Embed Size (px)
Citation preview

MASARYKOVA UNIVERZITAPŘÍRODOVĚDECKÁ FAKULTA
ÚSTAV MATEMATIKY A STATISTIKY
Bakalářská práce
BRNO 2015 ŠTĚPÁN VRTEK

MASARYKOVA UNIVERZITAPŘÍRODOVĚDECKÁ FAKULTA
ÚSTAV MATEMATIKY A STATISTIKY
Diferenční rovnice agenetikaBakalářská práce
Štěpán Vrtek
Vedoucí práce: doc. RNDr. Zdeněk Pospíšil, Dr. Brno 2015

Bibliografický záznam
Autor: Štěpán VrtekPřírodovědecká fakulta, Masarykova univerzitaÚstav matematiky a statistiky
Název práce: Diferenční rovnice a genetika
Studijní program: Matematika
Studijní obor: Finanční a pojistná matematika
Vedoucí práce: doc. RNDr. Zdeněk Pospíšil, Dr.
Akademický rok: 2014/2015
Počet stran: ix+ 36
Klíčová slova: Nelineární diferenční rovnice, genetika populací, selekce, FHWrovnice

Bibliographic Entry
Author: Štěpán VrtekFaculty of Science, Masaryk UniversityDepartment of mathematics and statistics
Title of Thesis: Difference equations and genetics
Degree Programme: Mathematics
Field of Study: Finance and insurance mathematics
Supervisor: doc. RNDr. Zdeněk Pospíšil, Dr.
Academic Year: 2014/2015
Number of Pages: ix+ 36
Keywords: Nonlinear difference equations, population genetics, selection,FHW equation

Abstrakt
Cílem této práce je studium nelineárních diferenčních rovnic a matematických modelův genetice populací s apelem na ty, jež nelineární diferenční rovnice využívají. Zejménaby práce měla řešit problém selekce a vyšetřit Fisher - Haldane - Wrightovu rovnici.
Abstract
The aim of this thesis is to study nonlinear difference equations and mathematicalmodels in population genetics, especially those using nonlinear difference equations. Inparticular, a problem of selection and behaviour of Fisher - Haldane - Wright’s equationshould be investigated.


Poděkování
Na tomto místě bych rád poděkoval doc. RNDr. Zdeňku Pospíšilovi, Dr. za vedení mépráce správným směrem, dobré rady a podnětné diskuse, prof. Ing. Josefovi Dvořákovi,CSc., dr.h.c. za konzultace v oblasti genetiky živočichů, Ing. Ireně Vrtkové za uvedení dopraxe genetiky populací a Bc. Jakubovi Fajnorovi za korekturu textu.
Prohlášení
Prohlašuji, že jsem tuto bakalářskou práci vypracoval samostatně s použitím literaturya pramenů uvedených v bibliografii.
Brno 27.5.2015 . . . . . . . . . . . . . . . . . . . . . . . . . .Štěpán Vrtek

Obsah
Úvod . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ix
Kapitola 1. Přípravné úvahy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11.1 Posloupnosti . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11.2 Operátor diference . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
Kapitola 2. Lineární diferenční rovnice . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32.1 Obecné řešení lineárních diferenčních rovnic . . . . . . . . . . . . . . . . . . . . . 32.2 Lineární homogenní rovnice s konstantními koeficienty . . . . . . . . . . . . . . 42.3 Nehomogenní lineární rovnice . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
2.3.1 Obecné řešení nehomogenních rovnic . . . . . . . . . . . . . . . . . . . . . . 62.3.2 Metoda variace konstant . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
Kapitola 3. Nelineární diferenční rovnice . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 103.1 Linearizace nelineárních rovnic . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
3.1.1 Riccatiho rovnice . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 103.1.2 Bernoulliho rovnice . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 113.1.3 Goniomterické a hyperbolické substituce . . . . . . . . . . . . . . . . . . . . 123.1.4 Homogenní rovnice . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 143.1.5 Logaritmicky lineární rovnice . . . . . . . . . . . . . . . . . . . . . . . . . . . 143.1.6 Linearizace pomocí Lieovy transformace . . . . . . . . . . . . . . . . . . . . 15
3.2 Grafické řešení . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 183.2.1 Rovnovážné body a jejich stabilita . . . . . . . . . . . . . . . . . . . . . . . . 183.2.2 Cykly . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
Kapitola 4. Populační genetika . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 224.1 Historie genetiky populací . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 224.2 Mendelistická genetika populací s nepřesahujícími
generacemi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 224.3 Selekce . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 244.4 Fisher - Haldane - Wrigthova rovnice . . . . . . . . . . . . . . . . . . . . . . . . . . 254.5 Další modely z genetiky populací . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
4.5.1 Překrývání generací . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 334.5.2 Mutace . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
– vii –

Závěr . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
Literatura . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36

Úvod
Dostává se Vám do rukou práce, jež se zabývá aplikací diferenčních rovnic v genetice. Di-ferenčními rovnicemi můžeme popisovat procesy živočišné říše, které se odehrávají v jasněvytyčených časových úsecích, které můžeme chápat jako jednotlivé kroky. Takovými rov-nicemi můžeme popisovat například populační dynamiku či genetiku populací.
A právě genetikou populací, která vyžaduje znalost nelineárních diferenčních rovnic,se budeme v této práci zabývat. Abychom dokázali explicitně řešit některé nelineárnídiferenční rovnice pomocí linearizace, uvedeme si v prních dvou kapitolách (ve kterýchbudeme čerpat zejména z [1], [2] a [3]) základní teorii k diferenčním rovnicím a metodyřešení lineárních rovnic.
Ve třetí kapitole se seznámíme s několika speciálními tvary diferenčních rovnic, kteréjsou linearizovatelné specifickými substitucemi, a uvedeme si obecnější metodu linearizacenelineární diferenční rovnice prvního řádu. Kapitola se opírá zejména o poznatky z [2] a[3].
Čtvrtá kapitola pojednává o aplikaci nelineárních diferenčních rovnic na genetiku po-pulací. Seznámíme se zejména s Fisher - Haldane - Wrightovou rovnicí, která nám umožnízjistit, jak se vyvíjí četnost zkoumaného genotypu při vlivu selekce. Kapitola čerpá z [4],[5] a [9].
V závěru se pak budeme věnovat diskusi o aplikovatelnosti vybraných modelů a jejichmodifikaci potřebné k uvedení do praxe.
– ix –

Kapitola 1
Přípravné úvahy
1.1 PosloupnostiPro celé číslo t0 ∈R označíme
Zt0 = {t0 + n : n ∈N0}= {t0, t0 + 1, t0 + 2, . . .}
množinu všech celých čísel větších či rovných číslu t0. Množinu celých čísel pak můžemeoznačit jako Z−∞.
Definice 1.1. Reálná posloupnost je zobrazení a z množiny celých čísel Z do množinycelých čísel R takové, že jeho definiční obor Doma je celá množina Z nebo některáz množin Zt0 .
Hodnotu posloupnosti a(t) budeme nazývat t-tý člen posloupnosti. Hodnotu t pakbudeme nazývat indexem posloupnosti. Pokud t0 > −∞ a Doma = Zt0 , řekneme, že t0 jepočáteční index posloupnosti.
Věta 1.2. Mějme t0 ∈Z∪{−∞} a posloupnosti a1,a2, . . . ,an takové, že Domai = Zt0 proi = 1, . . . ,n. Označme
C(t) =C(t;a1, . . . ,an) =
∣∣∣∣∣∣∣∣∣a1(t) a2(t) · · · an(t)
a1(t + 1) a2(t + 1) · · · an(t + 1)...
... . . . ...a1(t + n−1) a2(t + n−1) · · · an(t + n−1)
∣∣∣∣∣∣∣∣∣ . (1.1)
Pokud existuje t ∈ Zt0 takový index, že C(t) 6= 0, pak jsou posloupnosti a1,a2, . . . ,anlineárně nezávislé. Jsou-li a1,a2, . . . ,an lineárně závislé, pak C(t) = 0 pro všechny indexyt ∈Zt0 .
Důkaz. Nechť pro konstanty α1,α2, . . . ,αn platí
α1a1 +α2a2 + . . .+αnan = 0
a nechť t ∈Zt0 je takový index, že C(t) 6= 0. Z předchozí rovnosti plyne, že
α1a1(t)+α2a2(t)+ . . .+αnan(t) = 0
– 1 –

Kapitola 1. Přípravné úvahy 2
α1a1(t + 1)+α2a2(t + 1)+ . . .+αnan(t + 1) = 0...
α1a1(t + n−1)+α2a2(t + n−1)+ . . .+αnan(t + n−1) = 0,
což je homogenní soustava n lineárních rovnic o n neznámých α1,α2, . . . ,αn a C(t) je jejídeterminant. Odtud plyne, že tato soustava má jen triviální řešení, tj.
α1 = α2 = · · ·= αn = 0.
To znamená, že posloupnosti a1,a2, . . . ,an jsou lineárně nezávislé a první tvrzení je doká-záno.
Druhé tvrzení je bezprostředním důsledkem prvního.
Poznámka 1.3. Determinant C(t;a1,a2, . . . ,an) zavedený v předchozí větě se nazývá Ca-soratián posloupností a1,a2, . . . ,an v indexu t.
1.2 Operátor diferenceUveďme operátor diference a jeho základní vlastnosti.
4 x(t) = x(t + 1)− x(t), (1.2)
Lemma 1.4. Pro operátor diference platí
i.t−1
∑k=t0
4x(k) = x(t)− x(t0) (1.3)
ii.4 (t−1
∑k=t0
x(k)) = x(t) (1.4)
Důkaz.
i.t−1
∑k=t0
4x(k) =4x(t0)+4x(t0 + 1)+ . . .+4x(t−1) =
= x(t0 + 1)− x(t0)+ . . .+ x(t)− x(t−1) = x(t)− x(t0).
ii.4 (t−1
∑k=t0
x(k)) =t
∑k=t0
x(k)−t−1
∑k=t0
x(k) = x(t0)+ . . .+ x(t)− x(t0)− . . .− x(t−1) =
= x(t).

