
lower PMI in buffalo semen was reported by Singh et al. (2007). Difference in semen
parameters from these studies might be associated with differences in frequency and
procedure of semen collection, procedure of sample processing, expertise of the analyzer,

58
body size and weight, scrotum circumference, nutrition, age or breed of the bull and
season of semen collection.
MTT reduction rate of the present study revealed a direct correlation between the
reduction rate and spermatozoa viability under study. Iqbal et al. (2010) reported same
positive correlation for buffalo bulls and Aziz et al. (2005) for stallions. The present
assay is therefore, a better technique in evaluating the viability of spermatozoa whenever,
time, cost and achievability are important (Iqbal et al., 2010).
EXPERIMENT-I:
Experiment was designed to optimize the osmotic pressure of buffalo semen
extender. Tris citric acid extender (TCAE) and skim milk extender (SME) at <270
mOsm/kg osmotic pressure had significantly decreased post thaw spermatozoa motility.
And increasing trend in spermatozoa motility noted with increases in osmotic pressure of
SME. Decreased spermatozoa motility at osmotic pressures <270 mOsm/kg may be as
motility depends on ATP contents of spermatozoa mitochondria (Perchek et al., 1995)
which were damaged by hypo osmotic stress and consequently reduces the ATP
availability to spermatozoa (Meyers, 2005). Mitochondrial damage in hypo osmotic
extender is primarily because of increased intracellular ice formation during
cryopreservation. Spermatozoa motility observed at 300 mOsm/kg of TCAE were closely
related with the results reported for buffalo semen cryopreservation at 320 mOsm/kg
osmotic pressure by Ansari et al. (2010), Andrabi et al. (2008b) and Rasul et al. (2001).
Our study was in agreement with Liu et al. (1998) in which they has documented an
osmotic pressure of 305-375 mOsm/kg in tris extender suitable for cattle bull
cryopreservation. Similarly, Liu and Foote (1998) noted higher spermatozoa motility in

59
cattle bull semen at 300 mOsm/kg compared to hypo-osmotic tris extender i.e., 200
mOsm/kg. Liu and Foote also noted that spermatozoa motility had decreasing trend as
osmotic pressure gone up or down from 300 mOsm/kg in tris extender. Higher osmotic
requirement of cattle bull semen may be correlated to the fact that higher osmotic
pressure increments (35-40 mOsm/kg) were used in their study unlike our study having
increments of (10 mOsm/kg). Our results of SME were in agreement with Liu et al.
(1998) experiment in which osmotic pressure of (SME) 270- 340 mOsm/kg was
considered optimum without any difference in spermatozoa motility. Our results were
contrary of Ari et al. (2011) who noted higher spermatozoa motility in skim milk
extender for goat semen. These differences in results may be due to species difference.
Kommisrud et al. (1996) noted (17%) higher post thawed motility in cattle compared to
our study and in both studies comparable skim milk extenders were used. This difference
in spermatozoa motility may be due to difference in osmotic pressure which was not
considered in former study. Unlike other extenders no change in spermatozoa motility
noted in coconut water extender (CWE) at lower osmotic pressures partly may be
because the lowest and highest motility in CWE was 6% compared to other two extenders
where motility differences were 13%. Overall lower motility pattern in CWE may be
because of inadequate energy availability (Rasad and Simanjuntak, 2009). Vale et al.
(1997) used coconut water extender and reported an osmolality of 320 mOsm/kg for
buffalo semen cryopreservation. This difference may be due to the fact that they did not
use any other osmotic pressure level for buffalo semen cryopreservation. Luzardo et al.
(2010) found that cryopreserved boar semen diluted in coconut water based extender has
higher spermatozoa motility when semen extender has osmolality 381-480 mOsm/kg.

