Embed Size (px)
Citation preview

1 23
Biological Trace Element Research ISSN 0163-4984 Biol Trace Elem ResDOI 10.1007/s12011-014-0026-4
Dietary Supplementation of ZincNanoparticles and Its Influence on Biology,Physiology and Immune Responses ofthe Freshwater Prawn, MacrobrachiumrosenbergiiThirunavukkarasu Muralisankar,Periyakali Saravana Bhavan,Subramanian Radhakrishnan,Chandirasekar Seenivasan, et al.

1 23
Your article is protected by copyright and all
rights are held exclusively by Springer Science
+Business Media New York. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

Dietary Supplementation of Zinc Nanoparticles and Its Influenceon Biology, Physiology and Immune Responses of the FreshwaterPrawn, Macrobrachium rosenbergii
Thirunavukkarasu Muralisankar & Periyakali Saravana Bhavan &
Subramanian Radhakrishnan & Chandirasekar Seenivasan &
Narasimman Manickam & Veeran Srinivasan
Received: 6 March 2014 /Accepted: 21 May 2014# Springer Science+Business Media New York 2014
Abstract The present study was conducted to assess theinfluence of dietary zinc nanoparticles (size 50 nm) on thegrowth, biochemical constituents, enzymatic antioxidantlevels and the nonspecific immune response of the freshwaterprawn, Macrobrachium rosenbergii post larvae (PL). Theconcentrations of dietary supplement zinc nanoparticles(ZnNPs) were 0, 10, 20, 40, 60 and 80 mg kg−1 with the basaldiet, and the level of Zn in ZnNP-supplemented diets were0.71, 10.61, 20.73, 40.73, 60.61 and 80.60 mg kg−1, respec-tively. ZnNP-incorporated diets were fed toM. rosenbergii PL(initial body weight, 0.18±0.02 g) in a triplicate experimentalsetup for a period of 90 days. ZnNP supplemented feed fed PLup to 60 mg kg−1 showed significantly (P<0.05) improvedperformance in survival, growth and activities of digestiveenzymes (protease, amylase and lipase). The concentrationsof biochemical constituents (total protein, total amino acid,total carbohydrate and total lipid), total haemocyte count anddifferential haemocyte count were elevated in 10–60 mg kg−1
ZnNP supplemented feed fed PL. However, the PL fed with80 mg ZnNPs kg−1 showed negative results. Activities ofenzymatic antioxidants [superoxide dismutase (SOD) andcatalase (CAT)], metabolic enzymes [glutamate–oxaloacetatetransaminase (GOT) and glutamate–pyruvate transaminase(GPT)] and the process of lipid peroxidation (LPO) in thehepatopancreas and muscle showed no significant alterationsin 10–60 mg kg−1 ZnNP supplemented feed fed PL. Whereas,80 mg ZnNPs kg−1 supplemented feed fed PL showed signif-icant elevations in SOD, CAT, LPO, GOT and GPT.
Therefore, 80 mg ZnNPs kg−1 was found to be toxic toM. rosenbergii PL. Thus, the study suggests that up to60 mg ZnNPs kg−1 can be supplemented for regulating sur-vival, growth and immunity of M. rosenbergii.
Keywords Macrobrachium rosenbergii . ZnNPs . Growth .
Survival . Biochemical constituents . Antioxidant status .
Nonspecific immune response
Introduction
The giant freshwater prawn, Macrobrachium rosenbergii isone of the edible organisms among crustaceans. It is a mainprawn species for small- and large-scale freshwater farmingdue to its better environment tolerance, rapid growth, largersize, better meat quality, omnivorous feeding habit,established domestic and export markets in Asian countriesespecially in China, India, Bangladesh, Thailand, Malaysia,Vietnam, etc.
Zinc (Zn) is an essential micronutrient for stabilizing cel-lular membranes and component of all organs, tissues andfluids of an organism. It is involved in several metabolicpathways as cofactors in many enzyme systems and compo-nent of a large number of metalloenzymes (carbonicanhydrase, carboxypeptidase, alcohol dehydrogenase,glutamic dehydrogenase, D-superoxide dismutase etc.) [1]. Itregulates protein synthesis and acts as a regulator of energy,vitamin A and lipid metabolism. Zn-dependent physiologicalfunctions are influenced by its transport and storage. It isstored and metabolized via hepatic metallothionein proteinswhich are cytosolic proteins with high-cysteine content. Asingle metalloprotein like metallothionein-1 is capable ofbinding seven atoms of Zn [2–5]. The dietary Zn requirements
T. Muralisankar : P. S. Bhavan (*) : S. Radhakrishnan :C. Seenivasan :N. Manickam :V. SrinivasanCrustacean Biology Laboratory, Department of Zoology, School ofLife Sciences, Bharathiar University, Coimbatore 641046, TamilNadu, Indiae-mail: [email protected]
Biol Trace Elem ResDOI 10.1007/s12011-014-0026-4
Author's personal copy

as well the resultant effect of its deficiency or excess arespecies dependent. The role Zn in better survival, feed intake,growth, immune response, antimicrobial activities and stresstolerance has been reported in fishes supplemented with 40–120 mg Zn kg−1 [2–4] and crustaceans with 15–40 mg Znkg−1 [5, 6]. However, excess intake of Zn can be harmful tohuman beings and animals. The excess absorption of Znsuppresses the utilization of Cu and Fe which reduces thegrowth and reproductive performance in fishes and prawns[6, 7]. Similarly, the toxic effects of Zn-oxidized nanoparticles(ZnONPs) produce oxidative stress in the shrimp,Thamnocephalus platyurus and fish, Cyprinus carpio andDanio rerio [8–10].
The aquaculture industries can be revolutionized by usingnanotechnology with new tools to enhance the ability ofcultivable organisms to uptake drugs like hormones, vaccinesand nutrients [11]. The metal nanoparticles (NPs) such as Se,Al, Fe, FeO, and ZnO play a crucial role in aquacultureoperations [12]. The present study was focussed to understandthe influence of dietary Zn nanoparticles (ZnNPs) on survival,growth, activities of digestive enzymes (protease, amylase andlipase), contents of biochemical constituents (total protein,amino acid, carbohydrate and lipid), activities of antioxidantenzymes [superoxide dismutase (SOD) and catalase (CAT)],lipid peroxidation (LPO) activities of metabolic enzymes[glutamate–oxaloacetate transaminase (GOT) and gluta-mate–pyruvate transaminase (GPT)], total haemocyte count(THC) and differential haemocyte count (DHC) inM. rosenbergii PL.
Materials and Methods
Feed Formulation
Diets were prepared with locally available feed ingredients.Fishmeal (400 g kg−1) and soybean meal (200 g kg−1) wereused as protein sources; wheat flour (180 g kg−1) and tapiocaflour (150 g kg−1) were used as carbohydrate sources; codliver oil (20 ml kg−1) was used as lipid source; tapioca flourand egg albumin were served as binding agents; vitamin Bcomplex with vitamin C (thiamine mononitrate IP 10 mg,riboflavin IP 10 mg, pyridoxine hydrochloride IP 3 mg, vita-min B12 (as stablets 1:100) IP 15mcg, niacinamide IP 100mg,calcium pantothenate IP 50 mg, folic acid IP 1.5 mg, biotinUSP 100 mcg, ascorbic acid IP 150 mg) was also added. Zinc-free mineral mix (CuSO4·5H2O, 6 mg; CaCO3, 164 mg;NaH2PO4·2H2O, 148 mg; KH2PO4·2H2O, 337.6 mg; CaCl2,66.64 mg; MgSO4·7H2O, 80 mg; KCl, 22.40 mg;AlCl3·6H2O, 0.96 mg; MnSO4·H2O, 11.45 mg; FeSO4·7H2O,90 mg; COCl2·6H2O, 1.41 mg; KI, 1.81 mg; cellulose,69.74 mg) was also added. The graded concentrations of
dietary ZnNPs were designed according to dietary zinc (Zn)requirements in crustaceans [3].
