Embed Size (px)
Citation preview

Different Structural Correlates for Verbal Memory Impairment inTemporal Lobe Epilepsy with and Without Mesial Temporal LobeSclerosis
Susanne G. Mueller1,*, Kenneth D. Laxer2, Cathy Scanlon1, Paul Garcia3, William J.McMullen2, David W. Loring4, Kimford J. Meador4, and Michael W. Weiner1
1Department of Radiology, Center for Imaging of Neurodegenerative Diseases, University ofCalifornia, San Francisco2Pacific Epilepsy Program, California Pacific Medical Center, San Francisco3Department of Neurology, University of California, San Francisco4Department of Neurology, Emory University, Atlanta
AbstractObjectives—Memory impairment is one of the most prominent cognitive deficits in temporallobe epilepsy (TLE). The overall goal of this study was to explore the contribution of cortical andhippocampal (subfield) damage to impairment of auditory immediate recall (AIMrecall), auditorydelayed recall (ADMrecall), and auditory delayed recognition (ADMrecog) of the WechslerMemory Scale III (WMS-III) in TLE with (TLE–MTS) and without hippocampal sclerosis (TLE-no). It was hypothesized that volume loss in different subfields determines memory impairment inTLE–MTS and temporal neocortical thinning in TLE-no.
Methods—T1 whole brain and T2-weighted hippocampal magnetic resonance imaging andWMS-III were acquired in 22 controls, 18 TLE–MTS, and 25 TLE-no. Hippocampal subfieldswere determined on the T2 image. Free surfer was used to obtain cortical thickness averages oftemporal, frontal, and parietal cortical regions of interest (ROI). MANOVA and stepwiseregression analysis were used to identify hippocampal subfields and cortical ROI significantlycontributing to AIMrecall, ADMrecall, and ADMrecog.
Results—In TLE–MTS, AIMrecall was associated with cornu ammonis 3 (CA3) and dentate(CA3&DG) and pars opercularis, ADMrecall with CA1 and pars triangularis, and ADMrecog withCA1. In TLE-no, AIMrecall was associated with CA3&DG and fusiform gyrus (FUSI), andADMrecall and ADMrecog were associated with FUSI.
Conclusion—The study provided the evidence for different structural correlates of the verbalmemory impairment in TLE–MTS and TLE-no. In TLE–MTS, the memory impairment wasmainly associated by subfield-specific hippocampal and inferior frontal cortical damage. In TLE-no, the impairment was associated by mesial–temporal cortical and to a lesser degree hippocampaldamage.
KeywordsTLE; CA1; CA3; dentate gyrus; MRI; recognition; recall; mesial temporal sclerosis; fusiform
© 2011 Wiley-Liss, Inc.*Correspondence to: Susanne G. Mueller, Department of Veterans Affairs (DVA) Medical Center, Center for Imaging ofNeurodegenerative Diseases, Clement Street 4150, San Francisco, CA 94121, USA. [email protected] supporting information may be found in the online version of this article.
NIH Public AccessAuthor ManuscriptHum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
Published in final edited form as:Hum Brain Mapp. 2012 February ; 33(2): 489–499. doi:10.1002/hbm.21226.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

INTRODUCTIONMesial temporal lobe structures, particularly, the hippocampus, play a major role indeclarative memory [Eichenbaum, 2004]. Nonetheless, they are just one element in thewidespread networks of cortical and subcortical brain structures supporting declarativememory functions. Different subtypes of declarative memory use different brain structuresor networks although there is also some overlap. For example, posterior–inferior parietal,middle, and mesial temporal; dorsomedial, ventromedial, and inferior–lateral prefrontal; andposterior cingulate have been implicated in semantic memory, while hippocampus, mesialtemporal, anterior and inferior–lateral prefrontal, precuneus (PRE), lateral parietal, lingualregions, and anterior cingulate have been implicated in episodic memory [Binder et al.,2009; Burianova and Grady, 2007; Desganges et al., 1998; Moscovitch et al., 2005]. Withina network, different brain structures and sometimes even different parts of the same brainstructure are specialized in different aspects of the process. An example for specializationbetween different brain structures is the finding of functional imaging studies, showingactivity in the left prefrontal cortex during encoding of verbal information and in the rightprefrontal cortex during retrieval [Cabeza and Nyberg, 2000; Desgranges et al., 1998]. Thehippocampus on the other hand provides an example for specialization within a brainstructure. It consists of several histologically distinct subfields: subiculum, cornu ammonissectors (CA) 1–3, and dentate gyrus (DG) [Duvernoy, 2005]. These histological differencesalso translate into functional differences. Animal studies suggest that DG and CA3 areprimarily responsible for encoding and early retrieval while CA1 is responsible for delayedretrieval and novelty detection, that is, functions associated with recognition [Acsady andKali, 2007; Hunsaker and Kesner, 2008; Nakazawa et al., 2004; O’Reilly et al., 2001; Rollsand Kesner, 2006; Wan et al., 1999]. Considering the complexity of memory processes andthe widespread networks of specialized cortical and subcortical structures supporting them,any interruption within a network regardless of the localization can be expected to impairthe network performance and thus also the function supported by it. The severity of thedeficit though will vary depending on the function of the affected structure and the ability ofthe remaining network to compensate for its dysfunction.
Medial temporal lobe epilepsy (TLE) is characterized by seizures originating in mesialtemporal lobe structures, and, consequently, declarative memory deficits are one of the mostcommon and most prominent cognitive deficits in TLE [Jokeit and Schacher, 2004]. Basedon imaging and histopathologicial findings, two types of medial TLE are distinguished: TLEwith mesial–temporal lobe sclerosis (TLE–MTS, about 60–70%) is characterized by anatrophied hippocampus with MR signal abnormalities and severe neuron loss in thehistological examination (MTS), and TLE with a normal appearing hippocampus on themagnetic resonance imaging (MRI; TLE-no, about 30–40%) and no or mild neuron loss inthe histological examination. Impairment of declarative memory is found in both subtypesalthough the deficits tend to be more severe in TLE–MTS [Alessio et al., 2004; Dulay et al.,2004]. The pronounced ipsilateral hippocampal atrophy of TLE–MTS provides an obviousstructural correlate for the memory impairment in this TLE subgroup. Furthermore, giventhe functional specialization of different subfields, different patterns of subfield atrophy[Mueller et al., 2009a) might be associated with different aspects of memory impairment.However, recent studies [Berhardt et al., 2008, Lin et al., 2007] have demonstratedwidespread temporal and extratemporal cortical thinning in TLE–MTS, which also affectsmemory relevant cortical regions. It is therefore possible that damage to these corticalregions contributes to the memory impairment caused by the hippocampal damage.Widespread temporal lateral and extratemporal cortical thinning have also beendemonstrated in TLE-no [Mueller et al., 2009b]. This suggests cortical thinning in memoryrelevant regions as the structural correlate for the memory impairment in this subgroup.