Kapitola 2
Lineární diferenční rovnice
2.1 Obecné řešení lineárních diferenčních rovnicObecný tvar lineární diferenční rovnice k-tého řádu je
x(t + k)+ p1(t)x(t + k−1+ . . .+ pk(t)x(t) = g(t), (2.1)
kde pk(t) 6= 0 pro všechna t. Přidruženými počátečními podmínkami pro diferenční rovnici(2.1) nazýváme rovnosti
x(t0) = ξ0, x(t0 + 1) = ξ1, x(t0 + 2) = ξ2, . . . , x(t0 + k−1) = ξk−1, (2.2)
kde t0 ∈Z a ξ1,ξ2, . . . ,ξk−1 ∈R. Rovnice (2.1) s podmínkami (2.2) nazýváme počátečníúlohou.
Věta 2.1. Počáteční úloha (2.1), (2.2) má jednoznačné řešení x(t).
Důkaz. Plyne z postupného vypočítávání (2.1) pro t = t0, t0 + 1, . . .. Jelikož jsou hodnotyx(t0),x(t0 + 1), . . . ,x(t0 + k− 1) dané, existuje pouze jedna možná hodnota x(t0 + k).Obdobně pro x(t0 + k+ 1) jsou známy hodnoty x(t0 + 1),x(t0 + 2), . . . ,x(t0 + k) a x(t0 +k + 1) je jednoznačně určeno a tedy rekurentně x(t) je jednoznačně určeno hodnotamix(t− k),x(t− k−1), . . . ,x(t−1).
Zabývejme se nyní homogenními diferenčními rovnicemi k-tého řádu. Obecný tvartakovéto rovnice je
x(t + k)+ p1(t)x(t + k−1)+ p2(t)x(t + k−2)+ . . .+ pk(t)x(t) = 0. (2.3)
Řešením této rovnice bude posloupnost x(t) taková, která vyhovuje rovnici (2.3).
Lemma 2.2. Nechť x1(t) a x2(t) jsou řešení (2.3). Pak platí, že
x(t) = x1(t)+ x2(t)
je řešení rovnice (2.3) ax(t) = ax1(t)
je řešení (2.3) pro libovolnou konstantu a ∈R.
– 3 –

Kapitola 2. Lineární diferenční rovnice 4
Věta 2.3. (Princip superpozice) Pokud x1(t),x2(t), . . . ,xk(t) jsou řešení (2.3), pak x(t) =a1x1(t)+ a2x2(t)+ . . .+ akxk(t) je řešení (2.3) pro libovolné a1,a2, . . . ,ak.
Důkaz. Je přímým důsledkem 2.2
Věta 2.4. Množina S všech řešení rovnice (2.3) spolu s operacemi +, ·, teré splňují
(x+ y)(t) = x(t)+ y(t) pro x, y ∈ S, t ∈Z+,(a · x)(t) = a · x(t) pro x ∈ S, a ∈R,
tvoří vektorový prostor.
Důkaz. Důkaz se provede ověřením axiomů vektorového prostoru.
Poznámka 2.5. Množina k lineárně nezávislých řešení rovnice (2.3) se nazývá fundamen-tální řešení rovnice (2.3).
Poznámka 2.6. Nechť {x1(t),x2(t), . . . ,xk(t)} je fundamentální řešení (2.3). Pak obecnéřešení rovnice (2.3) je dáno jako
x(t) =k
∑i=1
axi(t), (2.4)
kde a ∈R.
Poznámka 2.7. Množina řešení x1(t),x2(t), . . . ,xk(t) rovnice (2.3) je fundamentálnímřešením právě tehdy, když pro nějaké t0 ∈Z+ je Casoratián C(t0) 6= 0.
Ve zbytku kapitoly se zaměříme na metody řešení homogenních a nehomogenníchlineárních diferenčních rovnic, které jsou potřebné při kompletním řešení nelineárníchrovnic.
2.2 Lineární homogenní rovnice s konstantními koefici-enty
Budeme se zabývat pouze diferenčními rovnicemi tvaru
x(t + k)+ p1x(t + k−1)+ . . .+ pkx(t) = 0, (2.5)
které mají konstantní koeficienty p1, p2, . . . , pk ∈R a kde pk 6= 0, jelikož lineárními dife-renčními rovnicemi s proměnnými koeficienty se v praktické části nebudeme zabývat.
Našim úkolem je najít fundamentální řešení této rovnice a následně najít obecné řešenírovnice (2.5). Uvažujme nejdříve lineární homogenní rovnici prvního řádu
x(t + 1)+ cx(t) = 0.

Kapitola 2. Lineární diferenční rovnice 5
Její řešení dostaneme z rekurentně zadané posloupnosti x(t + 1) = (−c)x(t) s počátečníhodnotou x(0) = x0 následovně
x(1) = (−c)x0,
x(2) = (−c)x(1) = (−c)2x0,...
x(t) = (−c)x(t−1) = . . .= (−c)tx0.
Analogicky budeme hledat řešení rovnice (2.5) ve tvaru x(t) = λ t , kde λ ∈C. Substitucíočekávaného tvaru řešení do rovnice získáváme
λk + p1λ
k−1 + . . .+ pk = 0. (2.6)
Tato rovnice se nazývá charakteristickou rovnicí rovnice (2.5) a její kořeny λi jsoucharakteristickými kořeny. Všimněme si, že jelikož pk 6= 0, pak žádný charakteristickýkořen není roven nule.
Mohou nastat dva případy.
i. Charakteristické kořeny λ1,λ2, . . . ,λk jsou různé. Ukážeme, že množina řešení{λ t
1,λ t2, . . . ,λ t
k} je fundamentálním řešením (2.6). Podle poznámky 2.7 stačí ukázat,že C(0) 6= 0, kde C(t) je Casoratián řešení v indexu t. Tedy
C(0) =
∣∣∣∣∣∣∣∣∣∣∣
1 1 · · · 1λ1 λ2 · · · λkλ 2
1 λ 22 · · · λ 2
k...
... . . . ...λ
k−11 λ
k−12 · · · λ
k−1k
∣∣∣∣∣∣∣∣∣∣∣. (2.7)
Tento determinant se nazývá Vandermondův determinant. Pro výpočet determinantuse použije vzorec
an−bn = (a−b)(an−1 + an−2b+ an−3b2 . . .+ abn−2 + bn−1.
Po vypočtení dostáváme
C(0) =k
∏i, j=0, i> j
(λi−λ j).
Jelikož jsou všechny charakteristické kořeny různé, pak C(0) 6= 0, což znamená, žemnožina {λ t
1,λ t2, . . . ,λkt} je fundamentální systém řešení (2.5). Obecným řešením
(2.5) je pak
x(t) =k
∑i=1
aiλti , kde ai ∈R.

Kapitola 2. Lineární diferenční rovnice 6
ii. Charakteristické kořeny λ1,λ2, . . . ,λs mají násobnosti m1,m2, . . . ,ms takové,
žes∑
i=1mi = k. Rovnice (2.5) se v tomto případě může psát ve tvaru
(E−λ1)m1 · (E−λ2)
m2 · . . . · (E−λs)msx(t) = 0, (2.8)
kde E je operátor posunu (tzv. shift operátor), který je definovaný vztahem
Ex(t) = x(t + 1),
a platí vztah (viz. [3] strana 55)
(E−λ )mx(t) =m
∑k=0
(mi
)(−λ )iEm−ix(t).
Pak pro jednotlivé kořeny platí
(E−λi)mix(t) = 0. (2.9)
Lemma 2.8. Množina Gi = {λ ti , (t
1)λt−1i , . . . , ( t
mi−1)λt−mi+1i } je fundamentálním řešením
(2.9).
Důkaz. Viz. [1].
Věta 2.9. Množina G =s⋃
i=1Gi je fundamentálním řešením (2.8).
Důkaz. Viz. [1]
Věta 2.10. Obecné řešení (2.8) je dáno vztahem
x(t) =s
∑i=1
λti (ai, 0 + ai, 1t + . . .+ ai, mi−1tmi−1).
Důkaz. Provedeme použitím 2.8 a 2.9. Viz [1].
2.3 Nehomogenní lineární rovnice
2.3.1 Obecné řešení nehomogenních rovnicUvažujme nehomogenní diferenční rovnici k-tého řádu
x(t + k)+ p1(t)x(t + k−1)+ . . .+ pk(t)x(t) = g(t), (2.10)
kde ∀t ≥ t0 platí, že pk(t) 6= 0.Uveďme nejdříve na příkladu, že obecné řešení nehomogenní rovnice netvoří vektorový
prostor.

Kapitola 2. Lineární diferenční rovnice 7
Příklad 2.11. Mějme diferenční rovnici
x(t + 2)−7x(t + 1)+ 6x(t) = t. (2.11)
Dosazením do rovnice (2.11) můžeme ukázat, že řešením jsou x1(t) = 350t− 1
10t2 + 3 ax2(t) = 6t + 3
50t− 110t2. Přitom x1(t)−x2(t) =−6t +3 není řešením rovnice (2.11), stejně
jako 5x1(t) = 310t− 1
2t2 + 15 není řešením (2.11).
Kromě zjištění, že obecné řešení (2.10) netvoří vektorový prostor, můžeme uvést ještějedno tvrzení.
Věta 2.12. Pokud x1(t) a x2(t) jsou řešením (2.10), pak x(t) = x1(t)− x2(t) je řešenímpřidružené homogenní rovnice
x(t + k)+ p1(t)x(t + k−1)+ . . .+ pkx(t) = 0. (2.12)
Důkaz. Platí, že
x1(t + k)+ p1(t)x1(t + k−1)+ . . .+ pk(t)x1(t) = g(t)
ax2(t + k)+ p1(t)x2(t + k−1) . . .+ pk(t)x2(t) = g(t).
Pokud odečteme druhou rovnici od první, dostáváme
x1(t + k)− x2(t + k)+ . . .+ pk(t)(x1(t)− x2(t)) = 0(x1− x2)(t + k)+ . . .+ pk(t)[(x1− x2)(t)] = 0,
tedy původní tvrzení je dokázáno.
Obecné řešení rovnice (2.12) budeme značit jako xc(t) a nazývat doplňkovým řešením.Řešení rovnice (2.10) budeme nazývat partikulárním řešením a značit xp(t).
Věta 2.13. Pro jakékoliv řešení x(t) rovnice (2.10) platí
x(t) = xp(t)+k
∑i=1
aixi(t),
kde {x1(t),x2(t), . . . ,xk(t)} je fundamentální systém řešení homogenní rovnice (2.12).
Důkaz. Viz. [1]
Uveďme si nyní metodu řešení nehomogenních rovnic. Nejdříve pouze zmiňme metoduneurčitých koeficientů, která je založena na šikovném odhadnutí partikulárního řešení nazákladě tvaru funkce g(t). Celá metoda je velice jasně a přehledně vysvětlena zde [1].

Kapitola 2. Lineární diferenční rovnice 8
2.3.2 Metoda variace konstantTato metoda je narozdíl od metody neurčitých koeficientů obecně použitelná a je obdobouvariace konstant k řešení diferenciálních rovnic. V diskrétním případě je ovšem operátorderivace nahrazen operátorem diference.
Předpokládejme nejdříve řešení rovnice (2.10) ve tvaru
x(t) =k
∑i=1
ai(t)xi(t), (2.13)
kde {x1(t),x2(t), . . . ,xk(t)} je fundamentální systém řešení (2.12) a a1(t),a2(t), . . . ,ak(t)jsou zatím neurčené posloupnosti. Vyjádříme dále
x(t + 1) =k
∑i=1
ai(t + 1)xi(t + 1) =k
∑i=1
[(4ai(t))xi(t + 1)+ ai(t)xi(t + 1)]
a požadujeme, aby a1(t),a2(t), . . . ,ak(t) splňovaly
k
∑i=1
[(4ai(t))xi(t + 1)] = 0.
Analogickou rovnost budeme požadovat až do kroku (k−1), celkově tedy požadujemesystém
k
∑i=1
[(4ai(t))xi(t + j)] = 0 pro j = 1,2, . . . ,k−1 (2.14)
a dostáváme
x(t + j) =k
∑i=1
ai(t)xi(t + j) pro j = 1,2, . . . ,k−1. (2.15)
Můžeme psát, že
x(t + k) =k
∑i=1
ai(t + 1)xi(t + k) =k
∑i=1
(4ai(t))xi(t + k)+k
∑i=1
ai(t)xi(t + k) =
=k
∑i=1
(4ai(t))xi(t + k)−k
∑i=1
ai(t) ·k−1
∑j=0
c j(t)xi(t + j). (2.16)
Současně platí, že x(t) má být řešením (2.10), tedy dostáváme
x(t + k) = b(t)−k−1
∑j=0
c j(t)x(t + j) = b(t)−k−1
∑j=0
c j(t) ·k
∑i=1
ai(t)xi(t + j) =
= b(t)−k
∑i=1
ai(t) ·k−1
∑j=0
c j(t)xi(t + j) (2.17)
a porovnáním (2.16) a (2.17) dostaneme
k
∑i=1
(4ai(t))xi(t + k) = g(t). (2.18)