In this study, 50-nm-sized ZnNPs (Sigma-Aldrich) weresupplemented with the basal diet at concentrations of 0, 10,20, 40, 60 and 80 mg kg−1. The content of Zn present in theseZnNP-supplemented diets was analysed [13] and found to be0.71, 10.61, 20.40, 40.73, 60.61 and 80.60 mg kg−1 respec-tively (Table 1). The diets were prepared by thoroughlymixing the ingredient powders in sterilized water. The doughwas then cooked in a closed aluminium container at 105 °C for20 min followed by cooling at room temperature. Cod liveroil, vitamins and egg albumin were added and thoroughlymixed until a stiff dough was obtained. The dough waspelletized by an indigenous hand pelletizer with mesh sizeof 0.1-mm diameter (Pigeon manufactures, Kolkata, India)and was cut into 3.0±0.97-mm-sized pieces. The pellets weredried at room temperature (27 °C) until constant weight wasreached. The prepared feed was stored individually in airtightplastic containers at −20 °C until used for the feeding trials.The formulated feed contains the proximate composition inthe following ratio with the energy value of 14.75 kJ g−1:protein, 41.78 %; carbohydrate, 29.37 %; lipid, 7.69 %; ash,11.46 %; moisture, 8.56 % [13].
Feeding Trial
M. rosenbergii PL (PL-5) were procured fromAqua Hatchery,Koovathur, Kanchipuram District, Tamilnadu, India. Theywere safely transported to the laboratory in plastic bags half-filled with hatchery water and well oxygenated. They wereacclimatized to ambient laboratory conditions for 3 weeks in alarge cement tank (1,000 L) with groundwater (pH, 7.10±0.20; total dissolved solids, 0.94±0.07 g L−1; dissolved oxy-gen, 7.20±0.36 mg L−1; BOD, 36.00±1.45 mg L−1; COD,125.0±7.00 mg L−1; ammonia, 0.018±0.004 mg L−1). Theprawns were provided aeration. During which they were fedwith boiled egg albumin, Artemia nauplii and control feedprepared with basal ingredients alternatively three times perday, and three fourths of aquarium water was changed daily.
Six groups of PL ranging from 1.42±0.35-cm length; 0.18±0.02-g weight was assigned in triplicate. A group was servedas control and fed with ‘0’ concentration of ZnNP-supplemented diet. The remaining five groups were fed with10, 20, 40, 60 and 80 mg kg−1 of ZnNP-supplemented dietsrespectively. Each group consisted of 40 PL in an aquariummaintained with 40 l of groundwater. The water medium wasrenewed every 24 hr by siphoning method without severedisturbance to the prawn and aerated adequately. The experi-mental prawns were fed with these feeds at 10 % of bodyweight two times per day (6.00 a.m. and 6.00 p.m.). Duringfeeding trial as well as acclimatization, the unfed feed, faecesand moult (if any) were removed on a daily basis while
Muralisankar et al.
Author's personal copy
renewing aquarium water. The experiment was performed fora period of 90 days.
Analysis of Survival, Growth and Nutritional Indices
At the end of the feeding trial, the survival rate and food indexparameters such as feed intake, length gain, weight gain,specific growth rate, feed conversion ratio and protein efficien-cy ratio were individually determined by the following equa-tions [14]:
Survival rate %ð Þ ¼ No:of live prawns=no:of prawns introduced� 100Length gain cmð Þ ¼ Final length cmð Þ − initial length cmð ÞWeight gain gð Þ ¼ Final weight gð Þ − initial weight gð ÞFeed intake g d−1
� � ¼ Feed eaten gð Þ = total number of daysSpecific growth rate %ð Þ¼logfinal weight gð Þ−loginitial weight gð Þ =
total number of days � 100Feed conversion ratio gð Þ ¼ feed intake gð Þ = weight gain gð ÞProtein efficiency ratio gð Þ ¼ weight gain gð Þ = protein intake gð Þ
Assay of Digestive Enzymes
Activities of digestive enzymes such as protease, amylase andlipase were assayed on initial and final days of feeding trial.The whole flesh except eyestalk, appendages and exoskeletonwas homogenized in ice-cold distilled water and centrifuged at9,300 g under 4 °C for 20 min. The supernatant was used as acrude enzyme source.
The activity of protease was estimated by the method ofFurne et al. [15]. One unit of enzyme activity represents the
amount of enzyme required to liberate 1 μg of tyrosine perminute under assay conditions. The reactionmixture consistedof 0.25 ml of casein at 1 % (w/v), 0.25 ml of 0.1 M glycine–NaOH buffer (pH 10.0) and 0.1 ml enzyme source. Thereaction was incubated for 1 h at 37 °C, then the reactionwas stopped by adding 0.6 ml 8 % (w/v) trichloroacetic acidsolution and kept for 1 h at 2 °C, then centrifuged at 1,800gfor 10 min, and the supernatant absorbance was measured at280 nm against blank. For the blank preparation, the enzymesource was added at the end of the incubation period, justbefore adding trichloroacetic acid. Tyrosine solution was usedas standard.
Amylase activity was assayed by starch hydrolysis methodof Bernfeld [16] in which the increase in reducing power ofbuffered starch solutions was measured. The specific activityof amylase was calculated as milligrams of maltose liberatedper gram of protein per hour (mg/g/h). The reaction mixtureconsisted of 0.125 ml of 2 % (w/v) starch solution, 0.125 ml of0.1 M citrate–phosphate buffer (pH 7.5) and 0.5 ml enzymesource. The reaction was incubated at 37 °C for 1 h, and theabsorbance was measured at 600 nm against a blank. For theblank, the enzyme source was added just after the incubationperiod. Maltose solution was used as standard.
The activity of lipase was assayed by the method of Furneet al. [15]. One unit of lipase activity was defined as theamount of free fatty acid released from triacylglycerol per unittime estimated by the amount of NaOH required to maintainpH constant and represented as mille equivalents of alkaliconsumed. A solution of 1 % polyvinyl alcohol (PVA) and5 ml of 0.1 N HCl in 1 l of distilled water was heated to 75–85 °C, cooled, filtered and adjusted to pH 8.0 with 0.1 N
Table 1 Composition of minerals in ZnNP-supplemented diets and carcass of M. rosenbergii fed with ZnNP-supplemented diets
Dietary ZnNP (mg kg−1) Cu Zn Fe Ca Mg Na K
0 Formulated diet (mg kg-1) 1.75±0.11a 0.71±0.04f 2.23±0.13a 1.84±0.10a 2.03±0.20a 1.49±0.21a 1.24±0.13a
10 1.74±0.15a 10.61±0.67e 2.11±0.21a 1.61±0.20a 2.05±0.23a 1.42±0.12a 1.32±0.17a
20 1.68±0.11a 20.73±1.59d 2.08±0.18a 1.73±0.16a 2.03±0.24a 1.48±0.10a 1.2±0.12a
40 1.76±0.14a 40.73±1.75c 2.06±0.21a 1.76±0.15a 2.1±0.11a 1.52±0.21a 1.26±0.16a
60 1.75±0.12a 60.61±1.24b 2.14±0.17a 1.56±0.16a 2.06±0.18a 1.38±0.11a 1.23±0.13a
80 1.75±0.15a 80.60±1.85a 2.03±0.12a 1.65±0.13a 2.06±0.16a 1.49±0.22a 1.29±0.18a
F value 0.077 1555.09 0.036 0.023 0.088 0.279 0.091
0 Carcass prawn (μg g−1) 37.05±1.28e 19.75±1.30f 26.25±2.01e 32.85±2.07f 78.50±3.11e 124.65±2.44f 146.28±3.01de
10 75.10±3.29d 69.05±4.01e 44.00±2.10d 117.35±5.04e 121.60±4.02d 144.65±2.84d 151.58±3.42d
20 81.15±4.30c 78.90±4.02d 50.45±2.35c 142.8±4.23d 133.60±4.65c 151.92±2.47c 162.27±3.21c
40 87.95±3.01b 138.60±4.07c 63.95±2.21b 156.1±5.32c 187.05±5.54b 174.49±3.14b 186.91±3.01b
60 95.25±3.19a 151.30±3.62b 68.4±2.35a 270.45±6.12a 196.65±5.76a 194.83±3.18a 203.78±4.61a
80 86.60±2.62bc 190.60±4.02a 45.95±1.83d 175.75±2.76b 137.60±3.54c 117.68±3.12e 156.92±3.91d
F value 136.405 892.317 149.263 894.295 277.667 321.743 195.752
Each value is mean ± SD of three individual observations; mean values within the same column sharing different alphabetical letter superscripts arestatistically significant at P<0.05 (one-way ANOVA and subsequent post hoc multiple comparisonwith DMRT); a–f, order of performance (a > b > c > d> e > f); some of the mean sharing two superscripts means that it falls into two ranks/columns
ZnNPs induced growth performance in prawn
Author's personal copy

NaOH. Virgin olive oil was added to an aliquot of this solutionfor obtaining 0.1 M substrate concentration. This mixture wasemulsified for 5 min. In addition, Mcllvaine buffer was pre-pared from 0.1 M citric acid and 0.2 M disodium phosphate.The reaction mixture consisted 1 ml of PVA-solution-emulsified substrate, 0.5 ml of Mcllvaine buffer (pH 8.0)and 0.5 ml enzyme source and was incubated at 37 °C for4 h. To stop the reaction and break the emulsion, 3 ml of 1:1ethanol-acetone solution was added. To the reaction mixture,phenolphthalein in ethanol 1 % (w/v) was added and titratedagainst 0.01 N NaOH. For the blanks, the same procedure wasfollowed but boiled enzyme was used.