60
Comparison of spermatozoa motility in TCAE was significantly higher (P<0.05)
than SME and CWE at 280, 290 and 300 mOsm/kg. Low visibility could be a factor for
low motility observation in SME extender as reported by Foote et al. (1993). Higher
spermatozoa motility in TCAE can be attributed to the zwitterions (tris) which were not
permeable to plasma membrane and dehydrate the spermatozoa more efficiently during
cryopreservation (Good et al., 1966) which decreases intracellular ice formation.
Spermatozoa viability at similar osmotic pressure of three extenders was
significantly higher (P<0.05) at 270 mOsm/kg of TCAE and CWE compared to SME.
That may be due to the fact that TCAE as zwitterions resist the changes in pH more
efficiently and also may be due to less intracellular enzyme leakage during
cryopreservation in tris based extenders as reported by Dhami and Kodagali, (1990).
Spermatozoa acrosomal integrity at 300 mOsm/kg of TCAE was significantly
(P<0.05) higher than <300 mOsm/kg. Similar to present study, Woelders et al. (1997)
noted that bull spermatozoa acrosomal integrity was significantly higher in isotonic
extender. Results of Singh et al., (2007) were close to our findings; however, a relatively
higher spermatozoa acrosomal integrity (24%) was reported by Andrabi et al., (2008b)
and 13% higher by Akhtar et al. (2010) by using buffer of osmotic pressure 320
mOsm/kg. The difference of spermatozoa acrosomal integrity might be due to the change
in osmotic pressure of the media. According to Soylu et al. (2007) ram semen acrosome
damage was lower at 400 mOsm/kg osmotic pressure in tris extender. Ram has higher
osmotic pressure requirements than bovines. Acrosomal integrity at 300 mOsm/kg of
SME was higher (P<0.05) compared to 260, 280 and 290 mOsm/kg whereas in CWE no
difference in acrosomal integrity was observed among various osmotic pressures.

61
Variation in spermatozoa acrosomal integrity in different extenders may be due to fact
that different spermatozoa parameters behave in a different way in various semen
extenders (Songsasen, 2002). Vale et al. (1997) reported 20 mOsm/kg higher optimum
osmolality than our study using coconut water extender for buffalo semen
cryopreservation. Lozardo et al. (2010) found that crypreserved boar semen diluted in
coconut water based extender has higher spermatozoa acrosomal integrity when semen
extender has osmolality 381-480 mOsm/kg. In this study higher percentage of acrosomal
integrity at 300 mOsm/kg may be due to relatively hypertonic extenders comparing to the
~293 mOsm/kg; reported by Ibrahim et al. (1985). Improved spermatozoa survival in
hypertonic extender as it enhances dehydration of the spermatozoa and reduces the
quantity of intracellular ice (Watson, 1979; Fiser et al., 1981; Pommer et al., 2002).
Comparative spermatozoa acrosomal integrity rate of three extenders at 290 mOsm/kg
osmotic pressures improved (P<0.05) in CWE compared to TCAE. Moreover, coconut
water has indole acetic acid and phytohormones that provide protection to spermatozoa
during cryopreservation (Nunes, 1993; Nunes et al., 1996).
In SME, lower spermatozoa plasma membrane integrity (PMI) rate at 260 than
290 mOsm/kg osmotic pressure may be due to damage of membrane ionic channels
(Kruger et al. 1984, Alavi et al. 2007), and due to structural damage that occur to the
plasma membrane (Meyers, 2005) at hypo-osmotic conditions.
In TCAE, spermatozoa DNA damage was higher (P<0.05) at both 260 and 290
mOsm/kg compared to other osmotic pressures as reported by Martin et al. (2007). A
higher rate of DNA fragmentation (35%) in buffalo bull semen was reported by Kumar et
al. (2011) by commet assay, higher DNA damage may be due to difference in assay