Mueller et al. Page 2
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

However, it cannot be excluded that subtle hippocampal damage, for example, mild DGdamage also contributes to the memory impairment in TLE-no. The overall goal of thisstudy was therefore to explore how cortical and hippocampal damage contribute to verbalmemory impairment as measured by the auditory immediate recall, delayed recall, andauditory delayed recognition scores of the Wechsler Memory Scale III (WMS-III). Inparticular, the following hypotheses were tested. (1) Memory impairment in TLE–MTS ismostly caused by hippocampal damage. Specifically, it will be tested if impaired immediatememory performance reflecting a deficient encoding process is associated with volume lossin CA3 and dentate and impaired recognition performance with volume loss in CA1.Cortical thinning, particularly in mesial temporal and prefrontal regions, that is, regions thatare involved in episodic memory [Desgranges et al., 1998; Nyberg, 1998] and are known tobe affected in TLE–MTS aggravates the memory impairment caused by the hippocampaldamage. (2) Memory impairment in TLE-no is associated with cortical thinning in mesialtemporal [fusiform and parahippocampal gyrus (PARA)] and prefrontal lateral regions.Subtle hippocampal damage in TLE-no contributes to the impairment of verbal memorycaused by the damage to cortical structures.
METHODSStudy Population
The committees of human research at the University of California, San Francisco (UCSF),California Pacific Medical Center, San Francisco, and VA Medical Center, San Franciscoapproved the study, and written informed consent was obtained from each subject accordingto the Declaration of Helsinki. Forty-three patients with drug-resistant TLE undergoingevaluation for epilepsy surgery who agreed to undergo a dedicated imaging researchprotocol at 4 Tesla were recruited between mid 2006 and end of 2009 from the PacificEpilepsy Program, California Pacific Medical Center and the Northern CaliforniaComprehensive Epilepsy Center, UCSF (Table I). Eighteen patients (mean age 41.2 ± 11.9;mean years of education: 14.3 ± 2.0; left TLE/right TLE: 7/11, females/males 12/6) hadevidence for mesial temporal lobe sclerosis on their 1.5-T MR images (TLE–MTS) by visualassessment. The lateralization of the MTS was concordant with the side of seizure onset in16 and discordant in 2. Twenty-five patients (mean age 39.9 ± 10.1; mean years ofeducation: 15.3 ± 1.9; left TLE/right TLE: 13/8, bilateral 4; females/ males: 10/15) hadnormal appearing hippocampi on their 1.5-T MR exams (TLE-no). Twenty-two TLE-no hadcompletely normal 1.5 T MR reads. Of the remaining TLE-no, one had a small cystic lesionin the left inferior temporal pole, another had an ectopic gray matter mass adjacent to theright amygdala, and one had a small left frontal venous angioma. The identification of theepileptogenic focus was based on seizure semiology and prolonged ictal and interictal video/EEG/telemetry recordings in all patients. All patients had been seizure free for at least 24 hbefore the MRI. TLE–MTS were significantly younger at the onset of epilepsy (TLE–MTS11.4 ± 9.5 years. TLE-no: 25.6 ± 12.6 years, P = 0.0003) and had a significantly longerduration of epilepsy (TLE–MTS: 29.8 ± 12.7 years, TLE-no: 14.4 ± 13.6 years, P = 0.0005)than TLE-no. The control population consisted of 22 healthy volunteers (mean age 34.9 ±11.4; mean years of education: 15.1 ± 2.2; females/ males: 17/5). All subjects underwent anextensive neuropsychological test battery to assess their cognitive function. TLE–MTSperformed significantly worse compared to controls but not to TLE-no regarding full-scaleintelligence and performance intelligence (Wechsler Adult Intelligence Scale: Full IQ:Controls: 112.3 ± 14.7; TLE–MTS: 98.6 ± 14.0; TLE-no: 104.1 ± 15.2, P = 0.016.Performance IQ: Controls: 112.5 ± 15.1; TLE–MTS: 96.8 ± 10.5; TLE-no: 105.4 ± 15.1).The three groups were not different regarding verbal intelligence performance (WechslerAdult Intelligence Scale Verbal IQ: Controls: 110.4 ± 13.9; TLE–MTS: 100.6 ± 16.3; TLE-no 102.4 ± 14.4, P = 0.08). From this battery, the following standard scores of the WMS-III
Mueller et al. Page 3
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

were selected for the purpose of this study: auditory immediate memory tested by recall(AIMrecall), auditory delayed memory tested by recall (ADMrecall), and auditoryrecognition delayed memory tested by recognition (ADMrecog). AIMrecall can be regardedas a measure of rapid encoding and early consolidation and ADMrecall as a measure ofretrieval performance while ADMrecog provides a measure of memory performance byrecognition.
MRI AcquisitionAll imaging was performed on a Bruker MedSpec 4 T system. The following sequenceswere acquired. (1) For the measurement of hippocampal subfields, a high-resolution T2-weighted fast spin echo sequence (TR/TE: 3,990/ 21 ms, echo train length 15, 18.6-ms echospacing, 160° flip angle, 100% oversampling in ky direction, 0.4 × 0.4 mm in planeresolution, 2-mm slice thickness, 24 interleaved slices without gap, acquisition time 5:30min, and angulated perpendicular to the long axis of the hippocampal formation). (2) For themeasurement of total hippocampal volume, a volumetric T1-weighted gradient echo MRI(MPRAGE) (TR/TE/TI = 2,300/3/950 ms, 7° flip angle, 1.0 × 1.0 × 1.0 mm3 resolution, andacquisition time, 5:17 min) and 3. For the determination of the intracranial volume (ICV), aT2-weighted turbospin echo sequence (TR/ TE 8,390/70 ms, 150° flip angle, 0.9 × 0.9 × 3-mm nominal resolution, 54 slices, and acquisition time 3:06 min).
Hippocampal Subfield VolumetryThe method used for subfield marking including assessment of measurement reliability andits limitations has been described in detail previously [Mueller et al., 2007, 2009a]. Tobriefly summarize it, the marking scheme depends on anatomical landmarks, particularly ona hypointense line representing myelinated fibers in the stratum moleculare/lacunosum[Eriksson et al. 2008], which can be reliably visualized on these high resolution images. Thedistance between this hypointense line and the outer boundary of the hippocampus providesan estimate of the cortical thickness of the hippocampus at this point. Additional externaland internal hippocampal landmarks are used to further subdivide the hippocampus intosubiculum, CA1, CA1-2 transition zone (CA1-2 transition), CA3, and DG. The latter twoare lumped together (CA3&DG), because there are no macroscopic landmarks to separatethem (cf. Fig. 1a). CA1-2 transition is in the dorsal medial region of the hippocampus andconsists mostly of CA2. However, due to the landmarks used for labeling it, its volume isinfluenced by the thickness of the dorsal CA1. To reflect this fact, the sector is called CA1-2transition rather than CA2. The volumes from the left and right hemisphere were combined,that is, added to provide a single measure from each subfield to be used in the analysis. ICVwas determined from the T2-weighted image, which was skull-stripped using the BETprogram (FMRIB Image Analysis Group, Oxford University, www.fmrib.ox.ac.uk/fsl). Tocorrect for volume differences due to different head sizes, all volumes were normalized tothe ICV using the following formula: normalized volume = raw volume × 1,000 ccm/ICVccm.