Kapitola 2. Lineární diferenční rovnice 9
Spojením rovnice (2.18) se soustavou (2.14) dostáváme systém, díky kterému spočtemeposloupnosti 4a1(t),4a2(t), . . . ,4ak(t). Determinant tohoto systému je Casoratiánemfundamentálního systému řešení rovnice (2.12) v indexu t +1. Je tedy nenulový a soustavaje jednoznačně řešitelná.
Označme di(t) jako
di(t) =
∣∣∣∣∣∣∣∣∣x1(t) · · · xi−1(t) xi+1(t) · · · xk(t)
x1(t + 1) · · · xi−1(t + 1) xi+1(t + 1) · · · xk(t + 1)... . . . ...
... . . . ...x1(t + k−2) · · · xi−1(t + k−2) xi+1(t + k−2) · · · xk(t + k−2)
∣∣∣∣∣∣∣∣∣ .Diference ai(t) pak můžeme pomocí Casoratiánu fundamentálního systému řešení rovnice(2.12) zapsat jako
4ai(t) =(−1)k+ig(t)di(t + 1)
C(t + 1)pro i = 1,2, . . . ,k.
Odtud dostáváme
ai(t) = ai(t0)+ (−1)k+it−1
∑j=t0
g( j)di( j+ 1)C( j+ 1)
pro i = 1,2, . . . ,k.
Při označení Ci = ai(t0) můžeme psát obecné řešení rovnice (2.10) ve tvaru
x(t) =k
∑i=1
Cixi(t)+ (−1)kt−1
∑j=t0
g( j)C( j+ 1)
k
∑i=1
(−1)idi( j+ 1)xi(t). (2.19)

Kapitola 3
Nelineární diferenční rovnice
V celé kapitole se budeme zabývat diferenčními rovnicemi k-tého řádu
x(t + k) = f (t,x(t), . . . ,x(t + k−1)), (3.1)
které jsou nelineární.Většina nelineárních diferenčních rovnic není explicitně řešitelná, a proto se přistupuje
k rekurzivnímu výpočtu ze známých hodnot v předešlém kroce. Některé nelineární rovniceje ovšem možné linearizovat.
3.1 Linearizace nelineárních rovnicZ diferenční rovnice (3.1) uhodneme tvar funkce f tak, aby substituce y(t) = f (x(t))převedla rovnici (3.1) na lineární rovnici. Nicméně je většinou velmi komplikované funkci fodhadnout. Můžeme tedy postupovat opačně a k lineární diferenční rovnici zvolit nelineárníprostou funkci g a substituovat g(x(t)) za x(t). Při označení z(t) = g(x(t)) dostanemenelineární diferenční rovnici, která je řešitelná substitucí x(t) = g−1(z(t)).
Uveďme nyní několik známých substitucí, které dokáží převést nelineární diferenčnírovnici na rovnici lineární.
3.1.1 Riccatiho rovniceRiccatiho rovnice je tvaru
p(t)x(t + 1)x(t)+ x(t + 1)+ q(t)x(t)+ r(t) = 0. (3.2)
Tuto rovnici můžeme vyřešit pomocí substituce
x(t) =(
y(t + 1)y(t)
−1)
1p(t)
. (3.3)
Dostáváme tak
y(t + 2)−(
1+p(t + 1)
p(t)(1+ q(t))
)y(t + 1)+
+
(p(t + 1)
p(t)(1+ q(t))− r(t)p(t + 1)
)y(t) = 0.
– 10 –

Kapitola 3. Nelineární diferenční rovnice 11
Pokud je r(t) ≡ 0, pak můžeme na místo (3.3) použít substituci
x(t) =1
y(t).
Příklad 3.1. Mějme rovnici Riccatiho typu
x(t + 1)x(t)+ x(t + 1)−3x(t)+ 1 = 0.
Zavedeme substituci (3.3) a dosadíme do původní rovnice.(y(t + 2)y(t + 1)
−1)(
y(t + 1)y(t)
−1)+
(y(t + 2)y(t + 1)
−1)−3(
y(t + 1)y(t)
−1)+ 1 = 0.
Po zjednodušení rovnice dostáváme
y(t + 2)−4y(t + 1)+ 4y(t) = 0,
což je homogenní lineární rovnice, kterou dále řešíme.
λ2−4λ + 4 = 0 ⇒ λ1, 2 = 2
y(t) = c12t + c2t2t .
Výrazem y(t) vyjádříme x(t) jako
x(t) =c12t+1 + c2(t + 1)2t+1
c1 + c2t2t −1
=c12+ c2(t + 1)2
c1 + c2t−1
=c1 + c2(t + 2)
c1 + c2t.
Při označení C = c1c2
můžeme psát řešení ve tvaru
x(t) =C+(t + 2)
C+ t.
3.1.2 Bernoulliho rovniceBernoulliho rovnici
x(t + 1) = [p(t)x(t)1−α + q(t)]1
1−α ,
řesíme substitucíx(t) = y(t)
11−α . (3.4)
Příklad 3.2. Mějme rovnici
x(t + 1) = (4x(t)−2 + t)−12 .

Kapitola 3. Nelineární diferenční rovnice 12
Zavedeme substituci (3.4) a dosadíme
y(t + 1)−12 = (4y(t)+ t)−
12
y(t + 1) = 4y(t)+ t ⇒ yc(t) = c14t ,
a metodou neurčitých koeficientů vypočteme yp(t)
yp(t) = a0 + a1t ⇒ a0 = −19
, a1 = −13
y(t) = c14t− 13
t− 19
,
tedy
x(t) =1√
c14t− 13t− 1
9
.
3.1.3 Goniomterické a hyperbolické substituceZabývejme se nyní substitucemi goniometrických a hyperbolických funkcí. U všech rovnicbudeme uvažovat počáteční podmínku
x(t0) = x0. (3.5)
Příklad 3.3. Mějme rovnici s počáteční podmínkou
x(t + 1) = 2x(t)2−1, x(t0) = x0.
Řešení této rovnice je pro libovolné t ≥ t0 určeno jednoznačně. Avšak hodnota x(t0− 1)musí splňovat x0 = 2[x(t0−1)]2−1. Řešením této podmínky buď není reálné číslo (pro x0 <−1), nebo jsou dvě různá reálná čísla (pro x0 >−1). Obecně je tedy úloha nejednoznačněřešitelná pro t < t0. Budeme tedy uvažovat pouze t ≥ t0.
Pokud ještě počáteční hodnota splňuje nerovnost |x0| ≤ 1, můžeme položit x(t) =cosy(t). Po dosazení dostáváme
cosy(t + 1) = 2cos2 y(t)−1= cos2y(t),
tedyy(t + 1) = ±2y(t)+ 2kπ , k ∈Z.
Každou rovnici tohoto spočetného systému lze řešit jako lineární nehomogenní rovniciprvního řádu. Tedy
y(t) = y0(±2)t−t0 + 2kπ(±2)t−t0−1±2−1
, k ∈Z,
kde y0 = y(t0), tedy cosy0 = x0, y0 = arccosx0.

Kapitola 3. Nelineární diferenční rovnice 13
Pokud je |x0| > 1, zvolíme substituci x(t) = coshy(t). S využitím vztahu cosh2α =2(coshα)2−1 dostáváme
coshy(t + 1) = 2(coshy(t))2−1= cosh2y(t).
Vzhledem k tomu, že je cosh sudá funkce na R a je ryze monotónní na intervalech[−∞,0] a [0,∞], platí
y(t + 1) = ±2y(t).
Řešením je tedyy(t) = y0(±2)t−t0 ,
kde y0 = y(t0), tedy coshy0 = x0 a
x0 = argcoshx0 = ln(|x0|+
√x2
0−1)
.
Řešení x(t) můžeme psát ve tvaru
x(t) =
x0, t = t0
cosh(
2t−t0 ln(|x0|+√
x20 + 1)
), t > t0.
Uveďme si ještě jeden příklad goniometrické substituce.
Příklad 3.4. Mějme rovnici s počáteční podmínkou
x(t + 1) =x(t)2−1
2x(t), x(t0) = x0.
Řešení, tak, aby bylo jednoznačné, má smysl pouze pro t ≥ t0. Substituci zvolíme jako
x(t) = cotgy(t)
cotgy(t + 1) =cotg2 y(t)−1
2cotgy(t)= cotg2y(t).
Řešením této rovnice je
y(t + 1) = 2y(t)+ kπ , k ∈Z
a řešením této diferenční rovnice je
y(t) = y02t−t0 + kπ(2t−t0−1),
kde y0 = arccotgx0. Dosazením do x(t) = cotgy(t) dostáváme řešení
x(t) = cotg[y02t−t0 + kπ(2t−t0−1)]
=cotg[arccotg(x02t−t0)]cotg[kπ(2t−t0−1)]−1cotg[arccotg(x02t−t0)]+ cotg[kπ(2t−t0−1)]
.

Kapitola 3. Nelineární diferenční rovnice 14
3.1.4 Homogenní rovniceHomogenní rovnicí k-tého řádu nazveme rovnici tvaru
F(
t,x(t + k)
x(t + k−1), . . . ,
x(t + 1)x(t)
)= 0,
kde F je funkce, která není konstantní ve své druhé a poslední proměnné. Rovnici převedemena diferenční rovnici (k−1)ního řádu pomocí substituce
y(t) =x(t + 1)
x(t). (3.6)
Příklad 3.5. Mějme diferenční rovnici s počáteční podmínkou
x(t + 2) =x3(t + 1)
x2(t), x(0) = x0, x(t0 + 1) = x1,
kterou upravíme na tvarx(t + 2)x(t + 1)
−[
x(t + 1)x(t)
]2
= 0
a substitucí (3.6) převedeme na rovnici
y(t + 1)− y2(t) = 0.
Vzhledem k povaze této diferenční rovnice se omezíme pouze na řešení v indexech t ≥ t0.Řešením je
y(t) = y2t−t00 ,
kde y0 =x1x0
. Zpětným dosazením do substituce dostáváme
x(t + 1) = y2t−t00 x(t),
tedy řešením je
x(t) =
[t−1
∏i=t0
(x1
x0
)2i−t0
x0
]
=
(x1
x0
)2t−t0−1
x0.
3.1.5 Logaritmicky lineární rovniceRovnice tvaru
x(t + k)rk(t) · x(t + k−1)rk−1(t) · . . . · x(t + 1)r1(t) · x(t)r0(t) = b(t),
jsou řešitelné logaritmizací. Tou dostaneme rovnici
k
∑i=0
ln[x(t + 1)ri(t)] = lnb(t)