62
technique. Nur et al. (2010) in ram and El-Sisy et al. (2010) in buffalo noted higher
spermatozoa DNA damage (4.5 and 10.4%, respectively) by acridine orange staining
technique. In these both studies osmotic pressure was not mentioned and an-isotonic
extender could be a reason for higher DNA damage of buffalo semen. In this study
finding of significantly lower (P<0.05) Spermatozoa DNA damage noted in SME and
CWE at 300 mOsm/kg than at osmotic pressures of 280 mOsm/kg and below was in line
with Yildiz et al. (2010) results that DNA damage was significantly lower in iso-osmotic
solutions (300 mOsm/kg) because mammalian spermatozoa had a limited ability to any
change of cell volume and were unable to maintain structural and functional integrity.
Waterhouse et al. (2010) noted higher buffalo spermatozoa DNA damage (7.7%) via
acridine orange technique in SME. As in this study at lower osmotic pressure higher
DNA damage was noted and an-isotonic extender in the Waterhouse et al. study as well
could be reason for higher DNA damage, similarly, higher unsaturated fatty acids
contents of buffalo plasma membrane enhance the vulnerability of plasma membrane to
oxidative stress triggered by hypo osmotic extenders that results into DNA damage
(Aitken and Krausz, 2001; Meyers, 2005). Over compaction of spermatozoa chromatin
(due to DNA strand separation, disulphide/covalent bonds formation; Cordova et al.
2002) occur (Glogowski et al. 1994) due to higher Intracellular ice (Royere et al., 1991)
and oxidative stress (Lewis and Aitken, 2005). Damaged spermatozoa did not attach to
oviduct, unable to decondence at fertilization (Ardon et al., 2008) and transmit defective
genome (Sakkas and Alvarez, 2010) that cause fertilization failure/embryo death/poor
embryo development.

63
Spermatozoa DNA damage was higher at 260 mOsm/kg osmotic pressures in both
TCAE and CWE than in SME. Similarly, at 290 mOsm/kg osmotic pressure spermatozoa
DNA damage in TCAE was higher than both SME and CWE. Lower spermatozoa DNA
damage in SME may be due to presence of natural antioxidants (casein proteins) in milk
(Salamon and Maxwell, 2000).
Minimum lipid per oxidation is associated with good semen quality (Chaudhari et
al., 2008). Semen lipid per oxidation response to three extenders at comparable osmotic
pressures (260, 280 and 300 mOsm/kg) was lower (P<0.05) in SME and TCAE than
CWE. Presence of natural antioxidants (casein proteins etc.) in skimmed milk (Salamon
and Maxwell, 2000) might be responsible for lower lipid per oxidation in SME.
Possibility for lower lipid per oxidation in TCAE may be due less intracellular ice
formation as TCAE dehydrate the spermatozoa more efficiently during cryopreservation
because impermeability of plasma membrane for tris.
Role of osmotic pressure difficult to assess by semen evaluation alone as it
damages the internal structures (mitochondria, acrosome and DNA) of the spermatozoa
which ultimately adversely affect the fertilizing ability of semen.
EXPERIMENT-II:
In this study various concentrations of antioxidants (butylated hydroxyl toluene
and L-cysteine) were added to determine their effects on semen characteristics. In current
study spermatozoa motility was not different from control group however, significantly
higher spermatozoa motility was noted at 2.0 mM BHT compared to other treatments.
Reason of no difference in spermatozoa motility from control group might be due to the
fact that osmotic pressure of semen extender used in this study was optimized in last

64
experiment. Lower spermatozoa motility noted at 2.25 mM compared to 2 mM BHT is
speculated due to damage of internal organs at higher BHT level, detail of phenomenon
involved is described in next paragraph. Similar findings were reported by Neagu et al.
(2010). However, Ijaz et al. (2009) reported similar buffalo spermatozoa motility at 1.0
and 2.0 mM BHT but in this study 2 mM BHT improved the spermatozoa motility than
both less and above levels of BHT. Similar results to present study were also reported by
Farshad et al. (2010) during ram semen cryopreservation. Comparatively higher buffalo
spermatozoa motility was observed at BHT 1.5 mM, than control (Pankaj et al., 2009) as
semen quality assessment in their study was carried out at room temperature over
different time periods (24 and 48 hours) instead of cryopreservation, at room temperature
spermatozoa metabolism rate was higher; after 48 hours BHT prevent oxidation more
effectively than after 24 hours. In current study semen was immediately cryopreserved
after addition of BHT and spermatozoa metabolism was not allowed to occur at higher
rate. On these bases it can be speculated that BHT may have an effective role after semen
deposition in female reproductive tract as antioxidants protect the spermatozoa integrity
after thawing. In buffalo Munir, (2011) also observed higher motility (%) at BHT 1.5
mM but in current study we did not use 1.5 mM of BHT. Ansari et al. (2011b) and Shoe
and Zamiri, (2008) results were also in our opposition in which 0.5-1.0 mM BHT
addition in cattle semen has higher spermatozoa motility. This difference in spermatozoa
motility may be due to higher buffalo plasma membrane contents of PUFA (Sansone et
al., 2000). It is suggested that antioxidants maintained the spermatozoa motility by
scavenging the ROS molecules (Bucak et al., 2008) which causes lipid peroxidation of
the spermatozoa plasma membrane (Urata et al., 2001). Membrane damage due to high