Cortical Thickness MeasurementAll T1 images were segmented using the Expectation Maximization algorithm [van Leemputet al., 1999a,b]. The bias field maps and tissue maps obtained from this process were usedfor bias correction and skull stripping of the T1 image. FreeSurfer (version 3.05,https://surfer.nmr.mgh.harvard.edu) was used for cortical surface reconstruction andestimation of the mean cortical thickness of 34 anatomical regions in each hemisphere [Daleet al., 1999; Desikan et al., 2006]. The following cortical regions commonly associated withepisodic verbal memory [Desgranges et al., 1998] of each hemisphere were selected ascortical regions of interest (ROI) for this project: PARA, fusiform gyrus (FUSI), parsorbitalis, pars opercularis (POP), pars triangularis (PT), caudal middle frontal and rostral
Mueller et al. Page 4
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

middle frontal, superior frontal gyrus, PRE, lobulus parietalis superior, lobulus parietalisinferior, and supramarginal gyrus (cf. Fig. 1b). The thickness measures from the left andright hemisphere were combined, that is, added to provide a single measure from eachregion. This approach was chosen (1) to account for the findings of functional imagingstudies in healthy controls [Desgranges et al., 1998], which did not show a clear preferencefor the left or right side in verbal tasks but rather provided evidence that both sides areinvolved although differently; (2) to account for potential compensatory inter-hemisphericshifts of the verbal memory function, which has been described in TLE [Bettus et al., 2009;Campo et al., 2009; Wagner et al., 2008; Richardson et al., 2003].
Statistical AnalysisMANCOVA tests with subfield volumes, cortical ROIs, and memory scores as dependentvariables were used to identify nuisance variables explaining a significant amount of thevariation of these parameters. In addition to group (TLE–MTS, TLE-no, and control), age,gender, and years of education (log transformed) were modeled as nuisance variables, andthus their contribution to the variability of each of the dependent variables (subfield, corticalROI, and verbal memory) was tested. Multiple linear regression analysis with group and theco-variates identified in the previous steps followed by Tukey’s post hoc tests were used totest for group differences of normalized hippocampal subfield volumes, cortical thicknessand of AIMrecall, ADMrecall, and ADMrecog.
The population was divided into two groups: (a) TLE– MTS and controls and (b) TLE-noand controls to test for associations between subfield volumes or thickness measures andmemory scores in each of the TLE subgroups. TLEs were combined with controls afterensuring that there was an overlap between the two groups by correlation plots to allow forthe assessment of the full range of structural– functional associations. In a first step, partialcorrelation analyses (corrected for log of years of education) were used to identify subfieldsand cortical ROIs, which were significantly correlated with the memory scores. However,subfield volumes and thickness measures of neighboring ROI were correlated with eachother (subfield volumes r: range, 0.02–0.76; cortical ROI r: range, 0.08–0.9). Therefore,stepwise linear regression analyses (mixed, probability to enter/ leave P = 0.05) with thememory score as dependent, and those subfield volumes, which had been identified as beingsignificantly correlated with this score in the previous analysis as independent variables,were performed in the next step. Years of education (log transformed) were forced into themodel as independent variable, so that only subfields explaining a significant amount of thevariation of the memory performance in addition to that explained by years of educationcould be included in the stepwise regression model. Group was not modeled, because it wasintended to test for structural–cognitive correlations over the whole range of memoryimpairment and subfield volumes represented in the two groups. The contribution of thecortical ROI to memory performance was tested with the same approach. Finally, todetermine if cortical ROI or the hippocampal subfields were stronger associated withmemory performance, a third stepwise regression was performed with the cortical ROIs andhippocampal subfields selected by the previous analysis as dependent variables. Given the apriori hypotheses outlined in the introduction, corrections for multiple comparisons duringthe initial partial correlation analyses were not performed for correlations between memoryscores and hippocampal subfields, mesial temporal, and frontal ROIs. For parietal ROIs, forwhich no a priori hypotheses existed, the threshold for significance was set at P < 0.004(Bonferroni correction based on 12 cortical ROI). All statistical analyses were done in JMP8 (SAS Institute).
Mueller et al. Page 5
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

RESULTSGroup Comparisons
The MANCOVA for subfields showed a significant effect for group [Roy’s Max Root 1.24,F(5, 55) = 13.66, P < 0.0001] but not for age, gender, or years of education. TheMANCOVA for cortical ROIs showed a trend for group [Roy’s Max Root 0.57, F(12, 49) =1.76, P = 0.08], there were no significant effects for gender, age, or years of education. TheMANCOVA for memory scores finally showed a significant effect for group [Roy’s MaxRoot: 0.58, F(3, 58) = 11.34; P = <0.0001] and years of education [0.33, F(3,57) = 6.25; P =0.001] but not for gender or age. AIMrecall and ADMrecall were significantly lower inTLE–MTS and TLE-no compared to controls but not different between the two TLE groups(cf. Fig. 2a); ADMrecog was worse in TLE–MTS compared to controls; TLE-no was notdifferent from the other two groups. Left and right TLE did not differ from each otherregarding verbal memory performance (cf. Fig. 2b). Figures 3 and 4 provide an overview ofthe subfield volumes and thickness measures in the three groups.
Associations Between Memory Scores and Brain StructuresPlease refer Figure 4 showing the corresponding scatter plots. TLE–MTS and controls:Table II displays the results of the partial correlation analyses. The results of the stepwiseregression analysis were as follows: on the hippocampal level, AIMrecall (β = 0.23, P =0.0012) was associated with CA3&DG. ADMrecall (β = 0.19, P = 0.0003) and ADMrecog(β = 0.11, P = 0.0261) were both associated with CA1. On the cortical level, AIMrecall andADMrecall were both associated with inferior lateral prefrontal cortical thickness.AIMrecall was associated with POP thickness (β = 32.17, P = 0.0140) and ADMrecall withPT thickness (β = 33.71, P = 0.004). When CA3&DG and POP were entered together in themodel, both were significantly associated with AIMrecall [CA3&DG: β = 0.20, P = 0.0037;R2 adjusted (education, CA3&DG) = 0.35, POP: β = 24.32; P = 0.0416, R2 adjusted(education, CA3&DG, POP) = 0.40]. Similarly, when CA1 and PT were both entered intothe model for ADMrecall, both fulfilled the selection criteria [CA1: β = 0.17, P = 0.0006, R2
adjusted (education, CA1) = 0.26) and PT: β = 27.35, P = 0.0074, R2 adjusted (education,CA1, PT) = 0.34].
TLE-no and controls: Table III displays the results of the partial correlation analysis. Theresults of the stepwise regression analysis were as follows: on the hippocampal level,AIMrecall was associated with CA3&DG (β = 0.15, P = 0.0431) and on the cortical level,with FUSI thickness (cf Table III). When CA3&DG and FUSI were both entered into themodel, both contributed significantly to AIMrecall variability and thus were selected by themodel [FUSI: β = 17.54, P = 0.0164, R2 adjusted (education, FUSI) = 0.30; CA3&DG: β =0.15, P = 0.0290, R2 adjusted (education, FUSI, CA3&DG) = 0.36]. FUSI was also thecortical region explaining most of the variability of ADMrecall (β = 24.2, P = 0.0012) andwas the only cortical region, which was significantly associated with ADMrecog (cf. TableIII).