Estimation of Biochemical Constituents and Carcass MineralContents
Analyses of total nitrogen, crude protein, moisture and ashcontents were performed according to standard AOAC proce-dures [13]. Dry matter was obtained by drying at 105 °C untila constant weight was achieved. Ash content was obtained byburning in a muffle furnace at 600 °C for 12 h. Total nitrogenand crude protein (N*6.25) were analysed after single aciddigestion using Kjeldhal apparatus (model: Kelplus DISTYL-BS, manufactured by Pelican Equipments Pvt. Ltd. Chennai,India).
Concentration of total protein was estimated by the methodof Lowry et al. [17], using ethanolic precipitated sample. Theblue colour was result of biuret reaction of protein with copperions in alkali solution and reduction of the phophomolybdic-phosphotungstic of Folin reagent by the tyrosine andtrphtophan present in the treated protein. Bovine serum albu-min (BSA) was used as a standard. This colour intensity wasmeasured at 650 nm against a blank which is devoid of proteinsample.
Concentration of total amino acid was estimated by themethod of Moore and Stein [18]. The total amino acid wasextracted with sodium tungstate and H2SO4. When aminoacids are heated with ninhydrine, they undergo deamination.The reaction of amino acid-hydratin complex with ninhydrinproduced purple colour, which was measured at 540 nm.Leusine was used as standard.
Concentration of total carbohydrate was estimated by themethod of Roe [19] using TCA extracted sample. Carbohy-drates are hydrolysed into simple sugars by diluted HCl in hotacidic medium. Glucose is dehydrated into hydroxyl-methylfurfural. This compound reacts with anthrone and produced agreen-coloured product, which was measured at 630 nm. Glu-cose was used as standard.
Total lipid was extracted with chloroform–methanol mix-ture following the method of Folch et al. [20] and estimated bythe method of Barnes and Blackstock [21]. Lipid reacts withvanillin in a medium of H2SO4 and phosphoric acid to form apink-coloured chromogen, which is proportional to the lipid
content of the sample and measured at 540 nm. Olive oil wasused as standard.
The carcass mineral contents including trace elements wereanalysed using the atomic absorption spectrophotometer(AAS) (PerkinElmer, model 2380) in air acetylene flame byadopting triple acid digestion method [13].
Total and Differential Haemocyte Counts
At the end of the experiment, 100 μl of haemolymph waswithdrawn from the ventral sinus in the first abdominal seg-ment using a 26-gauge hypodermic needle on a 1-ml syringe.Each syringe was pre-filled with 200 μl of anticoagulant(10 mM Tris-HCl, 250 mM sucrose, 100 mM sodium citrate,pH 7·6). More anticoagulants were added to make the volumeup to 1 ml in the anticoagulated haemolymph. Further, avolume of 200 μl anticoagulated haemolymph was fixed withan equal volume of formalin (10 %) for 30 min. The fixedhaemolymph was used to THC and DHC determinations.
For THC, 100 μl of fixed haemolymph was diluted at 1:2ratio (v/v) with ice-cold phosphate-buffered saline (PBS,20 mM, pH 7·2). The diluted haemolymph was stained with20 μl of Rose Bengal strain (1.2 % Rose Bengal in 50 %ethanol) and incubated at room temperature for 20 min. THCwas determined by haemocytometer (Neubauer improved,Germany) under the light microscope at RP 1000× (Labomed,CXR2).
THC �106cells ml−1� � ¼ Counted cells� depth of chamber � dilution factor
Number of 1−mm square
For DHC, fixed haemolymph was stained with Rose Ben-gal solution (10 %) for 10 min and smeared on a slide. Thenumbers of differential haemocytes, such as hyalinocytes,semigranulocytes and granulocytes, were characterised ac-cording to Tsing et al. [22], and 350–400 cells from eachsmear were counted under a Trinocular Inverted Microscope(model number INVERSO 3000) RP 1000×.
Activities of Enzymatic Antioxidants and Lipid Peroxidation
The muscles and hepatopancreas of test prawns were individ-ually homogenised (10 %w/v) in ice-cold 50-mM Tris buffer(pH 7.4), centrifuged at 9,300g for 20 min at 4 °C, and thesupernatant was used to assay the enzyme activities. Solubleprotein concentration was determined by the method of Lowryet al. [17] using BSA as the standard.
Superoxide dismutase (SOD) activity was measured usingpyrogallol (10 mM) autoxidation in Tris buffer (50 mM, pH7.0) [23]. The assay mixture contained 1.2 ml of sodiumpyrophosphate buffer, 0.1 ml of phenazine methosulphate(PMS), 0.3 ml of nitro blue tetrazolium (NBT), 0.2 ml of theenzyme prepared and water accounting to a total volume of2.8 ml. The reaction was initiated by the addition of 0.2 ml
Muralisankar et al.
Author's personal copy

NADH. The mixture was incubated at 30 °C for 90 s andarrested by the addition of 1.0 ml glacial acetic acid. Thereaction mixture was then shaken with 4.0 ml n-butanol,allowed to stand for 10 min and centrifuged. The intensity ofthe chromogen in the butanol layer was measured at 560 nmusing spectrophotometer. The specific activity of the enzymewas expressed in U/mg protein.
Catalase (CAT) activity was measured using H2O2 as thesubstrate in phosphate buffer [24]. The reaction was initiatedby the addition of 1.0 ml of phosphate buffer (0.01M, pH 7.1),0.5 ml of H2O2 (0.2 M) and 0.4 ml of distilled water succes-sively to 0.5 ml of tissue homogenate. After 60 s, the reactionwas stopped by the addition of 2.0 ml of dichromate-aceticacid reagent. Further, the tubes were kept in a boiling waterbath for 10 min and cooled at room temperature. The absor-bance of the chromophore was read at 620 nm. A systemdevoid of enzyme served as the control. The activity ofcatalase was expressed as micromoles of hydrogen peroxideconsumed per minute per milligram of protein.
Lipid peroxidation (LPO) in the tissue homogenates wasmeasured by estimating the formation of thiobarbituric-acid-reactive substances (TBARS) [25]. The tubes containing 1 mlof tissue homogenate (10 %w/v in 50 mM phosphate buffer,pH 7.4) was subsequently mixed with 1 ml of Tris buffer(0.02 M, pH 7.5), 1 ml of 10 % tricholoro acetic acid and1.5 ml of thiobarbituric acid (1.5%). The reactionmixture wasboiled for 15 min in the boiling water bath and cooled at roomtemperature. The content was centrifuged at 100g for 20 minand the supernatant was collected. The absorbance of thesupernatant was measured at 535 nm against the reagentblank. TBARS was expressed as nanomoles ofmalondialdehyde (MDA) per milligram of protein.