Kapitola 3. Nelineární diferenční rovnice 15
a substitucíx(t) = ey(t) (3.7)
dostávámek
∑i=0
ri(t)y(t + 1) = lnb(t).
Vzhledem k použití logaritmizace ovšem musí tato metoda splňovat podmínky pro svépočáteční hodnoty x0,x1, . . . ,xk−1
x(t0) = x0 > 0, x(t0 + 1) = x1 > 0, . . . , x(t0 + k−1) = xk−1 > 0.
Příklad 3.6. Mějme diferenční rovnici
x(t + 2) = x(t + 1)x2(t),
kterou zlogaritmujeme
lnx(t + 2) = lnx(t + 1)+ 2lnx(t)
a zavedeme substituci (3.7)
y(t + 2)− y(t + 1)−2y(t) = 0 ⇒ λ1 = 2, λ2 = −1y(t) = c12t + c2(−1)t ,
dosadíme y(t) do substitučního vztahu (3.7)
x(t) = ec12t+c2(−1)t
= A2t·B(−1)t
.
3.1.6 Linearizace pomocí Lieovy transformacePodívejme se nyní na obecnější techniku linearizace diferenčních rovnic. Tato technikaje založena na LTGM (Lie’s transformation group method). Pro jednoduchost omezímeúvahy na rovnice tvaru
x(t + 1) = f (x(t)).
Řekněme, že f (x) je diferencovatelná pro všechna x ∈ I. Předpokládejme, že řešení ξ (x)funkcionální rovnice
Dξ ( f (x)) = ξ (x)d fdx
(x) (3.8)
je pro nějaké D známé. Pak zavedeme novou závislou proměnnou y jako
dydx
(x) =1
ξ (x)(3.9)

Kapitola 3. Nelineární diferenční rovnice 16
pro x ∈ I tak, že ξ 6= 0. Použitím řetězového pravidla dostáváme
ddx
y( f (x)) =dydx
( f (x))d fdx
(x)
=1
ξ ( f (x))d fdx
(x) (z (3.9))
=D
ξ (x)(z (3.8))
= Ddydx
(x).
Nyní integrací dostávámey( f (x)) = Dy(x)+ c,
tedyy(x(t + 1)) = Dy(x(t))+ c,
což je lineární diferenční rovnice s konstantními koeficienty.
Příklad 3.7. Uvažujme logistickou rovnici
x(t + 1) = ax(t)(1− x(t)), kde a ∈R.
Funkce f (x) je tedy f (x) = ax(1− x) a rovnice (3.8) má podobu
Dξ (ax(1− x)) = ξ (x)a(1−2x)= aξ (x)−2axξ (x).
Z tvaru pravé strany rovnice můžeme usuzovat, že by hledaná funkce mohla být tvaruξ (x) = cx+ d. Dostáváme tedy
−Dacx2 +Dcax+Dd = −2acx2 +(ac−2ad)x+ ad.
Dostáváme soustavu rovnic
−Dca = −2ac, Dca = ac−2ad, Dd = ad,
z které vypočítáme, žeD = a = 2 a c = −2d.
Tedyξ (x) = −2dx+ d = d(1−2x).
Pak substituci zavedeme jako
dydx
=1
d(1−2x)∫dy =
∫ 1d(1−2x)
dx.

Kapitola 3. Nelineární diferenční rovnice 17
Jelikož je neurčitý integrál určený až na libovolnou konstantu, zvolme například
y = − 12d
ln[d(1−2x)],
z čehož si vyjádříme x jako
x =1− 1
d · e−2dy
2a daný vztah dosadíme do rovnice x(t + 1) = 2x(t)(1− x(t)).
1− 1d · e
−2dy(t+1)
2= 2
1− 1d · e
−2dy(t)
2·
(1−
1− 1d e−2dy(t)
2
)1− 1
d· e−2dy(t+1) = 1− 1
d2 · e−4dy(t)
−2dy(t + 1) = −4dy(t)− lnd
y(t + 1) = 2y(t)+lnd2d
.
Vyřešením této rovnice dostáváme řešení
y(t) = c02t +(2t−1)lnd2d
= c12t− lnd2d
,
které dosadíme do substitučního vztahu
x(t) =1− 1
d · e−2d(c12t− lnd
2d )
2
=1− 1
d · elnd · e−2dc12t
2
=12− (e−2dc1)2t
2
=1−A2t
2.
Všímněme si, že volbou ξ (x) = cx+ d jsme řešili rovnici x(t + 1) = ax(t)(1− x(t))pouze pro a = 2. Při výpočtu můžeme postupovat také obráceně, jak bylo naznačeno nazačátku této kapitoly, a vyjádřit nejdříve funkci ξ (x), k níž výpočtem nalezneme diferenčnírovnice, jež jsou pomocí této funkce řešitelné.
Příklad 3.8. Zvolme si za funkci ξ (x) funkci, kterou jsme intuitivně vybrali v předchozímpříkladě a zjistěme celou rodinu diferenčních rovnic, které jsou řešitelné pomocí substituce(3.9).
ξ (x) = cx+ d,
pak podle (3.8) platí
D(c f (x)+ d) = (cx+ d)d fdx
(x),

Kapitola 3. Nelineární diferenční rovnice 18
odtud
D∫ dx
cx+ d=∫ d f (x)
c f (x)+ d
D1c
ln(cx+ d)+ c∗ =1c
ln(c f (x)+ d)
eD ln(cx+d)+C = c f (x)+ d
(cx+ d)D · eC−dc
= f (x),
kde C = c∗ · c a C,D,c,d ∈R. Pokud zvolíme parametry jako
C = 0, D = 2, c = −2, d = 1,
pak získáme logistickou rovnici z předchozího příkladu.
3.2 Grafické řešeníUvažujeme-li nelineární diferenční rovnici prvního řádu
x(t + 1) = f (x(t) (3.10)
s počáteční podmínkou x(t0) = x0, můžeme danou rovnici pochopit jako zobrazení, kteréreálné hodnotě x(t) přiřadí hodnotu x(t+1). Dané zobrazení můžeme znázornit v souřadnérovině (na vodorovnou osu x naneseme hodnoty x(t), na svislou osu y hodnoty x(t + 1)).Nakreslíme tedy graf funkce f a najdeme na něm pro hodnotu x0 hodnotu x(1). Stejnýmzpůsobem nalezneme hodnotu x(2) – nejdříve přeneseme hodnotu x(1) na vodorovnouosu, a pak na grafu funkce f nalezneme příslušnou hodnotu x(2).
Tímto postupem můžeme odhadnout do jakých bodů posloupnost daná funkcí fsměřuje.
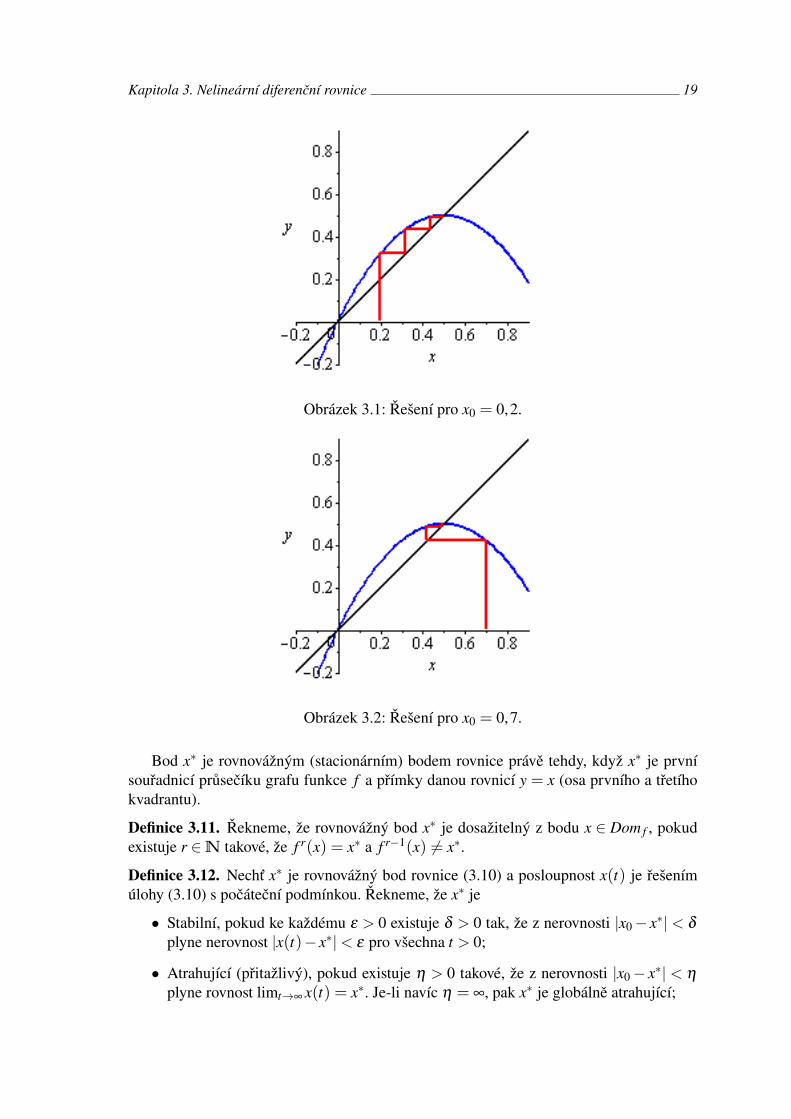
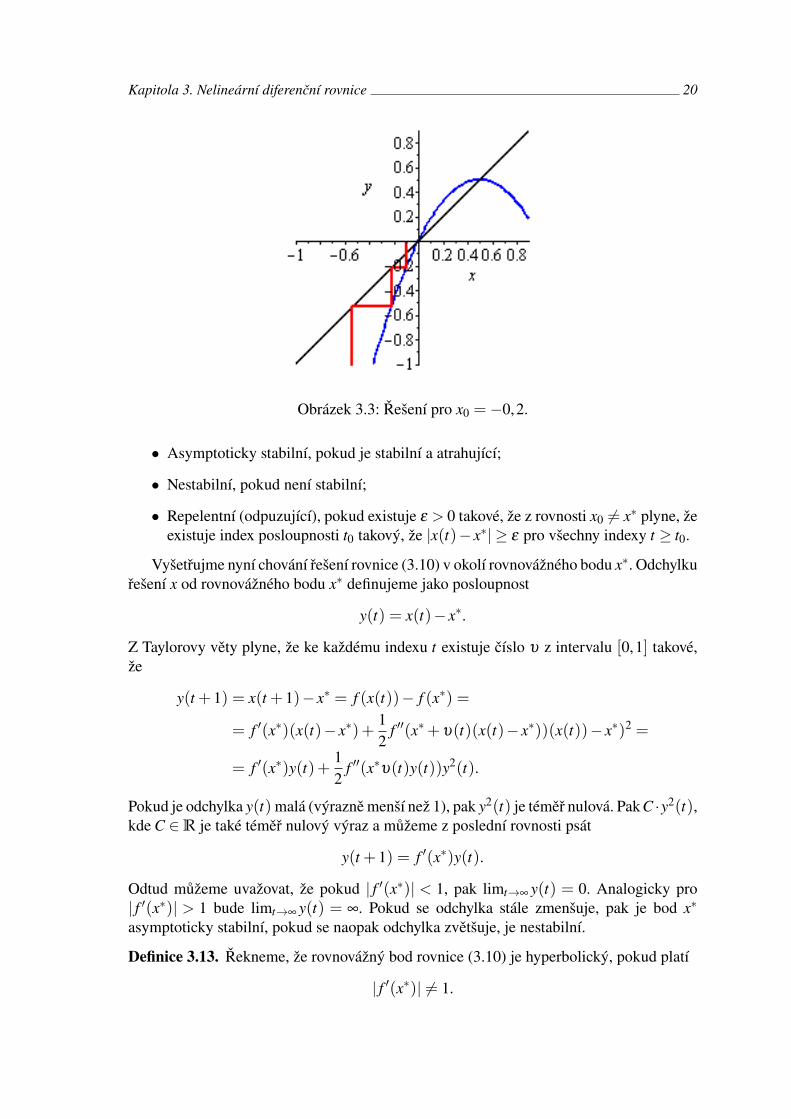
Příklad 3.9. Zabývejme se logistickou rovnicí s počáteční podmínkou
x(t + 1) = 2x(t)(1− x(t)), x(t0) = x0.
Řešení uveďme na obrázcích 3.1, 3.2 a 3.3 pro různá x0.
Touto metodou vyšetřujeme existenci, lokaci a stabilitu rovnovážných bodů. Uveďmeproto, co to rovnovážný bod je a jak určit jeho stabilitu.
3.2.1 Rovnovážné body a jejich stabilitaDefinice 3.10. Řekneme, že bod x∗ ∈Dom f je rovnovážný (stacionární) bod rovnice (3.10),pokud je pevným bodem funkce f , tedy pokud platí
f (x∗) = x∗.
Kapitola 3. Nelineární diferenční rovnice 19
Obrázek 3.1: Řešení pro x0 = 0,2.
Obrázek 3.2: Řešení pro x0 = 0,7.
Bod x∗ je rovnovážným (stacionárním) bodem rovnice právě tehdy, když x∗ je prvnísouřadnicí průsečíku grafu funkce f a přímky danou rovnicí y = x (osa prvního a třetíhokvadrantu).
Definice 3.11. Řekneme, že rovnovážný bod x∗ je dosažitelný z bodu x ∈ Dom f , pokudexistuje r ∈N takové, že f r(x) = x∗ a f r−1(x) 6= x∗.
Definice 3.12. Nechť x∗ je rovnovážný bod rovnice (3.10) a posloupnost x(t) je řešenímúlohy (3.10) s počáteční podmínkou. Řekneme, že x∗ je
• Stabilní, pokud ke každému ε > 0 existuje δ > 0 tak, že z nerovnosti |x0− x∗| < δ
plyne nerovnost |x(t)− x∗|< ε pro všechna t > 0;
• Atrahující (přitažlivý), pokud existuje η > 0 takové, že z nerovnosti |x0− x∗| < η
plyne rovnost limt→∞ x(t) = x∗. Je-li navíc η = ∞, pak x∗ je globálně atrahující;
Kapitola 3. Nelineární diferenční rovnice 20
Obrázek 3.3: Řešení pro x0 = −0,2.
• Asymptoticky stabilní, pokud je stabilní a atrahující;
• Nestabilní, pokud není stabilní;
• Repelentní (odpuzující), pokud existuje ε > 0 takové, že z rovnosti x0 6= x∗ plyne, žeexistuje index posloupnosti t0 takový, že |x(t)− x∗| ≥ ε pro všechny indexy t ≥ t0.
Vyšetřujme nyní chování řešení rovnice (3.10) v okolí rovnovážného bodu x∗. Odchylkuřešení x od rovnovážného bodu x∗ definujeme jako posloupnost
y(t) = x(t)− x∗.
Z Taylorovy věty plyne, že ke každému indexu t existuje číslo υ z intervalu [0,1] takové,že
y(t + 1) = x(t + 1)− x∗ = f (x(t))− f (x∗) =
= f ′(x∗)(x(t)− x∗)+12
f ′′(x∗+υ(t)(x(t)− x∗))(x(t))− x∗)2 =
= f ′(x∗)y(t)+12
f ′′(x∗υ(t)y(t))y2(t).
Pokud je odchylka y(t) malá (výrazně menší než 1), pak y2(t) je téměř nulová. Pak C ·y2(t),kde C ∈R je také téměř nulový výraz a můžeme z poslední rovnosti psát
y(t + 1) = f ′(x∗)y(t).
Odtud můžeme uvažovat, že pokud | f ′(x∗)| < 1, pak limt→∞ y(t) = 0. Analogicky pro| f ′(x∗)| > 1 bude limt→∞ y(t) = ∞. Pokud se odchylka stále zmenšuje, pak je bod x∗
asymptoticky stabilní, pokud se naopak odchylka zvětšuje, je nestabilní.
Definice 3.13. Řekneme, že rovnovážný bod rovnice (3.10) je hyperbolický, pokud platí
| f ′(x∗)| 6= 1.