DISCUSSIONThere were two major findings in this study. (1) As predicted by our a priori hypothesis,hippocampal volume loss was the strongest determinant of verbal memory deficits in TLE–MTS. CA3&DG was associated with impaired early recall (AIMrecall) and CA1 withimpaired delayed recall (ADMrecall) and recognition (ADMrecog), suggesting a functionalspecialization by different hippocampal subfields. Cortical thinning of inferior lateralprefrontal cortical regions contributed to AIMrecall and ADMrecall impairment. (2) Verbalmemory deficits in TLE-no were associated with mesial temporal cortical (FUSI) and to a
Mueller et al. Page 6
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

lesser degree also with hippocampal damage. The associations on the hippocampal levelwere in accordance with the subfield specialization demonstrated in TLE–MTS. Prefrontalcortical damage contributed less to the memory impairment in this group than in TLE–MTS.These findings suggest that verbal episodic memory impairment has different structuralcorrelates in TLE–MTS and TLE-no.
Structural Correlates of AIMrecall in TLE–MTS and TLE-noIn accordance with our a priori hypothesis regarding TLE–MTS, we found a strongassociation between hippocampal subfield volume loss and AIMrecall impairment. Themajor finding in this regard was that AIMrecall was associated with volume loss inCA3&DG but not with volume loss in CA1 even though the volume loss in the twosubfields was of equal severity. This supports the notion that CA3&DG plays a crucial rolein rapid encoding, early consolidation, and retrieval in TLE–MTS and thus providesevidence for a functional specialization, which is in accordance with the subfieldspecialization shown for the normal hippocampus in animal studies [Acsady and Kali, 2007;Florian and Roullet, 2004; Gloor, 1997; Hunsaker and Kesner, 2008; Kesner, 2007; Lee etal., 2005; Nakashiba et al., 2008; Nakazawa et al., 2004; Rajji et al., 2006].
Structural–functional correlations in TLE–MTS, however, were not restricted to thehippocampus. AIMrecall was also associated with inferior lateral prefrontal (POP) thicknesswhich was thinner in TLE–MTS than in controls although not significantly so. WhenCA3&DG and POP were entered together in the stepwise regression model predictingAIMrecall, both were selected. This finding supports the close interaction between mesialtemporal and prefrontal regions during memory processes which has been described byother studies [Andersen et al., 2010; Burianova and Grady, 2007; Desgranges et al., 1998;Grady et al., 2005]. Although the exact contribution of the prefrontal cortex to the memoryperformance is not fully understood, it is generally assumed that the prefrontal cortexmonitors retrieval processes and suppresses potential intrusions [Cruse and Wilding, 2009;Vallesi and Shallice, 2006]. Therefore, the significant association between hippocampal andinferior lateral prefrontal regions with AIMrecall shown in this study could indicate thatCA3&DG damage results in a less reliable retrieval on the hippocampal level, whichrequires an enhanced monitoring by prefrontal regions if the early verbal retrievalperformance is to be sustained. However, in TLE–MTS with hippocampal and inferiorlateral prefrontal damage this compensation mechanism fails and consequently AIMrecallperformance suffers.
Subfield volumes in TLE-no were within the control range. Nonetheless, TLE-no showed astrong association between AIMrecall and CA3&DG volume. The finding of such anassociation even without obvious structural damage further supports the hypothesis that CA3and DG play an important role in encoding and early consolidation. In accordance with our apriori hypothesis of a contribution of medial temporal cortical damage to memoryimpairment in TLE-no, AIMrecall was also strongly associated with FUSI, which wasthinner in TLE-no than in controls. When FUSI and CA3&DG were entered together in thestepwise regression model, both explained a significant amount of the variation ofAIMrecall. This suggests that AIMrecall impairment in TLE-no is related to the dysfunctionof a larger hippocampal–neocortical temporal region. In contrast to TLE–MTS, there was noevidence that prefrontal thinning contributed to the AIMrecall impairment in this group.
Structural Correlates of ADMrecall and ADMrecog in TLE–MTS and TLE-noOur a priori hypotheses that memory impairment is mostly determined by hippocampaldamage in TLE–MTS and by neocortical damage in TLE-no were also confirmed forADMrecall and ADMrecog, which were both associated with CA1 in TLE–MTS and with
Mueller et al. Page 7
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

FUSI in TLE-no. The fact that on the hippocampal level significant correlations withADMrecall and ADMrecog were limited to CA1 in TLE–MTS provides further evidence fora functional specialization of the different hippocampal subfields and is in accordance withanimal and computational studies, which also show a special role for CA1 in late retrieval/recognition [Lai et al., 2005; Leutgeb et al., 2004; Takahashi and Sakurai, 2009]. CA1 hasextensive projections into the mesial–inferior temporal lobe [Ichinohoe and Rockland,2005], a region that includes FUSI, that is, the only neocortical region showing significantassociations with ADMrecog in TLE-no. Interestingly, the Freesurfer label FUSIencompasses the perirhinal cortex in its anterior section. The perirhinal cortex is the corticalregion, which has consistently been implicated in recognition performance although there isstill a debate as to the exact contributions of the hippocampus and perirhinal cortex torecognition [Aggleton and Brown, 2006; Squire et al., 2007; Kumaran et al., 2007].
In TLE–MTS, ADMrecall impairment was associated with inferior lateral prefrontal corticalthinning (PT). When CA1 and PT were entered together into the stepwise regressionanalysis, both contributed significantly to ADMrecall thus indicating that inferior–lateralprefrontal damage might contribute to ADMrecall impairment similarly as it does toAIMrecall impairment. In TLE-no prefrontal cortical thinning correlated with ADMrecallimpairment in the initial partial correlation analyses, but these associations becamenonsignificant in the stepwise regression after FUSI was selected by the model. This findingcould indicate that in TLE-no prefrontal damage also contributes to ADMrecall impairmentbut to a lesser degree than it does in TLE–MTS.
LimitationsThis study has several limitations. (1) The number of patients in the study is small, and thefindings need to be validated in larger patient population. This will allow to test if thememory—structural associations found in the combined groups, that is, controls/TLE–MTSand controls/TLE-no are also found when each TLE group is analyzed separately. In thecurrent study, the patient groups were too small for these associations to reach significance.Furthermore, a larger population will also allow to analyze left TLE and right TLE groupsseparately and to assess the influence of side of seizure onset and language lateralization onmemory performance. (2) Only verbal memory impairment was assessed in this study. Oneof the main objectives of this study was to replicate the findings of animal studies showingan association of CA3 and DG with encoding and early retrieval and of CA1 withrecognition. AIMrecall and ADMrecog were the only scores in the whole clinicalneuropsychological test battery, which allowed to test this association in this population,because they measure not only early retrieval (AIMrecall) but also recognition (ADMrecog)of the same information. (3) Although we used a stepwise regression analysis and thus arelatively conservative statistical approach, it is important to keep in mind that a significantassociation between structure and function does not necessarily also imply a causalrelationship. (4) In contrast to functional neuroimaging studies, which detect not only thenetworks involved in a specific cognitive function but also potential compensatorymechanisms, structural studies only detect regions whose damage is associated withimpairment of a particular function, that is, the failure to find a structural–functionalcorrelation does not allow for the conclusion that a brain region is not involved in aparticular cognitive process. (5) There is evidence from functional studies that the anteriorand posterior hippocampus have different specializations [Giovanello et al., 2009]. In thisstudy, subfield measurements were only obtained from the anterior third of the hippocampalbody and so we were not able to study the anterior–posterior effects of MTS on memoryfunction. (6) Three TLE-no subjects had extrahippocampal lesions. Although these lesionswere not included in any of the ROIs assessed in this study, we can not exclude that they hadan influence on the memory impairment in these patients. When these subjects were
Mueller et al. Page 8
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

excluded from analysis the findings did not change, indicating that the structural–functionalcorrelations in the TLE-no group were not driven by these three subjects.