Activities of Metabolic Enzymes
The metabolic enzymes such as glutamic oxaloacetate trans-aminase (GOT) and glutamic pyruvate transaminase (GPT)were analysed according to the method of Reitman andFrankel [26] using a media source kit (Medsource OzoneBiomedicals Pvt. Ltd. Haryana, India). A total of 100 mg ofmuscle and hepatopancreas tissues was homogenized in0.25 M sucrose and centrifuged at 3,300g for 20 min in ahigh-speed cooling centrifuge at 4 °C. The supernatant wasused as the enzyme source.
GOT Analysis
The substrate solution, L-aspartic acid (500 μl, pH 7.4) wasadded with 100 μl sample and incubated at 37 °C for 1 h.Then, 500 μl of 2,4-dinitrophenyl hydrazine was added andallowed to stand for 20 min at room temperature, then 3 ml offreshly prepared 4 N sodium hydroxide solution was added tothe above solution. The colour development was read at
505 nm using spectrophotometer within 15 min. Sodiumpyruvate (160 U/l) was used as a calibrator. The activity ofGOTwas expressed as units per litre (U/l).
GPT Analysis
Buffered L-alanine, 2-oxoglutarate substrate (500 μl; pH 7.4)were added with 100-μl sample and incubated at 37 °C for20 min. With this, 500 μl of 2,4-dinitrophenyl hydrazine wasadded and allowed to stand at room temperature for 30 min,then 3 ml of freshly prepared 4 N sodium hydroxide solutionwas added. The colour development was read at 505 nm usingspectrophotometer within 15 min. Sodium pyruvate (170 U/l)was used as a calibrator. The activity of GPTwas expressed asunits per litre (U/l).
Statistical Analysis
The data were analysed by one-way analysis of variance(ANOVA) using SPSS (16.0), followed by Duncan’s multiplerange test to compare the differences among treatments wheresignificant differences (P<0.05) were observed. Data wereexpressed as mean ± SD.
Results
Content of Minerals
In formulated feeds, the levels of mineral salts, such as Cu, Fe,Ca, Mg, Na and K, were almost similar, whereas, in PL fedwith these diets, the levels of these mineral salts were signif-icantly increased (P<0.05) up to the level of 60 mg ZnNPskg−1 supplementation when compared with control (Table 1).
Survival, Growth, Nutritional Indices and Activitiesof Digestive Enzymes
In this study, the survival, growth, and other nutritional indicesparameters, such as feed intake, specific growth rate andprotein efficiency ratio, were significantly elevated (P<0.05)in PL fed with 10, 20, 40, 60 mg ZnNPs kg−1 supplementeddiets (Table 2) when compared with control. Among theseconcentrations, 60mg ZnNP kg−1 showed better performance.This was further confirmed through the observed lower feedconversion ratio in 60 mg ZnNP supplemented diet fed PLgroup when compared with control as well as other concen-trations of ZnNP supplemented diet fed PL.
In the present study, activities of protease, amylase andlipase were significantly increased (P<0.05) in PL fed with10–60 mg ZnNPs kg−1 incorporated diets when comparedwith control (Table 2). A maximum increase was recorded in
ZnNPs induced growth performance in prawn
Author's personal copy
60 mg ZnNPs kg−1 incorporated diet fed PL. Whereas, the PLfed with 80 mg ZnNPs kg−1 incorporated diet acts as nega-tively influenced on activities of these enzymes. The activityof protease was found to be significantly higher when com-pared with control, but significantly lower when comparedwith 60 mg ZnNPs kg−1 incorporated diet fed PL. The activityof amylase was found to be lower in 80 mg ZnNPs kg−1
incorporated diet fed PL when compared with control, butthis was not statistically significant, whereas the decrease wasstatistically significant when compared with 60 mg ZnNPskg−1 incorporated diet fed PL. In the case of lipase activity,insignificant difference was seen between control and 80 mgZnNPs kg−1 incorporated diet fed PL, whereas the activity oflipase in 80 mg ZnNPs kg−1 incorporated diet fed PL showeda significant decrease when compared with 60 mg ZnNPskg−1 (Table 2).
Concentrations of Biochemical Constituents
In the present study, the percentage of total nitrogen and crudeprotein in ZnNP supplemented diet fed PL groups are present-ed in Table 3, and their levels were found to be graduallyelevated up to 60 mg ZnNPs kg−1 supplementation whencompared with control (P<0.05), whereas, their levels werefound to be declined in 80 mg ZnNPs kg−1 supplementationdiet fed PL. Similarly, the concentrations of total protein, total
amino acids, total carbohydrate, total lipid and ash weresignificantly increased (P<0.05) in PL fed with 20, 40 and60 mg ZnNPs kg−1 supplemented diets (Table 3) when com-pared with control, whereas their levels were found to bedeclined in 80 mg ZnNPs kg−1 supplementation diet fed PL.
Haemocytes Population
THC and DHC (hyalinocytes, semigranulocytes andgranulocytes) were also found to be elevated significantly(P<0.05) in PL fed with 20–60mg ZnNPs kg−1 supplementeddiets when compared with control (Table 3). In the case of80 mg ZnNPs kg−1 supplementation diet fed PL, the THC andDHC were found to be declined (Table 3).
Activities of Antioxidant and Metabolic Enzymes
In this study, no significant changes in activities of SOD andCAT were recorded in PL fed with 10–60 mg ZnNPs kg−1
supplemented diets throughout the sampling days (Tables 4and 5). Similarly, no significant changes in activities of GOTand GPT were recorded (Tables 4 and 5). The level of LPOalso showed no significant alteration up to 60mg ZnNPs kg−1.In the case of 80 mg ZnNPs kg−1 supplemented diet fed PL,the activities of SOD, CAT, GOT and GPT, along with the
Table 2 Nutritional indices and activities of digestive enzymes in M. rosenbergii fed with ZnNP-supplemented diets
Parameters Dietary ZnNP (mg kg−1)
0 10 20 40 60 80 F value
SR (%) 75.83±1.84c 76.66±3.81c 78.33±3.81bc 83.33±2.88ab 87.5±2.50a 68.33±3.81d 12.993
Length (cm) Time 0 1.42±0.35 1.42±0.35 1.42±0.35 1.42±0.35 1.42±0.35 1.42±0.35 –
Final 3.64±0.28c 5.02±0.55b 5.20±0.33b 5.62±0.65b 6.56±0.69a 5.76±0.80b 13.735
Weight (g) Time 0 0.18±0.02 0.18±0.02 0.18±0.02 0.18±0.02 0.18±0.02 0.18±0.02 –
Final 0.58±0.13c 1.06±0.43bc 1.14±0.27bc 1.40±0.51b 2.28±0.74a 1.60±0.60b 7.190
LG (cm) 3.29±0.32e 4.67±0.27d 4.85±0.24cd 5.27±0.31bc 6.21±0.29a 5.41±0.30b 33.958
WG (g) 0.4±0.01f 0.88±0.02e 0.96±0.02d 1.22±0.03c 2.10±0.08a 1.42±0.04b 570.59
FR (g d−1) 0.39±0.01d 0.45±0.02c 0.47±0.01c 0.50±0.01b 0.66±0.02a 0.49±0.02b 69.641
SGR (%) 0.55±0.03d 0.84±0.07c 0.87±0.08bc 0.97±0.06b 1.21±0.07a 0.94±0.06bc 30.011
FCR (g) 2.69±0.35a 1.15±0.13b 1.10±0.21b 0.92±0.09bc 0.70±0.03d 0.78±0.07d 50.945
PER (%) 1.22±0.06c 3.14±0.15b 3.2±0.18b 3.58±0.32b 4.88±0.38a 4.58±0.27a 80.504
Digestive enzymes(U/mg protein)
Protease Time 0 0.27±0.06 0.27±0.06 0.27±0.06 0.27±0.06 0.27±0.06 0.27±0.06 –
Final 1.00±0.12c 1.37±0.16b 1.44±0.15b 1.59±0.19b 2.07±0.21a 1.34±0.20b 12.674
Amylase Time 0 0.16±0.04 0.16±0.04 0.16±0.04 0.16±0.04 0.16±0.04 0.16±0.04 –
Final 0.75±0.11c 0.81±0.10bc 0.96±0.12b 0.97±0.14b 1.47±0.21a 0.67±0.08c 16.693
Lipase × 102 Time 0 0.74±0.05 0.74±0.05 0.74±0.05 0.74±0.05 0.74±0.05 0.74±0.05 –
Final 0.22±0.02e 0.31±0.04d 0.37±0.02c 0.48±0.01b 0.55±0.02a 0.23±0.04e 71.680
Each value is mean ± SD of three individual observations; mean values within the same row sharing the different alphabetical letter superscripts arestatistically significant at P<0.05 (one-way ANOVA and subsequent post hoc multiple comparisonwith DMRT); a–f, order of performance (a > b > c > d> e > f); some of the mean sharing two superscripts means that it falls into two ranks/columns. SR, survival rate; LG, length gain;WG,weight gain; SGR,specific growth rate; FCR, feed conversion ratio; PER, protein efficiency ratio
Muralisankar et al.