Kapitola 3. Nelineární diferenční rovnice 21
Rozviňme nyní tyto úvahy a uveďme větu o stabilitě rovnice tvaru (3.10).
Věta 3.14. Nechť x∗ je rovnovážný bod rovnice (3.10) a funkce f je spojitě diferencovatelnáv bodě x∗. Pak platí
i. Je-li | f ′(x∗)|> 1, pak x∗ je nestabilní.
ii. Je-li | f ′(x∗)|< 1, pak x∗ je asymptoticky stabilní.
iii. Je-li f ′(x∗) = 1 a funkce f je v bodě x∗ dvakrát spojitě diferencovatelná, pak
a. Je-li f ′′(x∗) 6= 0, pak x∗ je nestabilní.
b. Je-li f ′′(x∗) = 0 a funkce f je třikrát spojitě diferencovatelná v bodě x∗, pak
I. Je-li f ′′′(x∗) > 0, pak x∗ je nestabilní.II. Je-li f ′′′(x∗) < 0, pak x∗ je asymptoticky stabilní.
iv. Je-li f ′(x∗) = −1 a funkce f je v bodě x∗ třikrát spojitě diferencovatelná, pak
a. Je-li f ′′′(x∗) < 32 [ f ′′(x∗)]2, pak x∗ je nestabilní.
b. Je-li f ′′′(x∗) > 32 [ f ′′(x∗)]2, pak x∗ je asymptoticky stabilní.
Důkaz. Viz. [2]
3.2.2 CyklyDefinice 3.15. Nechť b ∈Dom f , k ∈N, k > 1. Řekneme, že b je p-periodický bod rovnice(3.10), pokud f p(b) = b. V takovém případě se trajektorie
τ(b) = {b, f (b), f 2(b), . . . , f p−1(b)}
nazývá cyklus délky p (p-cyklus).Řekneme, že p-periodický bod je dosažitelný z bodu b, pokud existuje m ∈N,m≥ 1
takové, že f m(b) je p-periodický bod.
Bod b ∈Dom f je p-periodickým bodem rovnice (3.10) právě tehdy, když je rovnováž-ným bodem rovnice
x(t + 1) = f p(x(t)). (3.11)
Definice 3.16. Řekneme, že p-cyklus τ(b) rovnice (3.10) je
1. stabilní, pokud b je stabilní rovnovážný bod rovnice (3.11);
2. asymptoticky stabilní, pokud je b asymptoticky stabilní rovnovážný bod rovnice(3.11);
3. nestabilní, pokud b je nestabilní rovnovážný bod rovnice (3.11).
Věta 3.17. Nechť τ(b) = {b, f (b), f ( f (b)), . . . , f k−1(b)} = {x(0),x(1),x(2), . . . ,x(k−1)} je p-cyklus rovnice (3.10).
Je-li | f ′(x(0)) f ′(x(1)) f ′(x(2)) · · · f ′(x(p−1))|< 1, pak je τ(b) asymptoticky stabilní.Je-li | f ′(x(0)) f ′(x(1)) f ′(x(2)) · · · f ′(x(p−1))|> 1, pak je τ(b) nestabilní.
Důkaz. Viz. [2]

Kapitola 4
Populační genetika
4.1 Historie genetiky populacíGenetika populací je zaměřena obecně na zkoumání genetické struktury skupiny jedinců(populací), zejména pak na odhalení faktorů, které ovlivňují genetickou variabilitu v po-pulacích.
Charles Darwin svou prací „O původu druhů“ z roku 1859 položil základy pro studiumevoluce a vyvolal zájem o vysvětlení a význam genetické variability v populacích.
Gregor Johann Mendel, dnes po celém světě uznávaný jako zakladatel genetiky, vesvé klasické práci „Versuche über Pflanzen - Hybridea“ (1866) popsal princip segregacea princip kombinace alel (dědičných vloh). Dokumentoval, že pro počáteční populaci F0,která je odvozena ze dvou homozygotních linií (AA a aa) pro určitý dialelický gen, platí,že poměr genotypů (AA:Aa:aa) v t-té generaci je roven (2t−1):2:(2t−1).
V roce 1908 G. H. Hardy a nezávisle na něm i W. Weinberg vyřešili problém genetickéstruktury populací s náhodným oplozením. Jejich triviální zákon (například S. G. Wrighttento princip používal, aniž by někdy slyšel o jeho objevitelích) se stal základem genetikypopulací diploidních organismů.
Od první světové války do 50. let 20. století pole genetiky populací opanovala trojicegenetiků R. A. Fisher, J. B. S. Haldane a S. G. Wright. Tato trojice dala vzniknout známéa důležité rovnici genetiky populací: Fisher - Haldane - Wright (FHW ) rovnici, kterávyjadřuje pravděpodobnost frekvence alely v následující generaci.
4.2 Mendelistická genetika populací s nepřesahujícímigeneracemi
Genetika populací se zabývá organismy, které se sexuálně reprodukují. Dospělí jedinciprodukují samčí a samičí haploidní gamety (s jednou sadou autozomálních chromozomůz páru a jedním pohlavním chromozomem) z jejichž spojení vzniknou diploidní zygoty(s celým párem chromozomů).
První Mendelovo pravidlo tvrdí, že každá gameta může nést pouze jednu alelu z danéhoúseku DNA. Zygoty nesou alely dvě, jednu od každého z rodičů. Uvažujeme-li znak, jenžmá dvě alely (A a a), pak gameta nese vždy buď A, nebo a. Zygota pak může nést jednu ze
– 22 –

Kapitola 4. Populační genetika 23
tří dvojic AA, Aa a aa. Zygoty s genotypem AA a aa nazýváme homozygoty, genotyp Aa jeheterozygotem.
Označíme p četností alely A v populaci a q četností alely a. Platí, že p(t)+ q(t) = 1.Dále definujeme x, y a z jako četnosti genotypů AA, Aa a aa. Platí, že
p = x+12
y a q = z+12
y. (4.1)
Uveďme nyní předpoklady Hardyho-Weinbergova principu (zákona):
• organismy jsou diploidní;
• rozmnožování se děje pohlavní cestou;
• generace se nepřekrývají (potomci se rodí v době, kdy generace rodičů již nežije,což se děje například u některých druhů hmyzu);
• oplození je náhodné;
• početnost populace je dostatečně velká;
• migrace je zanedbatelná;
• mutace je zanedbatelná;
• na alely nepůsobí přírodní výběr.
Při splnění Hardyho-Weinbergova principu lze vyjádřit p a q v následující generacipomocí Punnettova čtverce (pojmenované po biologovi R.C. Punnettovi).
Frekvence alely u samičí gametyA a
Frekvence alely A p2 pqu samčí gamety a pq q2
Zapíšeme-li toto zjištění do diferenční rovnice, vyjde nám pro frekvenci p
p(t + 1) = p2(t)+12
2p(t)q(t) = p(t)(p(t)+ q(t)) = p(t),
tedy posloupnost p(t) je konstantní. Analogicky dojdeme k závěru, že q(t) je také kon-stantní. Zaveďme nyní t0 = 0 počáteční (rodičovskou) generaci. Pro t = 1 jsou genotypovéčetnosti x, y a z následující
x(t) = p2(t), y(t) = 2pq, z(t) = q2.