65
ROS further contributes in spermatozoa axosome and mitochondria damage and
ultimately affect spermatozoa motility as motility depends on ATP contents of
spermatozoa mitochondria (Perchek et al., 1995).
In the current study significantly lower spermatozoa viability, PMI rates and
higher DNA damage (%) at 2.25 mM BHT might be due to toxic effects of BHT as
excessive antioxidant decreases physiological level of oxidants which is considered
essential for normal spermatozoa function (Alvarez and Storey, 1983; Roca et al., 2004),
moreover, higher concentrations of BHT make spermatozoa vulnerable to cryo-injury by
increasing the plasma membrane fluidity (Shoe and Zamiri, 2008). As higher
concentrations of BHT decrease the oxidation rate of PUFA (in spermatozoa membrane)
and consequently enhance the membrane fluidity (Hammerstedt et al., 1976; Beconi et
al., 1993; Urata et al., 2001; Sikka, 2004). Enhanced fluidity of spermatozoa membrane
weakens the function of ionic channels/ATPases in membrane. As ionic
channels/ATPases regulate the entry of nutrients into the spermatozoa and damage to
these channels ultimately damages the spermatozoa. Higher oxidative stress during
buffalo semen cryopreservation due to its higher PUFA contents results into DNA
damage (Aitken and Krausz, 2001; Meyers, 2005). This is hypothesized that spermatozoa
damage is a cascade of events that decreases membrane integrity, damages DNA and
finally loss of spermatozoa viability, interestingly all these parameters of buffalo
spermatozoa were adversely affected at higher inclusion level of BHT. Similarly toxic
effect of higher BHT concentrations were reported in literature by Farshad et al. (2010),
Ijaz et al. (2009) and Shoe and Zamiri, (2008) in ram/buffalo and cattle, respectively.
However, our results of 2.25mM were not in agreement with work of Pankaj et al. (2009)

66
and Munir, (2011) on buffalo semen the difference may be due to the fact that BHT
above 2.0 mM was not used in these studies.
Significant lowest (P<0.05) lipid per oxidation was observed in BHT treated than
control group but difference among BHT treatment groups was not significantly different.
Similarly, Roca et al. (2004) claimed that 1.6 mM BHT addition to boar semen
significantly decrease the lipid peroxidation. In the present study lipid oxidation was
numerically decreased at BHT 2 mM compared to 1.75mM but surprisingly an increase
in peroxidation at 2.25 mM was noted. The relative increase in peroxidation might be due
to spermatozoa damage which was coinciding with significant decrease PMI and increase
in DNA damage and probably could be a reason for peroxidation even in the presence of
higher antioxidant level. Minimum lipid peroxidation is associated with good semen
quality and fertility (Chaudhari et al., 2008; Sinha et al., 1996; Stradaioli et al., 2007).
Buffalo spermatozoa membranes have relatively greater concentrations of membrane
PUFA and were more susceptible to peroxidative damage (Chatterjee and Gagnon, 2001),
that is why; antioxidants decreased the lipid peroxidation and improved semen quality.
Results of the present study have shown that DMSO did not have any effect as
antioxidant which rule out the role of DMSO as antioxidants in all BHT treated groups
because DMSO results for each spermatozoa parameter was same as that of control group
in this study. Based on the results of this study we can propose that 45 ul DMSO was not
an effective antioxidant. Secondly, DMSO might have poor antioxidant properties for the
mammalian spermatozoa cryopreservation. Therefore, DMSO could be used to dissolve
lipid soluble antioxidants instead of alcohol as we did in this study.