Author's personal copy
level of LPO, showed significant elevations (P<0.05) whencompared with control and other diets (Tables 4 and 5).
Discussion
Zinc has a generally stable association with macromolecules,like protein, nucleic acid, carbohydrate and lipid and enzymes,to carry out diverse biological functions. Zinc acts as anelectron acceptor, which contributes to its catalytic activityin metalloenzymes, such as DNA and RNA polymerases,alcohol dehydrogenase, carbonic anhydrase, and alkalinephosphatase. Zn plays an important role in the functions oftranscription factor, antioxidant defense system and DNArepair. Dietary deficiencies of zinc can contribute to single-and double-strand DNA breaks and oxidative modifications toDNA that increase risk for cancer development [27]. Zndeficiency increases the levels of lipid peroxidation in mito-chondrial and microsomal membranes and the osmotic fragil-ity of erythrocyte membranes, while the presence of Zn
prevents lipid peroxidation and thus plays an important rolein protecting the cells from oxidative stress [28]. According toHedemann et al. [29], Zn promotes the processes of tissuerepair in the small intestine and stimulates the synthesis ofdigestive enzymes, resulting in a better digestion and absorp-tion of nutrients and potentially improving growth perfor-mance in pig.
The results of mineral contents clearly indicated thatthe supplementation of ZnNPs promotes mineral absorp-tion based on its level of supplementation. Increased ironand phosphorus utilization have been reported in Haliotisdiscus hannai and Oncorhynchus mykiss due to Zn sup-plementation [3, 30]. The supplementation of Cu, Zn andK in the diet of Gadus morhua showed maximum utiliza-tion of Cu, Zn, K, Mg and Ca [5]. In the present study,80 mg ZnNPs kg−1 supplemented diet fed PL showedpoor utilization of minerals (Cu, Fe, Ca, Mg, Na and K)when compared with 40–60 mg ZnNPs kg−1. This clearlyindicates the fact that mineral utilization by prawn wasbased on ZnNP concentration. A study by Rani et al. [8]indicated that Zn supplemented feed fed Carassius
Table 3 Concentrations of biochemical constituents and population of haemocyte (THC and DHC) in M. rosenbergii fed with ZnNP-supplementeddiets
Parameters Dietary ZnNP (mg kg−1)
0 10 20 40 60 80 F value
Total Nitrogen (%) Time 0 4.10±0.18 4.10±0.18 4.10±0.18 4.10±0.18 4.10±0.18 4.10±0.18 –
Final 8.17±0.36e 8.49±0.32de 8.82±0.28cd 9.45±0.19b 10.89±0.14a 9.05±0.22bc 39.121
Crude protein (%) Time 0 25.66±1.16 25.66±1.16 25.66±1.16 25.66±1.16 25.66±1.16 25.66±1.16 –
Final 51.10±2.29e 53.08±2.02de 55.12±1.75cd 59.09±1.24b 68.10±0.87a 56.58±1.42bc 38.988
Protein(mg/g wet wt.)
Time 0 46.64±2.14 46.64±2.14 46.64±2.14 46.64±2.14 46.64±2.14 46.64±2.14 –
Final 148.79±2.41e 169.6±3.76d 185.01±4.62c 199.0±3.10b 256.0±4.49a 174.02±4.43d 183.28
Amino acid(mg/g wet wt.)
Time 0 25.33±2.30 25.33±2.30 25.33±2.30 25.33±2.30 25.33±2.30 25.33±2.30 –
Final 90.00±3.46e 103.40±3.07d 116.66±3.05c 125.40±4.13b 155.40±2.94a 107.33±3.05d 102.39
Carbohydrate(mg/g wet wt.)
Time 0 18.07±1.07 18.07±1.07 18.07±1.07 18.07±1.07 18.07±1.07 18.07±1.07 –
Final 32.07±1.88e 40.61±2.34d 44.36±1.82c 50.11±1.42b 58.56±2.99a 41.31±2.26d 51.600
Lipid(mg/g wet wt.)
Time 0 10.68±0.74 10.68±0.74 10.68±0.74 10.68±0.74 10.68±0.74 10.68±0.74 –
Final 18.03±2.69c 21.70±2.18bc 25.98±2.94ab 27.09±1.26b 29.23±1.42a 23.84±2.85ab 7.673
Ash (%) Time 0 10.40±0.69 10.40±0.69 10.40±0.69 10.40±0.69 10.40±0.69 10.40±0.69 –
Final 15.50±1.37b 17.00±1.04a 17.43±0.87a 17.44±0.92a 17.93±2.10a 15.73±1.85b 1.432
Moisture (%) Time 0 78.33±1.52 78.33±1.52 78.33±1.52 78.33±1.52 78.33±1.52 78.33±1.52 –
Final 75.00±4.00a 72.00±3.60a 70.23±2.35a 68.00±4.35a 63.33±2.51a 74.33±2.08a 3.415
THC (×106 cells ml−1 ) 4.49±0.13d 4.84±0.21cd 5.75±0.11bc 6.42±0.15b 8.61±0.32a 4.69±0.15d 18.760
DHC(×106 cells ml−1 )
Hyalinocytes 2.10±0.11d 2.30±0.13d 2.71±0.12d 3.11±0.17b 4.10±0.16a 2.26±0.10d 74.862
Semigranulocytes 1.40±0.10c 1.50±0.12c 1.75±0.11b 1.82±0.13b 2.50±0.17a 1.35±0.12c 33.251
Granulocytes 0.08±0.08c 1.01±0.11c 1.15±0.13b 1.23±0.12b 1.54±0.11a 0.94±0.10c 21.577
Each value is mean ± SD of three individual observations; mean values within the same row sharing the different alphabetical letter superscripts arestatistically significant atP<0.05 (one-wayANOVA and subsequent post hocmultiple comparison with DMRT); a–e, order of performance (a > b > c > d> e); some of the mean sharing two superscripts means that it falls into two ranks/columns
THC total haemocyte count, DHC differential haemocyte count
ZnNPs induced growth performance in prawn
Author's personal copy
auratus showed significant elevation in Fe and Cu con-tents only up to 60 mg Zn kg−1.
In the present study, the increased survival, weight gain,feed intake, specific growth rate and protein efficiency ratioindicate that ZnNPs have the ability to promote the survival,feed intake and growth of M. rosenbergii PL. The lower feedconversion ratio recorded in 60 mg ZnNPs kg−1 indicates theenhanced quality of the feed. The poor growth performance incontrol diet fed PL suggests deficiency of Zn. Zn deficiencyleads to cause slow growth and increased mortality in fish[31]. Poor survival, feed intake and growth recorded in 80 mgZnNPs kg−1 supplemented feed fed M. rosenbergii PL mightbe due to its toxicity associated with excess amount of ZnNPsin the diet. Concentration-dependent positive and negativeeffects of Zn-supplemented diets have been reported in fishes,Oreochromis niloticus, G. morhua, C. auratus and Cirrhinusmrigala, the shrimpPenaeus monodon and the Chinese mittencrab Eriochei sinensis [4–8, 32, 33].