Kapitola 4. Populační genetika 24
4.3 SelekcePři absenci selekce neexistuje evoluce. Selekce je tedy hlavním faktorem změny genofondupopulace a jedním z hlavních činitelů genetiky populací. Hardyho - Weinbergovy předpo-klady počítají s tím, že každý genotyp má stejnou schopnost reprodukce. Předpokládejmenyní, že alela A má nějakou výhodu, či nevýhodu. Řekněme, že genotyp AA má jinoupravděpodobnost dožití se reprodukčního věku než genotyp aa. Genotyp Aa může být takéovlivněn. Selektivní efekty se mohou objevovat i v jiných fázích životního cyklu (obrázek4.1), popřípadě se projevovat různě, což může vést k drobným změnám v rovnici. Nicméněprincipy zůstávají stejné.
Alelové a genotypové frekvence se mohou různit v závislosti na fázi životního cyklu.Musíme tedy stanovit, v jakém bodě cyklu budeme frekvence měřit. Většinou počítámealely a genotypy na konci gametální fáze tak, abychom uvažovali nejdříve pohlaví a potéselekci. Znovu musíme brát v potaz předpoklad náhodného páření, což nám umožňujepracovat pouze s alelovými frekvencemi, nikoliv s frekvencemi genotypů.
Obrázek 4.1: Životní cyklus organismu.
Řekněme, že p(t) a q(t) jsou alelové frekvence na konci gametální fáze t-té gene-race. Tedy na začátku zygotální fáze jsou (díky předpokladu náhodného páření) frekvencegenotypu dány jako
x(t) = p2(t), y(t) = 2p(t)q(t), z(t) = q2(t).
Označme poměr pravděpodobnosti dožití se zygoty daného genotypu fáze páření jakowx : wy : wz. Pak během páření je poměr genotypů AA, Aa a aa
wx p2(t) : wy2p(t)q(t) : wzq2(t)
a alelové frekvence A a a jsou v poměru
wx p2(t)+wy p(t)q(t) : wzq2(t)+wy p(t)q(t).
Vzhledem k zanedbání pravděpodobnosti plodnosti a pravděpodobnosti přežití gamety jepoměr alel stejný na konci gametální fáze generace t + 1. Dostáváme
p(t + 1) = f (p(t),q(t)) =(wx p(t)+wyq(t))p(t)
wx p2(t)+ 2wy p(t)q(t)+wzq2(t).
což je Fisher - Haldane - Wrightova rovnice. V následující části se budeme zabývat řešenímtéto rovnice.

Kapitola 4. Populační genetika 25
4.4 Fisher - Haldane - Wrigthova rovniceUvažujme FHW rovnici pro dialelický lokus (úsek DNA, který existuje ve dvou variantách).Rovnice
p(t + 1) =(wx p(t)+wy(1− p(t)))
wx p2(t)+ 2wy p(t)(1− p(t))+wz(1− p(t))2(t)· p(t) (4.2)
s podmínkami
wx,wy,wz ≥ 0, wx +wy +wz = 1, p(0) = p0 (4.3)
je explicitně řešitelná v několika speciálních případech. Zmiňme, že pevnými body rovnice(4.2) jsou body 0 a 1, tedy řešením (4.2) jsou vždy konstantní posloupnosti p(t) ≡ 0 ap(t) ≡ 1.
I. Položme wy = wz = 0. Potom FHW můžeme psát jako
p(t + 1) =wx p2(t)wx p(t)
= 1
p(t) = 1 pro t ≥ 1.
II. Položme wx = wy = 0. Potom FHW můžeme psát jako
p(t + 1) = 0p(t) = 0 pro t ≥ 1.
III. Položme wx = wz = 0. Potom FHW můžeme psát jako
p(t + 1) =wy(1− p(t))p(t)
2wy(1− p(t))p(t)=
12
p(t) =12
pro t ≥ 1.
IV. Položme wx = 0 6= wy. Potom FHW můžeme psát jako
p(t + 1) =wy(1− p(t))p(t)
2wy(1− p(t))p(t)+wz(1− p(t))2
=p(t)
2p(t)+ wzwy(1− p(t))
=p(t)
2p(t)+ wzwy(1− p(t))
.
Počítejme pro různé hodnoty wzwy
.

Kapitola 4. Populační genetika 26
• Pro wzwy
= 2
p(t + 1) =p(t)
2p(t) =
p0
2t .
• Pro wzwy6= 2 (
2− wz
wy
)p(t + 1)p(t)+
wz
wyp(t + 1)− p(t) = 0(
2wy
wz−1)
p(t + 1)p(t)+ p(t + 1)−wy
wzp(t) = 0,
což je Riccatiho rovnice s nulovým absolutním členem (r(t)≡ 0), kterou řešímesubstitucí
p(t) =1
y(t).
Po dosazení dostáváme(2wy
wz−1)
1y(t + 1)
1y(t)
+1
y(t + 1)−
wy
wz
1y(t)
= 0
y(t + 1) =wz
wyy(t)+
(2− wz
wy
),
tedy
y(1) =wz
wyy0 +
(2− wz
wy
)y(2) =
wz
wyy(1)+
(2− wz
wy
)=
(wz
wy
)2
y0 +wz
wy
(2− wz
wy
)+
(2− wz
wy
)...
y(t) =(
wz
wy
)t
y0 +
(2− wz
wy
) t−1
∑i=0
(wz
wy
)i
.
kde y0 =1p0
. Rozlišme nyní dva případy: wz 6= wy a wz = wy.
– Pro wz 6= wy pokračujeme v úpravě poslední rovnice a dostáváme
y(t) =(
wz
wy
)y0 +
(2− wz
wy
)(
wzwy
)t−1
wzwy−1
=
(1p0−
wzwy−2
wzwy−1
)(wz
wy
)t
+
wzwy−2
wzwy−1
=
=((1− p0)wz +(2p0−1)wy)
(wzwy
)t+ p0(wz−2wy)
p0(wz−wy).

Kapitola 4. Populační genetika 27
Řešení původní rovnice je tedy tvaru
p(t) =(wz−wy)p0
p0(wz−2wy)+ ((1− p0)wz +(2p0−1)wy)(
wzwy
)t
– Pro wz = wy dostáváme po substituci p(t) = 1y(t)
y(t + 1) =wz
wyy(t)+
(2− wz
wy
)= y(t)+ 1.
Řešením této rovnice je
y(t) = y0 + t
=1p0
+ t.
Potom x(t) jep(t) =
p0
1+ t p0.
V. Položme wz = 0 6= wy, pak FHW můžeme psát jako
p(t + 1) =wx p(t)+wy(1− p(t))
wx p(t)+ 2wy(1− p(t)).
Po úpravě dostáváme
(wx−2wy)
2wyp(t + 1)p(t)+ p(t + 1)+
wy−wx
2wyp(t)− 1
2= 0, (4.4)
což je Riccatiho rovnice, kterou řešíme substitucí (3.3)
p(t) =2wy
wx−2wy
(y(t + 1)
y(t)−1)
.
Vzniká rovnice
2wy
wx−2wyy(t + 2)+
3w2y−w2
x
w2x−3wywx + 2w2
yy(t + 1)+
12
wx
wx−2wy. (4.5)
Tuto rovnici již umíme řešit pomocí metod pro řešení lineárních diferenčních rovnicvyšších řádů. Označme fundamentální řešení rovnice (4.5) λ1 a λ2 a dále označmeC = c1
c2. Výsledek rovnice (4.4) pak můžeme psát jako
p(t) =2wy
wx−2wy
(Cλ
t+11 +λ
t+12
Cλ t1 +λ t
2−1
).
Vzhledem k substituci má ovšem řešení podmínky wxwy6= 2 a wx
wy6= 1. Rozeberme tedy
tyto dva případy zvlášť.

Kapitola 4. Populační genetika 28
• Pro wxwy
= 2 dostáváme
p(t + 1) =2− p(t)
4−3p(t)
− 34
p(t + 1)p(t)+ p(t + 1)+14
p(t)− 12= 0,
což je Riccatiho rovnice, kterou řešíme substitucí
p(t) =(−4
3
)(y(t + 1)
y(t)−1)
.
Dostávámey(t + 2)− 3
4y(t + 1)+
18
y(t) = 0.
Rovnice má řešeníλ1 =
12
, λ2 = −12
y(t) = c1
(12
)t
+ c2
(−1
2
)t
.
Označíme-li C = c1c2
, dostáváme po dosazení řešení
p(t) =
(C(1
2
)t+1+(1
2
)t+1
C(1
2
)t+(1
2
)t −1
)(−4
3
)= −2
3C+(−1)t+1
C+(−1)t +43
.
• Pro wxwy
= 1 dostáváme
p(t + 1) =1
2− p(t),
kterou řeším substitucí
p(t) = (−2)(
y(t + 1)y(t)
−1)
.
Dostávámey(t + 2)− y(t + 1)+
14= 0
a řešení této rovnice
y(t) = c1
(12
)t
+ c2
(12
)t
.
Po dosazení dostáváme řešení původní rovnice
p(t) = (−2)
(c1(1
2
)t+1+ c2t
(12
)t+1
c1(1
2
)t+ c2t
(12
)t −1
)= −c1 + c2t
c1 + c2t+ 2
= 1.

Kapitola 4. Populační genetika 29
VI. Zvolme wy = 0,wx 6= 0 a wz 6= 0. Pak FHW je tvaru
p(t + 1) =wx p2(t)
wx p2(t)wy(1− p2(t)), (4.6)
kterou dále řešíme pomocí logaritmizace
p(t + 1) =1
+1 wzwx
(1
p(t) −1)2
1p(t + 1)
−1 =wz
wx
(1
p(t)−1)2
ln(
1p(t + 1)
)= 2ln
(1
p(t)−1)+ ln
wz
wx
ln(
1p(t)−1)=
(ln(
1p0−1)+ ln
wz
wx
)2t− ln
wz
wx=
= lnwz
wx
(wz
wx
1− p0
p0
)2t
1p(t)−1 =
wz
wx
(wz
wx
1− p0
p0
)2t
p(t) =1
1+ wxwz
(wzwx
1−p0p0
)2t =wz
wz +wx
(wzwx
1−p0p0
)2t
=wz(wx p0)2t
wz(wx p0)2t +wx(wz(1− p0))2t .
VII. Položme wxwy
=wywz
. Pokud zavedeme K = wxwy
, můžeme FHW psát jako
p(t + 1) =K p(t)+ 1− p(t)
K p2(t)+ 2p(t)(1− p(t))+ 1K (1− p(t))2
p(t).
Rovnici upravíme na tvar
p(t + 1) =K p(t)
(K−1)p(t)+ 1,
což je Riccatiho rovnice
(K−1)p(t + 1)p(t)+ p(t + 1)−K p(t) = 0
s nulovou pravou stranou (r(t) ≡ 0), kterou řešíme substitucí
p(t) =1
y(t).