In conclusion, this study provided evidence for a different structural correlate of the verbalmemory impairment in TLE–MTS and TLE-no. In TLE–MTS, the impairment was mostlyassociated with neuron loss in the hippocampus but was exaggerated by prefrontal thinningwhile it was associated with damage to hippocampal–mesial–temporal cortical regions inTLE-no. The memory impairment however was of similar severity in both groups. Thisindicates that it is the interruption of the network supporting a specific function and less theanatomical localization of the interruption within the network, which determines thememory impairment. The finding of a different structural correlate for memory impairmentin the two TLE groups also further supports the notion that TLE-no is not just a mild form ofTLE–MTS but a different entity of TLE.
AcknowledgmentsContract grant sponsor: National Institutes of Health; Contract grant number: RO1-NS31966.
REFERENCESAcsady L, Kali S. Models, structure, function: The transformation of cortical signals in the dentate
gyrus. Prog Brain Res. 2007; 163:577–599. [PubMed: 17765739]Aggleton J, Brown MW. Interleaving brain system for episodic and recognition memory. Trends Cogn
Sci. 2006; 10:455–463. [PubMed: 16935547]Alessio A, Damasceno BP, Camargo CHP, Kobayashi E, Guerreiro CAM, Cendes F. Differences in
memory performance and other clinical characteristics in patients with mesial temporal lobeepilepsy with and without hippocampal atrophy. Epilepsy Behav. 2004; 5:22–27. [PubMed:14751202]
Anderson KL, Rajagovindan R, Ghacibeh GA, Meador KJ, Ding M. Theta oscillations mediateinteraction between prefrontal cortex and medial temporal lobe in human memory. Cereb Cortex.2010; 20:1604–1612. [PubMed: 19861635]
Bernhardt BC, Worsley KJ, Besson P, Concha L, Lerch JP, Evans AC, Bernasconi N. Mapping limbicnetwork organization in temporal epilepsy using morphometric correlations: Insights on the relationbetween mesiotemporal connectivity and cortical atrophy. Neuroimage. 2008; 42:515–524.[PubMed: 18554926]
Bettus G, Guedj E, Joyeux F, Sylviane Confort-Gouny, Soulier E, Laguitton V, Cozzone PJ, ChauvelP, Ranjeva JP, Bartolomei F, Guye M. Decreased basal fMRI functional connectivity inepileptogenic networks and contralateral compensatory mechanisms. Hum Brain Mapp. 2009;30:1580–1591. [PubMed: 18661506]
Binder JR, Desai RH, Graves WW, Conant LL. Where is the semantic system? A critical review andmeta-analysis of 120 functional neuroimaging studies. Cereb Cortex. 2009; 19:2767–2796.[PubMed: 19329570]
Burianova H, Grady CL. Common and unique neural activations in autobiographical, episodic andsemantic retrieval. J Cogn Neurosci. 2007; 19:1520–1534. [PubMed: 17714013]
Cabeza R, Nyberg L. Neural bases of learning and memory: Functional neuroimaging evidence. CurrOpin Neurol. 2000; 13:415–421. [PubMed: 10970058]
Campo P, Maestu F, Garcia-Morales I, Gil-Nagel A, Strange B, Morales M, Ortiz T. Modulation ofmedial temporal lobe activity in epilepsy patients with hippocampal sclerosis during verbalworking memory. J Int Neuropsychol Soc. 2009; 15:536–546. [PubMed: 19573272]
Cruse D, Wilding EL. Prefrontal cortex contributions to episodic retrieval monitoring and evaluation.Neuropsychologia. 2009; 47:2779–2789. [PubMed: 19523968]
Dale AM, Fischl B, Sereno MI. Cortical surface-based analysis I: Segmentation and surfacereconstruction. Neuroimage. 1999; 9:179–194. [PubMed: 9931268]
Mueller et al. Page 9
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Desgranges B, Baron JC, Eustache F. The functional neuroanatomy of episodic memory: The role ofthe frontal lobes, the hippocampal formation, and other areas. Neuroimage. 1998; 8:198–213.[PubMed: 9740762]
Desikan RS, Ségonne F, Fischl B, Quinn BT, Dickerson BC, Blacker D, Buckner RL, Dale AM,Maguire RP, Hyman BT, Albert MS, Killiany RJ. An automated labeling system for subdividingthe human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage. 2006;31:968–980. [PubMed: 16530430]
Dulay MF, Schefft BK, Fargo JD, Privitera MD, Yeh HS. Severity of depressive symptoms,hippocampal sclerosis, auditory memory and seizure focus in temporal lobe epilepsy. EpilepsyBehav. 2004; 5:522–531. [PubMed: 15256190]
Duvernoy, HM. The Human Hippocampus, Functional anatomy, Vascularization and Serial Sectionswith MRI. 3rd ed.. Berlin: Springer-Verlag; 2005. p. 232
Eichenbaum H. Hippocampus: Cognitive processes and neural representations that underlie declarativememory. Neuron. 2004; 44:109–120. [PubMed: 15450164]
Florian C, Roullet P. Hippocampal CA3-region is crucial for acquisition and memory consolidation inMorris water maze task in mice. Behav Brain Res. 2004; 154:365–374. [PubMed: 15313024]
Giovanello KS, Schnyer D, Verfaellie M. Distinct hippocampal regsions make unique contributions torelational memory. Hippocampus. 2009; 19:111–117. [PubMed: 18727049]
Gloor, P. The Temporal Lobe and Limbic System. New York: Oxford University Press; 1997.Grady CL, McIntosh AR, Craik FIM. Task-related activity in prefrontal cortex and its relation to
recognition memory performance in young and old adults. Neuropsychologia. 2005; 43:1466–1481. [PubMed: 15989937]
Hunsaker MR, Kesner RP. Evaluating the differential roles of the dorsal dentate gyrus, dorsal CA3,and dorsal CA1 during a temporal ordering for spatial locations task. Hippocampus. 2008; 18:955–964. [PubMed: 18493930]
Ichinohe N, Rockland KS. Zinc-enriched anygdalo- and hippocampo-cortical connections to theinferotemporal cortices in macaque monkey. Neurosci Res. 2005; 53:57–68. [PubMed: 16023236]
Jokeit H, Schacher M. Neuropsychological aspects of type of epilepsy and etiological factors in adults.Epilepsy Behav Suppl. 2004; 1:S14–S20.
Kesner RP. Behavioral functions of the CA3 subregion of the hippocampus. Learn Mem. 2007;14:771–781. [PubMed: 18007020]
Kumaran D, Maguire EA. Which computational mechanisms operate in the hippocampus duringnovelty detection? Hippocampus. 2007; 17:735–748. [PubMed: 17598148]
Lai WS, Ramiro LLR, Yu HA, Johnston RE. Recognition of familiar individuals in golden hamsters:A new method and functional neuroanatomy. J Neurosci. 2005; 25:11239–11247. [PubMed:16339019]
Lee I, Hunsaker MR, Kesner RP. The role of the hippocampal subregions in detecting spatial novelty.Behav Neuro-science. 2005; 119:145–153.