The increased activity of digestive enzymes recorded indi-cates that supplemented ZnNPs have influence on digestiveenzyme secretion in M. rosenbergii PL. In support of thisobservation, increased activities of protease, amylase andlipase have been reported in Zn supplemented feed fed tilapia,O. niloticus and Oreochromis aureus [34]. It has also been
reported in rats and weaning pigs that dietary Zn has elevatedthe activity of maltase, pepsin, amylase and protease [29, 35].It is important to mention here that the digestive enzyme incrustacean plays an essential role in nutritional physiology andregulates the growth and moult cycle directly or indirectly[36].
The elevation recorded in biochemical constituents, such asprotein, amino acid, carbohydrate, lipid and ash, suggests thatdietary ZnNPs have influenced the absorption of nutrients andincrease the synthesis of these basic biochemical constituentsin M. rosenbergii PL. Concentration-based increase and de-crease in protein, lipid, fatty acid and ash contents have beenreported in Zn supplemented diet fed fishes G. morhua andO. niloticus [4, 5], the crab E. sinensis [33] and the gastropodH. discus hannai [3]. It is worthy tomention here that Zn playsa vital role in lipid, protein, carbohydrate and nucleic acidmetabolism [37].
The health status of an organism is established by itsimmunological responses. In aquatic organisms, the immunestatus can be determined by studying the haematologicalparameters like blood cell counts. Few researchers have prov-en the immune stimulating ability of minerals in aquaticorganisms [38, 39]. In the present study, the elevated THCand DHC (hyalinocytes, semigranulocytes and granulocytes)
Table 4 Activities of antioxidant and metabolic enzymes in the muscle of M. rosenbergii fed with ZnNP-supplemented diets
Enzymes and LPO Dietary ZnNP (mg/100 g)
0 1 2 4 6 8 F value
Initial SOD (μmol/min/mg protein) 4.81±1.01 4.81±1.01 4.81±1.01 4.81±1.01 4.81±1.01 4.81±1.01 –
CAT (U/mg protein) 11.20±1.01 11.20±1.01 11.20±1.01 11.20±1.01 11.20±1.01 11.20±1.01 –
GOT (U/l) 6.32±0.67 6.32±0.67 6.32±0.67 6.32±0.67 6.32±0.67 6.32±0.67 –
GPT (U/l) 7.95±1.01 7.95±1.01 7.95±1.01 7.95±1.01 7.95±1.01 7.95±1.01 –
LPO (nmol MDA/mg protein) 0.13±0.010 0.13±0.01 0.13±0.01 0.13±0.01 0.13±0.01 0.13±0.01 –
30 days SOD (μmol/min/mg protein) 8.28±1.17b 8.29±1.43b 8.29±1.52b 8.31±1.06b 8.33±1.44b 12.72±1.48a 5.275
CAT (U/mg protein) 20.90±1.23b 20.92±1.32b 21.36±2.01b 21.36±2.00b 21.45±1.87b 26.37±2.31a 4.037
GOT (U/l) 8.53±1.11b 8.53±0.52b 8.53±0.60b 8.54±0.79b 8.50±1.08b 12.19±1.42a 7.032
GPT (U/l) 9.21±1.08b 9.25±1.28b 9.26±1.33b 9.25±1.17b 9.26±1.16b 12.23±0.79a 3.339
LPO (nmol MDA/mg protein) 0.63±0.02b 0.63±0.01b 0.64±0.01b 0.65±0.02b 0.67±0.02b 1.68±0.01a 989.11
60 Days SOD (μmol/min/mg protein) 8.27±1.24b 8.33±1.19b 8.40±1.23b 8.44±1.16b 8.52±1.06b 14.55±1.45a 12.602
CAT (U/mg protein) 20.92±1.32b 20.95±1.42b 21.37±1.54b 21.40±1.34b 21.46±1.97b 27.64±2.34a 7.698
GOT (U/l) 8.53±1.12b 8.52±0.49b 8.51±0.48b 8.53±0.74b 8.53±1.10b 14.51±2.28a 12.322
GPT (U/l) 9.22±0.83b 9.26±1.27b 9.26±1.08b 9.25±1.32b 9.26±1.26b 13.44±2.61a 3.867
LPO (nmol MDA/mg protein) 0.63±0.01b 0.63±0.01b 0.63±0.01b 0.64±0.01b 0.65±0.01b 1.91±0.16a 57.484
90 Days SOD (μmol/min/mg protein) 8.30±1.46b 8.44±1.01b 8.51±1.23b 8.59±1.11b 8.70±1.47b 15.77±1.82a 13.679
CAT (U/mg protein) 20.97±1.44b 21.42±1.35b 21.66±1.40b 21.68±1.50b 21.79±1.33b 29.52±2.03a 13.886
GOT (U/l) 8.54±1.13b 8.54±0.56b 8.54±0.61b 8.52±0.75b 8.53±1.07b 15.66±3.88a 8.112
GPT (U/l) 9.23±1.13b 9.25±1.30b 9.26±1.39b 9.26±1.26b 9.27±1.33b 16.62±2.93a 9.596
LPO (nmol MDA/mg protein) 0.63±0.02b 0.64±0.02b 0.65±0.01b 0.67±0.01b 0.68±0.02b 2.37±0.02a 1019.7
Each value is mean ± SD of three individual observations; mean values within the same row sharing the different alphabetical letter superscripts arestatistically significant at P<0.05 (one-way ANOVA and subsequent post hoc multiple comparison with DMRT); a–b, order of performance (a > b)
Muralisankar et al.
Author's personal copy
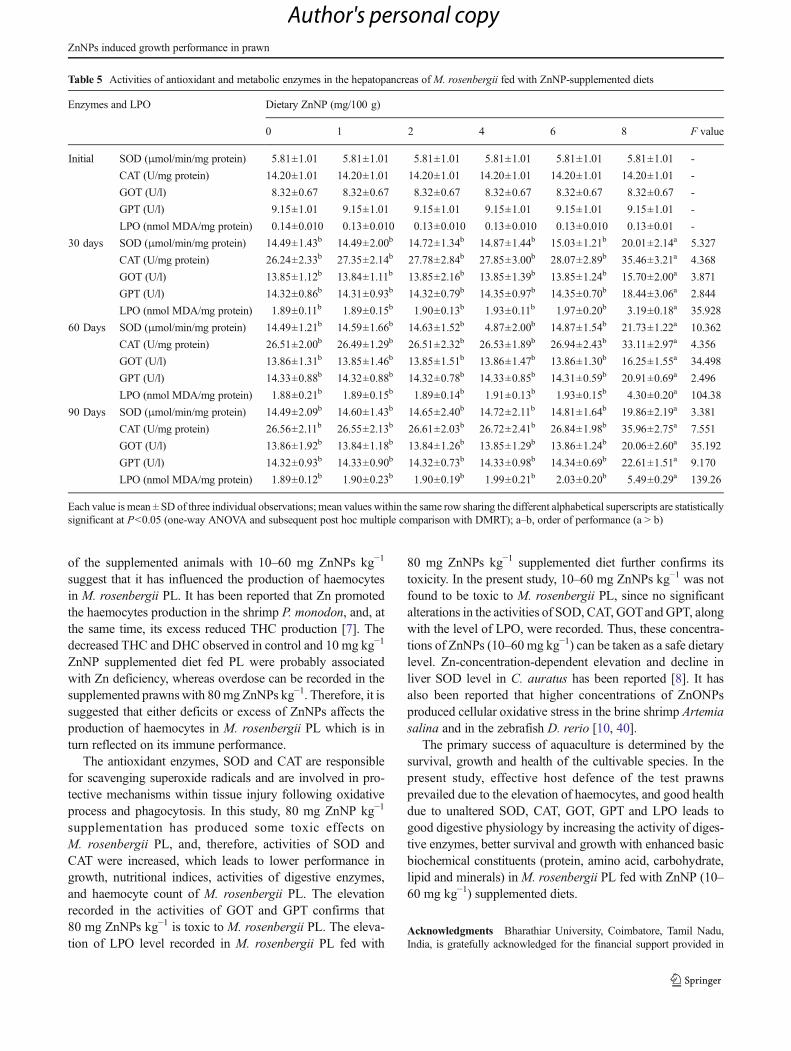
of the supplemented animals with 10–60 mg ZnNPs kg−1
suggest that it has influenced the production of haemocytesin M. rosenbergii PL. It has been reported that Zn promotedthe haemocytes production in the shrimp P. monodon, and, atthe same time, its excess reduced THC production [7]. Thedecreased THC and DHC observed in control and 10 mg kg−1
ZnNP supplemented diet fed PL were probably associatedwith Zn deficiency, whereas overdose can be recorded in thesupplemented prawns with 80mg ZnNPs kg−1. Therefore, it issuggested that either deficits or excess of ZnNPs affects theproduction of haemocytes in M. rosenbergii PL which is inturn reflected on its immune performance.