Kapitola 4. Populační genetika 30
Řešením rovnice
1y(t + 1)
=1
K−1K + 1
K y(t)
y(t + 1) =1K
y(t)+(
1− 1K
)je posloupnost
y(t) =1− p0
p0
(1K
)t
+ 1 =p0 +(1− p0)K−t
p0,
čili
p(t) =p0 +(1− p0)
(wxwy
)t
p0.
Zabývejme se nyní obecným řešením FHW rovnice. Explicitně jej nejsme schopnizískat, proto vyřešme rovnovážné body rovnice a jejich stabilitu.
Případy, kdy některý z koeficientů wx, wy, wz je nulový, jsme vyřešili v částech I. - V I.Proto předpokládejme, že wx 6= 0, wy 6= 0 a wz 6= 0. Označme K = wx
wy> 0 a k = wz
wy> 0.
Potom FHW je tvaru
p(t + 1) =(K p(t)+ 1− p(t))p(t)
K p2(t)+ 2p(t)(1− p(t))+ k(1− p(t))2
=1+(K−1)p(t)
1+(K−1)p2 +(k−1)(1− p(t))· p(t).
Rovnovážně body zjistíme vyřešením rovnice
p =1+(K−1)p
1+(K−1)p2 +(k−1)(1− p)p = f (p)
p(
1− 1+(K−1)p1+(K−1)p2 +(k−1)(1− p)
)= 0.
Jedním řešením je tedy p1 = 0 a druhé dostaneme řešením rovnice(1− 1+(K−1)p
1+(K−1)p2 +(k−1)(1− p)
)= 0
1+(K−1)p = 1+(K−1)p2 +(k−1)(1− p)
(K−1)p(p−1)+ (k−1)(1− p)2 = 0(1− p)(−(K−1)p+(k−1)(1− p)) = 0(1− p)(k−1− (K−1+ k−1)p) = 0.
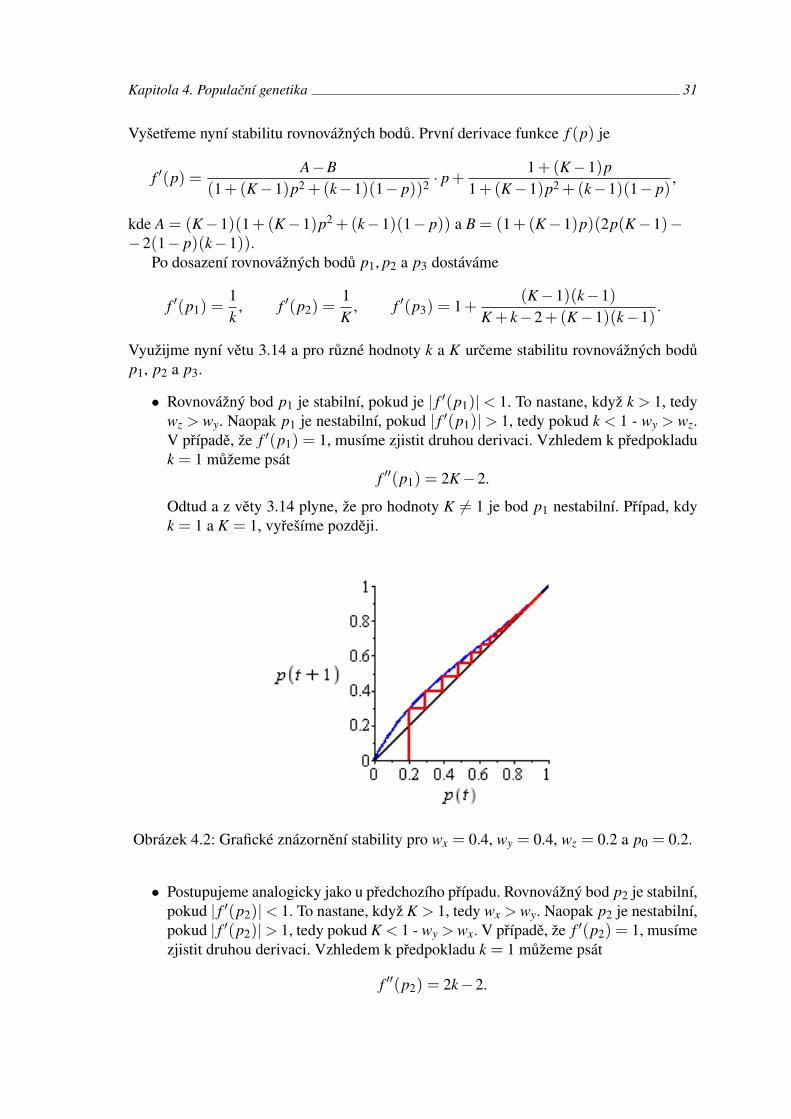
Odtud dostáváme rovnovážné body
p2 = 1, p3 =k−1
K + k−2.
Kapitola 4. Populační genetika 31
Vyšetřeme nyní stabilitu rovnovážných bodů. První derivace funkce f (p) je
f ′(p) =A−B
(1+(K−1)p2 +(k−1)(1− p))2 · p+1+(K−1)p
1+(K−1)p2 +(k−1)(1− p),
kde A = (K−1)(1+(K−1)p2 +(k−1)(1− p)) a B = (1+(K−1)p)(2p(K−1)−−2(1− p)(k−1)).
Po dosazení rovnovážných bodů p1, p2 a p3 dostáváme
f ′(p1) =1k
, f ′(p2) =1K
, f ′(p3) = 1+(K−1)(k−1)
K + k−2+(K−1)(k−1).
Využijme nyní větu 3.14 a pro různé hodnoty k a K určeme stabilitu rovnovážných bodůp1, p2 a p3.
• Rovnovážný bod p1 je stabilní, pokud je | f ′(p1)|< 1. To nastane, když k > 1, tedywz > wy. Naopak p1 je nestabilní, pokud | f ′(p1)|> 1, tedy pokud k < 1 - wy > wz.V případě, že f ′(p1) = 1, musíme zjistit druhou derivaci. Vzhledem k předpokladuk = 1 můžeme psát
f ′′(p1) = 2K−2.
Odtud a z věty 3.14 plyne, že pro hodnoty K 6= 1 je bod p1 nestabilní. Případ, kdyk = 1 a K = 1, vyřešíme později.
Obrázek 4.2: Grafické znázornění stability pro wx = 0.4, wy = 0.4, wz = 0.2 a p0 = 0.2.
• Postupujeme analogicky jako u předchozího případu. Rovnovážný bod p2 je stabilní,pokud | f ′(p2)|< 1. To nastane, když K > 1, tedy wx > wy. Naopak p2 je nestabilní,pokud | f ′(p2)|> 1, tedy pokud K < 1 - wy > wx. V případě, že f ′(p2) = 1, musímezjistit druhou derivaci. Vzhledem k předpokladu k = 1 můžeme psát
f ′′(p2) = 2k−2.
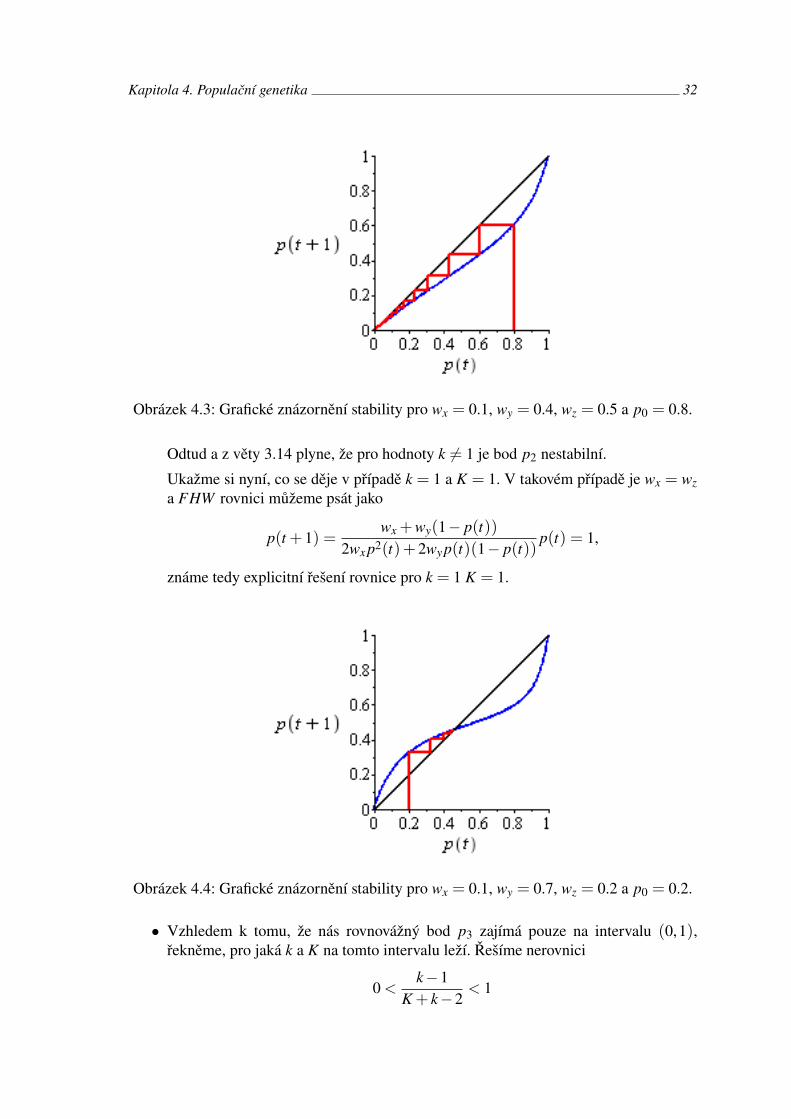
Kapitola 4. Populační genetika 32
Obrázek 4.3: Grafické znázornění stability pro wx = 0.1, wy = 0.4, wz = 0.5 a p0 = 0.8.
Odtud a z věty 3.14 plyne, že pro hodnoty k 6= 1 je bod p2 nestabilní.
Ukažme si nyní, co se děje v případě k = 1 a K = 1. V takovém případě je wx = wza FHW rovnici můžeme psát jako
p(t + 1) =wx +wy(1− p(t))
2wx p2(t)+ 2wy p(t)(1− p(t))p(t) = 1,
známe tedy explicitní řešení rovnice pro k = 1 K = 1.
Obrázek 4.4: Grafické znázornění stability pro wx = 0.1, wy = 0.7, wz = 0.2 a p0 = 0.2.
• Vzhledem k tomu, že nás rovnovážný bod p3 zajímá pouze na intervalu (0,1),řekněme, pro jaká k a K na tomto intervalu leží. Řešíme nerovnici
0 <k−1
K + k−2< 1
Kapitola 4. Populační genetika 33
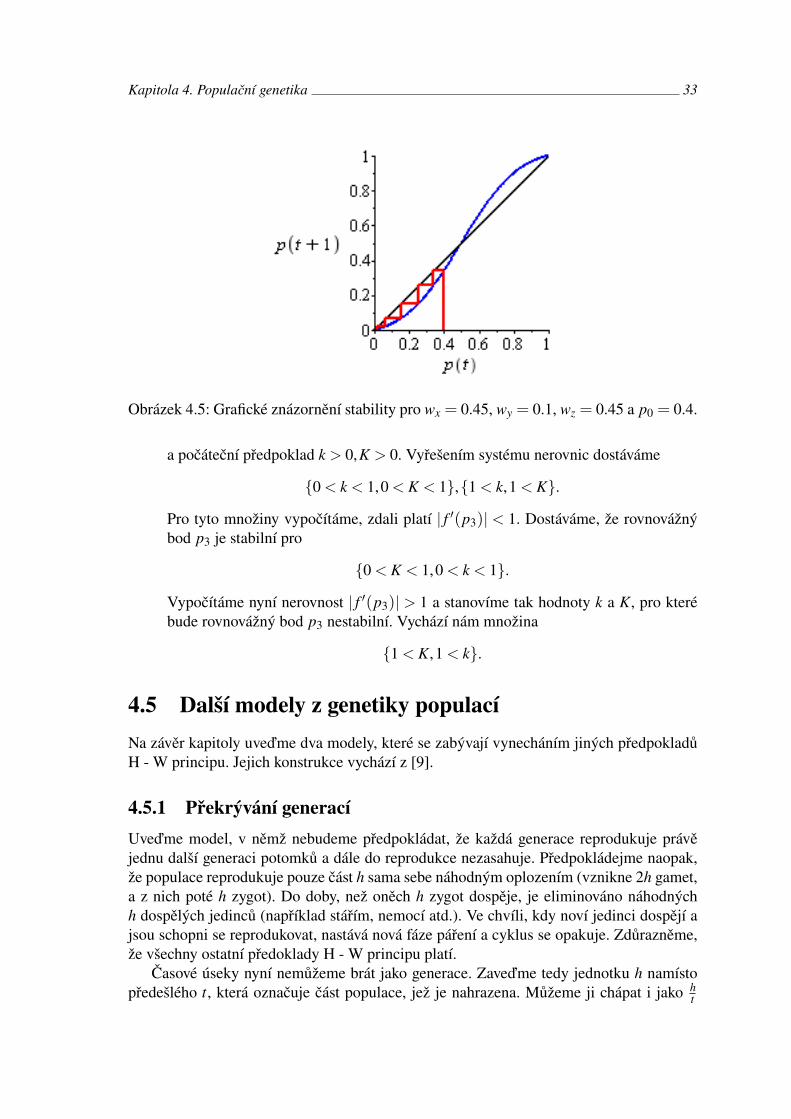
Obrázek 4.5: Grafické znázornění stability pro wx = 0.45, wy = 0.1, wz = 0.45 a p0 = 0.4.
a počáteční předpoklad k > 0,K > 0. Vyřešením systému nerovnic dostáváme
{0 < k < 1,0 < K < 1},{1 < k,1 < K}.
Pro tyto množiny vypočítáme, zdali platí | f ′(p3)| < 1. Dostáváme, že rovnovážnýbod p3 je stabilní pro
{0 < K < 1,0 < k < 1}.
Vypočítáme nyní nerovnost | f ′(p3)| > 1 a stanovíme tak hodnoty k a K, pro kterébude rovnovážný bod p3 nestabilní. Vychází nám množina
{1 < K,1 < k}.
4.5 Další modely z genetiky populacíNa závěr kapitoly uveďme dva modely, které se zabývají vynecháním jiných předpokladůH - W principu. Jejich konstrukce vychází z [9].
4.5.1 Překrývání generacíUveďme model, v němž nebudeme předpokládat, že každá generace reprodukuje právějednu další generaci potomků a dále do reprodukce nezasahuje. Předpokládejme naopak,že populace reprodukuje pouze část h sama sebe náhodným oplozením (vznikne 2h gamet,a z nich poté h zygot). Do doby, než oněch h zygot dospěje, je eliminováno náhodnýchh dospělých jedinců (například stářím, nemocí atd.). Ve chvíli, kdy noví jedinci dospějí ajsou schopni se reprodukovat, nastává nová fáze páření a cyklus se opakuje. Zdůrazněme,že všechny ostatní předoklady H - W principu platí.
Časové úseky nyní nemůžeme brát jako generace. Zaveďme tedy jednotku h namístopředešlého t, která označuje část populace, jež je nahrazena. Můžeme ji chápat i jako h
t