67
Similarly, L-cysteine caused spermatozoa DNA damage at highest level (i.e., 8
mM) and no difference found among rest of the groups. This may be due to the toxic
effect of cysteine as described earlier. Our results were contrary to Thuwanut, (2007)
work which may be primarily due to difference in extender composition, osmotic
pressure and species. Tuncer et al. (2010) noted higher cattle spermatozoa DNA damage
at 10 mM cysteine concentration compared to control as reported in this study.
Positive influence of BHT on post thaw semen quality as compared to L-cysteine
may be due to the fact that BHT protected plasma membrane fluidity due to its lipid
solubility (Khalifa et al., 2008). Second factor may be related to the fact that BHT is a
synthetic analogue of vitamin E and is a reactive oxygen species production chain
breaker not a scavenging antioxidant (Dad et al., 2006), thus it protects plasma membrane
integrity without affecting ROS production system (Sharma and Agarwal, 1996). Another
possible mechanism for BHT positive influence is due to its antiviral properties that
inactivate viruses in semen (Snipes et al., 1975; Hammerstedt et al., 1976).
Experiment-III:
Experiment was carried out to compare the pregnancy rate of the traditionally
used semen extender of S.P.U., Qadirabad, Pakistan (Control group) and tris citric acid
extender, selected on the basis of the results of experiment- II (Trial group). Pregnancy
rate was 24 and 42% in Control and Trial groups, respectively but difference was
statistically not significantly different. Less no of inseminations in buffaloes may be the
reason for the not significantly different results. Lower fertility rates in buffaloes in
Control group might be due to reduced motility, acrosome, PMI and DNA damage that
may be caused by osmotic and oxidative stresses (Aitken and Krausz, 2001; Meyers,

68
2005) as semen extender used in Control group was without any antioxidant and
moreover osmolality was higher. Reduced spermatozoa motility might be related to less
ATP availability due to mitochondrial damage. Secondly, glucose 6 phosphate
dehydrogenase (control ATP production via glycolysis pathway) intra-spermatozoa
concentration is decreased due to membrane damage that leads to reduced energy
availability and ultimately causes less spermatozoa motility. This reduced motility might
be responsible for lower fertility with cryopreserved semen as immotile spermatozoa
cannot reach to fertilization site (Cerolini et al, 2001; Gillan et al, 2004). Loss of plasma
membrane structural and functional integrity occur as proteins and fatty acyl chains
become deranged during cryopreservation (might be due to osmotic and oxidative
stresses) that decreases membrane fluidity which may be responsible for plasma
membrane rigidity and vesiculation (Medeiros et al, 2002; Parks and Lynch, 1992;
Watson, 2000). Spermatozoa with deranged plasma membrane cannot bind to cells of
female reproductive tract and oocyte (Lessard et al, 2000; Medeiros et al, 2002; Watson,
2000). Restructured plasma membrane is also responsible for increased calcium entry
into spermatozoa. This increased intracellular calcium leads to spermatozoa capacitation
(hyper activation) and acrosomal reaction. Atypical spermatozoa capacitation reduces
spermatozoa functionality and viability in female reproductive tract (Bailey et al, 2000;
Watson, 2000) and ultimately fertility. Phenomenon of DNA damage by osmotic and
oxidative stress and its effect on fertility as described in Experiment I. Results of the
present experiment were in agreement with Roca et al. (2004) in which they compared
fertility of boar semen tris extender supplementation with BHT. Our results were not in
agreement with Khalifa et al. (2008) in vivo fertility trial in goat and in vitro fertility trial