Leutgeb S, Leutgeb JK, Treves A, Moser M, Moser EI. Distinct ensemble codes in hippocampal areasCA3 and CA1. Science. 2004; 305:1296–1298.
Lin JJ, Salamon N, Lee AD, Dutton RA, Geaga JA, Hayashi KM, Luders E, Toga AW, Engel J,Thompson PM. Reduced cortical thickness and complexity mapped in mesial temporal lobeepilepsy with hippocampal sclerosis. Cereb Cortex. 2007; 17:2007–2018. [PubMed: 17088374]
Moscovitch M, Rosenbaum RS, Gilboa A, Addis DR, Westmacott R, Grady C, McAndrews MP,Levine B, Black S, Winocur G, Nadel L. Functional neuroanatomy of remote episodic, semanticand spatial memory: A unified account based on multiple trace theory. J Anat. 2005; 207:35–66.[PubMed: 16011544]
Mueller SG, Stables L, Du AT, Schuff N, Truran D, Cashdollar N, Weiner MW. Measurement ofhippocampal subfields and age-related changes with high resolution MRI at 4T. Neurobiol Aging.2007; 28:719–726. [PubMed: 16713659]
Mueller SG, Laxer KD, Barakos J, Cheong I, Garcia P, Weiner MW. Subfield atrophy pattern intemporal lobe epilepsy with and without mesial sclerosis detected by high resolution MRI at 4Tesla: Preliminary results. Epilepsia. 2009a; 50:1474–1483. [PubMed: 19400880]
Mueller et al. Page 10
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Mueller SG, Laxer KD, Barakos J, Cheong I, Garcia P, Weiner MW. Widespread neocorticalabnormalities in temporal lobe epilepsy with and without mesial sclerosis. Neuroimage. 2009b;46:353–359. [PubMed: 19249372]
Nakashiba T, Young JZ, McHugh TJ, Buhl DL, Tonegawa S. Transgenic inhibition of synaptictransmission reveals role of CA3 output in hippocampal learning. Science. 2008; 319:1260–1263.[PubMed: 18218862]
Nakazawa K, McHugh TJ, Wilson MA, Tonegawa S. NMDA receptors, place cells and hippocampalspatial memory. Nat Rev. 2004; 5:361–370.
Nyberg L. Mapping episodic memory. Behav Brain Res. 1998; 90:107–114. [PubMed: 9521543]O’Reilly CR, Rudy JW. Conjunctive representation in learning and memory: Principles of cortical and
hippocampal function. Psychol Rev. 2001; 108:311–345. [PubMed: 11381832]Rajjii T, Chapman D, Eichenbaum H, Greene R. The role of CA3 hippocampal NMDA receptors in
paired associate learning. J Neurosci. 2006; 26:908–915. [PubMed: 16421310]Richardson MP, Strange BA, Duncan JS, Dolan RJ. Preserved verbal memory function in left medial
temporal pathology involves reorganization of function to right medial temporal lobe.Neuroimage. 2003; 20:S112–S119. [PubMed: 14597304]
Rolls ET, Kesner RP. A computational theory of hippocampal function and empirical test of thetheory. Prog Neurobiol. 2006; 79:1–48. [PubMed: 16781044]
Squire LR, Wixted JT, Clark RE. Recognition memory and the medial temporal lobe: A newperspective. Nat Rev Neurosci. 2007; 8:872–883. [PubMed: 17948032]
Takahashi S, Sakurai Y. Information in small neuronal ensemble activity in the hippocampal CA1during delayed non-matching to sample performance in rats. BMC Neurosci. 2009; 10:115.[PubMed: 19751528]
Vallesi A, Shallice T. Prefrontal involvement in source memory: An electrophysiological investigationof accounts concerning confidence and accuracy. Brain Res. 2006; 1124:1111–1125.
Van Leemput K, Maes F, Vandermeulen D, Suetens P. Automated model-based bias field correction ofMR images of the brain. IEEE Trans Med Imaging. 1999a; 18:885–896. [PubMed: 10628948]
Van Leemput K, Maes F, Vandermeulen D, Suetens P. Automated model-based tissue classification ofMR images of the brain. IEEE Trans Med Imaging. 1999b; 18:897–908. [PubMed: 10628949]
Wagner K, Frings L, Halsband U, Everts R, Buller A, Spreer J, Zentner J, Schulze-Bonhage A.Hippocampal functional connectivity reflects verbal episodic memory network integrity.Neuroreport. 2007; 18:1719–1723. [PubMed: 17921875]
Wan H, Aggleton JP, Brown MW. Different contributions of the hippocampus and perirhinal cortex torecognition memory. J Neurosci. 1999; 19:1142–1148. [PubMed: 9920675]
Mueller et al. Page 11
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
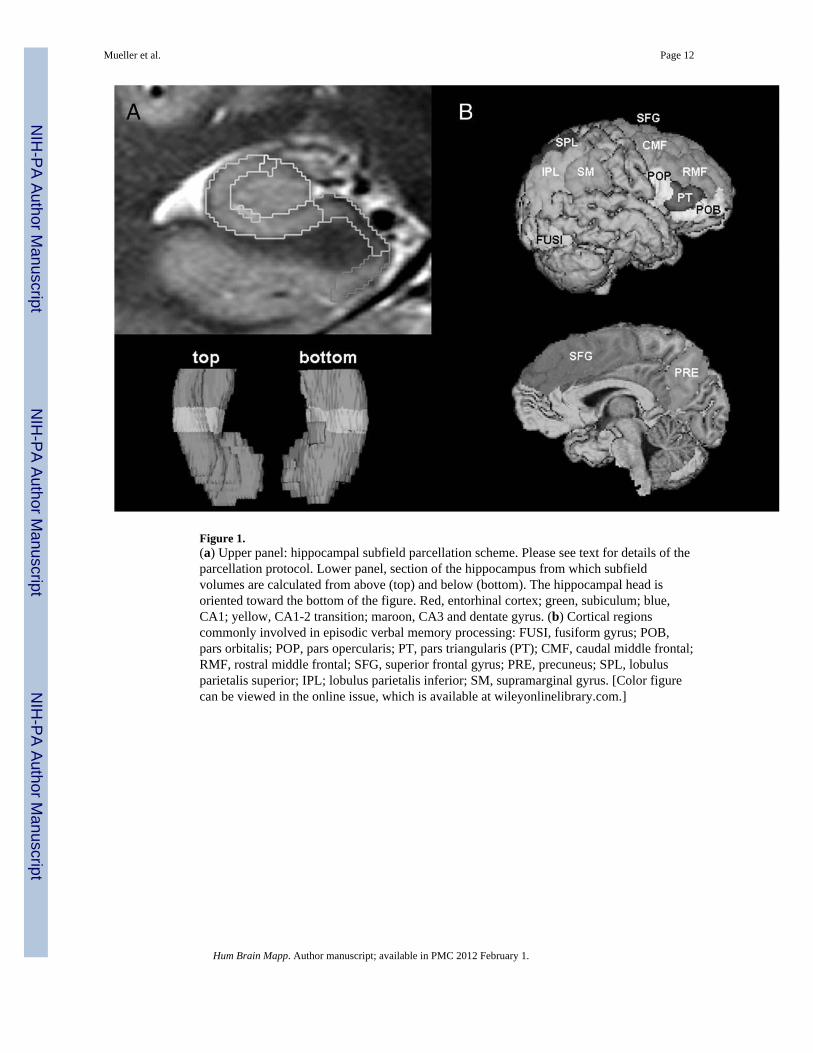
Figure 1.(a) Upper panel: hippocampal subfield parcellation scheme. Please see text for details of theparcellation protocol. Lower panel, section of the hippocampus from which subfieldvolumes are calculated from above (top) and below (bottom). The hippocampal head isoriented toward the bottom of the figure. Red, entorhinal cortex; green, subiculum; blue,CA1; yellow, CA1-2 transition; maroon, CA3 and dentate gyrus. (b) Cortical regionscommonly involved in episodic verbal memory processing: FUSI, fusiform gyrus; POB,pars orbitalis; POP, pars opercularis; PT, pars triangularis (PT); CMF, caudal middle frontal;RMF, rostral middle frontal; SFG, superior frontal gyrus; PRE, precuneus; SPL, lobulusparietalis superior; IPL; lobulus parietalis inferior; SM, supramarginal gyrus. [Color figurecan be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Mueller et al. Page 12
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2.Standardized auditory memory subscores from Wechsler Memory Scale III. AIMrecall,auditory immediate memory tested by recall; ADMrecall, auditory delayed memory testedby recall; ADMrecog, auditory delayed memory tested by recognition. (a) Memory scores inthe three study groups. Yellow, controls; orange; TLE with normal MRI; red, TLE withhippocampal sclerosis. Asterisks indicate scores significantly different from the controlgroup (P < 0.05). (b) Memory scores in TLE with right- and left-sided focus. Dark blue, leftTLE-MTS; bright blue, right TLE-MTS; orange, left TLE-no; yellow, right TLE-no. Therewere also no significant differences in the TLE subgroups (Mann–Whitney tests with
Mueller et al. Page 13
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Holm’s test to correct for multiple comparisons). [Color figure can be viewed in the onlineissue, which is available at wileyonlinelibrary.com.]