The antioxidant enzymes, SOD and CAT are responsiblefor scavenging superoxide radicals and are involved in pro-tective mechanisms within tissue injury following oxidativeprocess and phagocytosis. In this study, 80 mg ZnNP kg−1
supplementation has produced some toxic effects onM. rosenbergii PL, and, therefore, activities of SOD andCAT were increased, which leads to lower performance ingrowth, nutritional indices, activities of digestive enzymes,and haemocyte count of M. rosenbergii PL. The elevationrecorded in the activities of GOT and GPT confirms that80 mg ZnNPs kg−1 is toxic to M. rosenbergii PL. The eleva-tion of LPO level recorded in M. rosenbergii PL fed with
80 mg ZnNPs kg−1 supplemented diet further confirms itstoxicity. In the present study, 10–60 mg ZnNPs kg−1 was notfound to be toxic to M. rosenbergii PL, since no significantalterations in the activities of SOD, CAT, GOTandGPT, alongwith the level of LPO, were recorded. Thus, these concentra-tions of ZnNPs (10–60 mg kg−1) can be taken as a safe dietarylevel. Zn-concentration-dependent elevation and decline inliver SOD level in C. auratus has been reported [8]. It hasalso been reported that higher concentrations of ZnONPsproduced cellular oxidative stress in the brine shrimp Artemiasalina and in the zebrafish D. rerio [10, 40].
The primary success of aquaculture is determined by thesurvival, growth and health of the cultivable species. In thepresent study, effective host defence of the test prawnsprevailed due to the elevation of haemocytes, and good healthdue to unaltered SOD, CAT, GOT, GPT and LPO leads togood digestive physiology by increasing the activity of diges-tive enzymes, better survival and growth with enhanced basicbiochemical constituents (protein, amino acid, carbohydrate,lipid and minerals) in M. rosenbergii PL fed with ZnNP (10–60 mg kg−1) supplemented diets.
Acknowledgments Bharathiar University, Coimbatore, Tamil Nadu,India, is gratefully acknowledged for the financial support provided in
Table 5 Activities of antioxidant and metabolic enzymes in the hepatopancreas of M. rosenbergii fed with ZnNP-supplemented diets
Enzymes and LPO Dietary ZnNP (mg/100 g)
0 1 2 4 6 8 F value
Initial SOD (μmol/min/mg protein) 5.81±1.01 5.81±1.01 5.81±1.01 5.81±1.01 5.81±1.01 5.81±1.01 -
CAT (U/mg protein) 14.20±1.01 14.20±1.01 14.20±1.01 14.20±1.01 14.20±1.01 14.20±1.01 -
GOT (U/l) 8.32±0.67 8.32±0.67 8.32±0.67 8.32±0.67 8.32±0.67 8.32±0.67 -
GPT (U/l) 9.15±1.01 9.15±1.01 9.15±1.01 9.15±1.01 9.15±1.01 9.15±1.01 -
LPO (nmol MDA/mg protein) 0.14±0.010 0.13±0.010 0.13±0.010 0.13±0.010 0.13±0.010 0.13±0.01 -
30 days SOD (μmol/min/mg protein) 14.49±1.43b 14.49±2.00b 14.72±1.34b 14.87±1.44b 15.03±1.21b 20.01±2.14a 5.327
CAT (U/mg protein) 26.24±2.33b 27.35±2.14b 27.78±2.84b 27.85±3.00b 28.07±2.89b 35.46±3.21a 4.368
GOT (U/l) 13.85±1.12b 13.84±1.11b 13.85±2.16b 13.85±1.39b 13.85±1.24b 15.70±2.00a 3.871
GPT (U/l) 14.32±0.86b 14.31±0.93b 14.32±0.79b 14.35±0.97b 14.35±0.70b 18.44±3.06a 2.844
LPO (nmol MDA/mg protein) 1.89±0.11b 1.89±0.15b 1.90±0.13b 1.93±0.11b 1.97±0.20b 3.19±0.18a 35.928
60 Days SOD (μmol/min/mg protein) 14.49±1.21b 14.59±1.66b 14.63±1.52b 4.87±2.00b 14.87±1.54b 21.73±1.22a 10.362
CAT (U/mg protein) 26.51±2.00b 26.49±1.29b 26.51±2.32b 26.53±1.89b 26.94±2.43b 33.11±2.97a 4.356
GOT (U/l) 13.86±1.31b 13.85±1.46b 13.85±1.51b 13.86±1.47b 13.86±1.30b 16.25±1.55a 34.498
GPT (U/l) 14.33±0.88b 14.32±0.88b 14.32±0.78b 14.33±0.85b 14.31±0.59b 20.91±0.69a 2.496
LPO (nmol MDA/mg protein) 1.88±0.21b 1.89±0.15b 1.89±0.14b 1.91±0.13b 1.93±0.15b 4.30±0.20a 104.38
90 Days SOD (μmol/min/mg protein) 14.49±2.09b 14.60±1.43b 14.65±2.40b 14.72±2.11b 14.81±1.64b 19.86±2.19a 3.381
CAT (U/mg protein) 26.56±2.11b 26.55±2.13b 26.61±2.03b 26.72±2.41b 26.84±1.98b 35.96±2.75a 7.551
GOT (U/l) 13.86±1.92b 13.84±1.18b 13.84±1.26b 13.85±1.29b 13.86±1.24b 20.06±2.60a 35.192
GPT (U/l) 14.32±0.93b 14.33±0.90b 14.32±0.73b 14.33±0.98b 14.34±0.69b 22.61±1.51a 9.170
LPO (nmol MDA/mg protein) 1.89±0.12b 1.90±0.23b 1.90±0.19b 1.99±0.21b 2.03±0.20b 5.49±0.29a 139.26
Each value is mean ± SD of three individual observations; mean values within the same row sharing the different alphabetical superscripts are statisticallysignificant at P<0.05 (one-way ANOVA and subsequent post hoc multiple comparison with DMRT); a–b, order of performance (a > b)
ZnNPs induced growth performance in prawn
Author's personal copy

the form of University Research Fellowship to the first author. TheUniversity Grants Commission, Government of India, New Delhi, is alsogratefully acknowledged for the acquired laboratory facility by the secondauthor through aMajor Research Project operated (2009–2012) on prawnnutrition.