69
in boar with tris extender having BHT (Pursel, 1979). Difference in results from Pursel’s,
may be due to huge difference between in vitro and in vivo fertility. Pregnancy rate of our
study was lower from the results reported by Anwar et al. (2008) with tris citric acid
extender in buffalo. This fertility difference may be due to difference in insemination
seasons, cryopreservation procedure, nutrition and management of experimental animals.
Similar buffalo fertility rates were reported in tris based extender by Singh et al. (1980);
Dhami and Kodagali, (1990) and Akhtar et al. (2010).
Conclusion and Suggestions:
TCA extender having 300 mOsm/kg osmotic pressure and optimal inclusion of BHT
(2.0 mM) improved post thaw semen quality and yielded relatively better pregnancy rates.
Results of the study, clearly indicate that osmotic stress primarily damaged the spermatozoa
internal structures more severely (mitochondria, acrosome and DNA) rather than plasma
membrane.
Understanding of Semen Production Units of Pakistan to use semen extender with
optimum osmotic pressure (~300 mOsm/kg) and antioxidant inclusion (e.g. 2.0 mM BHT) may
improve buffalo pregnancy rates. Affect of >300 mOsm/kg osmotic pressure in buffalo semen
extenders may be important to determine. Furthermore, fertility trial with above mentioned
extenders at larger scale in the field is recommended in future.

70
Chapter VI
SUMMARY
Presently, buffalo farmers are dissatisfied with fertility rates of the frozen semen
used in the field and tend to use bulls. This study was designed to develop a suitable
semen extender for cryopreservation of Nili Ravi buffalo semen that can improve
conception rate in buffaloes.
Experiment-I, an attempt was made to develop semen extender with optimal
osmotic pressure for buffalo semen using tris citric acid (TCAE), skim milk (SME) and
coconut water (CWE) extenders (each extender have 260, 270, 280, 290 and 300
mOsm/kg osmotic pressure levels). In Experiment-II, best extender (TCAE: 300
mOsm/kg) of experiment-I was tried to improve post thaw spermatozoa characteristics by
supplementing antioxidants [0.0, 1.75, 2.0 and 2.25 mM butylated hydroxy toluene
(BHT) and 0.0, 2.0, 5.0 and 8.0 mM L-cysteine]. Post thaw spermatozoa motility,
viability, plasma membrane integrity (PMI), DNA damage rate and lipid peroxidation
were assessed in first two experiments. In Experiment-III, pregnancy rate assessment of
extended semen was carried out by using Trial extender (best of experiment II) or Control
extender of Semen Production Unit (SPU), Qadirabad, Pakistan (50 inseminations of
each extender).
Higher spermatozoa motility at ≥ 270 mOsm/kg was noted in TCAE than both
SME and CWE could be due to less intracellular ice formation in zwitterions extender.
Higher spermatozoa viability in TCAE and CWE compared to SME may be attributed to
extender effectiveness. Higher acrosomal integrity rate at 300 mOsm/kg in TCAE and
SME may be because of less intracellular ice formation in isotonic extenders. At 290

71
mOsm/kg, higher spermatozoa PMI in SME and lesser DNA damage in three extenders
might be due to lesser intracellular ice formation at cryopreservation. Decreased
spermatozoa DNA damage in SME might be due to the presence of natural antioxidants
i.e., casein. Higher lipid peroxidation in CWE than TCAE and SME may be due to
presence of natural antioxidants (in SME) and higher cell dehydration potential of TCAE.
Higher spermatozoa motility recorded at 2.0 mM BHT compared to other BHT
groups including DMSO might be due to fact that BHT protects spermatozoa
mitochondria by reducing oxidative stress. Lower spermatozoa viability, PMI rates and
higher DNA damage at 2.25 mM of BHT may be due to BHT toxic effects. Lower lipid
peroxidation in BHT treated groups compared to DMSO and BHT control groups might
be related to BHT strong antioxidant properties. L-cysteine caused higher spermatozoa
DNA damage at highest level (i.e., 8 mM) that could also be due to antioxidant’s toxic
effect.
Pregnancy rate 18 % higher was noted in Trial than Control semen extender;
however no significant difference have been noted that might be due to less no of
inseminations.
In conclusion, TCA extender (300 mOsm/kg) having BHT (2.0 mM) improved post
thaw semen quality and yielded numerically better pregnancy rates. Results of study indicated
that osmotic stress damaged the spermatozoa internal structures more severely than injury to
plasma membrane.





