Mueller et al. Page 14
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3.(a) Mean and standard deviation of hippocampal subfield volumes corrected for intracranialhead size (please see text for correction formula). Yellow, controls; orange; TLE withnormal MRI; red, TLE with hippocampal sclerosis. Asterisks indicate scores that aresignificantly different from the control group (P < 0.05). (b) Thickness of the corticalregions of interest (mean and standard deviation of the sum of both sides) in millimeters.PARA, parahippocampal gyrus; FUSI, fusiform gyrus; POB, pars orbitalis; POP, parsopercularis; PT, pars triangularis (PT); CMF, caudal middle frontal; RMF, rostral middlefrontal; SFG, superior frontal gyrus; PRE, precuneus; SPL, lobulus parietalis superior; IPL;lobulus parietalis inferior; SM, supramarginal gyrus. Yellow, controls; orange; TLE with
Mueller et al. Page 15
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

normal MRI; red, TLE with hippocampal sclerosis. [Color figure can be viewed in theonline issue, which is available at wileyonlinelibrary.com.]
Mueller et al. Page 16
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 4.Scatter plots of subfield volumes and cortical ROI thickness versus WMS-III scores(without accounting for years of education). Blue squares, controls; red squares, TLE-MTS;yellow squares, TLE-no; recall AIMrecall, auditory immediate recall; ADMrecall, auditorydelayed recall; ADMrecog, auditory delayed recognition; FUSI, fusiform gyrus; POP, parsopercularis; PT, pars triangularis; CA3&DG, CA3 and dentate gyrus. [Color figure can beviewed in the online issue, which is available at wileyonlinelibrary.com.]
Mueller et al. Page 17
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Mueller et al. Page 18
TAB
LE I
Patie
nts c
hara
cter
istic
s
No
Gen
der/
age
Age
at o
nset
Han
dedn
ess
Focu
s1.
5 M
RI
4 T
MR
I
1F/
2813
LR
RM
TSG
loba
l
2F/
497
RR
RM
TSC
A1
3M
/35
4R
LLM
TSG
loba
l
4F/
502
RL
LMTS
Glo
bal
5F/
235
RR
RM
TSC
A1&
DG
6M
/43
5L
RR
MTS
CA
1&D
G
7F/
359
LL
LMTS
CA
1&D
G
8M
/44
28R
LLM
TSG
loba
l
9F/
4813
RL
LMTS
CA
1&D
G
10F/
386
RL
RM
TSC
A1&
DG
11F/
5612
RR
RM
TSC
A1
12F/
3314
RL
LMTS
CA
1
13M
/63
6R
RR
MTS
CA
1
14F/
5328
RL
LMTS
CA
1&D
G
15F/
3224
RL
LMTS
CA
1
16M
/32
1L
RR
MTS
CA
1
17F/
5628
RL
RM
TSD
G
18M
/23
1L
LLM
TSC
A1
19M
/39
29R
LN
orm
alC
ystic
lesi
on le
ft in
ferio
r tem
pora
l pol
e
20M
/29
11R
RN
orm
alN
orm
al
21F/
3839
RL
Nor
mal
Nor
mal
22F/
4320
LR
Nor
mal
Nor
mal
23F/
2222
RR
Nor
mal
Left
fron
tal v
enou
s ang
iom
a
24M
/57
13R
LN
orm
alN
orm
al
25M
/45
30R
RN
orm
alN
orm
al
26F/
3030
ambi
RN
orm
alEc
topi
c gr
ay m
atte
r adj
acen
t rig
ht a
myg
dala
27F/
3532
RL
Nor
mal
Nor
mal
28F/
4221
RL
Nor
mal
Nor
mal
29F/
3837
RL
Nor
mal
Nor
mal
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Mueller et al. Page 19
No
Gen
der/
age
Age
at o
nset
Han
dedn
ess
Focu
s1.
5 M
RI
4 T
MR
I
30M
/39
13R
LN
orm
alN
orm
al
31F/
3322
RL
Nor
mal
Nor
mal
32F/
5118
RL
Nor
mal
Nor
mal
33M
/56
49R
RN
orm
alN
orm
al
34M
/41
40R
RN
orm
alN
orm
al
35M
/37
35R
biN
orm
alN
orm
al
36M
/26
1R
biN
orm
alN
orm
al
37F/
3130
RL
Nor
mal
Nor
mal
38F/
5343
Rbi
Nor
mal
Nor
mal
39F/
4812
Rbi
Nor
mal
Nor
mal
40F/
5347
LL
Nor
mal
Nor
mal
41F/
4815
RR
Nor
mal
Nor
mal
42M
/43
12R
LN
orm
alN
orm
al
43F/
2118
RL
Nor
mal
Nor
mal
F, fe
mal
e; M
, mal
e; R
, rig
ht; L
, lef
t; am
bi, a
mbi
dext
er; F
ocus
, lat
eral
izat
ion
of th
e fo
cus b
ased
on
VET
; 1.5
T M
RI;
Find
ings
on
the
1.5
T M
RI;
RM
TS ri
ght m
esia
l tem
pora
l scl
eros
is; L
MTS
, lef
t mes
ial
tem
pora
l scl
eros
is; 4
T H
ippo
; atro
phy
patte
rn o
n 4-
T hi
gh-r
esol
utio
n im
age.