References
1. Salgueiro MJ, Zubillaga M, Lysionek A, Sarabia MI, Caro R, PaoliDT, Hager A, Weill R, Boccio J (2000) Zinc as an essential micro-nutrient: a review. Nutr Res 20:737–755
2. Tan B, Mai K (2001) Zinc methionine and zinc sulfate as sources ofdietary zinc for juvenile abalone, Haliotis discus hannai Ino.Aquaculture 192:67–84
3. GammanpilaM, Age AY, Bart AN (2007) Evaluation of the effects ofdietary vitamin C, E and Zinc supplementation on reproductiveperformance of Nile tilapia (Oreochromis niloticus). Sri Lanka JoAquat Sci 12:39–60
4. Herland H, Cooper M (2011) Effects of dietary mineral supplemen-tation on quality of fresh and salt-cured fillets from farmed Atlanticcod Gadus morhua. J World Aquacult Soc 42:261–267
5. Davis DA, Lawrence AL, Galtin DM III (1993) Evaluation of thedietary zinc requirement of Penaeus vannamei and effects of phyticacid on zinc and phosphorus bioavailability. J World Aquacult Soc24:40–47
6. Shiau SY, Jiang LC (2006) Dietary zinc requirements of grassshrimp, Penaeus monodon, and effects on immune responses.Aquaculture 254:476–482
7. Rani B, Hasnat A, Kohli MPS, Chandraprakash G (2012) Zincsupplementation and its effect on thermal stress resistance inCarassius auratus Fry. Isr J Aquacult Bamidgeh 64:779–786
8. Manusadzianas L, Grigutyte R, Jurkoniene S, Karitonas R,Sadauskas K, Ferard JF, Cotelle S, Foucaud L (2009) Toxicity ofzinc oxide nanoparticle suspensions to aquatic biota. 14th interna-tional Symposium on Toxicity and Assement (ISTA-14).Universityof Metz France
9. Xiong D, Fang T, Yu L, Sima X, Zhu W (2011) Effects of nano-scaleTiO2, ZnO and their bulk counterparts on zebrafish: acute toxicity,oxidative stress and oxidative damage. Sci Total Environ 409:1444–1452
10. Hao L, Chen L, Hao J, Zhong N (2013) Bioaccumulation and sub-acute toxicity of zinc oxide nanoparticles in juvenile carp (Cyprinuscarpio): a comparative study with its bulk counterparts. EcotoxicolEnviron Saf 91:52–60
11. Rather MA, Sharma R, Aklakur M, Ahmad S, Kumar N, Khan M,Ramya VL (2011) Nanotechnology, a novel tool for aquaculture andfisheries development a prospective mini-review. Fish Aquacult J 16:1–5
12. Zhou X, Wang Y, Gu Q, Li W (2009) Effects of different dietaryselenium sources (selenium nanoparticle and selenomethionine) ongrowth performance muscle composition and glutathione peroxidaseenzyme activity of crucian carp (Carassius auratus gibelio).Aquaculture 29:78–81
13. AOAC (1995) Official methods of analysis, 16th edn. Arlinton, VA,AOAC International
14. Tekinay AA, Davies SJ (2001) Dietary carbohydrate level influenc-ing feed intake, nutrient utilization and plasma glucose concentrationin the rainbow trout, Oncorhynchus mykiss. Tur J Vet Anim Sci 25:657–666
15. Furne M, Hidalgo MC, Lopez A, Garcia-Gallego M, Morales AE,Domenzain A, Domezain J, Sanz A (2005) Digestive enzyme activ-ities in Adriatic sturgeon Acipenser naccarii and rainbow trout
Oncorhynchus mykiss A comparative study. Aquaculture 250:391–398
16. Bernfeld P (1955) Amylases alpha and beta. Methods Enzymol 1:149–158
17. Lowry OH, Rosenbrough WJ, Fair AL, Randall RJ (1951) Proteinmeasurement with the folinphenol reagent. J Biol Chem 193:265–275
18. Moore S, Stein WH (1948) Photometric ninhydrin method for use inthe chromatography of amino acid. J Biol Chem 176:367–388
19. Roe JH (1955) The determination of sugar and blood and spinal fluidwith anthrone reagent. J Biol Chem 212:335–343
20. Folch J, Lees M, Bloane-Stanley GH (1957) A simple method for theisolation and purification of total lipids from animal tissues. J BiolChem 266:497–509
21. Barnes H, Black Stock J (1973) Estimation of lipids in marinean ima l s and t i s s u e s De t a i l e d i nve s t i g a t i on o f th eSulphophosphovanillin method for total lipids. J Exper Mar BiolEcol 12:103–118
22. Tsing A, Arcier JM, Brehelin M (1989) Hemocytes of penaeid andpalaemonid shrimps: morphology, cytochemistry and hemograms. JInvert Pathol 53:64–77
23. Kakkar R, Kalra J, Mantha SV, Prasad K (1995) Lipid peroxidationand activity of antioxidant enzymes in diabetic rats. Mol CellBiochem 151:113–119
24. Sinha AK (1972) Colorimetric assay of catalase. Anal Biochem 47:389–394
25. Ohkawa H, Ohishi N, Yagi K (1979) Assay for lipid peroxides inanimal tissues by thiobarbituric acid reaction. Anal Biochem 95:351–358
26. Reitman S, Frankel S (1957) A colorimetric method for the determi-nation of serum glutamic oxalacetic and glutamic pyruvic transami-nases. Am J Clin Pathol 28:56–63
27. Ho E (2004) Zinc deficiency, DNA damage and cancer risk. J NutrBiochem 15:572–578
28. Tapiero H, Tew KD (2003) Trace elements in human physiology andpathology: zinc and metallothioneins. Biomed Pharmacother 57:399–411
29. Hedemann MS, Jensen BB, Poulsen HD (2006) Influence of dietaryzinc and copper on digestive enzyme activity and intestinal morphol-ogy in weaned pigs. J Anim Sci 84:3310–3320
30. Ramseyer L, Garling D, Hill G, Link J (1999) Effect of dietary zincsupplementation and phytase pre- treatment of soybean meal or corngluten meal on growth, zinc status and zinc-related metabolism inrainbow trout,Oncorhynchus mykiss. Fish Physiol Biochem 20:251–261
31. Satoh S, Izume K, Takeuchi T, Watanabe T (1987) Availability torainbow trout of zinc contained in various types of fishmeals. NipponSuisan Gakkaishi 53(1861–1):866
32. Mohanty M, Adhikari S, Mohanty P, Sarangi N (2009) Effect ofwaterborne zinc on survival, growth, and feed intake of Indianmajor carp, Cirrhinus mrigala (Hamilton). Water Air Soil Pollut201:3–7
33. Li WW, Gong YN, Jin XK, He L, Jiang H, Ren F, Wang Q (2010)The effect of dietary zinc supplementation on the growth hepatopan-creas fatty acid composition and gene expression in the Chinesemitten crab Eriocheir sinensis (H Milne-Edwards) (Decapoda,Grapsidae). Aquacult Res 41:828–837
34. Li JS, Li JL, Wu TT (2007) The effects of copper, iron and zinc ondigestive enzymeactivity in the hybrid tilapia Oreochromis niloticus(L.) Oreochromis aureus (Steindachner). J Fish Biol 71:1788–1798
35. JingMY, Sun JY,Weng XY,Wang JF (2009) Effects of zinc levels onactivities of gastrointestinal enzymes in growing rats. J Anim PhysiolAnim Nutr 93:606–612
36. Lovett DL, Felder DL (1990) Ontogenic change in digestive enzymeactivityof larval and postlarval white shrimp Penaeus setiferus(Crustacea, Decapoda, Penaeidae). Biol Bull 178:144–159
Muralisankar et al.
Author's personal copy

37. Lall SP (2002) The minerals. In: Halver JE, Hardy RD (eds) Fishnutrition, 3rd edn. Academic Press, New York, pp 259–308
38. Gatlin DM III, Wilson RP (1984) Dietary selenium requirements offingerling channel catfish. J Nutr 114:627–633
39. Ward JD, Spears JW, Kegley EB (1993) Effect of copper level andsource (copper lysine vs copper sulfate) on copper status performance
and immune response in growing steers fed diets with or withoutsupplemental molybdenum and sulfur. J Anim Sci 71:2748–2755
40. Ates A, Daniels J, Arslan Z, Farah IO, Rivera HF (2013)Comparative evaluation of impact of Zn and ZnO nanoparticles onbrine shrimp (Artemia salina) larvae: effects of particle size andsolubility on toxicity. Environ Sci Process Impact 15:225–233
ZnNPs induced growth performance in prawn
Author's personal copy





