Závěr 34
generaci. Pak k páření dochází v časech h,2h,3h, . . .. A tedy za dobu t = 1h se narodí celá
populace (což nutně neznamená, že zemřou všichni z původní generace, můžeme tedyvskutku mluvit o překrývání generací).
Označme nyní p(h), p(2H), . . . a q(h),q(2h), . . . alelové frekvence na konci každé fázepáření a stejně označme frekvence genotypů AA, Aa a aa x(h),x(2h), . . . atd. Zabývejme senyní genotypem AA a označme [t, (t+h)] časový úsek mezi dvěma pářeními. Z předchozíchúvah můžeme říct, že
x(t + h)− x(t) = h[p2(t)− x(t)].
Vzhledem k tomu, že p(t) je konstantní posloupností (jelikož neuvažujeme selekci nazkoumaný gen a dožití se reprodukce, smrt či další fáze cyklu jsou náhodné), můžemepsát, že
x(t + h) = [1−h]x(t)+ hp20.
Jelikož jsou fáze páření v časových úsecích h,2h, . . ., přepíšeme rovnici do tvaru
x(kh) = (1−h)k(x0− p20)+ p2
0
a tedyx(∞) = lim
k→∞(kh) = p2
0.
Analogicky dostáváme pro genotypy Aa a aa
y(∞) = 2p0q0, z(∞) = q20.
4.5.2 MutacePředpokládejme nyní všechny podmínky H - W principu kromě zanedbání mutace. K mu-taci dochází zejména při tvorbě gamety (bodové a chromozomální mutace) a může k nídojít i při tvorbě zygoty (chromozomální mutace). Gameta může obsahovat chybné či poš-kozené úseky DNA a genotyp zygoty se tedy může vyskytnout v jiné než předpokládanéformě. V našem případě se budeme zabývat situací, kdy alela A zmutuje v alelu a s prav-děpodobností u či obráceně s pravděpodobností v. Předpokládejme navíc, že 0 < u+ v.Označme PA(t + 1) pravděpodobnost, že potomek dostane alelu A od náhodně zvolenéhorodiče. K tomu, aby potomek dostal alelu A, dojde v případě, že alelu A zdědí a zároveňalela A nezmutuje nebo zdědí alelu a, která zmutuje. První nastává s pravděpodobností(1−u)PA(t). Pravděpodobnost druhé situace je v(1−PA(t)). Pak
PA(t + 1) = (1−u)PA(t)+ v(1−PA(t)) = v+(1−u− v)PA(t).
Vzniklou diferenční rovnici řešíme a dostáváme
PA(t) = (1−u− v)t(
PA(0)−v
u+ v
)+
vu+ v
.

Závěr
Genetika populací, jako součást genetiky, je vedle genomiky jedním z nejprogresivnějšíchoborů. Aplikovaná genetika populací může být zajímavým sociologickým nástrojem přianalýze lidských společností a jejich vývoje. FHW rovnice nám může pomoci říct (s větši-nou zachovaných předpokladů), zda i v sofistikované společnosti dochází k selekci. Jed-noznačně aplikovatelná je však u živočichů, zejména pak u těch, kteří lidem přinášejíužitek. U volně žijících druhů (jejichž parentální generaci nemůžeme vybírat) můžeme”pouze”zkoumat strukturu a vývoj jejich genofondu.
Genetika populací tak nabývá na svém významu u plemenných zvířat. U takových totižmůžeme vybírat genofond parentální generace a přímo ovlivňovat jeho vývoj. Z obecněekologického pohledu, který zahrnuje i trvalou udržitelnost tzv. genových zdrojů hospo-dářských zvířat, je nutné udržovat v takovýchto populacích co největší diversitu (různo-rodost) genofondu proto, aby se populace udržela životaschopná. Je tedy velice zajímavépomocí modelu optimalizovat diversitu genových zdrojů. Další významnou aplikací u ple-menných zvířat je tzv. genomická selekce. Díky detekci stovky tisíc alel, jejichž skupinyjsou označované jako haplotypy (genom gamety) a které mají vliv na užitkovost zvířat, jemožné za pomoci matematických modelů docílit maximální užitkovosti filiální generace.
Ovšem model, který by bylo možné použít v aplikaci pro chov plemenných zvířat,by musel projít mnoha změnami. V našich příkladech jsme se zabývali pouze jednímdialelickým genem, který by nám v aplikaci dozajista nestačil. Při ověřování parentityzvířete se zkoumá více úseků DNA (markerů nebo také lokusů). V České republice jeto například u prasat 12 lokusů [7] či u koní 17 lokusů [8]. Také se většinou nejednáo dialelické lokusy. Na daném lokusu může být více alel a tedy i více než tři různé genotypy.Dalším problémem je předpoklad dostatečně velké populace. [4] uvádí, že dostatečně velkápopulace čítá více než 500 jedinců. To je většinou příliš velké číslo, jelikož genová diversitanás zajímá především u genových zdrojů, které slouží k udržení původně chovaných plemenv daných regionech, které mohou nést vzácné, specifické a významné alely (jež se mohouuplatnit v budoucnosti), a které se vyskytují většinou v malých populacích, ve kterýchse může pravděpodobně projevit inbreeding, genetický drift nebo mutace. Kupodivu všakpraxe přináší i zjednodušení. Zkoumané úseky DNA jsou vybírány tak, aby měly conejmenší vliv na fenotyp. Je tedy možné zanedbat selekci na užitkovost a přírodní selekci.Na druhou stranu selekci můžeme do modelu zahrnout jako námi zvolené parametry(v našem případě parametry wx, wy a wz), které nastavujeme tak, aby rovnice splňovalanaše cíle (například diversitu či užitkovost filiální generace).
– 35 –

Literatura
[1] ELAYDI, Saber N. An introduction to difference equations. 3rd ed. New York: Sprin-ger, 2005. xxii, 539. s ISBN 0-387-23059-9
[2] POSPÍŠIL, Zdeněk. Diskrétní deterministické modely [online]. [cit. 2015-05-08].Dostupné z: http://is.muni.cz/el/1431/podzim2014/M8230/um/DiskrMod.pdf
[3] KELLEY, Walter G., PETERSON, Allan C., Difference Equations, Second Edition:An Introduction with Applications. Academic Press; 2 edition (May 19, 2000), 403 s.ISBN 978-0124033306
[4] RELICHOVÁ, Jiřina. Genetika populací. 1. vyd. Brno: Masarykova univerzita, 2009.188 s. ISBN 978-80-210-4795-2
[5] BRITTON, N. F. Essential mathematical biology. London: Springer, 2003. xv, 335 s.ISBN 1-85233-536-X
[6] BACAER, Nicolas, A Short History of Mathematical Population Dynamics. Springer-Verlag London, 2011, 153 s. ISBN 978-0-85729-114-1
[7] VRTKOVÁ, Irena. Genetic admixture analysis in Prestice Black-Pied pigs [on-line]. [cit. 2015-3-23]. Dostupné z: http://www.arch-anim-breed.net/58/115/2015/aab-58-115-2015.pdf
[8] VRTKOVÁ, Irena. Predikce zbarvení, validace parentit a prezentace EFABIS u po-pulace koní Kinských v ČR s využitím polymorfismů genomické DNA. 1. vyd. V Brně:Laboratoř agrogenomiky, Mendelova univerzita, 2011, 64 s. ISBN 978-80-7157-239-8.
[9] EWENS, W. Mathematical population genetics. 2nd ed. New York: Springer, ISBN03-872-0191-2.
[10] Vandermondův determinant. [online]. [cit. 2015-05-22]. Dostupné z: http://www.kolej.mff.cuni.cz/~lmotm275/skripta/sbirka/html/node80.html
– 36 –






