All
volu
mes
wer
e co
nver
ted
to z-
scor
es z-
scor
e =
volu
me
patie
nt –
mea
n vo
lum
e co
ntro
ls/s
tand
ard
devi
atio
n co
ntro
ls. S
ubfie
lds
with
a z-
scor
es ≤
−2
wer
e ar
bitra
rily
defin
ed a
s atro
phic
(cf.
Mue
ller e
t al.,
200
9). G
loba
l, C
A1,
CA
1-2
trans
ition
, and
CA
3&D
G a
troph
y. C
A1,
isol
ated
CA
1 at
roph
y; D
G, i
sola
ted
CA
3&D
G a
troph
y;C
A1&
DG
, atro
phy
of C
A1
and
CA
3&D
G.
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Mueller et al. Page 20
TAB
LE II
Parti
al c
orre
latio
ns v
erba
l mem
ory
scor
es w
ith su
bfie
ld v
olum
es a
nd c
ortic
al th
ickn
ess i
n TL
E-M
TS a
nd c
ontro
ls
ER
CSU
BC
A1
CA
1-2
CA
3&D
GSF
GR
MF
CM
FPO
BPO
PPT
PRE
SPL
IPL
SMPA
RA
FUSI
AIM
reca
lR
0.39
0.28
0.49
0.32
0.50
0.10
0.24
0.12
0.23
0.39
0.37
0.20
0.27
0.19
0.27
0.13
0.19
P<0
.01
0.04
<0.0
10.
02<0
.01
nsns
nsns
<0.0
1<0
.01
ns0.
04ns
0.05
nsns
AD
M re
cal
R0.
220.
200.
550.
320.
550.
130.
320.
150.
350.
410.
450.
280.
290.
310.
350.
240.
32
Pns
ns<0
.01
0.02
<0.0
1ns
0.02
0.02
0.01
<0.0
1<0
.01
0.04
0.04
0.02
0.01
ns0.
02
AD
M re
cog
R0.
030.
190.
360.
310.
300.
030.
15−0.12
0.05
0.03
0.04
0.11
0.14
−0.01
−0.01
0.10
0.24
Pns
ns0.
010.
020.
02ns
nsns
nsns
nsns
nsns
nsns
ns
AIM
reca
ll, a
udito
ry im
med
iate
ver
bal m
emor
y te
sted
by
reca
ll; A
DM
reca
ll, a
udito
ry d
elay
ed m
emor
y te
sted
by
reca
ll; A
DM
reca
ll, a
udito
ry d
elay
ed re
cogn
ition
test
ed b
y re
cogn
ition
; R, c
orre
latio
nco
effic
ient
; P, o
ne-s
ided
sign
ifica
nce;
ER
C, e
ntor
hina
l cor
tex;
SU
B, s
ubic
ulum
; CA
1-2;
CA
1-2
trans
ition
; CA
3&D
G, C
A3
and
dent
ate
gyru
s; S
FG, s
uper
ior f
ront
al g
yrus
; RM
F ro
stra
l mid
dle
fron
tal;
CM
F, c
auda
l mid
dle
fron
tal;
POB
, par
s orb
italis
; PO
P, p
ars o
perc
ular
is; P
T, p
ars t
riang
ular
is; P
RE,
pre
cune
us; S
PL; s
uper
ior p
arie
tal l
obul
e; IP
L, in
ferio
r par
ieta
l lob
ule;
SM
, sup
ram
argi
nal g
yrus
;; PA
RA
,pa
rahi
ppoc
ampa
l gyr
us; F
USI
, fus
iform
gyr
us. B
old,
sign
ifica
nt e
ffec
t in
first
and
seco
nd le
vel s
tepw
ise
regr
essi
on a
naly
sis (
hipp
ocam
pal o
r cor
tical
stru
ctur
es c
f tex
t); b
old
italic
, sig
nific
ant e
ffec
t in
seco
nd le
vel s
tepw
ise
regr
essi
on a
naly
sis (
hipp
ocam
pal a
nd c
ortic
al st
ruct
ures
cf t
ext).
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Mueller et al. Page 21
TAB
LE II
I
Parti
al c
orre
latio
ns v
erba
l mem
ory
scor
es w
ith su
bfie
ld v
olum
es a
nd c
ortic
al th
ickn
ess i
n TL
E-no
and
con
trols
ER
CSU
BC
A1
CA
1-2
CA
3&D
GSF
GR
MF
CM
FPO
BPO
PPT
PRE
SPL
IPL
SMPA
RA
FUSI
AIM
reca
llR
0.15
−0.01
0.16
0.29
0.30
0.02
0.11
0.00
0.15
0.21
0.07
0.10
0.04
0.10
0.13
−0.04
0.33
Pns
nsns
0.02
0.02
nans
Ns
nsns
nsns
nsns
nsns
0.01
AD
M re
call
R0.
05−0.06
0.13
0.20
0.17
0.18
0.26
0.14
0.30
0.33
0.24
0.30
0.18
0.27
0.30
0.12
0.46
Pns
nsns
nsns
ns0.
04N
s0.
020.
01ns
0.02
ns0.
030.
02ns
<0.0
1
AD
M re
cog
R0.
140.
200.
200.
170.
13−0.01
0.07
−0.05
0.05
0.02
−0.03
0.01
0.02
0.00
0.00
0.20
0.34
Pns
nsns
nsN
sns
nsN
sns
nsns
nsns
nsns
ns<0
.01
AIM
reca
ll, a
udito
ry im
med
iate
ver
bal m
emor
y te
sted
by
reca
ll; a
udito
ry d
elay
ed m
emor
y te
sted
by
reca
ll; a
udito
ry d
elay
ed re
cogn
ition
test
ed b
y re
cogn
ition
; R, c
orre
latio
n co
effic
ient
; P, o
ne-s
ided
sign
ifica
nce;
ER
C, e
ntor
hina
l cor
tex;
SU
B, s
ubic
ulum
; CA
1-2;
CA
1-2
trans
ition
; CA
3&D
G, C
A3
and
dent
ate
gyru
s; S
FG, s
uper
ior f
ront
al g
yrus
; RM
F ro
stra
l mid
dle
fron
tal;
CM
F, c
auda
l mid
dle
fron
tal;
POB
, par
s orb
italis
; PO
P, p
ars o
perc
ular
is; P
T, p
ars t
riang
ular
is; P
RE,
pre
cune
us; S
PL; s
uper
ior p
arie
tal l
obul
e; IP
L, in
ferio
r par
ieta
l lob
ule;
SM
, sup
ram
argi
nal g
yrus
; PA
RA
, par
ahip
poca
mpa
l gyr
us;
FUSI
, fus
iform
gyr
us; b
old,
sign
ifica
nt e
ffec
t in
first
and
seco
nd le
vel s
tepw
ise
regr
essi
on a
naly
sis (
hipp
ocam
pal o
r cor
tical
stru
ctur
es c
f tex
t), b
old
italic
, sig
nific
ant e
ffec
t in
seco
nd le
vel s
tepw
ise
regr
essi
on a
naly
sis (
hipp
ocam
pal a
nd c
ortic
al st
ruct
ures
cf t
ext).
Hum Brain Mapp. Author manuscript; available in PMC 2012 February 1.





























